Deep Research
Bioreason Pro
(rdgBbeta-deep-research-bioreason-sft.md)
Bioreason Pro
(rdgBbeta-deep-research-bioreason-sft.md)Thinking Trace
The architecture begins with IPR023393 (START-like domain superfamily, residues 2–263), a fold that creates a deep hydrophobic cavity for lipid sequestration and exchange. Nested within this scaffold is IPR055261 (Phosphatidylinositol transfer protein, N-terminal domain, residues 2–244), which defines the specific PITP topology that opens and closes a lipid-binding pocket. Multiple family-level signatures of the phosphatidylinositol transfer protein lineage are embedded across the sequence—IPR001666 (PITP family) at residues 3–254, 17–36, 84–104, 110–125, 193–208, and 213–232—marking conserved motifs that line the cavity and the gating elements that control lipid capture and release. The ordered nesting of a PITP-specific N-terminal domain inside a START-like superfamily fold causes high-affinity, monomeric lipid transfer between membranes without vesicle budding or fusion.
This PITP fold enforces a molecular function centered on selective lipid exchange. The conserved cavity and gating residues accommodate phosphatidylinositol and phosphatidylcholine, enabling GO:0008526 phosphatidylinositol transfer activity and GO:0008525 phosphatidylcholine transporter activity. Because the cavity is optimized for neutral and anionic glycerophospholipids, the mechanism is a cycle of membrane docking, lipid extraction into the hydrophobic pocket, transit through the cytosol, and deposition at a target membrane.
Lipid transfer of PI and PC has direct consequences for membrane composition and signaling during cell division. By delivering PI to specific membrane subdomains, the protein sustains phosphoinositide synthesis and curvature-sensitive recruitment of cytokinetic machinery; by shuttling PC, it maintains bilayer integrity and supports membrane expansion at the ingressing furrow. These activities drive the processes formalized as GO:0007110 meiosis I cytokinesis and GO:0007111 meiosis II cytokinesis, and extend to GO:0007112 male meiosis cytokinesis and GO:0048137 spermatocyte division, where precise membrane remodeling is essential. The same lipid supply and signaling flux coordinate with the actomyosin apparatus, promoting GO:0000916 actomyosin contractile ring contraction and GO:0036090 cleavage furrow ingression by ensuring a competent, lipid-rich cortex that can withstand contractile forces and by positioning PI pools that recruit small GTPases and effectors. Proper PI availability also influences microtubule–cortex communication and membrane–microtubule interfaces, contributing to GO:0000212 meiotic spindle organization.
The domain architecture lacks transmembrane segments and secretion signals, indicating a soluble protein that cycles on and off membranes. Its action requires proximity to sites of intense membrane turnover and signaling. Thus, it operates in the cytoplasm and transiently associates with the GO:0005794 Golgi apparatus to acquire and dispatch lipids, with the GO:0032154 cleavage furrow where cytokinesis occurs, and with the GO:0031965 nuclear membrane during meiotic stages when nuclear envelope dynamics and membrane continuity are critical. This distribution is consistent with a soluble lipid-transfer factor that concentrates at division sites and organelles engaged in lipid trafficking.
Mechanistically, the protein likely docks to donor membranes via basic surface patches and amphipathic elements, captures PI or PC into its START-like cavity, and releases the lipid at target membranes to sustain phosphoinositide synthesis and bilayer homeostasis. At the cleavage furrow and nuclear envelope, it plausibly collaborates with small GTPases (e.g., ARF and RAB family members) and phosphoinositide kinases to position PI pools that recruit actomyosin regulators and membrane-sculpting factors. It may also engage cytokinetic scaffolds at the furrow to synchronize lipid supply with contractile ring constriction and with spindle–cortex communication during meiosis.
## Functional Summary
A soluble lipid-transfer factor in fruit fly that uses a deep hydrophobic cavity to extract and shuttle phosphatidylinositol and phosphatidylcholine between membranes. By supplying these lipids to division sites and organelles, it sustains phosphoinositide signaling and bilayer integrity required for spindle organization, contractile ring function, and membrane expansion during meiotic and spermatocyte cytokinesis. It operates in the cytoplasm and transiently associates with the Golgi, the cleavage furrow, and the nuclear envelope to coordinate lipid flux with cell division mechanics.
## UniProt Summary
Catalyzes the transfer of phosphatidylinositol and phosphatidylcholine between membranes.
