Deep Research
Falcon
(BRI1-deep-research-falcon.md)
Falcon
(BRI1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: Arabidopsis thaliana BRI1 (BRASSINOSTEROID INSENSITIVE 1; UniProt O22476; At4g39400)
0) Target verification (required disambiguation)
The UniProt accession O22476 corresponds to Arabidopsis thaliana BRI1, a leucine-rich repeat receptor-like kinase (LRR-RLK) that functions as the canonical cell-surface brassinosteroid (BR) receptor and signals with SERK co-receptors (notably BAK1/AtSERK3). This identity, domain architecture (LRR ectodomain with an “island domain” plus intracellular kinase domain), and pathway placement are consistently described in authoritative review and primary literature, matching the provided UniProt description. (delesalle2024thecellsurface pages 24-28, wang2008sequentialtransphosphorylationof pages 1-2)
1) Key concepts and definitions (current understanding)
1.1 Brassinosteroids (BRs) and BR perception
Brassinosteroids are plant steroid hormones that regulate growth and development by being perceived at the plasma membrane (PM) by BRI1. Ligand binding (classically brassinolide, BL) is the initiating event that enables assembly/activation of a receptor complex and downstream phosphorylation signaling. (delesalle2024thecellsurface pages 20-24, wang2008sequentialtransphosphorylationof pages 1-2)
1.2 BRI1 as an LRR receptor kinase with an “island domain”
BRI1 is a cell-surface LRR-RLK whose ectodomain contains an island domain that participates directly in steroid binding; work on early BR signaling established that the island domain together with LRR22 forms the steroid-binding motif. (wang2008sequentialtransphosphorylationof pages 1-2)
1.3 Co-receptors and receptor-kinase complexes (SERKs/BAK1)
A core concept in BR signaling is that ligand binding promotes formation of an active receptor/co-receptor complex. In Arabidopsis, BRI1 forms a complex with a SERK family co-receptor, especially BAK1/AtSERK3, and the complex becomes active through sequential transphosphorylation between the two kinase domains. (delesalle2024thecellsurface pages 20-24, wang2008sequentialtransphosphorylationof pages 1-2)
1.4 Downstream signaling logic: RLCK relay → phosphatase → GSK3 inhibition → transcription factors
Activated BRI1 initiates signaling through receptor-proximal kinases (e.g., BSK1 and CDG1) that activate the phosphatase BSU1, which in turn inactivates the GSK3-like kinase BIN2. BIN2 normally represses BR output by phosphorylating the transcription factors BES1/BZR1; BR signaling shifts BES1/BZR1 toward nuclear accumulation and transcriptional reprogramming. (delesalle2024thecellsurface pages 20-24, kim2011thecdg1kinase pages 1-2)
2) Molecular function of BRI1 (enzyme activity, substrates, and specificity)
2.1 Catalytic activity: receptor kinase with dual specificity
BRI1 is a receptor kinase primarily classified as a Ser/Thr kinase, but it also displays tyrosine phosphorylation (dual-specificity behavior) as part of BR signaling regulatory logic. (delesalle2024thecellsurface pages 20-24)
2.2 Experimentally mapped phosphorylation sites and functional residues
A detailed LC–MS/MS study identified in vivo BRI1 phosphosites including S838, S858, T872, T880 (juxtamembrane), T982 (kinase domain), and S1168 (C-terminal region), with additional sites suggested in the activation loop. Functional mutagenesis showed that activation-loop residues T1049 and either S1044 or T1045 are essential for normal kinase activity/signaling, while T1039A and S1042A show intermediate effects. (wang2005identificationandfunctional pages 1-2, wang2005identificationandfunctional pages 10-11)
A notable quantitative enzyme-kinetic observation from this work is that the T872A mutation increases Vmax ~10-fold and reduces Km ~2-fold, strongly increasing catalytic efficiency relative to wild-type BRI1. (wang2005identificationandfunctional pages 13-15)
2.3 Direct downstream phosphotargets in early BR signaling
A key primary study established residue-level phosphorylation events linking BRI1 to downstream BR signaling:
- BRI1 phosphorylates BSK1 at Ser230, increasing BSK1 interaction with BSU1.
- BRI1 phosphorylates CDG1 at Ser44, Ser47, and Ser234, with Ser234 important for CDG1 activation.
- Activated CDG1 phosphorylates BSU1 at Ser764, enhancing BSU1 function.
- BSU1 promotes BIN2 inactivation by dephosphorylating BIN2 Tyr200.
(kim2011thecdg1kinase pages 1-2)
3) Ligand perception and receptor activation mechanism
3.1 Brassinolide binding and structural logic
Work on early BR signaling emphasizes that BL is perceived through the BRI1 ectodomain and that the island domain + LRR22 are integral to the steroid-binding motif. (wang2008sequentialtransphosphorylationof pages 1-2)
Recent structural/biophysical analysis of BR receptor complexes further supports the mechanism whereby BR binding creates a co-receptor (SERK) binding interface, enabling receptor–co-receptor association and activation. In this work, a BRI1–BL–SERK1 complex highlights polar contacts involving BL functional groups and residues including BRI1 Tyr642 and Lys601 and a co-receptor residue His62, illustrating how ligand chemistry supports co-receptor recruitment. (caregnato2025amechanisticframework pages 40-55)
3.2 Sequential transphosphorylation with BAK1/SERKs
A key mechanistic concept is sequential transphosphorylation: BR-dependent BRI1 activation precedes/permits association with BAK1, after which BRI1 and BAK1 transphosphorylate each other, enhancing signaling output. Conserved activation-loop residues (notably S1044 and T1049) are critical for BRI1 function in vitro and for BR signaling in planta. (wang2008sequentialtransphosphorylationof pages 1-2)
3.3 Relief of inhibition: BKI1 release
Receptor activation involves dissociation of an inhibitory factor, BKI1, triggered by tyrosine phosphorylation; this is an early step connecting ligand perception to propagation of the phosphorylation cascade. (qiu2025brassinosteroidsignalingdynamics pages 14-16, kim2011thecdg1kinase pages 1-2)
4) Subcellular localization, trafficking, and spatial organization
4.1 Primary site of action: the cell surface (plasma membrane)
A 2024 synthesis emphasizes that BR perception and key early signaling steps occur at the cell surface, with BRI1 functioning as a PM receptor whose abundance and activity are tightly controlled. (delesalle2024thecellsurface pages 5-8, delesalle2024thecellsurface pages 24-28)
4.2 Nanodomain organization
BRI1 is organized in PM nanodomains that are distinct from certain immune receptor nanodomains (e.g., FLS2). This spatial patterning is proposed to support signaling specificity, and BRI1-associated complexes can align with cortical microtubules. (delesalle2024thecellsurface pages 5-8)
4.3 Endocytosis and post-translational modification (PTM) control
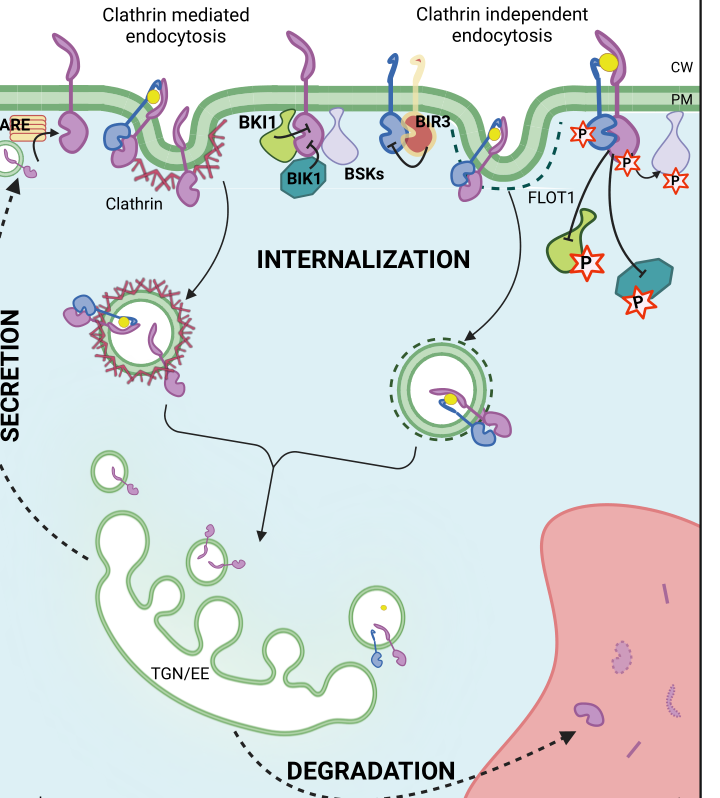
BRI1 undergoes regulated internalization by multiple routes including clathrin-mediated endocytosis (AP-2/T-PLATE-associated) and a clathrin-independent pathway (FLOT1-associated), followed by endosomal trafficking (e.g., via TGN/EE) and vacuolar degradation. (delesalle2024thecellsurface pages 5-8, delesalle2024thecellsurface media 76d771eb)
A major, current theme (highlighted in 2024) is that BRI1 PM abundance is controlled by competing PTMs:
- K63-linked polyubiquitination promotes endocytosis, involving PUB12/13 (E3 ligases) and UBC35/36 (E2 enzymes).
- Deubiquitination by UBP12/13 stabilizes BRI1 at the PM.
- SUMOylation stabilizes BRI1, whereas deSUMOylation by DeSi3a promotes internalization.
Moreover, mutation of 25 cytosolic lysines blocks internalization/degradation and causes hyperactivated BR signaling, consistent with extensive PTM control on overlapping Lys residues. (delesalle2024thecellsurface pages 5-8, delesalle2024thecellsurface media 76d771eb)
5) Pathways and biological processes impacted (focused on primary role)
5.1 Core BR signaling pathway
Integrating primary and review evidence, the core pathway can be summarized as:
1) BL binding to BRI1 ectodomain → 2) recruitment/activation with BAK1/SERKs (transphosphorylation) → 3) release of BKI1 inhibition → 4) phosphorylation of RLCKs (BSK1, CDG1) → 5) activation of BSU1 → 6) inactivation of BIN2 → 7) activation/nuclear accumulation of BES1/BZR1 → transcriptional regulation of BR-responsive genes. (delesalle2024thecellsurface pages 20-24, kim2011thecdg1kinase pages 1-2, wang2008sequentialtransphosphorylationof pages 1-2)
5.2 Quantitative and experimental condition examples
Primary experiments that illustrate BR-dependent complex formation used BRZ (biosynthesis inhibitor) followed by 0.1 mM BL for 90 minutes, where BL increased BAK1 co-immunoprecipitation with BRI1 and BRZ reduced it. (wang2005identificationandfunctional pages 2-3)
6) Recent developments and latest research (prioritizing 2023–2024)
6.1 2024 high-authority perspective: “cell surface is the place to be”
A 2024 Nature Plants review emphasizes that BR perception and response are organized at the PM and regulated by receptor trafficking, nanodomains, and PTM “codes” (ubiquitin/SUMO) that tune BRI1 abundance and signaling capacity. It also highlights gaps in understanding, such as how nanodomains are established/maintained and how PTM patterns are read to route receptors to distinct fates. (delesalle2024thecellsurface pages 5-8, delesalle2024thecellsurface pages 24-28)
6.2 2024 primary mechanism expansion: BON copines as facilitators of BRI1–SERK complex formation
A 2024 Nature Communications study identifies PM-associated copine/BON proteins as direct SERK interactors that promote effective BRI1–SERK interaction and transphosphorylation, adding a mechanistic layer upstream of the receptor complex activation step and pointing to new trait-engineering targets. (jing2024copineproteinsare pages 1-2)
7) Current applications and real-world implementations
7.1 Crop architecture engineering by tuning BRI1/BR perception
Although the target here is Arabidopsis BRI1, “BRI1” function is sufficiently conserved that applied work in crops leverages the same receptor logic. A 2025 peer-reviewed wheat study demonstrates that induced variation in TaBRI1 generates a compact, upright-leaf architecture, with quantitative phenotypes including:
- 57% and 27% shorter plants for two triple-mutant combinations, and one triple mutant at 34.3 cm vs 53.0 cm control (≈ 35% reduction).
