Deep Research
Falcon
(COP1-deep-research-falcon.md)
Falcon
(COP1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: Arabidopsis thaliana COP1 (UniProt P43254; At2g32950)
1) Identity verification and scope control (disambiguation)
The target is Arabidopsis thaliana CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1), an E3 ubiquitin-protein ligase that functions as a central repressor of photomorphogenesis. In the Arabidopsis literature used here, COP1 is consistently described as a ~76 kDa RING-type E3 ligase with N-terminal RING, central coiled-coil, and C-terminal WD40 repeats, matching the UniProt P43254 identity and its domain architecture. (boycheva2024howhistoneacetyltransferases pages 7-9, ponnu2021illuminatingthecop1spa pages 1-2, han2020thephotomorphogeniccentral pages 2-3, ponnu2021illuminatingthecop1spa pages 2-4)
2) Key concepts and definitions (current understanding)
2.1 COP1 as an E3 ubiquitin ligase in the UPS
COP1 is a RING-finger E3 ubiquitin ligase that catalyzes E2-dependent ubiquitin transfer to substrates, leading commonly to polyubiquitination and 26S proteasome-dependent degradation as a mechanism to repress light-induced developmental programs in darkness. (boycheva2024howhistoneacetyltransferases pages 7-9, ponnu2021illuminatingthecop1spa pages 1-2, han2020thephotomorphogeniccentral pages 2-3)
COP1 functions in vivo as part of a larger E3 ligase assembly: COP1 associates with SPA proteins to form COP1/SPA complexes and connects to a CUL4–DDB1–RBX1 scaffold, creating a multimeric E3 ligase module that supports substrate ubiquitylation in light signaling. (boycheva2024howhistoneacetyltransferases pages 7-9, ponnu2021illuminatingthecop1spa pages 1-2, ponnu2021illuminatingthecop1spa pages 2-4)
2.2 Domain architecture and substrate recognition logic
Arabidopsis COP1 is composed of:
- RING domain (E2 interaction/ubiquitin transfer module)
- Coiled-coil domain (mediates COP1 homo-oligomerization and interaction with SPA proteins)
- WD40 repeat domain (a seven-bladed β-propeller; major substrate-recognition surface) (boycheva2024howhistoneacetyltransferases pages 7-9, ponnu2021illuminatingthecop1spa pages 1-2, han2020thephotomorphogeniccentral pages 2-3, ponnu2021illuminatingthecop1spa pages 2-4)
A key organizing concept is that many COP1 substrates (and some photoreceptor regulators) contain a short VP (valine–proline) motif that is recognized by the COP1 WD40 pocket, enabling competitive binding and light-dependent rewiring of interactions. (ponnu2021illuminatingthecop1spa pages 1-2, ponnu2021illuminatingthecop1spa pages 2-4)
2.3 Canonical pathway role: repress photomorphogenesis in darkness
COP1 is a “central switch” of global light-responsive gene expression by promoting the turnover of multiple nuclear positive regulators of photomorphogenesis, including the bZIP transcription factor HY5 (direct COP1 substrate). (wang2024cop1controlslightdependent pages 1-2, han2020thephotomorphogeniccentral pages 2-3, han2020thephotomorphogeniccentral pages 1-2)
3) Molecular functions, biochemical activity, and substrate specificity
3.1 Primary biochemical function
Reaction class: COP1 is an E3 ubiquitin ligase (EC 2.3.2.27). Its biochemical role is to facilitate transfer of ubiquitin from an E2 enzyme to specific substrate proteins (often transcription factors or regulatory proteins), typically marking them for proteasomal degradation. (boycheva2024howhistoneacetyltransferases pages 7-9, ponnu2021illuminatingthecop1spa pages 1-2)
Substrate specificity: Specificity is largely mediated by the WD40 domain, including VP-motif-based recognition used by many substrates/photoreceptors. (ponnu2021illuminatingthecop1spa pages 1-2, ponnu2021illuminatingthecop1spa pages 2-4)
3.2 Validated substrates and regulatory targets (selected, evidence-backed)
- HY5 (ELONGATED HYPOCOTYL 5): COP1 ubiquitinates HY5, limiting its accumulation and repressing photomorphogenesis. (han2020thephotomorphogeniccentral pages 2-3, fang2024ubiquitinspecificproteaseubp14 pages 1-2)
- VIL1 (a Polycomb-associated PHD protein): COP1 directly ubiquitinates VIL1 and promotes its proteasome-dependent degradation in darkness, linking COP1 to chromatin remodeling control. (wang2024cop1controlslightdependent pages 1-2, wang2024cop1controlslightdependent pages 2-3)
- DCS1 (plant-specific spliceosomal component): COP1-dependent ubiquitination/degradation of DCS1 contributes to light-regulated intron retention and nuclear detainment of transcripts. (zhou2024lightregulatesnuclear pages 1-2)
Reviews additionally cite multiple light-pathway transcriptional regulators (e.g., HYH, LAF1, HFR1, BBX/CONSTANS family members) as COP1-associated targets/partners consistent with COP1’s broad control of nuclear photomorphogenesis regulators. (han2020thephotomorphogeniccentral pages 2-3, ponnu2021illuminatingthecop1spa pages 2-4)
4) Subcellular localization and dynamics (where COP1 acts)
COP1 functions prominently in the nucleus in darkness, where it targets nuclear substrates for ubiquitination. COP1 contains a bipartite nuclear localization signal (NLS) and an N-terminal cytoplasmic localization signal, enabling light-regulated nucleocytoplasmic partitioning. (ponnu2021illuminatingthecop1spa pages 1-2)
Nuclear COP1 can appear in punctate nuclear speckles/bodies where interaction partners can colocalize, consistent with subnuclear organization of light signaling. (ponnu2021illuminatingthecop1spa pages 1-2)
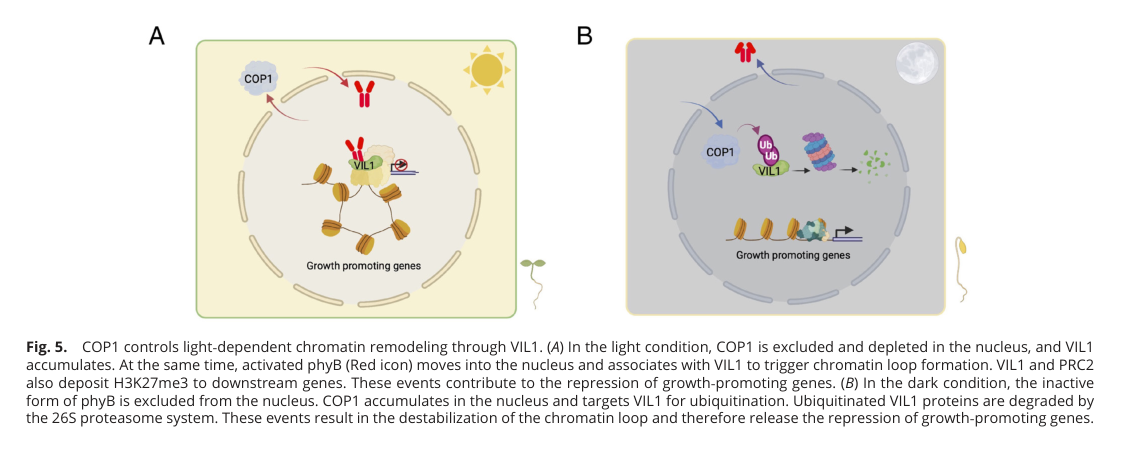
In the COP1–VIL1 chromatin pathway, a key current model is that COP1 is excluded/depleted from the nucleus in light, permitting VIL1 accumulation; in darkness, COP1 accumulates in the nucleus and ubiquitinates VIL1, reducing loop formation and altering repression at growth genes. (wang2024cop1controlslightdependent pages 6-8, wang2024cop1controlslightdependent media 3ca0080b)
5) Pathways and regulatory inputs
5.1 COP1/SPA as a hub downstream of photoreceptors
COP1 integrates signals from multiple photoreceptors and modulates substrate stability accordingly. For example, UV-B involves COP1 interaction with UVR8 in a pathway that supports HY5 stabilization, and COP1/SPA can cooperate with phytochromes in regulating PIF stability in light signaling. (ponnu2021illuminatingthecop1spa pages 1-2, ponnu2021illuminatingthecop1spa pages 2-4)
5.2 Light-regulated transcript processing via COP1–spliceosome control (2024)
A major 2024 development is that COP1 affects photomorphogenesis not only through transcription-factor turnover but also through alternative splicing/intron retention (IR) and nuclear detainment of intron-retained transcripts (IRTs). The Zhou et al. 2024 study reports that IR is prevalent and that COP1 and light regulate large numbers of nuclear IR events, with IRTs largely retained in the nucleus to prevent translation. (zhou2024lightregulatesnuclear pages 1-2, zhou2024lightregulatesnuclear pages 5-7)
6) Recent developments (prioritizing 2023–2024 primary research)
6.1 COP1 controls light-dependent chromatin remodeling via VIL1 (PNAS; Feb 2024)
Wang et al. (PNAS, Feb 2024, https://doi.org/10.1073/pnas.2312853121) established COP1 as the E3 ligase for VIL1, connecting COP1 to PRC2-associated chromatin regulation.
