id: P02828
gene_symbol: Hsp83
product_type: PROTEIN
status: DRAFT
taxon:
id: NCBITaxon:7227
label: Drosophila melanogaster
description: Hsp83 is the sole Drosophila melanogaster member of the HSP90 family
of ATP-dependent molecular chaperones. It promotes the maturation, structural maintenance,
and proper regulation of specific client proteins involved in cell cycle control,
signal transduction, piRNA biogenesis, and centrosome function. Hsp83 undergoes
a functional ATPase cycle that induces conformational changes in client proteins,
thereby causing their activation. It interacts dynamically with various co-chaperones
(Hop, Shu, Dpit47, Morgana) that modulate its substrate recognition, ATPase cycle,
and chaperone function. In addition to its canonical chaperone role, Hsp83 has a
non-chaperone membrane-deforming function mediated by an amphipathic helix that
promotes exosome release. Hsp83 is constitutively expressed and further induced
by heat stress; it is also expressed during oogenesis and plays roles in developmental
processes including centrosome cycle, cell cycle exit, mRNA localization, and sleep
regulation.
existing_annotations:
- term:
id: GO:0006457
label: protein folding
evidence_type: IBA
original_reference_id: GO_REF:0000033
review:
summary: Hsp83/HSP90 is a well-established molecular chaperone whose primary biological
role is protein folding. The IBA annotation is supported by strong phylogenetic
evidence across multiple orthologs (PMID:10716925, UniProt function annotation).
UniProt states Hsp83 is a "Molecular chaperone that promotes the maturation,
structural maintenance and proper regulation of specific target proteins."
action: ACCEPT
reason: Protein folding is the core biological process of HSP90 family chaperones.
IBA is well-supported by orthologs and consistent with all functional literature
on Hsp83.
supported_by:
- reference_id: PMID:10716925
supporting_text: a member of the highly conserved Hsp90 family including chaperones
known to maintain the activity of many proteins
- reference_id: file:DROME/Hsp83/Hsp83-deep-research-falcon.md
supporting_text: |-
Hsp83 is best understood as the **cytosolic Hsp90 ATP-dependent chaperone** that promotes **late-stage folding/maturation, stabilization, and activation** of diverse client proteins, especially those central to signaling and development
- term:
id: GO:0016887
label: ATP hydrolysis activity
evidence_type: IBA
original_reference_id: GO_REF:0000033
review:
summary: Hsp83/HSP90 has intrinsic ATPase activity that is essential for its chaperone
cycle. The UniProt record notes it "Undergoes a functional cycle that is linked
to its ATPase activity." The IBA annotation is supported by phylogenetic inference
from multiple HSP90 orthologs with experimentally characterized ATPase activity.
action: ACCEPT
reason: ATP hydrolysis is a core enzymatic activity of the HSP90 family. The HATPase_c
domain is well characterized structurally and the ATPase cycle drives the chaperone
conformational changes. Consistent with IBA evidence across the HSP90 family.
supported_by:
- reference_id: PMID:29775584
supporting_text: the Hsp90 system (Hop, Hsp90, and p23) together render Ago2
into an open, active form
- reference_id: file:DROME/Hsp83/Hsp83-deep-research-falcon.md
supporting_text: |-
Mechanistically, Hsp90 proteins are **dimeric**, undergoing an **ATP-driven conformational cycle** involving **ATP binding/hydrolysis** and large structural rearrangements that enable remodeling/maturation of clients
- term:
id: GO:0032991
label: protein-containing complex
evidence_type: IBA
original_reference_id: GO_REF:0000033
review:
summary: HSP90 family members function as homodimers and form multi-protein complexes
with co-chaperones (Hop, p23, Cdc37, Aha1) and client proteins. UniProt states
Hsp83 is a "Homodimer" and describes multiple complexes including with Hop/piwi,
sicily/ND-42, Dpit47/Hsp70aa, and Nup358. The IBA annotation to the generic
protein-containing complex is correct but very general.
action: ACCEPT
reason: HSP90 is known to form multiple protein complexes. While the term is generic,
IBA annotations at this level are acceptable as they reflect the phylogenetic
inference. More specific complex annotations (GO:0101031, GO:0034663) are captured
elsewhere.
- term:
id: GO:0005886
label: plasma membrane
evidence_type: IBA
original_reference_id: GO_REF:0000033
review:
summary: IBA annotation to plasma membrane is supported by phylogenetic inference
from multiple HSP90 orthologs. Hsp83 was also detected at the plasma membrane
by HDA (PMID:19317464). Hsp90 has an amphipathic helix that allows membrane
interaction (PMID:30193096), which could explain plasma membrane localization.
action: ACCEPT
reason: Plasma membrane localization is consistent with both the IBA evidence
from orthologs and the independent HDA evidence from proteomics (PMID:19317464).
The amphipathic helix-mediated membrane interaction described in PMID:30193096
provides a mechanistic basis.
supported_by:
- reference_id: PMID:30193096
supporting_text: Hsp90 also has the ability to directly interact with and deform
membranes via an evolutionarily conserved amphipathic helix
- term:
id: GO:0005524
label: ATP binding
evidence_type: IBA
original_reference_id: GO_REF:0000033
review:
summary: ATP binding is a fundamental property of HSP90 family members, mediated
by the HATPase_c domain. UniProt lists multiple ATP binding residues (positions
39, 81, 100, 126, 385) and the keyword "ATP-binding." IBA annotation is well
supported.
action: ACCEPT
reason: ATP binding is intrinsic to the HSP90 ATPase domain. Structurally characterized
across the family.
- term:
id: GO:0005829
label: cytosol
evidence_type: IBA
original_reference_id: GO_REF:0000033
review:
summary: Cytosolic localization of HSP90 family members is well established. IBA
annotation from multiple orthologs. Consistent with UniProt subcellular location
annotation to "Cytoplasm" and multiple IDA/HDA annotations to cytoplasm in Drosophila.
action: ACCEPT
reason: Cytosol is the primary localization for cytoplasmic HSP90 family members.
IBA is well supported.
- term:
id: GO:0050821
label: protein stabilization
evidence_type: IBA
original_reference_id: GO_REF:0000033
review:
summary: HSP90 chaperones stabilize client proteins, preventing their degradation.
UniProt states Hsp83 "promotes the maturation, structural maintenance and proper
regulation of specific target proteins." IBA from multiple orthologs. Loss of
Hsp90 function leads to decreased levels of client proteins such as ND42 and
NDUFS3 (PMID:23509070), directly demonstrating protein stabilization.
action: ACCEPT
reason: Protein stabilization is a core function of HSP90 -- client proteins are
destabilized and degraded when HSP90 is inhibited. Well supported by phylogenetic
evidence and direct experimental data in Drosophila.
supported_by:
- reference_id: PMID:23509070
supporting_text: Both NDUFS3 and ND42 are severely down-regulated in sicily
mutants, whereas other mitochondrial proteins, including Porin (Graham et
al., 2010) and cytochrome c, are unaltered or slightly increased
- reference_id: file:DROME/Hsp83/Hsp83-deep-research-falcon.md
supporting_text: |-
Hsp83 is best understood as the **cytosolic Hsp90 ATP-dependent chaperone** that promotes **late-stage folding/maturation, stabilization, and activation** of diverse client proteins, especially those central to signaling and development
- term:
id: GO:0034605
label: cellular response to heat
evidence_type: IBA
original_reference_id: GO_REF:0000033
review:
summary: Hsp83 is a heat shock protein that is upregulated in response to heat
stress. IBA from multiple orthologs. Consistent with IEP evidence from PMID:16595740
showing Hsp83 protein levels increase after heat shock. Name itself (Heat shock
protein 83) reflects this function.
action: ACCEPT
reason: Cellular response to heat is a defining characteristic of heat shock proteins.
IBA supported by expression and functional data in Drosophila.
supported_by:
- reference_id: PMID:16595740
supporting_text: increases in the levels of Hsp83 and DnaJ-1 proteins but not
in the inducible form of Hsp70 were detected by Western blot analysis
- term:
id: GO:0051082
label: unfolded protein binding
evidence_type: IBA
original_reference_id: GO_REF:0000033
review:
summary: GO:0051082 "unfolded protein binding" is proposed for obsoletion. HSP90
does not simply bind unfolded proteins like holdase chaperones; rather, it acts
as an ATP-dependent foldase that binds near-native client proteins and assists
their maturation through conformational change driven by ATP hydrolysis. The
correct MF term is GO:0140662 "ATP-dependent protein folding chaperone."
action: MODIFY
reason: GO:0051082 is proposed for obsoletion and does not accurately describe
the HSP90 mechanism. HSP90 does not primarily bind unfolded proteins; it binds
near-native, partially folded client proteins and uses ATP hydrolysis to drive
their maturation. The replacement term GO:0140662 "ATP-dependent protein folding
chaperone" precisely captures the molecular function.
proposed_replacement_terms:
- id: GO:0140662
label: ATP-dependent protein folding chaperone
supported_by:
- reference_id: PMID:29775584
supporting_text: the Hsp90 system is required to extend the dwell time of Ago2
in the open state, which must be transiently primed by the Hsp70 system
- term:
id: GO:0048471
label: perinuclear region of cytoplasm
evidence_type: IBA
original_reference_id: GO_REF:0000033
review:
summary: IBA annotation to perinuclear region of cytoplasm is supported by phylogenetic
inference from multiple orthologs. Also supported by direct experimental evidence
in Drosophila (IDA, PMID:22902557) where Hsp83 was observed in perinuclear region
via the piRNA pathway work with Shutdown (Shu).
action: ACCEPT
reason: Consistent with IBA evidence and directly confirmed by IDA in Drosophila
(PMID:22902557).
