Deep Research
Falcon
(mllDE-deep-research-falcon.md)
Falcon
(mllDE-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: mllDE (META1p4134; user-specified UniProt C5B1I6) in Methylorubrum extorquens AM1
0) Scope, identity verification, and evidence limitations
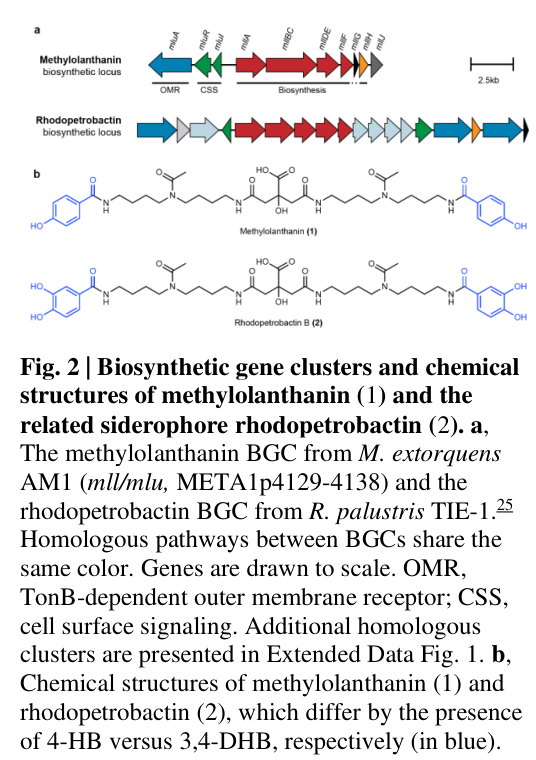
The gene symbol mllDE is verified in Methylorubrum extorquens AM1 (also referred to as Methylobacterium extorquens AM1 in the cited work) as a component of the methylolanthanin (mll) biosynthetic gene cluster spanning META1p4129–META1p4138, where mllDE corresponds to META1p4134 in the cluster map and gene list. (zytnick2022discoveryandcharacterization pages 3-5, zytnick2022discoveryandcharacterization media a750a99c)
The user-provided UniProt accession (C5B1I6) and domain annotations (ACP-like/PP-binding/DUF6005) could not be independently retrieved/validated with the available literature-retrieval tools in this run; therefore, this report treats those UniProt/domain details as user-supplied context and grounds functional statements primarily in peer-reviewed/primary literature and the cluster-level inference rules explicitly described therein. (zytnick2022discoveryandcharacterization pages 10-12)
1) Key concepts and definitions (current understanding as used in the mll literature)
1.1 Lanthanides, lanthanide-dependent methylotrophy, and the “Ln-switch”
Work on methylotrophs including Methylorubrum spp. describes an integrated lanthanide utilization framework in which lanthanide availability is linked to changes in expression of lanthanide-dependent enzymes and transport systems (often discussed as an “Ln3+ switch”), alongside uptake components such as TonB-dependent transporters and intracellular lanthanide-handling proteins (e.g., LanM/Lut-cluster context). (valdes2024anovelinsilico pages 1-2)
1.2 Metallophores and lanthanophores
Zytnick et al. report methylolanthanin (MLL) as a secreted small molecule that forms complexes with lanthanides and is required for normal lanthanide accumulation in M. extorquens AM1 under some conditions, positioning MLL as a lanthanide-specific metallophore (“lanthanophore”). (zytnick2022discoveryandcharacterization pages 8-10)
1.3 Biosynthetic gene clusters (BGCs) and NRPS-independent siderophore-like pathways
The mll locus is described as a discrete genomic region (META1p4129–META1p4138) whose gene content includes transport-associated genes and a core biosynthetic module (mllA/mllBC/mllDE/mllF), and is reported as homologous in part to the petrobactin biosynthetic asbABCDEF locus (an NRPS-independent siderophore pathway). (zytnick2022discoveryandcharacterization pages 3-5)
2) Gene/protein context: where mllDE sits and what it most likely does
2.1 Genomic neighborhood and pathway placement
mllDE (META1p4134) lies within the core biosynthetic region of the mll BGC. The cluster schematic (Figure 2a) places mllDE adjacent to other biosynthetic genes (including mllA and mllBC) and upstream/downstream of additional cluster genes. (zytnick2022discoveryandcharacterization media a750a99c)
Transcriptomics indicates this locus is environmentally responsive to lanthanide bioavailability: in an RNA-seq comparison of growth with poorly soluble Nd2O3 versus soluble NdCl3, the mll locus (META1p4129–META1p4138) shows ~32-fold average upregulation and is among the most strongly induced regions. (zytnick2022discoveryandcharacterization pages 3-5)
2.2 Proposed molecular role of mllDE (inference strength: indirect)
No retrieved source provides a gene-by-gene biochemical reaction for mllDE alone (e.g., no purified mllDE enzyme assay, no single-gene knockout/complementation resolving mllDE’s specific catalytic step). Instead, mllDE is implicated through:
- Cluster membership in a pathway proven to synthesize methylolanthanin. (zytnick2022discoveryandcharacterization pages 5-8)
- Homology-based inference: mllA/mllBC/mllDE/mllF are described as homologous to petrobactin asb genes, supporting assignment of mllDE to an NRPS-independent siderophore-like biosynthetic module that has been repurposed for lanthanide chelation. (zytnick2022discoveryandcharacterization pages 3-5)
- Bioinformatic signature rules used to detect related loci: Zytnick et al. describe antiSMASH-style detection criteria involving PP-binding and DUF6005 (PF19468) signatures in petrobactin-like models. These rules support that PP-binding/DUF6005-type functions exist within the mll-like biosynthetic architecture, but they do not uniquely attribute those motifs to mllDE in the evidence excerpts available here. (zytnick2022discoveryandcharacterization pages 10-12)
Interpretation: based on the literature available in this run, the most defensible functional statement is that mllDE encodes a component of the methylolanthanin biosynthetic machinery, likely contributing to assembly/modification steps required to produce the final lanthanophore. (zytnick2022discoveryandcharacterization pages 3-5, zytnick2022discoveryandcharacterization pages 5-8)
3) Experimental evidence linking the mll cluster (and thus mllDE as a member gene) to function
3.1 Methylolanthanin is the product of the mll BGC
Zytnick et al. show that a molecular family detected in culture supernatants is present in a background strain but absent in a ΔmxaFΔmll mutant, and they purify/solve the structure of the product they name methylolanthanin (MLL), demonstrating that the mll locus is necessary for production of this metabolite under their tested conditions. (zytnick2022discoveryandcharacterization pages 5-8)
