Deep Research
Bioreason Pro
(NEK3-deep-research-bioreason-sft.md)
Bioreason Pro
(NEK3-deep-research-bioreason-sft.md)Thinking Trace
I begin with the InterPro architecture. The polypeptide is assigned to IPR050660 (NEK Serine/Threonine Protein Kinase family, residues 1–518), which frames the entire sequence as a NIMA-related kinase with a conserved catalytic core and extended regulatory tail. Within this family envelope, the N-terminal half contains IPR011009 (Protein kinase-like domain superfamily, residues 1–301), and, more specifically, a canonical bilobal catalytic module defined by IPR000719 (Protein kinase domain, residues 4–257) and IPR000719 (Protein kinase domain, residues 4–258). The ATP-binding P-loop and adjacent glycine-rich segment are captured by IPR017441 (Protein kinase, ATP binding site, residues 10–33), while the catalytic HRD/DFG-centered machinery is pinpointed by IPR008271 (Serine/threonine-protein kinase, active site, residues 125–137). The ordered layout—ATP-binding segment leading into the catalytic loop within a NEK-family scaffold—causes ATP-dependent transfer of phosphate to serine/threonine residues on protein substrates. The NEK family context further implies regulation by activation-loop phosphorylation and docking interactions mediated by the C-terminal regulatory region (beyond residue ~257 up to 518), which typically confers subcellular targeting and substrate selection.
From this domain logic, the molecular function resolves to protein serine/threonine kinase activity (GO:0004674) driven by ATP binding (GO:0005524). The presence of broad interaction surfaces in NEK regulatory tails and the need to dock substrates and adaptors supports protein binding (GO:0005515). Together, these features define an enzyme that phosphorylates protein targets in a context-dependent manner.
NEK kinases are well known to control microtubule behavior. In plants, the cortical microtubule array dictates cell wall deposition patterns and anisotropic growth. A NEK with a long regulatory tail is well-suited to couple kinase activity to microtubule-associated proteins, thereby promoting microtubule bundle formation (GO:0001578). Phosphorylation of bundling factors, severing enzymes, or crosslinkers would stabilize parallel arrays and increase bundle longevity, which in turn drives directional expansion of specialized cells. In the male gametophyte, this control over cortical array stability and orientation provides the mechanical basis for polarized elongation, aligning with pollen development (GO:0009555). Thus, the catalytic architecture explains how the protein enforces microtubule bundle integrity and supports tip growth in pollen.
The cellular component follows from the same chain. A soluble kinase lacking transmembrane segments but equipped for cytoskeletal targeting will partition to the cytoplasm and associate with the microtubule cytoskeleton. The explicit functional tie to cortical microtubules places the protein at the cortical microtubule track (GO:0055028), embedded within the broader cytoskeleton (GO:0005856) and cytoplasm (GO:0005737). This localization allows direct access to substrates that shape the cortical array and to trafficking machinery that delivers wall materials.
A mechanistic hypothesis emerges: the kinase docks onto cortical microtubules via its NEK-family regulatory region, where it phosphorylates microtubule-associated proteins to promote bundle formation and persistence. SNAP29, a membrane trafficking factor at the plasma membrane–endomembrane interface, could act as a scaffold that positions the kinase near exocytic hotspots where cortical microtubules guide vesicle delivery. The T-complex protein 1 subunit epsilon (a CCT chaperonin component) likely assists in folding and assembly of the kinase and/or its microtubule-binding partners, ensuring a competent cortical complex. Beta-glucosidase 20, involved in cell wall carbohydrate remodeling, may be indirectly regulated through phosphorylation of trafficking or exocytosis components that control its delivery to growth sites. In aggregate, the kinase forms a cortical module that stabilizes microtubule bundles to steer vesicle traffic and cell wall deposition during pollen tube elongation.
## Functional Summary
A cytoplasmic, cortical-cytoskeleton–associated serine/threonine kinase that promotes microtubule bundle formation to support polarized growth during pollen development. Its N-terminal catalytic core binds ATP and phosphorylates protein substrates, while a NEK-family regulatory region targets the enzyme to cortical microtubules and provides docking interfaces for partners. By stabilizing microtubule bundles, it organizes vesicle delivery and cell wall deposition patterns required for elongating pollen tubes, likely operating in concert with membrane trafficking and protein-folding factors at the cell cortex.
## UniProt Summary
Involved in the regulation of cortical microtubule bundle integrity. Required for pollen tube elongation.