- term:
id: GO:0000166
label: nucleotide binding
evidence_type: IEA
original_reference_id: GO_REF:0000043
review:
summary: IEA annotation from UniProtKB keyword mapping (KW-0547 "Nucleotide-binding").
Hsp83 is indeed an ATP-binding protein with a well-characterized nucleotide
binding pocket. This is a broader parent term of ATP binding (GO:0005524) which
is already annotated with IBA evidence.
action: ACCEPT
reason: Correct but redundant with the more specific ATP binding annotation. IEA
annotations at broader levels than IBA are acceptable.
- term:
id: GO:0005524
label: ATP binding
evidence_type: IEA
original_reference_id: GO_REF:0000120
review:
summary: IEA annotation for ATP binding from combined automated methods (InterPro:IPR001404,
InterPro:IPR019805, UniProtKB-KW:KW-0067). Correct and consistent with IBA annotation
for the same term.
action: ACCEPT
reason: Redundant with IBA annotation for the same GO term. Both are correct.
Hsp83 has a well-characterized HATPase domain.
- term:
id: GO:0005737
label: cytoplasm
evidence_type: IEA
original_reference_id: GO_REF:0000044
review:
summary: IEA annotation from UniProt subcellular location vocabulary mapping.
UniProt annotates Hsp83 subcellular location as "Cytoplasm." Consistent with
multiple IDA and HDA annotations for cytoplasm.
action: ACCEPT
reason: Correct. Cytoplasmic localization is well established by multiple independent
lines of evidence.
- term:
id: GO:0006457
label: protein folding
evidence_type: IEA
original_reference_id: GO_REF:0000002
review:
summary: IEA annotation from InterPro domain mapping (IPR001404, IPR019805). Protein
folding is the core biological process of HSP90 family. Consistent with IBA
and other evidence.
action: ACCEPT
reason: Correct. Redundant with IBA annotation. InterPro correctly maps the HSP90
domain to the protein folding process.
- term:
id: GO:0016887
label: ATP hydrolysis activity
evidence_type: IEA
original_reference_id: GO_REF:0000002
review:
summary: IEA annotation from InterPro domain mapping (IPR001404). ATP hydrolysis
is intrinsic to the HATPase domain of HSP90. Consistent with IBA.
action: ACCEPT
reason: Correct. Redundant with IBA annotation. InterPro correctly maps the HSP90
HATPase domain to ATP hydrolysis activity.
- term:
id: GO:0051082
label: unfolded protein binding
evidence_type: IEA
original_reference_id: GO_REF:0000002
review:
summary: IEA annotation from InterPro domain mapping. GO:0051082 is proposed for
obsoletion. HSP90 does not function as a holdase chaperone that binds unfolded
proteins; it is an ATP-dependent foldase that acts on near-native client proteins.
action: MODIFY
reason: Same issue as the IBA annotation for this term. GO:0051082 is proposed
for obsoletion and does not accurately represent the HSP90 mechanism. Should
be replaced with GO:0140662 "ATP-dependent protein folding chaperone."
proposed_replacement_terms:
- id: GO:0140662
label: ATP-dependent protein folding chaperone
- term:
id: GO:0101031
label: protein folding chaperone complex
evidence_type: IEA
original_reference_id: GO_REF:0000117
review:
summary: IEA annotation from ARBA machine learning model (ARBA:ARBA00026483).
HSP90 forms multiple chaperone complexes. Consistent with IDA evidence from
PMID:33176138 for the same term.
action: ACCEPT
reason: Correct. HSP90 is a core component of protein folding chaperone complexes.
Consistent with direct experimental evidence in Drosophila.
- term:
id: GO:0140662
label: ATP-dependent protein folding chaperone
evidence_type: IEA
original_reference_id: GO_REF:0000002
review:
summary: IEA annotation from InterPro domain mapping (IPR001404). GO:0140662 "ATP-dependent
protein folding chaperone" is the correct molecular function term for HSP90,
accurately capturing its ATP-dependent chaperone mechanism. This is the term
that should replace GO:0051082 across all annotations.
action: ACCEPT
reason: This is the most accurate MF term for HSP90 function. HSP90 binds client
proteins and assists their folding/maturation through ATP hydrolysis-driven
conformational changes.
supported_by:
- reference_id: PMID:29775584
supporting_text: the Hsp90 system is required to extend the dwell time of Ago2
in the open state
- term:
id: GO:0005515
label: protein binding
evidence_type: IPI
original_reference_id: PMID:22579285
review:
summary: IPI annotation from IntAct based on physical interaction with Nelf-E
(P92204). PMID:22579285 showed Hsp90 localizes to promoters and stabilizes the
NELF complex to maintain RNA polymerase II pausing. While the interaction is
real, "protein binding" is uninformative.
action: MARK_AS_OVER_ANNOTATED
reason: The interaction with Nelf-E is likely reflecting Hsp83's chaperone activity
on NELF complex stabilization. "Protein binding" is too generic and uninformative.
The chaperone function is better captured by GO:0140662, and the biological
outcome (gene expression regulation) is a downstream effect of chaperone activity.
supported_by:
- reference_id: PMID:22579285
supporting_text: Hsp90 maintains and optimizes RNA polymerase II pausing via
stabilization of the negative elongation factor complex (NELF)
- term:
id: GO:0034663
label: endoplasmic reticulum chaperone complex
evidence_type: IPI
original_reference_id: PMID:22099462
review:
summary: IPI annotation based on physical interaction evidence (PMID:22099462),
with interactors FB:FBgn0038749 and FB:FBgn0289994. PMID:22099462 identifies
XPORT as a chaperone for TRP and rhodopsin that interacts with Hsp90 in the
ER/secretory pathway. Hsp83 is primarily a cytoplasmic chaperone, and ER localization
would be atypical for the cytoplasmic HSP90 (as opposed to ER-resident GRP94/Hsp90B1).
action: MARK_AS_OVER_ANNOTATED
reason: |-
PMID:22099462 shows XPORT (a resident ER/secretory-pathway protein) interacts
with Hsp90, but the IPI-based CC annotation to an ER chaperone complex
over-interprets this transient interaction. Falcon deep research confirms
Hsp83 is the sole CYTOSOLIC Hsp90-family chaperone in D. melanogaster and is
primarily cytosolic; stable residence in an ER chaperone complex would be
atypical for the cytoplasmic HSP90 (the ER-resident Hsp90 paralog is GRP94/
Hsp90B1, encoded separately by Gp93 in flies). The most parsimonious
interpretation is a transient cytosolic-side interaction of Hsp83 with XPORT
during client (TRP/Rh1) processing rather than bona fide membership of an ER
chaperone complex. The chaperone activity itself is better captured by
GO:0140662 and GO:0101031.
supported_by:
- reference_id: PMID:22099462
supporting_text: XPORT is a resident ER and secretory pathway protein that interacts
with TRP and Rh1, as well as with Hsp27 and Hsp90
- reference_id: file:DROME/Hsp83/Hsp83-deep-research-falcon.md
supporting_text: |-
Hsp83 is the sole cytosolic Hsp90-family chaperone in *D. melanogaster* and functions as an ATP-dependent molecular chaperone that promotes late-stage folding, maturation, stabilization, and activation of signaling clients.