- 56% reduction in flag-leaf angle (FLA) in a triple mutant.
- Trade-offs including 16% shorter spike length and reduced thousand-grain weight (e.g., 15.9% reduction in one genotype).
This illustrates a real-world implementation pattern: partial reduction of BR perception can improve canopy architecture but may impose yield penalties that breeding must mitigate. (gill2025inducedvariationin pages 6-9)
7.2 Chemical and hormonal implementations (growth and stress tolerance)
Recent reviews discuss the use of exogenous BR treatments (e.g., 2,4-epibrassinolide) and BR-pathway manipulation to improve stress tolerance and growth traits in horticultural contexts, and note that BR metabolism can modulate whether ligands fit the BRI1 pocket (thereby affecting perception). These strategies represent practical routes to modulate BRI1-pathway output without direct gene editing of BRI1. (gao2024brassinolidessignalingpathway pages 1-2)
8) Expert synthesis and analysis (authoritative-source interpretation)
The best-supported contemporary view is that BRI1 signaling output is determined not only by ligand availability and receptor–co-receptor phosphorylation chemistry, but also by spatial compartmentalization (nanodomains) and dynamic receptor trafficking regulated by PTMs (ubiquitin/SUMO). This implies that “functional annotation” for BRI1 should be framed as a system property: BRI1 is a kinase receptor whose core biochemical role is phosphorylation signaling, but whose biological function depends on regulated residence time at the PM, endosomal routing, and assembly with auxiliary proteins controlling complex formation (e.g., BON/SERKs). (delesalle2024thecellsurface pages 5-8, jing2024copineproteinsare pages 1-2)
9) Evidence map (summary table)
The following table provides a compact, citable map of key findings, residues, and mechanistic components.
| Topic | Key findings | Key molecules/residues | Evidence type | Year | DOI/URL |
|---|---|---|---|---|---|
| Identity/domains | Arabidopsis thaliana BRI1 (UniProt O22476; At4g39400) is consistently identified as a plasma-membrane leucine-rich repeat receptor-like kinase (LRR-RLK) with an extracellular LRR domain containing an island domain for steroid perception and an intracellular kinase domain; this matches the UniProt annotation. (delesalle2024thecellsurface pages 5-8, delesalle2024thecellsurface pages 24-28, wang2008sequentialtransphosphorylationof pages 1-2) | BRI1; LRR ectodomain; island domain; kinase domain | Review + primary | 2024, 2008 | https://doi.org/10.1038/s41477-024-01621-2 ; https://doi.org/10.1016/j.devcel.2008.06.011 |
| Ligand perception | BRI1 perceives brassinosteroids, especially brassinolide (BL), through the extracellular LRR/island domain; BL binding creates a SERK-binding surface and the island domain together with LRR22 directly forms the steroid-binding motif. Recent structural work further mapped BL-contacting chemistry important for co-receptor recruitment. (caregnato2025amechanisticframework pages 40-55, wang2008sequentialtransphosphorylationof pages 1-2, wang2005autoregulationandhomodimerization pages 1-2) | BL; island domain; LRR22; Tyr642; Lys601; His62 | Primary | 2025, 2008, 2005 | https://doi.org/10.1101/2025.08.08.669299 ; https://doi.org/10.1016/j.devcel.2008.06.011 ; https://doi.org/10.1016/j.devcel.2005.05.001 |
| Co-receptor complex | BR binding promotes BRI1 association with SERK co-receptors, especially BAK1/AtSERK3; the complex undergoes sequential transphosphorylation, and BRI1 can phosphorylate BAK1 before reciprocal activation further boosts signaling. BL also increases BAK1-GFP co-immunoprecipitation with BRI1, whereas BRZ reduces association. (qiu2025brassinosteroidsignalingdynamics pages 14-16, delesalle2024thecellsurface pages 20-24, wang2008sequentialtransphosphorylationof pages 1-2, wang2005identificationandfunctional pages 2-3) | BAK1/AtSERK3; SERK1; BRZ; BL | Review + primary | 2025, 2024, 2008, 2005 | https://doi.org/10.3390/ijms26104502 ; https://doi.org/10.1038/s41477-024-01621-2 ; https://doi.org/10.1016/j.devcel.2008.06.011 ; https://doi.org/10.1105/tpc.105.031393 |
| Kinase activity | BRI1 is a dual-specificity kinase: historically classified as Ser/Thr, but later shown to autophosphorylate on tyrosine as well. Autophosphorylation appears hierarchical (Ser > Thr > Tyr), and the juxtamembrane domain activates kinase output. (delesalle2024thecellsurface pages 24-28, delesalle2024thecellsurface pages 20-24) | Ser/Thr/Tyr phosphorylation; juxtamembrane domain | Review + primary | 2024, 2012, 2009 | https://doi.org/10.1038/s41477-024-01621-2 ; https://doi.org/10.3389/fpls.2012.00175 ; https://doi.org/10.1073/pnas.0810249106 |
| Kinase activity/phosphosites | In vivo phosphosites identified on BRI1 include S838, S858, T872, T880, T982, and S1168, with additional activation-loop sites detected by MS. Activation-loop residues are functionally critical: T1049 and S1044/T1045 are essential for kinase activity/signaling; T1039A and S1042A show intermediate rescue. T872A increased Vmax ~10-fold and lowered Km ~2-fold. (wang2005identificationandfunctional pages 10-11, wang2005identificationandfunctional pages 1-2, wang2005identificationandfunctional pages 13-15) | S838; S858; T872; T880; T982; S1168; T1039; S1042; S1044; T1045; T1049 | Primary | 2005 | https://doi.org/10.1105/tpc.105.031393 |
| Receptor regulation | BRI1 activation includes release of the inhibitor BKI1 via tyrosine phosphorylation; BKI1 dissociation is an early step linking receptor activation to downstream signaling. (qiu2025brassinosteroidsignalingdynamics pages 14-16, delesalle2024thecellsurface pages 24-28, kim2011thecdg1kinase pages 1-2) | BKI1; Tyr phosphorylation | Review + primary | 2025, 2024, 2011 | https://doi.org/10.3390/ijms26104502 ; https://doi.org/10.1038/s41477-024-01621-2 ; https://doi.org/10.1016/j.molcel.2011.05.037 |
| Early downstream signaling | Activated BRI1 phosphorylates RLCKs including BSK1 and CDG1; BSK1 is phosphorylated at Ser230, and CDG1 at Ser44, Ser47, and Ser234, with Ser234 important for CDG1 activation. (kim2011thecdg1kinase pages 1-2) | BSK1 Ser230; CDG1 Ser44/Ser47/Ser234 | Primary | 2011 | https://doi.org/10.1016/j.molcel.2011.05.037 |
| Core phosphorylation cascade | Activated CDG1 phosphorylates BSU1 at Ser764, enhancing BSU1-mediated dephosphorylation of BIN2 at Tyr200; BIN2 inactivation releases BES1/BZR1 to accumulate in the nucleus and regulate BR-responsive genes. (qiu2025brassinosteroidsignalingdynamics pages 14-16, delesalle2024thecellsurface pages 20-24, kim2011thecdg1kinase pages 1-2, wang2005autoregulationandhomodimerization pages 1-2) | BSU1 Ser764; BIN2 Tyr200; BES1; BZR1/BZR2 | Review + primary | 2025, 2024, 2011, 2005 | https://doi.org/10.3390/ijms26104502 ; https://doi.org/10.1038/s41477-024-01621-2 ; https://doi.org/10.1016/j.molcel.2011.05.037 ; https://doi.org/10.1016/j.devcel.2005.05.001 |
| Localization/nanodomains | BRI1 is primarily plasma-membrane localized and enriched in distinct PM nanodomains separate from FLS2; BRI1-BAK1-BIK1 complexes align with cortical microtubules, supporting spatially organized signaling. (delesalle2024thecellsurface pages 35-37, delesalle2024thecellsurface pages 5-8, delesalle2024thecellsurface media 76d771eb) | PM nanodomains; BIK1; cortical microtubules | Review | 2024 | https://doi.org/10.1038/s41477-024-01621-2 |
| Trafficking/endocytosis | BRI1 abundance is dynamically controlled by endocytosis and recycling. Internalization occurs via AP-2/T-PLATE-dependent clathrin-mediated endocytosis and a clathrin-independent FLOT1 pathway, followed by trafficking through TGN/EE and vacuolar degradation. (delesalle2024thecellsurface pages 5-8, delesalle2024thecellsurface media 76d771eb) | AP-2; T-PLATE; FLOT1; TGN/EE; vacuole | Review | 2024 | https://doi.org/10.1038/s41477-024-01621-2 |
| PTM control of receptor abundance | K63-polyubiquitination by PUB12/13 with E2s UBC35/36 promotes BRI1 endocytosis; deubiquitination by UBP12/13 stabilizes PM BRI1. Mutation of 25 cytosolic Lys residues abolishes internalization/degradation and causes hyperactive BR signaling. SUMOylation stabilizes BRI1, whereas deSUMOylation by DeSi3a promotes internalization. (delesalle2024thecellsurface pages 35-37, delesalle2024thecellsurface pages 5-8, delesalle2024thecellsurface media 76d771eb) | PUB12/13; UBC35/36; UBP12/13; 25 Lys residues; SUMO; DeSi3a | Review | 2024 | https://doi.org/10.1038/s41477-024-01621-2 |
| Functional/phenotypic evidence | Exogenous BR decreases root stele area in wild type, whereas perturbed BR signaling increases stele area; epidermis-specific BRI1 expression in bri1 mutants can rescue root growth and limit stele area. Recent chemical-biology assays used 100 nM BL in root growth inhibition tests (n = 30-35). (delesalle2024thecellsurface pages 16-20, caregnato2025amechanisticframework pages 40-55) | Root stele area; bri1 mutants; 100 nM BL; n=30-35 | Review + primary | 2024, 2025 | https://doi.org/10.1038/s41477-024-01621-2 ; https://doi.org/10.1101/2025.08.08.669299 |
Table: This table summarizes experimentally supported functional annotation evidence for Arabidopsis thaliana BRI1 (UniProt O22476), covering identity, ligand perception, receptor-complex formation, kinase activity, trafficking, and downstream signaling. It is useful as a compact evidence map linking molecular features to specific publications and residues.
10) Visual schematic evidence (cell-surface model)
A schematic model figure from the 2024 Nature Plants review summarizes (i) BR perception and BRI1–BAK1 complex formation at the PM, (ii) clathrin-mediated and clathrin-independent internalization routes, and (iii) ubiquitin/SUMO control of receptor dynamics and downstream signaling logic (BIK1, BIN2, BES1/BZR1). (delesalle2024thecellsurface media 76d771eb)
Key references (URLs and publication dates)
- Delesalle C, Vert G, Fujita S. The cell surface is the place to be for brassinosteroid perception and responses. Nature Plants (Feb 2024). https://doi.org/10.1038/s41477-024-01621-2 (delesalle2024thecellsurface pages 5-8)
- Jing T et al. Copine proteins are required for brassinosteroid signaling in maize and Arabidopsis. Nature Communications (Mar 2024). https://doi.org/10.1038/s41467-024-46289-6 (jing2024copineproteinsare pages 1-2)
- Kim T-W et al. The CDG1 kinase mediates brassinosteroid signal transduction from BRI1… to BSU1… and BIN2. Molecular Cell (Aug 2011). https://doi.org/10.1016/j.molcel.2011.05.037 (kim2011thecdg1kinase pages 1-2)
- Wang X et al. Sequential transphosphorylation of the BRI1/BAK1 receptor kinase complex… Developmental Cell (Aug 2008). https://doi.org/10.1016/j.devcel.2008.06.011 (wang2008sequentialtransphosphorylationof pages 1-2)
- Wang X et al. Identification and functional analysis of in vivo phosphorylation sites of BRI1. The Plant Cell (May 2005). https://doi.org/10.1105/tpc.105.031393 (wang2005identificationandfunctional pages 1-2)
- Gill MS et al. Induced variation in BRI1 confers a compact wheat architecture. BMC Plant Biology (May 2025). https://doi.org/10.1186/s12870-025-06762-w (gill2025inducedvariationin pages 6-9)
References
-
(delesalle2024thecellsurface pages 24-28): Charlotte Delesalle, Grégory Vert, and Satoshi Fujita. The cell surface is the place to be for brassinosteroid perception and responses. Nature Plants, 10:206-218, Feb 2024. URL: https://doi.org/10.1038/s41477-024-01621-2, doi:10.1038/s41477-024-01621-2. This article has 21 citations and is from a highest quality peer-reviewed journal.