## InterPro Domains
- IPR011009: Protein kinase-like domain superfamily (homologous_superfamily) [1-301]
- IPR050660: NEK Serine/Threonine Protein Kinase (family) [1-518]
- IPR000719: Protein kinase domain (domain) [4-257]
- IPR000719: Protein kinase domain (domain) [4-258]
- IPR017441: Protein kinase, ATP binding site (binding_site) [10-33]
- IPR008271: Serine/threonine-protein kinase, active site (active_site) [125-137]
## GO Term Predictions
### Molecular Function
### Biological Process
### Cellular Component
Falcon
(NEK3-deep-research-falcon.md)
Falcon
(NEK3-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: Arabidopsis thaliana NEK3 (AtNek3; At5g28290; UniProt Q8RX66)
0) Executive summary (scope and evidence limits)
Arabidopsis NEK3 (AtNek3; At5g28290; UniProt Q8RX66) is a plant NIMA-related serine/threonine protein kinase (EC 2.7.11.1) in the NEK family, defined by a conserved N-terminal protein-kinase catalytic domain and a long basic C-terminal extension typical of NIMA-related kinases. In the accessible literature retrieved here, the strongest AtNek3-specific evidence concerns gene identity, domain architecture, phylogenetic placement, and organ/tissue expression patterns; direct experimental evidence for AtNek3 subcellular localization, biochemical substrates, or mutant/overexpression phenotypes was not found. Therefore, mechanistic insights are largely inferred from the broader plant NEK family and are explicitly labeled as such. (vigneault2007membersofthe pages 1-2, vigneault2007membersofthe pages 2-4, vigneault2007membersofthe pages 4-6, vigneault2007membersofthe pages 8-10, vigneault2009caractérisationdela pages 130-134, vigneault2009caractérisationdela pages 70-74)
1) Key concepts and definitions (current understanding)
1.1 NIMA-related kinases (NEKs)
NIMA-related kinases (Neks/NEKs) are a conserved family of serine/threonine kinases originally linked to mitotic control in fungi and animals, but in plants they have been strongly connected to microtubule-associated regulation of cell expansion and organ morphogenesis (family-level concept). (vigneault2007membersofthe pages 1-2, motose2012nimarelatedkinasesregulate pages 1-3, motose2012nimarelatedkinasesregulate pages 3-4)
1.2 What “functional annotation” means here
For NEK3, functional annotation ideally includes: (i) catalytic activity and substrate specificity; (ii) cellular compartment(s) of action; (iii) pathway context (developmental/stress signaling modules); and (iv) phenotype under perturbation. In the retrieved evidence, AtNek3 is best annotated at (a) molecular class (Ser/Thr kinase) and (b) likely biological-process context (developmental/vascular and meristem-associated expression), while (i), (ii), and (iv) remain gaps for AtNek3 specifically. (vigneault2007membersofthe pages 4-6, vigneault2007membersofthe pages 8-10, vigneault2009caractérisationdela pages 130-134)
2) Target verification (critical gene/protein identification)
The gene symbol NEK3 is ambiguous across organisms. The sources retrieved explicitly map AtNek3 to Arabidopsis thaliana locus At5g28290, consistent with the UniProt target Q8RX66 and the NEK/NIMA-related Ser/Thr kinase family. (vigneault2007membersofthe pages 1-2, vigneault2007membersofthe pages 2-4, vigneault2009caractérisationdela pages 130-134)
3) Molecular features of AtNek3
3.1 Enzymatic class and domain architecture
Plant NEKs (including AtNek3) are described as serine/threonine protein kinases with a conserved N-terminal catalytic kinase domain and a longer C-terminal non-catalytic extension, consistent with the UniProt domain calls (protein kinase domain; ATP-binding and activation-segment motifs). Sequence excerpts in the plant-NEK thesis include conserved Ser/Thr kinase motifs for AtNek3 (At5g28290), supporting the enzyme-class assignment. (vigneault2007membersofthe pages 1-2, vigneault2009caractérisationdela pages 70-74, vigneault2009caractérisationdela pages 130-134)
3.2 Phylogenetic context
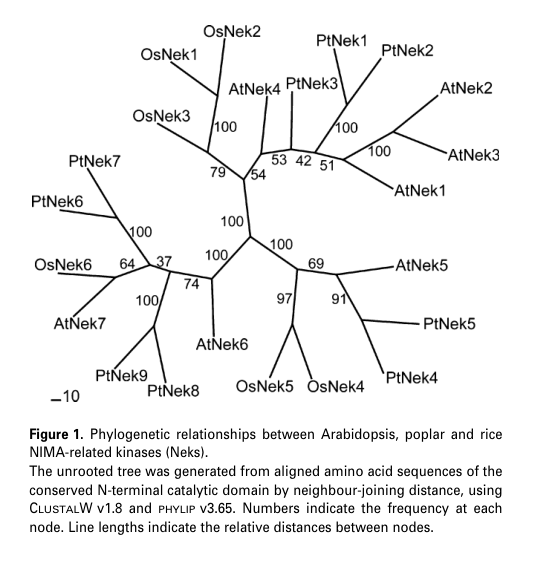
AtNek3 clusters with AtNek1–AtNek3 in a plant-specific clade distinct from fungal and mammalian NEKs, supporting the idea that plant NEKs have evolved specialized roles. (vigneault2007membersofthe pages 8-10, vigneault2007membersofthe media 26e7bf46, vigneault2009caractérisationdela pages 70-74)
4) Biological processes and pathway context supported for AtNek3
4.1 Expression-supported roles in development and vascularization