Key findings and data:
- COP1 limits H3K27me3 deposition at VIL1-dependent loci by degrading VIL1 in darkness. (wang2024cop1controlslightdependent pages 5-6)
- The study defined 3,368 genes as VIL1-dependent H3K27me3-enriched loci. (wang2024cop1controlslightdependent pages 5-6)
- H3K27me3 at these loci is significantly higher in the cop1-4 mutant (P = 3.8e−7). (wang2024cop1controlslightdependent pages 5-6)
- A co-regulated subset of 665 genes shows strong VIL1/COP1 co-regulation with highly elevated H3K27me3 in cop1-4 (P = 6.8e−10). (wang2024cop1controlslightdependent pages 5-6)
- Proteasome dependence: dark-induced VIL1 degradation can be inhibited by bortezomib (40 μM). (wang2024cop1controlslightdependent pages 2-3)
Mechanistic model (from Figure 5): in light, COP1 nuclear depletion allows VIL1 accumulation and phyB-associated chromatin loop formation and repression; in dark, COP1 targets VIL1 for ubiquitination and degradation, destabilizing the loop and releasing repression. (wang2024cop1controlslightdependent media 3ca0080b)
6.2 COP1 regulates nuclear detainment of intron-retained transcripts via spliceosome/DCS proteins (Nature Communications; Jun 2024)
Zhou et al. (Nature Communications, Jun 2024, https://doi.org/10.1038/s41467-024-49571-9) showed COP1 modulates IR/IRT detainment through spliceosome control, including COP1-dependent ubiquitination/degradation of a spliceosomal factor (DCS1). (zhou2024lightregulatesnuclear pages 1-2)
Key findings and data:
- 1,625 nuclear IR events were light-responsive and 1,594 were COP1-responsive. (zhou2024lightregulatesnuclear pages 5-7)
- Only about ~4% of detected IR events were in the cytoplasmic fraction, consistent with predominant nuclear detainment. (zhou2024lightregulatesnuclear pages 5-7)
- Around ~60% of IRTs (including those from PIF4, RVE1, ABA3) were upregulated in light-grown WT and dark-grown cop1-6 seedlings. (zhou2024lightregulatesnuclear pages 5-7)
- Phenotype linkage: a pif4 rve1 aba3 triple mutant had hypocotyl length reduced to ~76% of WT in the dark. (zhou2024lightregulatesnuclear pages 5-7)
- RNA-seq design: three biological replicates, differential expression criteria FDR ≤ 0.05 and fold change ≥ 2. (zhou2024lightregulatesnuclear pages 10-11)
6.3 Refining the COP1–HY5 module: UBP14 stabilizes HY5 by deubiquitination (PNAS; Aug 2024)
Fang et al. (PNAS, Aug 2024, https://doi.org/10.1073/pnas.2404883121) added an important layer to COP1-centered HY5 control by identifying UBP14 as a HY5 deubiquitinase that antagonizes COP1-mediated ubiquitination. (fang2024ubiquitinspecificproteaseubp14 pages 1-2, fang2024ubiquitinspecificproteaseubp14 pages 4-5)
Key findings and quantitative details:
- In vivo ubiquitination assays involve HY5-GFP and MYC-COP1 transient expression; UBP14 reduces HY5 ubiquitination in vivo. (fang2024ubiquitinspecificproteaseubp14 pages 4-5)
- HY5 stability experiments used cycloheximide (CHX) 1 mM ± proteasome inhibitor MG132 50 μM, sampled after dark-to-light transfer at 0, 2, 4 h, with quantification from three independent experiments (means ± SD) and significance at P < 0.05 (two-way ANOVA with Tukey). (fang2024ubiquitinspecificproteaseubp14 pages 4-5)
- Phospho-state preference: UBP14 binds and stabilizes nonphosphorylated HY5 (HY5S36A) more than phosphomimic HY5 (HY5S36D), with HY5S36A/S36D protein and ubiquitination time courses assessed at 0, 4, 8 h, quantified from three independent experiments and analyzed by ANOVA/Tukey (P < 0.05). (fang2024ubiquitinspecificproteaseubp14 pages 6-7)
7) Current applications and real-world implementations
COP1 is widely treated as an actionable regulatory node for engineering light-regulated traits because it controls protein stability of key transcription factors and, as of 2024, also controls chromatin and RNA-processing layers of gene regulation. The mechanistic expansion to chromatin (COP1→VIL1→H3K27me3/looping) and RNA-processing (COP1–spliceosome→IRTs) suggests new entry points for modifying growth programs that respond to light/dark transitions and photoreceptor state. (zhou2024lightregulatesnuclear pages 1-2, wang2024cop1controlslightdependent pages 5-6, wang2024cop1controlslightdependent media 3ca0080b)
Additionally, COP1-centered regulation is discussed in the context of secondary metabolite and phenylpropanoid/phenolic compound regulation through light signaling networks, indicating relevance for horticulture/food-production traits that depend on light-regulated transcriptional programs (review context). (boycheva2024howhistoneacetyltransferases pages 7-9)
8) Expert synthesis and analysis (authoritative perspectives)
Authoritative reviews describe COP1/SPA as a central hub that (i) uses VP-motif-dependent binding to coordinate multiple light inputs, (ii) employs regulated nuclear localization to gate access to nuclear substrates, and (iii) operates via multisubunit E3 ligase assemblies to enforce dark-state developmental programs. (ponnu2021illuminatingthecop1spa pages 1-2, han2020thephotomorphogeniccentral pages 2-3, ponnu2021illuminatingthecop1spa pages 2-4)
The 2024 primary studies collectively broaden COP1’s functional annotation from “E3 ligase that degrades transcription factors” to a more general “light-responsive proteostasis hub” that also:
- tunes chromatin repression architecture (via VIL1/PRC2/H3K27me3 and chromatin loops), and
- modulates spliceosome output and nuclear RNA availability (via DCS proteins and IRT detainment). (zhou2024lightregulatesnuclear pages 1-2, wang2024cop1controlslightdependent pages 5-6, wang2024cop1controlslightdependent media 3ca0080b)
9) Summary table (functional annotation snapshot)
The following table consolidates the most robust, evidence-supported annotation elements for Arabidopsis COP1, emphasizing 2024 findings and quantitative results.
| Functional role / process | Molecular mechanism & key partners | Substrates / targets | Localization / dynamics | Key quantitative data / statistics | Most relevant recent (2024) evidence |
|---|---|---|---|---|---|
| Core E3 ligase architecture and light-signaling repressor | Arabidopsis COP1 is a ~76 kDa RING-type E3 ligase with N-terminal RING (E2 interaction), central coiled-coil (COP1 homo-/heterodimerization with SPA proteins), and C-terminal WD40 β-propeller for substrate/photoreceptor binding; COP1 acts with SPA proteins in a CUL4-DDB1-RBX1 E3 module; many client proteins use a VP motif recognized by the COP1 WD40 pocket | Canonical light-signaling regulators include HY5, HYH, LAF1, HFR1, PIF1; table evidence also supports BBX1/CONSTANS and PP2Cs ABI1/AHG3 as COP1/SPA-associated targets/partners | COP1 contains bipartite NLS plus cytoplasmic localization signal; light regulates nuclear import/export; nuclear-localized GFP-COP1 forms punctate speckles / nuclear bodies where signaling partners colocalize | Complex size reported for COP1-SPA tetramer ~440 kDa; dark-grown seedlings contain COP1 in a ~700-kDa multimeric complex | 2024-focused evidence remains consistent with this established architecture; recent work extends COP1 functions beyond transcription factor turnover to chromatin and RNA-processing control (boycheva2024howhistoneacetyltransferases pages 7-9, ponnu2021illuminatingthecop1spa pages 1-2, han2020thephotomorphogeniccentral pages 2-3, ponnu2021illuminatingthecop1spa pages 2-4) |
| Chromatin remodeling in photomorphogenesis | COP1 directly binds, polyubiquitinates, and degrades VIL1 in the dark via the 26S proteasome; interaction maps to COP1 N-terminus (aa 1-282) and VIL1 N-terminus/PHD region; by removing VIL1 in darkness, COP1 limits VIL1/PRC2-dependent chromatin loop formation and H3K27me3 deposition at growth genes; phyB contributes to light-induced loop formation, while COP1 antagonizes this in darkness | VIL1 (direct substrate); downstream affected loci include growth-promoting genes such as ATHB2, EDF3, BIM1 | Model: in light, COP1 is excluded/depleted from nucleus while VIL1 accumulates and associates with active phyB to promote chromatin loops and repression; in dark, COP1 accumulates in nucleus, ubiquitinates VIL1, and loops destabilize | 3,368 genes identified as VIL1-dependent H3K27me3-enriched loci; 665 genes co-regulated by VIL1 and COP1; H3K27me3 significantly higher in cop1-4 at VIL1 loci (P = 3.8e−7) and in the 665-gene cluster (P = 6.8e−10); VIL1 degradation blocked by 40 μM bortezomib; quantification from 3 biological replicates; gene-expression assays used n = 3 biological replicates with 4 technical replicates each; hypocotyl assays measured 30 seedlings per line across 3 biological replicates | PNAS 2024 established COP1→VIL1 as a direct ubiquitination axis linking light signaling to Polycomb-associated chromatin remodeling and dynamic chromatin loop control (wang2024cop1controlslightdependent pages 1-2, wang2024cop1controlslightdependent pages 2-3, wang2024cop1controlslightdependent pages 8-9, wang2024cop1controlslightdependent pages 5-6, wang2024cop1controlslightdependent pages 6-8, wang2024cop1controlslightdependent media 3ca0080b) |
| RNA processing / spliceosome-dependent photomorphogenesis | Light-induced alternative splicing changes are mediated in part through a COP1-spliceosome axis; COP1-dependent ubiquitination/degradation of the plant-specific spliceosomal component DCS1 contributes to intron retention (IR) and nuclear detainment of intron-retained transcripts (IRTs), thereby reducing translation of light-signaling genes under photomorphogenic conditions | DCS1 (spliceosomal component regulated by COP1); IRT-regulated signaling transcripts highlighted include PIF4, RVE1, ABA3 | IRTs are predominantly nuclear-retained rather than cytoplasmic; dark-grown cop1-6 phenocopies light-grown WT for many IR features; DCS1 interacts with COP1 (Y2H/BiFC/Co-IP evidence in figure excerpt) | 1,625 nuclear IR events were light responsive and 1,594 were COP1 responsive; only ~4% of IR events were in cytoplasmic fraction; ~60% of IRTs including PIF4/RVE1/ABA3 were upregulated in light-grown WT and dark-grown cop1-6; ~55% overlap for nuclear IR events vs ~30% for cytoplasmic IR events; pif4 rve1 aba3 triple mutant hypocotyl length reduced to ~76% of WT in dark; RNA-seq used 3 biological replicates; DE genes called at adjusted FDR ≤ 0.05 and fold change ≥ 2 | Nature Communications 2024 expanded COP1 function from proteolysis of transcription factors to control of spliceosome activity and nuclear RNA detainment during photomorphogenesis (zhou2024lightregulatesnuclear pages 1-2, zhou2024lightregulatesnuclear pages 10-11, zhou2024lightregulatesnuclear pages 5-7) |
| HY5 proteostasis and antagonistic deubiquitination | COP1 ubiquitinates HY5, opposing photomorphogenesis; UBP14 directly binds HY5 and removes ubiquitin, stabilizing HY5, with stronger affinity for nonphosphorylated HY5 (HY5S36A) than phosphomimic HY5S36D; UBP14 and HY5 form a positive-feedback loop because HY5 promotes UBP14 expression/accumulation | HY5 (direct COP1 substrate; direct UBP14 substrate for deubiquitination) | Nuclear HY5 stabilization promotes light responses, especially during dark-to-light transition | In vivo HY5 stability assays used 1 mM cycloheximide with DMSO or 1 mM cycloheximide + 50 μM MG132 after dark-to-light transfer; sampled at 0, 2, 4 h; HY5S36A/S36D protein and ubiquitination time course sampled at 0, 4, 8 h; quantification from 3 independent experiments with means ± SD; significance tested by two-way ANOVA with Tukey multiple comparisons (P < 0.05) | PNAS 2024 sharpened the COP1-HY5 module by showing that HY5 abundance is also actively set by UBP14-mediated deubiquitination, especially for the nonphosphorylated active form (fang2024ubiquitinspecificproteaseubp14 pages 1-2, fang2024ubiquitinspecificproteaseubp14 pages 6-7, fang2024ubiquitinspecificproteaseubp14 pages 4-5) |
Table: This table summarizes experimentally supported functions, mechanisms, localization, and recent 2024 findings for Arabidopsis thaliana COP1 (UniProt P43254). It emphasizes direct evidence for COP1’s canonical E3 ligase architecture plus newer chromatin and RNA-processing roles.