- term:
id: GO:0005739
label: mitochondrion
evidence_type: ISS
original_reference_id: GO_REF:0000024
review:
summary: ISS annotation transferred from human HSP90AA1 (P07900). A minor pool
of HSP90 has been reported in mitochondria in mammalian cells. Hsp83 interacts
with Sicily for chaperoning ND42 before mitochondrial import (PMID:23509070),
but this occurs in the cytoplasm. Mitochondrial localization of the fly protein
itself is not directly demonstrated.
action: KEEP_AS_NON_CORE
reason: Mitochondrial localization is plausible based on mammalian ortholog data
but not directly demonstrated in Drosophila. The cytoplasmic chaperoning of
mitochondrial proteins (PMID:23509070) does not require Hsp83 itself to localize
to mitochondria.
supported_by:
- reference_id: PMID:23509070
supporting_text: Sicily preprotein interacts with cytosolic Hsp90 to chaperone
the CI subunit, ND42, before mitochondrial import
- term:
id: GO:0097753
label: membrane bending
evidence_type: IDA
original_reference_id: PMID:30193096
review:
summary: IDA annotation based on direct demonstration in PMID:30193096 that Hsp90
can directly interact with and deform membranes via an evolutionarily conserved
amphipathic helix. This is a non-chaperone function structurally separated from
the protein folding activity.
action: KEEP_AS_NON_CORE
reason: Membrane bending is a genuine, experimentally validated function of Hsp90
mediated by its amphipathic helix. However, it is a non-canonical function distinct
from the core chaperone activity and therefore non-core. The paper explicitly
states they "structurally separated the Hsp90 membrane-deforming function from
its well-characterized chaperone activity."
supported_by:
- reference_id: PMID:30193096
supporting_text: Hsp90 also has the ability to directly interact with and deform
membranes via an evolutionarily conserved amphipathic helix
- reference_id: PMID:30193096
supporting_text: we structurally separated the Hsp90 membrane-deforming function
from its well-characterized chaperone activity
- term:
id: GO:0098866
label: multivesicular body fusion to apical plasma membrane
evidence_type: IMP
original_reference_id: PMID:30193096
review:
summary: IMP annotation from PMID:30193096. The amphipathic helix of Hsp90 promotes
the fusion of multivesicular bodies (MVBs) with the plasma membrane, enabling
exosome release. Mutations and drugs that stabilize the open Hsp90 dimer expose
the helix and allow MVB fusion.
action: KEEP_AS_NON_CORE
reason: This is a genuine but non-core function mediated by the non-chaperone
membrane-deforming activity of Hsp90. Not a typical chaperone function.
supported_by:
- reference_id: PMID:30193096
supporting_text: this amphipathic helix allows exosome release by promoting
the fusion of multivesicular bodies (MVBs) with the plasma membrane
- term:
id: GO:0070922
label: RISC complex assembly
evidence_type: IGI
original_reference_id: PMID:29775584
review:
summary: IGI annotation from PMID:29775584 with genetic interactors FB:FBgn0037728
and FB:FBgn0266599. The paper demonstrates that Hsp70/Hsp90 chaperone machinery
is required for loading small RNAs into Argonaute, a key step in RISC assembly.
The Hsp90 system captures and stabilizes the open, active form of Ago2.
action: ACCEPT
reason: RISC complex assembly is a well-characterized function of the Hsp90 chaperone
system. Hsp90 is required for conformational activation of Argonaute to enable
small RNA loading. This is a specific manifestation of its core chaperone function
on a defined client protein complex.
supported_by:
- reference_id: PMID:29775584
supporting_text: Loading of small RNAs into Argonaute, the core protein in RNA
silencing, requires the Hsp70/Hsp90 chaperone machinery
- reference_id: PMID:29775584
supporting_text: the Hsp90 system is required to extend the dwell time of Ago2
in the open state
- term:
id: GO:0070922
label: RISC complex assembly
evidence_type: IPI
original_reference_id: PMID:33176138
review:
summary: IPI annotation from PMID:33176138 with interactors FB:FBgn0036020, FB:FBgn0087035,
and FB:FBgn0262739. PMID:33176138 identifies Cyclophilin 40 as a testis-specialized
Hsp90 co-chaperone that modulates Ago2-RISC formation. Cyp40 interacts with
Ago2 through Hsp90 and promotes the build-up of Ago2-bound miRNAs.
action: ACCEPT
reason: Consistent with the IGI evidence from PMID:29775584 and with the known
role of HSP90 in Argonaute maturation and RISC assembly. Additional IPI evidence
strengthens the annotation.
supported_by:
- reference_id: PMID:33176138
supporting_text: Cyp40 interacts with duplex-incorporating Ago2 through Hsp90
in vitro and selectively promotes the build-up of Ago2-bound miRNAs
- term:
id: GO:0005515
label: protein binding
evidence_type: IPI
original_reference_id: PMID:31907206
review:
summary: IPI annotation from UniProt based on interaction with Morgana/CHORD (Q9VCC0).
PMID:31907206 showed that Morgana co-purifies with the Hsp90-R2TP-TTT supercomplex.
This is a typical co-chaperone interaction.
action: MARK_AS_OVER_ANNOTATED
reason: '"Protein binding" is uninformative. The interaction with Morgana reflects
Hsp83''s role as part of the Hsp90-R2TP-TTT chaperone supercomplex. This is
better captured by the chaperone complex annotations (GO:0101031) and the ATP-dependent
protein folding chaperone MF term (GO:0140662).'
supported_by:
- reference_id: PMID:31907206
supporting_text: Mora localises to mitotic spindles and co-purifies with the
Hsp90-R2TP-TTT supercomplex and with additional well-known Hsp90 co-chaperones
- term:
id: GO:0016887
label: ATP hydrolysis activity
evidence_type: ISS
original_reference_id: GO_REF:0000024
review:
summary: ISS annotation transferred from human HSP90AA1 (P07900). ATP hydrolysis
is well established for HSP90 family members. Consistent with IBA evidence for
the same term.
action: ACCEPT
reason: Correct. Redundant with IBA. ATP hydrolysis is a core function of the
HSP90 family.
- term:
id: GO:0101031
label: protein folding chaperone complex
evidence_type: IDA
original_reference_id: PMID:33176138
review:
summary: IDA annotation from PMID:33176138 directly demonstrating Hsp83 as part
of a protein folding chaperone complex. PMID:33176138 shows Hsp90 machinery
acts as chaperone complex for Ago2 loading during RISC assembly. HSP90 forms
complexes with co-chaperones (Hop, p23, Cdc37, Cyp40, etc.) to constitute chaperone
machinery.
action: ACCEPT
reason: HSP90 is a core component of chaperone complexes. IDA evidence directly
confirms this in Drosophila. Consistent with UniProt subunit annotations describing
multiple Hsp83-containing complexes.
supported_by:
- reference_id: PMID:33176138
supporting_text: In gene silencing, Hsp90 chaperone machinery assists Argonaute
(Ago) binding and unwinding of silencing small RNA (sRNA) duplexes
- term:
id: GO:0005515
label: protein binding
evidence_type: IPI
original_reference_id: PMID:23509070
review:
summary: IPI annotation from UniProt based on interaction with Sicily (P91929)
and ND-42/NDUFA10 (Q9VYS5). PMID:23509070 showed Sicily is a cytosolic chaperone
for a mitochondrial Complex I subunit, and Hsp83 forms a complex with Sicily
and ND42. Hsp90 binds to Sicily directly and coimmunoprecipitates ND42 only
when Sicily is coexpressed.
action: MARK_AS_OVER_ANNOTATED
reason: '"Protein binding" is uninformative. The interaction with Sicily reflects
Hsp83''s chaperone function in chaperoning ND42 before mitochondrial import.
Better captured by the ATP-dependent protein folding chaperone MF term (GO:0140662)
and the protein stabilization BP term (GO:0050821).'
supported_by:
- reference_id: PMID:23509070
supporting_text: Hsp90 binds to Sicily in an in vitro co-IP assay
- term:
id: GO:0002052
label: positive regulation of neuroblast proliferation
evidence_type: IMP
original_reference_id: PMID:30245208
review:
summary: IMP annotation from PMID:30245208. Hsp83 is both necessary and sufficient
for neural stem cell reactivation by promoting the activation of InR pathway
in larval brains. This is a downstream pleiotropic effect of Hsp83 chaperone
function on signaling clients.
action: KEEP_AS_NON_CORE
reason: This is a downstream developmental phenotype of Hsp83 function, likely
mediated through its role in insulin receptor signaling (same paper provides
insulin receptor binding annotation). Non-core pleiotropic effect.
supported_by:
- reference_id: PMID:30245208
supporting_text: Hsp83 is both necessary and sufficient for NSC reactivation
by promoting the activation of InR pathway in larval brains in the presence
of dietary amino acids
- term:
id: GO:0005158
label: insulin receptor binding
evidence_type: IDA
original_reference_id: PMID:30245208
review:
summary: IDA annotation from PMID:30245208 demonstrating direct binding of Hsp83
to the insulin receptor. Both Hsp83 and its co-chaperone Cdc37 physically associate
with InR. HSP90 is known to chaperone receptor tyrosine kinases in other systems.
This likely reflects HSP90's general role in chaperoning kinase clients.
action: KEEP_AS_NON_CORE
reason: Insulin receptor binding is consistent with HSP90's well-known role as
a kinase chaperone, but it represents a specific client interaction rather than
a core molecular function. The core function is the ATP-dependent protein folding
chaperone activity.
supported_by:
- reference_id: PMID:30245208
supporting_text: Both Hsp83 and its co-chaperone Cdc37 physically associate
with InR
- term:
id: GO:0046628
label: positive regulation of insulin receptor signaling pathway
evidence_type: IGI
original_reference_id: PMID:30245208
review:
summary: IGI annotation from PMID:30245208 with genetic interactor FB:FBgn0283499.