A key AtNek3-specific result is its organ/tissue expression profile. In Arabidopsis, AtNek3 shows strong expression in root tips and relatively high expression in the shoot apex; expression is preferentially associated with young leaves and vascular elements, and it declines in senescent leaves. These patterns were interpreted by the authors as consistent with roles in organ development/tissue differentiation and vascularization (inference from expression and family context). (vigneault2007membersofthe pages 4-6, vigneault2007membersofthe pages 8-10, vigneault2007membersofthe media 26e7bf46, vigneault2007membersofthe media 77c1eb59)
4.2 Cell-cycle association (transcriptional)
A dissertation on Arabidopsis NIMA-like kinases reports that AtNIMA3 has an organ-specific transcription profile not associated with the mitotic cell cycle, suggesting AtNek3 transcription is not tightly coupled to mitosis (unlike many fungal/animal NEKs). (agueci2010characterizationofnimalike pages 5-9)
5) Subcellular localization and direct mechanism (evidence gaps for AtNek3)
No AtNek3-specific experimental evidence of subcellular localization (e.g., GFP fusions) or direct microtubule association was found in the retrieved evidence set. Consequently, AtNek3 localization cannot be asserted beyond domain-based speculation. (vigneault2007membersofthe pages 4-6, vigneault2009caractérisationdela pages 130-134)
6) Recent developments (prioritizing 2023–2024)
6.1 2023: Plant NEK functional insights from Marchantia (family-level inference)
A 2023 preprint in the liverwort Marchantia polymorpha investigated MpNEK1 using estradiol-inducible overexpression. Overexpression caused severe growth suppression (rhizoids and thalli), with dose sensitivity detectable even at 10–100 nM estradiol and MpNEK1 transcript induction of approximately 4–15× in responsive lines. Cell proliferation readouts (EdU labeling and reduced mitotic counts) supported suppressed proliferation, while microtubule arrays were not grossly disrupted. Importantly, kinase-deficient MpNEK1 variants still suppressed growth (milder), suggesting a potentially phosphorylation-independent component to NEK function in plants. These findings update the field’s thinking about how plant NEKs may operate, but they do not directly establish AtNek3 function in Arabidopsis. (mase2023overexpressionofnimarelated pages 5-8, mase2023overexpressionofnimarelated pages 1-5, mase2023overexpressionofnimarelated pages 8-11, mase2023overexpressionofnimarelated pages 11-13)
6.2 2023: Microtubule regulation and stress adaptation (context)
A 2023 review summarizes how microtubules participate in abiotic stress responses (heat, salinity, drought) through rapid reorganization and regulation by diverse microtubule-associated proteins. This contextualizes why plant NEKs—given their family-level links to microtubule function—remain relevant candidates for stress-adaptation biology, although the retrieved pages did not specifically mention AtNek3. (hsiao2023microtubuleregulationin pages 6-7, hsiao2023microtubuleregulationin pages 7-9, hsiao2023microtubuleregulationin pages 9-10)
7) Applications and real-world implementations (NEK family; not AtNek3-specific)
7.1 Stress tolerance and growth engineering in crops and model plants
Although AtNek3 itself lacks application-grade evidence in the retrieved set, the plant NEK family has been implemented in transgenic contexts:
- Soybean GmNEK1: Overexpression in Arabidopsis increased leaf growth with statistical reporting (n = 24, P < 0.01) and GmNEK1 co-localized with a tubulin marker (GFP–TUB6), supporting a microtubule-linked mechanism; the study also reports stress-responsive induction patterns (e.g., salt/cold) and improved tolerance phenotypes under stress assays. (pan2017soybeannimarelatedkinase1 pages 2-4, pan2017soybeannimarelatedkinase1 pages 8-9, pan2017soybeannimarelatedkinase1 pages 6-8)
- Arabidopsis NEK6: Overexpression and mutant analysis show altered growth and improved tolerance to osmotic/salt stress in controlled assays, indicating potential leverage points for engineering growth robustness. (zhang2011nimarelatedkinasenek6 pages 7-8, zhang2011nimarelatedkinasenek6 pages 1-2)
These examples support “real-world implementation” in the sense of genetic engineering strategies to modify growth and stress tolerance, but they should not be conflated with AtNek3-specific applications. (pan2017soybeannimarelatedkinase1 pages 2-4, zhang2011nimarelatedkinasenek6 pages 7-8)