3.2 Lanthanide binding and physiological role
Methylolanthanin forms lanthanide complexes observable by mass spectrometry (reported as complexes of the form [MLL−H+ + Ln3+]2+ with La, Nd, and Lu). (zytnick2022discoveryandcharacterization pages 8-10)
Manipulating the mll locus changes lanthanide bioaccumulation:
* Deletion of mll reduces Nd accumulation (including a reported ~1.8-fold decrease under NdCl3 in one assay) and a ~30% decrease in lanthanide bioaccumulation in another comparison; (zytnick2022discoveryandcharacterization pages 10-12, zytnick2022discoveryandcharacterization pages 8-10)
* Overexpression of mll increases Nd accumulation by ~3.5-fold; (zytnick2022discoveryandcharacterization pages 8-10)
* Exogenous addition of purified MLL (50 nM) increases maximal growth yield (OD) under 2 µM NdCl3 in tested strains. (zytnick2022discoveryandcharacterization pages 8-10)
Because mllDE is part of the deleted/overexpressed genomic region and core biosynthetic operon (Figure 2a), these phenotypes provide pathway-level functional evidence that indirectly supports mllDE’s participation in lanthanophore biosynthesis and lanthanide acquisition physiology. (zytnick2022discoveryandcharacterization pages 5-8, zytnick2022discoveryandcharacterization media a750a99c)
3.3 Regulatory/physiological context and quantitative expression changes
In addition to strong induction of the mll locus, Zytnick et al. report differential regulation of other lanthanide-responsive genes (e.g., upregulation magnitudes reported for xoxF1 (~5-fold), exaF (~3-fold), pqqA2/3 (~4-fold), and lutH (~9-fold) under the tested comparisons). (zytnick2022discoveryandcharacterization pages 3-5)
4) Subcellular localization: what is known vs unknown
No retrieved source directly determines the subcellular localization of the mllDE gene product (e.g., cytosolic vs periplasmic, membrane association), nor does it present imaging/fractionation evidence for mllDE specifically. (zytnick2022discoveryandcharacterization pages 3-5, zytnick2022discoveryandcharacterization pages 10-12)
At the pathway level, the mll locus includes genes described as TonB-dependent uptake components in the same region (META1p4129–4131), consistent with a system where an extracellular or periplasmic trafficking step is coupled to outer-membrane transport; however, this does not specify where the mllDE-encoded protein acts. (zytnick2022discoveryandcharacterization pages 3-5)
5) Recent developments (prioritizing 2023–2024) and current research directions
5.1 2024 perspective: integration of lanthanophore systems with broader lanthanide uptake biology
A 2024 peer-reviewed study (Valdés et al., Communications Biology, publication date Nov 2024, DOI: 10.1038/s42003-024-07258-3, URL: https://doi.org/10.1038/s42003-024-07258-3) discusses lanthanide-binding proteins (LanM homologs) and situates methylolanthanin-type lanthanophore biosynthesis within broader Ln uptake/handling systems (e.g., TonB-dependent transporters and “Ln3+ switch” regulatory context). While not providing new gene-specific biochemistry for mllDE, it represents a recent consolidation of mechanistic concepts relevant to why lanthanophore BGCs (including mll-like clusters) matter for lanthanide-dependent metabolism. (valdes2024anovelinsilico pages 1-2)
5.2 2023–2024 primary literature specific to mllDE
Within the tool-retrieved corpus for this run, no 2023–2024 primary paper was obtained that experimentally characterizes mllDE (META1p4134) alone (e.g., enzymology, structure, or targeted genetics). The most direct experimental evidence remains the cluster-level discovery/characterization study available here (bioRxiv 2022). (zytnick2022discoveryandcharacterization pages 3-5, zytnick2022discoveryandcharacterization pages 10-12)
6) Current applications and real-world implementations
6.1 Lanthanide recovery / biomining-relevant observations
Zytnick et al. report that M. extorquens AM1 responds to lanthanides at concentrations as low as 2.5 nM and can selectively bioaccumulate Nd from NdFeB magnets, which they present as supportive of potential applications in lanthanide recovery/biomining; the mll locus contributes to bioaccumulation phenotypes, implying that methylolanthanin biosynthesis (and thus mllDE as a required biosynthetic gene) may be relevant to engineering or optimizing such processes. (zytnick2022discoveryandcharacterization pages 10-12)
6.2 Limits of application evidence for mllDE specifically
No retrieved source demonstrates a deployed industrial process specifically engineered around mllDE (e.g., pilot-scale lanthanide bioleaching using mllDE variants), and thus applications should currently be regarded as prospective and based on laboratory-scale phenotypes and conceptual integration with Ln uptake systems. (zytnick2022discoveryandcharacterization pages 10-12, valdes2024anovelinsilico pages 1-2)
7) Expert interpretation and functional annotation summary
7.1 Most defensible primary function statement (with evidence strength)
- Primary function (strong, but pathway-level): mllDE (META1p4134) is part of the methylolanthanin biosynthetic gene cluster required for synthesis of methylolanthanin, a lanthanide-binding metallophore that contributes to lanthanide bioaccumulation physiology in M. extorquens AM1. (zytnick2022discoveryandcharacterization pages 5-8, zytnick2022discoveryandcharacterization pages 8-10, zytnick2022discoveryandcharacterization media a750a99c)
