Deep Research
Falcon
(AT5G52640-deep-research-falcon.md)
Falcon
(AT5G52640-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: Arabidopsis thaliana HSP90-1 (AT5G52640; UniProt P27323) functional annotation
Executive summary
Arabidopsis HSP90-1 (AtHSP90.1; AT5G52640; UniProt P27323) encodes a cytosolic Hsp90-family molecular chaperone that uses an ATP-driven conformational cycle and co-chaperones to stabilize and mature specific client proteins in multiple pathways. The best-supported primary functions are (i) innate immunity, where AtHSP90.1 forms functionally critical complexes with cochaperones RAR1 and SGT1 to support NLR-mediated effector-triggered immunity; and (ii) proteostasis and developmental plasticity, where recent work implicates cytosolic HSP90 (including HSP90.1) in heat acclimation networks and in stabilizing plasma-membrane ABCB auxin transporters through interaction with the immunophilin TWD1. (takahashi2003hsp90interactswith pages 1-2, takahashi2003hsp90interactswith pages 4-5, geisler2024hsp90providesplasticity pages 1-5, lefa2023proteomeanalysisof pages 9-11)
1. Target identity verification (critical disambiguation)
The symbol HSP90-1 is used across species, but the target here is unambiguously Arabidopsis thaliana cytosolic Hsp90 encoded by AT5G52640, matching UniProt P27323. A foundational Arabidopsis immunity study directly analyzes AtHSP90.1 and athsp90.1 T-DNA mutants in this locus context. (takahashi2003hsp90interactswith pages 4-5)
A recent 2024 study on auxin transport explicitly lists the hsp90.1 genotype as At5g52640 (SALK_007614) and uses HSP90.1-tagged constructs for interaction assays, further confirming identity consistency with UniProt P27323. (geisler2024hsp90providesplasticity pages 17-21)
2. Key concepts and definitions (current understanding)
2.1 Hsp90 molecular chaperone system
Hsp90 proteins are ATP-dependent molecular chaperones that act as dynamic scaffolds for folding/maturation and stabilization of a subset of regulatory proteins (“clients”). Arabidopsis AtHSP90.1 is described with the canonical three-domain architecture: an N-terminal ATPase domain, a middle domain implicated in client interactions, and a C-terminal dimerization region containing a conserved MEEVD motif that binds many TPR-domain cochaperones. (takahashi2003hsp90interactswith pages 2-3)
Co-chaperones regulate Hsp90’s cycle and specify clients. The Arabidopsis immune co-chaperone system includes RAR1 (non-TPR cochaperone) and SGT1 (TPR/CS-domain cochaperone), which interact with HSP90 via distinct binding modes (MEEVD/TPR interactions and additional N-terminal contacts). (takahashi2003hsp90interactswith pages 2-3, takahashi2003hsp90interactswith pages 5-6)
2.2 Client proteins and pathway buffering
A key conceptual point from plant Hsp90 literature is that Hsp90 often buffers the stability/function of signaling proteins, such that Hsp90 inhibition destabilizes clients and disrupts pathway outputs (e.g., immune receptors, transporters). This is directly supported in Arabidopsis by pharmacological Hsp90 inhibition (geldanamycin) and by athsp90.1 genetics affecting effector-triggered immunity. (takahashi2003hsp90interactswith pages 4-5)
3. Molecular function of AtHSP90.1
3.1 Biochemical function: ATP-dependent chaperone
AtHSP90.1 is a cytosolic Hsp90 chaperone with an N-terminal region containing the ATPase domain. Interaction mapping shows RAR1 binds the N-terminal (ATPase-containing) half of HSP90, reinforcing that the canonical Hsp90 ATPase module is present and functionally engaged in cochaperone binding. (takahashi2003hsp90interactswith pages 1-2, takahashi2003hsp90interactswith pages 2-3)
3.2 Co-chaperone interactions specifying function
RAR1: Identified in a yeast two-hybrid screen using RAR1 CHORD-I; deletion/pull-down mapping indicates the HSP90 N-terminal half is sufficient for direct RAR1 interaction and that CHORD-I mediates binding. (takahashi2003hsp90interactswith pages 2-3)
SGT1: SGT1 also interacts with HSP90, with evidence for both a C-terminal MEEVD/TPR-mediated interaction and an additional N-terminal binding site, consistent with multi-site engagement to tune the chaperone cycle. (takahashi2003hsp90interactswith pages 2-3, takahashi2003hsp90interactswith pages 5-6)
HOP/FKBPs: Cytosolic HSP90 cooperates with TPR cochaperones such as HOP and immunophilins (FKBPs) in additional pathways, including chloroplast preprotein targeting and heat acclimation networks. (fellerer2011cytosolichsp90cochaperones pages 7-9, lefa2023proteomeanalysisof pages 14-15)
4. Subcellular localization
AtHSP90.1 is consistently described as a cytosolic/cytoplasmic HSP90 isoform. Arabidopsis contains multiple Hsp90 paralogs, including organellar forms (chloroplast/mitochondrial/ER), but AtHSP90.1 is among the four cytosolic isoforms, distinguishing it from compartment-targeted Hsp90 family members. (takahashi2003hsp90interactswith pages 1-2, kahrizi2025hsp90mediatedstressresilience pages 1-4)
Functionally, “cytosolic” localization is supported by its roles in (i) cytosolic immune receptor maturation/signaling and (ii) interaction with cytosolic-facing trafficking/PM stability modules for ABCB transporters. (takahashi2003hsp90interactswith pages 4-5, geisler2024hsp90providesplasticity pages 21-28)
5. Biological processes and pathways
5.1 Innate immunity: RPS2/RPM1 effector-triggered immunity
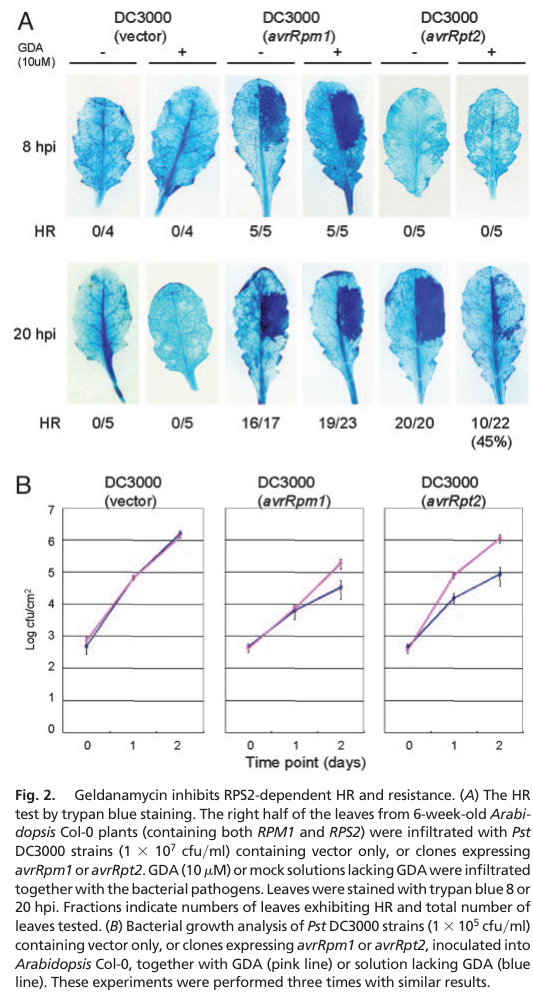
A landmark Arabidopsis study demonstrated AtHSP90.1 is required for full RPS2-mediated resistance and contributes to RPM1 resistance, acting with cochaperones RAR1 and SGT1. (takahashi2003hsp90interactswith pages 5-6, takahashi2003hsp90interactswith pages 4-5)
Quantitative phenotypes / statistics (pathogen growth assays):
- Pharmacological inhibition with the Hsp90 inhibitor geldanamycin (GDA) impaired RPS2-dependent hypersensitive response and increased bacterial titers for Pseudomonas syringae pv. tomato DC3000 carrying avrRpt2 by approximately 5–6× at 1 dpi and ~10× at 2 dpi; for avrRpm1, ~6× higher growth was observed at 2 dpi. (takahashi2003hsp90interactswith pages 4-5)
- athsp90.1 loss-of-function mutants showed ~5–20× higher bacterial growth by 3 dpi under RPS2-dependent conditions, with differences reported as significant at P < 0.05. (takahashi2003hsp90interactswith pages 4-5, takahashi2003hsp90interactswith pages 5-6)
A key interpretive model proposed is that HSP90 supports immune signaling by stabilizing resistance (R/NLR) proteins and/or their immediate signaling complexes, consistent with client destabilization when Hsp90 function is compromised. (takahashi2003hsp90interactswith pages 5-6)
Visual evidence (figure panels): The bacterial growth and disease phenotype panels supporting the above quantitative claims are shown in the extracted figure regions. (takahashi2003hsp90interactswith media 5dd24e95, takahashi2003hsp90interactswith media 14a3512f)
5.2 Protein targeting to chloroplasts (cytosolic chaperone escort)
Cytosolic HSP90 participates in a pathway for post-translational chloroplast preprotein targeting to the TOC import machinery. Co-immunoprecipitation data support a complex containing an FKBP (FKBP73), HSP90, and a chloroplast preprotein; peptide sequencing also identified HOP in preprotein pull-downs. A mechanistic model is proposed where preproteins initially bind HSP70, and a subset then recruit HSP90 (via HOP), supporting docking at TOC components (including Toc64), with FKBP73 potentially replacing HOP through TPR interactions. (fellerer2011cytosolichsp90cochaperones pages 7-9)
Quantitative descriptors: In that study, “more than half” of assessed preproteins used an HSP90+HSP70 route, and 12 tested preproteins specifically bound HSP90; HOP identification in the pull-down included ~9.9% sequence coverage for the wheat HOP homolog used as reference in peptide matching. (fellerer2011cytosolichsp90cochaperones pages 7-9)
5.3 Heat stress and proteostasis (including heat acclimation and recovery)
A 2023 Arabidopsis proteomics study of ROF1/ROF2 (FKBP immunophilins) mutants places HSP90-1 within heat-acclimation proteostasis networks. The authors report that heat acclimation at 37 °C induces heat response/protein folding pathways across genotypes, and that the rof1/rof2 double mutant accumulates significantly higher levels of many HSPs than WT following acclimation, including HSP90-1 as a heat-induced “large HSP”. (lefa2023proteomeanalysisof pages 14-15)
In the same dataset, lists of proteins preferentially accumulated after heat acclimation include HSP90-1, and ontology/KEGG analyses quantify pathway-level enrichment patterns; statistical analysis references FDR thresholds (0.05 and 0.1) for significance calls in comparative analyses. (lefa2023proteomeanalysisof pages 14-15, lefa2023proteomeanalysisof pages 8-9)
Mechanistically, this work also summarizes the Hsp90 cycle at the level of domain transitions and ATP-binding/dimerization, connecting Hsp90 activation to co-chaperone exchange (e.g., HOP dissociation and recruitment of additional cochaperones leading to an “ATPase-competent conformation”). (lefa2023proteomeanalysisof pages 14-15)
5.4 Developmental plasticity and auxin transport (2024 evidence)
Recent work proposes that HSP90 (including HSP90.1) contributes to developmental plasticity by stabilizing plasma membrane presence of ABCB-type auxin transporters, through interaction with the immunophilin cochaperone FKBP42/TWD1. (geisler2024hsp90providesplasticity pages 1-5, geisler2024hsp90providesplasticity pages 21-28)
Interaction and mechanism:
- HSP90.1 physically associates with TWD1 (co-IP and FRET-FLIM evidence), and ABCB1 can pull down HSP90.1/3 upon co-expression, supporting an HSP90–TWD1–ABCB module. (geisler2024hsp90providesplasticity pages 21-28, geisler2024hsp90providesplasticity pages 17-21)
- A prior estimate cited in this work reports HSP90.1–TWD1 binding in the low micromolar KD range. (geisler2024hsp90providesplasticity pages 1-5)
Experimental perturbations and quantitative descriptors:
- Geldanamycin 5 μM for 24 h was used as an HSP90 inhibitor condition in transporter stability/proteomics experiments. (geisler2024hsp90providesplasticity pages 21-28)
- Microsomal quantitative proteomics used TMT-based 16-plex multiplexing with n = 4 biological replicates (as described in the excerpted methods/results text) and statistical significance denoted (e.g., p < 0.05). (geisler2024hsp90providesplasticity pages 21-28)
- Root twisting phenotypes were quantified with n > 20 and reported as significantly increased in twd1, hsp90.1, hsp90.3 and HSP90RNAi backgrounds. (geisler2024hsp90providesplasticity pages 21-28)
These results expand AtHSP90.1 annotation beyond stress/immunity into hormone-regulated developmental processes, consistent with Hsp90 client specialization and co-chaperone-directed specificity. (geisler2024hsp90providesplasticity pages 21-28, geisler2024hsp90providesplasticity pages 12-15)
6. Current applications and real-world implementations
6.1 Chemical biology and inhibitor-based perturbation
Hsp90 inhibitors (e.g., geldanamycin) are routinely used as mechanistic probes in plant biology to reveal dependence of pathways on Hsp90 function. In Arabidopsis, GDA treatment phenocopies aspects of athsp90.1 defects in immunity and affects auxin transporter stability, demonstrating how Hsp90 can be modulated experimentally to dissect client dependence. (takahashi2003hsp90interactswith pages 4-5, geisler2024hsp90providesplasticity pages 21-28)
6.2 Crop improvement relevance (stress and disease resilience)
The network roles of HSP90.1 in thermotolerance/proteostasis and immune receptor stability make it an attractive—but high-risk—node for engineering. Evidence indicates that decreasing Hsp90 activity can compromise ETI, while modulating Hsp90/co-chaperone modules can shift stress acclimation outcomes (e.g., altered HSP accumulation patterns in ROF mutant networks). Thus, translational strategies are more likely to succeed by targeting specific co-chaperones (e.g., ROF1/ROF2; TWD1) or pathway-specific clients rather than globally suppressing Hsp90 activity. (takahashi2003hsp90interactswith pages 4-5, lefa2023proteomeanalysisof pages 14-15, geisler2024hsp90providesplasticity pages 21-28)
7. Expert opinion and authoritative synthesis (evidence-based analysis)
A coherent mechanistic model supported by the cited primary literature is that AtHSP90.1 acts as an ATP-dependent chaperone hub whose physiological specificity is largely determined by co-chaperones:
- In immunity, RAR1 and SGT1 provide a cochaperone module that couples AtHSP90.1 to NLR maturation/stability, explaining the observed increases in pathogen growth when Hsp90 is inhibited or genetically compromised. (takahashi2003hsp90interactswith pages 5-6, takahashi2003hsp90interactswith pages 4-5)
- In development, TWD1 appears to direct Hsp90 action toward ABCB transporter biogenesis/PM stability, giving a plausible mechanism for selective client regulation (ABCBs affected while some other transporters are less affected). (geisler2024hsp90providesplasticity pages 21-28, geisler2024hsp90providesplasticity pages 12-15)
- In heat acclimation, FKBP cochaperones ROF1/ROF2 and downstream HSF/HSP modules provide a route by which AtHSP90.1 participates in thermotolerance acquisition and recovery, consistent with Hsp90’s central role in proteostasis. (lefa2023proteomeanalysisof pages 14-15, lefa2023proteomeanalysisof pages 9-11)
8. Key statistics and data points (recent and classic)
- Immunity (Takahashi 2003, PNAS): ~5–6× (1 dpi) and ~10× (2 dpi) higher bacterial titers under GDA for avrRpt2-triggered resistance; ~5–20× higher bacterial growth by 3 dpi in athsp90.1 mutants; reported P < 0.05 significance for bacterial-growth differences. (takahashi2003hsp90interactswith pages 4-5, takahashi2003hsp90interactswith pages 5-6)
- Development/auxin (Geisler 2024 preprint): HSP90 inhibition with GDA 5 μM (24 h); proteomics n = 4; phenotype quantification n > 20; HSP90.1–TWD1 binding cited in low μM KD range. (geisler2024hsp90providesplasticity pages 21-28, geisler2024hsp90providesplasticity pages 1-5)
- Heat acclimation proteomics (Lefa 2023, ACS Omega): HSP90-1 is a heat-induced HSP and shows higher accumulation patterns in rof1/rof2 double mutant; analysis uses FDR thresholds 0.05/0.1 and reports pathway enrichment scores across conditions. (lefa2023proteomeanalysisof pages 14-15, lefa2023proteomeanalysisof pages 8-9)
- Chloroplast preprotein targeting (Fellerer 2011, Mol Plant): “More than half” of preproteins use an HSP90+HSP70 route; 12 tested preproteins bound HSP90; HOP peptide identification includes ~9.9% coverage. (fellerer2011cytosolichsp90cochaperones pages 7-9)
Evidence map table
| Functional aspect | Key findings | Evidence type | Quantitative/statistical details | Primary source (author, year, journal) with URL |
|---|---|---|---|---|
| Molecular function | AtHSP90.1 (At5g52640; UniProt P27323) is the cytosolic Hsp90 isoform in Arabidopsis with canonical Hsp90 architecture: N-terminal ATPase domain, middle client-binding region, and C-terminal dimerization/MEEVD motif for TPR co-chaperones; functions as an ATP-dependent molecular chaperone/scaffold for client protein maturation and stability (takahashi2003hsp90interactswith pages 2-3, takahashi2003hsp90interactswith pages 1-2) | Biochemistry, genetics | Domain-level mapping showed RAR1 binds the N-terminal ATPase-containing half; geldanamycin used at 10–50 μM in immune assays (takahashi2003hsp90interactswith pages 2-3) | Takahashi et al., 2003, PNAS. https://doi.org/10.1073/pnas.2033934100 |
| Localization | Evidence supports AtHSP90.1 as a cytosolic HSP90; Arabidopsis has four cytosolic HSP90 isoforms, with other family members targeted to chloroplast, mitochondrion, or ER, distinguishing AtHSP90.1 from organellar paralogs (takahashi2003hsp90interactswith pages 1-2, kahrizi2025hsp90mediatedstressresilience pages 1-4, prasad2010insilicoidentification pages 17-17) | Genetics, review/inference | No explicit localization percentage reported in cited excerpts; localization is consistently described as cytosolic/cytoplasmic (takahashi2003hsp90interactswith pages 1-2, kahrizi2025hsp90mediatedstressresilience pages 1-4) | Takahashi et al., 2003, PNAS. https://doi.org/10.1073/pnas.2033934100; Kahrizi et al., 2025, bioRxiv. https://doi.org/10.1101/2025.06.21.660849 |
| Key partners / co-chaperones | AtHSP90.1 physically interacts with RAR1 and SGT1; RAR1 binds the N-terminal ATPase region, while SGT1 engages HSP90 through TPR/CS-related features. These interactions define a plant HSP90–SGT1–RAR1 immune chaperone system for NLR stability/function (takahashi2003hsp90interactswith pages 2-3, takahashi2003hsp90interactswith pages 5-6, takahashi2003hsp90interactswith pages 1-2) | Biochemistry, genetics | Two interaction modes proposed for SGT1 with HSP90; deletion mapping identified HSP90 interaction regions including C-terminal MEEVD-dependent and N-terminal contacts (takahashi2003hsp90interactswith pages 2-3) | Takahashi et al., 2003, PNAS. https://doi.org/10.1073/pnas.2033934100 |
| Pathways / biological processes: immunity | AtHSP90.1 is essential for full RPS2-mediated disease resistance and contributes to RPM1 resistance; it supports hypersensitive response and likely stabilizes NLR immune receptors/complexes (takahashi2003hsp90interactswith pages 5-6, takahashi2003hsp90interactswith pages 1-2, takahashi2003hsp90interactswith pages 4-5) | Genetics, pharmacology, plant pathology | Geldanamycin caused ~5–6-fold higher Pst avrRpt2 bacterial titer at 1 dpi and ~10-fold at 2 dpi; ~6-fold increase for avrRpm1 at 2 dpi. athsp90.1 mutants showed 5–20-fold more bacterial growth by 3 dpi; significance reported at P < 0.05 (takahashi2003hsp90interactswith pages 4-5, takahashi2003hsp90interactswith pages 5-6) | Takahashi et al., 2003, PNAS. https://doi.org/10.1073/pnas.2033934100 |
| Pathways / biological processes: chloroplast preprotein targeting | Cytosolic HSP90, together with HOP and FKBP cochaperones, associates with freshly synthesized chloroplast preproteins and helps route a subset to the TOC import machinery, linking AtHSP90.1-like cytosolic HSP90 activity to post-translational plastid protein targeting (fellerer2011cytosolichsp90cochaperones pages 7-9) | Biochemistry, co-IP, pull-down | “More than half” of tested preproteins used the HSP90+HSP70 route; 12 tested preproteins specifically bound HSP90; TaHOP identified in pull-down with 9.9% sequence coverage (fellerer2011cytosolichsp90cochaperones pages 7-9) | Fellerer et al., 2011, Molecular Plant. https://doi.org/10.1093/mp/ssr037 |
| Pathways / biological processes: heat stress / thermotolerance | AtHSP90.1 participates in heat-stress responses with FKBP co-chaperones ROF1/ROF2 and the HsfA2 network; literature cited in 2023 work indicates ROF1 interacts with HSP90.1 to modulate thermotolerance and small HSP accumulation, and HSP90.1/ROF1 are targets of NBR1-mediated selective autophagy in heat-stress memory (lefa2023proteomeanalysisof pages 17-18) | Proteomics, genetics, review of primary studies | No direct fold-change for AtHSP90.1 in excerpt; rof1 mutants are heat sensitive after 45 °C challenge, whereas rof2 shows greater heat resistance; HSP90.1 listed among proteins enriched in rof1/rof2 double mutants (lefa2023proteomeanalysisof pages 11-14, lefa2023proteomeanalysisof pages 17-18) | Lefa et al., 2023, ACS Omega. https://doi.org/10.1021/acsomega.3c06773 |
| Pathways / biological processes: auxin transport / development (recent 2024) | 2024 work implicates HSP90.1 in development by stabilizing plasma-membrane ABCB auxin transporters through interaction with FKBP42/TWD1; HSP90 inhibition or mutation reduces ABCB abundance, perturbs polar auxin transport, increases root twisting, and alters root auxin distribution (geisler2024hsp90providesplasticity pages 1-5, geisler2024hsp90providesplasticity pages 21-28, geisler2024hsp90providesplasticity pages 17-21, geisler2024hsp90providesplasticity pages 5-7) | Biochemistry, pharmacology, genetics, proteomics | HSP90.1–TWD1 binding reported in low micromolar Kd range; 5 μM geldanamycin for 24 h reduced PM ABCB levels; TMT proteomics used n=4; root twisting quantified with n > 20; significance shown at P < 0.05 (geisler2024hsp90providesplasticity pages 1-5, geisler2024hsp90providesplasticity pages 21-28) | Geisler et al., 2024, Research Square preprint. https://doi.org/10.21203/rs.3.rs-4533687/v1 |