8) Expert opinions and authoritative analysis (what the field argues)
A peer-reviewed short review on Arabidopsis NEKs argues that plant NEKs regulate directional cell growth and organ development through microtubule function, with emphasis on NEK4/NEK5/NEK6 interactions and a model in which NEK6 destabilizes microtubules (potentially via β-tubulin phosphorylation). This is an authoritative synthesis for the NEK family, but it provides no direct AtNek3 mechanistic claims. (motose2012nimarelatedkinasesregulate pages 1-3, motose2012nimarelatedkinasesregulate pages 3-4, motose2012nimarelatedkinasesregulate pages 4-4)
9) Data and statistics (recent studies; with appropriate scope)
9.1 Quantitative data directly in retrieved evidence
- MpNEK1 (Marchantia) inducible overexpression: 4–15× induction of transcript upon estradiol treatment; phenotypes observed even at 10–100 nM estradiol; growth suppression reversible ≤3 days induction but irreversible after ≥7 days. (mase2023overexpressionofnimarelated pages 5-8, mase2023overexpressionofnimarelated pages 1-5)
- GmNEK1 (soybean) overexpression in Arabidopsis: increased leaf growth with n = 24, P < 0.01 (as reported in excerpt), supporting that NEK-family manipulation can yield measurable growth differences. (pan2017soybeannimarelatedkinase1 pages 2-4)
9.2 AtNek3-specific quantitative data
AtNek3-specific quantitative phenotype or biochemical data were not present in the retrieved evidence. The key quantitative-like outputs for AtNek3 are expression patterns across organs/tissues (qualitatively described and visually presented). (vigneault2007membersofthe pages 4-6, vigneault2007membersofthe media 26e7bf46, vigneault2007membersofthe media 77c1eb59)
10) Practical functional-annotation conclusions for AtNek3 (what can be stated with evidence)
- Molecular function (supported): AtNek3 is a NIMA-related serine/threonine protein kinase by sequence/domain architecture and family definition; reaction class is ATP-dependent phosphorylation of protein substrates (EC 2.7.11.1), but substrate identity and specificity remain unknown for AtNek3 based on retrieved evidence. (vigneault2007membersofthe pages 1-2, vigneault2009caractérisationdela pages 130-134, vigneault2009caractérisationdela pages 70-74)
- Biological process (supported/inferred): Expression patterns suggest roles in developmental programs associated with root tips, shoot apex, young leaves, and vascular tissues, consistent with proposed roles in organ development and vascularization (inference from expression). (vigneault2007membersofthe pages 4-6, vigneault2007membersofthe pages 8-10, vigneault2007membersofthe media 26e7bf46)
- Cellular location (unsupported for AtNek3): No AtNek3-specific localization evidence retrieved; cannot assert cytosolic/nuclear/microtubule localization for AtNek3 from this corpus. (vigneault2009caractérisationdela pages 130-134)
- Pathway context (inferred): While other plant NEKs (e.g., NEK4/5/6) participate in microtubule-based growth-direction control, AtNek3’s pathway membership is not established here; the most defensible hypothesis is that AtNek3 participates in plant-specific NEK signaling linked to developmental and/or vascular processes, pending direct experiments. (vigneault2007membersofthe pages 8-10, motose2012nimarelatedkinasesregulate pages 1-3, motose2012nimarelatedkinasesregulate pages 3-4)
Evidence summary table
| Claim/Aspect | Evidence summary | Source (author/year, journal) | URL/DOI | Notes/limitations |
|---|---|---|---|---|
| Identity | AtNek3 is explicitly identified as the Arabidopsis thaliana NIMA-related kinase family member encoded by locus At5g28290; the family description matches the UniProt target NEK3/Q8RX66. (vigneault2007membersofthe pages 1-2, vigneault2007membersofthe pages 2-4) | Vigneault et al., 2007, The Plant Journal | https://doi.org/10.1111/j.1365-313x.2007.03161.x | Confirms the correct Arabidopsis gene and distinguishes it from non-plant NEK3 proteins. |
| Domain architecture | Plant Neks, including AtNek3, are described as serine/threonine kinases with a conserved N-terminal catalytic kinase domain and a longer C-terminal non-catalytic extension; sequence-level evidence in the thesis shows conserved Ser/Thr kinase motifs in AtNek3. (vigneault2007membersofthe pages 1-2, vigneault2009caractérisationdela pages 130-134, vigneault2009caractérisationdela pages 70-74) | Vigneault et al., 2007, The Plant Journal; Vigneault, 2009, thesis | https://doi.org/10.1111/j.1365-313x.2007.03161.x; thesis DOI: 10.25673/202 | Strong family/domain inference, but no AtNek3-specific biochemical assay was reported in the gathered evidence. |
