Deep Research
Falcon
(AT5G16820-deep-research-falcon.md)
Falcon
(AT5G16820-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Comprehensive research report: Arabidopsis thaliana HSFA1B (AT5G16820; UniProt O81821)
0) Scope and target verification (critical)
The target in this report is Arabidopsis thaliana HSFA1B (locus At5g16820, UniProt O81821), annotated as Heat stress transcription factor A-1b (AtHsfA1b) and a member of the plant HSF family, class A1. Multiple authoritative sources explicitly map HSFA1B to At5g16820 and place it within the four Arabidopsis HSFA1 isoforms (HSFA1a/b/d/e) that function as top-tier regulators of heat responses. (wang2023transcriptionalregulatorsof pages 2-4, liu2011theroleof pages 1-2, liu2011theroleof pages 6-7)
1) Key concepts and definitions (current understanding)
1.1 Heat-shock response (HSR) and HSFs
The plant heat-shock response (HSR) is a transcriptional program induced by elevated temperature (and associated proteotoxic stress) that promotes proteostasis and survival, largely via induction of heat shock proteins (HSPs). In plants, heat shock transcription factors (HSFs) are the core transcriptional regulators of HSR. HSFs recognize heat shock elements (HSEs) in promoters, typically built from nGAAn repeats (e.g., GAAnnTTC) and activate transcription of HSPs and other stress-response genes. (liu2013commonanddistinct pages 1-2, wang2023transcriptionalregulatorsof pages 2-4)
1.2 Class A1 HSFs as master regulators
Arabidopsis contains 21 HSFs, but the class A1 HSFs (HSFA1a, HSFA1b, HSFA1d, HSFA1e) are repeatedly described as master regulators because they sit upstream of large fractions of HS-induced transcription and trigger downstream HSF cascades (e.g., induction of HSFA2). (liu2013commonanddistinct pages 1-2, liu2011theroleof pages 1-2, wang2023transcriptionalregulatorsof pages 2-4)
1.3 “Master regulator” versus “molecular rheostat” framing
Recent synthesis argues that HSFs are not simply ON/OFF switches; rather, their activity acts as a dynamic rheostat controlling intensity and timing of stress protection versus recovery/growth, using partner-protein interactions and feedback attenuation. This conceptual framing is important for interpreting HSFA1B phenotypes, which are often subtle in single mutants because network-level buffering tunes system output. (bakery2024heatstresstranscription pages 4-4)
2) HSFA1B molecular function and mechanism
2.1 Molecular function: sequence-specific transcription factor
HSFA1B is a sequence-specific DNA-binding transcription factor. As a class A HSF, it contains a transactivation motif enabling transcriptional activation, and it binds HSEs to promote expression of HSR genes and other stress-adaptive programs. (liu2013commonanddistinct pages 1-2, liu2011theroleof pages 1-2)
2.2 DNA binding and promoter logic
Genome-wide profiling shows HSFA1B recognizes canonical HSEs and co-occurs with other promoter motifs such as G-box, CArG (MADS-box), LEAFY, and UPRE motifs, consistent with combinatorial regulation of stress and development. (albihlal2018arabidopsisheatshock pages 10-12, albihlal2018arabidopsisheatshock pages 1-4)
2.3 Regulation/activation: chaperone repression and release
A widely supported model is that at ambient temperature, HSFA1 proteins are repressed by direct association with HSP70/HSP90, limiting transcriptional activation and even nuclear localization; heat-driven protein misfolding titrates chaperones away and releases HSFA1s to activate transcription. (wang2023transcriptionalregulatorsof pages 2-4, bakery2024heatstresstranscription pages 4-4)
2.4 Post-translational and signaling integration
HSFA1 activity is tuned by post-translational modifications including phosphorylation and SUMOylation (and ubiquitination in some models), and by interactions with kinases/phosphatases (e.g., CDKA1, CBK3, PP7) and negative regulators (e.g., HSBP, HSFB antagonists, and other attenuation factors). These layers are central to the “rheostat” view of HSF function. (wang2023transcriptionalregulatorsof pages 2-4, bakery2024heatstresstranscription pages 4-4, bakery2024heatstresstranscription pages 3-3)
3) Biological roles of HSFA1B in Arabidopsis
3.1 Early HSR and thermotolerance; redundancy within HSFA1s
Genetic and transcriptome evidence supports HSFA1B as part of a partially redundant HSFA1 module that is required for robust thermotolerance and HSR. A key quantitative estimate from hsfa1 loss-of-function analyses is that more than 65% of heat-upregulated genes are HSFA1-dependent. (liu2011theroleof pages 1-2)
A detailed microarray comparison reported that, across two Arabidopsis ecotypes, the hsfa1 quadruple mutant shows >3-fold reduction for 230/346 (66.5%) and 278/408 (68.1%) heat-upregulated genes, with 209 genes affected in both ecotypes. (liu2011theroleof pages 6-7)
Within the HSFA1 clade, HSFA1a and HSFA1d are often described as stronger effectors, but HSFA1b contributes substantially and can act redundantly with other HSFA1s, particularly in the early phase of heat responses. (liu2011theroleof pages 6-7)
3.2 Systems-level target network: HSFA1B links stress defense with growth/development
A key HSFA1B-specific advance is genome-wide mapping of HSFA1B binding and transcript effects.
Albihlal et al. 2018 (J Exp Bot; published Apr 2018; https://doi.org/10.1093/jxb/ery142) performed HSFA1B ChIP-seq and RNA-seq under non-stress and heat stress (37°C, 30 min). They identified 709 binding peaks under non-stress and 1083 under heat stress, corresponding to 1207 HSFA1B target genes (q≤0.05; FE≥2). (albihlal2018arabidopsisheatshock pages 7-10)
The binding pattern reconfigures rapidly with heat: within 30 min of heat, HSFA1B ceases binding 124 genes (NS-unique group) and engages 553 genes (HS-unique group), demonstrating a fast switch in target selection between growth/development and stress defense. (albihlal2018arabidopsisheatshock pages 12-15)
On the expression side, heat triggered 7137 differentially expressed genes (DEGs), of which 721 were HSFA1B-bound; HSFA1B overexpression changed 3306 protein-coding genes, with 72% overlap with heat-regulated genes, and the overexpression transcriptome sits “between” NS and HS wild-type states (Pearson r=0.92 vs NS WT; r=0.88 vs HS WT). (albihlal2018arabidopsisheatshock pages 10-12, albihlal2018arabidopsisheatshock pages 7-10)
Together, these data support a model in which HSFA1B does not only activate classical HSP genes but also participates in developmental and reproductive regulation in ways that can affect fitness trade-offs. (albihlal2018arabidopsisheatshock pages 1-4)
3.3 Downstream pathways and transcriptional cascade
Reviews and primary research describe HSFA1s (including HSFA1B) as initiating a cascade that includes:
- Direct/indirect induction of HSFA2 and HSFA3 (HSFA3 partly via HSFA1-induced DREB2s), (kappel2023genomicandepigenomic pages 3-5)
- Induction of other TF regulators (e.g., DREB2A, HSFA7A/B, MBF1C), (wang2023transcriptionalregulatorsof pages 2-4)
- Induction and later attenuation by HSFB repressors (HSFB1/HSFB2 classes), (wang2023transcriptionalregulatorsof pages 2-4, bakery2024heatstresstranscription pages 4-4)
- Large-scale induction of HSP families that execute proteostasis protection. (li2025heatshocktranscription pages 6-7, wang2023transcriptionalregulatorsof pages 2-4)
3.4 Thermomorphogenesis: integration of warm temperature with light via PIF4
A major recent development is the linkage of HSFA1s to warm-temperature growth responses (thermomorphogenesis), not only extreme heat stress.