10) Key references (with publication dates and URLs)
- Ponnu J, Hoecker U. Illuminating the COP1/SPA Ubiquitin Ligase… Frontiers in Plant Science (Mar 2021). https://doi.org/10.3389/fpls.2021.662793 (ponnu2021illuminatingthecop1spa pages 1-2, ponnu2021illuminatingthecop1spa pages 2-4)
- Han X, Huang X, Deng XW. The Photomorphogenic Central Repressor COP1… Plant Communications (May 2020). https://doi.org/10.1016/j.xplc.2020.100044 (han2020thephotomorphogeniccentral pages 2-3, han2020thephotomorphogeniccentral pages 1-2)
- Wang W et al. COP1 controls light-dependent chromatin remodeling PNAS (Feb 2024). https://doi.org/10.1073/pnas.2312853121 (wang2024cop1controlslightdependent pages 1-2, wang2024cop1controlslightdependent pages 2-3, wang2024cop1controlslightdependent pages 5-6, wang2024cop1controlslightdependent pages 6-8, wang2024cop1controlslightdependent media 3ca0080b)
- Zhou H et al. Light regulates nuclear detainment of intron-retained transcripts through COP1-spliceosome… Nature Communications (Jun 2024). https://doi.org/10.1038/s41467-024-49571-9 (zhou2024lightregulatesnuclear pages 1-2, zhou2024lightregulatesnuclear pages 10-11, zhou2024lightregulatesnuclear pages 5-7)
- Fang K et al. UBP14 stabilizes HY5 by deubiquitination to promote photomorphogenesis PNAS (Aug 2024). https://doi.org/10.1073/pnas.2404883121 (fang2024ubiquitinspecificproteaseubp14 pages 1-2, fang2024ubiquitinspecificproteaseubp14 pages 6-7, fang2024ubiquitinspecificproteaseubp14 pages 4-5)
Notes on evidence limitations
Within the retrieved full texts, detailed quantitative localization metrics for COP1 nuclear bodies/condensates and comprehensive substrate catalogs beyond the highlighted validated targets were not fully extractable. However, the included claims are restricted to those directly supported by the extracted review and 2024 primary-study evidence listed above.
References
-
(boycheva2024howhistoneacetyltransferases pages 7-9): Irina Boycheva, Georgi Bonchev, Vasilissa Manova, Lubomir Stoilov, and Valya Vassileva. How histone acetyltransferases shape plant photomorphogenesis and uv response. International Journal of Molecular Sciences, 25:7851, Jul 2024. URL: https://doi.org/10.3390/ijms25147851, doi:10.3390/ijms25147851. This article has 9 citations.
-
(ponnu2021illuminatingthecop1spa pages 1-2): Jathish Ponnu and Ute Hoecker. Illuminating the cop1/spa ubiquitin ligase: fresh insights into its structure and functions during plant photomorphogenesis. Frontiers in Plant Science, Mar 2021. URL: https://doi.org/10.3389/fpls.2021.662793, doi:10.3389/fpls.2021.662793. This article has 115 citations.
-
(han2020thephotomorphogeniccentral pages 2-3): Xue Han, Xi Huang, and Xing Wang Deng. The photomorphogenic central repressor cop1: conservation and functional diversification during evolution. May 2020. URL: https://doi.org/10.1016/j.xplc.2020.100044, doi:10.1016/j.xplc.2020.100044. This article has 186 citations and is from a peer-reviewed journal.
-
(ponnu2021illuminatingthecop1spa pages 2-4): Jathish Ponnu and Ute Hoecker. Illuminating the cop1/spa ubiquitin ligase: fresh insights into its structure and functions during plant photomorphogenesis. Frontiers in Plant Science, Mar 2021. URL: https://doi.org/10.3389/fpls.2021.662793, doi:10.3389/fpls.2021.662793. This article has 115 citations.
-
(wang2024cop1controlslightdependent pages 1-2): Wenli Wang, Junghyun Kim, Teresa S. Martinez, Enamul Huq, and Sibum Sung. Cop1 controls light-dependent chromatin remodeling. Proceedings of the National Academy of Sciences of the United States of America, Feb 2024. URL: https://doi.org/10.1073/pnas.2312853121, doi:10.1073/pnas.2312853121. This article has 22 citations and is from a highest quality peer-reviewed journal.
-
(han2020thephotomorphogeniccentral pages 1-2): Xue Han, Xi Huang, and Xing Wang Deng. The photomorphogenic central repressor cop1: conservation and functional diversification during evolution. May 2020. URL: https://doi.org/10.1016/j.xplc.2020.100044, doi:10.1016/j.xplc.2020.100044. This article has 186 citations and is from a peer-reviewed journal.
-
(fang2024ubiquitinspecificproteaseubp14 pages 1-2): Ke Fang, Xiuhong Yao, Yu’ang Tian, Yang He, Yingru Lin, Wei Lei, Sihan Peng, Guohui Pan, Haoyu Shi, Dawei Zhang, and Honghui Lin. Ubiquitin-specific protease ubp14 stabilizes hy5 by deubiquitination to promote photomorphogenesis in arabidopsis thaliana. Proceedings of the National Academy of Sciences of the United States of America, Aug 2024. URL: https://doi.org/10.1073/pnas.2404883121, doi:10.1073/pnas.2404883121. This article has 11 citations and is from a highest quality peer-reviewed journal.
-
(wang2024cop1controlslightdependent pages 2-3): Wenli Wang, Junghyun Kim, Teresa S. Martinez, Enamul Huq, and Sibum Sung. Cop1 controls light-dependent chromatin remodeling. Proceedings of the National Academy of Sciences of the United States of America, Feb 2024. URL: https://doi.org/10.1073/pnas.2312853121, doi:10.1073/pnas.2312853121. This article has 22 citations and is from a highest quality peer-reviewed journal.
-
(zhou2024lightregulatesnuclear pages 1-2): Hua Zhou, Haiyue Zeng, Tingting Yan, Sunlu Chen, Ying Fu, Guochen Qin, Xianhai Zhao, Yueqin Heng, Jian Li, Fang Lin, Dongqing Xu, Ning Wei, and Xing Wang Deng. Light regulates nuclear detainment of intron-retained transcripts through cop1-spliceosome to modulate photomorphogenesis. Nature Communications, Jun 2024. URL: https://doi.org/10.1038/s41467-024-49571-9, doi:10.1038/s41467-024-49571-9. This article has 29 citations and is from a highest quality peer-reviewed journal.
-
(wang2024cop1controlslightdependent pages 6-8): Wenli Wang, Junghyun Kim, Teresa S. Martinez, Enamul Huq, and Sibum Sung. Cop1 controls light-dependent chromatin remodeling. Proceedings of the National Academy of Sciences of the United States of America, Feb 2024. URL: https://doi.org/10.1073/pnas.2312853121, doi:10.1073/pnas.2312853121. This article has 22 citations and is from a highest quality peer-reviewed journal.
-
(wang2024cop1controlslightdependent media 3ca0080b): Wenli Wang, Junghyun Kim, Teresa S. Martinez, Enamul Huq, and Sibum Sung. Cop1 controls light-dependent chromatin remodeling. Proceedings of the National Academy of Sciences of the United States of America, Feb 2024. URL: https://doi.org/10.1073/pnas.2312853121, doi:10.1073/pnas.2312853121. This article has 22 citations and is from a highest quality peer-reviewed journal.
-
(zhou2024lightregulatesnuclear pages 5-7): Hua Zhou, Haiyue Zeng, Tingting Yan, Sunlu Chen, Ying Fu, Guochen Qin, Xianhai Zhao, Yueqin Heng, Jian Li, Fang Lin, Dongqing Xu, Ning Wei, and Xing Wang Deng. Light regulates nuclear detainment of intron-retained transcripts through cop1-spliceosome to modulate photomorphogenesis. Nature Communications, Jun 2024. URL: https://doi.org/10.1038/s41467-024-49571-9, doi:10.1038/s41467-024-49571-9. This article has 29 citations and is from a highest quality peer-reviewed journal.
-
(wang2024cop1controlslightdependent pages 5-6): Wenli Wang, Junghyun Kim, Teresa S. Martinez, Enamul Huq, and Sibum Sung. Cop1 controls light-dependent chromatin remodeling. Proceedings of the National Academy of Sciences of the United States of America, Feb 2024. URL: https://doi.org/10.1073/pnas.2312853121, doi:10.1073/pnas.2312853121. This article has 22 citations and is from a highest quality peer-reviewed journal.
-
(zhou2024lightregulatesnuclear pages 10-11): Hua Zhou, Haiyue Zeng, Tingting Yan, Sunlu Chen, Ying Fu, Guochen Qin, Xianhai Zhao, Yueqin Heng, Jian Li, Fang Lin, Dongqing Xu, Ning Wei, and Xing Wang Deng. Light regulates nuclear detainment of intron-retained transcripts through cop1-spliceosome to modulate photomorphogenesis. Nature Communications, Jun 2024. URL: https://doi.org/10.1038/s41467-024-49571-9, doi:10.1038/s41467-024-49571-9. This article has 29 citations and is from a highest quality peer-reviewed journal.
-
(fang2024ubiquitinspecificproteaseubp14 pages 4-5): Ke Fang, Xiuhong Yao, Yu’ang Tian, Yang He, Yingru Lin, Wei Lei, Sihan Peng, Guohui Pan, Haoyu Shi, Dawei Zhang, and Honghui Lin. Ubiquitin-specific protease ubp14 stabilizes hy5 by deubiquitination to promote photomorphogenesis in arabidopsis thaliana. Proceedings of the National Academy of Sciences of the United States of America, Aug 2024. URL: https://doi.org/10.1073/pnas.2404883121, doi:10.1073/pnas.2404883121. This article has 11 citations and is from a highest quality peer-reviewed journal.
-
(fang2024ubiquitinspecificproteaseubp14 pages 6-7): Ke Fang, Xiuhong Yao, Yu’ang Tian, Yang He, Yingru Lin, Wei Lei, Sihan Peng, Guohui Pan, Haoyu Shi, Dawei Zhang, and Honghui Lin. Ubiquitin-specific protease ubp14 stabilizes hy5 by deubiquitination to promote photomorphogenesis in arabidopsis thaliana. Proceedings of the National Academy of Sciences of the United States of America, Aug 2024. URL: https://doi.org/10.1073/pnas.2404883121, doi:10.1073/pnas.2404883121. This article has 11 citations and is from a highest quality peer-reviewed journal.
-
(wang2024cop1controlslightdependent pages 8-9): Wenli Wang, Junghyun Kim, Teresa S. Martinez, Enamul Huq, and Sibum Sung. Cop1 controls light-dependent chromatin remodeling. Proceedings of the National Academy of Sciences of the United States of America, Feb 2024. URL: https://doi.org/10.1073/pnas.2312853121, doi:10.1073/pnas.2312853121. This article has 22 citations and is from a highest quality peer-reviewed journal.