## InterPro Domains
- IPR023393: START-like domain superfamily (homologous_superfamily) [2-263]
- IPR055261: Phosphatidylinositol transfer protein, N-terminal (domain) [2-244]
- IPR001666: Phosphatidylinositol transfer protein (family) [3-254]
- IPR001666: Phosphatidylinositol transfer protein (family) [17-36]
- IPR001666: Phosphatidylinositol transfer protein (family) [84-104]
- IPR001666: Phosphatidylinositol transfer protein (family) [110-125]
- IPR001666: Phosphatidylinositol transfer protein (family) [193-208]
- IPR001666: Phosphatidylinositol transfer protein (family) [213-232]
## GO Term Predictions
### Molecular Function
### Biological Process
### Cellular Component
Falcon
(rdgBbeta-deep-research-falcon.md)
Falcon
(rdgBbeta-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: Functional annotation of rdgBbeta (CG17818; UniProt Q9U9P7) in Drosophila melanogaster
1) Target verification and gene/protein identity (critical disambiguation)
The UniProt target Q9U9P7 is annotated as Drosophila melanogaster rdgBbeta (CG17818), described as a cytoplasmic phosphatidylinositol transfer protein (PITP-family) and “retinal degeneration B homolog beta (RdgBβ)”. The PITP-family review by Hsuan & Cockcroft explicitly lists a Drosophila rdgBβ entry (and discusses the rdgBβ subfamily as distinct from the multi-domain rdgBα/Nir proteins), supporting that the symbol rdgBbeta refers to a soluble PITP-domain protein rather than the photoreceptor degeneration gene rdgB (rdgBα) commonly studied in Drosophila phototransduction. (hsuan2001thepitpfamily pages 2-3)
A key species caveat from the same review is that human RdgBβ has a short C-terminal extension whereas the Drosophila isoform does not, meaning some mammalian regulatory motifs in the C-terminus may not be conserved in Drosophila and must not be transferred uncritically. (hsuan2001thepitpfamily pages 2-3)
2) Key concepts and definitions (current understanding)
2.1 Phosphatidylinositol transfer proteins (PITPs)
PITPs are lipid-binding/transfer proteins that bind a single phospholipid molecule in a hydrophobic cavity and have long been studied for their ability to exchange lipids between membranes in vitro; they are now widely discussed as regulators of phosphoinositide metabolism, signaling, and membrane traffic. (hsuan2001thepitpfamily pages 2-3)
2.2 Class I vs Class II PITPs; where RdgBβ fits
A commonly used functional distinction in the literature is:
- Class I PITPs (e.g., PITPα/β) exchange PI and PC.
- Class II PITPs (RdgB proteins) are emphasized as PI and PA binders/transfer proteins (with minimal PC), and are often discussed in the context of phospholipase C (PLC) signaling and the need to couple PI metabolism with PA flux. (cockcroft2016rdgbαreciprocallytransfers pages 8-11, cockcroft2013potentialrolefor pages 6-8)
RdgBβ is explicitly described as a soluble class IIB PITP that binds other proteins (e.g., 14-3-3, ATRAP) for functional recruitment to membranes—distinct from multi-domain RdgBα proteins that localize to ER–PM contact sites via VAP. (cockcroft2016rdgbαreciprocallytransfers pages 8-11)
3) Primary molecular function: lipid binding/transfer and substrate specificity
3.1 Lipid ligands and inferred biochemical role
Expert review synthesis proposes that RdgBβ-class proteins preferentially bind/transfer phosphatidylinositol (PI) and phosphatidic acid (PA) with very little phosphatidylcholine (PC), and that RdgBβ can transfer PA robustly, a property not typical of class I PITPs. (cockcroft2013potentialrolefor pages 6-8, cockcroft2016rdgbαreciprocallytransfers pages 8-11)
Mechanistically, this PI/PA specialization is interpreted as fitting the lipid-transport demands that accompany PLC activation (consumption of PI(4,5)P2 at the plasma membrane and generation/turnover of DAG and PA), where exchange of PI and PA between compartments can support restoration of signaling lipids. (cockcroft2013potentialrolefor pages 6-8, cockcroft2016rdgbαreciprocallytransfers pages 8-11)
3.2 Quantitative biochemical evidence (mostly from mammalian PITPNC1/RdgBβ)
Direct quantitative assays in a Biochemical Journal primary paper on RdgBβ/PITPNC1 report very weak PI-transfer activity compared with PITPα: detectable activity required ~10–100 μg/mL RdgBβ vs ~200–500 ng/mL PITPα (≈100–250× higher protein concentration required), and endogenous/overexpressed RdgBβ in cell/tissue fractions showed no measurable PI-transfer activity under those conditions. (garner2011thephosphatidylinositoltransfer pages 8-10, garner2011thephosphatidylinositoltransfer pages 11-13, garner2011thephosphatidylinositoltransfer pages 1-4, garner2011thephosphatidylinositoltransfer pages 10-11)