-
(wang2008sequentialtransphosphorylationof pages 1-2): Xiaofeng Wang, Uma Kota, Kai He, Kevin Blackburn, Jia Li, Michael B. Goshe, Steven C. Huber, and Steven D. Clouse. Sequential transphosphorylation of the bri1/bak1 receptor kinase complex impacts early events in brassinosteroid signaling. Developmental cell, 15 2:220-35, Aug 2008. URL: https://doi.org/10.1016/j.devcel.2008.06.011, doi:10.1016/j.devcel.2008.06.011. This article has 668 citations and is from a highest quality peer-reviewed journal.
-
(delesalle2024thecellsurface pages 20-24): Charlotte Delesalle, Grégory Vert, and Satoshi Fujita. The cell surface is the place to be for brassinosteroid perception and responses. Nature Plants, 10:206-218, Feb 2024. URL: https://doi.org/10.1038/s41477-024-01621-2, doi:10.1038/s41477-024-01621-2. This article has 21 citations and is from a highest quality peer-reviewed journal.
-
(kim2011thecdg1kinase pages 1-2): Tae-Wuk Kim, Shenheng Guan, Alma L. Burlingame, and Zhi-Yong Wang. The cdg1 kinase mediates brassinosteroid signal transduction from bri1 receptor kinase to bsu1 phosphatase and gsk3-like kinase bin2. Molecular cell, 43 4:561-71, Aug 2011. URL: https://doi.org/10.1016/j.molcel.2011.05.037, doi:10.1016/j.molcel.2011.05.037. This article has 487 citations and is from a highest quality peer-reviewed journal.
-
(wang2005identificationandfunctional pages 1-2): Xiaofeng Wang, Michael B. Goshe, Erik J. Soderblom, Brett S. Phinney, Jason A. Kuchar, Jia Li, Tadao Asami, Shigeo Yoshida, Steven C. Huber, and Steven D. Clouse. Identification and functional analysis of in vivo phosphorylation sites of the arabidopsis brassinosteroid-insensitive1 receptor kinase. The Plant Cell, 17:1685-1703, May 2005. URL: https://doi.org/10.1105/tpc.105.031393, doi:10.1105/tpc.105.031393. This article has 492 citations.
-
(wang2005identificationandfunctional pages 10-11): Xiaofeng Wang, Michael B. Goshe, Erik J. Soderblom, Brett S. Phinney, Jason A. Kuchar, Jia Li, Tadao Asami, Shigeo Yoshida, Steven C. Huber, and Steven D. Clouse. Identification and functional analysis of in vivo phosphorylation sites of the arabidopsis brassinosteroid-insensitive1 receptor kinase. The Plant Cell, 17:1685-1703, May 2005. URL: https://doi.org/10.1105/tpc.105.031393, doi:10.1105/tpc.105.031393. This article has 492 citations.
-
(wang2005identificationandfunctional pages 13-15): Xiaofeng Wang, Michael B. Goshe, Erik J. Soderblom, Brett S. Phinney, Jason A. Kuchar, Jia Li, Tadao Asami, Shigeo Yoshida, Steven C. Huber, and Steven D. Clouse. Identification and functional analysis of in vivo phosphorylation sites of the arabidopsis brassinosteroid-insensitive1 receptor kinase. The Plant Cell, 17:1685-1703, May 2005. URL: https://doi.org/10.1105/tpc.105.031393, doi:10.1105/tpc.105.031393. This article has 492 citations.
-
(caregnato2025amechanisticframework pages 40-55): Alberto Caregnato, Houming Chen, Miroslav Kvasnica, Ulrich Hohmann, Jana Oklestkova, Karoll Ferrer, Larissa Broger, Ludwig A. Hothorn, Miroslav Strnad, and Michael Hothorn. A mechanistic framework for the recognition of chemically diverse brassinosteroids by bri1-family receptor kinases. bioRxiv, Aug 2025. URL: https://doi.org/10.1101/2025.08.08.669299, doi:10.1101/2025.08.08.669299. This article has 3 citations.
-
(qiu2025brassinosteroidsignalingdynamics pages 14-16): Riguang Qiu, Yan Zhou, and Juan Mao. Brassinosteroid signaling dynamics: ubiquitination-dependent regulation of core signaling components. International Journal of Molecular Sciences, 26:4502, May 2025. URL: https://doi.org/10.3390/ijms26104502, doi:10.3390/ijms26104502. This article has 2 citations.
-
(delesalle2024thecellsurface pages 5-8): Charlotte Delesalle, Grégory Vert, and Satoshi Fujita. The cell surface is the place to be for brassinosteroid perception and responses. Nature Plants, 10:206-218, Feb 2024. URL: https://doi.org/10.1038/s41477-024-01621-2, doi:10.1038/s41477-024-01621-2. This article has 21 citations and is from a highest quality peer-reviewed journal.
-
(delesalle2024thecellsurface media 76d771eb): Charlotte Delesalle, Grégory Vert, and Satoshi Fujita. The cell surface is the place to be for brassinosteroid perception and responses. Nature Plants, 10:206-218, Feb 2024. URL: https://doi.org/10.1038/s41477-024-01621-2, doi:10.1038/s41477-024-01621-2. This article has 21 citations and is from a highest quality peer-reviewed journal.
-
(wang2005identificationandfunctional pages 2-3): Xiaofeng Wang, Michael B. Goshe, Erik J. Soderblom, Brett S. Phinney, Jason A. Kuchar, Jia Li, Tadao Asami, Shigeo Yoshida, Steven C. Huber, and Steven D. Clouse. Identification and functional analysis of in vivo phosphorylation sites of the arabidopsis brassinosteroid-insensitive1 receptor kinase. The Plant Cell, 17:1685-1703, May 2005. URL: https://doi.org/10.1105/tpc.105.031393, doi:10.1105/tpc.105.031393. This article has 492 citations.
-
(jing2024copineproteinsare pages 1-2): Teng Jing, Yuying Wu, Yanwen Yu, Jiankun Li, Xiaohuan Mu, Liping Xu, Xi Wang, Guang Qi, Jihua Tang, Daowen Wang, Shuhua Yang, Jian Hua, and Mingyue Gou. Copine proteins are required for brassinosteroid signaling in maize and arabidopsis. Nature Communications, Mar 2024. URL: https://doi.org/10.1038/s41467-024-46289-6, doi:10.1038/s41467-024-46289-6. This article has 22 citations and is from a highest quality peer-reviewed journal.
-
(gill2025inducedvariationin pages 6-9): Manpartik S. Gill, Andrew L. Phillips, Danuše Tarkowská, John Addy, Patrycja Sokolowska, M. John Foulkes, Stephen Pearce, Stephen G. Thomas, and Peter Hedden. Induced variation in brassinosteroid insensitive 1 (bri1) confers a compact wheat architecture. BMC Plant Biology, May 2025. URL: https://doi.org/10.1186/s12870-025-06762-w, doi:10.1186/s12870-025-06762-w. This article has 2 citations and is from a peer-reviewed journal.
-
(gao2024brassinolidessignalingpathway pages 1-2): Yanlong Gao, Xiaolan Ma, Zhongxing Zhang, Xiaoya Wang, and Yanxiu Wang. Brassinolides signaling pathway: tandem response to plant hormones and regulation under various abiotic stresses. Horticulture Advances, Oct 2024. URL: https://doi.org/10.1007/s44281-024-00050-7, doi:10.1007/s44281-024-00050-7. This article has 24 citations.
-
(wang2005autoregulationandhomodimerization pages 1-2): Xuelu Wang, Xiaoqing Li, Jill Meisenhelder, Tony Hunter, Shigeo Yoshida, Tadao Asami, and Joanne Chory. Autoregulation and homodimerization are involved in the activation of the plant steroid receptor bri1. Developmental cell, 8 6:855-65, Jun 2005. URL: https://doi.org/10.1016/j.devcel.2005.05.001, doi:10.1016/j.devcel.2005.05.001. This article has 375 citations and is from a highest quality peer-reviewed journal.
-
(delesalle2024thecellsurface pages 35-37): Charlotte Delesalle, Grégory Vert, and Satoshi Fujita. The cell surface is the place to be for brassinosteroid perception and responses. Nature Plants, 10:206-218, Feb 2024. URL: https://doi.org/10.1038/s41477-024-01621-2, doi:10.1038/s41477-024-01621-2. This article has 21 citations and is from a highest quality peer-reviewed journal.
-
(delesalle2024thecellsurface pages 16-20): Charlotte Delesalle, Grégory Vert, and Satoshi Fujita. The cell surface is the place to be for brassinosteroid perception and responses. Nature Plants, 10:206-218, Feb 2024. URL: https://doi.org/10.1038/s41477-024-01621-2, doi:10.1038/s41477-024-01621-2. This article has 21 citations and is from a highest quality peer-reviewed journal.
Artifacts
Citations
- wang2008sequentialtransphosphorylationof pages 1-2
- delesalle2024thecellsurface pages 20-24
- wang2005identificationandfunctional pages 13-15
- caregnato2025amechanisticframework pages 40-55
- delesalle2024thecellsurface pages 5-8
- wang2005identificationandfunctional pages 2-3
- jing2024copineproteinsare pages 1-2
- gill2025inducedvariationin pages 6-9
- gao2024brassinolidessignalingpathway pages 1-2
- wang2005identificationandfunctional pages 1-2
- delesalle2024thecellsurface pages 24-28
- wang2005identificationandfunctional pages 10-11
- qiu2025brassinosteroidsignalingdynamics pages 14-16
- wang2005autoregulationandhomodimerization pages 1-2
- delesalle2024thecellsurface pages 35-37
- delesalle2024thecellsurface pages 16-20
- https://doi.org/10.1038/s41477-024-01621-2
- https://doi.org/10.1016/j.devcel.2008.06.011
- https://doi.org/10.1101/2025.08.08.669299
- https://doi.org/10.1016/j.devcel.2005.05.001
- https://doi.org/10.3390/ijms26104502
- https://doi.org/10.1105/tpc.105.031393
- https://doi.org/10.3389/fpls.2012.00175
- https://doi.org/10.1073/pnas.0810249106
- https://doi.org/10.1016/j.molcel.2011.05.037
- https://doi.org/10.1038/s41467-024-46289-6
- https://doi.org/10.1186/s12870-025-06762-w
- https://doi.org/10.1038/s41477-024-01621-2,
- https://doi.org/10.1016/j.devcel.2008.06.011,
- https://doi.org/10.1016/j.molcel.2011.05.037,
- https://doi.org/10.1105/tpc.105.031393,
- https://doi.org/10.1101/2025.08.08.669299,
- https://doi.org/10.3390/ijms26104502,
- https://doi.org/10.1038/s41467-024-46289-6,
- https://doi.org/10.1186/s12870-025-06762-w,
- https://doi.org/10.1007/s44281-024-00050-7,
- https://doi.org/10.1016/j.devcel.2005.05.001,
Manual
(BRI1-deep-research-manual.md)
Manual
(BRI1-deep-research-manual.md)Comprehensive Research Report on BRASSINOSTEROID INSENSITIVE 1 (BRI1) in Arabidopsis thaliana: A Gene Ontology Annotation-Focused Analysis
This comprehensive report synthesizes current experimental knowledge of the BRI1 gene in Arabidopsis thaliana, with emphasis on molecular function, cellular component, biological process, and evidence quality for Gene Ontology annotation curation. BRI1 encodes a leucine-rich repeat receptor-like kinase (LRR-RLK) that serves as the primary receptor for brassinosteroid hormones and plays fundamental roles in plant growth, development, and stress responses[2][14].