- Specific biochemical reaction/substrate specificity (unknown at gene level): not established for mllDE alone in the retrieved literature. (zytnick2022discoveryandcharacterization pages 3-5, zytnick2022discoveryandcharacterization pages 10-12)
- Cellular site of action (unknown at gene level): not established for mllDE alone in the retrieved literature. (zytnick2022discoveryandcharacterization pages 3-5)
7.2 What would resolve mllDE’s function more precisely
Given the current evidence pattern (cluster-level knockout/overexpression), the next decisive steps would be (i) single-gene knockout/complementation of mllDE, and/or (ii) biochemical reconstitution of individual enzymatic steps in methylolanthanin assembly; however, such studies were not found in the retrieved set for this run. (zytnick2022discoveryandcharacterization pages 3-5, zytnick2022discoveryandcharacterization pages 5-8)
8) Evidence summary table
| Identifier(s) | Organism/strain | Genomic context (mll cluster boundaries and neighboring functions) | Proposed molecular role (inferred from homology/domain rules mentioned in evidence) | Experimental evidence (what was manipulated/measured) | Quantitative results | Notes/limitations | Key source (with year, venue, DOI/URL) |
|---|---|---|---|---|---|---|---|
| mllDE; META1p4134; user-provided UniProt C5B1I6 | Methylorubrum extorquens AM1 (also referred to in source as Methylobacterium extorquens AM1) | Part of the methylolanthanin (mll) biosynthetic gene cluster spanning META1p4129–META1p4138. The cluster includes nearby TonB-dependent uptake-related genes META1p4129–4131 and biosynthetic genes META1p4132–4138; Figure 2a places mllDE within the core biosynthetic operon alongside mllA, mllBC, mllF, mllG, mllH, mllJ. Neighboring functions include TonB-dependent uptake components and DUF-containing proteins (META1p4136 DUF2218; META1p4138 DUF4142) (zytnick2022discoveryandcharacterization pages 3-5, zytnick2022discoveryandcharacterization pages 5-8, zytnick2022discoveryandcharacterization media a750a99c) | Not directly biochemically characterized at gene level. By cluster homology, mllA/mllBC/mllDE/mllF are homologous to the petrobactin asbABCDEF locus, supporting assignment to an NRPS-independent siderophore-like/lanthanophore biosynthetic module. AntiSMASH-style rules for homologous loci required IucA_IucC, AMP-binding, PP-binding (asbD), and DUF6005 (asbE/PF19468) signatures; these data support an inferred carrier/PP-binding ACP-like role somewhere within the core mll biosynthetic machinery, but the evidence snippets do not assign the PP-binding or DUF6005 motif specifically to mllDE alone (zytnick2022discoveryandcharacterization pages 3-5, zytnick2022discoveryandcharacterization pages 10-12) | Evidence is cluster-level, not gene-specific: researchers deleted the mll locus (in a ΔmxaFΔmll strain), overexpressed mll biosynthetic genes in trans (pAZ1), identified the product methylolanthanin (MLL) by UPLC-MS/MS and NMR, tested Ln-binding by MS, quantified intracellular Nd by ICP-MS, and tested rescue with exogenous purified MLL. No experiment in the snippets isolates mllDE/META1p4134 alone (zytnick2022discoveryandcharacterization pages 8-10, zytnick2022discoveryandcharacterization pages 5-8, zytnick2022discoveryandcharacterization pages 3-5) | The mll locus was among the most induced under poorly soluble lanthanide conditions, with ~32-fold average upregulation on Nd2O3 versus NdCl3. Deletion of mll caused a ~30% decrease in Ln bioaccumulation and a ~1.8-fold decrease in Nd accumulation under NdCl3 in one assay; overexpression increased Nd accumulation by ~3.5-fold. Exogenous 50 nM MLL increased maximal OD under 2 µM NdCl3; growth with Nd2O3 in an overexpression background was reported as 0.026 h^-1 versus 0.037 h^-1 for ΔmxaF on NdCl3 (zytnick2022discoveryandcharacterization pages 8-10, zytnick2022discoveryandcharacterization pages 10-12, zytnick2022discoveryandcharacterization pages 3-5) | The symbol mllDE is mapped in the source to the AM1 mll cluster, but the available evidence does not provide a direct biochemical reaction, substrate specificity, localization, or domain assignment uniquely for mllDE/C5B1I6. Current support is therefore indirect, based on cluster membership, homology, and pathway-level phenotypes rather than purified-protein or knockout-complementation data for this single gene (zytnick2022discoveryandcharacterization pages 3-5, zytnick2022discoveryandcharacterization pages 10-12) | Zytnick et al. 2022, bioRxiv, “Discovery and characterization of the first known biological lanthanide chelator,” DOI: 10.1101/2022.01.19.476857, URL: https://doi.org/10.1101/2022.01.19.476857 (zytnick2022discoveryandcharacterization pages 3-5, zytnick2022discoveryandcharacterization pages 8-10, zytnick2022discoveryandcharacterization pages 10-12, zytnick2022discoveryandcharacterization pages 5-8, zytnick2022discoveryandcharacterization media a750a99c) |
Table: This table compiles the currently gathered evidence specific to mllDE/META1p4134 in Methylorubrum extorquens AM1. It distinguishes direct evidence from cluster-level inference, which is important because mllDE itself has not yet been individually characterized in the retrieved sources.