These results are commonly interpreted to mean that RdgBβ’s function likely depends on context-specific membrane recruitment and/or regulation of the lipid-binding cavity, rather than acting as a high-flux bulk cytosolic PI-transfer factor. (garner2011thephosphatidylinositoltransfer pages 1-4, cockcroft2016rdgbαreciprocallytransfers pages 3-6)
4) Subcellular localization and interaction partners (where it acts in the cell)
4.1 Cytosolic protein with regulated membrane recruitment
RdgBβ/PITPNC1 is described as mainly cytosolic at baseline and can be recruited to membranes upon specific stimuli. In the primary study, treatment with PMA (activating PKC pathways) promoted membrane recruitment and co-localization with the integral membrane protein ATRAP/AGTRAP, which was proposed to serve as a recruitment factor. (garner2011thephosphatidylinositoltransfer pages 10-11, garner2011thephosphatidylinositoltransfer pages 1-4)
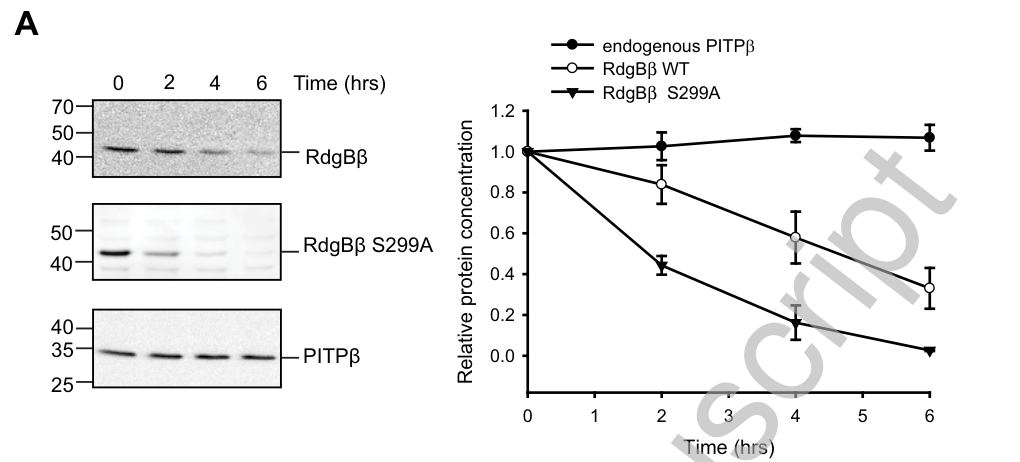
Quantitatively, the same study reports approximately 8-fold enrichment of wild-type RdgBβ in membranes after PMA, and an even larger (~14-fold) membrane increase for a mutant defective in 14-3-3 binding, supporting a model where 14-3-3 binding restrains membrane translocation. (garner2011thephosphatidylinositoltransfer pages 11-13, garner2011thephosphatidylinositoltransfer media 1c076168)
4.2 Protein–protein interactions and regulation
Two key binding partners repeatedly emphasized in authoritative sources are:
- 14-3-3 adaptor proteins, binding via phosphorylated motifs in an unstructured C-terminus (shown for mammalian PITPNC1/RdgBβ). (garner2011thephosphatidylinositoltransfer pages 8-10, garner2011thephosphatidylinositoltransfer pages 11-13)
- ATRAP/AGTRAP, binding via the PITP domain and promoting stimulus-dependent membrane recruitment (PMA/PKC-dependent). (garner2011thephosphatidylinositoltransfer pages 1-4, garner2011thephosphatidylinositoltransfer pages 10-11)
These interactions support an expert model in which RdgBβ executes function after protein-mediated recruitment to specific membrane regions, rather than via stable transmembrane anchoring. (cockcroft2016rdgbαreciprocallytransfers pages 8-11, garner2011thephosphatidylinositoltransfer pages 1-4)
5) Pathways and biological processes likely involving Drosophila rdgBbeta
5.1 Relationship to PLC/phosphoinositide signaling frameworks
The PITP/RdgB literature frames RdgB-family PITPs as central to phosphoinositide-cycle lipid flux that is topologically split between membranes (e.g., ER vs plasma membrane), requiring lipid transfer steps to maintain signaling competence during/after PLC activation. While this framework is most directly developed for multi-domain RdgBα at ER–PM contact sites, Class II PITPs (including RdgBβ-class) are explicitly described as PI/PA transfer proteins, and thus conceptually positioned to couple PI metabolism with PA handling during signaling. (cockcroft2016rdgbαreciprocallytransfers pages 8-11, cockcroft2013potentialrolefor pages 6-8)
5.2 Drosophila-specific functional evidence: current limitations
Within the retrieved corpus, direct primary mechanistic studies specific to Drosophila melanogaster rdgBbeta/CG17818/Q9U9P7 were limited, and much of the quantitative mechanistic evidence is from mammalian PITPNC1/RdgBβ. The strongest Drosophila-specific statement available is classification and structural distinction: Drosophila has a rdgBβ PITP-domain protein that lacks the human C-terminal extension, implying potential divergence in regulatory motifs. (hsuan2001thepitpfamily pages 2-3)
Accordingly, for functional annotation of Drosophila rdgBbeta, the most defensible current interpretation from this evidence set is:
- rdgBbeta encodes a soluble Class II (Class IIB) PITP-domain protein. (hsuan2001thepitpfamily pages 2-3, cockcroft2016rdgbαreciprocallytransfers pages 8-11)
- Its primary biochemical capability is most plausibly PI/PA binding/transfer (family inference), likely acting locally at membranes upon recruitment rather than as a bulk cytosolic transfer activity. (cockcroft2013potentialrolefor pages 6-8, cockcroft2016rdgbαreciprocallytransfers pages 3-6)