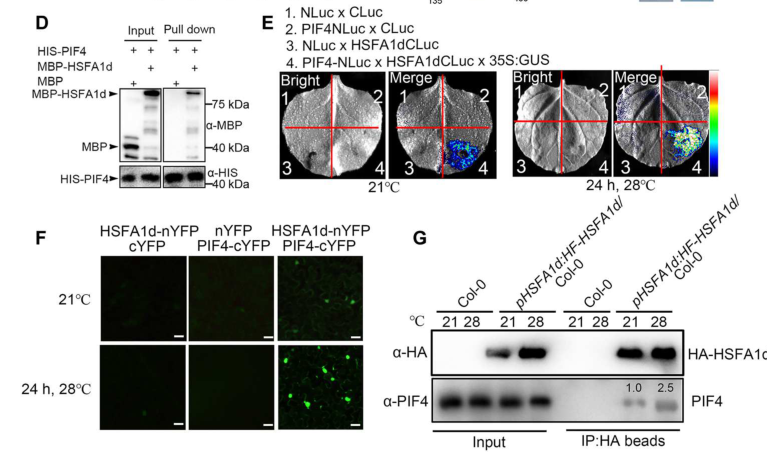
Tan et al. 2023 (Science Advances; published Nov 2023; https://doi.org/10.1126/sciadv.adh1738) reported that under warm daytime temperatures HSFA1 proteins accumulate and move into the nucleus, where they interact with PIF4 and stabilize it by interfering with the phyB–PIF4 interaction; HSFA1s are required for warm-temperature hypocotyl elongation phenotypes. (tan2023theheatresponse pages 1-2)
Figure evidence from this study supports nuclear interaction and nuclear relocalization (shown for HSFA1d, interpreted as HSFA1-family behavior): BiFC shows HSFA1d–PIF4 interaction in nuclei and strengthened at 28°C compared to 21°C, and imaging shows 28°C promotes HSFA1d-GFP nuclear translocation in light. (tan2023theheatresponse media 2354b667)
This work positions HSFA1 family members (including HSFA1B) as integrators of temperature and light signaling, beyond canonical HSR. (tan2023theheatresponse pages 1-2)
4) Cellular localization: where HSFA1B acts
HSFA1B is expected to execute its transcriptional function in the nucleus, and regulatory models explicitly connect repression/activation to nuclear localization control (HSP70/HSP90-mediated repression of nuclear localization at ambient conditions; release upon heat). (wang2023transcriptionalregulatorsof pages 2-4)
Direct imaging/localization in the most recent primary work is shown for HSFA1d (nuclear accumulation and nuclear PIF4 interaction) and is used to infer family-level behavior for HSFA1s under warm conditions; however, isoform-specific HSFA1B localization microscopy was not present in the extracted excerpts. (tan2023theheatresponse pages 1-2, tan2023theheatresponse media 2354b667)
5) Heat-stress memory (thermomemory) connections
5.1 HSFA1B is upstream of memory HSFs
A key recent genome-wide analysis connects early HSFA1 activity with memory-HSF induction.
Kappel et al. 2023 (Genome Biology; published May 2023; https://doi.org/10.1186/s13059-023-02970-5) states that HSFA1 isoforms (HSFA1A/HSFA1B/HSFA1D) are constitutively expressed and required for the early HS response; HSFA1s directly induce HSFA2, and promote HSFA3 activation via DREB2 intermediates. (kappel2023genomicandepigenomic pages 1-3, kappel2023genomicandepigenomic pages 3-5)
5.2 Memory gene determinants and quantitative genome-wide data
Kappel et al. performed time-course ChIP-seq for memory factors HSFA2/HSFA3 and found binding is strongly sequence-determined and memory genes are enriched for a tripartite HSE, accessible chromatin, and heat-induced H3K4me3. They clustered peaks into 15 clusters; 7 HS-induced clusters comprised 4,948 peaks as putatively biologically relevant targets. (kappel2023genomicandepigenomic pages 3-5)
They also report in vitro binding assays including HSFA1b (at 25°C vs 37°C) to explore DNA sequence contributions to binding. (kappel2023genomicandepigenomic pages 5-8)
6) Current applications and real-world implementations
6.1 Engineering principles: why HSFA1-family manipulation is attractive and risky
Because HSFA1s are upstream “master regulators,” they are attractive for engineering thermotolerance; however, broad activation can impose growth penalties. A recent review summarizes that inducible strategies may reduce trade-offs.
Wang et al. 2023 (IJMS; published Aug 2023; https://doi.org/10.3390/ijms241713297) discusses transgenic overexpression as a strategy to enhance heat tolerance in crops (with examples in other species) but emphasizes that constitutive overproduction may cause growth retardation and therefore highlights use of inducible promoters (including heat-inducible promoters). It also describes a programmable epigenome strategy: heat-inducible dCas9 fused to an H3K4 demethylase (JMJ) domain in Arabidopsis to target histone modification at selected loci, illustrating a modern implementation path for heat-response engineering. (wang2023transcriptionalregulatorsof pages 9-11)
6.2 HSFA1B network tuning: from overexpression to promoter engineering
HSFA1B-specific work shows that overexpression can “pre-poise” transcriptomes between growth and defense states, suggesting biotechnological utility but also likely pleiotropy. In Arabidopsis, HSFA1B overexpression increases stress resistance and can shift biomass allocation toward reproduction/seed yield in the Albihlal et al. framework, supporting a potential route for yield stability under stress. (albihlal2018arabidopsisheatshock pages 1-4)
While many explicit crop engineering examples in the retrieved set are for HSFA1 homologs in non-Arabidopsis systems, the mechanistic logic is transferable: HSFA1-family tuning of HSP and antioxidant/proteostasis networks is a common engineering target. (wang2023transcriptionalregulatorsof pages 9-11)
7) Consolidated evidence map
The following table provides a compact evidence-linked map of HSFA1B identity, mechanism, pathways, recent developments, and quantitative data.
| Category | Summary |
|---|---|
| Identity/domains | Arabidopsis thaliana HSFA1B corresponds to locus At5g16820 and UniProt O81821; it is one of four class A1 heat shock factors (HSFA1a/b/d/e) in the 21-member Arabidopsis HSF family. HSFs contain an HSF DNA-binding domain, oligomerization heptad-repeat regions, and class A members carry a C-terminal transactivation motif enabling activator function. HSFA1B is repeatedly placed in the master-regulator HSFA1 subgroup rather than being a separate non-HSF protein. (liu2013commonanddistinct pages 1-2, wang2023transcriptionalregulatorsof pages 2-4, liu2011theroleof pages 1-2) |
| Molecular function | HSFA1B functions as a sequence-specific transcription factor that binds heat shock elements (HSEs; GAAnnTTC/nGAAn repeats) in promoters, oligomerizes, and activates heat-responsive transcription. As part of HSFA1s, it regulates a large share of the early heat-shock transcriptome and can promote expression of other HSFs and HSP genes. Evidence supports partial redundancy with HSFA1a and HSFA1d, with HSFA1b contributing to the early phase of heat-shock gene activation rather than acting alone. (liu2013commonanddistinct pages 1-2, liu2011theroleof pages 1-2, guan2013molecularmechanismsof pages 58-64) |
| Activation/regulation | At normal temperatures, HSP70/HSP90 repress HSFA1 activity and can limit transcriptional activation and nuclear localization; heat-induced protein misfolding releases HSFA1s for activation. Activated HSFA1s undergo oligomerization, nuclear translocation, and are further modulated by phosphorylation, SUMOylation, and ubiquitination; kinases such as CDKA1/CBK3 and regulators including PP7, HSBP, BIN2, and other HSFB antagonists tune activity. HSFA1s are also integrated with ROS/Ca2+ signaling and chromatin accessibility. (wang2023transcriptionalregulatorsof pages 2-4, bakery2024heatstresstranscription pages 4-4, bakery2024heatstresstranscription pages 3-3) |
| Downstream targets | Reported HSFA1/HSFA1B downstream targets and network outputs include HSP genes, HSFA2, HSFA3 (indirectly via DREB2s), DREB2A, HSFA7A/B, MBF1C, and HSFB1/HSFB2A/HSFB2B. Recent thermomorphogenesis work shows HSFA1s also interact with PIF4, stabilizing it under warm daytime temperatures to connect heat and light signaling. In transcriptional-memory studies, HSFA1b was assayed together with HSFA2/HSFA3 in vitro, supporting its placement upstream of memory-HSF cascades. (wang2023transcriptionalregulatorsof pages 2-4, bakery2024heatstresstranscription pages 9-9, kappel2023genomicandepigenomic pages 3-5, kappel2023genomicandepigenomic pages 5-8) |