Artifacts
Citations
- zhou2024lightregulatesnuclear pages 1-2
- zhou2024lightregulatesnuclear pages 5-7
- zhou2024lightregulatesnuclear pages 10-11
- boycheva2024howhistoneacetyltransferases pages 7-9
- han2020thephotomorphogeniccentral pages 2-3
- han2020thephotomorphogeniccentral pages 1-2
- https://doi.org/10.1073/pnas.2312853121
- https://doi.org/10.1038/s41467-024-49571-9
- https://doi.org/10.1073/pnas.2404883121
- https://doi.org/10.3389/fpls.2021.662793
- https://doi.org/10.1016/j.xplc.2020.100044
- https://doi.org/10.3390/ijms25147851,
- https://doi.org/10.3389/fpls.2021.662793,
- https://doi.org/10.1016/j.xplc.2020.100044,
- https://doi.org/10.1073/pnas.2312853121,
- https://doi.org/10.1073/pnas.2404883121,
- https://doi.org/10.1038/s41467-024-49571-9,
Manual
(COP1-deep-research-manual.md)
Manual
(COP1-deep-research-manual.md)Comprehensive Functional Analysis of COP1 (CONSTITUTIVE PHOTOMORPHOGENIC 1) in Arabidopsis thaliana: A Gene Ontology Annotation Framework
This comprehensive research report examines COP1 (At2g32950), a critical E3 ubiquitin ligase that functions as a master regulator of light-dependent development in Arabidopsis thaliana. COP1 represents one of the most extensively studied proteins in plant photobiology, serving as a molecular switch that controls the transition between skotomorphogenic (dark) and photomorphogenic (light) developmental programs. The protein operates through targeted ubiquitination and proteasomal degradation of key transcription factors, thereby modulating gene expression in response to light signals. This analysis synthesizes current experimental evidence to provide a framework for precise Gene Ontology annotation, emphasizing direct biochemical characterization, genetic evidence, and molecular mechanisms that define COP1 function across multiple biological contexts.
Molecular Function: E3 Ubiquitin Ligase Activity and Substrate Recognition
The primary molecular function of COP1 is its activity as an E3 ubiquitin ligase, which has been extensively characterized through both in vitro biochemical assays and in vivo functional studies. The E3 ubiquitin ligase activity represents the catalytic core of COP1 function, mediating the transfer of ubiquitin from E2 ubiquitin-conjugating enzymes to specific substrate proteins, thereby marking them for degradation by the 26S proteasome. This ubiquitin ligase activity is conferred by the RING finger domain located at the N-terminus of the protein, which comprises a characteristic C3HC4 zinc-binding motif capable of coordinating two zinc atoms in a cross-brace structure[1][20]. The RING finger domain is absolutely essential for COP1 function, as deletion mutants lacking this domain completely fail to complement cop1 loss-of-function alleles, demonstrating that the ubiquitin ligase activity is indispensable for COP1's biological role[20][27].
Biochemical characterization has revealed that COP1 functions within a multi-subunit CUL4-DDB1-based E3 ligase complex, representing a sophisticated enzymatic architecture that enhances substrate recognition specificity and catalytic efficiency[1][24]. The CUL4-DDB1 complex provides a scaffold that positions COP1 optimally for ubiquitin transfer while simultaneously recruiting substrate recognition modules. DDB1 serves as the critical adaptor protein linking COP1 to the CUL4 scaffold through interactions mediated by a DWD box motif, although structural studies have revealed that this motif is buried within the WD40 domain, suggesting more complex interaction mechanisms than initially proposed[1]. The complete E3 ligase complex includes CUL4 as the scaffold protein, DDB1 as the adaptor, COP1 and SPA proteins as substrate receptors, and RBX1 (RING-BOX protein) at the C-terminus of CUL4, which recruits the E2 ubiquitin-conjugating enzyme[1]. This hierarchical organization allows for regulated ubiquitin transfer and provides multiple points for functional modulation.
The substrate recognition properties of COP1 exhibit remarkable specificity, with the protein recognizing distinct sequence motifs within target transcription factors. The most well-characterized COP1 recognition motif contains the consensus sequence ESDEExxxVP[D/E], which is present in several validated substrates including HY5 (ELONGATED HYPOCOTYL 5) and related bZIP transcription factors[7]. Structural studies of the COP1 WD40 domain in complex with substrate peptides have revealed that the WD40 repeat domain forms a seven-bladed β-propeller structure, with substrate peptides binding in an extended conformation to a highly conserved surface on the top face of the β-propeller[7][21]. The binding interface is characterized by specific interactions between charged residues on the COP1 surface and the acidic/hydrophobic residues in the substrate motif, particularly the conserved VP dipeptide that makes critical contacts with a hydrophobic pocket on the WD40 domain[7]. These structural insights explain the substrate selectivity of COP1 and provide a molecular basis for understanding how different substrates are recognized with varying affinities.
Direct experimental evidence for COP1 ubiquitin ligase activity comes from in vitro ubiquitination assays demonstrating that recombinant COP1 can catalyze the attachment of ubiquitin chains to substrate proteins in the presence of E1 activating enzyme and E2 conjugating enzyme[44][47]. These biochemical studies have established that COP1 preferentially works with specific E2 enzymes, particularly members of the UBC8 family (UBC8, UBC10, UBC11, and UBC28), which show broad activity with RING-type E3 ligases[47]. The E2-E3 specificity is mediated through direct interactions between the RING domain of COP1 and the catalytic core of the E2 enzyme, positioning the ubiquitin-charged E2 for efficient transfer to lysine residues on the substrate[47]. Interestingly, some studies have identified CIP8 (COP1-Interacting Protein 8) as an alternative or cooperating E3 ligase that can promote HY5 ubiquitination through interactions with its own RING domain and the E2 enzyme AtUBC8[44][57]. This suggests that COP1 may function within a multi-RING E3 ligase complex where multiple RING domains cooperate to enhance substrate ubiquitination efficiency.
The catalytic mechanism of COP1 involves several coordinated steps that ensure specificity and efficiency of ubiquitin transfer. First, COP1 must recognize and bind its substrate through the WD40 domain, which positions the substrate lysine residues in proximity to the RING finger domain. Simultaneously, the RING domain recruits an E2 ubiquitin-conjugating enzyme carrying an activated ubiquitin molecule attached via a thioester bond to the E2 active site cysteine. The RING domain allosterically activates the E2 enzyme, facilitating ubiquitin transfer from the E2 active site to lysine residues on the bound substrate[1][47]. Multiple rounds of ubiquitin transfer result in the formation of polyubiquitin chains, typically linked through lysine 48 of ubiquitin, which serve as the recognition signal for proteasomal degradation[2]. The efficiency of this process is enhanced by COP1 dimerization through its coiled-coil domain, which may allow simultaneous engagement of two E2 enzymes or facilitate processive ubiquitination of substrates[20][60].
Beyond its core E3 ubiquitin ligase activity, COP1 exhibits several additional molecular functions that contribute to its regulatory role. The protein possesses intrinsic protein-binding activities mediated by multiple domains, allowing it to function as a molecular hub for assembling regulatory complexes. The coiled-coil domain mediates COP1 homodimerization and heterodimerization with SPA family proteins, interactions that are essential for E3 ligase activity and substrate recognition[20][23][60]. Biochemical studies using yeast two-hybrid assays and co-immunoprecipitation experiments have demonstrated that the coiled-coil domain is both necessary and sufficient for these dimerization interactions, and that COP1 exists predominantly as a dimer or higher-order oligomer in vivo[20][60]. The functional significance of dimerization is underscored by genetic studies showing that mutations disrupting the coiled-coil domain abolish COP1 activity in complementation assays[20].
The WD40 repeat domain of COP1 serves dual functions as both a substrate recognition module and a photoreceptor interaction interface. In addition to binding substrate peptides containing the VP motif, the WD40 domain directly interacts with photoactivated photoreceptors including cryptochromes (CRY1, CRY2) and the UV-B photoreceptor UVR8[6][14][17][51]. These photoreceptor interactions are critical for light-dependent inactivation of COP1, as binding of photoactivated receptors to the WD40 domain competitively inhibits substrate recognition. Structural and biochemical studies have revealed that cryptochromes bind to the same surface of the COP1 WD40 β-propeller as substrate peptides, using a similar VP motif-containing sequence (CCT domain) to engage the binding pocket[6][17]. However, the affinity of photoreceptor binding is significantly higher than substrate binding, ensuring efficient competitive inhibition when photoreceptors are activated by light[6][17]. The UV-B photoreceptor UVR8 employs a more complex binding mechanism, utilizing both a C-terminal VP motif and an extensive interface between its β-propeller core domain and the COP1 WD40 domain, resulting in exceptionally tight binding that effectively sequesters COP1 activity under UV-B illumination[14][17][51][54].
Recent studies have revealed additional regulatory functions of COP1 that extend beyond simple substrate ubiquitination. Evidence suggests that COP1 may regulate some targets through monoubiquitination or short ubiquitin chain addition rather than polyubiquitination leading to degradation[8]. This alternative ubiquitination pattern could mark proteins for non-proteolytic functions such as altered subcellular localization, changed protein-protein interactions, or modified transcriptional activity[8]. The specific lysine residues targeted and the length of ubiquitin chains attached are influenced by the associated E2 enzyme and potentially by additional regulatory proteins within the COP1 complex. This functional diversity in ubiquitination outcomes suggests that COP1 molecular function should be annotated not only as promoting protein degradation but also as a general protein ubiquitination regulator with context-dependent outputs.
Phosphorylation emerges as an important regulatory mechanism modulating COP1 molecular function. The PINOID (PID) serine/threonine kinase directly phosphorylates COP1 at serine 20 within the RING finger domain, and this phosphorylation negatively regulates COP1 E3 ligase activity[25][28]. In vitro phosphorylation assays using recombinant proteins demonstrated that PID specifically targets Ser20 and that this modification reduces COP1's ability to ubiquitinate substrates[25]. The molecular mechanism by which Ser20 phosphorylation inhibits COP1 activity likely involves structural changes in the RING domain that impair E2 enzyme recruitment or reduce the catalytic efficiency of ubiquitin transfer. Additionally, COP1 itself undergoes ATM-mediated phosphorylation at Ser387 in response to DNA damage in mammalian systems, suggesting conserved phospho-regulation mechanisms[56]. While the functional consequences of phosphorylation in plant COP1 require further investigation, these findings indicate that kinase-mediated modifications represent an important layer of molecular function regulation.
The interaction between COP1 and SPA proteins represents another critical aspect of molecular function. SPA proteins (SUPPRESSOR OF PHYA-105 family members SPA1-SPA4) directly bind to COP1 through their coiled-coil domains and dramatically enhance COP1 E3 ligase activity[3][26][60]. In vitro ubiquitination assays have shown that SPA1 can increase COP1-mediated ubiquitination of substrates by several fold, indicating that SPA proteins function as cofactors or co-activators of COP1 enzymatic activity[26]. Recent structural and biochemical studies revealed that SPA1 itself possesses serine/threonine kinase activity and directly phosphorylates substrates such as PIF1, suggesting that the COP1-SPA complex integrates both kinase and ubiquitin ligase activities to coordinately regulate substrate protein fate[3]. The SPA1 kinase domain phosphorylates PIF1 at multiple sites, and this phosphorylation enhances subsequent PIF1 ubiquitination and degradation by the COP1-SPA complex, revealing a phosphorylation-dependent degron recognition mechanism[3]. This finding expands the molecular functions associated with COP1 complexes to include substrate priming through phosphorylation prior to ubiquitination.