Molecular Function: Biochemical Activities and Binding Properties
Dual-Specificity Kinase Activity
BRASSINOSTEROID INSENSITIVE 1 exhibits dual-specificity kinase activity, phosphorylating substrates on both serine/threonine and tyrosine residues[3][43][46]. This unusual property distinguishes BRI1 from most plant receptor kinases, which are typically classified as serine/threonine kinases. Experimental evidence demonstrates that recombinant cytoplasmic domains of BRI1 autophosphorylate on tyrosine residues in addition to serine and threonine residues[43][46]. Mass spectrometry analysis identified specific tyrosine autophosphorylation sites: Tyr-831 in the juxtamembrane domain and Tyr-956 in kinase subdomain V[43][46]. The Tyr-956 residue proved essential for catalytic activity, as the Y956F mutant was completely inactive, preventing simple mutagenesis manipulation of this site[43][46]. In contrast, Tyr-831 was not essential for kinase activity but played important roles in brassinosteroid signaling in vivo, as demonstrated by transgenic complementation studies in bri1-5 mutant plants[43][46].
Liquid chromatography-tandem mass spectrometry analysis of BRI1 immunoprecipitated from brassinosteroid-treated Arabidopsis seedlings identified at least eleven sites of in vivo phosphorylation in the juxtamembrane, kinase domain, and C-terminal domains[3][14][37]. Many of these phosphorylation sites showed brassinosteroid-dependent regulation[3][14][37]. Functional characterization through biochemical and genetic analyses revealed that highly conserved kinase activation loop residues, specifically Ser-1044 and Thr-1049, were critical for kinase function in vitro and proper brassinosteroid signaling in vivo[14][37]. Multiple juxtamembrane and C-terminal residues were required for optimal substrate phosphorylation by the BRI1 kinase domain[14][37]. Structural analysis of phosphorylated BRI1 kinase domain crystals provided detailed three-dimensional insight into the kinase mechanism and identified phosphorylation sites including Thr-1039, Ser-1042, Ser-1044, and Ser-1060 in the activation loop[6][57].
The BRI1 kinase domain adopts a conformation similar to that observed in the IRAK-4 structure, and based on activation loop conformation, BRI1 has been suggested to function as a dual-specificity kinase[6][57]. Phosphorylation of Thr-1039, Ser-1042, and Ser-1060 likely affects activation loop orientation and thus BRI1 kinase activity and substrate interaction[6][57]. Consistently, plants carrying Thr-1039-Ala and Ser-1042-Ala mutations exhibited intermediate brassinosteroid signaling phenotypes[6][57]. A cluster of phosphorylation sites in the N-terminal extension of the N-lobe, including Thr-872, Thr-880, and Ser-887, were shown to negatively regulate BRI1 activity, as phosphorylation at these sites could disrupt essential interactions between the N-terminal extension and the conserved N-lobe region[6][57].
Brassinosteroid Binding Activity
BRI1 functions as the primary brassinosteroid receptor in Arabidopsis, directly binding the most active brassinosteroid brassinolide with high affinity[2][5][23]. Using biotin-tagged photoaffinity castasterone, researchers demonstrated that brassinosteroids directly bind to the extracellular domain of BRI1 in Arabidopsis and to recombinant BRI1[5][23]. The minimal brassinosteroid-binding region consists of a 70-amino acid island domain and the carboxy-terminal flanking leucine-rich repeat 22 (ID-LRR22)[5][17][23]. Brassinosteroid binding assays using recombinant BRI1 fragments indicated that this ID-LRR22 domain specifically binds brassinolide and castasterone with high affinity[5][17][23]. The crystal structure of the BRI1 extracellular domain revealed that the island domain forms a separate folded structure anchored to the inner surface of the leucine-rich repeat solenoid[17]. Brassinolide binds in a hydrophobic surface groove created between the island domain and LRR22, with hormone recognition occurring through an induced-fit mechanism involving stabilization of interdomain loops[17].
Structural studies demonstrated that brassinosteroids act as "molecular glue" to enhance association between BRI1 and its coreceptor BAK1[11][17]. Both BRI1 and BAK1 make direct contact with the brassinosteroid molecule, qualifying them as a coreceptor pair[2][17]. The brassinolide-binding site within the BRI1 ectodomain is located approximately five leucine-rich repeat units away from the plasma membrane surface, coinciding spatially with the tip of the BAK1 leucine-rich repeat domain[17][55]. This spatial arrangement facilitates hormone-induced heterooligomerization of the receptor and coreceptor[17][55]. Quantitative measurements using analytical ultracentrifugation confirmed that brassinolide-bound BRI1 ectodomain behaves as a monomer in solution, while addition of the SERK1 ectodomain results in formation of a very stable heterodimer[57].
Substrate Phosphorylation and Downstream Signaling
BRI1 phosphorylates multiple substrate proteins to transduce brassinosteroid signals. The activated BRI1 receptor phosphorylates receptor-like cytoplasmic kinases including brassinosteroid-signaling kinase 1 (BSK1), BSK2, and BSK3[8]. In vitro kinase assays using recombinant proteins demonstrated that BRI1 directly phosphorylates BSK1, BSK2, and BSK3[8]. Extended analysis showed that BRI1 phosphorylates additional BSK family members including BSK5, BSK6, BSK8, and BSK11 in vitro[11]. These BSK kinases interact with BRI1 in vivo, and genetic and transgenic studies confirmed that BSKs represent a small family of kinases that activate brassinosteroid signaling downstream of BRI1[8]. Proteomic studies using plasma membrane protein preparations led to the identification of these BSK proteins as key early signaling components[8].
BRI1 also phosphorylates constitutive differential growth 1 (CDG1), another receptor-like cytoplasmic kinase[9][12]. Biochemical assays showed strong interaction between CDG1 and the BRI1 kinase domain, but not with the BAK1 kinase domain[9]. Co-immunoprecipitation experiments using microsomal fractions from transgenic Arabidopsis co-expressing BRI1-GFP and CDG1-myc confirmed in vivo interaction[9]. In vitro kinase assays demonstrated that BRI1 kinase domain, but not BAK1 kinase domain, strongly phosphorylates CDG1[9][12]. Mass spectrometry analysis identified BRI1 phosphorylation sites in CDG1, with Ser-234 being particularly important for CDG1 activation[12]. Pre-incubation of CDG1 with BRI1 and ATP increased CDG1 binding to and phosphorylation of the phosphatase BSU1, suggesting that BRI1 phosphorylation of CDG1 at Ser-234 activates CDG1 kinase activity[9][12].
BRI1 transphosphorylates its coreceptor BAK1 on specific kinase-domain residues critical for BAK1 function[3][10]. Reciprocally, BAK1 transphosphorylates BRI1, quantitatively increasing BRI1 kinase activity toward specific substrates[3][10]. Sequential transphosphorylation experiments demonstrated that brassinosteroid-dependent activation of BRI1 precedes association with BAK1 in planta, and that BRI1 positively regulates BAK1 phosphorylation levels in vivo[3][10]. This sequential transphosphorylation model proposes that BRI1 controls signaling specificity through direct brassinosteroid binding followed by substrate phosphorylation, with coreceptor BAK1 then activated by BRI1-dependent transphosphorylation and subsequently enhancing signaling output through reciprocal BRI1 transphosphorylation[3][10].
The Arabidopsis transthyretin-like protein (TTL) was identified as a potential BRI1 substrate through yeast two-hybrid screening using the BRI1 kinase domain[27]. TTL interacts with BRI1 in a kinase-dependent manner in yeast and is phosphorylated by BRI1 in vitro[27]. TTL displays similar expression patterns to BRI1 and associates with the plasma membrane[27]. Overexpression of TTL results in phenotypes observed in weak bri1 mutants and null bak1 mutants, while T-DNA insertional mutations in TTL promote plant growth and enhance brassinosteroid sensitivity[27]. These findings suggest that TTL functions as a negative regulator of brassinosteroid signaling, potentially regulating certain biochemical activities near the plasma membrane to control plant growth[27].
Guanylate Cyclase Activity
Beyond its kinase activity, BRI1 possesses guanylate cyclase activity capable of generating cyclic GMP[15][18]. Using liquid chromatography-tandem mass spectrometry, researchers confirmed that the recombinant cytoplasmic domain of BRI1 generates picomol amounts of cGMP per microgram protein, with a preference for magnesium over manganese as a cofactor[15][18]. Importantly, functional BRI1 kinase activity is essential for optimal cGMP generation[15][18]. The guanylate cyclase activity of BRI1 is modulated by its kinase activity, while cGMP, the product of guanylate cyclase, in turn inhibits BRI1 kinase activity[15][18]. This reciprocal regulation creates a feedback mechanism that fine-tunes brassinosteroid signaling[15][18].
Using Arabidopsis root cell cultures, researchers demonstrated that cGMP rapidly potentiates phosphorylation of the downstream substrate brassinosteroid-signaling kinase 1 (BSK1)[15][18]. These results suggest that cGMP acts as a modulator that enhances downstream signaling while dampening signal generation from the receptor[15][18]. The dual enzymatic activity of BRI1 as both a protein kinase and guanylate cyclase represents an unusual feature among plant receptor kinases and provides an additional layer of complexity to brassinosteroid signal transduction[15][18]. The intracellular kinase domain of BRI1 thus encapsulates a guanylate cyclase catalytic center alongside its kinase function[15][18].
Protein-Protein Interactions and Complex Formation
BRI1 exists as a monomer or homodimer at the plasma membrane under basal conditions[2][5][14]. In the absence of brassinosteroids, BRI1 is held in an inactive state by BRI1 kinase inhibitor 1 (BKI1)[2][25]. Structural and biochemical studies revealed that BKI1 binds to the BRI1 kinase domain through its BRI1-interacting motif (BIM), with this interaction being essential for inhibiting BRI1 activity[25]. The BIM peptide of BKI1 binds to the αG region of the BRI1 kinase domain, and this binding is sufficient to tightly interact with BRI1 with a dissociation constant of approximately 5.4 μM[25]. When brassinosteroid binds to BRI1, it reduces the stability of the BRI1:BKI1 complex and promotes binding of BRI1 to BAK1[2][25]. Phosphorylated BKI1 has lower affinity for BRI1 than dephosphorylated BKI1, facilitating release of the inhibitor upon receptor activation[25].
BRI1 forms heterodimers with BAK1 in endosomes upon brassinosteroid perception[2][13][55]. Both BRI1 and BAK1 make contact with the brassinosteroid molecule in the receptor-coreceptor complex, qualifying them as a coreceptor pair[2][11][17]. In the BRI1:BAK1 complex, the two receptor kinases sequentially phosphorylate each other in their kinase domains, resulting in full activation of BRI1[2][3][10]. BRI1 also interacts with SERK1 and SERK4/BKK1, which play partially redundant roles with BAK1 in brassinosteroid signaling[23][55]. Component analysis revealed that BRI1 is part of a SERK1 signaling complex composed of KAPP, CDC48A, GRF6 or GRF7, SERK1, SERK2, SERK3/BAK1, and BRI1[55].
BRI1 interacts with BIK1 (botrytis-induced kinase 1), a plasma membrane-localized receptor-like cytoplasmic kinase[11][49]. Co-immunoprecipitation assays demonstrated that BIK1 associates with both BRI1 and BAK1[11][49]. Interestingly, the association of BIK1 with BRI1 appears to be reduced upon brassinolide treatment, suggesting that BIK1 might be released from the receptor complex upon brassinosteroid perception[49]. In vitro kinase assays confirmed that BRI1 directly phosphorylates BIK1, and brassinolide treatment enhanced BRI1 phosphorylation of BIK1[49]. BIK1 functions as a signaling hub downstream of multiple pattern recognition receptors and appears to connect brassinosteroid signaling with immune responses[49][52].