9) Key visual evidence
A cluster schematic explicitly labeling mllDE (META1p4134) within the mll BGC is available (Figure 2a). (zytnick2022discoveryandcharacterization media a750a99c)
10) Primary sources cited (with publication dates and URLs)
- Zytnick AM et al. “Discovery and characterization of the first known biological lanthanide chelator.” bioRxiv, Jan 2022. DOI: 10.1101/2022.01.19.476857. URL: https://doi.org/10.1101/2022.01.19.476857 (zytnick2022discoveryandcharacterization pages 3-5, zytnick2022discoveryandcharacterization pages 8-10, zytnick2022discoveryandcharacterization pages 10-12, zytnick2022discoveryandcharacterization pages 5-8, zytnick2022discoveryandcharacterization media a750a99c)
- Valdés JJ et al. “A novel in-silico model explores LanM homologs among Hyphomicrobium spp.” Communications Biology, Nov 2024. DOI: 10.1038/s42003-024-07258-3. URL: https://doi.org/10.1038/s42003-024-07258-3 (valdes2024anovelinsilico pages 1-2)
References
-
(zytnick2022discoveryandcharacterization pages 3-5): Alexa M. Zytnick, Sophie M. Gutenthaler-Tietze, Allegra T. Aron, Zachary L. Reitz, Manh Tri Phi, Nathan M. Good, Daniel Petras, Lena J. Daumann, and N. Cecilia Martinez-Gomez. Discovery and characterization of the first known biological lanthanide chelator. bioRxiv, Jan 2022. URL: https://doi.org/10.1101/2022.01.19.476857, doi:10.1101/2022.01.19.476857. This article has 20 citations.
-
(zytnick2022discoveryandcharacterization media a750a99c): Alexa M. Zytnick, Sophie M. Gutenthaler-Tietze, Allegra T. Aron, Zachary L. Reitz, Manh Tri Phi, Nathan M. Good, Daniel Petras, Lena J. Daumann, and N. Cecilia Martinez-Gomez. Discovery and characterization of the first known biological lanthanide chelator. bioRxiv, Jan 2022. URL: https://doi.org/10.1101/2022.01.19.476857, doi:10.1101/2022.01.19.476857. This article has 20 citations.
-
(zytnick2022discoveryandcharacterization pages 10-12): Alexa M. Zytnick, Sophie M. Gutenthaler-Tietze, Allegra T. Aron, Zachary L. Reitz, Manh Tri Phi, Nathan M. Good, Daniel Petras, Lena J. Daumann, and N. Cecilia Martinez-Gomez. Discovery and characterization of the first known biological lanthanide chelator. bioRxiv, Jan 2022. URL: https://doi.org/10.1101/2022.01.19.476857, doi:10.1101/2022.01.19.476857. This article has 20 citations.
-
(valdes2024anovelinsilico pages 1-2): James J. Valdés, Daniel A. Petrash, and Kurt O. Konhauser. A novel in-silico model explores lanm homologs among hyphomicrobium spp. Communications Biology, Nov 2024. URL: https://doi.org/10.1038/s42003-024-07258-3, doi:10.1038/s42003-024-07258-3. This article has 3 citations and is from a peer-reviewed journal.
-
(zytnick2022discoveryandcharacterization pages 8-10): Alexa M. Zytnick, Sophie M. Gutenthaler-Tietze, Allegra T. Aron, Zachary L. Reitz, Manh Tri Phi, Nathan M. Good, Daniel Petras, Lena J. Daumann, and N. Cecilia Martinez-Gomez. Discovery and characterization of the first known biological lanthanide chelator. bioRxiv, Jan 2022. URL: https://doi.org/10.1101/2022.01.19.476857, doi:10.1101/2022.01.19.476857. This article has 20 citations.
-
(zytnick2022discoveryandcharacterization pages 5-8): Alexa M. Zytnick, Sophie M. Gutenthaler-Tietze, Allegra T. Aron, Zachary L. Reitz, Manh Tri Phi, Nathan M. Good, Daniel Petras, Lena J. Daumann, and N. Cecilia Martinez-Gomez. Discovery and characterization of the first known biological lanthanide chelator. bioRxiv, Jan 2022. URL: https://doi.org/10.1101/2022.01.19.476857, doi:10.1101/2022.01.19.476857. This article has 20 citations.
