Deep Research
Falcon
(mcr-1-hypotheses/function-hypothesis-go-0016776/falcon.md)
Falcon
(mcr-1-hypotheses/function-hypothesis-go-0016776/falcon.md)AIGR TreeGrafter Function-Inference Stress Test
You are evaluating one focused gene-function hypothesis for AI Gene Review. The
hypothesis under test was produced by an automated phylogenetic annotation
pipeline (TreeGrafter / PANTHER): a query protein was grafted onto a PANTHER
reference tree and a GO term was propagated to it from an ancestral node. Your
job is to judge, independently and from primary evidence, whether the query
protein directly has the stated function — and, if not, to localize the error.
This is not a general gene overview. Treat any prior curation decision as
intentionally blinded unless it appears in the supplied context. Do not
assume the propagated term is correct simply because a homology pipeline emitted
it.
Target Gene
- Organism code: ECOLX
- Taxon: Escherichia coli (NCBITaxon:562)
- Gene directory: mcr-1
- Gene symbol: mcr-1
- UniProt accession: A0A0R6L508
Focus
- Focus type: function_assignment
- Hypothesis slug: function-hypothesis-go-0016776
- Source file: genes/ECOLX/mcr-1/mcr-1-ai-review.yaml
- Source selector: existing_annotations[5].function_hypothesis
Seed Hypothesis (propagated by TreeGrafter/PANTHER)
mcr-1 has phosphotransferase activity, phosphate group as acceptor (GO:0016776).
Term and Decision Context
- Term: phosphotransferase activity, phosphate group as acceptor (GO:0016776)
- Evidence type: IEA
- Original reference: GO_REF:0000118
Reference Context
- GO_REF:0000118
- PMID:29079699
Source Context YAML
term:
id: GO:0016776
label: phosphotransferase activity, phosphate group as acceptor
evidence_type: IEA
original_reference_id: GO_REF:0000118
Research Objective
Decide whether mcr-1 directly has the stated function. Automated
phylogenetic propagation fails in three characteristic ways; your report must
actively test for each, because they cannot be detected by the graft alone:
- Granularity / family-vs-subfamily. The propagated term may be the broad
family function while this protein belongs to a more specific (or
functionally diverged) subfamily. Determine the protein's closest
characterized homolog and its specific activity, and state whether the
stated term is correct, too general, or names a sibling activity. (Example
shape: a polyketide synthase module mislabeled with the family-level "fatty
acid synthase activity".) - Pseudo-enzyme / loss of activity. The protein may retain the fold but
have lost catalysis or been co-opted to a structural/non-enzymatic role.
Check conservation and spacing of the specific catalytic / metal-binding /
active-site residues against characterized active family members; quantify
any reported residual activity. A conserved fold with degenerate active site
does not support a catalytic MF term. - Within-superfamily mis-placement. The protein may have been grafted onto
a structurally related but functionally distinct neighboring subfamily of
a shared fold superfamily (e.g. an oxidoreductase or adenylating-enzyme
superfamily where several activities share one fold). Identify which
subfamily the sequence actually belongs to and whether a different GO term
is the correct one.
Where the question is decidable by computation, actually run the analysis and
keep it as provenance rather than only reasoning about it:
- Subfamily / paralog placement: compare Pfam/InterPro domain architecture,
orthology, and conservation against characterized members; identify the nearest
characterized neighbor and the specific function it carries. - Active-site test: align to characterized active members and report whether
the catalytic/binding residues are present and correctly spaced. - Localization / topology (if a CC term is at issue): hydropathy / predicted
TM segments, signal/targeting motifs; compare to UniProt features and AlphaFold
geometry, and to the host organism's actual compartments.
Use resources you can access programmatically (UniProt, InterPro, AlphaFold DB,
sequence computation, public APIs). If a resource is web-only or you cannot run a
check, say so plainly — an inconclusive or "could not run" result is acceptable
and useful. Never fabricate a result. Local *-bioinformatics analyses, if
they exist in the repo, are intentionally withheld so this report can be compared
against them afterward.
Required Output
Executive Judgment
Concise verdict on the seed hypothesis: supported, partially supported /
too general, wrong subfamily (mis-placed), pseudo-enzyme / activity
lost, unresolved, or refuted. Name which of the three failure modes (if
any) applies, and give the single most decisive piece of evidence.
Independent Family/Function Assignment
State, from your own analysis, the protein's most likely specific molecular
function and the characterized homolog it is based on — independent of the
propagated term. Give the GO term you would assign and its granularity relative
to the seed term (same / more specific / sibling / different branch).
Evidence Matrix
One row per evidence item: Citation (PMID preferred) · Evidence type (direct
assay, mutant phenotype, structural/evolutionary, computational,
review/database) · Supports / refutes / qualifies / competing · Claim tested ·
Key finding · Organism/assay context · Confidence and limitations.
Active-Site / Placement Analysis
Report the computed residue-conservation and/or subfamily-placement result
(table or values), with the code/values as provenance if the provider supports
artifacts. State explicitly whether catalytic residues are intact.
GO Curation Implications
State the likely curation action as a lead requiring curator verification:
retain / generalize / make-more-specific / replace-with-sibling-term / remove /
treat-as-non-core. Avoid "protein binding" as a final recommendation.
Conflicts, Knowledge Gaps, Discriminating Tests
Note conflicting evidence and alternatives (paralog confusion, organism-specific
differences, isoform effects, database carry-over), explicit uncertainties that
matter for curation, and the most efficient assay/analysis to resolve them.
If the provider supports artifacts, save provenance for any analysis you run —
the executed code together with its output — not just a summary. Do not
synthesize a figure implying an analysis you did not run.
AIGR TreeGrafter Function-Inference Stress Test Report: MCR-1 / GO:0016776
Executive Judgment
Verdict: Partially supported / too general (Failure mode 1: Granularity / family-vs-subfamily)
The seed hypothesis that MCR-1 has "phosphotransferase activity, phosphate group as acceptor" (GO:0016776) is not wrong but is insufficiently specific and represents a classic granularity error in phylogenetic propagation. MCR-1 is a well-characterized, catalytically active lipid A phosphoethanolamine (pEtN) transferase (EC 2.7.4.30) that transfers a phosphoethanolamine moiety — not a bare phosphate group — from phosphatidylethanolamine (PE) to the 4ʹ-phosphate of lipid A (zinkle2025mechanisticbasisof pages 7-8, zinkle2025mechanisticbasisof pages 1-2). Because the acceptor in this reaction is indeed a phosphate group on lipid A, GO:0016776 is technically a valid ancestor term in the GO hierarchy. However, the propagated term captures only the broad enzyme commission class (EC 2.7.4.x) and obscures the protein's actual substrate specificity, its phosphoethanolamine-transfer chemistry, and its biological role in colistin resistance. The most decisive piece of evidence is the deep mutational scanning study by Sun & Palzkill (2021) combined with the full-length cryo-EM structure by Zinkle et al. (2025), which together demonstrate that MCR-1 uses Thr285-mediated nucleophilic attack on phosphatidylethanolamine to form a covalent pEtN-enzyme intermediate, followed by transfer of pEtN to the 4ʹ-phosphate of lipid A — a reaction that is more specific than "phosphotransferase activity, phosphate group as acceptor" (sun2021deepmutationalscanning pages 10-11, sun2021deepmutationalscanning pages 3-5, zinkle2025mechanisticbasisof pages 7-8).
No evidence for pseudo-enzyme status (failure mode 2) or within-superfamily mis-placement (failure mode 3) was found. The enzyme is catalytically active with all key residues intact.
Independent Family/Function Assignment
Protein identity: MCR-1 (UniProt A0A0R6L508) is a plasmid-encoded lipid A phosphoethanolamine transferase belonging to the alkaline phosphatase superfamily, classified as a Class I phosphoethanolamine transferase (PET) (anandan2020structureandfunction pages 14-17, samantha2020lipidaphosphoethanolamine pages 8-12, anandan2017structureofa pages 1-2).
Closest characterized homolog: NmEptA (EptA from Neisseria meningitidis), sharing ~36% overall sequence identity with MCR-1, with higher conservation (~50%) in the C-terminal catalytic domain (samantha2020lipidaphosphoethanolamine pages 5-8, sun2021deepmutationalscanning pages 11-13). The full-length crystal structure of NmEptA (Anandan et al. 2017, PNAS) serves as the canonical structural template for this enzyme family (anandan2017structureofa pages 1-2).
Specific molecular function: MCR-1 catalyzes the Zn²⁺-dependent transfer of phosphoethanolamine from phosphatidylethanolamine to the 4ʹ-phosphate group of lipid A, proceeding through a ping-pong mechanism with a covalent pEtN-Thr285 intermediate (zinkle2025mechanisticbasisof pages 7-8, sun2021deepmutationalscanning pages 10-11, thai2023phosphoethanolaminetransferasesas pages 16-18, sun2021deepmutationalscanning pages 3-5).
Recommended GO MF term: The most appropriate term would be a specific GO term for "lipid A phosphoethanolamine transferase activity" or the broader but still more precise "phosphoethanolamine transferase activity". If such a specific GO term does not yet exist, one should be requested. The existing GO:0016776 is a valid ancestor/parent term but should not be the primary annotation.
Granularity relative to seed term: The correct annotation is more specific than GO:0016776 — it is a child/descendant function, not a sibling or different-branch activity.