BRI1 interacts with multiple B subunits of protein phosphatase 2A (PP2A), including B'α, B'β, B'γ, and B'η[50][53]. These PP2A regulatory subunits target the phosphatase complex to BRI1 for dephosphorylation, playing important roles in brassinosteroid signaling regulation[50][53]. The B'η family members prevent formation of the BRI1-BAK1 heterodimer by promoting BRI1 dephosphorylation, creating a negative feedback mechanism that blocks excessive signaling[50]. Conversely, B'α and B'β target PP2A to downstream transcription factors BZR1 and BES1 for dephosphorylation, which activates these transcription factors to switch on brassinosteroid-responsive genes[50].
Receptor-like protein 44 (RLP44) was recently identified as an important BRI1 interacting partner[7][24][35]. RLP44 physically associates with BRI1 and is required for proper brassinosteroid signaling[7][24][35]. Loss of RLP44 results in ectopic xylem differentiation in root vascular tissues, a phenotype shared with BRI1 null mutants[24][35]. Importantly, BRI1 is required for RLP44 expression and function, creating a regulatory circuit where BRI1 controls the abundance and activity of RLP44, which in turn modulates BRI1 signaling specificity[7][24][35]. RLP44 promotes complex formation between the phytosulfokine receptor PSKR1 and its coreceptor, connecting brassinosteroid and phytosulfokine signaling pathways[24][35].
Enzyme Kinetics and Catalytic Properties
Detailed kinetic analysis of BRI1 catalytic activity revealed important properties of this dual-specificity kinase. The kinase domain of BRI1 exhibits robust autophosphorylation activity, with phosphorylation occurring on both serine/threonine and tyrosine residues[3][14][43]. Kinetic parameters for substrate phosphorylation have been determined through in vitro assays using recombinant BRI1 kinase domain and various substrates[3][6][12]. The Michaelis constant (Km) for BRI1-catalyzed BKI1 C-terminal phosphorylation was measured, with the BRI1-interacting motif being a major determinant for substrate specificity[25]. Deletion of the BIM region from BKI1 resulted in more than an 80-fold decrease in phosphorylation rate compared to intact BKI1, demonstrating the importance of this interaction motif for efficient catalysis[25].
The activation state of BRI1 is tightly controlled by multiple phosphorylation events. Phosphorylation of the activation loop is essential for full kinase activity, with mutations at key activation loop residues severely impairing or abolishing catalytic function[6][14][57]. The C-terminal region of BRI1 plays an inhibitory role, as deletion of the C-terminal 40 amino acids increased receptor activity, indicated by enhanced phosphorylation of BRI1 and increased growth response in transgenic plants[5]. The inhibitory function of the C-terminal region appears to be reduced by phosphorylation, as kinase activity increased when several serine/threonine residues in this region were mutated to aspartate (mimicking phosphorylation) and decreased when mutated to alanine (preventing phosphorylation)[5][14].
Crystal structures of the phosphorylated BRI1 kinase domain in complex with nucleotides provided molecular details of the catalytic mechanism[6][57]. The structures revealed that the ATP-binding pocket of BRI1 adopts a canonical configuration typical of active protein kinases, with proper positioning of catalytic residues for phosphotransfer[6][57]. The kinase domain structure rationalized the effects of previously characterized genetic missense alleles, many of which map to residues critical for maintaining proper protein folding, ATP binding, or substrate recognition[6][14][57]. These structural insights have been essential for understanding how specific mutations cause brassinosteroid insensitivity and developmental defects in Arabidopsis[6][21][34][57].
Cellular Component: Subcellular Localization and Trafficking
Plasma Membrane Localization and Function
BRI1 localizes primarily to the plasma membrane, where it functions as the major site for brassinosteroid perception and signal initiation[2][13][14][44]. Confocal microscopy analysis of full-length BRI1 fused to green fluorescent protein demonstrated clear plasma membrane localization in various cell types[14][44]. The plasma membrane-localized BRI1 is the active form of the receptor that initiates brassinosteroid signaling[13][44]. Genetic and biochemical evidence strongly supports the plasma membrane as the primary site of BRI1 signaling activity, as mutations affecting proper plasma membrane trafficking of BRI1 result in severe brassinosteroid insensitivity and developmental defects[13][44][47].
BRI1 is an integral membrane protein with a single-pass transmembrane domain[2][14]. The protein topology consists of an extensive extracellular domain containing 25 leucine-rich repeats, a single transmembrane helix, and an intracellular portion containing the kinase domain[2][14][17]. The extracellular leucine-rich repeat domain forms a highly curved solenoid structure with an overall rotation angle of approximately 360 degrees, creating a complete right-handed helical turn[17]. This unusual twisted architecture distinguishes BRI1 from other leucine-rich repeat proteins that typically adopt horseshoe-shaped structures[17]. The transmembrane domain anchors BRI1 in the plasma membrane and may play roles in signal transduction by transmitting conformational changes from the extracellular to intracellular domains upon ligand binding[17][55].
At the plasma membrane, BRI1 exists in dynamic equilibrium between monomeric and oligomeric states[5][55]. In the absence of brassinosteroids, BRI1 may exist as preformed inactive homo-oligomers, with kinase activity maintained at basal levels through cis- or trans-repression by the C-terminal domain[5]. Steroid binding to the extracellular domain induces conformational changes in the kinase domain that allow trans-phosphorylation of the C-terminal region, enhancing BRI1 kinase activity and promoting further phosphorylation[5]. This brassinosteroid-induced activation facilitates recruitment and phosphorylation of BAK1, leading to full activation of the receptor complex[3][5][10][55].
Importantly, the level of BRI1 at the plasma membrane directly correlates with brassinosteroid signaling output and plant growth[26][56]. Overexpression of BRI1 in Arabidopsis promotes plant growth with elongated petioles, whereas BRI1 deficiency leads to dwarfism with shorter petioles[56]. The dynamic equilibrium of BRI1 at the plasma membrane is maintained by delivery of newly synthesized protein via the normal secretory pathway, brassinosteroid-dependent and brassinosteroid-independent endocytosis leading to vacuolar degradation, and recycling back to the plasma membrane[26][56]. Cytosolic proteasomal degradation also contributes to BRI1 quality control[26][56].
Endosomal Localization and Signaling
In addition to plasma membrane localization, BRI1 localizes to endosomal compartments, including trans-Golgi network/early endosomes (TGN/EE) and multivesicular bodies[13][44][47]. Live-cell imaging studies using BRI1-GFP fusion proteins expressed at endogenous levels revealed BRI1 in both plasma membrane and highly mobile, pleiomorphic intracellular structures in root meristem cells[13][44][47]. These BRI1-positive intracellular structures colocalize with the endocytic tracer FM4-64 after 5-10 minutes of uptake and display partial colocalization with VHA-a1, a marker for TGN/early endosomes, confirming their identity as early endosomes[13][44][47].
Importantly, BRI1 trafficking between the plasma membrane and endosomes occurs constitutively, independent of ligand binding[13][44][47]. Neither exogenous brassinolide treatment nor brassinosteroid depletion using the biosynthesis inhibitor brassinazole affected BRI1 localization or protein levels[13][44][47]. This ligand-independent trafficking contrasts with many animal receptor tyrosine kinases that exhibit ligand-induced endocytosis and degradation[13][44]. The constitutive cycling of BRI1 between compartments suggests that brassinosteroid signaling does not primarily rely on receptor trafficking as a regulatory mechanism, but rather on changes in receptor activation state[13][44][47].
Multiple lines of evidence indicate that endosomal BRI1 represents an active signaling pool[13][44]. The inhibitory protein BKI1 does not accumulate in endosomal structures, suggesting that endosomal BRI1 may be in an activated state not bound by this negative regulator[13][44]. Furthermore, increasing the ratio of endosomal to plasma membrane-localized BRI1 significantly enhances signaling activity[13][44]. Treatment with brefeldin A (BFA), which blocks endosomal trafficking and causes BRI1 accumulation in enlarged endosomal compartments, results in increased brassinosteroid signaling[13][44]. These observations led to the proposal that downstream targets of activated BRI1 may localize preferentially to endosomal compartments[13][44].
The distribution of BRI1 between plasma membrane and endosomal compartments is regulated by multiple factors. Pulse-chase analysis using heat-shock-inducible BRI1-YFP determined the half-life of BRI1 to be approximately 5 hours, indicating relative stability of the receptor[13][44]. This moderate turnover rate suggests that BRI1 either recycles during its lifetime or that only a minor fraction engages in endocytic transport to the vacuole, with the majority remaining at the plasma membrane[13][44]. Ubiquitination of BRI1 by E3 ubiquitin ligases PUB12 and PUB13 promotes endocytosis and subsequent degradation[26][56]. In pub12pub13 double mutants, BRI1 protein levels increase, endocytosis is impaired, and residence time at the plasma membrane is prolonged, resulting in brassinosteroid hypersensitivity[26][56].
Endoplasmic Reticulum Quality Control
Proper folding and quality control of BRI1 occurs in the endoplasmic reticulum before trafficking to the plasma membrane[21][34][39]. Several bri1 mutant alleles, including bri1-5 and bri1-9, produce misfolded proteins that are retained in the endoplasmic reticulum and fail to reach the plasma membrane[21][34]. These mutations typically affect the extracellular leucine-rich repeat domain and result in severe brassinosteroid insensitivity and developmental defects[21][34]. The endoplasmic reticulum quality control machinery recognizes misfolded BRI1 variants and targets them for endoplasmic reticulum-associated degradation[21][34].
Recent work identified DAY (DE-ETIOLATION IN THE DARK AND YELLOWING IN THE LIGHT) as a chaperone-like protein containing a DnaJ-like domain that plays critical roles in BRI1 stability and trafficking[39]. DAY localizes to both endomembrane compartments and chloroplasts via its first transmembrane domain and chloroplast transit peptide[39]. In endomembrane compartments, DAY interacts with BRI1 and stabilizes the receptor, promoting its proper folding and trafficking to the plasma membrane[39]. Knockdown of DAY results in reduced BRI1 protein abundance and brassinosteroid insensitivity[39]. This work revealed that DAY acts as a molecular chaperone specifically for BRI1, facilitating receptor maturation and quality control in the secretory pathway[39].
The leucine carboxylmethyltransferase SBI1 (SUPPRESSOR OF BRI1) regulates BRI1 stability and degradation through its effects on protein phosphatase 2A[53]. SBI1 methylates the catalytic subunit of PP2A, facilitating its association with activated BRI1 at membranes[53]. This methylation-dependent recruitment of PP2A leads to BRI1 dephosphorylation and subsequent degradation, providing negative feedback regulation of brassinosteroid signaling[53]. The sbi1 mutant shows selective accumulation of brassinosteroid-activated BRI1, indicating that SBI1 specifically targets the active, phosphorylated form of the receptor for turnover[53]. This sophisticated quality control mechanism ensures appropriate receptor levels and prevents excessive brassinosteroid signaling[53].
Protein Complex Assembly and Membrane Microdomains
BRI1 functions as part of multi-protein complexes assembled at the plasma membrane and endosomal compartments[2][24][35][55]. The core signaling complex consists of BRI1, the coreceptor BAK1 (or SERK1/SERK4), and various receptor-like cytoplasmic kinases including BSKs, CDG1, and BIK1[2][8][9][11][55]. Formation of this signaling complex is dynamically regulated by brassinosteroid availability, with hormone binding promoting heterooligomerization of BRI1 with SERK family coreceptors[3][11][17][55]. Fluorescence resonance energy transfer (FRET) and bimolecular fluorescence complementation (BiFC) studies confirmed direct interactions between BRI1 and these signaling partners at the plasma membrane[9][11][55].
Recent evidence suggests that BRI1 and other receptor kinases may localize to distinct membrane microdomains or nanodomains that contribute to signaling specificity[2]. BRI1 shares signaling components with the flagellin receptor FLS2, including the coreceptor BAK1 and downstream kinases like BIK1, yet these receptors elicit very different cellular responses[2][49]. Spatial separation of receptor complexes into different membrane nanodomains has been proposed to account for their distinct signaling outputs despite shared components[2]. This compartmentalization may be achieved through differential lipid associations, scaffolding proteins, or other mechanisms that create functionally distinct signaling platforms at the plasma membrane[2].