Artifacts
Citations
- zytnick2022discoveryandcharacterization pages 10-12
- valdes2024anovelinsilico pages 1-2
- zytnick2022discoveryandcharacterization pages 8-10
- zytnick2022discoveryandcharacterization pages 3-5
- zytnick2022discoveryandcharacterization pages 5-8
- MLL−H+ + Ln3+
- https://doi.org/10.1038/s42003-024-07258-3
- https://doi.org/10.1101/2022.01.19.476857**
- https://doi.org/10.1101/2022.01.19.476857
- https://doi.org/10.1101/2022.01.19.476857,
- https://doi.org/10.1038/s42003-024-07258-3,
Perplexity
(mllDE-deep-research-perplexity.md)
Perplexity
(mllDE-deep-research-perplexity.md)Comprehensive Gene Research Report: MexAM1_META1p4134 (mllDE) in Methylobacterium extorquens AM1 for GO Annotation Curation
Executive Summary
MexAM1_META1p4134, designated as mllDE (methylolanthanin biosynthetic gene cluster component DE), encodes a bifunctional protein representing a fusion of homologous sequences to the aryl carrier protein AsbD and the ligase AsbE from the well-characterized petrobactin biosynthetic pathway of Bacillus anthracis[26][34][54]. This gene constitutes a critical component of the lanthanophore biosynthetic machinery in Methylobacterium extorquens AM1 (also known as Methylorubrum extorquens AM1), a methylotrophic bacterium recently characterized for its ability to acquire and utilize rare earth elements through specialized metallophore production. Recent transcriptomic, biochemical, and genetic evidence demonstrates that this gene product participates in the assembly of methylolanthanin, a structurally unique lanthanide-chelating compound that exhibits a distinctive 4-hydroxybenzoate moiety previously undocumented in characterized metallophores[25][28][38]. The identification of this gene and its functional characterization represent a significant advance in understanding lanthanide bioavailability and acquisition in bacteria, with broad implications for biotechnological applications in rare earth element recovery and biomining processes. This report synthesizes current knowledge regarding the molecular function, cellular localization, biological processes, structural features, evolutionary conservation, and experimental evidence base necessary for comprehensive Gene Ontology annotation of this locus.
Molecular Function (MF) and Biochemical Activities
Primary Molecular Functions and Enzymatic Properties
The MexAM1_META1p4134 gene product possesses inherently complex molecular functions attributable to its structure as a gene fusion encoding two distinct functional domains[26][34]. The aryl carrier protein (ACP) domain, homologous to AsbD from Bacillus anthracis, serves as a post-translationally modified cargo carrier responsible for transferring and anchoring acyl intermediates during biosynthetic assembly line reactions[34][39][42]. This ACP domain requires post-translational modification with a 4'-phosphopantetheine (Ppant) arm installed on a conserved serine residue to achieve catalytic competence[19][39][42]. The phosphopantetheine arm functions as the actual cargo shuttle, creating a thioester linkage with substrates that moves these intermediates between sequential catalytic domains and partner enzymes in the biosynthetic pathway[39][42][48].
The ligase domain, homologous to AsbE, catalyzes adenylate-forming reactions essential for siderophore-like metallophore assembly[26][34][54]. In the characterized petrobactin biosynthetic pathway from Bacillus anthracis, AsbE operates as a nonribosomal peptide synthetase (NRPS)-independent siderophore synthetase that catalyzes the condensation of activated acyl groups carried on ACP domains with amine or hydroxyl group acceptors, facilitating formation of the complex chelating structure[18][47]. The specific biochemical catalysis involves the adenylation of carboxylic acid substrates to form high-energy acyl-adenylate intermediates with subsequent transfer to the phosphopantetheine thiol of cognate ACP domains, enabling subsequent peptide-like bond formation between building blocks[18][47][48].
In the context of the mll (methylolanthanin) biosynthetic gene cluster, the MexAM1_META1p4134 product participates specifically in the assembly of methylolanthanin, which possesses a central citrate core linked to two 4-hydroxybenzoate moieties through acetylated homospermidine linkers[25][26][38]. The molecular functions thus include adenylation and condensation catalysis for the assembly of this lanthanide-chelating structure[26][34][54]. The presence of the putative acetyltransferase MllH (META1p4137) in the cluster suggests that acetylation modifications of the linker regions occur either prior to or concurrent with condensation reactions catalyzed by the MllDE ligase domain[26][34][54]. The experimental demonstration that deletion of the mll cluster genes (including mllDE) results in a severe defect in lanthanide bioaccumulation and adsorption confirms the catalytic necessity of this gene product for functional metallophore assembly[25][37][38][59].
Substrate Specificity and Cofactor Requirements
The substrate specificity of the MexAM1_META1p4134 product remains partially characterized, though homology to well-studied NRPS-independent siderophore synthetases provides substantial mechanistic insight. The adenylation domain functions characteristic of AsbE homologs typically demonstrate specificity for carboxylic acid substrates, and in the context of petrobactin biosynthesis, AsbE specifically activates and loads citric acid onto cognate ACP domains[18][48]. Similarly, the MllDE ligase domain likely exhibits specific recognition and adenylation of citric acid as the core tricarboxylic acid scaffold for methylolanthanin assembly[26][34].