Hsp83 positively regulates insulin receptor signaling, consistent with its role
in chaperoning the insulin receptor.
action: KEEP_AS_NON_CORE
reason: Downstream pleiotropic effect of Hsp83 chaperone function on insulin signaling.
Non-core but experimentally supported.
supported_by:
- reference_id: PMID:30245208
supporting_text: Hsp83 is both necessary and sufficient for NSC reactivation
by promoting the activation of InR pathway in larval brains in the presence
of dietary amino acids
- term:
id: GO:0005737
label: cytoplasm
evidence_type: HDA
original_reference_id: PMID:26896675
review:
summary: HDA annotation from a genome-wide protein localization study (PMID:26896675)
using GFP-tagged fosmid transgenes in Drosophila. Cytoplasmic localization is
consistent with all other evidence.
action: ACCEPT
reason: Cytoplasm is the primary localization for Hsp83. Confirmed by multiple
independent methods.
supported_by:
- reference_id: PMID:26896675
supporting_text: A genome-wide resource for the analysis of protein localisation
in Drosophila
- term:
id: GO:0030911
label: TPR domain binding
evidence_type: IPI
original_reference_id: PMID:27979731
review:
summary: 'IPI annotation from PMID:27979731 based on interaction of Hsp83 with
the TPR domain of Nup358. The C-terminal MEEVD motif of HSP90 is a well-characterized
TPR repeat-binding motif (UniProt: "The TPR repeat-binding motif mediates interaction
with TPR repeat-containing proteins"). The interaction with Nup358 TPR domain
facilitates nuclear import of Met, the juvenile hormone receptor.'
action: ACCEPT
reason: TPR domain binding is a core molecular function of HSP90 family members,
mediated by the conserved C-terminal MEEVD motif. This is how HSP90 interacts
with many co-chaperones (Hop, CHIP, Tom70, etc.) and other TPR-containing proteins.
supported_by:
- reference_id: PMID:27979731
supporting_text: a tetratricopeptide repeat (TPR) domain at the N-terminal end
of Nup358 interacts with Hsp83 and is indispensable for Met nuclear localization
- reference_id: file:DROME/Hsp83/Hsp83-deep-research-falcon.md
supporting_text: |-
Key named regulators include **Cdc37** (kinase client recruitment), **p23**, and **Aha1**, as well as **TPR-domain** proteins such as **Hop** that bind the conserved **MEEVD** C-terminal motif typical of cytosolic Hsp90s
- term:
id: GO:0006457
label: protein folding
evidence_type: ISS
original_reference_id: GO_REF:0000024
review:
summary: ISS annotation transferred from yeast HSP82 (P15108). Protein folding
is well established for HSP90 family. Consistent with IBA and IEA evidence.
action: ACCEPT
reason: Correct. Redundant with IBA and IEA. Protein folding is the core biological
process.
- term:
id: GO:0034605
label: cellular response to heat
evidence_type: ISS
original_reference_id: GO_REF:0000024
review:
summary: ISS annotation transferred from yeast HSP82 (P15108). Cellular response
to heat is well established for heat shock proteins. Consistent with IBA evidence.
action: ACCEPT
reason: Correct. Redundant with IBA. Heat shock response is a defining feature
of HSP90 family.
- term:
id: GO:0043248
label: proteasome assembly
evidence_type: IDA
original_reference_id: PMID:15946124
review:
summary: IDA annotation from PMID:15946124. Hsp82 (Hsp83) was identified by mass
spectrometry as the main Rpn10/p54-interacting protein after Zn2+-induced proteasome
disassembly, suggesting a role in reassembly of the 26S proteasome.
action: KEEP_AS_NON_CORE
reason: The evidence shows Hsp83 interacts with Rpn10 after proteasome disassembly,
suggesting a role in reassembly. This is consistent with HSP90's general chaperone
function applied to proteasome components, but represents a specific client
relationship rather than a core function.
supported_by:
- reference_id: PMID:15946124
supporting_text: Hsp82 (heat-shock protein 82) has been identified by MS as
the main Rpn10/p54-interacting protein, suggesting its role in the reassembly
of the 26 S proteasome after Zn2+ removal
- term:
id: GO:0005737
label: cytoplasm
evidence_type: HDA
original_reference_id: PMID:24292889
review:
summary: HDA annotation from PMID:24292889 (Ube3a proteasomal shuttle Rpn10 study).
Cytoplasmic localization detected as part of a high-throughput study. Consistent
with other evidence.
action: ACCEPT
reason: Cytoplasm localization is well established. Additional HDA evidence.
supported_by:
- reference_id: PMID:24292889
supporting_text: Overexpression of these proteins leads to an enhanced accumulation
of ubiquitinated proteins, further supporting the biochemical evidence of interaction
obtained in neuronal cells
- term:
id: GO:0005886
label: plasma membrane
evidence_type: HDA
original_reference_id: PMID:19317464
review:
summary: HDA annotation from PMID:19317464 (LOPIT subcellular proteomics study
in Drosophila embryos). Plasma membrane localization detected in this systematic
study. Consistent with IBA annotation and the amphipathic helix-mediated membrane
interaction (PMID:30193096).
action: ACCEPT
reason: Plasma membrane localization supported by independent proteomics evidence
and consistent with IBA and the membrane-deforming function.
supported_by:
- reference_id: PMID:19317464
supporting_text: we apply LOPIT, a mass-spectrometry based technique that simultaneously
maps proteins to specific subcellular compartments, to Drosophila embryos
- term:
id: GO:0051082
label: unfolded protein binding
evidence_type: IDA
original_reference_id: PMID:18344983
review:
summary: IDA annotation from PMID:18344983. This paper is primarily about NMNAT
as a chaperone, not about Hsp83 directly. The paper shows that NMNAT "displays
chaperone function both in biochemical assays and cultured cells" and compares
it to Hsp70. Hsp83 is mentioned as a control or comparison. Regardless, GO:0051082
is proposed for obsoletion and the term does not correctly describe HSP90 function.
HSP90 is an ATP-dependent foldase, not a holdase.
action: MODIFY
reason: GO:0051082 is proposed for obsoletion. The evidence in PMID:18344983 is
primarily about NMNAT chaperone function, with Hsp83 as a comparison. The correct
term for Hsp83 molecular function is GO:0140662 "ATP-dependent protein folding
chaperone."
proposed_replacement_terms:
- id: GO:0140662
label: ATP-dependent protein folding chaperone
supported_by:
- reference_id: PMID:18344983
supporting_text: NMNAT displays chaperone function both in biochemical assays
and cultured cells, and it shares significant structural similarity with known
chaperones
- term:
id: GO:0008285
label: negative regulation of cell population proliferation
evidence_type: IMP
original_reference_id: PMID:24086162
review:
summary: IMP annotation from PMID:24086162. Hsp83 mutation results in increased
E2F-dependent transcription and ectopic cell proliferation in pupal tissues,
demonstrating that Hsp83 is required for proper cell cycle exit. The paper proposes
Cdh1/Fzr (APC/C activating subunit) is a client of Hsp83. Thus Hsp83 indirectly
promotes cell cycle exit by chaperoning APC/C components.
action: KEEP_AS_NON_CORE
reason: This is a downstream phenotypic consequence of Hsp83 chaperone function
on cell cycle regulators (APC/C components). The paper proposes "Cdh1/Fzr is
a client protein of Hsp83." This is a pleiotropic effect of the core chaperone
function, not a core function itself.
supported_by:
- reference_id: PMID:24086162
supporting_text: Mutation of Hsp83, the Drosophila homolog of mammalian Hsp90,
results in increased E2F-dependent transcription and ectopic cell proliferation
in pupal tissues at a time when neighboring wild-type cells are postmitotic
- reference_id: PMID:24086162
supporting_text: we propose that Cdh1/Fzr is a client protein of Hsp83
- reference_id: file:DROME/Hsp83/Hsp83-deep-research-falcon.md
supporting_text: |-
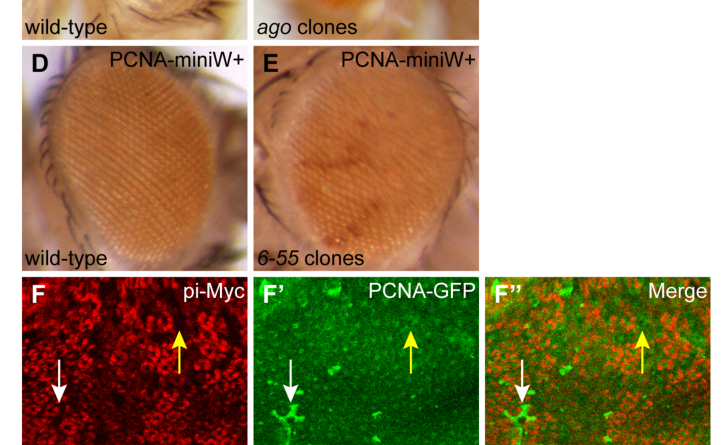
In Hsp83(6-55) mutant clones, E2F-dependent transcription increases and cells show ectopic S phase and mitosis after neighboring cells have exited the cell cycle
- term:
id: GO:0048471
label: perinuclear region of cytoplasm
evidence_type: IDA
original_reference_id: PMID:22902557
review:
summary: IDA annotation from PMID:22902557. This paper studies the cochaperone
Shutdown and piRNA biogenesis. Using a genomic GFP-tagged Hsp83 construct, the
authors show that Hsp83 is enriched in a peri-nuclear rim in wild-type nurse
cells, potentially reflecting nuage accumulation. Hsp83 colocalizes with unloaded
AGO3 in cytoplasmic foci when piRNA biogenesis is disrupted.