- Any proposed 14-3-3/ATRAP regulatory module should be treated as hypothesis-by-homology unless conserved motifs are verified in the Drosophila protein sequence, because of the explicit Drosophila/mammal C-terminal difference. (hsuan2001thepitpfamily pages 2-3)
6) Recent developments (prioritizing 2023–2024) and current research directions
The most directly relevant recent paper captured here for the broader RdgB/PITP field in Drosophila is a 2024 genetic screen focused on the multi-domain RDGB (RdgBα) lipid transfer protein at membrane contact sites, motivated by neurodegeneration/retinal degeneration phenotypes upon RDGB loss. Although it is not rdgBbeta-specific, it reflects active 2024 research emphasis on lipid-transfer mechanisms at contact sites and on identifying regulatory networks that modulate PITP-family lipid transfer function in vivo. (Life Science Alliance; publication date Mar 2024; https://doi.org/10.26508/lsa.202302525) (cockcroft2016rdgbαreciprocallytransfers pages 8-11)
For rdgBbeta specifically, the most recent information in the retrieved corpus remains primarily review-level synthesis and mammalian mechanistic studies rather than 2023–2024 Drosophila CG17818-focused primary work. (hsuan2001thepitpfamily pages 2-3, cockcroft2016rdgbαreciprocallytransfers pages 8-11)
7) Applications and real-world implementations
7.1 Functional genomics and membrane-contact-site biology
RdgB-family PITPs (including Class II PITPs) are used conceptually and experimentally as paradigms for how lipid transfer supports signaling homeostasis across organelles, especially in models of PLC-driven phosphoinositide turnover and membrane contact site function. This is a widely adopted framework in cell biology and signaling research. (cockcroft2016rdgbαreciprocallytransfers pages 8-11, cockcroft2013potentialrolefor pages 6-8)
7.2 Translational relevance via mammalian PITPNC1 (contextual, not Drosophila-specific)
While outside the Drosophila gene itself, authoritative expert sources discuss mammalian PITPNC1/RdgBβ in the context of regulated secretion and disease models (e.g., cancer secretory phenotypes), providing a translational motivation for mechanistic studies of RdgBβ-class proteins. (cockcroft2012thediversefunctions pages 206-209)
8) Key quantitative statistics/data (from primary evidence)
Quantitative findings most directly supporting mechanistic annotation (measured for mammalian PITPNC1/RdgBβ) include:
- Protein stability: ~4 h half-life for WT vs ~2 h for a 14-3-3-binding-defective mutant; degradation is proteasome-dependent and associated with ubiquitination. (garner2011thephosphatidylinositoltransfer pages 8-10, garner2011thephosphatidylinositoltransfer pages 1-4, garner2011thephosphatidylinositoltransfer media 1c076168)
- Stimulus-dependent membrane recruitment: PMA induces ~8× membrane enrichment for WT and ~14× for a 14-3-3-binding-defective mutant. (garner2011thephosphatidylinositoltransfer pages 11-13, garner2011thephosphatidylinositoltransfer media 1c076168)
- Relative PI-transfer assay performance: PITPα active at ~200–500 ng/mL, whereas RdgBβ required ~10–100 μg/mL for detectable activity (≈100–250× weaker by concentration). (garner2011thephosphatidylinositoltransfer pages 8-10, garner2011thephosphatidylinositoltransfer pages 11-13, garner2011thephosphatidylinositoltransfer pages 10-11)
Evidence summary table
| Claim (short) | Evidence/Details | System/Organism | Source (paper + year + DOI URL) | Context ID(s) |
|---|---|---|---|---|
| Family classification: soluble Class IIB PITP | RdgBβ is described as a soluble class IIB phosphatidylinositol transfer protein within the RdgB/PITP family, distinct from multidomain RdgBα/Class IIA proteins; Drosophila has an rdgBβ subfamily member and assignment is based on PITP-domain similarity. | Family-level annotation; Drosophila + mammalian comparison | Hsuan & Cockcroft 2001, Genome Biology, https://doi.org/10.1186/gb-2001-2-9-reviews3011; Cockcroft et al. 2016, Biochem Soc Trans, https://doi.org/10.1042/bst20150228 | (hsuan2001thepitpfamily pages 2-3, cockcroft2016rdgbαreciprocallytransfers pages 8-11) |