| Phenotypes / quantitative data | athsp90.1 loss-of-function mutants show no obvious morphology under normal growth but are compromised in effector-triggered immunity; pharmacological HSP90 inhibition similarly impairs HR and resistance. This supports a specialized stress/immune buffering role rather than a strong constitutive developmental defect (takahashi2003hsp90interactswith pages 4-5, takahashi2003hsp90interactswith media 5dd24e95, takahashi2003hsp90interactswith media 14a3512f) | Genetics, pharmacology | Pathogen assays used Pst inocula of 10^5–10^7 cfu/mL; bacterial growth measured immediately and 3 dpi; chlorosis increased by 6 dpi in mutants; figure evidence documents CFU/cm² increases in mutants and geldanamycin-treated plants (takahashi2003hsp90interactswith pages 4-5, takahashi2003hsp90interactswith media 5dd24e95) | Takahashi et al., 2003, PNAS. https://doi.org/10.1073/pnas.2033934100 |
| Recent 2023–2024 findings | 2023 proteomics linked HSP90-1 abundance to ROF-FKBP-dependent heat acclimation networks; 2024 studies/reviews extended HSP90.1 relevance to selective autophagy/heat recovery and to auxin-transporter stabilization, broadening its annotation beyond immunity alone (lefa2023proteomeanalysisof pages 11-14, geisler2024hsp90providesplasticity pages 21-28, lefa2023proteomeanalysisof pages 17-18) | Proteomics, systems biology, review, preprint | In rof datasets, HSP90-1 was differentially accumulated/enriched in double mutants; in auxin work, HSP90-dependent microsomal proteomics used 16-plex TMT and GDA 5 μM for 24 h (geisler2024hsp90providesplasticity pages 21-28, lefa2023proteomeanalysisof pages 9-11) | Lefa et al., 2023, ACS Omega. https://doi.org/10.1021/acsomega.3c06773; Geisler et al., 2024, Research Square preprint. https://doi.org/10.21203/rs.3.rs-4533687/v1 |
| Applications / real-world relevance | AtHSP90.1 is a plausible target for engineering stress resilience and disease resistance, but also a cautionary node because HSP90 inhibition can suppress immunity and auxin-dependent development. Plant HSP90 biology is already used conceptually in chemical biology (geldanamycin/radicicol-class inhibition) and in crop stress-improvement strategies through chaperone/co-chaperone manipulation (takahashi2003hsp90interactswith pages 4-5, fellerer2011cytosolichsp90cochaperones pages 7-9, geisler2024hsp90providesplasticity pages 21-28) | Pharmacology, translational inference | Practical perturbants include geldanamycin 5–50 μM depending on assay; recent work links HSP90-dependent proteostasis to developmental plasticity and thermotolerance pathways, making the system relevant for crop engineering (takahashi2003hsp90interactswith pages 4-5, geisler2024hsp90providesplasticity pages 21-28) | Takahashi et al., 2003, PNAS. https://doi.org/10.1073/pnas.2033934100; Geisler et al., 2024, Research Square preprint. https://doi.org/10.21203/rs.3.rs-4533687/v1 |
Table: This table summarizes functional annotation evidence for Arabidopsis thaliana HSP90-1/AtHSP90.1 (At5g52640; UniProt P27323), including molecular role, localization, interaction partners, pathways, phenotypes, and recent 2023–2024 findings. It is useful as a compact evidence map linking specific claims to experimental modalities, quantitative details, and source URLs.
References (URLs and publication dates)
- Takahashi A. et al. 2003-09. PNAS. “HSP90 interacts with RAR1 and SGT1 and is essential for RPS2-mediated disease resistance in Arabidopsis.” https://doi.org/10.1073/pnas.2033934100 (takahashi2003hsp90interactswith pages 4-5, takahashi2003hsp90interactswith pages 2-3)
- Fellerer C. et al. 2011-11. Molecular Plant. “Cytosolic HSP90 cochaperones HOP and FKBP interact with freshly synthesized chloroplast preproteins of Arabidopsis.” https://doi.org/10.1093/mp/ssr037 (fellerer2011cytosolichsp90cochaperones pages 7-9)
- Lefa P. et al. 2023-12. ACS Omega (vol 9 issue year listed as 2024 in PDF header; article year 2023). “Proteome Analysis of the ROF-FKBP Mutants…” https://doi.org/10.1021/acsomega.3c06773 (lefa2023proteomeanalysisof pages 14-15, lefa2023proteomeanalysisof pages 8-9)
- Geisler M. et al. 2024-06. Research Square (preprint). “HSP90 provides plasticity to plant development by gradually stabilizing plasma membrane presence of ABCB-type auxin transporters.” https://doi.org/10.21203/rs.3.rs-4533687/v1 (geisler2024hsp90providesplasticity pages 21-28, geisler2024hsp90providesplasticity pages 1-5)
Limitations of this report
Despite targeted searching, some classic Arabidopsis HSP90.1 client/pathway papers (e.g., circadian client ZTL stabilization) were not retrievable in full text within the current tool session; therefore, circadian roles are not described here beyond general Hsp90 client concepts. Claims above are restricted to evidence available in the cited sources.
References
-
(takahashi2003hsp90interactswith pages 1-2): Akira Takahashi, Catarina Casais, Kazuya Ichimura, and Ken Shirasu. Hsp90 interacts with rar1 and sgt1 and is essential for rps2-mediated disease resistance in arabidopsis. Proceedings of the National Academy of Sciences of the United States of America, 100:11777-11782, Sep 2003. URL: https://doi.org/10.1073/pnas.2033934100, doi:10.1073/pnas.2033934100. This article has 655 citations and is from a highest quality peer-reviewed journal.
-
(takahashi2003hsp90interactswith pages 4-5): Akira Takahashi, Catarina Casais, Kazuya Ichimura, and Ken Shirasu. Hsp90 interacts with rar1 and sgt1 and is essential for rps2-mediated disease resistance in arabidopsis. Proceedings of the National Academy of Sciences of the United States of America, 100:11777-11782, Sep 2003. URL: https://doi.org/10.1073/pnas.2033934100, doi:10.1073/pnas.2033934100. This article has 655 citations and is from a highest quality peer-reviewed journal.
-
(geisler2024hsp90providesplasticity pages 1-5): Markus Geisler, Tashi Tsering, Martin Di Donato, Despina Samakovli, Dimitra Milioni, Elisa Azarello, Stefano Mancuso, Vendula Pukyšová, Marta Zwiewka, Tomasz Nodzynski, Michael Stumpe, Jutta Ludwig-Müller, Aurelien Bailly, and Polydefkis Hatzopoulos. Hsp90 provides plasticity to plant development by gradually stabilizing plasma membrane presence of abcb-type auxin transporters. Unknown journal, Jun 2024. URL: https://doi.org/10.21203/rs.3.rs-4533687/v1, doi:10.21203/rs.3.rs-4533687/v1.
-
(lefa2023proteomeanalysisof pages 9-11): Paraskevi Lefa, Martina Samiotaki, and Theodora Farmaki. Proteome analysis of the rof-fkbp mutants reveals functional relations among heat stress responses, plant development, and protein quality control during heat acclimation in arabidopsis thaliana. ACS Omega, 9:2391-2408, Dec 2023. URL: https://doi.org/10.1021/acsomega.3c06773, doi:10.1021/acsomega.3c06773. This article has 6 citations and is from a peer-reviewed journal.
-
(geisler2024hsp90providesplasticity pages 17-21): Markus Geisler, Tashi Tsering, Martin Di Donato, Despina Samakovli, Dimitra Milioni, Elisa Azarello, Stefano Mancuso, Vendula Pukyšová, Marta Zwiewka, Tomasz Nodzynski, Michael Stumpe, Jutta Ludwig-Müller, Aurelien Bailly, and Polydefkis Hatzopoulos. Hsp90 provides plasticity to plant development by gradually stabilizing plasma membrane presence of abcb-type auxin transporters. Unknown journal, Jun 2024. URL: https://doi.org/10.21203/rs.3.rs-4533687/v1, doi:10.21203/rs.3.rs-4533687/v1.
-
(takahashi2003hsp90interactswith pages 2-3): Akira Takahashi, Catarina Casais, Kazuya Ichimura, and Ken Shirasu. Hsp90 interacts with rar1 and sgt1 and is essential for rps2-mediated disease resistance in arabidopsis. Proceedings of the National Academy of Sciences of the United States of America, 100:11777-11782, Sep 2003. URL: https://doi.org/10.1073/pnas.2033934100, doi:10.1073/pnas.2033934100. This article has 655 citations and is from a highest quality peer-reviewed journal.
-
(takahashi2003hsp90interactswith pages 5-6): Akira Takahashi, Catarina Casais, Kazuya Ichimura, and Ken Shirasu. Hsp90 interacts with rar1 and sgt1 and is essential for rps2-mediated disease resistance in arabidopsis. Proceedings of the National Academy of Sciences of the United States of America, 100:11777-11782, Sep 2003. URL: https://doi.org/10.1073/pnas.2033934100, doi:10.1073/pnas.2033934100. This article has 655 citations and is from a highest quality peer-reviewed journal.
-
(fellerer2011cytosolichsp90cochaperones pages 7-9): Christine Fellerer, Regina Schweiger, Katharina Schöngruber, Jürgen Soll, and Serena Schwenkert. Cytosolic hsp90 cochaperones hop and fkbp interact with freshly synthesized chloroplast preproteins of arabidopsis. Molecular plant, 4 6:1133-45, Nov 2011. URL: https://doi.org/10.1093/mp/ssr037, doi:10.1093/mp/ssr037. This article has 109 citations and is from a highest quality peer-reviewed journal.
-
(lefa2023proteomeanalysisof pages 14-15): Paraskevi Lefa, Martina Samiotaki, and Theodora Farmaki. Proteome analysis of the rof-fkbp mutants reveals functional relations among heat stress responses, plant development, and protein quality control during heat acclimation in arabidopsis thaliana. ACS Omega, 9:2391-2408, Dec 2023. URL: https://doi.org/10.1021/acsomega.3c06773, doi:10.1021/acsomega.3c06773. This article has 6 citations and is from a peer-reviewed journal.
-
(kahrizi2025hsp90mediatedstressresilience pages 1-4): Zahra Kahrizi, Christos Michailidis, Karel Raabe, Jan Fíla, Božena Klodová, Jiří Rudolf, Petra Procházková Schrumpfová, David Honys, and Sotirios Fragkostefanakis. Hsp90-mediated stress resilience in male gametophyte of arabidopsis thaliana. bioRxiv, Jun 2025. URL: https://doi.org/10.1101/2025.06.21.660849, doi:10.1101/2025.06.21.660849. This article has 2 citations.