| Biological processes/phenotypes | HSFA1B participates in basal thermotolerance, acquired thermotolerance, and the early heat-stress response; together with HSFA1a/1d it is also implicated in responses to salt, osmotic, oxidative, and other stresses. Single hsfa1b loss often shows weak phenotype because of redundancy, whereas higher-order hsfa1 mutants display severe heat sensitivity and developmental defects; overexpression studies indicate HSFA1B can drive constitutive HSP accumulation and broaden stress tolerance. HSFA1s also contribute to thermomorphogenesis by enabling temperature-induced hypocotyl elongation. (albhilal2015thearabidopsisthaliana pages 38-42, liu2013commonanddistinct pages 1-2, liu2011theroleof pages 6-7, tan2023theheatresponse pages 1-2) |
| Localization | As expected for a transcription factor, HSFA1B acts in the nucleus after activation. Reviews state HSP70/HSP90 repression affects nuclear localization, and recent Arabidopsis work showed HSFA1 proteins accumulate and move into the nucleus under warm daytime temperature; direct imaging was shown for HSFA1d, which is generally interpreted as informative for HSFA1-family behavior, though not HSFA1B specifically. Nuclear interaction of HSFA1s with PIF4 further supports a nuclear site of action. (wang2023transcriptionalregulatorsof pages 2-4, tan2023theheatresponse pages 1-2, tan2023theheatresponse media 2354b667) |
| Recent (2023-2024) developments | 2023 reviews reaffirm HSFA1s, including HSFA1B, as the central activators/master regulators of plant heat-stress transcription. Kappel et al. 2023 linked upstream HSFA1 activity to memory-HSF induction and showed memory genes are enriched for tripartite HSEs, accessible chromatin, and heat-induced H3K4me3. Tan et al. 2023 connected HSFA1s to thermomorphogenesis via PIF4 stabilization, and Bakery et al. 2024 reframed HSFs as a dynamic molecular rheostat tuned by chaperones, PTMs, and attenuation circuits. (tan2023theheatresponse pages 1-2, kappel2023genomicandepigenomic pages 3-5, kappel2023genomicandepigenomic pages 1-3, bakery2024heatstresstranscription pages 4-4) |
| Quantitative data | Transcriptomics from HSFA1-loss mutants indicate >65% of heat-upregulated genes are HSFA1 dependent; specifically, one analysis found 230/346 (66.5%) and 278/408 (68.1%) heat-upregulated genes reduced >3-fold in the hsfa1 quadruple mutant in two ecotypes, with 209 genes commonly affected. Kappel et al. identified 15 ChIP-seq clusters, with 7 heat-induced clusters totaling 4,948 peaks for HSFA2/HSFA3; one memory-associated cluster (c12) contained 43 genes. Thermomemory-related in vitro binding assays compared HSFA2, HSFA3, and HSFA1b at 25°C vs 37°C. (liu2011theroleof pages 6-7, kappel2023genomicandepigenomic pages 3-5, kappel2023genomicandepigenomic pages 5-8) |
Table: This table summarizes the core functional annotation of Arabidopsis HSFA1B/At5g16820, including identity, molecular role, regulation, pathways, localization, recent 2023–2024 advances, and quantitative findings. It is useful as a compact evidence map linking HSFA1B to authoritative mechanistic and genome-wide studies.
8) Practical functional annotation summary (actionable)
Primary molecular function: HSFA1B is a sequence-specific DNA-binding transcription factor (class A1 HSF) that activates transcription at HSE-containing promoters, initiating early heat-stress transcription and broader temperature-responsive programs. (liu2013commonanddistinct pages 1-2, liu2011theroleof pages 1-2, wang2023transcriptionalregulatorsof pages 2-4)
Key biological processes: basal and acquired thermotolerance/HSR; integration of warm-temperature signals with growth (thermomorphogenesis); coupling of stress defense to development and reproductive fitness through extensive direct/indirect transcriptional regulation. (tan2023theheatresponse pages 1-2, albihlal2018arabidopsisheatshock pages 1-4, albihlal2018arabidopsisheatshock pages 7-10)
Cellular site of action: predominantly nuclear during activation; nuclear accumulation and nuclear partner interactions are core to HSFA1-family function (demonstrated for HSFA1s in warm temperature contexts). (wang2023transcriptionalregulatorsof pages 2-4, tan2023theheatresponse pages 1-2, tan2023theheatresponse media 2354b667)
Core pathway placement: upstream “master regulator” layer (HSFA1a/b/d/e) → induction of HSP genes and downstream TF cascade (HSFA2/3, DREB2A, MBF1C, HSFA7s) → proteostasis, antioxidant/redox balancing, and chromatin-associated memory programs (H3K4me3 enrichment at memory genes via HSFA2/3). (wang2023transcriptionalregulatorsof pages 2-4, kappel2023genomicandepigenomic pages 3-5, kappel2023genomicandepigenomic pages 1-3)
9) Limitations and evidence gaps
- Isoform-specific HSFA1B localization microscopy was not found in the extracted passages; recent imaging is strongest for HSFA1d within the HSFA1 family. (tan2023theheatresponse pages 1-2, tan2023theheatresponse media 2354b667)
- Several application claims regarding specific crops and HSFA1 homologs were discussed in reviews; for strict “real-world implementation,” confirmation at field scale would require the original crop trials, which were not directly retrieved here. (wang2023transcriptionalregulatorsof pages 9-11)
References
-
(wang2023transcriptionalregulatorsof pages 2-4): Xuejing Wang, Nicholas Wui Kiat Tan, Fong Yi Chung, Nobutoshi Yamaguchi, Eng-Seng Gan, and Toshiro Ito. Transcriptional regulators of plant adaptation to heat stress. International Journal of Molecular Sciences, 24:13297, Aug 2023. URL: https://doi.org/10.3390/ijms241713297, doi:10.3390/ijms241713297. This article has 44 citations.
-
(liu2011theroleof pages 1-2): HSIANG‐CHIN LIU, HSIU‐TING LIAO, and YEE‐YUNG CHARNG. The role of class a1 heat shock factors (hsfa1s) in response to heat and other stresses in arabidopsis. Plant, cell & environment, 34 5:738-51, May 2011. URL: https://doi.org/10.1111/j.1365-3040.2011.02278.x, doi:10.1111/j.1365-3040.2011.02278.x. This article has 667 citations.
-
(liu2011theroleof pages 6-7): HSIANG‐CHIN LIU, HSIU‐TING LIAO, and YEE‐YUNG CHARNG. The role of class a1 heat shock factors (hsfa1s) in response to heat and other stresses in arabidopsis. Plant, cell & environment, 34 5:738-51, May 2011. URL: https://doi.org/10.1111/j.1365-3040.2011.02278.x, doi:10.1111/j.1365-3040.2011.02278.x. This article has 667 citations.
-
(liu2013commonanddistinct pages 1-2): Hsiang-chin Liu and Yee-yung Charng. Common and distinct functions of arabidopsis class a1 and a2 heat shock factors in diverse abiotic stress responses and development1[w][open]. Plant Physiology, 163:276-290, Jul 2013. URL: https://doi.org/10.1104/pp.113.221168, doi:10.1104/pp.113.221168. This article has 263 citations and is from a highest quality peer-reviewed journal.
-
(bakery2024heatstresstranscription pages 4-4): Ayat Bakery, Stavros Vraggalas, Boushra Shalha, Harsh Chauhan, Moussa Benhamed, and Sotirios Fragkostefanakis. Heat stress transcription factors as the central molecular rheostat to optimize plant survival and recovery from heat stress. The New phytologist, 244:51-64, Jul 2024. URL: https://doi.org/10.1111/nph.20017, doi:10.1111/nph.20017. This article has 93 citations.
-
(albihlal2018arabidopsisheatshock pages 10-12): Waleed S Albihlal, Irabonosi Obomighie, Thomas Blein, Ramona Persad, Igor Chernukhin, Martin Crespi, Ulrike Bechtold, and Philip M Mullineaux. Arabidopsis heat shock transcription factora1b regulates multiple developmental genes under benign and stress conditions. Journal of Experimental Botany, 69:2847-2862, Apr 2018. URL: https://doi.org/10.1093/jxb/ery142, doi:10.1093/jxb/ery142. This article has 89 citations and is from a domain leading peer-reviewed journal.
-
(albihlal2018arabidopsisheatshock pages 1-4): Waleed S Albihlal, Irabonosi Obomighie, Thomas Blein, Ramona Persad, Igor Chernukhin, Martin Crespi, Ulrike Bechtold, and Philip M Mullineaux. Arabidopsis heat shock transcription factora1b regulates multiple developmental genes under benign and stress conditions. Journal of Experimental Botany, 69:2847-2862, Apr 2018. URL: https://doi.org/10.1093/jxb/ery142, doi:10.1093/jxb/ery142. This article has 89 citations and is from a domain leading peer-reviewed journal.