| Phylogenetic placement | AtNek3 clusters with the AtNek1–AtNek3 subgroup within a plant-specific Nek clade, distinct from fungal and mammalian Neks; plant Neks were inferred to descend from an ancestral plant Nek. (vigneault2007membersofthe pages 8-10, vigneault2009caractérisationdela pages 70-74, vigneault2007membersofthe media 26e7bf46) | Vigneault et al., 2007, The Plant Journal; Vigneault, 2009, thesis | https://doi.org/10.1111/j.1365-313x.2007.03161.x; thesis DOI: 10.25673/202 | Evolutionary evidence supports likely conserved plant-specific function, but does not by itself establish mechanism. |
| Expression pattern in Arabidopsis organs/tissues | AtNek3 transcripts are reported as strong in root tips and relatively high in the shoot apex, with expression preferentially associated with young leaves and vascular elements; AtNek3 expression is very low in hypocotyl, xylem, and cork extracts and decreases strongly in senescent leaves. (vigneault2007membersofthe pages 4-6, vigneault2007membersofthe pages 8-10, vigneault2007membersofthe media 26e7bf46) | Vigneault et al., 2007, The Plant Journal | https://doi.org/10.1111/j.1365-313x.2007.03161.x | Evidence is expression-based and tissue-level; it does not prove subcellular localization or direct function. |
| Cell-cycle association | A dissertation summary states that AtNIMA3/AtNek3 shows an organ-specific transcription profile not associated with the mitotic cell cycle, contrasting with some NEKs in fungi/animals. (agueci2010characterizationofnimalike pages 5-9) | Agueci, 2010, dissertation | https://doi.org/10.25673/202 | Mapping to At5g28290/Q8RX66 is inferential within the gathered evidence; no direct AtNek3 cell-cycle assay was cited here. |
| Inferred biological role | Based on expression in root tips, shoot apex, young leaves, and vascular tissues, the authors infer that AtNek3 is involved more in organ development, tissue differentiation, and vascularization than in core mitotic control. (vigneault2007membersofthe pages 4-6, vigneault2007membersofthe pages 8-10) | Vigneault et al., 2007, The Plant Journal | https://doi.org/10.1111/j.1365-313x.2007.03161.x | This is an inference from comparative expression and phylogeny; no direct AtNek3 mutant or overexpression phenotype was provided. |
| Subcellular localization | No AtNek3-specific experimental subcellular localization was identified in the gathered evidence. Localization data in the available literature concern other plant NEKs such as PNek1, not AtNek3. (vigneault2009caractérisationdela pages 74-79) | Vigneault, 2009, thesis | thesis DOI: 10.25673/202 | Important evidence gap: localization for AtNek3 remains unresolved in the retrieved sources. |
| Enzymatic activity / substrates | Although AtNek3 is classified as a serine/threonine protein kinase by family/domain features, no AtNek3-specific kinase assay, substrate, or substrate specificity was found in the gathered evidence. (vigneault2009caractérisationdela pages 130-134, vigneault2009caractérisationdela pages 70-74) | Vigneault, 2009, thesis | thesis DOI: 10.25673/202 | Functional annotation at the reaction/substrate level is currently unsupported by direct evidence in the retrieved literature. |
| Mutant or overexpression phenotype | The gathered evidence did not provide a direct AtNek3 loss-of-function or overexpression phenotype in Arabidopsis. Family-level papers discuss developmental functions for plant NEKs, but AtNek3-specific phenotype data were not reported in the retrieved excerpts. (vigneault2007membersofthe pages 4-6, vigneault2007membersofthe pages 1-2) | Vigneault et al., 2007, The Plant Journal | https://doi.org/10.1111/j.1365-313x.2007.03161.x | Another major evidence gap; conclusions for AtNek3 remain largely inferential. |
Table: This table summarizes what the gathered evidence supports for Arabidopsis thaliana NEK3/AtNek3 (At5g28290; UniProt Q8RX66). It highlights that identity, kinase-family assignment, and expression-based developmental inferences are supported, while direct localization, substrates, and phenotype evidence remain limited.