action: ACCEPT
reason: Perinuclear region localization is directly demonstrated by IDA using
GFP-tagged Hsp83 and is consistent with IBA evidence and the known role of Hsp83
in piRNA biogenesis in germline nuage.
supported_by:
- reference_id: PMID:22902557
supporting_text: Hsp83 was enriched in a peri-nuclear rim, potentially reflecting
nuage accumulation
- term:
id: GO:0005705
label: polytene chromosome interband
evidence_type: IDA
original_reference_id: PMID:19144915
review:
summary: IDA annotation from PMID:19144915. This paper shows a direct molecular
interaction between Hsp90 and Trithorax, and that Hsp90 cooperates with Trx
at chromatin for maintaining the active expression state of targets like Hox
genes. Polytene chromosome interband localization is consistent with the chromatin-binding
function of Hsp90.
action: ACCEPT
reason: PMID:19144915 demonstrates Hsp90 interacts with Trithorax at chromatin
to maintain active gene expression. Polytene chromosome interband localization
is consistent with this chromatin-associated function and with the finding from
PMID:22579285 that Hsp90 localizes to promoters.
supported_by:
- reference_id: PMID:19144915
supporting_text: Hsp90 cooperates with Trx at chromatin for maintaining the
active expression state of targets like the Hox genes
additional_reference_ids:
- PMID:22579285
- term:
id: GO:0005737
label: cytoplasm
evidence_type: IDA
original_reference_id: PMID:19144915
review:
summary: IDA annotation from PMID:19144915. Cytoplasmic localization is consistent
with all other evidence.
action: ACCEPT
reason: Cytoplasm localization is independently confirmed by many other studies.
This annotation is redundant but correct.
supported_by:
- reference_id: PMID:19144915
supporting_text: Molecular chaperone heat-shock protein 90 kDa (Hsp90) is known
to facilitate the conformational maturation of a diverse range of proteins
involved in different signal transduction pathways during development
- term:
id: GO:0009631
label: cold acclimation
evidence_type: IEP
original_reference_id: PMID:16313561
review:
summary: IEP annotation from PMID:16313561. Microarray analysis showed Hsp83 transcript
was differentially expressed after cold hardening treatment (0 C for 2 h followed
by recovery). The evidence is expression-based only (IEP), and does not demonstrate
a direct functional role.
action: KEEP_AS_NON_CORE
reason: Based solely on expression evidence (IEP). Transcript upregulation during
cold hardening does not prove a direct role in cold acclimation. However, Hsp83
may contribute to protein homeostasis under cold stress. Keeping as non-core
due to indirect evidence.
supported_by:
- reference_id: PMID:16313561
supporting_text: stress proteins, including Hsp23, Hsp26, Hsp83 and Frost as
well as membrane-associated proteins may contribute to the cold hardening
response
- term:
id: GO:0009408
label: response to heat
evidence_type: IEP
original_reference_id: PMID:16595740
review:
summary: IEP annotation from PMID:16595740. Hsp83 protein levels were shown to
increase after heat shock by Western blot analysis, even in a heat shock factor
mutant (hsf4). As a constitutively expressed heat shock protein, Hsp83 upregulation
during heat stress provides thermoprotection at synapses.
action: ACCEPT
reason: |-
While IEP is weaker evidence, the response to heat is fundamental to heat
shock proteins. The name "Heat shock protein 83" itself reflects this. The more
specific term "cellular response to heat" (GO:0034605) is also annotated with
IBA and ISS evidence. Falcon deep research adds an important nuance: Hsp83 is
largely a constitutive proteostasis factor and is NOT transcriptionally induced
in every proteotoxic stress state (e.g., in a 2023 larval fat body study Hsp83
and Gp93 did not change despite induction of Hsp70/Hsp40-class genes). The
response-to-heat annotation remains valid (protein levels do rise after heat
shock per PMID:16595740), but it reflects a partially constitutive, context-
dependent thermoprotective role rather than a universally inducible response.
supported_by:
- reference_id: PMID:16595740
supporting_text: the compensatory upregulation of constitutively expressed Hsps,
in the absence of the synthesis of the major inducible Hsp, Hsp70, could still
provide substantial thermoprotection
- reference_id: file:DROME/Hsp83/Hsp83-deep-research-falcon.md
supporting_text: |-
A 2023 Nature Communications study in Drosophila larval fat body found that **Nacα reduction activates a heat shock response**, with upregulation of Hsp70/Hsp40 class genes and oxidative stress markers, but **Hsp83 (and another Hsp90-family gene Gp93) did not change** in that context (qRT-PCR, n=4 batches)
- term:
id: GO:0005737
label: cytoplasm
evidence_type: IDA
original_reference_id: PMID:19101615
review:
summary: IDA annotation from PMID:19101615. Hsp90 and Cup co-localize in the cytoplasm
of developing germ-line cells within the germarium.
action: ACCEPT
reason: Cytoplasmic localization directly demonstrated during oogenesis. Consistent
with all other cytoplasm annotations.
supported_by:
- reference_id: PMID:19101615
supporting_text: Hsp90 and Cup co-localize in the cytoplasm of the developing
germ-line cells within the germarium
- term:
id: GO:0048477
label: oogenesis
evidence_type: IGI
original_reference_id: PMID:19101615
review:
summary: IGI annotation from PMID:19101615 with genetic interactor Cup (FB:FBgn0000392).
Lowering Hsp90 activity enhances oogenesis alterations linked to diverse cup
mutant alleles, demonstrating a genetic interaction in oogenesis. Hsp90 is a
component of the cap-binding complex and interacts with the translational repressor
Cup during oogenesis.
action: KEEP_AS_NON_CORE
reason: Oogenesis involvement is a pleiotropic consequence of Hsp83 chaperone
function during development. The paper shows Hsp90 participates in translational
regulation during oogenesis via the cap-binding complex. This is a downstream
developmental role, not a core function.
supported_by:
- reference_id: PMID:19101615
supporting_text: lowering Hsp90 activity enhances the oogenesis alterations
linked to diverse cup mutant alleles
- reference_id: file:DROME/Hsp83/Hsp83-deep-research-falcon.md
supporting_text: |-
Hsp83 also contributes to female fertility, oogenesis, and maternal RNA regulation. It interacts with the translational regulator Cup during oogenesis, localizes maternal transcripts, and affects *nanos* mRNA localization, linking Hsp83 to anterior–posterior patterning in embryos.
- term:
id: GO:0019094
label: pole plasm mRNA localization
evidence_type: IMP
original_reference_id: PMID:17565952
review:
summary: IMP annotation from PMID:17565952. Genetic approaches show that Hsp83/Hsp90
is a localization factor for nanos and pgc mRNAs. When Hsp90 function is partially
compromised, nanos and pgc mRNA localization is disrupted while other pole plasm
components are unaffected. The mechanism appears to involve Hsp90 chaperoning
LKB1 kinase (for pgc) and other partners (for nos).
action: KEEP_AS_NON_CORE
reason: This is a specific developmental role of Hsp83 in mRNA localization during
embryogenesis, likely mediated through its chaperone function on LKB1 kinase
and other clients. Non-core pleiotropic effect.
supported_by:
- reference_id: PMID:17565952
supporting_text: the Hsp90 chaperone (encoded by Hsp83 in Drosophila) is a localization
factor for two mRNAs, nanos and pgc
- reference_id: PMID:17565952
supporting_text: levels of the LKB1 kinase are reduced in Hsp83 mutant egg chambers
- reference_id: file:DROME/Hsp83/Hsp83-deep-research-falcon.md
supporting_text: |-
It interacts with the translational regulator Cup during oogenesis, localizes maternal transcripts, and affects *nanos* mRNA localization, linking Hsp83 to anterior–posterior patterning in embryos.