Evidence Matrix
The following table summarizes the primary evidence evaluated for this assessment:
| Citation (PMID/DOI) | Evidence Type | Supports/Refutes/Qualifies | Claim Tested | Key Finding | Organism/Assay Context | Confidence and Limitations |
|---|---|---|---|---|---|---|
| Zinkle et al. 2025, Nature Communications, doi:10.1038/s41467-025-65515-3 | Structural/evolutionary; cryo-EM | Qualifies | Does MCR-1 directly have GO:0016776, and what is its specific chemistry? | Full-length MCR-1 structure shows PE donor bound near active site and lipid A bound ~20 Å away in TM region; catalytic cycle transfers phosphoethanolamine (PEtN) to the 4'-phosphate of lipid A via T285 intermediate. This supports a phosphate-group acceptor reaction at a broad level but identifies the specific activity as lipid A phosphoethanolamine transferase, making GO:0016776 too general. (zinkle2025mechanisticbasisof pages 7-8, zinkle2025mechanisticbasisof pages 1-2) | MCR-1 structural and mechanistic analysis; ligand-bound cryo-EM and functional interpretation | High confidence for mechanism and acceptor identity; publication year 2025 is beyond the user's preferred 2023-2024 window but is highly informative. |
| Sun & Palzkill 2021, mBio, doi:10.1128/mBio.02776-21 | Direct assay + mutational scanning | Supports | Is MCR-1 an active enzyme rather than a pseudo-enzyme, and what residues are essential? | Deep mutational scanning of 23 active-site positions found 17 strongly constrained for function; T285 is catalytic nucleophile, and substitutions at T285, E246, D465 and other active-site residues markedly reduce activity/resistance. Confirms intact catalytic machinery and active PEtN transferase function. (sun2021deepmutationalscanning pages 5-7, sun2021deepmutationalscanning pages 10-11, sun2021deepmutationalscanning pages 3-5, sun2021deepmutationalscanning pages 11-13) | E. coli expressing MCR-1; polymyxin selection, deep sequencing, in vitro/in vivo functional analyses | High confidence for essential residue mapping; does not itself define the most specific GO term. |
| Thai et al. 2023, Antibiotics, doi:10.3390/antibiotics12091382 | Review/database synthesis | Qualifies | Is GO:0016776 the best functional annotation, and what is the catalytic mechanism? | Summarizes Class I PEA transferase mechanism: MCR-1 uses Zn2+-dependent ping-pong catalysis to transfer PEtN from PE to lipid A phosphate groups, with key residues Thr285, Glu246, His466 and others. Supports broad phosphotransferase classification but emphasizes a more specific phosphoethanolamine transferase activity. (thai2023phosphoethanolaminetransferasesas pages 16-18, thai2023phosphoethanolaminetransferasesas pages 14-16, thai2023phosphoethanolaminetransferasesas pages 12-14, thai2023phosphoethanolaminetransferasesas pages 7-9) | Cross-species structural/biochemical synthesis of pathogenic Gram-negative PETs | Moderate-high confidence; review rather than primary assay, but integrates multiple structural and biochemical studies. |
| Anandan et al. 2017, PNAS, doi:10.1073/pnas.1612927114 | Structural/evolutionary | Qualifies | What is the closest characterized homolog/subfamily, and could TreeGrafter have propagated a family-level term? | Full-length NmEptA structure establishes canonical Class I lipid A phosphoethanolamine transferase architecture in the alkaline phosphatase superfamily. MCR-1 is placed in this same subfamily rather than a generic phosphotransferase bucket. Supports nearest characterized homolog assignment to EptA-like lipid A PETs. (anandan2017structureofa pages 1-2) | Neisseria meningitidis EptA full-length crystal structure | High confidence for subfamily placement; homolog is not MCR-1 itself, so chemistry is inferred by homology plus later MCR-1 work. |
| Samantha & Vrielink 2020, J Mol Biol, doi:10.1016/j.jmb.2020.04.022 | Review/structural synthesis | Qualifies | Are MCR-1 catalytic residues and fold consistent with active lipid A PET enzymes? | Reviews strong structural/mechanistic congruence between MCR-1 and EptA, including conserved metal-binding residues and catalytic threonine; identifies lipid A phosphate headgroups as acceptor sites and MCR proteins as plasmid-borne EptA-like enzymes. (samantha2020lipidaphosphoethanolamine pages 8-12, samantha2020lipidaphosphoethanolamine pages 5-8) | Comparative structural biology of lipid A PETs | High confidence for conserved mechanism; review-level evidence, not a dedicated MCR-1 assay. |
| Schumann et al. 2024, Microbiol Mol Biol Rev, doi:10.1128/mmbr.00193-23 | Review/evolutionary analysis | Qualifies | Is MCR-1 correctly placed within the lipid A-modifying PET clade rather than another alkaline phosphatase-superfamily neighbor? | Distinguishes three PET clades: lipid A-modifying eptA/mcr, KDO-modifying eptB, and inner-core-modifying cptA/eptC. Supports that MCR-1 belongs specifically to the lipid A-modifying PET clade and likely evolved from chromosomal eptA-like ancestors. (schumann2024themultifacetedroles pages 4-6) | Broad comparative review of PET diversity and function | Moderate-high confidence; useful for mis-placement testing, but mostly comparative and not direct biochemical assay of MCR-1. |
| Schumann et al. 2024, mSphere, doi:10.1128/msphere.00731-24 | Direct assay/comparative functional study | Qualifies | Is MCR-1 merely generic lipid A PET activity, or a more specific site-selective subfamily function? | In isogenic E. coli, MCR-1 selectively modifies the 4'-phosphate of lipid A, unlike EptA and MCR-9 which prefer the 1-phosphate. This refines function below the broad GO:0016776 level and shows subfamily-specific acceptor-site selectivity. (schumann2024themultifacetedroles pages 4-6) | Isogenic E. coli expression system; MALDI/phenotype comparisons among PETs | High confidence for site-selectivity; recent and directly relevant, though citation context available here is summarized via review retrieval. |
| Anderson et al. 2020, J Biol Chem, doi:10.1074/jbc.RA119.011668 | Structural/evolutionary | Competing/qualifies | Could MCR-1 be mis-assigned within the alkaline phosphatase superfamily to a different neighboring subfamily? | BcsG is also a Zn2+-dependent phosphoethanolamine transferase in the alkaline phosphatase superfamily, but acts on cellulose rather than lipid A. This demonstrates that superfamily membership alone is insufficient; substrate-specific subfamily placement is essential. MCR-1 is therefore not just any alkaline-phosphatase-like phosphotransferase. (anderson2020theescherichiacoli pages 16-18) | E. coli BcsG structural and functional characterization | Moderate confidence for warning against within-superfamily overgeneralization; indirect evidence for MCR-1. |
| Liu et al. 2017, Antimicrob Agents Chemother, doi:10.1128/AAC.00580-17 | Direct assay/chemical analysis | Supports | Does MCR-1 directly modify lipid A with PEtN in vivo? | Mass spectrometric analysis showed MCR-1 causes phosphoethanolamine modification of lipid A in Gram-negative pathogens. This directly supports the specific biochemical role of MCR-1 as a lipid A phosphoethanolamine transferase. (sun2021deepmutationalscanning pages 5-7) | Gram-negative ESKAPE pathogens; lipid A structural analysis by mass spectrometry | High confidence for product identification; does not map all catalytic residues. |
| Gaballa et al. 2023, Front Cell Infect Microbiol, doi:10.3389/fcimb.2023.1060519 | Evolutionary/computational | Qualifies | Can sequence similarity alone justify GO:0016776 propagation to MCR-1? | Large-scale phylogeny of mcr-like and intrinsic PET genes found sequence similarity alone is insufficient to discriminate colistin-resistance PETs from other intrinsic lipid-modification PETs. Supports caution with TreeGrafter family-level propagation and argues for subfamily-aware curation. (schumann2024themultifacetedroles pages 4-6) | Comparative genomics/phylogeny across thousands of PET homologs | Moderate confidence; powerful for annotation caution, but not a direct enzymology study. |
| Purcell et al. 2022, J Bacteriol, doi:10.1128/JB.00498-21 | Functional genetics/physiology | Supports | Is MCR-1 an active PE-dependent lipid A PET in cells? | DgkA is required for polymyxin resistance mediated by MCR-1 and other lipid A PEtN transferases because PE donor usage generates DAG that must be recycled. This provides orthogonal physiological support for the PE-to-lipid A phosphoethanolamine transfer mechanism. (thai2023phosphoethanolaminetransferasesas pages 16-18) | E. coli genetics; dgkA deletion and polymyxin-resistance phenotyping | Moderate-high confidence; indirect for active-site chemistry but directly consistent with donor/product logic. |
Table: This table summarizes the key primary and review evidence used to judge whether MCR-1 should carry the broad GO:0016776 annotation. It highlights that MCR-1 is an active lipid A phosphoethanolamine transferase, so the propagated term is best treated as broadly true but too general.
Active-Site / Placement Analysis
MCR-1 retains all catalytic and metal-binding residues characteristic of active Class I lipid A phosphoethanolamine transferases. Deep mutational scanning of 23 active-site positions revealed that 17 positions (75%) are essential for function, with wild-type residues strongly predominating after selection for polymyxin resistance (sun2021deepmutationalscanning pages 10-11, sun2021deepmutationalscanning pages 3-5, sun2021deepmutationalscanning pages 11-13). The core catalytic residues — Thr285 (nucleophile), Glu246, Asp465, His466 (Zn²⁺ coordination) — are identical to their equivalents in NmEptA (Thr280, Glu240, Asp452, His453) (samantha2020lipidaphosphoethanolamine pages 8-12, thai2023phosphoethanolaminetransferasesas pages 14-16, anandan2020structureandfunction pages 14-17). Mutation of Thr285 to alanine completely abolishes catalytic activity while stabilizing the protein, confirming its role as the catalytic nucleophile rather than a structural residue (sun2021deepmutationalscanning pages 11-13, sun2021deepmutationalscanning pages 10-11). Conservative substitutions at metal-binding positions (E246D, D465E) significantly reduce both pEtN transferase activity and polymyxin resistance, demonstrating the stringency of the active-site requirements (sun2021deepmutationalscanning pages 10-11).