Key cited sources (with publication dates and URLs)
- Vigneault F. et al. 2007-07. The Plant Journal. “Members of the plant NIMA-related kinases are involved in organ development and vascularization in poplar, Arabidopsis and rice.” https://doi.org/10.1111/j.1365-313x.2007.03161.x (vigneault2007membersofthe pages 1-2, vigneault2007membersofthe pages 4-6, vigneault2007membersofthe pages 8-10, vigneault2007membersofthe media 26e7bf46, vigneault2007membersofthe media 77c1eb59)
- Agueci F. 2010-01. Dissertation. “Characterization of NIMA-like kinases in Arabidopsis thaliana.” https://doi.org/10.25673/202 (agueci2010characterizationofnimalike pages 5-9)
- Motose H. et al. 2012-12. Plant Signaling & Behavior. “NIMA-related kinases regulate directional cell growth and organ development through microtubule function in Arabidopsis thaliana.” https://doi.org/10.4161/psb.22412 (motose2012nimarelatedkinasesregulate pages 1-3, motose2012nimarelatedkinasesregulate pages 3-4)
- Mase H. et al. 2023-07 (preprint). bioRxiv. “Overexpression of NIMA-related kinase suppresses cell proliferation and tip growth in a liverwort Marchantia polymorpha.” https://doi.org/10.1101/2023.01.25.525476 (mase2023overexpressionofnimarelated pages 5-8, mase2023overexpressionofnimarelated pages 1-5, mase2023overexpressionofnimarelated pages 8-11)
- Hsiao A.-S., Huang J.-Y. 2023-03. Biomolecules. “Microtubule Regulation in Plants: From Morphological Development to Stress Adaptation.” https://doi.org/10.3390/biom13040627 (hsiao2023microtubuleregulationin pages 6-7, hsiao2023microtubuleregulationin pages 7-9, hsiao2023microtubuleregulationin pages 9-10)
- Pan W.-J. et al. 2017-07. Plant and Cell Physiology. “Soybean NIMA-Related Kinase1 Promotes Plant Growth and Improves Salt and Cold Tolerance.” https://doi.org/10.1093/pcp/pcx060 (pan2017soybeannimarelatedkinase1 pages 2-4)
- Zhang B. et al. 2011-12. The Plant Journal. “NIMA-related kinase NEK6 affects plant growth and stress response in Arabidopsis.” https://doi.org/10.1111/j.1365-313x.2011.04733.x (zhang2011nimarelatedkinasenek6 pages 7-8, zhang2011nimarelatedkinasenek6 pages 1-2)
References
-
(vigneault2007membersofthe pages 1-2): Frédéric Vigneault, Denis Lachance, Monikca Cloutier, Gervais Pelletier, Caroline Levasseur, and Armand Séguin. Members of the plant nima‐related kinases are involved in organ development and vascularization in poplar, arabidopsis and rice. The Plant Journal, 51:575-588, Jul 2007. URL: https://doi.org/10.1111/j.1365-313x.2007.03161.x, doi:10.1111/j.1365-313x.2007.03161.x. This article has 56 citations.
-
(vigneault2007membersofthe pages 2-4): Frédéric Vigneault, Denis Lachance, Monikca Cloutier, Gervais Pelletier, Caroline Levasseur, and Armand Séguin. Members of the plant nima‐related kinases are involved in organ development and vascularization in poplar, arabidopsis and rice. The Plant Journal, 51:575-588, Jul 2007. URL: https://doi.org/10.1111/j.1365-313x.2007.03161.x, doi:10.1111/j.1365-313x.2007.03161.x. This article has 56 citations.
-
(vigneault2007membersofthe pages 4-6): Frédéric Vigneault, Denis Lachance, Monikca Cloutier, Gervais Pelletier, Caroline Levasseur, and Armand Séguin. Members of the plant nima‐related kinases are involved in organ development and vascularization in poplar, arabidopsis and rice. The Plant Journal, 51:575-588, Jul 2007. URL: https://doi.org/10.1111/j.1365-313x.2007.03161.x, doi:10.1111/j.1365-313x.2007.03161.x. This article has 56 citations.
-
(vigneault2007membersofthe pages 8-10): Frédéric Vigneault, Denis Lachance, Monikca Cloutier, Gervais Pelletier, Caroline Levasseur, and Armand Séguin. Members of the plant nima‐related kinases are involved in organ development and vascularization in poplar, arabidopsis and rice. The Plant Journal, 51:575-588, Jul 2007. URL: https://doi.org/10.1111/j.1365-313x.2007.03161.x, doi:10.1111/j.1365-313x.2007.03161.x. This article has 56 citations.
-
(vigneault2009caractérisationdela pages 130-134): F Vigneault. Caractérisation de la famille des protéines kinases de type nima chez les plantes et analyse fonctionnelle de pnek1, une nek du peuplier (populus tremula x p …. Unknown journal, 2009.
-
(vigneault2009caractérisationdela pages 70-74): F Vigneault. Caractérisation de la famille des protéines kinases de type nima chez les plantes et analyse fonctionnelle de pnek1, une nek du peuplier (populus tremula x p …. Unknown journal, 2009.
-
(motose2012nimarelatedkinasesregulate pages 1-3): Hiroyasu Motose, Shogo Takatani, Tatsuya Ikeda, and Taku Takahashi. Nima-related kinases regulate directional cell growth and organ development through microtubule function inarabidopsis thaliana. Plant Signaling & Behavior, 7:1552-1555, Dec 2012. URL: https://doi.org/10.4161/psb.22412, doi:10.4161/psb.22412. This article has 30 citations and is from a peer-reviewed journal.
-
(motose2012nimarelatedkinasesregulate pages 3-4): Hiroyasu Motose, Shogo Takatani, Tatsuya Ikeda, and Taku Takahashi. Nima-related kinases regulate directional cell growth and organ development through microtubule function inarabidopsis thaliana. Plant Signaling & Behavior, 7:1552-1555, Dec 2012. URL: https://doi.org/10.4161/psb.22412, doi:10.4161/psb.22412. This article has 30 citations and is from a peer-reviewed journal.