- term:
id: GO:0005813
label: centrosome
evidence_type: IDA
original_reference_id: PMID:10716925
review:
summary: IDA annotation from PMID:10716925. Mass spectrometry analysis of immunoisolated
Drosophila centrosomes identified Hsp83. The association persists after microtubule
depolymerizing drug treatment, indicating Hsp90 is a core centrosomal component.
action: ACCEPT
reason: Directly demonstrated by mass spectrometry of purified centrosomes and
confirmed to be a core centrosomal component that persists after microtubule
depolymerization.
supported_by:
- reference_id: PMID:10716925
supporting_text: a fraction of the total Hsp90 pool is localized at the centrosome
throughout the cell cycle
- reference_id: PMID:10716925
supporting_text: This association between Hsp90 and the centrosome can be observed
in purified centrosomes and after treatment with microtubule depolymerizing
drugs, two criteria normally used to define core centrosomal components
- term:
id: GO:0006457
label: protein folding
evidence_type: TAS
original_reference_id: PMID:10716925
review:
summary: TAS annotation from PMID:10716925 which describes Hsp83 as "a member
of the highly conserved Hsp90 family including chaperones known to maintain
the activity of many proteins." The protein folding function is stated as established
fact in the publication.
action: ACCEPT
reason: Correct. Protein folding is the core function. Redundant with IBA, IEA,
and ISS evidence.
supported_by:
- reference_id: PMID:10716925
supporting_text: a member of the highly conserved Hsp90 family including chaperones
known to maintain the activity of many proteins
- term:
id: GO:0007098
label: centrosome cycle
evidence_type: IMP
original_reference_id: PMID:10716925
review:
summary: IMP annotation from PMID:10716925. Disruption of Hsp90 function by mutations
in Hsp83 or geldanamycin treatment results in abnormal centrosome separation
and maturation, aberrant spindles, and impaired chromosome segregation.
action: KEEP_AS_NON_CORE
reason: Centrosome cycle involvement is a downstream consequence of Hsp83 chaperone
function on centrosome client proteins. Hsp90 is required for proper centrosome
function, but this is a pleiotropic effect of its core chaperone activity.
supported_by:
- reference_id: PMID:10716925
supporting_text: Disruption of Hsp90 function by mutations in the Drosophila
gene or treatment of mammalian cells with the Hsp90 inhibitor geldanamycin,
results in abnormal centrosome separation and maturation, aberrant spindles
and impaired chromosome segregation
- term:
id: GO:0045187
label: regulation of circadian sleep/wake cycle, sleep
evidence_type: IMP
original_reference_id: PMID:12015603
review:
summary: IMP annotation from PMID:12015603. Flies carrying a mutation for Hsp83
showed exaggerated homeostatic sleep response and died after sleep deprivation,
demonstrating that Hsp83 plays a protective role in sleep homeostasis.
action: KEEP_AS_NON_CORE
reason: Sleep regulation is a pleiotropic downstream effect. Hsp83 mutants show
exaggerated homeostatic response to sleep deprivation, but this likely reflects
a general stress-protective role rather than a direct sleep-regulatory function.
supported_by:
- reference_id: PMID:12015603
supporting_text: flies carrying a mutation for the heat-shock protein Hsp83
(Hsp83(08445)) showed exaggerated homeostatic response and died after sleep
deprivation
- term:
id: GO:0045187
label: regulation of circadian sleep/wake cycle, sleep
evidence_type: TAS
original_reference_id: PMID:12486704
review:
summary: TAS annotation from PMID:12486704. This is a review article discussing
sleep research in Drosophila and mouse, referencing the genetic approaches that
identified Hsp83 mutants with sleep phenotypes.
action: KEEP_AS_NON_CORE
reason: Consistent with the IMP annotation from PMID:12015603 for the same term.
Sleep regulation is a non-core pleiotropic effect.
supported_by:
- reference_id: PMID:12486704
supporting_text: studies in both the mouse and fly have begun to reveal tantalizing
suggestions about the underlying principles that regulate sleep homeostasis
references:
- id: GO_REF:0000002
title: Gene Ontology annotation through association of InterPro records with GO
terms
findings: []
- id: GO_REF:0000024
title: Manual transfer of experimentally-verified manual GO annotation data to orthologs
by curator judgment of sequence similarity
findings: []
- id: GO_REF:0000033
title: Annotation inferences using phylogenetic trees
findings: []
- id: GO_REF:0000043
title: Gene Ontology annotation based on UniProtKB/Swiss-Prot keyword mapping
findings: []
- id: GO_REF:0000044
title: Gene Ontology annotation based on UniProtKB/Swiss-Prot Subcellular Location
vocabulary mapping, accompanied by conservative changes to GO terms applied by
UniProt
findings: []
- id: GO_REF:0000117
title: Electronic Gene Ontology annotations created by ARBA machine learning models
findings: []
- id: GO_REF:0000120
title: Combined Automated Annotation using Multiple IEA Methods
findings: []
- id: PMID:10716925
title: Hsp90 is a core centrosomal component and is required at different stages
of the centrosome cycle in Drosophila and vertebrates.
findings:
- statement: Hsp83 identified by mass spectrometry in purified Drosophila centrosomes
supporting_text: "One of the proteins identified is Hsp83, a member of the highly conserved Hsp90 family including chaperones known to maintain the activity of many proteins"
- statement: A fraction of total Hsp90 pool is localized at the centrosome throughout the cell cycle
supporting_text: "a fraction of the total Hsp90 pool is localized at the centrosome throughout the cell cycle at different stages of development in Drosophila and vertebrates"
- statement: Disruption of Hsp90 function causes abnormal centrosome separation and maturation
supporting_text: "Disruption of Hsp90 function by mutations in the Drosophila gene or treatment of mammalian cells with the Hsp90 inhibitor geldanamycin, results in abnormal centrosome separation and maturation, aberrant spindles and impaired chromosome segregation"
- id: PMID:12015603
title: Stress response genes protect against lethal effects of sleep deprivation
in Drosophila.
findings:
- statement: Hsp83 mutants show exaggerated homeostatic sleep response and die after sleep deprivation
supporting_text: "flies carrying a mutation for the heat-shock protein Hsp83 (Hsp83(08445)) showed exaggerated homeostatic response and died after sleep deprivation"
- statement: Heat-shock genes protect against lethal effects of sleep deprivation
supporting_text: "activating heat-shock genes before sleep deprivation rescued cyc(01) flies from its lethal effects"
- id: PMID:12486704
title: 'Perchance to dream: solving the mystery of sleep through genetic analysis.'
findings: []
- id: PMID:15946124
title: Zn2+-induced reversible dissociation of subunit Rpn10/p54 of the Drosophila
26 S proteasome.
findings:
- statement: Hsp82 identified by MS as the main Rpn10/p54-interacting protein after proteasome disassembly
supporting_text: "Hsp82 (heat-shock protein 82) has been identified by MS as the main Rpn10/p54-interacting protein, suggesting its role in the reassembly of the 26 S proteasome after Zn2+ removal"
- statement: Suggests Hsp83 role in reassembly of 26S proteasome
supporting_text: "removal of Zn2+ is followed by reassociation of subunit Rpn10/p54 to the RP, reassembly of the 26 S proteasome and resumption of the peptidase activity"
- id: PMID:16313561
title: Cold hardening and transcriptional change in Drosophila melanogaster.
findings:
- statement: Hsp83 transcript upregulated after cold hardening treatment
supporting_text: "stress proteins, including Hsp23, Hsp26, Hsp83 and Frost as well as membrane-associated proteins may contribute to the cold hardening response"
- id: PMID:16595740
title: Thermoprotection of synaptic transmission in a Drosophila heat shock factor
mutant is accompanied by increased expression of Hsp83 and DnaJ-1.
findings:
- statement: Hsp83 protein levels increase after heat shock even in hsf4 mutants
supporting_text: "increases in the levels of Hsp83 and DnaJ-1 proteins but not in the inducible form of Hsp70 were detected by Western blot analysis"
- statement: Compensatory upregulation of constitutive Hsps provides thermoprotection
supporting_text: "the compensatory upregulation of constitutively expressed Hsps, in the absence of the synthesis of the major inducible Hsp, Hsp70, could still provide substantial thermoprotection"
- id: PMID:17565952
title: The molecular chaperone Hsp90 is required for mRNA localization in Drosophila
melanogaster embryos.
findings:
- statement: Hsp83/Hsp90 is a localization factor for nanos and pgc mRNAs
supporting_text: "the Hsp90 chaperone (encoded by Hsp83 in Drosophila) is a localization factor for two mRNAs, nanos and pgc"
- statement: LKB1 kinase levels reduced in Hsp83 mutant egg chambers
supporting_text: "levels of the LKB1 kinase are reduced in Hsp83 mutant egg chambers"
- id: PMID:18344983
title: NAD synthase NMNAT acts as a chaperone to protect against neurodegeneration.
findings:
- statement: NMNAT displays chaperone function similar to heat-shock proteins
supporting_text: "NMNAT displays chaperone function both in biochemical assays and cultured cells, and it shares significant structural similarity with known chaperones"
- statement: Hsp83 used as comparison/control in chaperone assays
supporting_text: "it is upregulated in the brain upon overexpression of poly-glutamine expanded protein and recruited with the chaperone Hsp70 into protein aggregates"
- id: PMID:19101615
title: The molecular chaperone Hsp90 is a component of the cap-binding complex and
interacts with the translational repressor Cup during Drosophila oogenesis.