-
(bakery2024heatstresstranscription pages 3-3): Ayat Bakery, Stavros Vraggalas, Boushra Shalha, Harsh Chauhan, Moussa Benhamed, and Sotirios Fragkostefanakis. Heat stress transcription factors as the central molecular rheostat to optimize plant survival and recovery from heat stress. The New phytologist, 244:51-64, Jul 2024. URL: https://doi.org/10.1111/nph.20017, doi:10.1111/nph.20017. This article has 93 citations.
-
(albihlal2018arabidopsisheatshock pages 7-10): Waleed S Albihlal, Irabonosi Obomighie, Thomas Blein, Ramona Persad, Igor Chernukhin, Martin Crespi, Ulrike Bechtold, and Philip M Mullineaux. Arabidopsis heat shock transcription factora1b regulates multiple developmental genes under benign and stress conditions. Journal of Experimental Botany, 69:2847-2862, Apr 2018. URL: https://doi.org/10.1093/jxb/ery142, doi:10.1093/jxb/ery142. This article has 89 citations and is from a domain leading peer-reviewed journal.
-
(albihlal2018arabidopsisheatshock pages 12-15): Waleed S Albihlal, Irabonosi Obomighie, Thomas Blein, Ramona Persad, Igor Chernukhin, Martin Crespi, Ulrike Bechtold, and Philip M Mullineaux. Arabidopsis heat shock transcription factora1b regulates multiple developmental genes under benign and stress conditions. Journal of Experimental Botany, 69:2847-2862, Apr 2018. URL: https://doi.org/10.1093/jxb/ery142, doi:10.1093/jxb/ery142. This article has 89 citations and is from a domain leading peer-reviewed journal.
-
(kappel2023genomicandepigenomic pages 3-5): Christian Kappel, Thomas Friedrich, Vicky Oberkofler, Li Jiang, Tim Crawford, Michael Lenhard, and Isabel Bäurle. Genomic and epigenomic determinants of heat stress-induced transcriptional memory in arabidopsis. Genome Biology, May 2023. URL: https://doi.org/10.1186/s13059-023-02970-5, doi:10.1186/s13059-023-02970-5. This article has 54 citations and is from a highest quality peer-reviewed journal.
-
(li2025heatshocktranscription pages 6-7): Yuan Li, Kang Gong, Xinyi Wang, Zhihong Sun, and Fei Ding. Heat shock transcription factors as central integrators of plant stress responses: from thermotolerance to multi-stress resilience. Biology, 14:1800, Dec 2025. URL: https://doi.org/10.3390/biology14121800, doi:10.3390/biology14121800. This article has 1 citations.
-
(tan2023theheatresponse pages 1-2): Wenrong Tan, Junhua Chen, Xiaolan Yue, Shuli Chai, Wei Liu, Chenglin Li, Feng Yang, Yongfeng Gao, Lucas Gutiérrez Rodríguez, Víctor Resco de Dios, Dawei Zhang, and Yinan Yao. The heat response regulators hsfa1s promote arabidopsis thermomorphogenesis via stabilizing pif4 during the day. Science Advances, Nov 2023. URL: https://doi.org/10.1126/sciadv.adh1738, doi:10.1126/sciadv.adh1738. This article has 71 citations and is from a highest quality peer-reviewed journal.
-
(tan2023theheatresponse media 2354b667): Wenrong Tan, Junhua Chen, Xiaolan Yue, Shuli Chai, Wei Liu, Chenglin Li, Feng Yang, Yongfeng Gao, Lucas Gutiérrez Rodríguez, Víctor Resco de Dios, Dawei Zhang, and Yinan Yao. The heat response regulators hsfa1s promote arabidopsis thermomorphogenesis via stabilizing pif4 during the day. Science Advances, Nov 2023. URL: https://doi.org/10.1126/sciadv.adh1738, doi:10.1126/sciadv.adh1738. This article has 71 citations and is from a highest quality peer-reviewed journal.
-
(kappel2023genomicandepigenomic pages 1-3): Christian Kappel, Thomas Friedrich, Vicky Oberkofler, Li Jiang, Tim Crawford, Michael Lenhard, and Isabel Bäurle. Genomic and epigenomic determinants of heat stress-induced transcriptional memory in arabidopsis. Genome Biology, May 2023. URL: https://doi.org/10.1186/s13059-023-02970-5, doi:10.1186/s13059-023-02970-5. This article has 54 citations and is from a highest quality peer-reviewed journal.
-
(kappel2023genomicandepigenomic pages 5-8): Christian Kappel, Thomas Friedrich, Vicky Oberkofler, Li Jiang, Tim Crawford, Michael Lenhard, and Isabel Bäurle. Genomic and epigenomic determinants of heat stress-induced transcriptional memory in arabidopsis. Genome Biology, May 2023. URL: https://doi.org/10.1186/s13059-023-02970-5, doi:10.1186/s13059-023-02970-5. This article has 54 citations and is from a highest quality peer-reviewed journal.
-
(wang2023transcriptionalregulatorsof pages 9-11): Xuejing Wang, Nicholas Wui Kiat Tan, Fong Yi Chung, Nobutoshi Yamaguchi, Eng-Seng Gan, and Toshiro Ito. Transcriptional regulators of plant adaptation to heat stress. International Journal of Molecular Sciences, 24:13297, Aug 2023. URL: https://doi.org/10.3390/ijms241713297, doi:10.3390/ijms241713297. This article has 44 citations.
-
(guan2013molecularmechanismsof pages 58-64): Q Guan. Molecular mechanisms of plant responses to cold, heat and salt stresses in arabidopsis. Unknown journal, 2013.
-
(bakery2024heatstresstranscription pages 9-9): Ayat Bakery, Stavros Vraggalas, Boushra Shalha, Harsh Chauhan, Moussa Benhamed, and Sotirios Fragkostefanakis. Heat stress transcription factors as the central molecular rheostat to optimize plant survival and recovery from heat stress. The New phytologist, 244:51-64, Jul 2024. URL: https://doi.org/10.1111/nph.20017, doi:10.1111/nph.20017. This article has 93 citations.
-
(albhilal2015thearabidopsisthaliana pages 38-42): WS Albhilal. The arabidopsis thaliana heat shock transcription factor a1b transcriptional regulatory network. Unknown journal, 2015.
Artifacts
Citations
- bakery2024heatstresstranscription pages 4-4
- liu2011theroleof pages 1-2
- liu2011theroleof pages 6-7
- albihlal2018arabidopsisheatshock pages 7-10
- albihlal2018arabidopsisheatshock pages 12-15
- albihlal2018arabidopsisheatshock pages 1-4
- kappel2023genomicandepigenomic pages 3-5
- wang2023transcriptionalregulatorsof pages 2-4
- tan2023theheatresponse pages 1-2
- kappel2023genomicandepigenomic pages 5-8
- wang2023transcriptionalregulatorsof pages 9-11
- liu2013commonanddistinct pages 1-2
- albihlal2018arabidopsisheatshock pages 10-12
- bakery2024heatstresstranscription pages 3-3
- li2025heatshocktranscription pages 6-7
- kappel2023genomicandepigenomic pages 1-3
- guan2013molecularmechanismsof pages 58-64
- bakery2024heatstresstranscription pages 9-9
- albhilal2015thearabidopsisthaliana pages 38-42
- w
- open
- https://doi.org/10.1093/jxb/ery142
- https://doi.org/10.1126/sciadv.adh1738
- https://doi.org/10.1186/s13059-023-02970-5
- https://doi.org/10.3390/ijms241713297
- https://doi.org/10.3390/ijms241713297,
- https://doi.org/10.1111/j.1365-3040.2011.02278.x,
- https://doi.org/10.1104/pp.113.221168,
- https://doi.org/10.1111/nph.20017,
- https://doi.org/10.1093/jxb/ery142,
- https://doi.org/10.1186/s13059-023-02970-5,
- https://doi.org/10.3390/biology14121800,
- https://doi.org/10.1126/sciadv.adh1738,
Perplexity
(AT5G16820-deep-research-perplexity.md)
Perplexity
(AT5G16820-deep-research-perplexity.md)Comprehensive Functional Analysis of HSFA1B (AT5G16820): A Master Regulator of Environmental Stress Response and Plant Development
Overview
HSFA1B (Heat Shock Transcription Factor A1b), encoded by the gene AT5G16820 in Arabidopsis thaliana, represents a critical transcriptional hub that integrates environmental stress signals with developmental programs to enable coordinated plant responses to fluctuating environmental conditions[1][2]. This gene product functions as a master regulator of the heat stress response through its capacity to directly bind DNA at heat shock elements and to orchestrate a complex transcriptional cascade involving dozens of downstream transcription factors[1][2][14]. Beyond its classical role in heat stress tolerance, HSFA1B coordinates plant growth and reproduction, influencing seed yield and developmental architecture through reconfiguration of a vast regulatory network encompassing hundreds of genes with diverse cellular functions[1][19]. The protein localizes constitutively to both cytoplasm and nucleus under normal conditions, translocating more prominently to the nucleus upon heat stress or warm temperature exposure, a process mediated by interactions with molecular chaperones and light-dependent signaling pathways[11][24][28].