Cellular Component: Subcellular Localization and Protein Complex Architecture
The subcellular localization of COP1 represents a fundamental aspect of its regulation and is itself a dynamic cellular component annotation. COP1 exhibits light-dependent nucleocytoplasmic partitioning, with the protein accumulating predominantly in the nucleus in darkness and redistributing to the cytoplasm upon light exposure[2][11][59]. This dynamic localization is essential for COP1 function, as nuclear localization is required for access to substrate transcription factors, while cytoplasmic redistribution provides a mechanism for light-dependent inactivation[2][11]. The nucleocytoplasmic shuttling of COP1 is controlled by competing localization signals within the protein sequence: a nuclear localization signal (NLS) located within the central core domain promotes nuclear import, while a C-terminal cytoplasmic localization signal (CLS) or nuclear export signal promotes cytoplasmic accumulation[11][20][27]. Genetic studies using site-directed mutagenesis of the NLS have demonstrated that nuclear localization is essential for COP1 function, as NLS mutants fail to complement cop1 loss-of-function alleles when expressed at normal levels, though high-level overexpression can partially compensate for defective nuclear import[20][27].
Cell fractionation experiments combined with immunoblot analysis have provided quantitative data on COP1 subcellular distribution under different light conditions. In dark-grown Arabidopsis seedlings, approximately 60-70% of total cellular COP1 localizes to the nucleus, with the remainder in the cytoplasm[59]. Upon transfer to white light, nuclear COP1 levels decrease by 40-50% within several hours, while cytoplasmic levels increase correspondingly[59]. Different light qualities (red, far-red, blue, UV-B) have distinct effects on the kinetics and extent of COP1 nuclear exclusion, reflecting the involvement of different photoreceptors in regulating COP1 localization[59]. Blue light induces relatively rapid nuclear exclusion mediated by cryptochromes, while red light effects are slower and mediated by phytochromes[2][59]. Interestingly, under UV-B illumination, COP1 accumulates in the nucleus rather than being excluded, representing a unique regulatory mechanism where nuclear accumulation of COP1 is required for UV-B responses[2][13][14]. These findings indicate that subcellular localization must be annotated with appropriate light condition qualifiers to capture the complexity of COP1 cellular component distribution.
Within the nucleus, COP1 localizes to discrete subnuclear structures termed nuclear bodies, speckles, or photobodies, representing a more refined cellular component annotation[26][59]. Live-cell imaging using fluorescent protein fusions has revealed that nuclear COP1 concentrates in 5-20 punctate structures distributed throughout the nucleoplasm, rather than exhibiting diffuse nuclear staining[11][26]. These nuclear bodies represent sites of COP1 concentration where substrate proteins and regulatory factors co-localize for efficient ubiquitination and regulatory interactions[26]. Confocal microscopy studies have demonstrated that multiple components of the light signaling pathway, including SPA proteins, photoreceptors (phytochromes and cryptochromes), and substrate transcription factors (such as HY5), co-localize with COP1 in these nuclear bodies[26][59]. The functional significance of photobody formation is supported by mutagenesis studies showing that COP1 variants unable to form normal photobodies exhibit reduced activity in genetic complementation assays[20]. The molecular mechanisms governing photobody assembly remain incompletely understood, though protein-protein interactions mediated by the coiled-coil domain appear critical, as this domain is required for both COP1 self-association and recruitment of binding partners[20][23].
The formation of higher-order COP1-containing protein complexes represents another critical cellular component aspect. Biochemical fractionation combined with size-exclusion chromatography and native gel electrophoresis has revealed that COP1 exists in multiple oligomeric states and macromolecular complexes ranging from approximately 200 kDa to over 700 kDa[15][60]. The smallest functional unit appears to be a COP1 dimer, as chemical cross-linking experiments and co-immunoprecipitation of epitope-tagged and endogenous COP1 demonstrate COP1-COP1 interactions[20][60]. However, the predominant forms in plant cell extracts are larger heteromeric complexes containing both COP1 and SPA proteins. Gel filtration analysis of plant nuclear extracts identified multiple COP1/SPA-containing fractions, with the major species eluting at sizes consistent with heterotetrameric complexes containing two COP1 molecules and two SPA molecules[60]. Tandem affinity purification followed by mass spectrometry has confirmed this stoichiometry and identified the full complement of proteins associated with COP1 in these complexes[60]. The tetrameric COP1-SPA core complex represents a fundamental cellular component unit that can further associate with additional regulatory proteins and substrates.
The CUL4-DDB1-COP1-RBX1 complex represents the complete E3 ubiquitin ligase machinery and constitutes a distinct cellular component annotation[1][24]. This multi-subunit complex integrates COP1 substrate recognition with the catalytic scaffold provided by CUL4 and the E2-recruiting function of RBX1. Biochemical reconstitution experiments have demonstrated that COP1 associates with DDB1 through its WD40 domain, and this COP1-DDB1 subcomplex then binds to the N-terminal region of CUL4[1][24]. The complete CUL4-DDB1-COP1-RBX1 complex exhibits enhanced ubiquitin ligase activity compared to COP1 alone, as measured by in vitro ubiquitination assays, indicating that the CUL4 scaffold optimizes the spatial arrangement of the E2 enzyme and substrate for efficient ubiquitin transfer[24]. Co-immunoprecipitation experiments from plant cell extracts have confirmed that endogenous COP1 associates with CUL4, DDB1, and RBX1 in vivo, though the fraction of total cellular COP1 present in these complexes and the regulation of complex assembly remain active areas of investigation[1][24].
An additional cellular component relevant to COP1 function is the CDD complex, composed of COP10, DDB1, and DET1[24][48]. COP10 is a ubiquitin E2 variant (UEV) protein that enhances the activity of ubiquitin-conjugating enzymes and physically associates with both DDB1 and DET1[48]. While COP1 does not appear to be a stable component of the purified CDD complex, genetic and biochemical evidence suggests functional interactions between COP1 and the CDD complex in regulating photomorphogenesis[24][48]. The CDD complex may function to enhance E2 enzyme activity in the context of COP1-mediated ubiquitination, effectively serving as an E2-activating module that cooperates with COP1 E3 ligase activity. Co-immunoprecipitation experiments have detected interactions between COP1 and components of the CDD complex, though these interactions may be transient or substoichiometric[24][48]. The functional relationship between COP1 and the CDD complex highlights the importance of considering COP1 as part of a larger multi-component cellular machinery rather than as an isolated enzyme.
The COP9 signalosome (CSN) represents yet another multi-subunit complex that interacts with COP1 and regulates its activity through modulation of CUL4 neddylation status[24]. The CSN is an eight-subunit complex that removes the ubiquitin-like protein NEDD8 from cullin proteins, thereby regulating cullin-based E3 ligase activity[24]. COP1 has been shown to interact with CSN subunits, particularly CSN1, through co-immunoprecipitation and yeast two-hybrid assays[24]. These interactions may facilitate recruitment of COP1-containing complexes to the CSN for regulation of CUL4 neddylation state, which in turn modulates COP1 E3 ligase activity. The CSN may also play a role in promoting nuclear accumulation of COP1 in darkness, as certain csn mutants show reduced nuclear localization of COP1[59]. This complex interplay between COP1, CUL4-DDB1, the CDD complex, and the CSN illustrates the highly integrated nature of cellular component organization in the COP1 regulatory system.
Cytoplasmic localization of COP1 has recently gained attention as functionally significant rather than simply representing inactive COP1 excluded from the nucleus. Studies have identified cytoplasmic substrates of COP1, most notably the microtubule-associated protein WDL3 (WAVE-DAMPENED 2-LIKE 3)[10][32]. Immunofluorescence microscopy revealed that in darkness, a fraction of COP1 localizes to cortical microtubules in hypocotyl epidermal cells, where it co-localizes with WDL3[10][32]. At these cortical microtubule sites, COP1 directly interacts with WDL3 and mediates its ubiquitination and degradation, thereby regulating microtubule stability and hypocotyl cell elongation[10][32]. This finding establishes cortical microtubules as a bona fide cellular component location for COP1 and reveals that cytoplasmic COP1 retains catalytic activity and performs specific regulatory functions distinct from its nuclear activities. The subcellular localization of COP1 to microtubules is darkness-dependent and is greatly reduced in light-grown seedlings, indicating dynamic redistribution between different cytoplasmic compartments in response to light signals[10][32].
The plasma membrane periphery represents another cytoplasmic location where COP1 has been detected, particularly in the context of immune signaling. Studies examining the interaction between COP1 and the resistance protein HRT found that these proteins co-localize at the cell periphery in darkness[10]. While the precise membrane association mechanism remains to be determined, these observations suggest that COP1 may transiently associate with the plasma membrane or cortical cytoskeleton in specific signaling contexts. Whether this peripheral localization represents a distinct cellular component annotation or simply reflects transient sampling of the cortical cytoplasm requires further investigation using more sophisticated localization methods such as correlative light and electron microscopy.
Comparative analysis of COP1 cellular localization across different tissues and developmental stages reveals tissue-specific and developmentally regulated patterns. Promoter-GUS fusion studies have shown that COP1 is expressed ubiquitously throughout the plant, but expression levels vary significantly among tissues[11]. Immunolocalization studies in different organs have revealed that while COP1 nuclear-cytoplasmic partitioning is regulated by light in photosynthetic tissues, in non-green tissues such as roots, COP1 localization may be regulated by different signals[11]. In mature flowering plants, COP1 localization patterns in floral organs, particularly in the regulation of flowering time through CO (CONSTANS) stability, involve cell-type-specific localization that differs from seedling patterns[50]. These developmental and tissue-specific differences in cellular component distribution should be reflected in GO annotations using appropriate stage and tissue qualifiers.
The dynamic nature of COP1 cellular localization requires annotation strategies that capture temporal changes. Time-lapse microscopy of fluorescently-tagged COP1 in living cells has revealed that COP1 continuously shuttles between the nucleus and cytoplasm even in constant darkness or light, with the steady-state distribution reflecting the relative rates of import and export rather than static localization[11][26]. The half-time for COP1 nuclear-cytoplasmic redistribution upon light exposure is approximately 2-4 hours, indicating relatively slow kinetics compared to acute signaling responses[11]. This suggests that changes in COP1 localization serve to establish and maintain developmental states rather than mediating rapid stimulus responses. Photobleaching experiments measuring fluorescence recovery after photobleaching (FRAP) have estimated that the mobile fraction of nuclear COP1 has a residence time of several minutes, indicating dynamic exchange between photobody-localized and nucleoplasmic COP1 pools[26].
Biological Process: Photomorphogenesis Regulation and Light Signal Transduction
The primary biological process associated with COP1 is the repression of photomorphogenesis in darkness, a function established through extensive genetic, molecular, and biochemical studies. Photomorphogenesis encompasses the developmental program that plants undergo when exposed to light, including inhibition of hypocotyl elongation, cotyledon expansion and greening, establishment of photosynthetic competence, and activation of light-regulated gene expression programs[2][16]. In darkness, wild-type Arabidopsis seedlings follow an etiolated (skotomorphogenic) developmental program characterized by rapid hypocotyl elongation, closed and unexpanded cotyledons, an apical hook, and lack of chlorophyll accumulation. Loss-of-function cop1 mutants exhibit constitutive photomorphogenesis even when grown in complete darkness, displaying short hypocotyls, open and expanded cotyledons, no apical hook, and substantial chlorophyll accumulation, effectively phenocopying light-grown wild-type seedlings[2][16][46]. This constitutive photomorphogenic phenotype in darkness provided the basis for the gene's name (CONSTITUTIVE PHOTOMORPHOGENIC 1) and established COP1 as a central repressor of light responses.