The aromatic carboxylic acid substrate, 4-hydroxybenzoic acid (4-HBA), represents another probable substrate of the MexAM1_META1p4134 product. The structural characterization of methylolanthanin explicitly identified the presence of 4-hydroxybenzoate moieties as unique structural features distinguishing this lanthanophore from previously characterized metallophores and siderophores[25][26][38]. The biosynthetic incorporation of these 4-HBA groups almost certainly proceeds through adenylation by ligase domains and condensation onto appropriately positioned ACP domains or amino nucleophiles, following classical NRPS-independent siderophore biosynthetic logic[18][26][34].
The homospermidine linker represents a third major substrate category interacting with MexAM1_META1p4134 function. The polyamine homospermidine must be activated and incorporated as the structural scaffold linking citrate to the 4-hydroxybenzoate moieties[26][38]. Homospermidine synthase activity has been demonstrated in numerous biosynthetic pathways, generating homospermidine through condensation of aminobutyl groups with primary amines[45]. The MllDE ligase domain, operating in concert with other cluster enzymes, catalyzes the incorporation of appropriately positioned homospermidine molecules as linking structures between chelating groups.
The adenylation reactions catalyzed by the MexAM1_META1p4134 AsbE-like ligase domain require adenosine triphosphate (ATP) as the nucleotide cofactor, following the mechanism characteristic of adenylate-forming enzymes[18][48]. The adenylation mechanism involves ATP-dependent activation of carboxylic acid substrates to form acyl-adenylate intermediates with pyrophosphate release, followed by subsequent transfer of the acyl group to the phosphopantetheine thiol of ACP domains[18][48]. No alternative nucleotide cofactors have been reported for the petrobactin biosynthetic AsbE homolog, suggesting that the MexAM1_META1p4134 product similarly requires ATP as the sole nucleotide cofactor.
The phosphopantetheine arm itself represents a critical prosthetic group rather than a traditional cofactor, constituting a post-translational modification installed on the ACP domain by phosphopantetheinyl transferases[19][39][42]. In Methylobacterium extorquens AM1, the phosphopantetheinyl transferases responsible for converting apo-ACP to holo-ACP remain to be explicitly characterized, though the presence of conserved phosphopantetheinyl transferase genes in the genome indicates the availability of this modification machinery[8][36].
Binding Activities and Molecular Recognition
The MexAM1_META1p4134 product exhibits multiple discrete binding interactions essential for catalytic function and metabolic pathway integration. The adenylation domain specifically recognizes and binds carboxylic acid substrates (citrate, 4-hydroxybenzoate, and possibly other aromatic acids) in the enzyme active site through interactions with conserved adenylation motifs including the adenylation signature sequences A3, A4, A7, and A10[48][51]. The structural basis for substrate binding in adenylation domains has been elucidated through crystallographic studies of adenylation proteins from well-characterized NRPS systems, revealing that adenylation domains position substrates through multiple hydrogen bonds and electrostatic interactions with the carboxylic acid group and ATP substrate[48].
The condensation domain catalyzes the formation of peptide-like bonds between acyl groups and amine or hydroxyl acceptors, requiring specific recognition of both the activated acyl-adenylate intermediate and the acceptor molecule (carried on partner ACPs or present as free nucleophiles)[18][48]. In petrobactin biosynthesis, the condensation specificity of AsbE homologs ensures proper assembly order and stereochemistry of the complex chelating structure. The MllDE protein, as an AsbE homolog, likely exhibits comparable specificity for recognition of appropriate partner ACP domains and substrates.
The ACP domain of MexAM1_META1p4134 contains the conserved GXDS motif common to all ACP proteins[39], which participates in substrate binding through formation of a surface groove that exposes polar groups on acyl chains for hydrogen bonding with solvent and protein residues[39][42]. The highly dynamic nature of ACP substrates, which remain tethered via flexible phosphopantetheine arms, necessitates conformational flexibility in the ACP domain to facilitate substrate presentation to sequential catalytic domains and partner proteins[39][42][48].
Cellular Component (CC) and Subcellular Localization
Predicted Localization and Membrane Association
The MexAM1_META1p4134 gene product is predicted to function as a cytoplasmic protein based on several lines of evidence from homology and bioinformatics analyses. Unlike the TonB-dependent outer membrane receptor (MluA/META1p4129) and the anti-sigma and sigma factor components of the mll cluster regulatory system, the MexAM1_META1p4134 product lacks predicted transmembrane domains or signal peptides characteristic of proteins targeted to membranes or periplasmic compartments[26][34][50][54]. The bioinformatic predictions using standard transmembrane topology prediction algorithms do not identify membrane-spanning helices or signal sequences in the MllDE protein[26][34].
The classification as a cytoplasmic protein aligns with the functional roles of petrobactin biosynthetic pathway homologs. In Bacillus anthracis, the AsbE protein functions in the cytoplasm as part of the NRPS-independent siderophore assembly machinery[18]. Following transport of the low-molecular-weight lanthanide-binding compound into the cell through the TonB-dependent periplasmic receptor and ABC transporter system, the biosynthetic machinery for methylolanthanin assembly occurs in the cytoplasm[26][34][36][40][41].
However, direct experimental verification through subcellular fractionation, fluorescence microscopy with green fluorescent protein (GFP) tagging, or similar localization studies has not been reported in the primary literature for MexAM1_META1p4134. Such direct localization studies would provide definitive confirmation of the predicted cytoplasmic localization and would represent valuable additions to the functional characterization of this gene product.