-
(vigneault2007membersofthe media 26e7bf46): Frédéric Vigneault, Denis Lachance, Monikca Cloutier, Gervais Pelletier, Caroline Levasseur, and Armand Séguin. Members of the plant nima‐related kinases are involved in organ development and vascularization in poplar, arabidopsis and rice. The Plant Journal, 51:575-588, Jul 2007. URL: https://doi.org/10.1111/j.1365-313x.2007.03161.x, doi:10.1111/j.1365-313x.2007.03161.x. This article has 56 citations.
-
(vigneault2007membersofthe media 77c1eb59): Frédéric Vigneault, Denis Lachance, Monikca Cloutier, Gervais Pelletier, Caroline Levasseur, and Armand Séguin. Members of the plant nima‐related kinases are involved in organ development and vascularization in poplar, arabidopsis and rice. The Plant Journal, 51:575-588, Jul 2007. URL: https://doi.org/10.1111/j.1365-313x.2007.03161.x, doi:10.1111/j.1365-313x.2007.03161.x. This article has 56 citations.
-
(agueci2010characterizationofnimalike pages 5-9): Francesco Agueci. Characterization of nima-like kinases in arabidopsis thaliana. Dissertation, Jan 2010. URL: https://doi.org/10.25673/202, doi:10.25673/202. This article has 0 citations.
-
(mase2023overexpressionofnimarelated pages 5-8): Hikari Mase, Yoshihiro Yoshitake, Takayuki Kohchi, Taku Takahashi, and Hiroyasu Motose. Overexpression of nima-related kinase suppresses cell proliferation and tip growth in a liverwort marchantia polymorpha. bioRxiv, Jul 2023. URL: https://doi.org/10.1101/2023.01.25.525476, doi:10.1101/2023.01.25.525476. This article has 0 citations.
-
(mase2023overexpressionofnimarelated pages 1-5): Hikari Mase, Yoshihiro Yoshitake, Takayuki Kohchi, Taku Takahashi, and Hiroyasu Motose. Overexpression of nima-related kinase suppresses cell proliferation and tip growth in a liverwort marchantia polymorpha. bioRxiv, Jul 2023. URL: https://doi.org/10.1101/2023.01.25.525476, doi:10.1101/2023.01.25.525476. This article has 0 citations.
-
(mase2023overexpressionofnimarelated pages 8-11): Hikari Mase, Yoshihiro Yoshitake, Takayuki Kohchi, Taku Takahashi, and Hiroyasu Motose. Overexpression of nima-related kinase suppresses cell proliferation and tip growth in a liverwort marchantia polymorpha. bioRxiv, Jul 2023. URL: https://doi.org/10.1101/2023.01.25.525476, doi:10.1101/2023.01.25.525476. This article has 0 citations.
-
(mase2023overexpressionofnimarelated pages 11-13): Hikari Mase, Yoshihiro Yoshitake, Takayuki Kohchi, Taku Takahashi, and Hiroyasu Motose. Overexpression of nima-related kinase suppresses cell proliferation and tip growth in a liverwort marchantia polymorpha. bioRxiv, Jul 2023. URL: https://doi.org/10.1101/2023.01.25.525476, doi:10.1101/2023.01.25.525476. This article has 0 citations.
-
(hsiao2023microtubuleregulationin pages 6-7): An-Shan Hsiao and Ji-Ying Huang. Microtubule regulation in plants: from morphological development to stress adaptation. Biomolecules, 13:627, Mar 2023. URL: https://doi.org/10.3390/biom13040627, doi:10.3390/biom13040627. This article has 28 citations.
-
(hsiao2023microtubuleregulationin pages 7-9): An-Shan Hsiao and Ji-Ying Huang. Microtubule regulation in plants: from morphological development to stress adaptation. Biomolecules, 13:627, Mar 2023. URL: https://doi.org/10.3390/biom13040627, doi:10.3390/biom13040627. This article has 28 citations.
-
(hsiao2023microtubuleregulationin pages 9-10): An-Shan Hsiao and Ji-Ying Huang. Microtubule regulation in plants: from morphological development to stress adaptation. Biomolecules, 13:627, Mar 2023. URL: https://doi.org/10.3390/biom13040627, doi:10.3390/biom13040627. This article has 28 citations.
-
(pan2017soybeannimarelatedkinase1 pages 2-4): Wen-Jia Pan, Jian-Jun Tao, Tong Cheng, Ming Shen, Jin-Biao Ma, Wan-Ke Zhang, Qin Lin, Biao Ma, Shou-Yi Chen, and Jin-Song Zhang. Soybean nima-related kinase1 promotes plant growth and improves salt and cold tolerance. Plant and Cell Physiology, 58:1268–1278, Jul 2017. URL: https://doi.org/10.1093/pcp/pcx060, doi:10.1093/pcp/pcx060. This article has 30 citations and is from a domain leading peer-reviewed journal.