| Lipid ligands/specificity: PI/PA vs PC | Class II/RdgBβ proteins are reported to bind and transfer PI and PA, with very little PC; under PLC/PLD-stimulated conditions RdgBβ shifts toward greater PA binding, and PA transfer is described as robust relative to Class I PITPs. | Conserved RdgBβ/PITPNC1 family conclusion | Cockcroft & Garner 2013, Adv Biol Regul, https://doi.org/10.1016/j.jbior.2013.07.007; Cockcroft et al. 2016, Biochem Soc Trans, https://doi.org/10.1042/bst20150228 | (cockcroft2013potentialrolefor pages 6-8, cockcroft2016rdgbαreciprocallytransfers pages 8-11) |
| 14-3-3 docking sites and stability control | Two phosphorylated serines in the disordered C-terminus (Ser274 and Ser299) form the 14-3-3 docking module; mutation of either site abolishes 14-3-3 binding. 14-3-3 shields nearby PEST sequences and stabilizes RdgBβ. | Human PITPNC1/RdgBβ experimental system; family-relevant inference | Garner et al. 2011, Biochem J, https://doi.org/10.1042/bj20110649 | (garner2011thephosphatidylinositoltransfer pages 8-10, garner2011thephosphatidylinositoltransfer pages 11-13, garner2011thephosphatidylinositoltransfer pages 13-14, garner2011thephosphatidylinositoltransfer media 1c076168) |
| Proteasome turnover / half-life | RdgBβ is ubiquitinated and degraded via the proteasome. Wild-type protein has an approximate 4 h half-life, whereas a 14-3-3-binding-defective mutant is reduced to about 2 h, indicating 14-3-3 binding protects against turnover. | Human PITPNC1/RdgBβ in cells | Garner et al. 2011, Biochem J, https://doi.org/10.1042/bj20110649 | (garner2011thephosphatidylinositoltransfer pages 8-10, garner2011thephosphatidylinositoltransfer pages 11-13, garner2011thephosphatidylinositoltransfer pages 1-4, garner2011thephosphatidylinositoltransfer media 1c076168) |
| ATRAP interaction and PKC/PMA dependence | The PITP domain of RdgBβ interacts with the integral membrane protein ATRAP/AGTRAP at a site distinct from the 14-3-3 site. Interaction and membrane recruitment increase after PMA treatment and are reduced by the PKC inhibitor BIM, supporting PKC-dependent regulation. | Human PITPNC1/RdgBβ; mechanistic family inference | Garner et al. 2011, Biochem J, https://doi.org/10.1042/bj20110649; Cockcroft & Garner 2013, Adv Biol Regul, https://doi.org/10.1016/j.jbior.2013.07.007 | (garner2011thephosphatidylinositoltransfer pages 8-10, garner2011thephosphatidylinositoltransfer pages 4-5, garner2011thephosphatidylinositoltransfer pages 10-11, cockcroft2013potentialrolefor pages 6-8, garner2011thephosphatidylinositoltransfer media 1c076168) |
| Membrane recruitment fold change | Upon PMA treatment, wild-type RdgBβ shows about an 8-fold increase in the membrane fraction, whereas a 14-3-3-binding-deficient mutant shows about a 14-fold increase, indicating 14-3-3 restrains membrane translocation. | Human PITPNC1/RdgBβ in COS-7 cells | Garner et al. 2011, Biochem J, https://doi.org/10.1042/bj20110649 | (garner2011thephosphatidylinositoltransfer pages 11-13, garner2011thephosphatidylinositoltransfer media 1c076168) |
| Very low/undetectable in vitro PI transfer | RdgBβ is far weaker than canonical PITPα in PI transfer assays. Reported thresholds: PITPα active at ~200–500 ng/ml, while RdgBβ requires ~10–100 μg/ml for detectable/significant activity (roughly 100–250-fold less active by concentration). Endogenous/overexpressed RdgBβ fractions typically show no detectable PI transfer under tested conditions. | Human PITPNC1/RdgBβ; rat heart cytosol | Garner et al. 2011, Biochem J, https://doi.org/10.1042/bj20110649; Cockcroft et al. 2016, Biochem Soc Trans, https://doi.org/10.1042/bst20150228 | (garner2011thephosphatidylinositoltransfer pages 8-10, garner2011thephosphatidylinositoltransfer pages 11-13, garner2011thephosphatidylinositoltransfer pages 1-4, garner2011thephosphatidylinositoltransfer pages 10-11, cockcroft2016rdgbαreciprocallytransfers pages 3-6, garner2011thephosphatidylinositoltransfer media 1c076168) |
| Tissue enrichment / localization clue | RdgBβ is reported as enriched in heart (and also brain in review discussion); cytosolic activity peaks attributable to RdgBβ were not observed, leading to the proposal that it acts locally at membranes/contact sites after recruitment rather than as a bulk soluble transfer activity. | Rat heart / review interpretation | Cockcroft & Garner 2013, Adv Biol Regul, https://doi.org/10.1016/j.jbior.2013.07.007; Cockcroft et al. 2016, Biochem Soc Trans, https://doi.org/10.1042/bst20150228 | (cockcroft2013potentialrolefor pages 8-9, cockcroft2016rdgbαreciprocallytransfers pages 3-6) |