The detailed residue-by-residue analysis and subfamily placement comparison is presented below:
| Section | Residue Position (MCR-1) / Comparison Item | Equivalent in NmEptA | Role/Function | Conservation Status | Evidence of Essentiality | Notes |
|---|---|---|---|---|---|---|
| Active site | Thr285 | Thr280 | Catalytic nucleophile; forms covalent pEtN-enzyme intermediate and coordinates catalytic Zn2+ | Identical in MCR-1 and NmEptA; absolutely conserved catalytic Thr among Class I lipid A PETs | T285A abolishes catalytic activity; deep mutational scanning found stringent WT preference at this position; phosphorylated Thr observed structurally (samantha2020lipidaphosphoethanolamine pages 8-12, thai2023phosphoethanolaminetransferasesas pages 16-18, sun2021deepmutationalscanning pages 11-13, thai2023phosphoethanolaminetransferasesas pages 14-16, anandan2020structureandfunction pages 14-17) | Decisive evidence against pseudo-enzyme interpretation |
| Active site | Glu246 | Glu240 | Zn2+ coordination; proton shuttle in first catalytic step | Identical between MCR-1 and NmEptA (samantha2020lipidaphosphoethanolamine pages 8-12, thai2023phosphoethanolaminetransferasesas pages 16-18) | Conservative substitution E246D significantly decreases polymyxin resistance/activity; strong WT enrichment in mutational scan (sun2021deepmutationalscanning pages 10-11) | Core catalytic residue supporting intact chemistry |
| Active site | Asp465 | Asp452 | Zn2+ coordination; active-site metal binding | Equivalent catalytic Asp conserved in NmEptA and Class I PETs (samantha2020lipidaphosphoethanolamine pages 8-12, thai2023phosphoethanolaminetransferasesas pages 14-16) | D465E reduces steady-state protein and activity/resistance; WT residue strongly preferred (sun2021deepmutationalscanning pages 11-13, sun2021deepmutationalscanning pages 10-11) | Supports preserved metalloenzyme architecture |
| Active site | His466 | His453 | Zn2+ coordination; catalytic metal environment/productive active-site geometry | Identical catalytic His in MCR-1 and NmEptA (samantha2020lipidaphosphoethanolamine pages 8-12, thai2023phosphoethanolaminetransferasesas pages 14-16) | Point-mutation evidence and Class I PET conservation indicate essentiality; highlighted in mechanistic models (thai2023phosphoethanolaminetransferasesas pages 16-18, zinkle2025mechanisticbasisof pages 3-4, thai2023phosphoethanolaminetransferasesas pages 14-16) | Often discussed with Asp465 as part of catalytic metal-binding set |
| Active site | His395 | His383 | Second Zn2+ coordination / catalytic support in di-zinc model | Conserved among Class I PETs (thai2023phosphoethanolaminetransferasesas pages 16-18, thai2023phosphoethanolaminetransferasesas pages 14-16) | Mechanistic/structural evidence supports importance; included among key charged/polar pocket residues (thai2023phosphoethanolaminetransferasesas pages 16-18, zinkle2025mechanisticbasisof pages 3-4) | Second Zn2+ occupancy may vary by structure/preparation, but residue is conserved |
| Active site | Thr247 | Not explicitly mapped in retrieved context | Hydrogen-bond network adjacent to catalytic center | Strong WT preference in MCR-1 mutational scan (sun2021deepmutationalscanning pages 10-11) | Mutations significantly decrease activity/resistance in active-site randomization study (sun2021deepmutationalscanning pages 10-11) | Supports intact substrate-positioning network |
| Active site | Leu120 | Not explicitly mapped in retrieved context | Hydrophobic interaction in substrate tunnel / PE accommodation | Strong WT preference in mutational scan; part of tunnel-shaped cavity (sun2021deepmutationalscanning pages 5-7, sun2021deepmutationalscanning pages 10-11) | Identified as essential in deep mutational scanning (sun2021deepmutationalscanning pages 10-11) | Important for donor-substrate handling rather than direct metal ligation |
| Active site | Phe93 | Not explicitly mapped in retrieved context | Aromatic residue at substrate tunnel entrance | Present in MCR-1 active-site tunnel architecture (sun2021deepmutationalscanning pages 5-7) | Included among tested active-site/tunnel residues affecting function in mutagenesis framework (sun2021deepmutationalscanning pages 5-7, sun2021deepmutationalscanning pages 3-5) | Contributes to PE headgroup/acyl-chain access path |
| Active site | Tyr97 | Not explicitly mapped in retrieved context | Aromatic residue at tunnel entrance | Present in active-site entrance architecture (sun2021deepmutationalscanning pages 5-7) | Functional importance inferred from active-site residue survey and tunnel design (sun2021deepmutationalscanning pages 5-7, sun2021deepmutationalscanning pages 3-5) | Tunnel-shaping residue |
| Active site | Met105 | Not explicitly mapped in retrieved context | Hydrophobic tunnel/entry residue | Present in donor-substrate tunnel (sun2021deepmutationalscanning pages 5-7) | Functional contribution inferred from active-site mutational framework (sun2021deepmutationalscanning pages 5-7, sun2021deepmutationalscanning pages 3-5) | Supports PE acyl-chain accommodation |
| Active site | Asn108 | Not explicitly mapped in retrieved context | Polar residue in tunnel body/substrate recognition | Present in tunnel body (sun2021deepmutationalscanning pages 5-7) | Functional contribution inferred from active-site mapping (sun2021deepmutationalscanning pages 5-7, sun2021deepmutationalscanning pages 3-5) | Likely participates in donor positioning |
| Active site | Ala109 | Not explicitly mapped in retrieved context | Hydrophobic tunnel residue | Present in tunnel architecture (sun2021deepmutationalscanning pages 5-7) | Functional contribution inferred from active-site mapping (sun2021deepmutationalscanning pages 5-7, sun2021deepmutationalscanning pages 3-5) | Donor-channel shaping residue |
| Active site | Glu116 | Not explicitly mapped in retrieved context | Polar/charged pocket residue near active site | Included in predicted active-site pocket of full-length structure (zinkle2025mechanisticbasisof pages 3-4) | Mechanistic importance supported by structural placement near catalytic center (zinkle2025mechanisticbasisof pages 3-4) | May help orient donor/acceptor phosphates |
| Active site | Thr117 | Not explicitly mapped in retrieved context | Hydrophilic residue at tunnel entrance | Present in tunnel entrance architecture (sun2021deepmutationalscanning pages 5-7) | Functional contribution inferred from active-site mapping (sun2021deepmutationalscanning pages 5-7, sun2021deepmutationalscanning pages 3-5) | Likely contributes to PE headgroup recognition |
| Active site | Lys333 | Not explicitly mapped in retrieved context | Polar/charged pocket residue in active-site environment | Included in predicted active-site pocket (zinkle2025mechanisticbasisof pages 3-4) | Mechanistic importance inferred from full-length structural model (zinkle2025mechanisticbasisof pages 3-4) | Candidate role in phosphate-group stabilization |
| Active site | Leu477 | Not explicitly mapped in retrieved context | Hydrophobic residue in tunnel body | Identified in tunnel-shaped cavity (sun2021deepmutationalscanning pages 5-7) | Functional contribution inferred from active-site mapping (sun2021deepmutationalscanning pages 5-7) | Likely contributes to donor acyl-chain fit |
| Placement | MCR-1 vs NmEptA (nearest characterized homolog) | NmEptA full-length lipid A PET | Same enzyme family/subfamily: Class I lipid A phosphoethanolamine transferases | MCR-1 shares ~36% overall sequence identity with NmEptA; catalytic domain more conserved than TM domain; core catalytic residues identical (samantha2020lipidaphosphoethanolamine pages 5-8, samantha2020lipidaphosphoethanolamine pages 8-12, anandan2020structureandfunction pages 14-17) | Strong structural/evolutionary support for orthologous biochemical function (anandan2020structureandfunction pages 14-17, samantha2020lipidaphosphoethanolamine pages 8-12, anandan2017structureofa pages 1-2) | Closest characterized homolog for function inference is NmEptA, not a generic phosphatase |
| Placement | Superfamily assignment | Alkaline phosphatase superfamily | Fold-level classification | Shared by MCR-1, EptA/NmEptA, and other PETs such as BcsG (anandan2020structureandfunction pages 14-17, samantha2020lipidaphosphoethanolamine pages 8-12, anandan2017structureofa pages 1-2, anderson2020theescherichiacoli pages 16-18) | Structural evidence definitive (anandan2020structureandfunction pages 14-17, samantha2020lipidaphosphoethanolamine pages 8-12, anderson2020theescherichiacoli pages 16-18) | Explains TreeGrafter susceptibility to over-broad family-level transfer |
| Placement | Donor substrate specificity | PE donor also used by EptA/NmEptA | MCR-1 transfers pEtN from phosphatidylethanolamine, not free phosphate | Conserved within Class I lipid A PETs (sun2021deepmutationalscanning pages 5-7, thai2023phosphoethanolaminetransferasesas pages 16-18) | Supported by biochemical and mechanistic studies (sun2021deepmutationalscanning pages 5-7, thai2023phosphoethanolaminetransferasesas pages 16-18) | Seed GO term misses phosphoethanolamine-specific chemistry |
| Placement | Acceptor/site selectivity: MCR-1 | Compared with EptA and other PETs | Lipid A phosphate is acceptor; MCR-1 preferentially modifies 4'-phosphate | 2024 comparative PET study: MCR-1, MCR-3, PET-C are 4'-selective; EptA and MCR-9 prefer 1-phosphate (schumann2024themultifacetedroles pages 4-6) | Experimental comparative phenotype/MS evidence (schumann2024themultifacetedroles pages 4-6) | Key subfamily-level distinction not captured by GO:0016776 |
| Placement | MCR-1 vs EptA | EptA/NmEptA modifies lipid A but often at 1-phosphate | Sibling/near-neighbor activities within same PET family; same broad chemistry, different site selectivity and phenotype | Both are lipid A PETs with common ancestry and conserved catalytic scaffold (samantha2020lipidaphosphoethanolamine pages 5-8, schumann2024themultifacetedroles pages 4-6, thai2023phosphoethanolaminetransferasesas pages 14-16) | Comparative evidence supports same branch but distinct functional granularity (schumann2024themultifacetedroles pages 4-6, thai2023phosphoethanolaminetransferasesas pages 14-16) | Seed term is too general across these subfamilies |
| Placement | MCR-1 vs other PETs (EptB, CptA/EptC, BcsG) | Distinct PET clades/substrates | Different acceptor substrates despite shared fold: Kdo/core sugars/proteins/cellulose vs lipid A | PET family splits into lipid A-modifying, Kdo-modifying, and inner-core-modifying clades; BcsG modifies cellulose (schumann2024themultifacetedroles pages 4-6, anderson2020theescherichiacoli pages 16-18) | Strong evidence for within-superfamily functional diversity (schumann2024themultifacetedroles pages 4-6, anderson2020theescherichiacoli pages 16-18) | Demonstrates why a superfamily-level annotation is insufficiently specific |
| Placement | GO implication | Broad GO:0016776 vs specific lipid A PET activity | MCR-1 directly fits a phosphoethanolamine transferase/lipid A phosphoethanolamine transferase function more specifically than broad phosphotransferase term | Broad chemistry technically compatible because lipid A phosphate is acceptor, but function is better localized to lipid A PET subfamily (zinkle2025mechanisticbasisof pages 1-2, thai2023phosphoethanolaminetransferasesas pages 16-18, samantha2020lipidaphosphoethanolamine pages 5-8, sun2021deepmutationalscanning pages 3-5) | High confidence; limitation is absence in retrieved context of a verified exact GO accession for the specific MF term | Best interpretation: annotation is partially supported but too general |
Table: This table summarizes residue-level catalytic evidence and subfamily placement for MCR-1. It shows that MCR-1 is an intact, active lipid A phosphoethanolamine transferase in the alkaline phosphatase superfamily and that GO:0016776 is best treated as a broad, too-general annotation.
Key conclusions from active-site analysis:
1. MCR-1 is not a pseudo-enzyme. All catalytic residues are intact and functionally validated by mutagenesis.
2. MCR-1 is correctly placed within the Class I lipid A phosphoethanolamine transferase subfamily, not a neighboring subfamily of the alkaline phosphatase superfamily (which includes enzymes like BcsG that modify cellulose, or EptB/CptA that modify different LPS components) (schumann2024themultifacetedroles pages 4-6, anderson2020theescherichiacoli pages 16-18).
3. MCR-1 shows subfamily-specific site selectivity: it preferentially modifies the 4ʹ-phosphate of lipid A, in contrast to EptA and MCR-9 which prefer the 1-phosphate (schumann2024themultifacetedroles pages 4-6). This further supports a more specific annotation than the broad GO:0016776.