-
(pan2017soybeannimarelatedkinase1 pages 8-9): Wen-Jia Pan, Jian-Jun Tao, Tong Cheng, Ming Shen, Jin-Biao Ma, Wan-Ke Zhang, Qin Lin, Biao Ma, Shou-Yi Chen, and Jin-Song Zhang. Soybean nima-related kinase1 promotes plant growth and improves salt and cold tolerance. Plant and Cell Physiology, 58:1268–1278, Jul 2017. URL: https://doi.org/10.1093/pcp/pcx060, doi:10.1093/pcp/pcx060. This article has 30 citations and is from a domain leading peer-reviewed journal.
-
(pan2017soybeannimarelatedkinase1 pages 6-8): Wen-Jia Pan, Jian-Jun Tao, Tong Cheng, Ming Shen, Jin-Biao Ma, Wan-Ke Zhang, Qin Lin, Biao Ma, Shou-Yi Chen, and Jin-Song Zhang. Soybean nima-related kinase1 promotes plant growth and improves salt and cold tolerance. Plant and Cell Physiology, 58:1268–1278, Jul 2017. URL: https://doi.org/10.1093/pcp/pcx060, doi:10.1093/pcp/pcx060. This article has 30 citations and is from a domain leading peer-reviewed journal.
-
(zhang2011nimarelatedkinasenek6 pages 7-8): Bo Zhang, Hao‐Wei Chen, Rui‐Ling Mu, Wang‐Ke Zhang, Ming‐Yu Zhao, Wei Wei, Fang Wang, Hui Yu, Gang Lei, Hong‐Feng Zou, Biao Ma, Shou‐Yi Chen, and Jin‐Song Zhang. Nima-related kinase nek6 affects plant growth and stress response in arabidopsis. The Plant journal : for cell and molecular biology, 68 5:830-43, Dec 2011. URL: https://doi.org/10.1111/j.1365-313x.2011.04733.x, doi:10.1111/j.1365-313x.2011.04733.x. This article has 55 citations.
-
(zhang2011nimarelatedkinasenek6 pages 1-2): Bo Zhang, Hao‐Wei Chen, Rui‐Ling Mu, Wang‐Ke Zhang, Ming‐Yu Zhao, Wei Wei, Fang Wang, Hui Yu, Gang Lei, Hong‐Feng Zou, Biao Ma, Shou‐Yi Chen, and Jin‐Song Zhang. Nima-related kinase nek6 affects plant growth and stress response in arabidopsis. The Plant journal : for cell and molecular biology, 68 5:830-43, Dec 2011. URL: https://doi.org/10.1111/j.1365-313x.2011.04733.x, doi:10.1111/j.1365-313x.2011.04733.x. This article has 55 citations.
-
(motose2012nimarelatedkinasesregulate pages 4-4): Hiroyasu Motose, Shogo Takatani, Tatsuya Ikeda, and Taku Takahashi. Nima-related kinases regulate directional cell growth and organ development through microtubule function inarabidopsis thaliana. Plant Signaling & Behavior, 7:1552-1555, Dec 2012. URL: https://doi.org/10.4161/psb.22412, doi:10.4161/psb.22412. This article has 30 citations and is from a peer-reviewed journal.
-
(vigneault2009caractérisationdela pages 74-79): F Vigneault. Caractérisation de la famille des protéines kinases de type nima chez les plantes et analyse fonctionnelle de pnek1, une nek du peuplier (populus tremula x p …. Unknown journal, 2009.
Artifacts
Citations
- agueci2010characterizationofnimalike pages 5-9
- vigneault2007membersofthe pages 1-2
- vigneault2007membersofthe pages 2-4
- vigneault2007membersofthe pages 4-6
- vigneault2007membersofthe pages 8-10
- motose2012nimarelatedkinasesregulate pages 1-3
- motose2012nimarelatedkinasesregulate pages 3-4
- mase2023overexpressionofnimarelated pages 5-8
- mase2023overexpressionofnimarelated pages 1-5
- mase2023overexpressionofnimarelated pages 8-11
- mase2023overexpressionofnimarelated pages 11-13
- hsiao2023microtubuleregulationin pages 6-7
- hsiao2023microtubuleregulationin pages 7-9
- hsiao2023microtubuleregulationin pages 9-10
- motose2012nimarelatedkinasesregulate pages 4-4
- https://doi.org/10.1111/j.1365-313x.2007.03161.x
- https://doi.org/10.1111/j.1365-313x.2007.03161.x;
- https://doi.org/10.25673/202
- https://doi.org/10.4161/psb.22412
- https://doi.org/10.1101/2023.01.25.525476
- https://doi.org/10.3390/biom13040627
- https://doi.org/10.1093/pcp/pcx060
- https://doi.org/10.1111/j.1365-313x.2011.04733.x
- https://doi.org/10.1111/j.1365-313x.2007.03161.x,
- https://doi.org/10.4161/psb.22412,
- https://doi.org/10.25673/202,
- https://doi.org/10.1101/2023.01.25.525476,
- https://doi.org/10.3390/biom13040627,
- https://doi.org/10.1093/pcp/pcx060,
- https://doi.org/10.1111/j.1365-313x.2011.04733.x,