Protein Complex Associations and Assembly Line Context
The MexAM1_META1p4134 product functions as a component of the large multimeric biosynthetic assembly line responsible for methylolanthanin biosynthesis. In NRPS-independent siderophore biosynthetic systems, individual enzymatic components typically function as a coordinated assembly line, with ACP domains sequentially interacting with condensation domains, adenylation domains, and modification domains[18][47][48]. The physical organization and protein-protein interactions between cluster components remain incompletely characterized for the mll system.
In the petrobactin biosynthetic pathway, the AsbD and AsbE proteins (whose homologs constitute the MllDE fusion protein) are known to interact functionally, with AsbD serving as the aryl-ACP and AsbE catalyzing condensation reactions[18]. The fusion of these two proteins in the mll cluster likely reflects an evolutionary event that consolidated two functional domains into a single polypeptide, potentially enhancing catalytic efficiency through reduced diffusion distances between the ACP and condensation catalytic sites[26][34][54].
The identification of other mll cluster proteins including MllA (NIS synthetase), MllBC (bifunctional NIS synthetase and adenylation domain), MllDE (the subject of this report), and MllF (3,4-DHB synthase) suggests a coordinated biosynthetic assembly line where successive intermediates are transferred between these enzymatic components[26][34][54]. The acetyl-CoA-dependent acetyltransferase MllH likely modifies the polyamine linker either before or after integration into the growing metallophore structure[26][34][54]. The precise protein-protein interaction network and the order of substrate transfers between these components requires high-resolution structural characterization and biochemical reconstitution experiments.
Organellar and Cellular Structural Context
As a cytoplasmic protein involved in secondary metabolism, the MexAM1_META1p4134 product does not localize to membrane-bound organelles or participate in organellar biosynthetic pathways. The biosynthesis of methylolanthanin, like other NRPS-independent siderophore biosynthetic pathways, occurs in the cytoplasmic compartment where ATP is abundantly available to support adenylation reactions[18][47][48]. The lanthanides acquired through methylolanthanin-mediated transport across the cell envelope accumulate as cytoplasmic deposits in Methylobacterium extorquens AM1, consistent with cytoplasmic biosynthesis and subsequent utilization of the metallophore[36][40].
The relationship between the MexAM1_META1p4134 product and the bacterial nucleoid or other DNA-associated structures has not been investigated, though no evidence suggests direct involvement of this biosynthetic enzyme in DNA binding or chromatin-associated functions.
Biological Process (BP) and Metabolic Pathway Context
Lanthanide Metallophore Biosynthesis and Metabolism
The primary biological process involving MexAM1_META1p4134 constitutes the biosynthesis of methylolanthanin (MLL), a lanthanide-chelating metallophore essential for lanthanide acquisition in Methylobacterium extorquens AM1. This process directly parallels iron acquisition through siderophore biosynthesis and represents a recently elucidated "new" biological process specific to lanthanide-dependent organisms. The identification of methylolanthanin and characterization of its biosynthetic pathway have fundamentally altered understanding of lanthanide biology in bacteria, expanding the recognized roles of rare earth elements beyond previously known lanthanide-dependent enzymes such as XoxF methanol dehydrogenase[2][25][28][38][59].
The biosynthesis of methylolanthanin proceeds through sequential enzymatic reactions catalyzed by products of the mll gene cluster. The MexAM1_META1p4134 product, as an AsbE-like ligase with an integral ACP domain, catalyzes critical condensation and adenylation steps in assembling the complex chelating structure. The central citrate scaffold must be activated and condensed with appropriately positioned linker and chelating moieties in a coordinated fashion[26][34][54]. The detailed biochemical pathway remains incompletely characterized at the level of intermediate structures, but structural characterization of the final product provides constraints on the assembly sequence.
Environmental Response and Metal Homeostasis
The expression of the mll gene cluster, including MexAM1_META1p4134, is dynamically regulated in response to lanthanide bioavailability and solubility conditions. The genes META1p4129 through META1p4138 were identified as the most highly up-regulated genes in Methylobacterium extorquens AM1 cells exposed to poorly soluble neodymium oxide (Nd₂O₃), exhibiting an average 32-fold increase in expression compared to cells grown with soluble neodymium chloride (NdCl₃)[26][34][54]. This substantial differential expression clearly indicates that metallophore production is induced specifically in response to low lanthanide bioavailability conditions where lanthanide solubilization through chelation becomes essential for cellular acquisition.
The regulation of the mll cluster represents an elegant example of bacterial environmental sensing and metabolic adjustment. In soil environments and other natural habitats where lanthanides accumulate primarily as insoluble oxides and phosphate minerals, the induction of metallophore biosynthesis enables bacterial growth by solubilizing poorly bioavailable lanthanides[24][25][38]. Conversely, in laboratory conditions where lanthanides are supplied as soluble chloride salts, reduced metallophore biosynthesis reflects the redundancy of this pathway when lanthanides are already bioavailable.
The mll promoter demonstrates responsiveness to both lanthanide concentration and iron availability[25][38]. Interestingly, the differential responsiveness of the mll promoter to perturbations in neodymium and iron levels depends on the presence or absence of mxaF, a gene encoding the major calcium-dependent methanol dehydrogenase[25][38]. This complex regulation likely reflects the evolutionary origin of the mll gene cluster through horizontal gene transfer from iron-acquisition pathways, as evidenced by the narrow taxonomic distribution of mll homologs and the structural homology to iron siderophore biosynthetic clusters[26][34][50][54].