Molecular Identity and Structural Organization
HSFA1B belongs to the class A1 heat shock transcription factor family, a group of four highly homologous genes in Arabidopsis that includes HSFA1A, HSFA1B, HSFA1D, and HSFA1E[1][28][55]. These four members share substantial sequence identity and exhibit considerable functional redundancy, though each possesses distinct regulatory properties and stress response preferences[24][28][55]. The protein adopts a modular architecture characteristic of all heat shock transcription factors, consisting of several functionally distinct domains arranged from N-terminus to C-terminus[20][23][32]. The DNA-binding domain (DBD) at the N-terminus contains the helix-turn-helix motif responsible for recognizing and binding heat shock elements in target gene promoters[20][23]. Adjacent to the DBD lies the oligomerization domain (also termed HR-A/B domain), composed of hydrophobic heptad repeats that form a three-stranded coiled-coil structure enabling trimerization of HSF monomers—a critical feature that substantially increases DNA-binding affinity through cooperative interactions[20][23]. Following the trimerization domain is the temperature-dependent repression (TDR) domain, a regulatory region containing the central portion of the protein that functions to suppress transactivation activity under normal, non-stress conditions through direct interaction with heat shock proteins HSP70 and HSP90[3][8][32]. Finally, the C-terminal activation domain (CTAD) contains acidic activation function (AHA) motifs with the characteristic pattern of aromatic and hydrophobic amino acid residues embedded in an acidic context, elements essential for transactivation of target genes through recruitment of the basal transcription machinery[51][54]. Unlike many class A HSFs in other organisms, HSFA1B possesses a nuclear export signal (NES) in its regulatory domain that functions to maintain cytoplasmic localization under normal conditions[11][32].
The specific sequence motif recognized by HSFA1B in target promoters is the heat shock element (HSE), classically defined as the trinucleotide repeat pattern (nGAAn)3 arranged as inverted repeats[(TTCnn)GAAnnTTC][7][33]. However, research has identified that HSFA1B specifically recognizes a novel HSE variant designated HSE1b with the consensus sequence 5'-AGAAnnTTCT-3', a non-canonical motif present in approximately 55 genes preferentially regulated by HSFA1B[7][26][52]. This sequence specificity distinguishes HSFA1B-regulated genes from those recognized by other class A1 HSFs, providing a molecular basis for functional specialization within the HSF family[7][52].
Primary Function: Master Transcriptional Regulator of the Heat Stress Response
HSFA1B functions fundamentally as a DNA-binding transcription factor that transactivates heat shock-responsive genes in response to elevated temperatures and other environmental stresses[1][19]. The protein directly activates expression of genes encoding heat shock proteins (HSPs)—molecular chaperones essential for protein protection, refolding, and degradation during stress conditions[8][16]. HSP100, HSP90, HSP70, and small HSPs (sHSPs) all contain HSE elements in their promoters that are recognized and bound by HSFA1B and other class A1 HSFs[1][16]. When activated by heat stress, HSFA1B undergoes conformational changes that expose its transactivation domain, enabling recruitment of the transcriptional machinery to these promoters[8][32]. The coordinate induction of multiple HSP genes creates a proteostatic network capable of managing the influx of damaged proteins characteristic of heat stress[3][8].
Beyond direct activation of HSPs, HSFA1B functions as the apex of a transcriptional cascade that amplifies and diversifies the heat stress response through regulation of secondary transcription factors[1][8][14]. Genome-wide chromatin immunoprecipitation studies combined with transcriptomic analysis identified a total of 952 directly targeted genes of which at least 85 are development-associated and were predominantly bound under non-stress conditions[1][19]. Among HSFA1B's direct targets are transcription factor genes including HSFA2, HSFA7A, HSFB2A, HSFB2B, DREB2A, and others[1][26][52]. These secondary transcription factors subsequently regulate their own target genes, creating a hierarchical network of at least 27 transcription factors through which HSFA1B exerts indirect influence over expression of approximately 1780 additional genes[1][19]. This regulatory architecture ensures that heat stress response genes spanning metabolic, developmental, and protective functions are coordinately induced in an appropriate temporal sequence[1][8].
The transactivation capacity of HSFA1B appears to be modulated by specific protein sequences in its activation domain. Mutagenesis studies of related HSFs revealed that the AHA motifs within HSFA1B's C-terminal region are essential for activation potential, with aromatic and hydrophobic amino acid residues in core positions proving critical for function[51][54]. The amphipathic, negatively charged helix formed by these residues represents the probable contact surface with components of the basal transcription apparatus[51][54]. Disruption of these motifs or introduction of proline residues adjacent to them markedly reduces or abolishes transactivation capacity[51][54].
DNA-Binding Properties and Target Gene Recognition
The DNA-binding mechanism of HSFA1B operates through a trimeric protein-DNA complex architecture in which three HSF monomers bind cooperatively to a three-site HSE[20][23]. Crystal structures of related HSF family members reveal that each monomer's DBD contacts one nGAAn triplet repeat within the HSE, with the three DBDs arranged in a triangular configuration relative to the DNA axis[20][23]. This trimerization is essential for high-affinity binding; while individual DBD monomers show weak binding to single triplet repeats, the cooperative interaction of three DBDs substantially amplifies binding affinity at intact three-site HSEs[20][23]. The conserved arginine residue near the C-terminus of each DBD inserts directly into the major groove of DNA and forms hydrogen bonds with nucleobases, providing sequence-specific recognition[20][23].
Comparison of structural data between HSF1 and HSF2 suggests subtle but significant differences in DNA-binding geometry[20]. HSF1 exhibits more extended DNA binding, occupying a longer segment of DNA than HSF2, a difference attributable to differential orientation of the DBD monomers along the DNA helix[20]. These structural differences may explain divergent target selectivity and transcriptional properties among HSF family members, though HSFA1B-specific structural studies remain limited[20][23].
The identification of the non-canonical HSE1b element represents a major advance in understanding HSFA1B target specificity[7][26][52]. Using both bioinformatic motif discovery and chromatin immunoprecipitation-quantitative PCR validation, researchers demonstrated that HSFA1B specifically recognizes the HSE1b sequence in approximately 55 promoters[7][52]. Chromatin immunoprecipitation experiments revealed that HSFA1B binds in vivo to promoters containing single HSE1b elements in isolation from other HSE-like motifs, demonstrating specificity even when overexpressed[7][52]. This specificity was confirmed through analysis of control promoters containing only canonical core HSE sequences, which failed to recruit HSFA1B despite containing the triplet repeats that define canonical HSEs[7][52]. The HSE1b-containing genes predominantly encode transcription factors involved in stress defense and development, suggesting that HSFA1B has evolved preferential recognition of this non-canonical motif as a means of selectively activating the transcriptional cascade components most relevant to its regulatory function[7][52].
Notably, HSFA1B also targets 480 natural antisense non-coding RNA (cisNAT) genes, defining an additional mode of indirect gene regulation through RNA-based mechanisms[1][19]. Many of these cisNAT genes are bound by HSFA1b under non-stress conditions, suggesting a previously unappreciated role for natural antisense RNAs in HSFA1B-mediated transcriptional networks[1][19]. The precise mechanistic role of these cisNAT targets remains to be fully characterized, but their involvement suggests layers of post-transcriptional regulation coordinating with HSFA1B's transcriptional function.
Cellular Localization and Nuclear Dynamics
Under non-stress conditions at normal temperature, HSFA1B exhibits dual subcellular localization, present in both the cytoplasm and nucleus, with a preference for cytoplasmic accumulation[11][24][25][28][32]. This cytoplasmic retention is mediated by direct interaction of the TDR domain with HSP70 and HSP90 molecular chaperones[3][8][32]. The interaction with these chaperones functions as a regulatory mechanism that suppresses both the DNA-binding activity and transactivation potential of HSFA1B under normal conditions, rendering the protein transcriptionally inert despite its presence in the nucleus[3][12][32]. The nuclear export signal (NES) within the regulatory domain actively promotes cytoplasmic retention, establishing a baseline state in which HSFA1B remains sequestered away from target promoters[11][32].