| Distinction from RdgBα | RdgBβ is a small soluble PITP with a short disordered tail and protein-partner-mediated membrane recruitment, whereas RdgBα is a multidomain protein with FFAT/DDHD/LSN2-related modules that localizes to ER–PM contact sites via VAP. This distinction is important to avoid confusing Drosophila rdgBbeta with rdgB/rdgBα phototransduction literature. | Family comparison | Hsuan & Cockcroft 2001, Genome Biology, https://doi.org/10.1186/gb-2001-2-9-reviews3011; Cockcroft et al. 2016, Biochem Soc Trans, https://doi.org/10.1042/bst20150228 | (hsuan2001thepitpfamily pages 2-3, cockcroft2016rdgbαreciprocallytransfers pages 8-11) |
| Drosophila-specific C-terminus difference | The Drosophila rdgBβ isoform is explicitly noted to lack the short carboxy-terminal extension present in human RdgBβ. Thus, C-terminal regulatory findings from mammalian PITPNC1/RdgBβ (e.g., Ser274/Ser299 14-3-3 docking) should be transferred to Drosophila CG17818/Q9U9P7 cautiously. | Drosophila-specific annotation | Hsuan & Cockcroft 2001, Genome Biology, https://doi.org/10.1186/gb-2001-2-9-reviews3011 | (hsuan2001thepitpfamily pages 2-3) |
Table: This table compiles experimentally supported properties of the RdgBβ/PITPNC1 family most relevant to annotating Drosophila rdgBbeta (CG17818; UniProt Q9U9P7). It highlights where evidence is direct versus family-based inference and flags the important Drosophila-specific C-terminal difference.
Figures (primary evidence snapshots)
Cropped figure panels from the primary RdgBβ/PITPNC1 study document (i) the 14-3-3 phosphosite module and binding assays, (ii) protein half-life differences, (iii) low PI-transfer activity relative to PITPα, and (iv) PMA/ATRAP-associated membrane recruitment. (garner2011thephosphatidylinositoltransfer media 1c076168, garner2011thephosphatidylinositoltransfer media f519b69c, garner2011thephosphatidylinositoltransfer media 13836492, garner2011thephosphatidylinositoltransfer media 54c46ab2, garner2011thephosphatidylinositoltransfer media caf2756a, garner2011thephosphatidylinositoltransfer media e2864aea)
Conclusion (functional annotation best supported by current evidence set)
Within the available and retrieved literature, Drosophila rdgBbeta (CG17818; UniProt Q9U9P7) is best annotated as a soluble Class IIB PITP-domain protein in the PtdIns transfer protein family, distinct from the multi-domain RdgBα phototransduction protein. (hsuan2001thepitpfamily pages 2-3, cockcroft2016rdgbαreciprocallytransfers pages 8-11)
Its primary molecular function is most plausibly binding/transfer of PI and PA (not PC) and participation in lipid homeostasis coupled to phosphoinositide signaling, likely acting locally at membranes upon recruitment (family-level inference), but direct Drosophila CG17818-specific mechanistic evidence remains limited in the retrieved corpus; therefore, mammalian PITPNC1/RdgBβ regulatory details (e.g., C-terminal 14-3-3 module) should be applied to Drosophila only after conservation is confirmed, especially given the reported Drosophila-vs-human C-terminal difference. (cockcroft2013potentialrolefor pages 6-8, cockcroft2016rdgbαreciprocallytransfers pages 3-6, hsuan2001thepitpfamily pages 2-3)
References
-
(hsuan2001thepitpfamily pages 2-3): Justin Hsuan and Shamshad Cockcroft. The pitp family of phosphatidylinositol transfer proteins. Genome Biology, 2:reviews3011.1-reviews3011.8, Aug 2001. URL: https://doi.org/10.1186/gb-2001-2-9-reviews3011, doi:10.1186/gb-2001-2-9-reviews3011. This article has 93 citations and is from a highest quality peer-reviewed journal.
-
(cockcroft2016rdgbαreciprocallytransfers pages 8-11): Shamshad Cockcroft, Kathryn Garner, Shweta Yadav, Evelyn Gomez-Espinoza, and Padinjat Raghu. Rdgbα reciprocally transfers pa and pi at er-pm contact sites to maintain pi(4,5)p2 homoeostasis during phospholipase c signalling in drosophila photoreceptors. Biochemical Society transactions, 44 1:286-92, Feb 2016. URL: https://doi.org/10.1042/bst20150228, doi:10.1042/bst20150228. This article has 38 citations and is from a peer-reviewed journal.
-
(cockcroft2013potentialrolefor pages 6-8): Shamshad Cockcroft and Kathryn Garner. Potential role for phosphatidylinositol transfer protein (pitp) family in lipid transfer during phospholipase c signalling. Advances in biological regulation, 53 3:280-91, Sep 2013. URL: https://doi.org/10.1016/j.jbior.2013.07.007, doi:10.1016/j.jbior.2013.07.007. This article has 44 citations and is from a peer-reviewed journal.