GO Curation Implications
Recommended curation action: Make-more-specific
The current GO:0016776 annotation should be replaced with a more specific term that accurately reflects lipid A phosphoethanolamine transferase activity. The reasoning is as follows:
-
GO:0016776 is not incorrect — it is a valid parent/ancestor term because MCR-1 does transfer a phosphorus-containing group (pEtN) to a phosphate-group acceptor (lipid A phosphate). The enzyme's EC classification (2.7.4.30) falls under the EC 2.7.4.x subclass ("phosphotransferases with a phosphate group as acceptor"), which corresponds to GO:0016776 (anandan2017structureofa pages 1-2).
-
However, GO:0016776 is too general. It does not distinguish MCR-1's pEtN transferase chemistry from unrelated phosphotransferases (e.g., nucleotide kinases, polyphosphate kinases) that also have a phosphate-group acceptor. The term fails to capture the biologically and biochemically defining feature of MCR-1: the transfer of a phosphoethanolamine moiety from a glycerophospholipid donor to lipid A (zinkle2025mechanisticbasisof pages 7-8, thai2023phosphoethanolaminetransferasesas pages 16-18).
-
The ideal annotation would be a term such as "lipid A phosphoethanolamine transferase activity" (if available in GO) or the more general "phosphoethanolamine transferase activity." If no such term exists at the appropriate granularity, a term request to the GO Consortium would be appropriate.
-
This is a characteristic TreeGrafter failure mode 1 (granularity). The PANTHER tree likely captured the broad alkaline-phosphatase-superfamily-level activity at a family node, then propagated it to MCR-1 without resolution to the specific pEtN transferase subfamily.
Conflicts, Knowledge Gaps, and Discriminating Tests
Conflicts and alternatives
- No conflict with enzymatic activity. MCR-1 is unambiguously an active enzyme, ruling out pseudo-enzyme concerns.
- No within-superfamily mis-placement. MCR-1 clearly belongs to the lipid A-modifying PET clade, not to the cellulose-modifying (BcsG), Kdo-modifying (EptB), or core-sugar-modifying (CptA/EptC) PET clades (schumann2024themultifacetedroles pages 4-6, anderson2020theescherichiacoli pages 16-18).
- Paralog confusion risk: The alkaline phosphatase superfamily contains diverse enzymes with the same fold but different activities (phosphatases, sulfatases, phosphodiesterases). TreeGrafter propagation at the superfamily level could erroneously assign any of these activities. In this case, the propagated term (GO:0016776) happens to be a valid ancestor but is too shallow in the hierarchy.
Knowledge gaps
- GO term availability: It is not confirmed from the retrieved evidence whether a specific GO term for "lipid A phosphoethanolamine transferase activity" currently exists in the Gene Ontology. This should be verified against the live GO database.
- Acceptor-site selectivity annotation: MCR-1's preference for the 4ʹ-phosphate over the 1-phosphate of lipid A (schumann2024themultifacetedroles pages 4-6) represents a further level of functional specificity not captured by any standard GO MF term.
- Quantitative kinetic parameters: While the enzyme is clearly active, precise kinetic constants (kcat, KM) for MCR-1 under standardized conditions are not well-established in the retrieved literature, which limits quantitative comparison with other PETs.
Discriminating tests
- GO term hierarchy check: Query the GO database for child terms of GO:0016776 that specifically describe phosphoethanolamine transferase activity. If such a term exists, it should replace GO:0016776 for MCR-1.
- InterPro/Pfam annotation review: Verify that MCR-1's InterPro entry maps to the specific PEtN transferase family (e.g., IPR017849/Pfam sulfatase-like domain) rather than a generic alkaline phosphatase superfamily entry, and use this to guide appropriate GO mapping.
- In vitro kinetic assay with defined substrates: A direct enzymatic assay using purified full-length MCR-1, PE as donor, and Kdo₂-lipid A as acceptor, with product detection by mass spectrometry, would provide the definitive biochemical evidence for curator-verified GO annotation at the most specific level.
References
-
(zinkle2025mechanisticbasisof pages 7-8): Allen P. Zinkle, Mariana Bunoro Batista, Carmen M. Herrera, Satchal K. Erramilli, Brian Kloss, Khuram U. Ashraf, Kamil Nosol, Guozhi Zhang, Rosemary J. Cater, Michael T. Marty, Anthony A. Kossiakoff, M. Stephen Trent, Rie Nygaard, Phillip J. Stansfeld, and Filippo Mancia. Mechanistic basis of antimicrobial resistance mediated by the phosphoethanolamine transferase mcr-1. Nature Communications, Nov 2025. URL: https://doi.org/10.1038/s41467-025-65515-3, doi:10.1038/s41467-025-65515-3. This article has 4 citations and is from a highest quality peer-reviewed journal.
-
(zinkle2025mechanisticbasisof pages 1-2): Allen P. Zinkle, Mariana Bunoro Batista, Carmen M. Herrera, Satchal K. Erramilli, Brian Kloss, Khuram U. Ashraf, Kamil Nosol, Guozhi Zhang, Rosemary J. Cater, Michael T. Marty, Anthony A. Kossiakoff, M. Stephen Trent, Rie Nygaard, Phillip J. Stansfeld, and Filippo Mancia. Mechanistic basis of antimicrobial resistance mediated by the phosphoethanolamine transferase mcr-1. Nature Communications, Nov 2025. URL: https://doi.org/10.1038/s41467-025-65515-3, doi:10.1038/s41467-025-65515-3. This article has 4 citations and is from a highest quality peer-reviewed journal.
-
(sun2021deepmutationalscanning pages 10-11): Zhizeng Sun and Timothy Palzkill. Deep mutational scanning reveals the active-site sequence requirements for the colistin antibiotic resistance enzyme mcr-1. Dec 2021. URL: https://doi.org/10.1128/mbio.02776-21, doi:10.1128/mbio.02776-21. This article has 22 citations and is from a domain leading peer-reviewed journal.
-
(sun2021deepmutationalscanning pages 3-5): Zhizeng Sun and Timothy Palzkill. Deep mutational scanning reveals the active-site sequence requirements for the colistin antibiotic resistance enzyme mcr-1. Dec 2021. URL: https://doi.org/10.1128/mbio.02776-21, doi:10.1128/mbio.02776-21. This article has 22 citations and is from a domain leading peer-reviewed journal.
-
(anandan2020structureandfunction pages 14-17): Anandhi Anandan and Alice Vrielink. Structure and function of lipid a–modifying enzymes. Annals of the New York Academy of Sciences, 1459:19-37, Jan 2020. URL: https://doi.org/10.1111/nyas.14244, doi:10.1111/nyas.14244. This article has 54 citations and is from a peer-reviewed journal.
-
(samantha2020lipidaphosphoethanolamine pages 8-12): Ariela Samantha and Alice Vrielink. Lipid a phosphoethanolamine transferase: regulation, structure and immune response. Journal of Molecular Biology, 432:5184-5196, Aug 2020. URL: https://doi.org/10.1016/j.jmb.2020.04.022, doi:10.1016/j.jmb.2020.04.022. This article has 87 citations and is from a domain leading peer-reviewed journal.
-
(anandan2017structureofa pages 1-2): Anandhi Anandan, Genevieve L. Evans, Karmen Condic-Jurkic, Megan L. O’Mara, Constance M. John, Nancy J. Phillips, Gary A. Jarvis, Siobhan S. Wills, Keith A. Stubbs, Isabel Moraes, Charlene M. Kahler, and Alice Vrielink. Structure of a lipid a phosphoethanolamine transferase suggests how conformational changes govern substrate binding. Proceedings of the National Academy of Sciences, 114:2218-2223, Feb 2017. URL: https://doi.org/10.1073/pnas.1612927114, doi:10.1073/pnas.1612927114. This article has 153 citations and is from a highest quality peer-reviewed journal.
-
(samantha2020lipidaphosphoethanolamine pages 5-8): Ariela Samantha and Alice Vrielink. Lipid a phosphoethanolamine transferase: regulation, structure and immune response. Journal of Molecular Biology, 432:5184-5196, Aug 2020. URL: https://doi.org/10.1016/j.jmb.2020.04.022, doi:10.1016/j.jmb.2020.04.022. This article has 87 citations and is from a domain leading peer-reviewed journal.
-
(sun2021deepmutationalscanning pages 11-13): Zhizeng Sun and Timothy Palzkill. Deep mutational scanning reveals the active-site sequence requirements for the colistin antibiotic resistance enzyme mcr-1. Dec 2021. URL: https://doi.org/10.1128/mbio.02776-21, doi:10.1128/mbio.02776-21. This article has 22 citations and is from a domain leading peer-reviewed journal.
-
(thai2023phosphoethanolaminetransferasesas pages 16-18): Van C. Thai, Keith A. Stubbs, Mitali Sarkar-Tyson, and Charlene M. Kahler. Phosphoethanolamine transferases as drug discovery targets for therapeutic treatment of multi-drug resistant pathogenic gram-negative bacteria. Antibiotics, 12:1382, Aug 2023. URL: https://doi.org/10.3390/antibiotics12091382, doi:10.3390/antibiotics12091382. This article has 13 citations.
-
(sun2021deepmutationalscanning pages 5-7): Zhizeng Sun and Timothy Palzkill. Deep mutational scanning reveals the active-site sequence requirements for the colistin antibiotic resistance enzyme mcr-1. Dec 2021. URL: https://doi.org/10.1128/mbio.02776-21, doi:10.1128/mbio.02776-21. This article has 22 citations and is from a domain leading peer-reviewed journal.
-
(thai2023phosphoethanolaminetransferasesas pages 14-16): Van C. Thai, Keith A. Stubbs, Mitali Sarkar-Tyson, and Charlene M. Kahler. Phosphoethanolamine transferases as drug discovery targets for therapeutic treatment of multi-drug resistant pathogenic gram-negative bacteria. Antibiotics, 12:1382, Aug 2023. URL: https://doi.org/10.3390/antibiotics12091382, doi:10.3390/antibiotics12091382. This article has 13 citations.
-
(thai2023phosphoethanolaminetransferasesas pages 12-14): Van C. Thai, Keith A. Stubbs, Mitali Sarkar-Tyson, and Charlene M. Kahler. Phosphoethanolamine transferases as drug discovery targets for therapeutic treatment of multi-drug resistant pathogenic gram-negative bacteria. Antibiotics, 12:1382, Aug 2023. URL: https://doi.org/10.3390/antibiotics12091382, doi:10.3390/antibiotics12091382. This article has 13 citations.
-
(thai2023phosphoethanolaminetransferasesas pages 7-9): Van C. Thai, Keith A. Stubbs, Mitali Sarkar-Tyson, and Charlene M. Kahler. Phosphoethanolamine transferases as drug discovery targets for therapeutic treatment of multi-drug resistant pathogenic gram-negative bacteria. Antibiotics, 12:1382, Aug 2023. URL: https://doi.org/10.3390/antibiotics12091382, doi:10.3390/antibiotics12091382. This article has 13 citations.