-
(geisler2024hsp90providesplasticity pages 21-28): Markus Geisler, Tashi Tsering, Martin Di Donato, Despina Samakovli, Dimitra Milioni, Elisa Azarello, Stefano Mancuso, Vendula Pukyšová, Marta Zwiewka, Tomasz Nodzynski, Michael Stumpe, Jutta Ludwig-Müller, Aurelien Bailly, and Polydefkis Hatzopoulos. Hsp90 provides plasticity to plant development by gradually stabilizing plasma membrane presence of abcb-type auxin transporters. Unknown journal, Jun 2024. URL: https://doi.org/10.21203/rs.3.rs-4533687/v1, doi:10.21203/rs.3.rs-4533687/v1.
-
(takahashi2003hsp90interactswith media 5dd24e95): Akira Takahashi, Catarina Casais, Kazuya Ichimura, and Ken Shirasu. Hsp90 interacts with rar1 and sgt1 and is essential for rps2-mediated disease resistance in arabidopsis. Proceedings of the National Academy of Sciences of the United States of America, 100:11777-11782, Sep 2003. URL: https://doi.org/10.1073/pnas.2033934100, doi:10.1073/pnas.2033934100. This article has 655 citations and is from a highest quality peer-reviewed journal.
-
(takahashi2003hsp90interactswith media 14a3512f): Akira Takahashi, Catarina Casais, Kazuya Ichimura, and Ken Shirasu. Hsp90 interacts with rar1 and sgt1 and is essential for rps2-mediated disease resistance in arabidopsis. Proceedings of the National Academy of Sciences of the United States of America, 100:11777-11782, Sep 2003. URL: https://doi.org/10.1073/pnas.2033934100, doi:10.1073/pnas.2033934100. This article has 655 citations and is from a highest quality peer-reviewed journal.
-
(lefa2023proteomeanalysisof pages 8-9): Paraskevi Lefa, Martina Samiotaki, and Theodora Farmaki. Proteome analysis of the rof-fkbp mutants reveals functional relations among heat stress responses, plant development, and protein quality control during heat acclimation in arabidopsis thaliana. ACS Omega, 9:2391-2408, Dec 2023. URL: https://doi.org/10.1021/acsomega.3c06773, doi:10.1021/acsomega.3c06773. This article has 6 citations and is from a peer-reviewed journal.
-
(geisler2024hsp90providesplasticity pages 12-15): Markus Geisler, Tashi Tsering, Martin Di Donato, Despina Samakovli, Dimitra Milioni, Elisa Azarello, Stefano Mancuso, Vendula Pukyšová, Marta Zwiewka, Tomasz Nodzynski, Michael Stumpe, Jutta Ludwig-Müller, Aurelien Bailly, and Polydefkis Hatzopoulos. Hsp90 provides plasticity to plant development by gradually stabilizing plasma membrane presence of abcb-type auxin transporters. Unknown journal, Jun 2024. URL: https://doi.org/10.21203/rs.3.rs-4533687/v1, doi:10.21203/rs.3.rs-4533687/v1.
-
(prasad2010insilicoidentification pages 17-17): Bishun D. Prasad, Shilpi Goel, and Priti Krishna. In silico identification of carboxylate clamp type tetratricopeptide repeat proteins in arabidopsis and rice as putative co-chaperones of hsp90/hsp70. PLoS ONE, 5:e12761, Sep 2010. URL: https://doi.org/10.1371/journal.pone.0012761, doi:10.1371/journal.pone.0012761. This article has 117 citations and is from a peer-reviewed journal.
-
(lefa2023proteomeanalysisof pages 17-18): Paraskevi Lefa, Martina Samiotaki, and Theodora Farmaki. Proteome analysis of the rof-fkbp mutants reveals functional relations among heat stress responses, plant development, and protein quality control during heat acclimation in arabidopsis thaliana. ACS Omega, 9:2391-2408, Dec 2023. URL: https://doi.org/10.1021/acsomega.3c06773, doi:10.1021/acsomega.3c06773. This article has 6 citations and is from a peer-reviewed journal.
-
(lefa2023proteomeanalysisof pages 11-14): Paraskevi Lefa, Martina Samiotaki, and Theodora Farmaki. Proteome analysis of the rof-fkbp mutants reveals functional relations among heat stress responses, plant development, and protein quality control during heat acclimation in arabidopsis thaliana. ACS Omega, 9:2391-2408, Dec 2023. URL: https://doi.org/10.1021/acsomega.3c06773, doi:10.1021/acsomega.3c06773. This article has 6 citations and is from a peer-reviewed journal.
-
(geisler2024hsp90providesplasticity pages 5-7): Markus Geisler, Tashi Tsering, Martin Di Donato, Despina Samakovli, Dimitra Milioni, Elisa Azarello, Stefano Mancuso, Vendula Pukyšová, Marta Zwiewka, Tomasz Nodzynski, Michael Stumpe, Jutta Ludwig-Müller, Aurelien Bailly, and Polydefkis Hatzopoulos. Hsp90 provides plasticity to plant development by gradually stabilizing plasma membrane presence of abcb-type auxin transporters. Unknown journal, Jun 2024. URL: https://doi.org/10.21203/rs.3.rs-4533687/v1, doi:10.21203/rs.3.rs-4533687/v1.
Artifacts
Citations
- lefa2023proteomeanalysisof pages 14-15
- lefa2023proteomeanalysisof pages 17-18
- lefa2023proteomeanalysisof pages 9-11
- lefa2023proteomeanalysisof pages 8-9
- prasad2010insilicoidentification pages 17-17
- lefa2023proteomeanalysisof pages 11-14
- https://doi.org/10.1073/pnas.2033934100
- https://doi.org/10.1073/pnas.2033934100;
- https://doi.org/10.1101/2025.06.21.660849
- https://doi.org/10.1093/mp/ssr037
- https://doi.org/10.1021/acsomega.3c06773
- https://doi.org/10.21203/rs.3.rs-4533687/v1
- https://doi.org/10.1021/acsomega.3c06773;
- https://doi.org/10.1073/pnas.2033934100,
- https://doi.org/10.21203/rs.3.rs-4533687/v1,
- https://doi.org/10.1021/acsomega.3c06773,
- https://doi.org/10.1093/mp/ssr037,
- https://doi.org/10.1101/2025.06.21.660849,
- https://doi.org/10.1371/journal.pone.0012761,
Perplexity
(AT5G52640-deep-research-perplexity.md)
Perplexity
(AT5G52640-deep-research-perplexity.md)The proposed structure will allow for a comprehensive exploration of the gene's characteristics, from its molecular details to its broader biological implications. I'll ensure a logical progression that connects molecular mechanisms to physiological outcomes.
Heat Shock Protein 90-1 (HSP90-1, AT5G52640) in Arabidopsis thaliana: Molecular Function and Physiological Roles
The heat shock protein 90 (HSP90) family represents one of the most abundant and highly conserved molecular chaperone systems across eukaryotic organisms, and HSP90-1 (AT5G52640) constitutes a central component of this machinery in Arabidopsis thaliana. As a 90-kiloDalton ATP-dependent molecular chaperone, HSP90-1 functions as a sophisticated folding machine that facilitates the maturation, stability, and regulation of a diverse array of client proteins involved in signal transduction, developmental control, and stress responses[2][14]. In Arabidopsis, this gene encodes one of four cytoplasmic HSP90 isoforms that exhibit functional redundancy yet display distinct tissue-specific expression patterns and regulatory roles[2]. Unlike the constitutively low expression of heat shock proteins in prokaryotic systems, HSP90-1 expression is developmentally regulated and responsive to multiple environmental stimuli, including heat stress, heavy metals, and light transitions, positioning it as a critical nexus for cellular adaptation to changing conditions[2][16]. The protein performs its chaperone function through a remarkable ATP-dependent conformational cycle that enables it to recognize, bind, and process structurally diverse client proteins while maintaining them in near-native conformations poised for activation by specific signals such as ligand binding or phosphorylation. This comprehensive report synthesizes current knowledge regarding the molecular mechanisms, biochemical properties, cellular localization, client protein networks, and diverse physiological functions of HSP90-1, with emphasis on the precise molecular details that distinguish this protein as an integrator of signal transduction, developmental programming, and stress adaptation in plant systems.
Molecular Structure, Domain Organization, and Biochemical Properties of HSP90-1
The HSP90-1 protein exhibits the characteristic architecture conserved throughout the HSP90 family of molecular chaperones, comprising three major functional domains that coordinate its ATP-dependent chaperone activity and cochaperone interactions[2][26]. The N-terminal domain (NTD) encompasses the primary ATP-binding pocket, a distinctive Bergerat fold that represents a non-conventional nucleotide-binding site unique to the GHKL superfamily of ATPases[29][50]. This domain contains a highly conserved adenosine triphosphate-binding site that exhibits ATP-binding and hydrolysis activities essential for the chaperone cycle[7][26]. The central middle domain connects the N-terminal and C-terminal regions and plays critical roles in both substrate binding and conformational reorganization during the ATPase cycle[50]. The C-terminal domain (CTD) terminates with the highly conserved pentapeptide motif MEEVD, which serves as the primary binding platform for cochaperone proteins containing tetratricopeptide repeat (TPR) domains[2]. Critically, the HSP90-1 sequence includes this C-terminal pentapeptide motif characteristic of cytoplasmic HSP90 isoforms, distinguishing it from the plastidial (HSP90-5), mitochondrial (HSP90-6), and endoplasmic reticulum-resident (HSP90-7) family members[2]. Additionally, the HSP90-1 protein contains an adjacent charged linker domain positioned between the N-terminal domain and the middle domain, a feature that appears common in eukaryotic and cytoplasmic HSP90 sequences but is notably absent in bacterial homologs[2][26].
The functional properties of HSP90-1 fundamentally depend on its capacity to bind and hydrolyze ATP in a manner that drives conformational transitions essential for client protein processing[7]. In vitro biochemical studies have demonstrated that HSP90-1, like other members of the HSP90 family, possesses inherent ATPase activity that is sensitive to inhibition by the HSP90-specific antibiotic geldanamycin, a natural product compound that binds specifically to the ATP-binding pocket of the N-terminal domain[7][42]. ATP binding to HSP90-1 induces a transition from an open, nucleotide-free conformation toward a more closed state characterized by N-terminal domain dimerization and juxtaposition of the middle domains[50]. This ATP-induced conformational transition represents a critical mechanistic feature that enables HSP90-1 to form the characteristic "molecular clamp" configuration proposed to accommodate diverse client proteins[50]. However, single-molecule fluorescence studies have revealed that, in contrast to the deterministic mechanism observed in prokaryotic HSP90 homologs such as Escherichia coli HtpG, the conformational dynamics of eukaryotic cytoplasmic HSP90 proteins including Arabidopsis HSP90-1 are substantially more stochastic in nature[8]. Rather than ATP binding and hydrolysis unidirectionally determining specific conformations, HSP90-1 exists in a dynamic equilibrium between multiple conformational states, with ATP binding and hydrolysis shifting the equilibrium distribution toward different populations of states rather than locking the protein into discrete conformations[8][50].