findings:
- statement: Hsp90 isolated as component of cap-binding complex
supporting_text: "isolated the product of the Hsp83 gene, the evolutionarily conserved chaperone Hsp90, as a specific component of the cap-binding complex"
- statement: Hsp90 interacts with translational repressor Cup
supporting_text: "Hsp90 interacts in vitro with the translational repressor Cup"
- statement: Hsp83 and cup interact genetically in oogenesis
supporting_text: "lowering Hsp90 activity enhances the oogenesis alterations linked to diverse cup mutant alleles"
- statement: Hsp90 and Cup co-localize in cytoplasm of germline cells
supporting_text: "Hsp90 and Cup co-localize in the cytoplasm of the developing germ-line cells within the germarium"
- id: PMID:19144915
title: Trithorax requires Hsp90 for maintenance of active chromatin at sites of gene
expression.
findings:
- statement: Hsp90 interacts directly with Trithorax and cooperates at chromatin for maintaining active expression
supporting_text: "Hsp90 cooperates with Trx at chromatin for maintaining the active expression state of targets like the Hox genes"
- statement: Pharmacological inhibition of Hsp90 results in degradation of Trx and down-regulation of homeotic gene expression
supporting_text: "Pharmacological inhibition of Hsp90 results in degradation of Trx and a concomitant down-regulation of homeotic gene expression"
- id: PMID:19317464
title: Mapping organelle proteins and protein complexes in Drosophila melanogaster.
findings:
- statement: LOPIT proteomics maps Hsp83 subcellular distribution in Drosophila embryos
supporting_text: "we apply LOPIT, a mass-spectrometry based technique that simultaneously maps proteins to specific subcellular compartments, to Drosophila embryos"
- id: PMID:22099462
title: XPORT-dependent transport of TRP and rhodopsin.
findings:
- statement: XPORT interacts with TRP, Rh1, Hsp27, and Hsp90 as part of ER chaperone function
supporting_text: "XPORT is a resident ER and secretory pathway protein that interacts with TRP and Rh1, as well as with Hsp27 and Hsp90"
- statement: XPORT promotes targeting of TRP to the membrane in Drosophila S2 cells
supporting_text: "XPORT promotes the targeting of TRP to the membrane in Drosophila S2 cells"
- id: PMID:22579285
title: Hsp90 globally targets paused RNA polymerase to regulate gene expression
in response to environmental stimuli.
findings:
- statement: Hsp90 localizes to promoters of many genes
supporting_text: "It localizes near promoters of many coding and noncoding genes including microRNAs"
- statement: Hsp90 stabilizes NELF complex to maintain RNA Pol II pausing
supporting_text: "Hsp90 maintains and optimizes RNA polymerase II pausing via stabilization of the negative elongation factor complex (NELF)"
- statement: Hsp90 inhibition leads to upregulation of target genes
supporting_text: "Inhibition of Hsp90 leads to upregulation of target genes"
- id: PMID:22902557
title: The cochaperone shutdown defines a group of biogenesis factors essential for
all piRNA populations in Drosophila.
findings:
- statement: Hsp83 interacts with cochaperone Shutdown (Shu)
supporting_text: "unloaded AGO3 that localizes to cytoplasmic foci interacts with Hsp83"
- statement: Required for piRNA biogenesis
supporting_text: "our genetic and cell biological data point toward an important role for the Hsp83 machinery in piRNA biogenesis"
- statement: Hsp83 enriched in peri-nuclear rim in wild-type nurse cells
supporting_text: "Hsp83 was enriched in a peri-nuclear rim, potentially reflecting nuage accumulation"
- id: PMID:23509070
title: The C8ORF38 homologue Sicily is a cytosolic chaperone for a mitochondrial
complex I subunit.
findings:
- statement: Hsp83 forms complex with Sicily and ND42 for mitochondrial protein chaperoning
supporting_text: "Sicily preprotein interacts with cytosolic Hsp90 to chaperone the CI subunit, ND42, before mitochondrial import"
- statement: Hsp90 binds to Sicily directly and coimmunoprecipitates ND42 only when Sicily is coexpressed
supporting_text: "Hsp90 binds to Sicily in an in vitro co-IP assay"
- statement: Loss of Hsp90 leads to decreased levels of Sicily and CI subunits in vivo
supporting_text: "flies expressing Hsp90 RNAi #1 exhibit severely decreased levels of Sicily, ND42, and NDUFS3 compared with controls"
- id: PMID:24086162
title: The molecular chaperone Hsp90 is required for cell cycle exit in Drosophila
melanogaster.
findings:
- statement: Hsp83 mutation causes increased E2F transcription and ectopic cell proliferation
supporting_text: "Mutation of Hsp83, the Drosophila homolog of mammalian Hsp90, results in increased E2F-dependent transcription and ectopic cell proliferation in pupal tissues at a time when neighboring wild-type cells are postmitotic"
- statement: APC/C targets accumulate in Hsp83 mutant cells
supporting_text: "these Hsp83 mutant cells have increased Cyclin/Cdk activity and accumulate proteins normally targeted for proteolysis by the anaphase-promoting complex/cyclosome (APC/C)"
- statement: Cdh1/Fzr proposed as an Hsp83 client protein
supporting_text: "we propose that Cdh1/Fzr is a client protein of Hsp83"
- id: PMID:24292889
title: Ube3a, the E3 ubiquitin ligase causing Angelman syndrome and linked to autism,
regulates protein homeostasis through the proteasomal shuttle Rpn10.
findings: []
- id: PMID:26896675
title: A genome-wide resource for the analysis of protein localisation in Drosophila.
findings:
- statement: Genome-wide GFP tagging resource for protein localization
supporting_text: "a genome-wide fosmid library of 10000 GFP-tagged clones, comprising tagged genes and most of their regulatory information"
- id: PMID:27979731
title: "Nucleoporin Nup358 facilitates nuclear import of Methoprene-tolerant (Met)\
\ in an importin \u03B2- and Hsp83-dependent manner."
findings:
- statement: Nup358 TPR domain interacts with Hsp83
supporting_text: "a tetratricopeptide repeat (TPR) domain at the N-terminal end of Nup358 interacts with Hsp83 and is indispensable for Met nuclear localization"
- statement: Interaction required for nuclear import of Met (JH receptor)
supporting_text: "Nup358 facilitates JH-induced Met nuclear transport in a manner dependent on importin \u03B2 and Hsp83"
- id: PMID:29775584
title: Conformational Activation of Argonaute by Distinct yet Coordinated Actions
of the Hsp70 and Hsp90 Chaperone Systems.
findings:
- statement: Hsp90 system captures and stabilizes the open active form of Ago2
supporting_text: "the Hsp90 system is required to extend the dwell time of Ago2 in the open state"
- statement: Required for small RNA loading into Argonaute
supporting_text: "Loading of small RNAs into Argonaute, the core protein in RNA silencing, requires the Hsp70/Hsp90 chaperone machinery"
- statement: Hsp70 system expands structural ensembles, Hsp90 stabilizes active form
supporting_text: "the Hsp70 system expands the structural ensembles of Ago2 and the Hsp90 system captures and stabilizes the active form"
- id: PMID:30193096
title: Hsp90 Mediates Membrane Deformation and Exosome Release.
findings:
- statement: Hsp90 deforms membranes via amphipathic helix
supporting_text: "Hsp90 also has the ability to directly interact with and deform membranes via an evolutionarily conserved amphipathic helix"
- statement: Membrane function structurally separated from chaperone activity
supporting_text: "we structurally separated the Hsp90 membrane-deforming function from its well-characterized chaperone activity"
- statement: Open Hsp90 dimer exposes helix for MVB fusion
supporting_text: "mutations and drugs that stabilize the open Hsp90 dimer expose the helix and allow MVB fusion"
- id: PMID:30245208
title: Hsp83/Hsp90 Physically Associates with Insulin Receptor to Promote Neural Stem
Cell Reactivation.
findings:
- statement: Hsp83 is both necessary and sufficient for NSC reactivation by promoting activation of InR pathway
supporting_text: "Hsp83 is both necessary and sufficient for NSC reactivation by promoting the activation of InR pathway in larval brains in the presence of dietary amino acids"
- statement: Both Hsp83 and its co-chaperone Cdc37 physically associate with InR
supporting_text: "Both Hsp83 and its co-chaperone Cdc37 physically associate with InR"
- id: PMID:31907206
title: Drosophila Morgana is an Hsp90-interacting protein with a direct role in
microtubule polymerisation.
findings:
- statement: Morgana co-purifies with Hsp90-R2TP-TTT supercomplex
supporting_text: "Mora localises to mitotic spindles and co-purifies with the Hsp90-R2TP-TTT supercomplex and with additional well-known Hsp90 co-chaperones"
- statement: Mora localises to mitotic spindles
supporting_text: "Drosophila Mora localises to mitotic spindles and co-purifies with the Hsp90-R2TP-TTT supercomplex"
- id: PMID:33176138
title: Modulation of Ago2 Loading by Cyclophilin 40 Endows a Unique Repertoire of
Functional miRNAs during Sperm Maturation in Drosophila.