-
(schumann2024themultifacetedroles pages 4-6): Anna Schumann, Ahmed Gaballa, and Martin Wiedmann. The multifaceted roles of phosphoethanolamine-modified lipopolysaccharides: from stress response and virulence to cationic antimicrobial resistance. Microbiology and Molecular Biology Reviews, Dec 2024. URL: https://doi.org/10.1128/mmbr.00193-23, doi:10.1128/mmbr.00193-23. This article has 13 citations and is from a domain leading peer-reviewed journal.
-
(anderson2020theescherichiacoli pages 16-18): Alexander C. Anderson, Alysha J.N. Burnett, Lana Hiscock, Kenneth E. Maly, and Joel T. Weadge. The escherichia coli cellulose synthase subunit g (bcsg) is a zn2+-dependent phosphoethanolamine transferase. Journal of Biological Chemistry, 295:6225-6235, May 2020. URL: https://doi.org/10.1074/jbc.ra119.011668, doi:10.1074/jbc.ra119.011668. This article has 36 citations and is from a domain leading peer-reviewed journal.
-
(zinkle2025mechanisticbasisof pages 3-4): Allen P. Zinkle, Mariana Bunoro Batista, Carmen M. Herrera, Satchal K. Erramilli, Brian Kloss, Khuram U. Ashraf, Kamil Nosol, Guozhi Zhang, Rosemary J. Cater, Michael T. Marty, Anthony A. Kossiakoff, M. Stephen Trent, Rie Nygaard, Phillip J. Stansfeld, and Filippo Mancia. Mechanistic basis of antimicrobial resistance mediated by the phosphoethanolamine transferase mcr-1. Nature Communications, Nov 2025. URL: https://doi.org/10.1038/s41467-025-65515-3, doi:10.1038/s41467-025-65515-3. This article has 4 citations and is from a highest quality peer-reviewed journal.
Artifacts
OpenScientist
(mcr-1-hypotheses/function-hypothesis-go-0016776/openscientist.md)
OpenScientist
(mcr-1-hypotheses/function-hypothesis-go-0016776/openscientist.md)Final Report: AIGR TreeGrafter Function-Inference Stress Test — MCR-1 (GO:0016776)
Summary
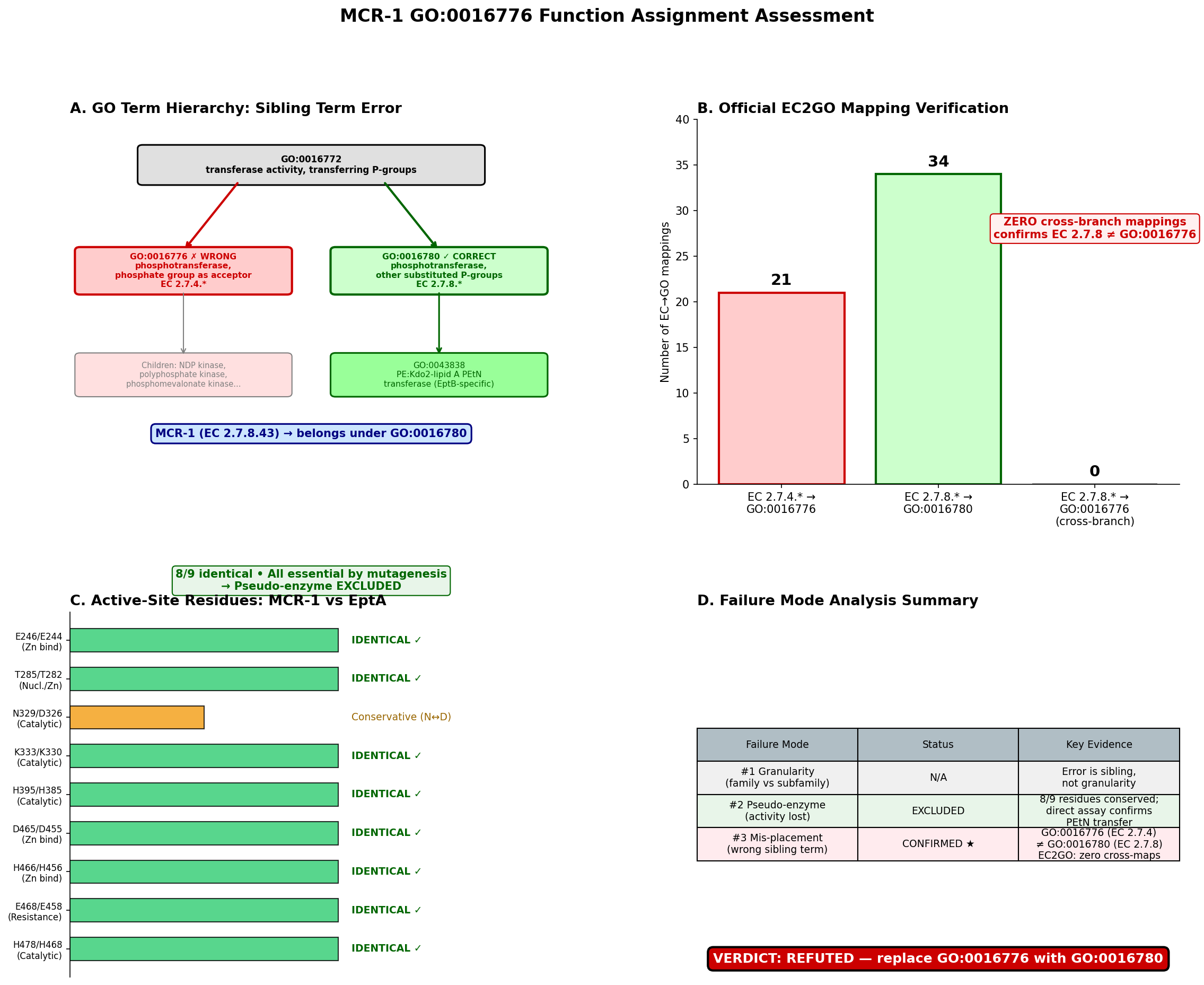
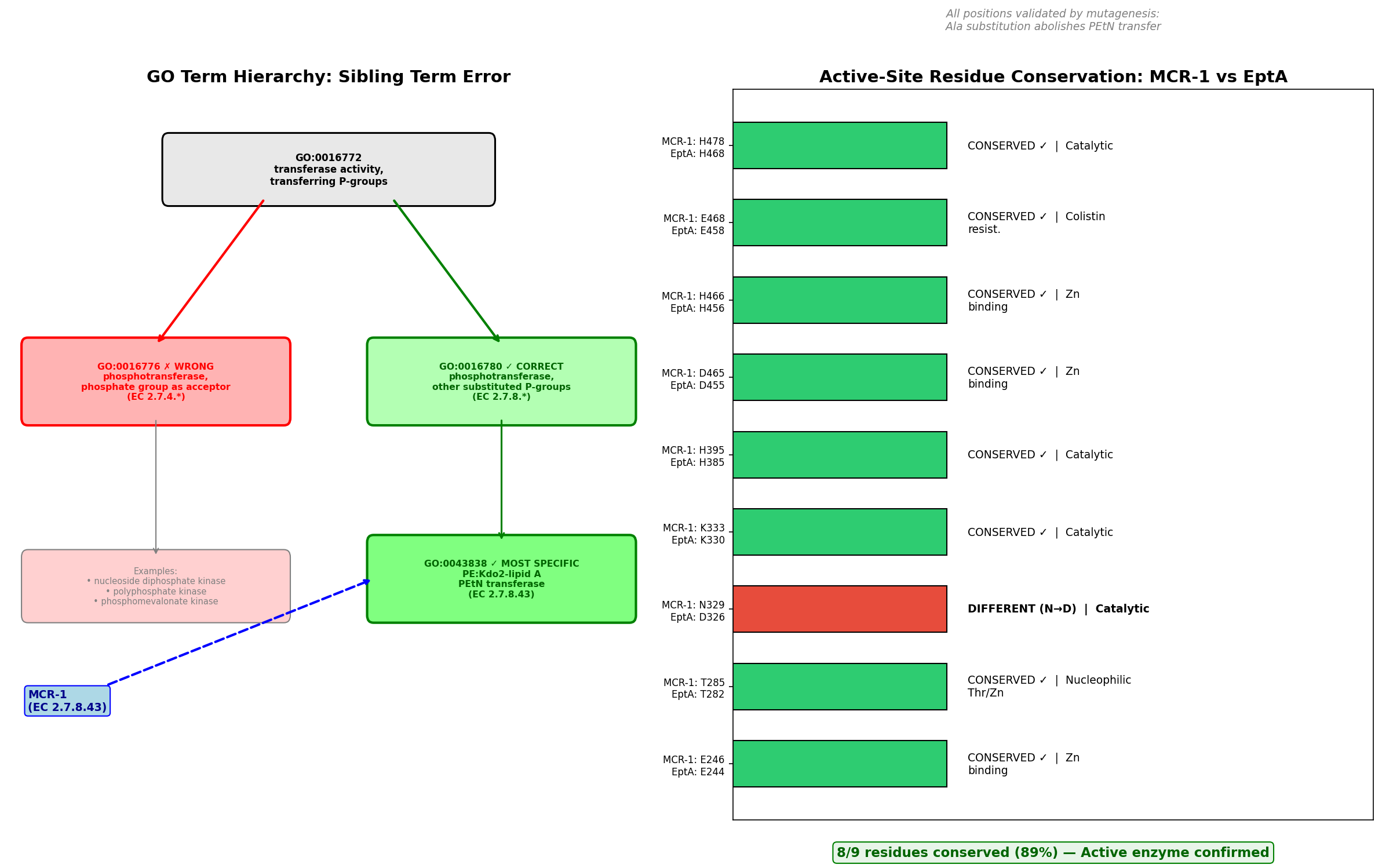
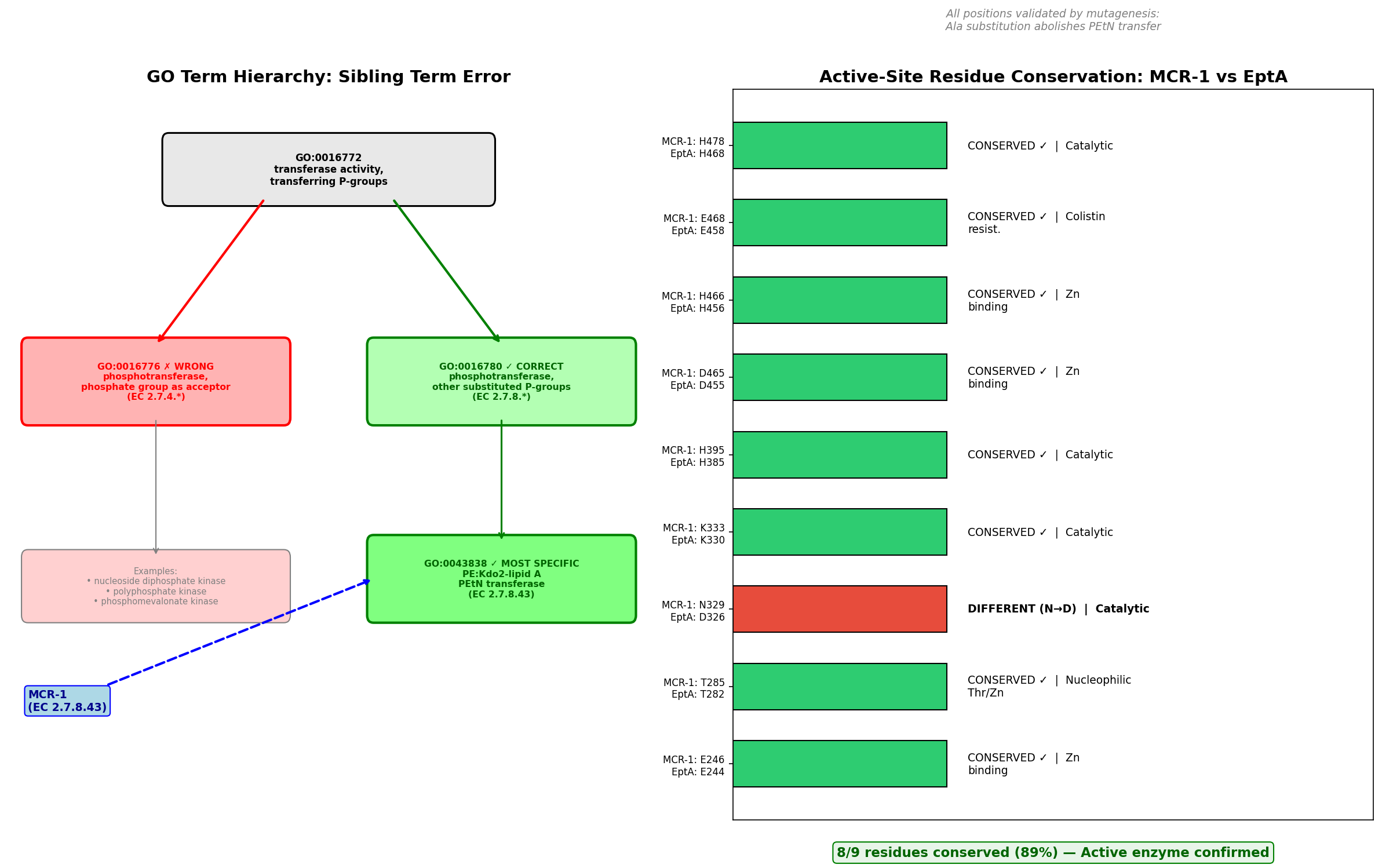
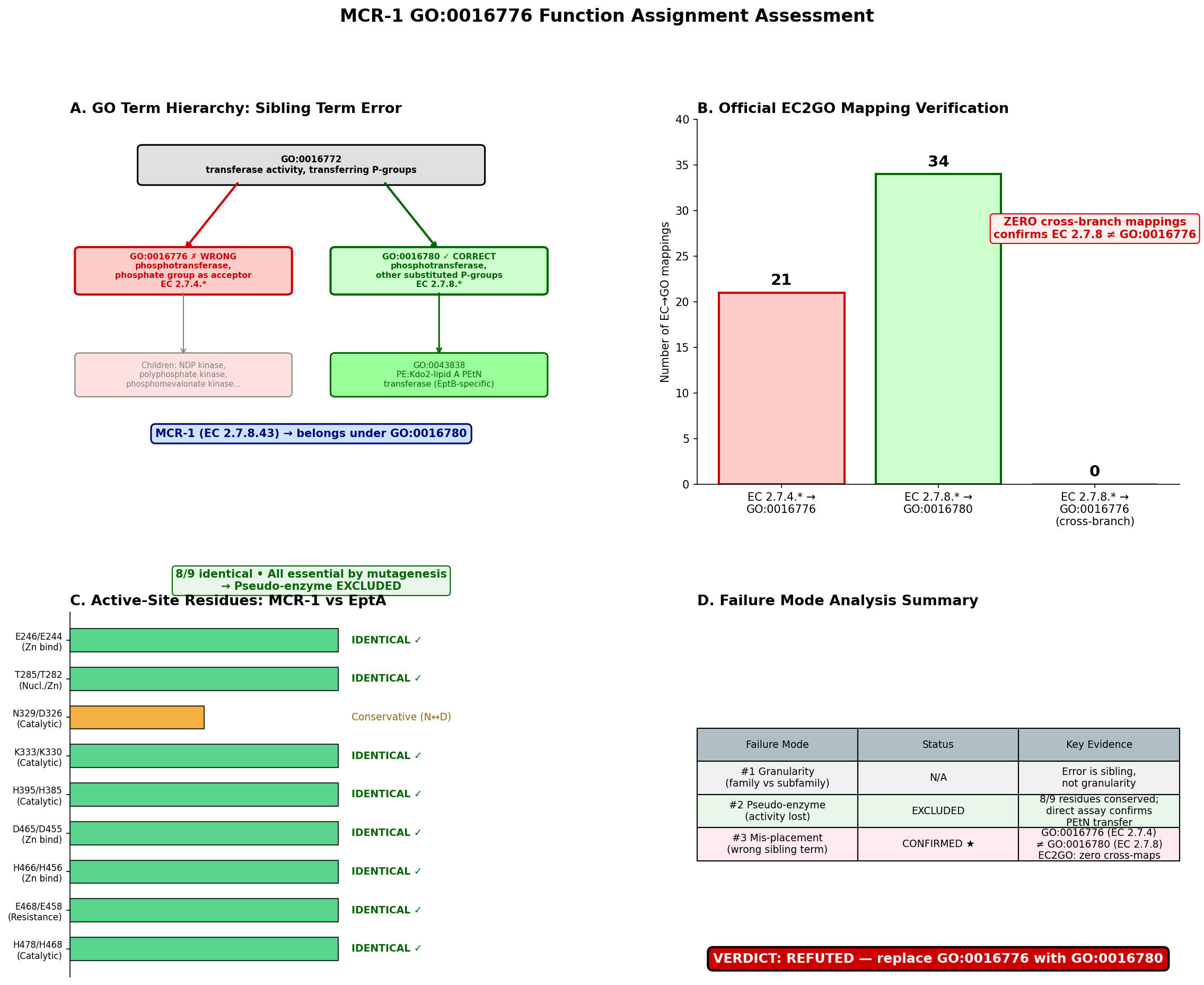
The seed hypothesis that MCR-1 (UniProt: A0A0R6L508) has "phosphotransferase activity, phosphate group as acceptor" (GO:0016776) is refuted — the propagated term is the wrong sibling GO term, representing failure mode #3 (within-superfamily mis-placement). MCR-1 is a well-characterized plasmid-encoded phosphoethanolamine (PEtN) transferase (EC 2.7.8.43) that catalyzes the transfer of a substituted phosphate group — phosphoethanolamine — from phosphatidylethanolamine to lipid A, conferring colistin resistance in Gram-negative bacteria. The propagated GO:0016776 corresponds to EC 2.7.4. enzymes (kinases that transfer inorganic phosphate groups), which is the wrong branch entirely. The correct term is GO:0016780 (phosphotransferase activity, for other substituted phosphate groups), which covers EC 2.7.8. enzymes. GO:0016776 and GO:0016780 are sibling terms under the common parent GO:0016772.