The charged linker region connecting the N-terminal domain and middle domain appears to play a specialized role in regulating the substrate-binding properties of HSP90-1[26]. Biochemical analysis of N-terminal fragments containing the charged linker region (N272) compared to the isolated N-terminal domain (N210) revealed that the charged linker increases the substrate affinity of the N-terminal chaperone site while simultaneously establishing crosstalk between peptide and ATP binding[26]. Specifically, peptide binding to the charged linker-containing fragment decreases its affinity for ATP and the HSP90-specific inhibitor geldanamycin, whereas ATP-binding properties of the isolated N-terminal domain remain uninfluenced by peptide binding[26]. This crosstalk mechanism suggests that the eukaryotic-specific charged linker domain represents a functional innovation that allows more sophisticated regulation of HSP90-1's chaperone activity compared to simpler prokaryotic systems. Furthermore, the HSP90-1 protein functions as a homodimer in its active state, with dimerization mediated through both N-terminal and C-terminal interfaces[2][50]. The C-terminal dimer interface was traditionally viewed as constitutively stable; however, recent single-molecule studies revealed that this interface undergoes rapid opening and closing dynamics on the timescale of seconds[43]. These C-terminal dynamics are modulated by nucleotide occupation at the N-terminal domain and cochaperone binding, demonstrating allosteric communication between the two terminal domains of the HSP90-1 protein[43]. The C-terminal open-closed dynamics observed for HSP90-1 have significant implications for cochaperone and client protein recruitment, as the dynamic opening of the C-terminus may facilitate the association and dissociation of TPR-containing cochaperones bearing the C-terminal MEEVD-binding domain[43].
Subcellular Localization and Tissue-Specific Expression Patterns
The HSP90-1 protein is localized to the cytoplasm and nucleus of Arabidopsis cells, consistent with its role as a cytoplasmic molecular chaperone[2][20]. The gene encoding HSP90-1 (AT5G52640) is positioned on chromosome 5 of the Arabidopsis genome[1][16]. The expression of HSP90-1 displays remarkable developmental and tissue-specific regulation that differs substantially from the other cytoplasmic HSP90 isoforms in Arabidopsis[2][49]. During unstressed conditions in seedlings, HSP90-1 transcripts could be detected only in the root tissues of control Arabidopsis plants; however, transcripts became abundant throughout all plant organs following exposure to heat shock or treatment with heavy metals[2]. This root-preferential expression pattern in unstressed conditions suggests that HSP90-1 may play specialized roles in root development and function under normal growth conditions. The developmental analysis of HSP90-1 expression reveals that its promoter displays low activity throughout early stages of embryo development but exhibits markedly elevated activity just before embryo maturation, with expression most prominent in cotyledon tissues[49]. This pattern indicates a critical role for HSP90-1 in late embryonic development and the preparation of tissues for postembryonic growth.
In contrast to HSP90-1, the other cytoplasmic isoforms HSP90-2 and HSP90-3 display distinct expression patterns. HSP90-2 and HSP90-3 transcripts can be detected in all plant organs under unstressed conditions, with particular abundance in roots and flowers[2]. Following heat shock treatment, the transcript levels of HSP90-2 and HSP90-3 increase modestly, whereas these isoforms show substantial upregulation following treatment with the phytohormone indoleacetic acid (auxin), high salt concentration, or heavy metals[2]. The developmental expression pattern of HSP90-3 is nearly constant and restricted to root and cotyledon tips of the embryo under unstressed conditions, with this pattern dramatically altered upon heat stress exposure[49]. These differential expression patterns suggest that while HSP90 family members exhibit functional redundancy, they have been evolutionarily partitioned to assume distinct regulatory roles during particular developmental stages or in response to specific environmental signals. The spatial distribution of HSP90-1 expression in vascular tissues, leaf veins, siliques, and pollen sacs becomes prominent following heat stress, suggesting that stress conditions reorient HSP90-1 expression toward tissues with elevated metabolic demands or where protein stability is particularly critical[49].
The nuclear-cytoplasmic partitioning of HSP90-1 function has been experimentally demonstrated through nuclear translocation studies in mammalian cells that involved targeted nuclear localization of the chicken Hsp90 homolog[19][22]. These studies established that HSP90 proteins can physically interact with nuclear client proteins and participate in their functional maturation within the nuclear compartment[19]. In Arabidopsis, the TIR1 auxin receptor represents a known client of HSP90-1 that localizes to the nucleus, and the HSP90-TIR1 complex forms in both cytoplasmic and nuclear pools, suggesting that HSP90-1 shuttles between compartments or maintains persistent nuclear pools required for specific functions[21][24]. The balance between cytoplasmic and nuclear HSP90-1 pools appears to be dynamic and responsive to cellular conditions, as the stabilization of the nuclear auxin receptor TIR1 by HSP90-1 occurs in temperature-dependent manner, with increased HSP90 protein levels observed following temperature elevation to 29°C[24].
ATP-Dependent Conformational Cycle and Mechanistic Basis of Chaperone Function
The HSP90-1 protein employs an ATP-dependent conformational cycle as the fundamental mechanism underlying its role as a molecular chaperone[7][50]. This cycle represents a highly conserved mechanism shared across HSP90 orthologs from bacteria to humans, though the specific kinetics and dynamics of transitions differ between prokaryotic and eukaryotic forms[53]. The ATP-dependent cycle can be conceptualized as beginning from an open, nucleotide-free conformation characterized by separation of the N-terminal domains and exposure of hydrophobic binding surfaces on the periphery of the protein dimer[50]. In this open state, HSP90-1 exhibits low affinity for ATP and displays enhanced accessibility for client protein binding. The addition of ATP to HSP90-1 induces a conformational transition toward a more compact, closed state where the N-terminal domains approximate one another and form an intimate interface[43][50]. ATP binding and hydrolysis establish directionality to the functional cycle while simultaneously modulating the conformational populations sampled by HSP90-1[50]. The ATP-bound closed state is stabilized by formation of an interface between the N-terminal domain "lids" that fold over the ATP-binding pockets in each monomer[46]. Following ATP hydrolysis to ADP, HSP90-1 transitions to a compact state where hydrophobic surfaces that were accessible in the open conformation become buried at the dimer interface, resulting in the release of bound client proteins and cochaperones[50].
The remarkable feature of HSP90-1's conformational dynamics is that these transitions are not deterministic but rather probabilistic in nature, with the protein sampling an ensemble of distinct conformational states at any given moment[8][50]. Single-molecule fluorescence resonance energy transfer (smFRET) measurements have revealed that even in the presence of ATP or non-hydrolyzable ATP analogs, HSP90-1 continues to populate an array of distinct open and closed states rather than adopting a single discrete conformation[8][53]. This inherent conformational heterogeneity represents a fundamental distinction between eukaryotic HSP90 proteins like HSP90-1 and the more rigid prokaryotic HtpG, which displays more deterministic conformational transitions coupled to ATP hydrolysis[8][53]. The conformational dynamics of HSP90-1 are further modulated by the binding of cochaperones and client proteins, with different cochaperones establishing distinct effects on the equilibrium between open and closed states[8][43]. For instance, the cochaperone Sti1 (yeast Hop homolog in plants) acts to stabilize the open conformation of HSP90-1 by binding to the C-terminal MEEVD motifs and inhibiting the ATPase activity[43]. In contrast, other cochaperones such as Aha1 promote the closed conformation and accelerate the ATPase cycle[8].
The mechanistic basis by which HSP90-1 facilitates client protein folding and activation remains incompletely understood, as the molecular details of how the conformational transitions directly promote client maturation have proven challenging to resolve experimentally[6]. The classical model proposed that HSP90-1 functions as a "molecular clamp" by enclosing client proteins within the lumen formed between N-terminally dimerized monomers, thus keeping multidomain clients physically separated and facilitating their independent folding[6]. However, high-resolution structural studies and single-molecule imaging revealed that many HSP90-1 clients bind to the external surfaces of the N-terminally open protein conformation rather than within the closed lumen, suggesting that multiple distinct binding modes accommodate the diverse clientele[6]. The identification of multiple client-binding sites on HSP90-1, including regions within the N-terminal domain, middle domain, and at the dimer interface, provides a mechanistic explanation for how HSP90-1 achieves specificity despite binding structurally and functionally diverse substrates[6]. Some clients appear to be held by HSP90-1 in a partially unfolded state, a role termed "holdase" activity, which prevents the formation of aggregation-prone intermediates and toxic oligomeric species[6]. For kinase clients, HSP90-1 has been proposed to facilitate reactivation of locally destabilized kinases by maintaining them in a conformation competent for activation by phosphorylation or other posttranslational modifications[6].
Recent investigations have revealed that ATP plays a structural role in HSP90-1 function distinct from its traditional role as a driving force for conformational changes[10]. These studies demonstrated that ATP binding to the N-terminal domain induces allosteric effects on distal regions of the protein, thereby modulating overall protein dynamics and the propensity to sample particular conformational states[10]. This ATP-structural role complements the ATP-chemical energy function previously characterized, suggesting that HSP90-1 has evolved multiple mechanisms through which nucleotide binding influences its functional properties. The precise timing and order of individual molecular events within the HSP90-1 cycle, including the sequence of cochaperone binding, ATP hydrolysis, client protein binding, and product release, appear to be fluid and reversible rather than strictly unidirectional[8]. This flexibility enables HSP90-1 to accommodate the distinct biochemical requirements of its diverse clientele and to respond dynamically to the cellular environment and cofactor availability.
Client Protein Recognition and Substrate Specificity of HSP90-1
The HSP90-1 protein exhibits remarkable specificity in recognizing and binding its client proteins, despite the apparent structural and functional diversity of known clients ranging from protein kinases to transcription factors to transport proteins[11][12][38]. The substrate specificity of HSP90-1 is not intrinsic to the chaperone itself but rather is conferred and regulated by the associated network of cochaperone proteins that interact with both HSP90-1 and the client proteins[11]. The main classes of characterized HSP90-1 client proteins in Arabidopsis and other eukaryotic systems include protein kinases, hormone receptors, and a diverse array of signaling proteins, many of which are direct or indirect regulators of developmental processes and stress responses[38]. Unlike the general molecular chaperones Hsp70 and Hsp40, which recognize and bind unfolded polypeptide chains in extended conformations, HSP90-1 characteristically maintains client proteins in a nearly completely folded conformation that is poised to respond to an activation signal such as ligand binding, phosphorylation, or association with regulatory proteins[12][38]. This fundamental difference in client binding modality positions HSP90-1 as a specialized chaperone dedicated to the maturation and activation of proteins already substantially folded rather than to the initial folding of newly synthesized proteins.
The protein kinase family represents one of the most well-characterized classes of HSP90-1 clients in plant systems. In Arabidopsis, kinase clients of HSP90-1 require the kinase-specific cochaperone Cdc37 for efficient chaperoning and maturation[38]. The interaction between HSP90-1 and kinase clients appears to be mediated through Cdc37, which functions as an adaptor protein that stabilizes kinases in an open conformation competent for HSP90-1 binding[9]. The cochaperone Cdc37 exhibits an unfoldase activity specific for kinases, distinguishing it from the more general HSP90 cochaperones[9]. The maturation of kinase clients by HSP90-1 requires multiple cochaperones beyond Cdc37, including the conserved cochaperones p23 (Sba1) and Aha1[9][25][28]. During the kinase chaperoning cycle, both HSP90-1 and Cdc37 undergo post-translational modifications including phosphorylation that regulate progression through the chaperone cycle and confer specificity for particular kinase substrates[9]. Specifically, phosphorylation of Cdc37 at Y298 results in partial unfolding of the C-terminal domain of Cdc37 and population of folding intermediates that facilitate HSP90-1 phosphorylation at Y197, which then serves as a docking site to recruit non-receptor tyrosine kinases to the HSP90-1-Cdc37 complex via their SH2 domains[9].