-
(garner2011thephosphatidylinositoltransfer pages 8-10): Kathryn Garner, Michelle Li, Natalie Ugwuanya, and Shamshad Cockcroft. The phosphatidylinositol transfer protein rdgbβ binds 14-3-3 via its unstructured c-terminus, whereas its lipid-binding domain interacts with the integral membrane protein atrap (angiotensin ii type i receptor-associated protein). The Biochemical journal, 439 1:97-111, Oct 2011. URL: https://doi.org/10.1042/bj20110649, doi:10.1042/bj20110649. This article has 42 citations.
-
(garner2011thephosphatidylinositoltransfer pages 11-13): Kathryn Garner, Michelle Li, Natalie Ugwuanya, and Shamshad Cockcroft. The phosphatidylinositol transfer protein rdgbβ binds 14-3-3 via its unstructured c-terminus, whereas its lipid-binding domain interacts with the integral membrane protein atrap (angiotensin ii type i receptor-associated protein). The Biochemical journal, 439 1:97-111, Oct 2011. URL: https://doi.org/10.1042/bj20110649, doi:10.1042/bj20110649. This article has 42 citations.
-
(garner2011thephosphatidylinositoltransfer pages 1-4): Kathryn Garner, Michelle Li, Natalie Ugwuanya, and Shamshad Cockcroft. The phosphatidylinositol transfer protein rdgbβ binds 14-3-3 via its unstructured c-terminus, whereas its lipid-binding domain interacts with the integral membrane protein atrap (angiotensin ii type i receptor-associated protein). The Biochemical journal, 439 1:97-111, Oct 2011. URL: https://doi.org/10.1042/bj20110649, doi:10.1042/bj20110649. This article has 42 citations.
-
(garner2011thephosphatidylinositoltransfer pages 10-11): Kathryn Garner, Michelle Li, Natalie Ugwuanya, and Shamshad Cockcroft. The phosphatidylinositol transfer protein rdgbβ binds 14-3-3 via its unstructured c-terminus, whereas its lipid-binding domain interacts with the integral membrane protein atrap (angiotensin ii type i receptor-associated protein). The Biochemical journal, 439 1:97-111, Oct 2011. URL: https://doi.org/10.1042/bj20110649, doi:10.1042/bj20110649. This article has 42 citations.
-
(cockcroft2016rdgbαreciprocallytransfers pages 3-6): Shamshad Cockcroft, Kathryn Garner, Shweta Yadav, Evelyn Gomez-Espinoza, and Padinjat Raghu. Rdgbα reciprocally transfers pa and pi at er-pm contact sites to maintain pi(4,5)p2 homoeostasis during phospholipase c signalling in drosophila photoreceptors. Biochemical Society transactions, 44 1:286-92, Feb 2016. URL: https://doi.org/10.1042/bst20150228, doi:10.1042/bst20150228. This article has 38 citations and is from a peer-reviewed journal.
-
(garner2011thephosphatidylinositoltransfer media 1c076168): Kathryn Garner, Michelle Li, Natalie Ugwuanya, and Shamshad Cockcroft. The phosphatidylinositol transfer protein rdgbβ binds 14-3-3 via its unstructured c-terminus, whereas its lipid-binding domain interacts with the integral membrane protein atrap (angiotensin ii type i receptor-associated protein). The Biochemical journal, 439 1:97-111, Oct 2011. URL: https://doi.org/10.1042/bj20110649, doi:10.1042/bj20110649. This article has 42 citations.
-
(cockcroft2012thediversefunctions pages 206-209): Shamshad Cockcroft. The diverse functions of phosphatidylinositol transfer proteins. Current topics in microbiology and immunology, 362:185-208, Jan 2012. URL: https://doi.org/10.1007/978-94-007-5025-8_9, doi:10.1007/978-94-007-5025-8_9. This article has 53 citations and is from a peer-reviewed journal.
-
(garner2011thephosphatidylinositoltransfer pages 13-14): Kathryn Garner, Michelle Li, Natalie Ugwuanya, and Shamshad Cockcroft. The phosphatidylinositol transfer protein rdgbβ binds 14-3-3 via its unstructured c-terminus, whereas its lipid-binding domain interacts with the integral membrane protein atrap (angiotensin ii type i receptor-associated protein). The Biochemical journal, 439 1:97-111, Oct 2011. URL: https://doi.org/10.1042/bj20110649, doi:10.1042/bj20110649. This article has 42 citations.
-
(garner2011thephosphatidylinositoltransfer pages 4-5): Kathryn Garner, Michelle Li, Natalie Ugwuanya, and Shamshad Cockcroft. The phosphatidylinositol transfer protein rdgbβ binds 14-3-3 via its unstructured c-terminus, whereas its lipid-binding domain interacts with the integral membrane protein atrap (angiotensin ii type i receptor-associated protein). The Biochemical journal, 439 1:97-111, Oct 2011. URL: https://doi.org/10.1042/bj20110649, doi:10.1042/bj20110649. This article has 42 citations.