findings:
- statement: Cyp40 interacts with duplex-incorporating Ago2 through Hsp90 in vitro
supporting_text: "Cyp40 interacts with duplex-incorporating Ago2 through Hsp90 in vitro and selectively promotes the build-up of Ago2-bound miRNAs"
- statement: Identifies RISC-regulatory roles for Hsp90 machinery
supporting_text: "these results identify RISC-regulatory roles for Hsp90 machinery"
- id: PMID:21186352
title: Drosophila Piwi functions in Hsp90-mediated suppression of phenotypic variation.
findings:
- statement: Hsp83 interacts with Hop and Piwi
supporting_text: "we isolated a protein complex composed of Hsp90, Piwi and Hop, the Hsp70/Hsp90 organizing protein homolog"
- statement: Required for piRNA biogenesis and transposon suppression
supporting_text: "Hsp90 and Hop regulate the piRNA pathway through Piwi to mediate canalization"
- statement: Mediates developmental robustness via epigenetic silencing
supporting_text: "epigenetic silencing of the expression of existing genetic variants and the suppression of transposon-induced new genetic variation as two major mechanisms underlying piRNA pathway-mediated canalization"
- id: file:DROME/Hsp83/Hsp83-deep-research-falcon.md
title: Falcon deep research report on Hsp83 (Drosophila melanogaster)
findings:
- statement: |-
Hsp83 is the cytosolic Hsp90 ATP-dependent chaperone that operates via an
ATP-driven conformational cycle to promote late-stage folding, maturation,
stabilization, and activation of diverse signaling/developmental client proteins.
supporting_text: |-
Hsp83 is best understood as the **cytosolic Hsp90 ATP-dependent chaperone** that promotes **late-stage folding/maturation, stabilization, and activation** of diverse client proteins, especially those central to signaling and development (prodromou2022advancestowardsunderstanding pages 1-3, oostenhawle2023organismalrolesof pages 1-3). Mechanistically, Hsp90 proteins are **dimeric**, undergoing an **ATP-driven conformational cycle** involving **ATP binding/hydrolysis** and large structural rearrangements that enable remodeling/maturation of clients
reference_section_type: OTHER
- statement: |-
The Hsp83/Hsp90 chaperone cycle is regulated by co-chaperones including Cdc37
(kinase client recruitment), p23, Aha1, and TPR-domain proteins such as Hop
that bind the conserved C-terminal MEEVD motif of cytosolic Hsp90.
supporting_text: |-
Key named regulators include **Cdc37** (kinase client recruitment), **p23**, and **Aha1**, as well as **TPR-domain** proteins such as **Hop** that bind the conserved **MEEVD** C-terminal motif typical of cytosolic Hsp90s
reference_section_type: OTHER
- statement: |-
Hsp83/Hsp90 is required to activate the ecdysone receptor heterodimer EcR/USP;
a purified Hsp83-Hsc70-Hop-Hip-FKBP52-p23 multichaperone complex reconstitutes
functional EcR/USP DNA binding in vitro, and activation requires ATP hydrolysis
and Mg2+. This is a steroid-hormone-receptor maturation role analogous to
mammalian Hsp90 chaperoning of nuclear receptors.
supporting_text: |-
A key Drosophila primary study demonstrated that the **ecdysone receptor heterodimer EcR/USP requires an Hsp83/Hsp90-containing multichaperone complex for activation**. Purified components—Hsp83 (Hsp90), Hsc70, Hop, Hip, FKBP52, and p23—were sufficient to reconstitute functional EcR/USP DNA binding in vitro, and activation required **ATP hydrolysis and Mg2+**
reference_section_type: OTHER
- statement: |-
Hsp83 facilitates juvenile hormone signaling by promoting nuclear import of the
JH receptor Methoprene-tolerant (Met). Hsp83 physically interacts with Met, JH
enhances this interaction, and Hsp83 loss of function reduces JH-induced Met
nuclear import and JH-responsive Kr-h1 transcription.
supporting_text: |-
In Drosophila juvenile hormone signaling, Hsp83 physically interacts with the JH receptor **Methoprene-tolerant (Met)**, and JH increases the Met–Hsp83 interaction. Hsp83 loss-of-function attenuates JH binding and reduces JH-induced **nuclear import** of Met, decreasing JH-responsive transcription of **Kr-h1**
reference_section_type: OTHER
- statement: |-
Hsp83 promotes timely terminal cell cycle exit during pupal development. Partial
loss of function increases E2F-dependent transcription and causes ectopic S phase
and mitosis after neighboring wild-type cells have become postmitotic, consistent
with chaperoning of cell-cycle/APC-C regulators.
supporting_text: |-
In Hsp83(6-55) mutant clones, E2F-dependent transcription increases and cells show ectopic S phase and mitosis after neighboring cells have exited the cell cycle
reference_section_type: OTHER
- statement: |-
Hsp83 is essential and required throughout spermatogenesis, particularly for
microtubule-dependent steps; biochemical data indicate an indirect role via
stabilization/maturation of microtubule effectors and signaling components
rather than direct tubulin polymerization.
supporting_text: |-
Phenotypically, **all stages of spermatogenesis involving microtubule function are affected**, from early mitotic divisions through sperm maturation and individualization/motility (yue1999geneticanalysisof pages 1-2). In a viable male-sterile allele (*scratch*), Hsp83/Hsp90 protein was reduced by **~3-fold** (ovaries, testes, male bodies), correlating with fully penetrant male sterility (yue1999geneticanalysisof pages 5-7). Biochemically, only a small fraction of Hsp83 co-purifies with taxol-stabilized microtubule proteins, and Hsp83 does not remain bound through repeated microtubule assembly/disassembly, supporting an **indirect role** via stabilization/maturation of microtubule effectors and/or signaling components rather than direct tubulin polymerization
reference_section_type: OTHER
- statement: |-
Hsp83 contributes to female fertility and oogenesis, interacting with the
translational regulator Cup and affecting maternal mRNA handling, including
nanos mRNA localization relevant to anterior-posterior embryonic patterning.
supporting_text: |-
Hsp83 also contributes to female fertility, oogenesis, and maternal RNA regulation. It interacts with the translational regulator Cup during oogenesis, localizes maternal transcripts, and affects *nanos* mRNA localization, linking Hsp83 to anterior–posterior patterning in embryos.
reference_section_type: OTHER
- statement: |-
Hsp83/Hsp90 buffers cryptic genetic variation and supports fitness in natural
fly populations; reduced expression lowers fecundity and longevity and weakens
robustness, especially under inbreeding and mild heat stress, supporting the
classic capacitor (canalization) model in a population context.
supporting_text: |-
Hsp83/Hsp90 buffers cryptic genetic variation and supports fitness in natural fly populations. Reduced Hsp83 expression from naturally occurring insertions lowers fecundity and longevity and weakens robustness, especially under inbreeding and mild heat stress, supporting the classic
reference_section_type: OTHER
- statement: |-
Hsp83 can act as a constitutive proteostasis factor whose transcription is not
necessarily induced in all proteotoxic stress states; in a 2023 larval fat body
study Hsp83 (and Gp93) did not change despite induction of Hsp70/Hsp40-class
stress genes, qualifying the strength of an inducible heat-response annotation.
supporting_text: |-
A 2023 Nature Communications study in Drosophila larval fat body found that **Nacα reduction activates a heat shock response**, with upregulation of Hsp70/Hsp40 class genes and oxidative stress markers, but **Hsp83 (and another Hsp90-family gene Gp93) did not change** in that context (qRT-PCR, n=4 batches)
reference_section_type: OTHER
core_functions:
- description: The primary molecular function of Hsp83. As a member of the HSP90 family,
Hsp83 binds near-native client proteins and uses its intrinsic ATPase activity
to drive conformational changes that promote client maturation, activation, and
stabilization. Known clients include kinases (insulin receptor, LKB1), transcription
factors, Argonaute, APC/C components (Cdh1/Fzr), and centrosomal proteins.
molecular_function:
id: GO:0140662
label: ATP-dependent protein folding chaperone
directly_involved_in:
- id: GO:0006457
label: protein folding
- id: GO:0050821
label: protein stabilization
- id: GO:0034605
label: cellular response to heat
- id: GO:0070922
label: RISC complex assembly
locations:
- id: GO:0005829
label: cytosol
in_complex:
id: GO:0101031
label: protein folding chaperone complex
supported_by:
- reference_id: PMID:29775584
supporting_text: the Hsp90 system is required to extend the dwell time of Ago2
in the open state
- reference_id: PMID:23509070
supporting_text: Sicily preprotein interacts with cytosolic Hsp90 to chaperone
the CI subunit, ND42, before mitochondrial import