The hormone receptor family represents another critical class of HSP90-1 clients in Arabidopsis, particularly for the auxin receptor TIR1 and its homologs, which belong to the F-box protein family of E3 ubiquitin ligase components[21][24]. HSP90-1 associates with TIR1 in plant cells and is required for the nuclear localization and functional activity of the auxin receptor[21]. The HSP90-1-mediated stabilization of TIR1 is temperature-dependent, with HSP90 levels increasing dramatically within one hour of temperature shifts to 29°C, accompanied by rapid stabilization of TIR1 protein levels[24]. The HSP90-1-SGT1b cochaperone complex functions as the chaperone system for TIR1 in the nucleus, with SGT1b displaying reduced binding to TIR1 in auxin-resistant eta3 mutants, suggesting direct involvement of this cochaperone in client recognition[24]. The HSP90-1 regulation of TIR1 stability and function represents a mechanism for integrating environmental temperature signals with auxin-dependent developmental responses, linking the HSP90-1 chaperone system to temperature-controlled plant development[16][24].
Beyond kinases and hormone receptors, HSP90-1 maintains functional relationships with a diverse array of client proteins in Arabidopsis that participate in distinct biological processes. These clients include the F-box protein ZEITLUPE (ZTL), a central component of the plant circadian clock that is stabilized by HSP90-1 through an ATPase-dependent mechanism[51]. The interaction between HSP90-1 and ZTL represents a unique instance where HSP90 functions within the core oscillator of the circadian clock rather than at the periphery, placing HSP90-1 in a particularly central role for circadian timekeeping in plants[51]. The resistance (R) proteins that mediate plant immunity to pathogenic microorganisms represent another class of HSP90-1 clients, with HSP90-1 functioning together with the cochaperones RAR1 and SGT1 to stabilize and regulate these immune surveillance proteins[20][33]. The specific HSP90-1 isoform, AtHSP90.1, is rapidly induced upon pathogen challenge and is required for full RPM1-mediated resistance to the bacterial pathogen Pseudomonas syringae[33].
The ABCB-type auxin transporters represent a more recently identified class of HSP90-1 clients with important roles in polar auxin transport and developmental patterning[35]. HSP90-1 works in conjunction with the cochaperone TWD1 to stabilize ABCB1, ABCB4, and ABCB19 proteins at the plasma membrane, thereby regulating their steady-state abundance and subcellular localization[35]. The regulation of ABCB transporters by HSP90-1 appears to be temporally, spatially, and mechanistically distinct from HSP90-1's regulation of the nuclear auxin receptor TIR1[35]. Whereas TIR1 stabilization by HSP90-1 appears to be stress-related and responsive to temperature elevation, the ABCB stabilization by HSP90-1 appears constitutive rather than stress-dependent[35]. The differential sensitivities of ABCB proteins to HSP90 inhibition follow the order ABCB1 >> ABCB19 > ABCB4, suggesting that HSP90-1 exerts client-specific effects on protein stability and trafficking[35].
The mechanisms of client recognition by HSP90-1 remain only partially resolved, but emerging evidence suggests that HSP90-1 recognizes amino acid sequence motifs or structural features shared among diverse clients[6]. The identification of multiple classes of Hsp90-specific inhibitors including geldanamycin derivatives and alternative inhibitor scaffolds has demonstrated that some inhibitors prevent HSP90-1 activity toward certain clients while sparing others, indicating that the N-terminal ATP-binding domain exhibits conformational or dynamic states that are client-specific[25]. The availability of HSP90-1 can also be rate-limiting for client maturation under certain conditions, as evidenced by the demonstration that geldanamycin-induced inhibition of HSP90-1 leads to accumulation of clients in inactive, partially folded forms followed by their targeting to proteasome-mediated degradation[12][21][24]. The structural basis of client recognition appears to involve recognition of biochemical properties such as net charge, hydrophobicity patterns, or folding status rather than a strict consensus sequence, explaining the ability of HSP90-1 to accommodate structurally and functionally diverse clients while maintaining specificity through cochaperone-mediated client selection.
Cochaperone Networks and Regulatory Complexity of HSP90-1 Function
The functional activity of HSP90-1 is fundamentally dependent on and regulated by an extensive network of cochaperone proteins that have been termed collectively as the "HSP90 chaperone machinery"[11][25][37][40]. These cochaperones function through multiple distinct mechanisms including modulation of HSP90-1 ATPase activity, direction of HSP90-1 toward specific client proteins, stabilization of the open conformation required for client loading, and facilitation of client activation following maturation[11][25]. The tetratricopeptide repeat (TPR)-containing cochaperones represent the most extensively characterized family of HSP90-1 regulators, with multiple members identified in Arabidopsis that exhibit distinct interaction specificities and functional roles. These TPR-containing cochaperones including Hop (Sti1), FKBP52, and plant homologs recognize and bind to the conserved C-terminal MEEVD motif of HSP90-1, establishing a primary interaction interface[37][40].
The cochaperone Hop (Sti1 in yeast) plays a critical role in recruiting client proteins from Hsp70 to HSP90-1 and in inhibiting the ATPase activity of HSP90-1 to stabilize the open conformation required for client loading[28][37][40]. The structural analysis of Hop complexed with HSP90-1 revealed that Hop adopts a "butterfly-like" quaternary structure with multiple interaction surfaces on HSP90-1 including the C-terminal MEEVD motifs and additional sites within the C-terminal and middle domains[40]. The Hop-HSP90-1 complex is stabilized in the nucleotide-free, open state of HSP90-1, but ATP binding induces conformational changes in HSP90-1 that weaken the Hop-HSP90-1 interaction, allowing the progression toward the closed conformation during the ATPase cycle[40]. The interaction between Hop and HSP90-1 exhibits nucleotide dependence, with Hop binding preferentially to the ATP-free form of HSP90-1 and being displaced upon ATP binding[40]. The dynamic nature of Hop binding to HSP90-1 enables the cyclical recruitment and release of Hop during successive rounds of the ATPase cycle, thereby establishing directionality to the chaperone cycle progression[40].
The cochaperone p23 (Sba1 in yeast, which functions as an acidic domain-containing co-chaperone) represents another critical component of the HSP90-1 regulatory network that exhibits distinct functional roles compared to Hop[25][37]. The p23 cochaperone stabilizes the ATP-bound, closed conformation of HSP90-1, thereby promoting extended residence times in conformational states that facilitate client protein maturation[25]. Unlike Hop, which inhibits the ATPase activity of HSP90-1, p23 exhibits weak interactions with HSP90-1 and does not substantially affect the ATPase rate. Rather, p23 appears to stabilize particular conformational intermediates along the ATPase-driven cycle, effectively "clamping" the protein in conformations conducive to productive client interactions[25]. The binding of p23 to HSP90-1 is modulated by the nucleotide status, with p23 exhibiting enhanced binding to the ATP-bound closed conformation[25]. The cochaperone Aha1 (Activator of Hsp90 ATPase) exhibits an opposite functional role compared to p23 and Hop, promoting the ATPase activity of HSP90-1 and accelerating the rate of the conformational cycle[25].
The SGT1 (Suppressor of G2 transition) family of cochaperones represents plant-specific regulators of HSP90-1 that have been implicated in diverse developmental and immune functions[14][20][33]. SGT1 contains a conserved SGS (SGT1-specific) domain and tetratricopeptide repeat domains that mediate interactions with both HSP90-1 and client proteins[33]. The HSP90-1-SGT1 complex plays particularly critical roles in mediating disease resistance responses triggered by R proteins, with studies demonstrating that deletion of SGT1 compromises the function of multiple R protein-mediated disease resistance pathways[33]. SGT1 also associates with HSP90-1 in the context of auxin signaling, with the HSP90-1-SGT1 chaperone system required for the temperature-dependent stabilization of the TIR1 auxin receptor[24]. The interaction between SGT1 and HSP90-1 appears to be modulated by the ATPase state of HSP90-1, with SGT1 exhibiting differential binding affinities to distinct nucleotide-bound forms[25].
The recruitment cochaperone Cdc37 represents a kinase-specific cochaperone with demonstrated essential roles in HSP90-1-mediated maturation of protein kinase clients[9][25][28]. Cdc37 specifically recognizes kinase substrates through an unfoldase activity that partially unfolds kinases to expose surfaces competent for HSP90-1 binding[9]. The Cdc37-HSP90-1 interaction is stabilized through multiple contact points involving the N-terminal ATP-binding domain of HSP90-1 and the TPR domains of Cdc37[9][25]. The cochaperone Cdc37 undergoes phosphorylation at specific residues including S13 (by CK2), Y298 (by tyrosine kinases), and S339 (by Ulk1) that modulate its interaction with protein kinase clients and its association with HSP90-1[9]. The phosphorylation events on Cdc37 establish a molecular mechanism for temporal and spatial regulation of kinase client maturation by the HSP90-1-Cdc37 complex[9].
The RAR1 (Required for Mla12 Resistance) protein represents another plant-specific cochaperone that associates with HSP90-1 and participates in disease resistance signaling[20][33]. RAR1 contains zinc-binding CHORD domains that mediate protein-protein interactions and associates with both HSP90-1 and SGT1 to form a tripartite complex critical for R protein function[20][33]. The structural analysis revealed that the CHORD-I domain of RAR1 interacts directly with the N-terminus of HSP90-1, specifically with the ATPase domain containing the ATP-binding pocket[33]. The HSP90-1-RAR1-SGT1 complex has been demonstrated to associate with disease resistance proteins and to modulate their stability and signaling activities through mechanisms that remain to be fully elucidated[20][33].
The cochaperone TWD1 (Twisted Dwarf 1, also known as Fkbp42) represents a plant-specific TPR-containing cochaperone that associates with HSP90-1 in the context of regulating plasma membrane auxin transporters[35]. The TWD1-HSP90-1 chaperone system stabilizes ABCB-type auxin transporters at the plasma membrane, a function distinct from the HSP90-1-SGT1 regulation of the nuclear TIR1 receptor[35]. The interaction between TWD1 and HSP90-1 is established through the conserved C-terminal MEEVD motif of HSP90-1 and the TPR domain of TWD1, and confocal imaging revealed nearly perfect colocalization of HSP90-1 and TWD1 in root tissues[35]. The TWD1-HSP90-1 complex exhibits differential preferences for distinct ABCB isoforms, suggesting that cochaperones can provide client specificity through selective recognition mechanisms[35].
Post-translational modifications of HSP90-1 itself represent an additional layer of regulation controlling its chaperone function and client interactions. HSP90-1 undergoes phosphorylation at multiple residues in response to diverse stimuli, with these modifications modulating interaction profiles with clients and cochaperones[9][30]. The phosphorylation of HSP90-1 at Y197 by non-receptor tyrosine kinases recruited through Cdc37 specifically regulates the interaction between HSP90-1 and Cdc37, thereby affecting the chaperoning of kinase clients while leaving non-kinase clients unaffected[9]. Additional post-translational modifications of HSP90-1 potentially include acetylation, S-nitrosylation, and ubiquitination, though these have been less thoroughly characterized in plants compared to mammalian systems[9]. The phosphorylation-mediated regulation of HSP90-1 provides a mechanism for coupling HSP90-1 chaperone activity to cell signaling events, enabling HSP90-1 to integrate diverse cellular signals and respond dynamically to changing physiological conditions.