The receptor-like protein RLP44 plays a critical role in organizing BRI1 signaling complexes and connecting brassinosteroid signaling with other pathways[7][24][35]. RLP44 interacts with both BRI1 and the phytosulfokine receptor PSKR1, promoting association of PSKR1 with its coreceptor BAK1[24][35]. This molecular bridge allows coordination between brassinosteroid and phytosulfokine signaling pathways, demonstrating how membrane-localized scaffold proteins can integrate multiple hormone and peptide signals[24][35]. Loss of RLP44 results in altered vascular cell fate determination, highlighting the importance of proper complex assembly for developmental outcomes[24][35].
BRI1 also associates with other plasma membrane proteins beyond the core brassinosteroid signaling machinery. Direct interaction has been demonstrated between BRI1 and the plasma membrane H+-ATPase AHA2, suggesting roles in coordinating growth responses with cellular energetics and pH homeostasis[39]. These broader interactions position BRI1 as a signaling hub that integrates brassinosteroid perception with multiple cellular processes at the plasma membrane[39]. The full complement of BRI1-interacting proteins and their organization into functional complexes continues to be an active area of investigation[58].
Biological Process: Developmental and Physiological Roles
Cell Elongation and Expansion
BRI1 plays fundamental roles in regulating cell elongation and expansion throughout plant development[2][14][19][31][37]. Loss-of-function bri1 mutants exhibit severe dwarfism primarily caused by defects in cell elongation rather than cell number[14][19]. Microscopic examination of bri1 mutants reveals that reduced cell size, particularly in hypocotyls and petioles, accounts for the dwarf phenotype[14][19]. Conversely, overexpression of BRI1 or enhanced brassinosteroid signaling promotes excessive cell elongation, resulting in elongated hypocotyls, petioles, and inflorescence stems[37][56]. These observations establish BRI1 as a critical positive regulator of cell elongation across multiple tissue types[2][31][37].
The molecular mechanisms by which BRI1 promotes cell elongation involve activation of transcription factors BZR1 and BES1, which regulate expression of genes controlling cell wall modification, expansin activity, and cytoskeletal organization[23][29][37][38]. Brassinosteroid signaling through BRI1 enhances cell wall loosening and expansion by inducing expression of xyloglucan endotransglycosylases, expansins, and other cell wall-modifying enzymes[23][37]. BRI1 signaling also coordinates cell expansion with cellular metabolism and ion transport, as evidenced by its interaction with the plasma membrane H+-ATPase[39]. The proton pump creates the electrochemical gradient necessary for nutrient uptake and drives cell expansion through acidification of the cell wall, which activates expansins[39].
Genetic analysis using double mutants revealed epistatic relationships between BRI1 and other regulators of cell elongation. The dwarf phenotype of bri1 mutants cannot be rescued by mutations that enhance gibberellin signaling, indicating that brassinosteroid signaling through BRI1 acts independently of gibberellin in promoting cell elongation in most tissues[20][32]. However, in specific developmental contexts such as vascular patterning, interactions between brassinosteroid and gibberellin pathways do occur[20][32]. The balance between cell division and cell elongation is also influenced by BRI1 activity, with brassinosteroid signaling generally promoting elongation over division in most cell types[31][37].
Vascular Development and Patterning
BRI1 plays essential roles in vascular tissue development and patterning in Arabidopsis[2][24][31][32][35]. Loss of BRI1 function results in abnormal vascular development characterized by altered numbers and arrangements of vascular bundles in stems and roots[24][31][32]. The bri1 null mutants show ectopic xylem differentiation in positions normally occupied by procambium, indicating that BRI1 is required for maintaining procambial cell fate and preventing premature xylem differentiation[24][35]. This vascular cell fate determination function appears to be independent of canonical brassinosteroid signaling outputs, as intermediate bri1 alleles and BR biosynthesis mutants do not show the same vascular phenotypes[24][35].
BRI1 homologs BRL1 and BRL3 play specialized roles in vascular development, with functions restricted primarily to vascular tissues[2][31][32]. These BRI1-family proteins are expressed at high levels in developing vascular tissues and can bind brassinosteroids, functioning as brassinosteroid receptors in the vasculature[2][31]. The triple mutant bri1 brl1 brl3 shows more severe vascular defects than single mutants, demonstrating partially redundant functions in vascular development[2][31]. However, even in vascular tissues, BRI1 plays a more dominant role than BRL1 or BRL3[2][31]. Expression of BRL1 or BRL3 under the BRI1 promoter can rescue bri1 mutant phenotypes, suggesting that tissue-specific expression patterns rather than inherent functional differences account for their specialized roles[23][31].
Mathematical modeling combined with experimental validation demonstrated that brassinosteroid signaling through BRI1 and auxin polar transport coordinately control vascular bundle patterning in Arabidopsis inflorescence stems[32]. Brassinosteroids promote the number of cells in the provascular ring, while auxin maxima established by polar transport position the individual vascular bundles across this ring[32]. This model was supported by analysis of various brassinosteroid biosynthesis and signaling mutants, which showed altered numbers of vascular bundles correlating with brassinosteroid levels and BRI1 activity[32]. These findings revealed that BRI1 serves as a promoting signal for provascular cell number and suggest roles in controlling procambial cell divisions during vascular pattern formation[32].
The mechanism by which BRI1 controls vascular cell fate involves regulation of RLP44 expression and function[24][35]. BRI1 is required for proper RLP44 protein accumulation, and RLP44 in turn mediates interactions between BRI1 and phytosulfokine signaling pathways[24][35]. This regulatory circuit connects multiple signaling systems to coordinate vascular development[24][35]. Loss of either BRI1 or RLP44 results in similar vascular cell fate defects, indicating that they function in the same pathway for this process[24][35]. However, the vascular phenotype is independent of downstream brassinosteroid signaling components like BIN2 and BZR1, suggesting that BRI1 has signaling outputs beyond the canonical pathway in the context of vascular development[24][35].
Photomorphogenesis and Light Responses
BRI1 plays important roles in photomorphogenesis, the light-mediated development of plants[5][39][42]. Brassinosteroid-deficient and brassinosteroid-insensitive mutants including bri1 alleles exhibit constitutive photomorphogenic phenotypes when grown in darkness, displaying shortened hypocotyls and opened cotyledons characteristic of light-grown seedlings[5][14][19]. This observation indicates that brassinosteroid signaling through BRI1 normally promotes skotomorphogenic development (etiolation) and must be suppressed for proper photomorphogenesis to occur[5][42]. The constitutive photomorphogenic phenotype of bri1 mutants suggests that BRI1 serves as a developmental bridge between skotomorphogenesis and photomorphogenesis[5].
The mechanism by which BRI1 regulates photomorphogenesis involves interactions between brassinosteroid signaling and light signaling pathways[37][42]. BZR1 and BES1, the master transcription factors activated by BRI1, interact with phytochrome-interacting factors (PIFs) to coordinately regulate light-responsive gene expression[37][42]. BZR1 negatively regulates genes encoding positive light-signaling components such as phytochrome B and phototropin 1, while inducing expression of negative regulators including COP1 and SPA1[23][37]. This regulatory network positions BRI1 as an antagonist of photomorphogenesis and a positive regulator of skotomorphogenic development[23][37][42].
Recent work demonstrated that pure green light promotes hypocotyl elongation through activation of brassinosteroid signaling via BRI1[42]. Green light exposure increases DNA binding of BES1 to target gene promoters, activating expression of brassinosteroid-responsive genes that promote hypocotyl elongation[42]. Mutants deficient in brassinosteroid biosynthesis or perception, including bri1 alleles, are insensitive to green light and fail to show enhanced hypocotyl elongation[42]. These findings reveal that BRI1 is required for the shade avoidance response mediated by green light, demonstrating functional connections between light quality perception and brassinosteroid signaling[42].
The chaperone-like protein DAY provides a molecular link between BRI1-mediated brassinosteroid signaling and chloroplast development during photomorphogenesis[39]. DAY localizes to both endomembrane compartments, where it stabilizes BRI1, and to chloroplasts, where it stabilizes the chlorophyll biosynthesis enzyme protochlorophyllide oxidoreductase (POR)[39]. DAY knockdown plants show both reduced brassinosteroid signaling and impaired chloroplast development[39]. This dual function of DAY reveals how a single factor can coordinate brassinosteroid signaling and chloroplast biogenesis, two processes that must be integrated during the transition from skotomorphogenesis to photomorphogenesis[39].
Stress Responses and Environmental Adaptation
BRI1 mediates plant responses to various abiotic and biotic stresses through brassinosteroid signaling[33][36]. Brassinosteroid treatment enhances tolerance to drought, salt, cold, and heat stresses, and these protective effects require functional BRI1[33]. Expression analysis of BRI1 and brassinosteroid biosynthesis genes shows dynamic regulation in response to stress conditions[33][36]. However, the relationship between BRI1 activity and stress tolerance is complex, as both brassinosteroid-deficient and brassinosteroid-insensitive mutants can show altered stress responses[33].
Under drought stress, brassinosteroid signaling through BRI1 interacts with abscisic acid (ABA) signaling in complex ways[33]. Exogenous brassinosteroid treatment can enhance drought tolerance by improving photosynthetic efficiency, leaf water content, and antioxidant defenses[33]. However, the kinase BIN2, which is inhibited by BRI1-mediated signaling, positively regulates ABA responses and stress tolerance by phosphorylating ABA-responsive transcription factors[33]. This creates an antagonistic relationship where brassinosteroid signaling through BRI1 must be balanced with ABA signaling for optimal stress adaptation[33].
Salt stress affects BRI1 localization and function through modulation of endoplasmic reticulum-associated protein degradation pathways[33]. The bri1-9 mutant, which produces a misfolded receptor retained in the endoplasmic reticulum, shows partial recovery from salt hypersensitivity when the ER quality control pathway is suppressed[33]. High salinity also prevents nuclear accumulation of BZR1, thereby blocking brassinosteroid signaling outputs that would otherwise promote growth under non-optimal conditions[33]. This growth suppression under salt stress represents an adaptive response that allows resources to be redirected toward stress tolerance mechanisms[33].
BRI1 signaling intersects with immune responses through shared components including the coreceptor BAK1 and the receptor-like cytoplasmic kinase BIK1[2][49][52]. BAK1 functions as a coreceptor for both BRI1 and multiple pathogen-associated molecular pattern (PAMP) receptors including the flagellin receptor FLS2[2][45][49]. This shared use of BAK1 creates potential trade-offs between growth promotion mediated by BRI1 and immunity mediated by pattern recognition receptors[45][49]. Recent evidence demonstrates that BIK1 acts as a negative regulator of brassinosteroid signaling while serving as a positive regulator of immune responses, providing a molecular mechanism for balancing growth and immunity[49].
Reproductive Development and Fertility
BRI1 is essential for multiple aspects of reproductive development including flowering time, organ development, pollen formation, and seed production[14][19][22][37][45][47]. Loss-of-function bri1 mutants exhibit delayed flowering, reduced fertility, and abnormal flower morphology[14][19][22]. The severe bri1 alleles show complete male sterility, preventing genetic crosses and complicating genetic analysis[14][19][21]. Intermediate bri1 alleles that retain partial function show delayed flowering and reduced seed set, indicating that proper brassinosteroid signaling through BRI1 is required for normal reproductive development[21][37][47].
The mechanism by which BRI1 regulates flowering time involves repression of FLOWERING LOCUS C (FLC), a major floral repressor[37][47]. BRI1-dependent brassinosteroid signaling promotes flowering by reducing FLC expression, allowing activation of floral integrators including FLOWERING LOCUS T (FT) and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1)[37][47]. The transcription factor BES1 interacts with REF6 (RELATIVE OF EARLY FLOWERING 6), a Jumonji domain-containing protein that removes repressive histone marks from the FLC locus[37]. This interaction provides a mechanism for BRI1-mediated brassinosteroid signaling to affect chromatin structure and gene expression at the FLC locus[37].