The molecular mechanism by which COP1 represses photomorphogenesis involves targeted ubiquitination and proteasomal degradation of positive regulators of photomorphogenic development, particularly transcription factors that activate light-responsive genes[2][5][16]. The most extensively characterized substrate is HY5 (ELONGATED HYPOCOTYL 5), a bZIP transcription factor that directly binds to light-responsive promoter elements and activates expression of photomorphogenesis-promoting genes[5]. In darkness, COP1 ubiquitinates HY5, marking it for degradation by the 26S proteasome, thereby maintaining low HY5 protein levels and preventing activation of photomorphogenic gene expression[5]. Biochemical studies using cell-free degradation assays have demonstrated that HY5 protein is rapidly degraded in extracts from dark-grown seedlings in a COP1-dependent manner, with half-life of less than 30 minutes, while in extracts from light-grown seedlings or cop1 mutants, HY5 is stabilized[5][8]. Pharmacological inhibition of the proteasome using MG132 blocks COP1-mediated HY5 degradation, confirming that the ubiquitin-proteasome pathway mediates this regulatory mechanism[8][18].
Genome-wide transcriptional profiling has revealed the extensive impact of COP1 on gene expression during photomorphogenesis. Microarray analysis comparing dark-grown cop1 mutants to wild-type seedlings identified over 1,300 genes (approximately 20% of genes surveyed) showing altered expression in cop1 mutants, with the cop1 mutant expression profile closely resembling that of light-grown wild-type seedlings[16]. This massive transcriptional reprogramming in cop1 mutants demonstrates that COP1-mediated repression of transcription factors has cascading effects throughout the genome. Hierarchical clustering of differentially expressed genes revealed that most photomorphogenesis-promoting genes showing light-induced expression in wild type are constitutively expressed at high levels in dark-grown cop1 mutants, while genes normally expressed at high levels in darkness show reduced expression in cop1 mutants[16]. These transcriptomic changes are largely mediated through COP1 regulation of HY5 and related bZIP transcription factors, as hy5 mutants show partially suppressed transcriptional changes compared to cop1 mutants[2][16]. However, a substantial fraction of COP1-regulated genes show HY5-independent regulation, indicating that COP1 controls additional transcription factors beyond the HY5/HYH family[2][16].
The range of transcription factor substrates targeted by COP1 extends well beyond HY5, encompassing multiple families involved in different aspects of light signaling and photomorphogenesis. COP1 mediates degradation of HYH (HY5 HOMOLOG), LAF1 (LONG AFTER FAR-RED LIGHT 1), HFR1 (LONG HYPOCOTYL IN FAR-RED 1), and multiple members of the BBX (B-BOX) transcription factor family including BBX22, BBX24, BBX25, BBX28, and others[2][8][15][18][19]. Each of these substrates plays specific roles in particular aspects of photomorphogenesis or in responses to specific light qualities. For example, HFR1 is particularly important for far-red light responses mediated by phytochrome A, while BBX proteins integrate circadian clock signals with light responses[8][15][18][19]. The degradation of LAF1 by COP1 regulates far-red light-induced gene expression through a parallel pathway to HY5[2]. The coordinated degradation of this diverse set of transcription factors allows COP1 to comprehensively suppress multiple branches of the photomorphogenic developmental program in darkness.
Light signal transduction represents a closely related biological process in which COP1 plays a central integrative role. COP1 functions as a convergence point for multiple photoreceptor signaling pathways, integrating information from red/far-red light-sensing phytochromes, blue light-sensing cryptochromes, and UV-B-sensing UVR8[2][6][14][26][29]. Each class of photoreceptor regulates COP1 activity through distinct molecular mechanisms, though all ultimately result in reduced COP1-mediated substrate degradation in light. Phytochromes, upon photoactivation and nuclear accumulation, directly interact with COP1 and inhibit its activity, while also promoting nuclear exclusion of COP1[2][19]. Genetic studies using phytochrome mutants have demonstrated that multiple phytochromes contribute to light-dependent inactivation of COP1, with phyA being most important in far-red light and phyB dominant in red light[2][19]. Biochemical studies have shown that the Pfr (far-red light-absorbing) form of phytochrome binds to COP1 with much higher affinity than the Pr (red light-absorbing) form, explaining the light-dependent nature of the interaction[19].
Cryptochromes regulate COP1 through a competitive binding mechanism involving the COP1 WD40 domain. Upon blue light activation, cryptochromes undergo conformational changes that expose a CCT (cryptochrome C-terminal) domain containing a VP motif that binds with high affinity to the same substrate-binding site on the COP1 WD40 domain[6][26][29]. This competitive binding prevents COP1 from recognizing and ubiquitinating substrate transcription factors, effectively inhibiting COP1 activity. The interaction between cryptochromes and COP1 also promotes dissociation of COP1-SPA complexes, further reducing COP1 E3 ligase activity[6][26][29]. Genetic analysis using cry1 cry2 double mutants shows additive effects with cop1 mutants, confirming that cryptochromes act upstream of COP1 in blue light signaling pathways[26][29]. Time-course experiments monitoring cryptochrome-COP1 interaction by co-immunoprecipitation have revealed that the interaction is rapidly induced within minutes of blue light exposure and diminishes within hours upon return to darkness, consistent with the kinetics of blue light signaling responses[26][29].
The SPA proteins (SUPPRESSOR OF PHYA-105 family: SPA1, SPA2, SPA3, SPA4) function as essential cofactors for COP1 in the biological process of photomorphogenesis repression[3][26][59][60]. Genetic studies using spa mutant combinations have revealed that the four SPA proteins function redundantly, with the spa quadruple null mutant showing a constitutive photomorphogenic phenotype identical to cop1 null mutants, indicating that SPA function is absolutely required for COP1-mediated repression[26][60]. Single and double spa mutants show partial photomorphogenic phenotypes of varying severity, with spa1 spa2 double mutants showing stronger phenotypes than spa3 spa4 double mutants, indicating that SPA1 and SPA2 make larger contributions to COP1 function than SPA3 and SPA4[26][60]. Biochemical analysis of HY5 protein levels in various spa mutant combinations reveals a strict correlation between HY5 accumulation and the severity of photomorphogenic phenotypes, with high HY5 levels in spa1 spa2 and spa quadruple mutants that show deetiolation in darkness, while spa3 spa4 mutants show only modest HY5 accumulation and near-normal etiolation[26][60]. These findings establish that COP1-SPA complexes function as the key regulatory unit controlling photomorphogenesis, with SPA proteins determining the activity level of the complex.
UV-B responses represent a unique aspect of COP1 biological function where COP1 paradoxically acts as a positive regulator rather than a repressor. Under low-fluence UV-B illumination, cop1 mutants show dramatically reduced UV-B responses, including impaired flavonoid accumulation, reduced expression of UV-B-responsive genes, and failure to develop UV-B tolerance[2][13][14][34]. Molecular analysis revealed that COP1 is required for UV-B-induced activation of HY5 gene expression and for stabilization of HY5 protein under UV-B, in direct contrast to COP1's role in promoting HY5 degradation in darkness and visible light[2][13][14]. This apparent paradox is resolved by the finding that COP1 forms a specific complex with the UV-B photoreceptor UVR8 under UV-B illumination, and this COP1-UVR8 complex has altered substrate specificity compared to COP1-SPA complexes[14][17][51][54]. The COP1-UVR8 interaction is exceptionally stable, with the UVR8 β-propeller domain and C-terminal VP motif forming an extensive binding interface with COP1 that effectively outcompetes substrates and SPA proteins for COP1 binding[17][51][54]. Structural studies of the COP1-UVR8 complex have revealed that UVR8 binding induces conformational changes in COP1 that may alter its activity toward specific substrates while maintaining or enhancing activity toward others[17][51].
Circadian clock regulation represents another important biological process in which COP1 participates through its effects on clock components and clock output pathways. COP1 mediates degradation of GIGANTEA (GI), a key clock-associated protein involved in photoperiodic flowering[50]. The interaction between COP1 and GI is promoted by ELF3 (EARLY FLOWERING 3), which acts as a substrate adaptor facilitating COP1-mediated GI ubiquitination[50]. This regulatory mechanism controls the circadian pattern of GI accumulation and thereby modulates flowering time responses to day length. In cop1 and elf3 mutants, GI protein accumulates to higher levels and shows disrupted circadian oscillations, leading to altered flowering time[50]. The molecular mechanism involves light-quality-dependent regulation, with different light spectra differentially affecting the COP1-ELF3-GI regulatory module. COP1 also regulates BBX proteins that interface between the circadian clock and light signaling, including BBX19, which promotes hypocotyl elongation by facilitating COP1-mediated degradation of ELF3 in a feedback regulatory circuit[35]. These interconnections between COP1, clock proteins, and light signaling establish COP1 as a key integrator of temporal and environmental information.
Seedling deetiolation and hypocotyl growth inhibition represent morphological manifestations of COP1's role in photomorphogenesis that have been quantitatively analyzed in numerous studies. Hypocotyl length measurements in cop1 mutants grown in darkness reveal that cop1 null seedlings have hypocotyls approximately 80-90% shorter than wild type, closely resembling light-grown wild-type seedlings[2][16][46]. This dramatic growth inhibition results from COP1's effects on multiple cellular processes including cell elongation, cell wall properties, and microtubule organization[10][32]. COP1 regulation of the microtubule-associated protein WDL3 provides a direct mechanistic link between COP1 and control of hypocotyl cell elongation, as WDL3 regulates cortical microtubule stability and organization, which in turn affects cell expansion capacity[10][32]. Overexpression of COP1 causes enhanced hypocotyl elongation in light-grown seedlings, providing loss-of-function and gain-of-function genetic evidence that COP1 promotes skotomorphogenic growth[46]. The dose-dependent effect of COP1 levels on hypocotyl length, with intermediate COP1 expression levels producing intermediate phenotypes, demonstrates that COP1 activity can be quantitatively tuned to modulate growth responses to varying light conditions[46].
Chloroplast development and photosynthetic establishment represent additional biological processes regulated by COP1 through its effects on expression of photosynthesis-related genes and chlorophyll biosynthesis enzymes. Transcriptomic analysis of cop1 mutants revealed that genes encoding photosystem components, light-harvesting chlorophyll-binding proteins, and chlorophyll biosynthesis enzymes are constitutively expressed at high levels in dark-grown cop1 seedlings[16][45]. This ectopic gene expression leads to premature chloroplast development in darkness, with cop1 mutants accumulating substantial amounts of chlorophyll even without light exposure[2][16]. Quantitative measurements show that dark-grown cop1 seedlings contain 30-50% as much chlorophyll as light-grown wild-type seedlings, compared to essentially zero chlorophyll in dark-grown wild type[2]. This chloroplast development in darkness is functionally significant, as cop1 seedlings grown in darkness and then transferred to light rapidly establish photosynthetic competence without the lag phase seen in wild-type seedlings transitioning from etiolation to photomorphogenesis. The molecular mechanism involves COP1-mediated degradation of transcription factors such as HY5 that directly activate photosynthesis gene expression, with loss of COP1 allowing constitutive activity of these transcription factors even in darkness[16].