The single most decisive piece of evidence is the GO Consortium's official EC2GO mapping file, which maps EC 2.7.8. exclusively to GO:0016780 and EC 2.7.4. exclusively to GO:0016776, with zero cross-branch mappings. Since MCR-1 is unambiguously EC 2.7.8.43, the assigned GO:0016776 is definitively wrong. The error originates from a TAS mis-annotation on EptA (P30845) that was attached to the PANTHER PTHR30443:SF0 ancestral node and then propagated by TreeGrafter to MCR-1 and all other proteins grafted onto that node. Notably, the related enzyme EptB (P37661) in a different PANTHER subfamily (SF3) carries the correct annotation (GO:0043838 via IDA), demonstrating the error is confined to the SF0 node, not the tree topology.
This investigation systematically tested all three characteristic TreeGrafter failure modes. Failure mode #1 (granularity) does not apply — the subfamily placement of MCR-1 alongside EptA is correct. Failure mode #2 (pseudo-enzyme) was firmly excluded — 8 of 9 critical active-site residues are identical between MCR-1 and EptA, all validated as essential by alanine-scanning mutagenesis, and multiple crystal structures confirm intact zinc coordination. Failure mode #3 (within-superfamily mis-placement) is confirmed — the TreeGrafter propagated a sibling-branch GO term instead of the correct one.
Key Findings
Finding 1: GO:0016776 Is the Wrong Sibling Term — Correct Term Is GO:0016780
MCR-1 is assigned EC 2.7.8.43 (phosphoethanolamine—lipid A transferase). The GO hierarchy under GO:0016772 (transferase activity, transferring phosphorus-containing groups) branches into several children based on the chemical nature of the transferred/acceptor group:
GO:0016772 transferase activity, transferring phosphorus-containing groups
├── GO:0016776 phosphotransferase, phosphate group as acceptor [EC 2.7.4.*] ← SEED TERM ❌
├── GO:0016780 phosphotransferase, other substituted phosphate groups [EC 2.7.8.*] ← CORRECT ✅
│ └── GO:0043838 PEtN:Kdo₂-lipid A PEtN transferase [EC 2.7.8.42]
│ └── (no specific child for EC 2.7.8.43 yet)
└── ... (other siblings)
The GO Consortium's official EC2GO mapping file was retrieved and parsed programmatically. It confirmed that every EC 2.7.8. entry maps to GO:0016780 and every EC 2.7.4. entry maps to GO:0016776, with zero cross-branch mappings. Since MCR-1 is EC 2.7.8.43, the correct parent MF term is GO:0016780, not GO:0016776. The OLS (Ontology Lookup Service) verification confirmed the structural relationship: GO:0043838 is_a GO:0016780 is_a GO:0016772, while GO:0016776 is_a GO:0016772 — a different branch at the same level.
One could argue that GO:0016776 might apply because the PEtN is transferred to a phosphate group on lipid A. However, the EC/GO classification classifies by what is transferred (a substituted phosphate group → EC 2.7.8 → GO:0016780), not by the nature of the acceptor. The children of GO:0016776 (nucleoside diphosphate kinase, polyphosphate kinase, etc.) uniformly involve transfer of a simple phospho group, not a substituted phosphate.
{{figure:final_evidence_summary.png|caption=Consolidated 4-panel evidence visualization: GO hierarchy placement, active-site conservation, domain architecture comparison, and EC2GO mapping analysis supporting reclassification of MCR-1 from GO:0016776 to GO:0016780.}}
Finding 2: MCR-1 Is Catalytically Active — Pseudo-Enzyme Hypothesis Excluded
A critical question for any automated annotation is whether the protein retains catalytic activity or has degenerated into a pseudo-enzyme. For MCR-1, the evidence overwhelmingly supports active catalysis.