BRI1 regulates male fertility through control of anther development and pollen formation[37][45]. Brassinosteroid signaling promotes expression of genes involved in tapetum development, pollen wall formation, and pollen maturation[37][45]. The transcription factors BZR1 and BES1 directly bind to promoters of genes required for anther and pollen development, activating their expression[37][45]. Loss of BRI1 function results in defective anthers with abnormal tapetum differentiation and pollen development, leading to male sterility in severe mutants and reduced pollen viability in weaker alleles[37][45].
Brassinosteroid signaling through BRI1 also affects female reproductive development, although these effects are generally less severe than impacts on male fertility[37][45]. BRI1 influences ovule development, embryo sac formation, and seed development[37]. The partial female sterility observed in some bri1 mutants may result from defects in flower development, pistil structure, or embryo development[37]. Analysis of BRI1 expression patterns shows that the receptor is expressed in developing flowers, ovules, and seeds, consistent with direct roles in female reproductive tissues[14][37].
Root Development and Architecture
BRI1 plays important roles in root development and architecture, although these effects are generally less severe than impacts on shoot development[31][35][40]. Root growth inhibition assays using exogenous brassinosteroids demonstrated that BRI1 is required for brassinosteroid perception in roots[14][19]. Loss-of-function bri1 mutants show altered root length, lateral root formation, and root hair development[31][35]. However, the most severe bri1 null mutants have relatively mild root phenotypes compared to their dramatic shoot defects, suggesting that brassinosteroid-independent functions or redundant receptors may operate in roots[31][35].
The relatively mild root phenotypes of bri1 null mutants are explained by functional redundancy with BRI1 homologs BRL1 and BRL3[31]. These BRI1-family proteins are highly expressed in roots and can bind brassinosteroids, functioning as brassinosteroid receptors in root tissues[31]. Analysis of single, double, and triple mutants revealed that BRL1 and BRL3 have partially redundant functions with BRI1 in root development[31]. The triple mutant bri1 brl1 brl3 shows more severe root defects than single mutants, demonstrating that multiple brassinosteroid receptors contribute to root development[2][31]. Castasterone accumulation was only 1.5-fold higher in bri1 mutant roots compared to wild-type, much less than the 30-fold increase in shoots, consistent with compensation by other receptors[31].
BRI1 regulates root vascular patterning through mechanisms distinct from canonical brassinosteroid signaling[24][35]. Loss of BRI1 results in ectopic xylem differentiation in root vascular tissues, with metaxylem cells forming in positions normally occupied by procambium[24][35]. This cell fate defect is specific to BRI1 and not observed in brassinosteroid biosynthesis mutants or signaling mutants, indicating a signaling output independent of BIN2 and BZR1[24][35]. The root vascular phenotype requires RLP44, which functions downstream of BRI1 to maintain procambial cell identity[24][35]. This brassinosteroid-independent function of BRI1 in root vascular development demonstrates that the receptor has signaling outputs beyond the canonical pathway[24][35].
Root gravitropism and growth responses are modulated by BRI1-mediated brassinosteroid signaling[32][45]. Brassinosteroids influence root growth direction through effects on auxin transport and distribution[32][45]. BRI1 interacts with components of auxin signaling pathways to coordinate hormone responses in roots[32]. The ethylene signaling pathway also intersects with BRI1-mediated brassinosteroid signaling in roots, with protein phosphatase 2A serving as a convergence point for these pathways[50]. These interactions allow integration of multiple hormone signals to fine-tune root growth and development[50].
Senescence and Aging
BRI1 influences leaf senescence and aging processes in Arabidopsis[2][14][37]. Loss-of-function bri1 mutants exhibit delayed leaf senescence compared to wild-type plants, with leaves remaining green longer and showing delayed yellowing and deterioration[2][14]. This delayed senescence phenotype is characteristic of brassinosteroid-deficient and brassinosteroid-insensitive mutants, indicating that brassinosteroid signaling through BRI1 normally promotes senescence[2][14]. Conversely, enhanced brassinosteroid signaling can accelerate senescence under certain conditions[2][37].
The mechanism by which BRI1 regulates senescence involves transcriptional control of senescence-associated genes through BZR1 and BES1[37][38]. These transcription factors regulate expression of genes involved in chlorophyll degradation, nutrient remobilization, and programmed cell death during senescence[37][38]. Brassinosteroid signaling through BRI1 coordinates senescence with other developmental programs and environmental responses, ensuring that nutrient remobilization occurs at appropriate developmental stages[37].
BRI1 also affects lifespan and aging at the whole-plant level[2][14]. The severely dwarfed bri1 mutants have prolonged vegetative phases and delayed transition to reproductive development, resulting in extended overall lifespans[14][37]. However, this extended lifespan reflects developmental delay rather than true aging suppression[37]. The relationship between BRI1 activity, growth rate, and aging demonstrates trade-offs between growth and longevity in plant development[2][37].
Experimental Evidence Quality and Types
Direct Biochemical Evidence
Extensive direct biochemical evidence supports the molecular functions of BRI1. In vitro kinase assays using purified recombinant BRI1 kinase domain demonstrated that BRI1 is an active protein kinase capable of autophosphorylation and substrate phosphorylation[3][6][8][9][12][14][25][43]. These studies used radioactive ATP labeling and mass spectrometry to identify specific phosphorylation sites and measure kinase activity[3][6][14][43]. The dual-specificity kinase activity of BRI1, phosphorylating both serine/threonine and tyrosine residues, was confirmed using phosphoamino acid analysis and phospho-specific antibodies[43][46]. Kinetic parameters including Km and Vmax values were determined for various substrates[25].
Brassinosteroid binding activity of BRI1 was directly demonstrated using photoaffinity labeling approaches[5][23]. Biotin-tagged photoaffinity castasterone was shown to specifically cross-link to BRI1 in Arabidopsis extracts and to recombinant BRI1 protein[5][23]. Binding assays using recombinant fragments of BRI1 identified the minimal brassinosteroid-binding region as the island domain plus LRR22[5][17][23]. Equilibrium binding studies and surface plasmon resonance analysis measured binding affinities and kinetics[17]. These direct binding studies definitively established BRI1 as the brassinosteroid receptor[5][17][23].
Protein-protein interaction studies used multiple complementary biochemical approaches. Co-immunoprecipitation experiments from plant tissue extracts and transiently transfected protoplasts demonstrated interactions between BRI1 and BAK1, BSKs, CDG1, BIK1, BKI1, and other partners[3][8][9][11][25][49]. Pull-down assays using recombinant proteins confirmed direct interactions[8][9][25]. Surface plasmon resonance and isothermal titration calorimetry measured binding affinities and thermodynamic parameters[25]. These quantitative binding studies provided Kd values and established the stoichiometry and dynamics of complex formation[25].
Guanylate cyclase activity of BRI1 was demonstrated using liquid chromatography-tandem mass spectrometry to detect and quantify cyclic GMP produced by recombinant BRI1 kinase domain[15][18]. This sensitive analytical approach confirmed that BRI1 generates picomol amounts of cGMP per microgram protein and established cofactor requirements[15][18]. The functional significance of cGMP was validated by showing that it inhibits BRI1 kinase activity and enhances phosphorylation of downstream substrates[15][18].
Genetic Evidence
Comprehensive genetic evidence supports BRI1 function through analysis of numerous mutant alleles. More than 40 distinct bri1 alleles have been isolated and characterized in Arabidopsis[14][19][21][34]. These alleles range from null mutations causing complete loss of function to weak alleles with partial activity[14][19][21]. The severity of developmental phenotypes correlates with residual BRI1 activity, providing strong genetic evidence for dose-dependent effects[14][19][21][34]. Null alleles like bri1-4 exhibit the most severe dwarfism, male sterility, and brassinosteroid insensitivity, confirming that BRI1 is essential for brassinosteroid responses[14][19].
Complementation studies definitively established that mutations in the BRI1 gene cause the brassinosteroid-insensitive phenotype[14][19][27]. Expression of wild-type BRI1 under its endogenous promoter fully rescues bri1 mutant phenotypes, restoring normal growth, development, and brassinosteroid sensitivity[14][19][21]. Domain-swapping experiments using chimeric receptors demonstrated that both the extracellular brassinosteroid-binding domain and the intracellular kinase domain are required for function[5][14]. Mutations affecting either domain cause brassinosteroid insensitivity, confirming that both brassinosteroid binding and kinase activity are essential[14][21][34].
Genetic interaction studies revealed epistatic relationships between BRI1 and downstream signaling components. Double mutant analysis showed that bri1 is epistatic to mutations affecting brassinosteroid biosynthesis, confirming that BRI1 acts downstream of brassinosteroid production[19][22]. Conversely, dominant gain-of-function mutations in downstream components like BZR1 and BES1 can partially suppress bri1 mutant phenotypes, placing these transcription factors downstream of BRI1[22][23][37]. The bin2 kinase mutant also suppresses bri1, confirming that BIN2 acts downstream and that its inactivation is a key output of BRI1 signaling[22].
Analysis of BRI1 orthologs in other plant species provided evolutionary evidence for functional conservation. BRI1 orthologs have been identified in rice, barley, tomato, pea, and other species, and mutations in these orthologs cause similar developmental phenotypes[31][34][40]. The high sequence conservation of BRI1, particularly in the kinase domain and brassinosteroid-binding regions, indicates strong evolutionary constraint consistent with essential function[31][34]. Complementation experiments showed that BRI1 orthologs from other species can partially rescue Arabidopsis bri1 mutants, demonstrating functional conservation across species[31][34].
Structural Evidence
High-resolution crystal structures provide detailed molecular understanding of BRI1 function. The structure of the BRI1 extracellular domain revealed a unique helical solenoid architecture formed by 25 leucine-rich repeats, with an island domain anchored to the inner surface[17]. The brassinolide-binding site was identified as a hydrophobic groove between the island domain and LRR22[17]. Co-crystal structures with bound brassinosteroids showed how hormone binding induces conformational changes that create a platform for BAK1 recruitment[11][17]. These structures explained how brassinosteroids act as "molecular glue" to stabilize receptor-coreceptor heterodimers[11][17].
Crystal structures of the phosphorylated BRI1 kinase domain in complex with nucleotides provided insights into catalytic mechanism[6][57]. The structures revealed the architecture of the ATP-binding pocket, the conformation of the activation loop, and the positions of critical catalytic residues[6][57]. A three-dimensional map of phosphorylation sites showed clusters in the activation loop, N-terminal extension, and juxtamembrane region[6][57]. These structures rationalized effects of point mutations found in bri1 alleles, showing how specific amino acid changes disrupt protein folding, ATP binding, or catalytic activity[6][21][34][57].
NMR spectroscopy and analytical ultracentrifugation studies characterized the oligomeric state of BRI1[17][55][57]. These analyses showed that the BRI1 ectodomain exists as a monomer in solution in the absence of ligand, but forms stable heterodimers with SERK ectodomains in the presence of brassinosteroids[17][57]. Size-exclusion chromatography coupled with multi-angle light scattering confirmed molecular weights consistent with monomeric and dimeric species[57]. These biophysical studies established that BRI1 undergoes ligand-induced heterodimerization rather than homodimerization[17][55][57].
Cryo-electron microscopy studies of membrane-embedded BRI1 complexes are beginning to provide insights into full-length receptor architecture and conformational changes upon activation[55]. These structural studies complement crystallographic analyses by revealing receptor organization in a more native membrane environment[55]. Hydrogen-deuterium exchange mass spectrometry has been used to probe conformational dynamics and identify regions that undergo structural changes upon brassinosteroid binding or phosphorylation[6]. Together, these structural approaches provide a comprehensive picture of BRI1 architecture and function[6][17][57].
Localization and Imaging Evidence
Extensive microscopy evidence documents BRI1 subcellular localization and trafficking. Confocal laser scanning microscopy of plants expressing BRI1-GFP fusion proteins showed clear plasma membrane localization in various cell types including root epidermis, hypocotyl, and leaf cells[13][14][44][47]. These studies used multiple independent transgenic lines, including lines expressing BRI1-GFP at endogenous levels, to ensure that observed localization was not an artifact of overexpression[13][44]. Z-stack imaging and 3D reconstruction confirmed plasma membrane localization[13][44].