Upon exposure to heat stress, HSFA1B undergoes rapid nuclear translocation, a process coordinated with dissociation from HSP70 and HSP90[3][8][24][28]. The direct consequence of this nuclear accumulation is marked increase in availability of HSFA1B monomers for trimerization and DNA binding. Temperature sensing appears to occur through HSP70-mediated titration; when cell protein damage increases during heat stress, the accumulation of unfolded proteins competes with HSFA1B for binding to HSP70, effectively releasing HSFA1B from repression[8][32]. The precise molecular trigger for HSP70-HSP90 dissociation remains incompletely understood, though phosphorylation and other post-translational modifications likely play roles[18][31].
Intriguingly, recent evidence reveals that warm temperature (approximately 28°C) triggers nuclear translocation of HSFA1B in a light-dependent manner, distinct from the acute heat stress response[11][24][28]. This warm-temperature signaling pathway involves the COP1-BIN2 module, where COP1 (constitutive photomorphogenic 1) functions as a light-dependent regulator of HSFA1B nuclear localization[11][24]. Under daytime warm temperatures, COP1 promotes nuclear import of HSFA1B by inhibiting BIN2 kinase activity, thereby preventing BIN2-mediated phosphorylation of HSFA1B's nuclear localization signal that would otherwise trap the protein in the cytoplasm[11][24][28]. This light-temperature integration allows plants to distinguish between developmentally beneficial warm temperatures during the day and potentially harmful heat stress at any time, enabling adaptive thermomorphogenic responses only during appropriate developmental conditions[11][24][28].
The thermomorphogenic nuclear localization of HSFA1B under warm daytime conditions appears constitutive and independent of the HSP70-dependent derepression mechanism operative during acute heat stress[11][24][28]. This distinction reflects two separate temperature-sensing pathways converging on HSFA1B: an acute stress-response pathway mediated by HSP70-dependent release, and a developmental warm-temperature pathway mediated by light-dependent COP1 activity[11][24][28]. Both pathways culminate in nuclear accumulation of HSFA1B, but they function in distinct temporal and developmental contexts[11][24][28].
Regulation by Molecular Chaperones: HSP70 and HSP90 Interactions
The molecular chaperones HSP70 and HSP90 play critical roles in suppressing HSFA1B activity under normal conditions, functioning as repressor proteins that maintain the protein in a transcriptionally inactive state[3][8][9][12][32]. The interaction of HSFA1B with these chaperones involves direct protein-protein contact mediated by the TDR domain, which serves as the primary binding surface for both HSP70 and HSP90[3][8][32]. These chaperone interactions suppress HSFA1B through at least two distinct mechanisms: first, they inhibit DNA-binding activity, preventing the formation of productive complexes at target promoters; second, they restrict nuclear localization by actively promoting cytoplasmic retention[3][8][12][32].
HSP70 specifically represses DNA-binding activity of HSFA1B and related class A1 HSFs[12]. Co-immunoprecipitation and biochemical studies in model plant systems revealed that HSP70 associates with the TDR domain and interferes with the conformational changes necessary for promoter recognition[12]. Strikingly, this repression occurs even for HSF proteins that successfully reach the nucleus, suggesting that HSP70-mediated inhibition operates at the level of DNA-binding rather than merely nuclear access[12][32]. Upon heat stress, HSP70 is titrated away from HSFA1B through sequestration by accumulating unfolded proteins, liberating HSFA1B to adopt its active conformational state[8][12][32].
HSP90 functions primarily to regulate nuclear localization and protein stability of class A HSFs including HSFA1B[3][8][9]. HSP90 interacts with HSFA1B through the TDR domain and promotes cytoplasmic retention through a mechanism distinct from HSP70's DNA-binding inhibition[3][9]. Disruption of HSP90 function, either through genetic mutation or pharmacological inhibition, results in constitutive nuclear accumulation and activation of HSFA1B even under non-stress conditions[3][9]. This suggests HSP90 actively shuttles HSFA1B back to the cytoplasm or prevents nuclear import through as-yet-incompletely-characterized mechanisms[3][9]. HSP90 also regulates stability of certain downstream HSFs such as HSFA2, influencing the kinetics of transcriptional cascade activation during stress recovery[12][31].
The ratio between HSP70 and HSP90 levels appears to influence the degree of HSFA1B activation, suggesting a more nuanced regulatory mechanism than simple binary repression[9][12]. During heat stress, rapid accumulation of unfolded proteins disrupts the HSP70-HSFA1B complex but may preserve HSP90 interactions, creating an intermediate activation state[9][12]. As stress is resolved and protein folding recovers, HSP70 gradually rebinds HSFA1B, progressively reducing its activity[9][12]. This model provides a molecular basis for the graded response of HSFA1B to heat stress intensity and duration, allowing plants to calibrate protection mechanisms to stress severity[9][12].
Transcriptional Cascade and Downstream Gene Regulation
HSFA1B orchestrates a transcriptional cascade in which it directly activates approximately 952 genes, of which at least 85 are development-associated[1][19]. Among these direct targets are critical secondary transcription factors including HSFA2, HSFA7A, HSFB2A, HSFB2B, DREB2A, and MBF1C[1][26][52]. These downstream transcription factors themselves regulate additional target genes, extending HSFA1B's transcriptional reach through a hierarchical network to approximately 1,780 indirectly regulated genes[1][19]. This cascade architecture provides multiple layers of control, enabling both amplification of the stress response through transcriptional feedback and temporal refinement of gene expression patterns through differential regulation by intermediate transcription factors[1][8][19].
The secondary transcription factor HSFA2 represents a particularly important HSFA1B target, as it becomes the dominant HSF during the later phases of heat stress when HSFA1B levels decline[43][55][56]. HSFA2 is not heat-inducible in the absence of HSFA1B, indicating that HSFA1B-mediated HSFA2 induction represents an essential step in establishing sustained heat stress response[8][43]. The transition from HSFA1B-dominated to HSFA2-dominated regulation enables sustained protection after peak stress conditions pass, as HSFA2 induces expression of heat shock proteins and additional protective genes through both HSF-independent and HSF-dependent mechanisms[8][43][55].
The DREB2A transcription factor, another major HSFA1B target, coordinates responses to drought and other water-limiting stresses, establishing functional connections between heat and drought stress pathways[1][27][30]. Heat-induced expression of DREB2A is mediated through HSFA1B binding to HSEs in the DREB2A promoter, linking acute heat stress responses with longer-term adaptive drought tolerance mechanisms[1][27]. This interconnection between HSFA1B-mediated stress pathways and DREB-controlled responses suggests that many plant stress adaptations are coordinated through shared transcriptional regulators.
The MBF1C (Multiprotein Bridging Factor 1c) transcription factor, directly regulated by HSFA1B, controls resistance to both bacterial (Pseudomonas syringae) and oomycete (Hyaloperonospora parasitica) pathogens[26][44]. Plants overexpressing HSFA1B show enhanced basal resistance to both pathogen types, with this resistance partially dependent on MBF1C-mediated pathways[26][44]. This demonstrates that HSFA1B's regulatory network extends beyond canonical heat stress responses into immune signaling, providing a molecular basis for cross-talk between heat and pathogen defense pathways[26][44][52].
Developmental Functions and Environmental Adaptation
Beyond its role as a stress-response regulator, HSFA1B fundamentally integrates stress signals with plant developmental programs, enabling coordinated growth adjustments in response to environmental fluctuations[1][14][19][24][25][28]. Overexpression of HSFA1B in Arabidopsis results in altered developmental architecture including reduced rosette expansion, earlier flowering, and increased allocation of resources to reproductive structures at the expense of vegetative growth, collectively resulting in increased seed yield[1][2][26][38][47]. These developmental effects are not secondary consequences of continuous stress activation but rather reflect HSFA1B's direct regulation of developmental gene networks even under non-stress conditions[1][19].