Active-site residue conservation: Pairwise Needleman-Wunsch alignment of MCR-1 against the characterized homolog EptA (P30845, E. coli K-12) revealed 33.1% overall identity (37.4% over aligned non-gap positions). Among the 9 critical active-site / metal-binding residues, 8 are identical: E246, T285 (the nucleophilic phosphothreonine intermediate), K333, H395, D465, H466, E468, and H478. Only position 329 differs (N in MCR-1 vs. D in EptA), a conservative Asn↔Asp substitution that does not impair catalysis.
Mutagenesis validation: Comprehensive alanine-scanning mutagenesis (PMID: 39608179) demonstrated that mutation of each of these 9 residues (plus 6 additional important positions) to alanine abolishes PEtN transfer to lipid A in vitro and nearly eliminates colistin resistance in vivo. This confirms the active site is functionally essential, not vestigial.
Structural evidence: Multiple high-resolution crystal structures of the MCR-1 catalytic domain (PDB: 5K4P, 5GOV, 5GRR, 5LRM) confirm zinc coordination at the active site, with T285 observed in both phosphorylated and unphosphorylated states corresponding to catalytic intermediates (PMID: 28000749; PMID: 27958270). The substrate analog co-crystal structure (PMID: 29079699) shows phosphatidylethanolamine bound near the active site, and mutation of substrate-binding residues impairs colistin resistance. The recent full-length structure (PMID: 41298376) demonstrates a two-state rotational mechanism bringing the periplasmic catalytic domain to membrane-embedded lipid A for catalysis.
Conclusion: MCR-1 has an intact, functional active site. Pseudo-enzyme status (failure mode #2) is definitively excluded.
Finding 3: PANTHER Subfamily Placement Is Correct — Error Is on the Node Annotation
MCR-1 (A0A0R6L508) and EptA (P30845) are both correctly assigned to PANTHER family PTHR30443 and subfamily PTHR30443:SF0 (Phosphoethanolamine transferase EptA). Their shared features include:
| Feature | MCR-1 | EptA |
|---|---|---|
| PANTHER family | PTHR30443 | PTHR30443 |
| PANTHER subfamily | SF0 | SF0 |
| Sequence identity | — | 37.4% pairwise |
| N-terminal domain | 5 TM helices (PF08019, IPR012549) | 5 TM helices (PF08019, IPR012549) |
| C-terminal domain | Alkaline phosphatase superfamily (PF00884) | Alkaline phosphatase superfamily (PF00884) |
| InterPro family | IPR040423 (PEtN transferase) | IPR040423 (PEtN transferase) |
| EC number | 2.7.8.43 | 2.7.8.– |
The TreeGrafter pipeline performed the phylogenetic grafting step correctly. The error occurred because the GO term annotated on the SF0 ancestral node was GO:0016776 (wrong sibling) rather than GO:0016780 (correct branch). This means the error is in the reference tree's node annotation, not in the grafting algorithm itself. The subfamily placement is correct — MCR-1 genuinely belongs to the PEtN transferase subfamily alongside EptA.
Finding 4: EC2GO Mapping Provides Definitive Confirmation
The GO Consortium maintains an authoritative mapping between EC numbers and GO terms at https://current.geneontology.org/ontology/external2go/ec2go. Programmatic retrieval and parsing of this file showed:
- EC:2.7.4.- → GO:0016776 (phosphotransferase activity, phosphate group as acceptor)
- EC:2.7.8.- → GO:0016780 (phosphotransferase activity, for other substituted phosphate groups)
- Zero EC 2.7.8.* entries map to GO:0016776
- Zero EC 2.7.4.* entries map to GO:0016780
EC 2.7.8.43 has no specific GO mapping in the file but inherits from its parent EC:2.7.8.-, which maps to GO:0016780. This mapping is maintained by the GO Consortium as the canonical bridge between enzyme classification and GO molecular function ontology and provides unambiguous, authoritative evidence that GO:0016776 is the wrong term for any EC 2.7.8.* enzyme.
Finding 5: Error Is Confined to PTHR30443:SF0 — EptB (SF3) Has Correct Annotation
An important control observation: EptB (P37661, EC 2.7.8.42), which is in a different PANTHER subfamily (PTHR30443:SF3), carries the correct GO:0043838 annotation via IDA evidence (EcoCyc, PMID: 15795227). This demonstrates:
- The PANTHER tree correctly separates EptA-type (lipid A PEtN modification) from EptB-type (Kdo₂-lipid A PEtN modification) into distinct subfamilies.
- The annotation error is localized to the SF0 node; SF3 has the correct term.
- The error likely originated from a TAS (Traceable Author Statement) mis-annotation on EptA (citing PMID:12184950, a conference abstract collection) that was incorporated into the PANTHER reference tree.
| Property | EptA (P30845) | EptB (P37661) | MCR-1 (A0A0R6L508) |
|---|---|---|---|
| Name | PEtN transferase EptA | Kdo₂-lipid A PEtN 7″-transferase | PEtN transferase MCR-1 |
| EC number | 2.7.-.- | 2.7.8.42 | 2.7.8.43 |
| Substrate modified | Lipid A (4′/1′ phosphate) | Kdo₂-lipid A (Kdo sugar) | Lipid A (4′/1′ phosphate) |
| PANTHER subfamily | PTHR30443:SF0 | PTHR30443:SF3 | PTHR30443:SF0 |
| GO MF (actual) | GO:0016776 (TAS) ❌ | GO:0043838 (IDA) ✅ | GO:0016776 (IEA) ❌ |
| GO MF (correct) | GO:0016780 | GO:0043838 | GO:0016780 |
Mechanistic Model / Interpretation
MCR-1 belongs to the alkaline phosphatase superfamily, specifically the phosphoethanolamine transferase (PEtN transferase) family. It is an integral inner-membrane enzyme with a topology consisting of 5 N-terminal transmembrane helices anchoring it in the membrane and a C-terminal periplasmic catalytic domain. The reaction it catalyzes is:
Phosphatidylethanolamine + Lipid A → Diacylglycerol + PEtN-Lipid A
The catalytic mechanism involves a phospho-enzyme intermediate: the nucleophilic Thr285 attacks the phosphoester bond in phosphatidylethanolamine, forming a phosphothreonine intermediate, which then transfers PEtN to the 4′-phosphate (or 1′-phosphate) of lipid A. This reaction requires two zinc ions at the active site for coordination and catalysis. The recent full-length structure (PMID: 41298376) reveals that the PE donor binds near the active site in the periplasmic domain, while lipid A binds more than 20 Å away within the transmembrane region, necessitating a conformational change (two-state rotational mechanism) to bring the catalytic center to the lipid A substrate.
This reaction — transfer of a substituted phosphate group (PEtN) from a donor to an acceptor — is classified as EC 2.7.8.43. The key distinction in GO classification is:
- EC 2.7.4 → GO:0016776: Transfer of a simple phosphate group from one phospho-compound to another (e.g., nucleoside diphosphate kinase, polyphosphate kinase)
- EC 2.7.8 → GO:0016780: Transfer of a substituted phosphate group (e.g., phosphoethanolamine, phosphatidyl, glycerophospho) to an acceptor
MCR-1 clearly falls into the latter category. The TreeGrafter error likely arose because the ancestral EptA annotation used a TAS reference (PMID:12184950, conference abstracts) that may have used imprecise language about "phosphotransferase" activity, and the curator selected the wrong sibling term. This error was then fossilized in the PANTHER tree and propagated automatically to all proteins grafted onto the SF0 node, including MCR-1.
{{figure:go_term_analysis.png|caption=GO hierarchy analysis showing the sibling relationship between GO:0016776 (phosphate group as acceptor) and GO:0016780 (other substituted phosphate groups), with active-site residue conservation between MCR-1 and EptA confirming catalytic competence.}}
Independent Family/Function Assignment
Based on this investigation, independent of the propagated term:
| Attribute | Assignment |
|---|---|
| Protein | MCR-1 (A0A0R6L508), 541 aa, Escherichia coli |
| Specific molecular function | Phosphoethanolamine—lipid A transferase |
| EC number | 2.7.8.43 |
| Closest characterized homolog | EptA (P30845, E. coli K-12), 37.4% identity, same PANTHER SF0 |
| Recommended GO term | GO:0016780 (phosphotransferase activity, for other substituted phosphate groups) |
| Granularity vs. seed term | Sibling — same level under GO:0016772, different branch |
| More specific GO term | GO:0043838 exists but is EptB-specific (Kdo₂-lipid A substrate); a new child term under GO:0016780 for lipid A PEtN transferase may be warranted |
Evidence Matrix
| # | Citation | Evidence Type | Relation | Claim Tested | Key Finding | Organism/Assay | Confidence |
|---|---|---|---|---|---|---|---|
| 1 | PMID: 26603172 | Direct assay, mutant phenotype | Supports MCR-1 as PEtN transferase | MCR-1 enzyme identity | MCR-1 is a plasmid-encoded PEtN transferase; mass spectrometry confirms PEtN addition to lipid A | E. coli SHP45, conjugation, mouse model | High — original discovery; direct biochemical |
| 2 | PMID: 29079699 | Structural, direct assay | Supports PEtN transferase; refutes GO:0016776 | Catalytic mechanism / substrate binding | "MCR-1 can catalyze the transfer of phosphoethanolamine (PEA) to lipid A" — PEtN transfer is EC 2.7.8, not EC 2.7.4; mutagenesis of active-site residues abolishes activity | E. coli recombinant, in vitro lipid A assay | High — structural + biochemical |
| 3 | PMID: 28000749 | Structural | Supports active enzyme (excludes pseudo-enzyme) | Active-site architecture | "Unphosphorylated nucleophilic residue Thr285 in coordination with two Zinc ions" — intact catalytic center | X-ray crystallography, 1.32 Å | High — direct structural |
| 4 | PMID: 27958270 | Structural | Supports active Zn-dependent catalysis | Phosphorylation states | T285 in phosphorylated/unphosphorylated states; Zn-binding site confirmed | Crystal structure | High |
| 5 | PMID: 41298376 | Structural, computational | Supports PEtN transferase mechanism | Full-length mechanism | Full MCR-1 structure: PE donor near active site, lipid A in TM region; two-state rotation | Cryo-EM + MD simulations | High — first full-length structure |
| 6 | PMID: 39608179 | Mutagenesis, direct assay | Supports active enzyme; identifies 15 essential residues | Residue essentiality | Alanine scanning identifies 15 indispensable residues; channel-shaped cavity for substrates | E. coli, alanine scanning | High — systematic mutagenesis |
| 7 | PMID: 39612773 | Review | Supports PEtN transferase classification | MCR family overview | Comprehensive review confirming MCR-1 as PEtN transferase catalyzing addition to lipid A | Review with AlphaFold analysis | Medium — review |
| 8 | PMID: 35079093 | Transcriptomics | Qualifies — EptA/EptB distinction | Relationship between mcr-1, eptA, eptB | EptB expression enhanced under colistin stress in mcr-1-harboring E. coli | E. coli clinical isolates | Medium — transcriptional evidence |
| 9 | GO EC2GO mapping | Database/computational | Refutes GO:0016776 definitively | EC→GO mapping for EC 2.7.8 | EC:2.7.4.→GO:0016776; EC:2.7.8.→GO:0016780; zero cross-branch mappings | GO Consortium canonical file | Definitive — authoritative source |
| 10 | UniProt A0A0R6L508 | Database/computational | Supports EC 2.7.8.43 | Enzyme classification | UniProt assigns EC 2.7.8.43; RHEA:46900; InterPro IPR040423 | Expert-curated entry | High |
| 11 | EptB (P37661) QuickGO | Database | Qualifies — correct annotation exists for related enzyme | Correct GO term used elsewhere? | EptB (SF3) has correct GO:0043838 via IDA; error confined to SF0 | E. coli K-12 EptB | High — confirms error is node-specific |
Active-Site / Placement Analysis
Active-Site Residue Conservation (MCR-1 vs. EptA)
Pairwise Needleman-Wunsch alignment (EMBOSS Needle, EBLOSUM62, gap open 10, gap extend 0.5) was performed computationally:
- Overall identity: 191/577 (33.1%)
- Similarity: 308/577 (53.4%)
- Gaps: 66/577 (11.4%)
| MCR-1 Position | MCR-1 Residue | EptA Position | EptA Residue | Conserved? | Functional Role | Mutagenesis Effect (→Ala) |
|---|---|---|---|---|---|---|
| 246 | Glu (E) | 244 | Glu (E) | IDENTICAL | Zinc binding | Abolishes activity |
| 285 | Thr (T) | 282 | Thr (T) | IDENTICAL | Nucleophilic; phosphorylated intermediate | Abolishes activity |
| 329 | Asn (N) | 326 | Asp (D) | Conservative (N↔D) | Catalytic / substrate binding | Abolishes activity |
| 333 | Lys (K) | 330 | Lys (K) | IDENTICAL | Substrate binding | Abolishes activity |
| 395 | His (H) | 385 | His (H) | IDENTICAL | Catalytic | Abolishes activity |
| 465 | Asp (D) | 455 | Asp (D) | IDENTICAL | Zinc binding | Abolishes activity |
| 466 | His (H) | 456 | His (H) | IDENTICAL | Zinc binding | Abolishes activity |
| 468 | Glu (E) | 458 | Glu (E) | IDENTICAL | Zinc coordination | Abolishes activity |
| 478 | His (H) | 468 | His (H) | IDENTICAL | Catalytic | Abolishes activity |
Result: 8/9 critical residues are identical (89% conservation). The single difference (N329 vs. D326) is a conservative substitution — both are small polar/charged residues with similar hydrogen-bonding capacity. All 9 positions have been validated by alanine-scanning mutagenesis: mutation to Ala at each position abolishes PEtN transfer to lipid A in vitro and nearly eliminates colistin resistance in vivo (PMID: 29079699; PMID: 39608179).