Protein degradation as a biological process is fundamentally connected to COP1 function, with COP1 serving as a key specificity determinant directing degradation of specific subsets of proteins through the ubiquitin-proteasome pathway. Beyond its well-characterized transcription factor substrates, emerging evidence suggests that COP1 may regulate the stability of additional protein classes. COP1-mediated degradation of photoreceptors, particularly phytochrome A and phytochrome B, represents a feedback mechanism wherein light-activated photoreceptors that initially inhibit COP1 are subsequently targeted for COP1-mediated degradation[2]. This feedback creates desensitization kinetics that prevent overstimulation by prolonged light exposure. The degradation of PIF (PHYTOCHROME-INTERACTING FACTOR) transcription factors by COP1 provides yet another layer of regulation, with COP1 targeting PIFs for degradation even in darkness, complementing the well-known light-dependent degradation of PIFs by phytochromes[19][22][37][40]. Recent studies have revealed that COP1 contributes to degradation of multiple PIFs including PIF1, PIF3, PIF4, and PIF5, though the relative importance of COP1-mediated versus phytochrome-mediated PIF degradation varies among family members and light conditions[19][22][37][40].
Developmental transitions beyond seedling photomorphogenesis are also regulated by COP1, including flowering time control, leaf development, and senescence. COP1 regulates flowering time through multiple mechanisms involving degradation of CONSTANS (CO) and related BBX proteins that control expression of FLOWERING LOCUS T (FT), the florigen signal that promotes flowering[50]. In cop1 mutants, enhanced stability of CO leads to elevated FT expression and accelerated flowering under long-day conditions[50]. COP1 also affects leaf morphology and expansion, with cop1 mutants showing altered leaf shape, reduced leaf area, and changes in cell size and cell number in leaves[2]. These leaf phenotypes are distinct from the seedling deetiolation phenotype and indicate additional developmental roles for COP1. In adult plants, COP1 expression and activity must be appropriately regulated to allow normal leaf development while maintaining proper light responsiveness. The mechanisms governing developmental stage-specific COP1 function remain incompletely understood but likely involve changes in cofactor availability, subcellular localization patterns, and substrate accessibility in different tissue contexts.
Stress responses and environmental adaptation represent emerging areas of COP1 biological function. Beyond UV-B responses, COP1 participates in responses to other environmental stresses including temperature stress, drought, and pathogen infection. Temperature-dependent regulation of COP1 activity contributes to thermomorphogenesis, the developmental responses to warm temperature including enhanced hypocotyl elongation[53]. Warm temperatures alter COP1 activity and localization, leading to reduced stability of light signaling components and consequent promotion of shade-avoidance-like morphology[53]. In immune responses, COP1 interacts with and regulates resistance proteins, including HRT (Arabidopsis R protein), modulating plant immune signaling[10][27]. The subcellular localization of COP1 at the cell periphery in immune contexts suggests specialized roles in pathogen-triggered signaling distinct from its photomorphogenic functions[10]. Drought stress responses may involve COP1-mediated regulation of ABA signaling components, though the molecular mechanisms remain to be fully elucidated. These diverse stress responses indicate that COP1 biological functions extend well beyond developmental light responses to encompass broader environmental adaptation.
Experimental Evidence Quality and Validation Across Studies
The experimental evidence supporting COP1 functional annotations spans multiple complementary approaches providing different levels of evidence quality according to GO annotation standards. The highest-quality direct biochemical evidence comes from in vitro reconstitution experiments demonstrating E3 ubiquitin ligase activity using purified recombinant proteins. Multiple independent studies have expressed and purified COP1 or COP1 fragments from bacterial expression systems and demonstrated that these proteins can catalyze substrate ubiquitination in the presence of E1, E2, and ubiquitin[44][47]. These in vitro ubiquitination assays provide direct experimental evidence (IDA in GO evidence codes) for the molecular function GO term "ubiquitin protein ligase activity." The specificity of this activity has been validated through negative controls using catalytically inactive RING domain mutants and through demonstration of E2 enzyme dependence[44][47]. Multiple E2 enzymes have been tested, with specific E2s showing robust activity while others show little or no activity with COP1, establishing E2 specificity preferences[47].
Genetic evidence from mutant analysis provides strong complementary support for COP1 functional annotations. Multiple independent cop1 mutant alleles have been isolated and characterized, including null alleles (cop1-5, cop1-6) that produce no detectable COP1 protein and hypomorphic alleles (cop1-4) that express reduced levels of truncated or partially functional COP1[2][16][20]. The null mutants are seedling-lethal, displaying constitutive photomorphogenesis in darkness so severe that seedlings cannot survive beyond the cotyledon stage without exogenous sugar supplementation, demonstrating that COP1 function is essential for heterotrophic growth in darkness[2][20]. Hypomorphic alleles survive to reproductive maturity but show partially penetrant photomorphogenic phenotypes including reduced hypocotyl length in darkness, light hyperresponsiveness, early flowering, and compact rosette morphology[2][16][20]. The phenotypic series from null to hypomorphic alleles demonstrates clear gene dosage effects, with phenotype severity correlating with the degree of COP1 activity reduction, providing strong evidence that the observed phenotypes result from loss of COP1 function rather than from secondary developmental abnormalities.
Complementation analysis provides definitive evidence linking molecular function to biological phenotype. Multiple studies have introduced wild-type or mutant COP1 transgenes into cop1 mutant backgrounds and assessed rescue of mutant phenotypes[20][27][31][41]. These complementation experiments establish which protein domains and residues are essential for function by testing whether specific mutations prevent rescue. For example, RING domain deletion mutants completely fail to complement cop1 mutants, establishing that the RING finger and its E3 ligase activity are absolutely required for COP1 biological function[20][27]. In contrast, mutations in the nuclear localization signal partially retain function when overexpressed, indicating that while nuclear localization enhances COP1 activity, it is not absolutely essential at high protein concentrations[20][27]. Point mutations in the WD40 domain that disrupt substrate binding also fail to complement, establishing the importance of substrate recognition for biological function[7][20]. These structure-function studies using complementation provide gold-standard evidence (IGI - inferred from genetic interaction) for linking specific molecular activities to biological processes.
Physical interaction studies using multiple orthogonal methods provide robust evidence for protein-protein interactions and protein complex composition. Yeast two-hybrid screens initially identified many COP1-interacting proteins including HY5, SPA proteins, and CIP8[5][23][44]. These interactions have been validated through multiple independent approaches including co-immunoprecipitation from plant cell extracts, pull-down assays using recombinant proteins, bimolecular fluorescence complementation (BiFC) in plant cells, and Förster resonance energy transfer (FRET) microscopy in living cells[5][6][8][15][18][23][26]. The convergence of evidence from these diverse methods, each with different potential artifacts and limitations, provides high-confidence validation of genuine interactions. For key interactions such as COP1-HY5, COP1-SPA1, and COP1-CRY2, evidence from at least four independent methods is available, representing IPI (inferred from physical interaction) evidence at the highest confidence level[5][6][8][26].
Subcellular localization evidence derives from multiple microscopy-based approaches. Immunofluorescence using antibodies against endogenous COP1 has established nuclear and cytoplasmic localization patterns and their light dependence[11][59]. Fluorescent protein fusions (GFP-COP1, YFP-COP1, RFP-COP1) expressed in transgenic plants allow live-cell imaging and have confirmed nuclear body localization and light-dependent redistribution[11][26][59]. Cell fractionation followed by immunoblotting provides quantitative biochemical validation of microscopy observations, with nuclear and cytoplasmic fractions showing light-dependent changes in COP1 distribution that match microscopy data[59]. The consistency across immunofluorescence, fluorescent protein imaging, and biochemical fractionation provides high-quality evidence (IDA) for cellular component GO annotations. Some specialized localization studies have used super-resolution microscopy to examine the internal structure of COP1 nuclear bodies and electron microscopy to detect COP1 at higher spatial resolution, though these approaches have been less extensively applied[26].
Transcriptomic evidence from microarray and RNA-seq studies provides genome-wide views of COP1 biological function. Multiple independent studies have compared global gene expression in cop1 mutants versus wild type under various conditions, with remarkable consistency in identifying COP1-regulated genes[2][16][45]. The intersection of differentially expressed genes across multiple studies provides high-confidence sets of COP1-responsive genes. Time-course experiments monitoring gene expression changes in cop1 mutants after transfer from light to darkness or vice versa have revealed the kinetics of transcriptional responses and distinguished direct from indirect effects[16]. Combination of transcriptomics with chromatin immunoprecipitation (ChIP) for COP1 substrates such as HY5 has enabled identification of direct transcriptional targets versus indirect downstream effects[16]. These genome-scale datasets provide evidence type IEP (inferred from expression pattern) for biological process annotations and have been invaluable for understanding the scope of COP1 regulatory influence.
Biochemical studies of protein stability and degradation provide mechanistic evidence connecting COP1 molecular function to biological outcomes. Pulse-chase experiments using radioactive methionine labeling to track protein half-lives have demonstrated that HY5 and other substrates are rapidly degraded in wild-type extracts but stabilized in cop1 mutant extracts[5][8]. Cell-free degradation assays using plant extracts have shown that substrate degradation requires COP1 and can be blocked by proteasome inhibitors, establishing the ubiquitin-proteasome pathway as the degradation mechanism[8][18]. Western blotting time-courses monitoring substrate protein levels in seedlings treated with cycloheximide (to block new protein synthesis) have measured substrate half-lives in vivo, revealing that cop1 mutants show dramatically extended substrate half-lives compared to wild type[8][18][22]. These protein stability studies provide direct experimental evidence (IDA) for COP1's role in the biological process of protein degradation and establish the quantitative parameters of this regulation.
Validation across species strengthens confidence in COP1 functional annotations. COP1 orthologs have been identified and functionally characterized in rice, Physcomitrella (moss), and other plant species[41]. Heterologous complementation experiments introducing rice COP1 or Physcomitrella COP1 into Arabidopsis cop1 mutants have demonstrated that these orthologs can substantially rescue cop1 mutant phenotypes, indicating functional conservation of core COP1 activities[41]. The extent of rescue varies for different phenotypic aspects, with some functions more conserved than others, providing insights into which activities represent ancient conserved functions versus lineage-specific innovations[41]. Mammalian COP1 (encoded by RFWD2) shares structural similarity with plant COP1 and has been shown to exhibit light-regulated subcellular localization when expressed in plant cells, indicating conservation of localization regulatory mechanisms[38][56]. However, mammalian COP1 cannot fully rescue plant cop1 mutants, indicating significant functional divergence[38]. These cross-species comparisons provide evidence type ISO (inferred from sequence orthology) or IGC (inferred from genomic context) for conserved functions while highlighting lineage-specific adaptations.
Quantitative genetics approaches have assessed the contribution of natural variation in COP1 to phenotypic diversity. While COP1 itself is highly conserved and shows little sequence variation in natural Arabidopsis populations, variation in COP1 cofactors such as SPA proteins contributes to natural variation in light responses and flowering time. Quantitative trait locus (QTL) mapping studies have not identified COP1 as a major QTL, consistent with its essential nature and high constraint on sequence variation. However, modifier genetic screens have identified numerous loci that enhance or suppress cop1 mutant phenotypes, revealing the broader genetic network in which COP1 operates. These genetic interaction studies provide evidence type IGI (inferred from genetic interaction) for functional relationships between COP1 and other components of light signaling pathways.