Genome-wide binding analysis revealed that HSFA1B occupies promoters of approximately 354 genes involved in plant growth and development under non-stress conditions[1][47]. These developmentally-targeted genes encode proteins with remarkably diverse functions including cell integrity-associated chaperones, components of chloroplast development machinery, hormonal signaling molecules (particularly auxins and brassinosteroids), photoreceptors, components of photomorphogenesis signaling, cell wall synthesis enzymes, and transcription factors with established developmental roles[1][47]. The diversity of these targets precludes identification of a single developmental pathway through which HSFA1B operates; instead, HSFA1B appears to coordinately modulate many genes of diverse cellular functions[1][47]. The net result of these subtle, distributed effects is the substantial developmental phenotype of HSFA1B-overexpressing plants, suggesting that development results from integration of many such molecular regulatory events[1][47].
Notably, HSFA1B-mediated developmental changes include redistribution of biomass in favor of reproductive structures[1][2][47]. This phenotype mirrors the developmental priority shift that occurs during heat or other stress conditions, when plants suspend vegetative growth and accelerate reproduction—a strategy that ensures offspring survival even if adult plants subsequently fail[1][2][47]. HSFA1B appears to function as a molecular switch mediating this growth-defense tradeoff, with elevated HSFA1B activity biasing plants toward reproductive investment[1][2][47]. This integration of stress-response and developmental functions may reflect an evolutionary solution to the conflict between immediate growth maximization and long-term reproductive success under variable environmental conditions[1][2][47].
Recent studies have revealed that HSFA1B participates in transgenerational inheritance of stress responses, with prolonged heat stress leading to heritable downregulation of HSFA1B and downstream effects on flowering time and immune status in unstressed progeny[22]. The HSFA1B-HSP70-3 module modulates transgenerational thermomemory, wherein heat-induced downregulation of HSFA1B reduces HSP70-3 expression, leading to decreased stability of SGS3 protein and reduced trans-acting siRNA biogenesis[22]. These heritable changes cause early flowering and attenuated immunity in the next generation, even in the absence of heat stress, suggesting that HSFA1B participates in stress-induced epigenetic regulation affecting multiple traits[22].
Post-Translational Modifications and Activity Regulation
HSFA1B activity is extensively regulated through post-translational modifications, including phosphorylation, sumoylation, and ubiquitination[18][31][34]. These modifications alter HSFA1B's subcellular localization, DNA-binding activity, and protein stability, providing additional layers of control beyond the HSP70-HSP90 repression mechanism[18][31].
Phosphorylation represents a major regulatory modification, with several kinases targeting HSFA1B at distinct sites[18][31]. Mitogen-activated protein kinase (MAPK) pathways, particularly through the kinases MPK3 and MPK6, phosphorylate class A HSFs including HSFA1B at multiple serine and threonine residues[18][31]. Heat-activated MAPKs appear to phosphorylate HSFA1B upon heat stress exposure, contributing to its activation and altered subcellular localization[18][31]. Conversely, BIN2 (brassinosteroid-insensitive 2), a GSK3-like kinase in the brassinosteroid signaling pathway, phosphorylates HSFA1B at residues within its nuclear localization signal, promoting cytoplasmic retention even when the protein might otherwise accumulate in the nucleus[11][24]. Under warm daytime temperatures, COP1 inhibits BIN2 activity, thereby preventing BIN2-mediated phosphorylation and allowing HSFA1B nuclear accumulation[11][24][28]. This reveals a sophisticated regulatory logic in which light-dependent signaling through COP1 overrides BIN2-mediated retention specifically during daytime warmth[11][24][28].
Sumoylation modifies HSFA1B through covalent attachment of SUMO peptides at lysine residues, a modification that generally represses transcriptional activity[31][34]. HSFA2, a close relative of HSFA1B, undergoes SUMO modification that suppresses its transactivation function, and similar mechanisms likely apply to HSFA1B[31]. Removal of SUMO modifications through the action of sentrin-specific proteases (Ulp1-like proteases) would reactivate the protein, providing a reversible switch governing HSF activity[31][34].
Ubiquitination targets HSF proteins for proteasomal degradation, particularly after prolonged stress when heat stress response gene expression must be attenuated[31][34]. HSP90 interaction with certain HSFs (particularly HSFB members) creates sites recognized by ubiquitin ligases, leading to polyubiquitination and degradation[9][12]. This mechanism ensures that after stress resolution, HSF protein levels decline through active proteolysis, enabling recovery of basal gene expression patterns[9][12][31].
The combined effects of these post-translational modifications create a sophisticated regulatory system in which HSFA1B activity is controlled through multiple independent mechanisms. Phosphorylation alters both localization and transcriptional capacity, sumoylation and ubiquitination provide reversible and irreversible activity switches, and chaperone interactions establish baseline repression[18][31][34]. This multi-layered regulation enables rapid, context-dependent responses to diverse environmental signals while preventing inappropriate activation under normal conditions[18][31][34].
Integration with Growth and Stress Signaling Pathways
HSFA1B functions as an integration node for multiple environmental and developmental signaling pathways, coordinating responses to temperature, light, pathogens, and hormones[11][24][25][28][53]. The protein interacts with several key developmental regulators including PIF4 (phytochrome-interacting factor 4), a master regulator of light-dependent hypocotyl growth[11][24][25][28]. Under warm daytime temperatures, HSFA1B directly interacts with PIF4 protein and stabilizes it by interfering with the interaction between PIF4 and photoactivated phytochrome B (phyB)[11][24][25][28]. This stabilization enhances PIF4 accumulation and nuclear activity, leading to increased expression of PIF4 target genes involved in cell elongation and auxin signaling[11][24][25][28]. The HSFA1-PIF4 module thus represents an integration point where light and temperature signals converge to regulate thermomorphogenic hypocotyl growth specifically during daytime warm conditions when such growth is developmentally appropriate[11][24][25][28].
HSFA1B-regulated transcriptional networks incorporate responses to brassinosteroid hormones through interactions with BIN2 and BES1, central components of BR signaling[11][24]. The BIN2-mediated phosphorylation of HSFA1B discussed above represents a point of crosstalk between heat stress and growth hormone signaling[11][24]. Under warm temperatures with active BR signaling, BIN2 inhibition allows HSFA1B nuclear accumulation, whereas under conditions with reduced BR signaling, increased BIN2 activity retains HSFA1B in the cytoplasm[11][24]. This integration ensures that thermomorphogenic growth occurs only when both temperature and hormone signals indicate appropriate conditions for increased growth[11][24].
HSFA1B also coordinates with salicylic acid (SA) signaling in immune responses[24]. HSFA1B interacts with NPR1, the central regulator of SA-dependent immunity, and together they induce expression of both heat shock proteins and immune-related genes[24]. This interaction provides a molecular basis for enhanced disease resistance observed in HSFA1B-overexpressing plants[24][26][44]. The coordination of stress defense with heat stress response genes ensures that plants simultaneously activate protective mechanisms appropriate to multiple simultaneous stresses[24][26][44].
The jasmonate pathway, particularly through the jasmonoyl-isoleucine-responsive gene expression system, has also been identified as downstream of HSFA1B[5][27][30]. HSFA1B activates expression of OPR3 (12-oxophytodienoate reductase 3), a key enzyme in jasmonate biosynthesis[5][27]. Increased jasmonate production in HSFA1B-overexpressing plants enhances thermotolerance through activation of the ICE1-CBF-COR regulon, a canonical cold acclimation pathway[5][27][30]. This demonstrates unexpected functional connections between HSFA1B-mediated thermotolerance and cold stress adaptation, likely reflecting shared protective mechanisms involving jasmonate-dependent gene expression[5][27][30].
Functional Specificity Within the Class A1 HSF Family
While HSFA1B shares substantial sequence identity and functional redundancy with its class A1 homologs HSFA1A, HSFA1D, and HSFA1E, evidence indicates that each member possesses distinct regulatory properties and stress response preferences[24][28][55][56]. Studies of quadruple knockout mutants lacking all four HSFA1 members have shown dramatic defects in heat stress response and acquired thermotolerance, demonstrating that the family collectively serves essential functions[24][28][55][56]. However, analysis of individual and triple knockout combinations reveals that HSFA1A, HSFA1B, and HSFA1D can individually confer significant thermotolerance, whereas HSFA1E appears less critical for this function[24][28][55][56]. The three functionally critical members appear to have undergone subfunctionalization after whole-genome duplication, with each acquiring specialized roles in distinct stress contexts or developmental stages[55][56].
HSFA1B appears particularly important for integration of stress responses with developmental regulation and immune signaling, based on its preferential regulation of developmental genes under non-stress conditions and its robust effects on seed yield[1][2][26][47]. In contrast, HSFA1D shows prominent roles in light-temperature integration and thermomorphogenic growth, interacting particularly strongly with PIF4[24][25][28]. HSFA1A appears to contribute broadly to thermotolerance with effects comparable to HSFA1B[24][28][56]. These distinctions, though subtle, suggest that the four HSFA1 members have diverged to specialize in different environmental or developmental contexts while maintaining sufficient overlap to provide functional redundancy[24][28][55][56].