-
(cockcroft2013potentialrolefor pages 8-9): Shamshad Cockcroft and Kathryn Garner. Potential role for phosphatidylinositol transfer protein (pitp) family in lipid transfer during phospholipase c signalling. Advances in biological regulation, 53 3:280-91, Sep 2013. URL: https://doi.org/10.1016/j.jbior.2013.07.007, doi:10.1016/j.jbior.2013.07.007. This article has 44 citations and is from a peer-reviewed journal.
-
(garner2011thephosphatidylinositoltransfer media f519b69c): Kathryn Garner, Michelle Li, Natalie Ugwuanya, and Shamshad Cockcroft. The phosphatidylinositol transfer protein rdgbβ binds 14-3-3 via its unstructured c-terminus, whereas its lipid-binding domain interacts with the integral membrane protein atrap (angiotensin ii type i receptor-associated protein). The Biochemical journal, 439 1:97-111, Oct 2011. URL: https://doi.org/10.1042/bj20110649, doi:10.1042/bj20110649. This article has 42 citations.
-
(garner2011thephosphatidylinositoltransfer media 13836492): Kathryn Garner, Michelle Li, Natalie Ugwuanya, and Shamshad Cockcroft. The phosphatidylinositol transfer protein rdgbβ binds 14-3-3 via its unstructured c-terminus, whereas its lipid-binding domain interacts with the integral membrane protein atrap (angiotensin ii type i receptor-associated protein). The Biochemical journal, 439 1:97-111, Oct 2011. URL: https://doi.org/10.1042/bj20110649, doi:10.1042/bj20110649. This article has 42 citations.
-
(garner2011thephosphatidylinositoltransfer media 54c46ab2): Kathryn Garner, Michelle Li, Natalie Ugwuanya, and Shamshad Cockcroft. The phosphatidylinositol transfer protein rdgbβ binds 14-3-3 via its unstructured c-terminus, whereas its lipid-binding domain interacts with the integral membrane protein atrap (angiotensin ii type i receptor-associated protein). The Biochemical journal, 439 1:97-111, Oct 2011. URL: https://doi.org/10.1042/bj20110649, doi:10.1042/bj20110649. This article has 42 citations.
-
(garner2011thephosphatidylinositoltransfer media caf2756a): Kathryn Garner, Michelle Li, Natalie Ugwuanya, and Shamshad Cockcroft. The phosphatidylinositol transfer protein rdgbβ binds 14-3-3 via its unstructured c-terminus, whereas its lipid-binding domain interacts with the integral membrane protein atrap (angiotensin ii type i receptor-associated protein). The Biochemical journal, 439 1:97-111, Oct 2011. URL: https://doi.org/10.1042/bj20110649, doi:10.1042/bj20110649. This article has 42 citations.
-
(garner2011thephosphatidylinositoltransfer media e2864aea): Kathryn Garner, Michelle Li, Natalie Ugwuanya, and Shamshad Cockcroft. The phosphatidylinositol transfer protein rdgbβ binds 14-3-3 via its unstructured c-terminus, whereas its lipid-binding domain interacts with the integral membrane protein atrap (angiotensin ii type i receptor-associated protein). The Biochemical journal, 439 1:97-111, Oct 2011. URL: https://doi.org/10.1042/bj20110649, doi:10.1042/bj20110649. This article has 42 citations.
Artifacts
Citations
- hsuan2001thepitpfamily pages 2-3
- cockcroft2012thediversefunctions pages 206-209
- cockcroft2013potentialrolefor pages 6-8
- garner2011thephosphatidylinositoltransfer pages 8-10
- garner2011thephosphatidylinositoltransfer pages 11-13
- garner2011thephosphatidylinositoltransfer pages 1-4
- garner2011thephosphatidylinositoltransfer pages 10-11
- garner2011thephosphatidylinositoltransfer pages 13-14
- garner2011thephosphatidylinositoltransfer pages 4-5
- cockcroft2013potentialrolefor pages 8-9
- https://doi.org/10.26508/lsa.202302525
- https://doi.org/10.1186/gb-2001-2-9-reviews3011;
- https://doi.org/10.1042/bst20150228
- https://doi.org/10.1016/j.jbior.2013.07.007;
- https://doi.org/10.1042/bj20110649
- https://doi.org/10.1042/bj20110649;
- https://doi.org/10.1016/j.jbior.2013.07.007
- https://doi.org/10.1186/gb-2001-2-9-reviews3011
- https://doi.org/10.1186/gb-2001-2-9-reviews3011,
- https://doi.org/10.1042/bst20150228,
- https://doi.org/10.1016/j.jbior.2013.07.007,
- https://doi.org/10.1042/bj20110649,
- https://doi.org/10.1007/978-94-007-5025-8_9,