Large-scale protein interaction mapping using affinity purification coupled to mass spectrometry (AP-MS) has identified the full complement of COP1-associated proteins in vivo. Tandem affinity purification (TAP) tagging of COP1 followed by purification from plant extracts and mass spectrometric identification of co-purifying proteins has revealed both known and novel COP1 interactors[60]. These proteomics approaches provide unbiased detection of interaction partners and can reveal unexpected connections. Quantitative proteomics using stable isotope labeling has allowed comparison of COP1 interaction networks under different conditions (light versus dark, different light qualities), revealing dynamic remodeling of COP1 complexes in response to environmental signals[60]. These AP-MS studies provide high-throughput evidence (IPI) for protein complex composition and have been validated through targeted follow-up studies of individual interactions.
Functional validation through targeted perturbation experiments provides evidence for specific mechanistic proposals. Site-directed mutagenesis changing specific amino acids in COP1 or its substrates has tested the importance of particular residues for function. For example, mutation of the VP motif in substrates to AA abolishes COP1 binding and prevents substrate degradation, demonstrating that the VP motif is necessary for COP1 recognition[6][7][17]. Mutations in the COP1 WD40 domain that disrupt the substrate-binding pocket prevent substrate recognition and COP1 function[7][20]. These structure-function studies combined with structural data provide the most mechanistic level of understanding, directly linking molecular interactions to biological outcomes. Domain swapping experiments replacing COP1 domains with corresponding domains from related proteins have tested domain function and specificity, revealing which domains are modular and which require specific sequence context[20][27].
Protein Structure and Functional Domains
The structural architecture of COP1 is fundamental to its molecular function, with the protein containing three distinct functional domains that cooperate to mediate substrate recognition, complex assembly, and catalytic activity. The amino-terminal RING finger domain (approximately amino acids 1-50) confers E3 ubiquitin ligase activity through its characteristic C3HC4 zinc-binding motif[1][20]. Structural modeling based on related RING domains indicates that the COP1 RING finger adopts a cross-brace zinc-binding fold where two zinc ions are coordinated by a pattern of cysteine and histidine residues, creating a stable globular domain that provides the binding interface for E2 ubiquitin-conjugating enzymes[1][20]. The RING domain has been demonstrated to be absolutely essential for COP1 function through deletion mutagenesis studies showing complete loss of activity in complementation assays when the RING is deleted[20][27]. Point mutations disrupting individual zinc-coordinating residues also abolish COP1 function, confirming that the intact RING fold is required rather than simply the presence of the N-terminal region[20].
The central coiled-coil domain (approximately amino acids 50-282) mediates protein-protein interactions essential for COP1 oligomerization and SPA protein binding[20][23]. Secondary structure prediction algorithms identify this region as having high probability of forming α-helical coiled-coil structures, and circular dichroism spectroscopy of purified COP1 fragments has confirmed high α-helical content in this region[20]. The coiled-coil domain is both necessary and sufficient for COP1-COP1 self-interaction, as demonstrated by yeast two-hybrid assays showing that isolated coiled-coil domains from two COP1 molecules interact strongly while deletion of the coiled-coil abolishes self-interaction[20][60]. The functional importance of the coiled-coil for COP1 activity has been established through complementation experiments showing that deletion of the coiled-coil region prevents rescue of cop1 mutants[20][27]. Interestingly, the coiled-coil domain together with the RING finger (amino acids 1-282, termed N282 or the N-terminal module) retains significant COP1 activity and can provide basal function allowing survival of cop1 mutants to reproductive maturity, though full activity requires the C-terminal domain as well[20][27][31][43].
The C-terminal WD40 repeat domain (approximately amino acids 283-675) forms a seven-bladed β-propeller structure that serves as the primary substrate recognition module[7][12][21]. High-resolution crystal structures of the COP1 WD40 domain from both Arabidopsis and human sources have revealed the three-dimensional architecture of this domain[7][12][21]. Each blade of the β-propeller consists of a four-stranded antiparallel β-sheet arranged in a radial fashion around a central axis, creating a toroid-shaped structure approximately 40 Å in diameter[7][12][21]. The substrate-binding site is located on the top face of the β-propeller, within a shallow groove formed at the interface between blades[7][21]. Structural studies of the WD40 domain in complex with substrate peptides containing the VP motif have revealed that substrates bind in an extended conformation, with the valine and proline residues of the VP motif making critical hydrophobic contacts with a conserved pocket formed by tryptophan and other aromatic residues from multiple WD40 repeats[7][21]. Adjacent to the VP-binding pocket, acidic residues in the substrate make electrostatic interactions with positively charged lysine and arginine residues on the COP1 surface, contributing to binding specificity and affinity[7][21].
A notable structural feature unique to COP1 among WD40 domain proteins is an inserted loop on the bottom face of the first blade, which may play regulatory roles in complex assembly or substrate presentation[7][21]. This insertion is conserved across plant and animal COP1 orthologs but is not present in structurally related WD40 proteins such as βTrCP or FBW7, suggesting COP1-specific functional requirements[7][21]. The structural basis for how this insertion affects COP1 function remains to be fully elucidated, though its conservation implies functional importance. Additionally, the structures revealed that the DWD box motif, previously proposed to mediate DDB1 binding, is buried within the WD40 fold and inaccessible, indicating that the mechanism of COP1-DDB1 interaction differs from the canonical DWD box-dependent recognition and requires reassessment[1].
Nuclear localization signals represent critical structural elements controlling COP1 subcellular distribution. The primary nuclear localization signal (NLS) is located within the central core region (approximately amino acids 200-250) and consists of a cluster of basic amino acids (lysine and arginine residues) that conform to a bipartite NLS consensus sequence recognized by importin proteins[11][20][27]. Site-directed mutagenesis converting lysine and arginine residues to a
Perplexity
(COP1-deep-research-perplexity-lite.md)
Perplexity
(COP1-deep-research-perplexity-lite.md)The COP1 gene in Arabidopsis thaliana (ARATH) encodes a central regulator of light signaling, acting as a key E3 ubiquitin ligase that controls photomorphogenesis and other developmental processes by targeting specific proteins for degradation[3][6][12].
Molecular Function
- E3 ubiquitin ligase activity: COP1 functions as an E3 ubiquitin-protein ligase, mediating the ubiquitination and subsequent proteasomal degradation of target proteins, most notably the transcription factor HY5, which is a positive regulator of photomorphogenesis[3][6][11].
- Protein-protein interactions: COP1 interacts with a variety of substrates and regulatory proteins, including SPA proteins (SUPPRESSOR OF PHYA-105), which modulate its activity, and other transcription factors such as ABI4 and ABI5 involved in abscisic acid (ABA) signaling[11][13].
Biological Processes
- Photomorphogenesis: COP1 is a negative regulator of photomorphogenesis in darkness, repressing light-induced development by targeting positive regulators for degradation. In light, COP1 activity is suppressed, allowing photomorphogenic development[3][12].
- UV-B response: COP1 acts as a positive regulator in UV-B signaling, coordinating HY5-dependent and independent pathways for UV-B tolerance[3].
- Hormone signaling: COP1 integrates light and ABA signaling, influencing seed germination, seedling establishment, stomatal closure, and abiotic stress responses by modulating the stability of ABA-related transcription factors and co-receptors[11].
- Root and shoot growth coordination: COP1 regulates the transcription of PIN1, affecting auxin transport and thus coordinating shoot-to-root growth responses to light[9].
Cellular Localization
- Nuclear localization: COP1 predominantly localizes to the nucleus in darkness, where it represses photomorphogenesis by targeting nuclear proteins for degradation[4][6].
- Cytoplasmic localization: In the presence of light, COP1 is exported to the cytoplasm, reducing its nuclear activity and allowing light-responsive gene expression[4][6].
- Localization signals: COP1 contains a bipartite nuclear localization signal (NLS) in its core domain and a cytoplasmic localization signal (CLS) overlapping the coiled-coil domain, which regulate its subcellular distribution[4].
Protein Domains
| Domain | Location (Amino Acids) | Function |
|---|---|---|
| RING-finger motif | N-terminal | Zinc-binding, essential for E3 ligase activity and protein interactions |
| Coiled-coil domain | Adjacent to RING | Mediates dimerization and protein-protein interactions |
| Core domain | Central | Contains NLS, essential for nuclear import |
| WD-40 repeats | C-terminal | Protein-protein interactions, substrate recognition |
- The modular structure allows COP1 to interact with multiple partners and substrates, conferring its pleiotropic regulatory roles[4][13].
Known Interactions
- HY5: Direct substrate, targeted for degradation to repress photomorphogenesis[3][6].
- SPA proteins (SPA1–SPA4): Form complexes with COP1, modulating its activity in light and dark conditions[3][11].
- ABI4 and ABI5: Transcription factors in ABA signaling, targeted by COP1 for degradation, integrating light and hormone signaling[11].
- PIN1: COP1 regulates its transcription, affecting auxin transport and root-shoot growth coordination[9].
- CIP1: COP1-interacting protein, promotes ABA signaling[11].
Disease Associations
- No direct human disease associations are reported for Arabidopsis COP1. However, the mammalian homolog (COP1/ RFWD2) is implicated in cancer and other diseases due to its role in protein degradation and gene regulation[6].
- In Arabidopsis, mutations in COP1 result in constitutive photomorphogenic phenotypes (de-etiolated seedlings in darkness), seedling lethality in severe alleles, and altered responses to environmental stresses[4][12].
Recent Developments (2023–2024)
- Chromatin remodeling: Recent studies show COP1 controls light-dependent chromatin remodeling, further linking it to epigenetic regulation of gene expression[5].
- Expanded regulatory network: New research highlights COP1’s integration of light, hormone, and stress signaling, emphasizing its central role in plant adaptability[11].
Key References
- UniProt entry for COP1 (P43254): Detailed domain structure, function, and interactions[6].
- Recent reviews and primary research articles (2021–2024) on COP1’s roles in light, hormone, and stress signaling[5][11][12].
Summary Table: COP1 in Arabidopsis thaliana
| Aspect | Details |
|---|---|
| Molecular Function | E3 ubiquitin ligase, protein-protein interactions |
| Biological Processes | Photomorphogenesis, UV-B response, ABA signaling, root-shoot growth coordination |
| Cellular Localization | Nucleus (dark), cytoplasm (light); regulated by NLS and CLS |
| Protein Domains | RING-finger, coiled-coil, core (NLS), WD-40 repeats |
| Known Interactions | HY5, SPA1–SPA4, ABI4, ABI5, PIN1, CIP1 |
| Disease Associations | No direct plant disease; mammalian homolog linked to cancer |
Key URLs and Dates:
- UniProt: https://www.uniprot.org/uniprotkb/P43254/entry (accessed 2024)
- Frontiers in Plant Science review: https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2021.662793/full (2021)
- PNAS chromatin remodeling study: https://www.pnas.org/doi/10.1073/pnas.2312853121 (2024)
Citations
- https://www.uniprot.org/uniprotkb/P43254/entry
- https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2021.662793/full
- https://www.pnas.org/doi/10.1073/pnas.2312853121