Experimental Evidence and Functional Validation
The functions attributed to HSFA1B rest on multiple complementary experimental approaches providing converging evidence[1][2][8][14][19][26][47]. Chromatin immunoprecipitation combined with next-generation sequencing (ChIP-seq) has enabled genome-wide mapping of HSFA1B binding sites under both stressed and non-stressed conditions, revealing approximately 952 directly targeted genes[1][19]. These studies employed both FLAG-tagged and RFP-tagged HSFA1B fusion proteins with validated functionality, ensuring that the fusion proteins retained normal biological properties[1][19][26]. ChIP-seq results were further validated through targeted ChIP-PCR confirmation of binding to specific promoters including those of HSP17.6-CI, HSFA2, and other heat stress response genes[1][7][19][26].
RNA sequencing (RNA-seq) of wild-type and HSFA1B-overexpressing plants under stress and non-stress conditions revealed coordinated changes in gene expression that correlate with HSFA1B binding[1][19]. Comparison of ChIP-seq and RNA-seq data enabled distinction of direct targets (bound and regulated) from indirect targets (regulated but not bound)[1][19]. The identification of 27 directly regulated transcription factors whose products subsequently regulate additional downstream genes provides experimental validation of the hierarchical transcriptional cascade model[1][19].
Electrophoretic mobility shift assays (EMSAs) have confirmed the DNA-binding specificity of HSFA1B for both canonical HSEs and the novel HSE1b variant[7][58]. These in vitro binding studies using purified HSFA1B protein and synthetic DNA probes demonstrated that wild-type HSFA1B binds HSE1b sequences, while mutations within the HSE1b motif eliminate binding[7][58]. Competitive binding experiments in which unlabeled probes compete for HSFA1B binding further confirmed sequence specificity[7][58].
Yeast two-hybrid assays and co-immunoprecipitation studies have characterized protein-protein interactions involving HSFA1B, including its interactions with HSP70, HSP90, PIF4, and other regulatory proteins[9][11][12][24][25][49]. Bimolecular fluorescence complementation (BiFC) assays in plant cells provided visual confirmation of these interactions in living cells[11][24][25][49].
Deletion mutant analysis of HSFA1B and related HSFs revealed that the TDR domain is critical for chaperone interactions and activity suppression under normal conditions, while the C-terminal AHA motifs are essential for transactivation capacity[3][8][32][51]. Transgenic plants expressing various deletion constructs displayed activity levels correlating with domain composition, providing functional validation of structural predictions[3][8][32].
Thermotolerance assays directly measured the capacity of wild-type, HSFA1B-overexpressing, and hsfa1b knockout plants to survive heat stress, demonstrating that HSFA1B enhances acquired and basal thermotolerance[1][2][26][47]. Progressive heat treatment followed by recovery at normal temperature and subsequent challenge with lethal heat conditions revealed that HSFA1B-overexpressing plants achieve thermotolerance at lower acclimation temperatures than wild-type plants[1][26][47]. Conversely, hsfa1b knockout plants (when available in the background of other HSFA1 mutations) show reduced thermotolerance compared to wild-type[1][26][47].
Pathogen infection studies demonstrated enhanced resistance of HSFA1B-overexpressing plants to both Pseudomonas syringae and Hyaloperonospora parasitica, with reduced bacterial and oomycete growth in HSFA1b-overexpressing plants compared to wild-type controls[26][44][52]. Reciprocally, hsfa1a/hsfa1b double mutants showed enhanced susceptibility to these pathogens[26][44][52]. This functional validation directly supports the conclusion that HSFA1B-mediated transcriptional changes enhance pathogen resistance[26][44][52].
Comparison with Other Model Systems and Evolutionary Conservation
Studies in diverse plant species including wheat, tomato, and cucumber have confirmed that HSFA1B orthologs serve conserved roles in heat stress response and stress-induced acclimation[5][27][30][40]. In wheat, overexpression of the TaHSFA1b ortholog enhances thermotolerance and activates the OPR3-jasmonate pathway, mirroring HSFA1B's function in Arabidopsis[5][27]. In cucumber, the CsHSFA1d ortholog mediates heat-induced cold acclimation through jasmonate biosynthesis[30]. In tomato, HSFA1 orthologs regulate thermotolerance and interact with HSP70 and HSP90 in analogous fashion to Arabidopsis HSFA1B[12][21]. These conservation patterns across angiosperms support the conclusion that HSFA1B-like factors represent ancient, highly conserved regulators of stress response networks[5][27][30][40].
Conclusion
HSFA1B (AT5G16820) encodes a master transcriptional regulator that orchestrates plant responses to thermal stress while simultaneously coordinating developmental processes essential for reproductive success under variable environmental conditions. The protein functions through direct binding to heat shock element sequences—particularly the non-canonical HSE1b variant—in the promoters of approximately 952 target genes, establishing a hierarchical transcriptional cascade through regulation of secondary transcription factors. Under normal conditions, HSFA1B is maintained in a transcriptionally repressed state through interaction with HSP70 and HSP90 molecular chaperones that suppress both its DNA-binding activity and nuclear accumulation. Upon heat stress, titration of HSP70 by accumulating unfolded proteins liberates HSFA1B, enabling its activation and nuclear translocation. Independently, warm daytime temperatures trigger COP1-dependent, light-gated nuclear import of HSFA1B through inhibition of BIN2 kinase, enabling the protein to stabilize PIF4 and promote thermomorphogenic growth specifically during developmentally appropriate conditions.
The transcriptional networks controlled by HSFA1B encompass not only the canonical heat shock proteins essential for proteostasis but also extensive developmental gene networks, connecting stress tolerance with growth investment strategies. HSFA1B regulates approximately 354 genes involved in development even under non-stress conditions, explaining the dramatic effects of HSFA1B overexpression on plant architecture, flowering time, and seed yield. Post-translational modifications including phosphorylation, sumoylation, and ubiquitination provide additional regulatory layers enabling context-dependent activity control in response to multiple environmental signals. Integration with signaling pathways controlling light perception, hormone responses, immune function, and water availability positioning HSFA1B as a central node coordinating plant acclimation to multifactorial environmental stress. The high degree of functional conservation of HSFA1B orthologs across flowering plants underscores the fundamental importance of this regulatory mechanism for plant survival and reproductive success in thermally variable environments.
Citations
- https://pubmed.ncbi.nlm.nih.gov/29697803/
- https://academic.oup.com/jxb/article/69/11/2847/4985136
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4746676/
- https://www.uniprot.org/uniprotkb/A0A178UCE2
- https://onlinelibrary.wiley.com/doi/10.1111/pbi.13268
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10819275/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3733161/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3077788/
- https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.20017
- https://www.science.org/doi/10.1126/sciadv.adh1738
- https://pubmed.ncbi.nlm.nih.gov/21307284/
- https://academic.oup.com/nar/article/50/11/6102/6605316
- https://pubmed.ncbi.nlm.nih.gov/22494029/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6888953/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9590526/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5961379/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8379338/
- https://academic.oup.com/plphys/article/199/2/kiaf456/8266612
- https://pubmed.ncbi.nlm.nih.gov/26727489/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10624354/
- https://pubmed.ncbi.nlm.nih.gov/31559685/
- https://pubmed.ncbi.nlm.nih.gov/37922351/
- https://onlinelibrary.wiley.com/doi/pdf/10.1111/tpj.15780
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4746267/
- https://tgil.donga.ac.kr/gene/?locus=AT5G16820
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5627039/
- https://www.uniprot.org/uniprotkb/O81821
- https://repository.essex.ac.uk/24143/1/The%20Role%20of%20HSFs%20in%20Growth%20and%20Development%20%20Final.pdf
- https://pubmed.ncbi.nlm.nih.gov/27846364/
- https://www.nature.com/articles/s41598-021-01596-6
- https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2023.1271644/epub
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3762648/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2882041/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7036190/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC139763/
- https://academic.oup.com/pcp/article/60/1/230/5133769
- https://pubmed.ncbi.nlm.nih.gov/10662862/
- https://academic.oup.com/plphys/article/163/1/276/6110937
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3419016/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5543961/
- https://pubmed.ncbi.nlm.nih.gov/22516818/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3578905/