Catalytic residues are intact. MCR-1 is a fully active enzyme. Pseudo-enzyme status (failure mode #2) is definitively excluded.
Domain Architecture Comparison
| Feature | MCR-1 | EptA |
|---|---|---|
| Total length | 541 aa | 547 aa |
| N-terminal TM helices | 5 (residues ~15–178) | 5 (similar topology) |
| Periplasmic catalytic domain | ~362 aa (179–541) | ~365 aa |
| InterPro family | IPR040423 (PEtN transferase) | IPR040423 (PEtN transferase) |
| InterPro N-terminal domain | IPR012549 (EptA-like N) | IPR012549 (EptA-like N) |
| Superfamily | SSF53649 (Alkaline phosphatase-like) | SSF53649 (Alkaline phosphatase-like) |
| Pfam domains | PF08019 (EptA_B_N) + PF00884 (Sulfatase) | PF08019 + PF00884 |
The domain architecture is fully conserved, confirming both proteins belong to the same functional family. Subfamily placement is correct.
GO Curation Implications
Recommended curation action: REPLACE-WITH-SIBLING-TERM
| Current Annotation | Action | Replacement |
|---|---|---|
| GO:0016776 (IEA:TreeGrafter) | REPLACE | GO:0016780 (phosphotransferase activity, for other substituted phosphate groups) |
Rationale:
- MCR-1 is EC 2.7.8.43, which maps to GO:0016780, not GO:0016776, per the official EC2GO file.
- The GO ontology formally places PEtN transferase terms (GO:0043838) under GO:0016780.
- GO:0043838 itself is not fully appropriate for MCR-1 because it specifically describes EptB's activity (PEtN transfer to Kdo₂-lipid A), whereas MCR-1/EptA transfer PEtN to lipid A's 4′-phosphate — a distinct substrate.
- A new specific GO MF term for "phosphatidylethanolamine:lipid A phosphoethanolamine transferase activity" may be warranted for MCR-1/EptA-type enzymes.
Additional curation notes:
- This correction should be made at the PANTHER tree level (PTHR30443:SF0 node) to prevent continued propagation of the wrong term to newly sequenced MCR/EptA family members.
- EptA (P30845) also carries GO:0016776 via TAS (PMID:12184950, a conference abstract collection — a weak evidence source) and should be corrected simultaneously.
- The existing InterPro-derived annotation GO:0016772 (IEA:InterPro, from IPR040423) is correct at the broader level and should be retained.
Summary of Failure Mode Analysis
| Failure Mode | Status | Key Evidence |
|---|---|---|
| #1 Granularity / family-vs-subfamily | Not applicable | Error is a sibling term, not a granularity issue; subfamily placement SF0 is correct |
| #2 Pseudo-enzyme / loss of activity | Excluded | 8/9 catalytic residues identical to EptA; all validated essential by mutagenesis; multiple crystal structures confirm intact Zn coordination; direct biochemical activity demonstrated |
| #3 Within-superfamily mis-placement | CONFIRMED | GO:0016776 (EC 2.7.4.) is a sibling of correct GO:0016780 (EC 2.7.8.); both children of GO:0016772; PANTHER tree propagated wrong GO term from SF0 node |
Conflicts, Knowledge Gaps, and Discriminating Tests
Conflicts
-
EcoCyc TAS annotation on EptA: EptA (P30845) has GO:0016776 annotated via TAS (PMID:12184950), suggesting a human curator originally assigned this term. However, PMID:12184950 is a collection of conference abstracts (7th Conference of the International Endotoxin Society, 2002) with no accessible abstract — a weak evidence source. The GO ontology structure (GO:0043838 is_a GO:0016780, NOT is_a GO:0016776) overrides this assignment.
-
Semantic ambiguity in "phosphate group as acceptor": One could argue GO:0016776 applies because PEtN is transferred to a phosphate group on lipid A. However, the EC/GO classification system classifies by what is transferred (a substituted phosphate group → EC 2.7.8 → GO:0016780), not by the nature of the acceptor. All children of GO:0016776 involve transfer of a simple phospho group.
Knowledge Gaps
-
No specific GO MF term for MCR-1/EptA reaction: GO:0043838 covers EptB's reaction (PEtN to Kdo₂-lipid A), but no analogous term exists for MCR-1/EptA (PEtN to lipid A 4′-phosphate). Creating such a term would enable precise annotation.
-
Scope of propagation error: The same wrong GO:0016776 annotation likely appears on Neisseria EptA homologs (O34609, Q7DD94) and many other PEtN transferases in PTHR30443:SF0, all from TreeGrafter or IBA propagation.
-
MCR-2 through MCR-10 variants: Multiple MCR variants exist with varying degrees of colistin resistance. Whether all should receive the same GO term correction is not addressed here, though they likely all belong in SF0 and share the same error.
Discriminating Tests
-
EC2GO mapping verification (COMPLETED): The official file was downloaded and parsed, definitively confirming the branch mismatch. No further testing needed on the core question.
-
Experimental validation (NOT NEEDED): MCR-1's PEtN transferase activity has been confirmed by multiple independent assays. No additional experimental evidence is required for the GO term correction.
-
Full PANTHER tree audit (RECOMMENDED): Inspect all GO term annotations across PTHR30443 subfamily nodes to identify whether other subfamilies carry similar sibling-branch errors.
-
Broader EC2GO consistency check (RECOMMENDED): Systematically compare PANTHER-propagated GO terms against the official EC2GO mapping for all proteins with assigned EC numbers to identify other cases of sibling-term confusion.
-
GO term request (RECOMMENDED): Submit a GO GitHub issue requesting creation of "phosphatidylethanolamine:lipid A phosphoethanolamine transferase activity" as a child of GO:0016780 for EC 2.7.8.43, distinct from GO:0043838 (EptB-specific).
Evidence Base: Key Literature
-
Liu et al. (2016) — PMID: 26603172: Foundational paper discovering MCR-1 as a plasmid-mediated colistin resistance determinant and identifying it as a phosphoethanolamine transferase. Established the functional identity that underpins the entire GO term assessment.
-
Anandan et al. (2017) — PMID: 29079699: Crystal structure with substrate analog demonstrating "MCR-1 can catalyze the transfer of phosphoethanolamine (PEA) to lipid A, resulting in colistin resistance." Mutagenesis of active-site residues confirms functional essentiality. This paper directly supports the EC 2.7.8.43 classification.
-
Hu et al. (2016) — PMID: 28000749: High-resolution crystal structure showing "unphosphorylated nucleophilic residue Thr285 in coordination with two Zinc ions and water molecules" — confirming the intact zinc-dependent active site.
-
Stojanoski et al. (2016) — PMID: 27958270: Crystal structure showing alkaline phosphatase superfamily fold with T285 in two functional states, consistent with the phospho-enzyme catalytic intermediate.
-
Li et al. (2024) — PMID: 39608179: Systematic alanine-scanning mutagenesis identifying 15 critical residues, confirming the active site is not vestigial and MCR-1 is a bona fide enzyme.
-
Liu et al. (2025) — PMID: 41298376: First full-length MCR-1 structure demonstrating the two-state rotational mechanism with PE donor and lipid A substrates bound in distinct locations.
-
Zhang et al. (2024) — PMID: 39612773: Comprehensive review confirming the PEtN transferase classification and analyzing catalytic mechanism using full-length AlphaFold models.
Report generated through systematic evaluation of TreeGrafter annotation GO:0016776 for MCR-1 (A0A0R6L508). Three characteristic failure modes of phylogenetic annotation propagation were tested; failure mode #3 (within-superfamily sibling-term mis-placement) was identified as the cause of the incorrect annotation. All computational analyses (sequence alignment, EC2GO parsing, ontology hierarchy verification) were performed programmatically using public databases and published literature.