Role of HSP90-1 in Developmental Processes and Morphogenesis
The HSP90-1 protein plays essential roles in multiple developmental processes in Arabidopsis through its function as a molecular chaperone for key regulators of growth and differentiation. The involvement of HSP90-1 in early embryonic development is evidenced by the dramatic increase in HSP90-1 transcript levels just before embryo maturation, with expression becoming most prominent in developing cotyledons[2][49]. The high expression of HSP90-1 during late embryonic stages suggests critical roles in the preparation of tissues for postembryonic growth and suggests that HSP90-1 may be involved in the maturation and stabilization of proteins required for the transition from embryonic to postembryonic development. The T-DNA insertion mutant alleles and conditional knockdown approaches have revealed that HSP90-1 is required for proper embryo development, as complete loss of HSP90 function during embryogenesis results in embryonic lethality[14].
The role of HSP90-1 in hypocotyl elongation during skotomorphogenesis (seedling development in darkness) and thermomorphogenesis (growth at elevated ambient temperatures) represents a well-characterized developmental function of this chaperone[15][18]. In Arabidopsis, HSP90-1 participates in light and temperature signaling through its chaperone activity toward the transcription factor PHYTOCHROME-INTERACTING FACTOR 4 (PIF4), which promotes hypocotyl elongation in response to darkness and high ambient temperature[15]. When HSP90 activities are impaired by knockdown of HSP90 expression or application of the HSP90-specific inhibitor geldanamycin, the expression levels and protein abundance of PIF4 markedly decrease[15]. Furthermore, HSP90-1 interacts with and destabilizes the EARLY FLOWERING 3 (ELF3) protein, and this destabilization is modulated by the CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1) protein[15][18]. HSP90-1 enhances the interaction between COP1 and ELF3, thereby reducing the functional effects of ELF3 on PIF4 and ultimately modulating hypocotyl elongation during both skotomorphogenesis and thermomorphogenesis[15]. The HSP90-1-mediated regulation of the COP1-ELF3-PIF4 module represents an elegant example of how a molecular chaperone can be integrated into developmental regulatory networks to modulate environmental responses during postembryonic growth.
The critical role of HSP90-1 in root development has been demonstrated through studies examining the effects of HSP90 inhibition on root architecture and gravitropism[52][57]. HSP90-1 affects the asymmetrical distribution of the PIN1 auxin efflux transporter at the plasma membrane and influences PIN1 expression in the distal cell niches of the root meristem[52]. Depletion of HSP90-1 through genetic knockouts or pharmacological inhibition distorts the polar distribution of auxin, resulting in highly impaired acropetal (root-to-shoot) auxin transport[52]. This disruption of auxin transport leads to defective root gravitropism and compromised lateral root formation[52]. The essential role of HSP90-1 in auxin homeostasis is evident from early development, as HSP90-1 depletion affects embryo development and the pattern formation of veins in cotyledons through disruption of PIN1 distribution and auxin signaling[52]. The mechanism by which HSP90-1 regulates PIN1 distribution likely involves direct chaperoning of PIN1 and related ABCB transporters, with HSP90-1 working in conjunction with the cochaperone TWD1 to stabilize these proteins at the plasma membrane[35][52].
The involvement of HSP90-1 in seed germination and seedling vigor has been established through studies examining seed germination under normal and stress conditions. Plants overexpressing HSC70-1 or with reduced HSP90.2 activity exhibit hypersensitivity to abscisic acid (ABA) in seed germination assays, suggesting that HSP90-1 chaperone complexes with distinct substrates tune tissue-specific responses to ABA and other biotic and abiotic stimuli[30]. The regulation of seed germination by HSP90-1 likely involves chaperoning of signaling proteins in the ABA response pathway, though the specific client proteins and mechanisms remain to be fully characterized.
HSP90-1 in Environmental Stress Response and Adaptation
The HSP90-1 protein plays central roles in mediating plant responses to diverse environmental stresses through multiple mechanisms including direct chaperoning of stress-responsive proteins and modulation of stress signal transduction pathways. The expression of HSP90-1 is dramatically elevated in response to heat stress, with transcript levels increasing rapidly following exposure to elevated temperatures and remaining elevated during extended heat shock[2][14]. This heat-inducible expression pattern is mediated by heat shock transcription factors (Hsfs) that recognize heat shock elements (HSEs) in the HSP90-1 promoter region. The molecular chaperone Hsf1, a well-established master regulator of the heat shock response, is itself a client of HSP90-1, creating a regulatory feedback loop in which HSP90-1 both responds to heat stress and regulates its own expression through chaperoning of Hsf1[31][34]. Studies with cells lacking Hsp90 demonstrated that Hsp90 binding keeps Hsf1 inactive under non-stressed conditions by titrating Hsf1 monomers and preventing their trimerization and nuclear accumulation[31]. During heat stress, reduced Hsp90 binding to Hsf1 allows the accumulation of Hsf1 trimers and their transit to the nucleus, where they bind heat shock elements and activate transcription of heat shock protein genes including HSP90-1[31].
The stabilization of temperature-sensitive proteins represents another mechanism through which HSP90-1 contributes to heat tolerance and temperature adaptation. The role of HSP90-1 in regulating temperature-dependent seedling growth is mediated through its chaperoning of the TIR1 auxin receptor, with HSP90 levels increasing dramatically in response to temperature elevation and stabilizing TIR1 at higher temperatures[16][24]. This HSP90-1-dependent mechanism allows plants to integrate environmental temperature cues with developmental responses through modulation of auxin signaling, thereby enabling proper responses to ambient temperature changes[16][24]. The temperature-dependent stabilization of TIR1 by HSP90-1 occurs through a mechanism involving the co-chaperone SGT1b, which associates with TIR1 in the context of the HSP90-1 chaperone complex[24].
The role of HSP90-1 in mediating responses to heavy metal stress has been demonstrated through the observation that HSP90-1 transcript levels increase substantially following treatment of plants with heavy metals including cadmium and other toxic ions[2]. The specific mechanisms by which HSP90-1 contributes to heavy metal tolerance remain to be fully characterized but likely involve chaperoning of metal-responsive proteins or facilitation of detoxification pathways. The expression of HSP90-1 in response to arsenic exposure has been documented, suggesting roles in managing stress responses to metalloid toxins[1].
HSP90-1 in Disease Resistance and Immune Signaling
The HSP90-1 protein plays critical roles in plant defense responses through its association with resistance (R) proteins that mediate effector-triggered immunity to pathogenic microorganisms. The identification of HSP90-1 as a critical component of R-protein-mediated immunity came from genetic screens that revealed that specific mutations in HSP90-1 (specifically in the HSP90.2 isoform) compromise the function of certain R proteins such as RPM1 without affecting the function of other disease resistance proteins[20]. This finding demonstrated that HSP90-1 is not simply a general requirement for cell viability but rather functions in a selective manner toward specific R protein clients[20]. The RPM1 resistance protein represents a nucleotide-binding leucine-rich repeat (NB-LRR) type R protein that activates disease resistance responses following recognition of the bacterial Type III secretion system effector protein AvrRpm1[20]. The association between HSP90-1 and RPM1 has been demonstrated through co-immunoprecipitation analyses, confirming that these proteins form a complex within plant cells[20].
The cochaperone RAR1 and SGT1 associate with HSP90-1 in the context of R protein function, with studies demonstrating that all three proteins interact in plant cells and are required for full RPM1-mediated disease resistance[20][33]. The structural analysis revealed that RAR1 contains two zinc-binding CHORD domains that interact with distinct regions of HSP90-1, and that SGT1 associates with RAR1 through its CS and TPR domains[33]. The RAR1-SGT1-HSP90-1 complex functions to stabilize R proteins such as RPM1, preventing their degradation by the proteasome and maintaining them at functional levels required for effective defense responses[20][33]. The Arabidopsis protein RPS2, another nucleotide-binding leucine-rich repeat type R protein, was similarly shown to require HSP90-1 for full resistance against the bacterial pathogen Pseudomonas syringae carrying the avrRpt2 effector[33]. The specific HSP90-1 isoform is rapidly induced upon pathogen challenge, demonstrating that R-protein-dependent immunity recruits HSP90-1 as a component of the defense response[33].
The role of HSP90-1 in disease resistance extends beyond NB-LRR type R proteins to other R protein classes and also to basal immunity responses triggered by recognition of pathogen-associated molecular patterns (PAMPs). Studies in rice demonstrated that the RAR1-HSP90-RAC1 complex participates in innate immune responses to both compatible and incompatible pathogenic races of the blast fungus Magnaporthe grisea and the bacterial blight pathogen Xanthomonas oryzae[36]. The Rac1 GTPase, a Rho-family small GTPase, forms a complex with RAR1, HSP90, and Hsp70 that promotes PAMP-triggered immune responses, with the Rac1-RAR1-HSP90 complex essential for proper functioning of innate immunity[36]. The functional link between HSP90-1 and Rac1 represents an intersection between developmental signaling and immunity signaling in plants, with the Rac1-RAR1-HSP90 complex modulating both developmental processes and immune responses[36].
The role of HSP90-1 in regulating disease resistance proteins includes modulation of their subcellular localization and conformational states required for activation. The involvement of HSP90-1 in nuclear-cytoplasmic trafficking of R proteins has been suggested through studies demonstrating that HSP90 associates with R proteins and influences their localization, though direct evidence for HSP90-1-mediated nuclear transport of R proteins remains limited[19]. The stabilization of R proteins by HSP90-1 appears to involve maintenance of properly folded conformations that are competent for recognition of pathogen effectors and subsequent signal transduction.
HSP90-1 as a Capacitor of Genetic Variation and Evolutionary Adaptation
One of the most profound and paradigm-shifting functions of HSP90-1 is its role as a "capacitor" of genetic variation, meaning its ability to buffer the phenotypic expression of cryptic genetic variants that accumulate in populations under normal conditions[32][55][58]. This function was first discovered in metazoan systems through observations that perturbation of HSP90 function in Drosophila flies revealed phenotypic variation absent in the wild-type population, and subsequent work in plants revealed that HSP90-1 performs an analogous function in buffering genetic variation in Arabidopsis[32][55]. The mechanisms underlying HSP90-1's capacitor function involve its molecular chaperone activity: HSP90-1 can accommodate and stabilize proteins bearing mutations that would otherwise compromise their folding or stability, thereby masking the phenotypic effects of these mutations until HSP90-1 levels or activity are reduced[21][32][55].
The evidence that HSP90-1 buffers genetic variation in Arabidopsis comes from studies employing populations with constitutively reduced HSP90 levels that were subsequently used for quantitative genetic mapping[32]. These studies demonstrated that HSP90-dependent natural variation is present at high frequencies in A. thaliana, with the expectation that at least one HSP90-dependent polymorphism will affect nearly every quantitative trait in progeny of different wild lines[32]. The analysis of HSP90-buffered variation affecting multiple life-history traits revealed that nine independent genomic regions displayed significant HSP90-buffering effects for at least one trait, with an average expectation of 0.5 HSP90-dependent quantitative trait loci per chromosome per trait[32]. The phenotypic effects of HSP90-dependent variants were particularly pronounced for certain traits including leaf length, plant height, seed mass, and flowering time[32].