Time-lapse live-cell imaging revealed dynamic trafficking of BRI1 between plasma membrane and endosomal compartments[13][44]. High-speed confocal imaging captured movement of BRI1-positive vesicles and their fusion with target membranes[13][44]. Photobleaching experiments including fluorescence recovery after photobleaching (FRAP) and fluorescence loss in photobleaching (FLIP) measured lateral diffusion rates and exchange kinetics between compartments[13][44]. These quantitative imaging approaches established that BRI1 undergoes constitutive cycling independent of ligand[13][44].
Co-localization studies using fluorescent markers for specific organelles identified BRI1-containing compartments[13][44][47]. BRI1 showed co-localization with FM4-64, an endocytic tracer, after short uptake periods, confirming presence in endocytic vesicles[13][44]. Partial co-localization with VHA-a1, a TGN/early endosome marker, established BRI1 presence in these compartments[13][44]. Co-localization with late endosomal and vacuolar markers was observed at later time points, consistent with trafficking to degradative compartments[13][44].
Brefeldin A treatment studies demonstrated effects of perturbed trafficking on BRI1 localization and signaling[13][44]. BFA caused accumulation of BRI1 in enlarged endosomal aggregates, and quantitative image analysis showed increased ratio of endosomal to plasma membrane BRI1[13][44]. Importantly, BFA treatment enhanced brassinosteroid signaling despite reducing plasma membrane BRI1 levels, suggesting that endosomal compartments support active signaling[13][44]. Super-resolution microscopy approaches including stimulated emission depletion (STED) microscopy have begun to resolve BRI1 organization at the nanoscale, potentially revealing membrane microdomains[2].
Expression Pattern Evidence
Detailed expression analysis established when and where BRI1 functions during development. Quantitative RT-PCR analysis showed that BRI1 is expressed in all major organs including roots, shoots, leaves, flowers, and siliques[14][37]. Expression levels are highest in actively growing tissues such as meristems, elongating stems, and developing flowers[14][37]. In situ hybridization localized BRI1 transcripts to specific cell types including vascular tissues, shoot apical meristem, and floral organ primordia[14]. These expression patterns are consistent with the diverse developmental roles of BRI1[14][37].
Promoter-reporter gene fusions provided detailed spatial expression information[14]. Plants expressing β-glucuronidase (GUS) under control of the BRI1 promoter showed activity in seedling hypocotyls, cotyledons, roots, rosette leaves, cauline leaves, stems, flowers, and siliques[14]. Within roots, BRI1 promoter activity was observed in epidermis, cortex, endodermis, and vascular tissues[14][24]. In flowers, expression occurred in sepals, petals, stamens, and carpels[14]. This widespread expression pattern supports roles for BRI1 throughout plant development[14][37].
Microarray and RNA-seq studies characterized BRI1 expression under various conditions[33][36][38]. BRI1 transcript levels show relatively modest changes in response to developmental transitions or environmental stimuli, consistent with housekeeping roles[33][36]. However, expression of brassinosteroid biosynthesis genes and downstream targets is highly dynamic, reflecting feedback regulation of the pathway[19][33][36]. Single-cell RNA-seq studies have begun to reveal cell-type-specific expression patterns and identify rare cell types where BRI1 expression is particularly high or low[24].
Protein-level expression analysis using immunoblotting and immunolocalization confirmed that BRI1 protein expression generally follows transcript patterns[14][26][56]. However, post-translational regulation significantly affects BRI1 protein levels through effects on protein stability and degradation[26][56]. Treatment with proteasome inhibitors or mutations affecting E3 ubiquitin ligases like PUB12 and PUB13 increase BRI1 protein abundance, demonstrating active proteolytic regulation[26][56]. These studies established that BRI1 levels are controlled at both transcriptional and post-translational levels[26][56].
Post-Translational Modifications and Regulation
Phosphorylation Networks
BRI1 is subject to extensive phosphorylation regulation involving multiple kinases and phosphatases. Mass spectrometry-based phosphoproteomics identified more than 20 phosphorylation sites on BRI1 across the juxtamembrane, kinase, and C-terminal domains[3][6][14][37]. Many of these sites show brassinosteroid-dependent regulation, with increased phosphorylation upon hormone treatment[3][6][14]. The activation loop residues Thr-1039, Ser-1042, Ser-1044, and Thr-1049 are critical for kinase activity, and their phosphorylation state directly controls BRI1 enzymatic function[6][14][37][57].
Autophosphorylation represents a major mechanism of BRI1 activation[3][6][14]. BRI1 phosphorylates itself on multiple residues in trans when receptor molecules are brought into proximity through ligand-induced dimerization or oligomerization[3][6][14]. This autophosphorylation enhances kinase activity toward downstream substrates and creates docking sites for interacting proteins[3][6][14]. The juxtamembrane domain contains several autophosphorylation sites that regulate interactions with negative regulators like BKI1[14][25]. Phosphorylation of these sites reduces BKI1 binding affinity, facilitating release of the inhibitor[25].
Transphosphorylation between BRI1 and BAK1 plays essential roles in receptor activation[3][10]. BRI1 phosphorylates BAK1 on specific residues in the kinase domain, enhancing BAK1 activity[3][10]. Reciprocally, BAK1 phosphorylates BRI1, further increasing BRI1 kinase activity toward downstream substrates[3][10]. This sequential transphosphorylation creates a positive feedback loop that amplifies brassinosteroid signals[3][10]. Mass spectrometry identified specific residues on each kinase that are phosphorylated by the partner kinase, and mutagenesis studies confirmed functional importance of these sites[3][10].
Protein phosphatase 2A (PP2A) dephosphorylates BRI1, providing negative regulation of receptor activity[50][53]. PP2A interacts with BRI1 and removes phosphate groups from activation loop and other regulatory sites[50][53]. The regulatory B subunits of PP2A, particularly the B'η family, target PP2A specifically to BRI1[50]. Methylation of the PP2A catalytic subunit by the leucine carboxylmethyltransferase SBI1 enhances PP2A association with membranes and interaction with BRI1[53]. Dephosphorylation by PP2A promotes BRI1 internalization and degradation, creating a negative feedback mechanism that prevents excessive signaling[53].
Ubiquitination and Degradation
Ubiquitination regulates BRI1 stability and turnover through multiple E3 ubiquitin ligases. PUB12 and PUB13, plant U-box E3 ligases, directly ubiquitinate BRI1 in vitro and in vivo[26][56]. These E3 ligases interact with BRI1 in a brassinosteroid-dependent manner, with hormone treatment enhancing the BRI1-PUB13 association[26][56]. Importantly, BRI1 phosphorylates PUB13 on Ser-344, and this phosphorylation regulates the association between PUB13 and BRI1, creating intertwined regulation of protein phosphorylation and ubiquitination[26][56].
PUB12 and PUB13 mediate BRI1 polyubiquitination, and loss of these E3 ligases in the pub12pub13 double mutant results in increased BRI1 protein levels[26][56]. The accumulated BRI1 in pub12pub13 plants shows prolonged residence time at the plasma membrane an
Perplexity
(BRI1-deep-research-perplexity-lite.md)
Perplexity
(BRI1-deep-research-perplexity-lite.md)The BRI1 gene in Arabidopsis thaliana encodes a leucine-rich repeat receptor-like kinase (LRR-RLK) that functions as the primary cell surface receptor for brassinosteroids (BRs), which are essential plant steroid hormones regulating growth and development[6][5][3].
Key Concepts and Definitions
-
Molecular Function:
BRI1 acts as a transmembrane receptor kinase. Upon binding brassinosteroids at its extracellular domain, BRI1 undergoes autophosphorylation and initiates a signaling cascade that regulates gene expression related to plant development, light response, and stress adaptation[6][3][5][9]. -
Biological Processes:
BRI1 is central to: - Cell elongation and division
- Seed germination
- Flowering time regulation
- Male fertility
- De-etiolation (growth in darkness)
-
Stress responses
Mutations in BRI1 or BR biosynthetic enzymes cause dwarfism, delayed flowering, reduced germination, and sterility[5][10][6][3]. -
Cellular Localization:
BRI1 is primarily localized at the plasma membrane[2][4][5]. It is synthesized in the endoplasmic reticulum (ER), undergoes ER quality control, and is trafficked to the plasma membrane. Some mutant forms (e.g., bri1-5, bri1-9) are retained in the ER due to misfolding, but wild-type and certain mutant alleles (e.g., bri1-301 at 22°C) localize mainly to the plasma membrane[2].
Protein Domains
- Extracellular Domain:
Contains 25 leucine-rich repeats (LRRs) forming a superhelical structure. A unique 70-amino acid "island domain" is inserted between the 21st and 22nd LRRs, which is critical for brassinosteroid binding[5][3]. - Transmembrane Domain:
Anchors BRI1 in the plasma membrane[5]. - Cytoplasmic Kinase Domain:
Functions as a serine/threonine kinase, essential for downstream signaling[9][5].
Known Interactions
- BAK1 (BRI1-Associated Receptor Kinase 1):
Upon BR binding, BRI1 heterodimerizes with BAK1, which is required for full activation of the signaling pathway[5][3]. - BRS1 (Brassinosteroid Suppressor 1):
A secreted serine carboxypeptidase that modulates BRI1 signaling, possibly by processing an extracellular protein involved in BR perception[1][3]. - BIN2 kinase and BSU1 phosphatase:
Downstream regulators of transcription factors BES1 and BZR1, which mediate BR-responsive gene expression[3][5]. - TRIP-1 and SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASE1 complex:
BRI1 is part of larger protein complexes involved in embryogenesis and other developmental processes[7].
Disease Associations
- Plant Phenotypes:
Mutations in BRI1 cause brassinosteroid insensitivity, resulting in severe developmental defects such as dwarfism, delayed flowering, and sterility[5][6][3].
There are no known direct associations with human diseases, as BRI1 is plant-specific.
Recent Developments (2023–2024)
- Cellular Safeguarding:
Recent studies highlight BRI1's role in safeguarding cell capacity to respond to differential brassinosteroid cues, ensuring robust developmental responses at the single-cell level[8]. - Quality Control Mechanisms:
New research has clarified the ER and plasma membrane quality control systems that regulate BRI1 stability and trafficking, especially for mutant alleles[2]. - Evolutionary Conservation:
The serine residues in BRI1's LRR motifs are evolutionarily conserved and critical for receptor function and BR binding[5].
Relevant Statistics and Data
- Gene ID: At4g39400 (Arabidopsis Genome Initiative)[10].
- Protein Length: 1,199 amino acids[6].
- Mutant Phenotypes:
- bri1-5, bri1-9: ER retention, severe dwarfism[2][5].
- bri1-301: Partial ER retention, temperature-sensitive instability[2].
Authoritative Sources
- UniProtKB (O22476): Comprehensive protein annotation, including domains and function[6].
- Frontiers in Plant Science (2020): Structural and functional analysis of BRI1[5].
- PNAS (2001): BRS1 interaction and genetic suppression studies[1].
- Science Advances (2023): Single-cell developmental safeguarding by BRI1[8].
- NCBI Gene (830095): Interaction networks and gene function[7].
URLs and Publication Dates:
- UniProtKB: https://www.uniprot.org/uniprotkb/O22476/entry (2024)
- Frontiers in Plant Science: https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2020.00032/full (2020)
- PNAS: https://www.pnas.org/doi/10.1073/pnas.091065998 (2001)
- Science Advances: https://www.science.org/doi/10.1126/sciadv.adq3352 (2023)
- NCBI Gene: https://www.ncbi.nlm.nih.gov/gene/830095 (2024)
If you need more detailed structural data or specific mutant analyses, please specify.
Citations
- https://www.uniprot.org/uniprotkb/O22476/entry
- https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2020.00032/full
- https://www.pnas.org/doi/10.1073/pnas.091065998
- https://www.science.org/doi/10.1126/sciadv.adq3352
- https://www.ncbi.nlm.nih.gov/gene/830095