A specific example of HSP90-1's capacitor function comes from studies of the TIR1 auxin receptor with the tir1-1 mutation bearing the G147D amino acid substitution[21]. This mutation causes a cryptic auxin response defect that is normally masked by HSP90-1 buffering; however, when HSP90-1 activity is reduced through inhibition with geldanamycin or through HSP90-1 knockdown, the phenotypic consequences of the tir1-1 mutation become visible[21]. The mechanism appears to involve HSP90-1 providing enhanced chaperoning activity that allows the mutated TIR1G147D protein to recover functional properties through assisted folding, thereby buffering the mutation[21]. The tir1-10 null mutant, by contrast, displays obvious root growth defects even in the presence of functional HSP90-1, because HSP90-1 cannot generate function from a completely absent protein[21].
The evolutionary implications of HSP90-1's capacitor function are profound, as this mechanism provides a molecular basis for understanding how cryptic genetic variation can be exposed under stress conditions, thereby increasing the effective mutation rate available to natural selection[32][55][58]. The hypothesis proposes that under environmental stress conditions where HSP90-1 function is compromised—for example through heat shock or through competition for HSP90-1 chaperoning resources by stress-denatured proteins—the buffering capacity of HSP90-1 is reduced, and previously silent genetic variants become phenotypically expressed[55]. This "genetic assimilation" mechanism allows natural selection to act on previously cryptic variation, thereby enabling rapid evolutionary adaptation in response to environmental change[32][55]. Studies in Mexican cavefish (Astyanax mexicanus) have demonstrated that HSP90 buffers developmental variation in eye size, and that under stress conditions, this buffering is compromised, allowing variation in developmental processes to be expressed and selected upon[58]. This mechanism provides a molecular explanation for the observation that periods of environmental stress are often accompanied by morphological diversification and rapid evolutionary change.
Circadian Clock Regulation and HSP90-1-Dependent Temporal Organization
The HSP90-1 protein plays unique and central roles in the plant circadian clock through its chaperoning of the F-box protein ZEITLUPE (ZTL), a key component of the circadian oscillator[51]. The circadian clock represents a system of autoregulatory feedback loops that maintain approximate 24-hour periodicity of physiological processes in response to environmental light-dark cycles and temperature changes. The ZTL protein functions as a component of an SCF ubiquitin ligase complex that targets the PRR5 (PSEUDO-RESPONSE REGULATOR 5) protein for proteasomal degradation as part of a negative feedback loop that generates circadian oscillations[51]. The discovery that HSP90-1 is essential for ZTL function came from observations that HSP90-1 inhibition by geldanamycin or RNAi-mediated knockdown reduces ZTL protein levels and lengthens the circadian period, phenotypes identical to those of ztl loss-of-function mutants[51]. The HSP90-1-mediated stabilization of ZTL appears to involve holdase activity that prevents ZTL aggregation and promotes its proper folding and assembly into functional complexes[51].
The in vitro holdase assay, typically used to demonstrate chaperone activity with general substrate proteins, demonstrated that ZTL can be effectively bound by HSP90-1 and its aggregation prevented in an ATP-dependent manner[51]. The functional consequence of HSP90-1-mediated ZTL maturation is that the levels of the proteolytic targets of the SCF(ZTL) complex, specifically TOC1 and PRR5, are elevated when HSP90-1 is inhibited, confirming that HSP90-1 acts upstream of the SCF(ZTL) complex in the circadian circuit[51]. Unlike metazoan circadian clock systems where HSP90 functions at the periphery of the clock network, HSP90-1 in plants functions within the core oscillator itself, representing a unique and more central role for this molecular chaperone in maintaining temporal organization[51]. The integration of HSP90-1 into the core circadian oscillator positions the plant circadian clock to directly sense environmental stresses or changes in cellular proteostasis that affect HSP90-1 availability, thereby allowing the clock to adjust its period in response to stress conditions affecting protein homeostasis.
Evolutionary Conservation and Divergence of HSP90-1 Function
The HSP90-1 protein exhibits remarkable conservation across eukaryotic organisms, with the Arabidopsis HSP90-1 sharing over 80% amino acid sequence identity with mammalian and yeast HSP90 homologs[14][53]. This high degree of sequence conservation reflects the essential nature of HSP90 function and the ancient evolutionary origins of this protein family, with HSP90 orthologs recognizable in archaeal and eubacterial lineages[14]. However, recent evolutionary studies have revealed that despite this high sequence identity, significant functional divergence has occurred between prokaryotic and eukaryotic HSP90 proteins, particularly with respect to their conformational dynamics and regulatory complexity[53].
The comparison of yeast and human HSP90 proteins revealed that while the overall conformational transitions coupled to the ATPase cycle are conserved, the cycle timing and conformational dynamics differ substantially[53]. Human HSP90 is characterized by broad ensembles of conformational states that are sampled irrespective of the absence or presence of ATP, whereas yeast HSP90 displays more restricted conformational sampling with more deterministic ATP-dependent changes[53]. The molecular basis for these evolutionary differences was traced to two residues within conserved structural elements that trigger structural changes in response to ATP binding[53]. The exchange of these two amino acid residues between human and yeast Hsp90 proteins was sufficient to swap the ATPase rate and conformational transition characteristics, demonstrating that evolution of HSP90 function has involved fine-tuning of specific amino acids that modulate protein dynamics rather than large-scale architectural changes[53].
The evolutionary adaptations of HSP90 toward increased conformational flexibility and dynamics in higher eukaryotes appear to correlate with an expanded repertoire of cochaperones and client proteins in these organisms[53]. The increased conformational heterogeneity of eukaryotic HSP90 proteins may reflect adaptation to accommodate an increased diversity of client proteins and to integrate signals from a more complex suite of cochaperone regulators[53]. The plant HSP90-1 exhibits intermediate characteristics between prokaryotic HtpG and mammalian HSP90 proteins, suggesting that the evolution of HSP90 function proceeded gradually toward increased conformational complexity and regulatory sophistication[8][53].
The expansion of the HSP90 family in plants compared to bacteria represents another aspect of evolutionary adaptation, with Arabidopsis encoding seven HSP90 isoforms compared to a single HtpG gene in E. coli[2][14]. The subcellular compartmentalization of HSP90 function through targeting of distinct isoforms to different cellular compartments (cytoplasm, chloroplast, mitochondria, endoplasmic reticulum) represents an evolutionary innovation in plants that allows specialization of chaperone function for compartment-specific proteostasis requirements[2]. The cytoplasmic HSP90-1 and its three cytoplasmic paralogs exhibit distinct tissue-specific expression patterns and stress-responsiveness, suggesting that gene duplication followed by regulatory divergence has partitioned HSP90 function among specialized isoforms with distinct developmental and stress-response roles.
Conclusion
The HSP90-1 protein represents a paradigmatic example of a multifunctional molecular chaperone whose seemingly simple biochemical function—ATP-dependent protein folding and stabilization—belies its extraordinary complexity and centrality to diverse developmental and physiological processes in Arabidopsis thaliana. Through its capacity to interact with structurally and functionally diverse client proteins in an ATP-dependent manner, HSP90-1 has become integrated into developmental regulatory networks controlling hypocotyl elongation, root growth, seed germination, and vascular patterning. The chaperoning of key signal transduction proteins including kinases, hormone receptors, and resistance proteins by HSP90-1 positions this molecular chaperone as a critical integrator of environmental signals including temperature, light, and pathogenic challenge into appropriate physiological responses. The HSP90-1 protein achieves remarkable specificity in client protein recognition through cooperative interactions with an extensive network of cochaperones including the plant-specific regulators SGT1, RAR1, and TWD1, which collectively establish a sophisticated regulatory system for selective client maturation and activation.
Perhaps most intriguingly, HSP90-1 functions as a capacitor of genetic variation, buffering the phenotypic expression of cryptic mutations and thereby masking genetic diversity under normal conditions while allowing this variation to be revealed under stress conditions when HSP90-1 function is compromised[32][55][58]. This capacity positions HSP90-1 as a potential evolutionary innovation that accelerates adaptation to environmental change by enabling the rapid phenotypic expression of accumulated cryptic genetic variants when environmental stress compromises HSP90-1 function. The integration of HSP90-1 into the core circadian oscillator reveals an additional layer of complexity, whereby the proteostasis machinery directly influences temporal organization of physiological processes. Future investigations leveraging structural biology, single-molecule imaging, quantitative proteomics, and evolutionary comparative approaches promise to further illuminate the precise molecular mechanisms through which HSP90-1 achieves its remarkable functional versatility and physiological importance in plant development and stress adaptation.
Citations
- https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=835341
- https://pmc.ncbi.nlm.nih.gov/articles/PMC434405/
- https://pubmed.ncbi.nlm.nih.gov/9465043/
- https://www.uniprot.org/uniprotkb/P27323/entry
- https://www.uniprot.org/uniprotkb/P27323/publications
- https://elifesciences.org/articles/102666
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1170812/
- https://www.nature.com/articles/s41467-017-02711-w
- https://www.nature.com/articles/s41467-025-61962-0
- https://pubmed.ncbi.nlm.nih.gov/15763706/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2810645/
- https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.17528
- https://pubmed.ncbi.nlm.nih.gov/23443089/
- https://pubmed.ncbi.nlm.nih.gov/36707919/
- https://febs.onlinelibrary.wiley.com/doi/10.1002/1873-3468.13499
- https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.18776
- https://pmc.ncbi.nlm.nih.gov/articles/PMC42943/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC275404/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5501234/
- https://www.pnas.org/doi/10.1073/pnas.91.1.340
- https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.12760
- https://www.nature.com/articles/ncomms10269
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4838542/
- https://www.pnas.org/doi/10.1073/pnas.96.4.1297
- https://pmc.ncbi.nlm.nih.gov/articles/PMC379275/
- https://pubmed.ncbi.nlm.nih.gov/11805114/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3135925/
- https://elifesciences.org/articles/88658
- https://www.pnas.org/doi/10.1073/pnas.0712210105
- https://www.pnas.org/doi/10.1073/pnas.2033934100
- https://pubmed.ncbi.nlm.nih.gov/38059913/
- https://www.nature.com/articles/s41467-025-63780-w
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2217650/
- https://www.pnas.org/doi/10.1073/pnas.1106261108
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5078872/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11122155/
- https://pubmed.ncbi.nlm.nih.gov/18485364/
- https://www.picard.ch/downloads/Hsp90interactors.pdf
- https://pubs.acs.org/doi/10.1021/acs.jmedchem.4c01048
- https://www.pnas.org/doi/10.1073/pnas.1000916107
- https://pubmed.ncbi.nlm.nih.gov/39652595/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3546657/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC302038/
- https://journals.plos.org/plosgenetics/article?id=10.1371%2Fjournal.pgen.1011508
- https://pubmed.ncbi.nlm.nih.gov/15582930/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5070531/
- https://pubmed.ncbi.nlm.nih.gov/21949396/
- https://www.nature.com/articles/s41467-024-52995-y
- https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.15518
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6281791/
- https://pubmed.ncbi.nlm.nih.gov/29091774/
- https://pubmed.ncbi.nlm.nih.gov/34086995/
- https://www.nature.com/articles/s41467-025-65027-0
- https://www.nature.com/articles/s41392-024-02080-z