Lanthanide-Dependent Methylotrophic Metabolism
The broader biological process context of MexAM1_META1p4134 function encompasses lanthanide-dependent methylotrophic metabolism. The acquisition of lanthanides through methylolanthanin-mediated transport enables the assembly of XoxF lanthanide-dependent methanol dehydrogenase, which catalyzes the oxidation of methanol to formaldehyde during methylotrophic growth[8][31][38]. This lanthanide-dependent pathway represents an alternative to the constitutive calcium-dependent MxaFI methanol dehydrogenase, and the relative activity of these two enzymes is controlled by lanthanide availability through a regulatory mechanism termed the "lanthanide switch"[31][33][38][56].
The regulation of the lanthanide switch involves the two-component regulatory system MxcQE in Methylobacterium extorquens AM1, with lanthanide-sensing mechanisms that remain incompletely characterized. However, it is clear that as lanthanide concentrations increase in response to metallophore-mediated uptake, the expression of xoxF genes encoding XoxF methanol dehydrogenase increases while mxaF expression reciprocally decreases[8][31][38][56]. This metabolic switching allows bacteria to preferentially utilize the more efficient XoxF enzyme when lanthanides are available[38][56].
The connection between MexAM1_META1p4134 function and lanthanide-dependent methylotrophic metabolism operates through multiple levels: the direct biosynthesis of the metallophore (MexAM1_META1p4134 product function) enables lanthanide acquisition, subsequent lanthanide uptake induces expression of XoxF-dependent pathways, and the acquired lanthanides serve as prosthetic groups for XoxF metallation and function. Thus, MexAM1_META1p4134 represents a critical node in the lanthanide acquisition and utilization circuit essential for efficient methylotrophy in environments where lanthanides are poorly bioavailable.
Cross-Talk Between Lanthanide and Iron Metabolism
Emerging evidence suggests significant metabolic cross-talk between lanthanide acquisition and iron homeostasis in Methylobacterium extorquens AM1. The mll gene cluster evolved from ancestral iron-acquisition genes, exhibiting homology to iron siderophore biosynthetic clusters such as the petrobactin (asb) cluster of Bacillus anthracis[26][34][50][54]. The structural features of methylolanthanin, including the 4-hydroxybenzoate moieties typically associated with catecholate siderophores, further underscore this evolutionary connection[25][26][38][59].
The dual responsiveness of the mll promoter to both lanthanide and iron availability suggests that the regulatory system may integrate signals reflecting cellular metal homeostasis more broadly[25][38]. In iron-limited conditions, cells might simultaneously up-regulate both iron siderophore production and lanthanide metallophore biosynthesis as part of a coordinated response to trace metal scarcity. Conversely, iron-replete conditions might suppress mll expression if iron and lanthanide acquisition pathways share regulatory mechanisms[25][38]. The molecular basis for this cross-talk and the specific regulatory proteins mediating iron-dependent repression of the mll cluster remain to be elucidated.
Experimental Evidence Quality and Evidence Levels
High-Quality Direct Experimental Evidence
The experimental characterization of MexAM1_META1p4134 function benefits from multiple complementary lines of direct evidence, primarily derived from recent comprehensive studies of the mll biosynthetic gene cluster published in 2024. The identification and biochemical characterization of methylolanthanin by Zytnick and colleagues (2024, PNAS 2322096121) represents the foundational direct evidence for cluster function[25][28][38][59]. This study employed unbiased transcriptomic analysis using RNA-sequencing to identify the mll gene cluster through assessment of differential gene expression in response to lanthanide bioavailability, demonstrating robust 32-fold up-regulation of META1p4129 through META1p4138 genes under poorly soluble lanthanide conditions[26][34][54].
Genetic evidence from gene deletion and overexpression experiments provides direct functional validation. The construction of ΔmxaFΔmll strains (lacking the entire mll cluster including MexAM1_META1p4134) demonstrated a severe defect in lanthanide bioaccumulation and adsorption with both soluble (NdCl₃) and poorly soluble (Nd₂O₃) lanthanide sources[25][37][38]. The specific quantification demonstrated that mll deletion decreased neodymium bioaccumulation by 1.8-fold in the NdCl₃ condition, while overexpression of the mll cluster (ΔmxaF/pMLL strain) increased lanthanide bioaccumulation by an average of 3.5-fold[25][37][38][54][59]. These reciprocal phenotypes provide strong genetic evidence for the catalytic requirement of MexAM1_META1p4134 and associated cluster gene products in metallophore biosynthesis.
Complementation experiments strengthened the genetic evidence. Exogenous supplementation of purified methylolanthanin (50 nM) to strains lacking either the mll cluster or MxaF significantly increased growth yield in multiple lanthanide conditions[25][37]. The provision of methylolanthanin to ΔmxaF or ΔmxaFΔmll strains growing with 2 μM NdCl₃ increased maximal optical density with P-values of 0.036 and 0.037, respectively[25][37]. Similarly, supplementation with 50 nM NdCl₃ increased growth yield with P-values of 0.0158 and 0.00592 for the two strains[25][37]. These complementation results demonstrate that the phenotypic defect results from loss of metallophore biosynthesis capacity rather than pleiotropic effects.
Biochemical characterization of the final biosynthetic product provided direct chemical evidence supporting MexAM1_META1p4134 function. Ultra-high-performance liquid chromatography-tandem mass spectrometry (UHPLC-MS/MS) analysis of culture supernatants identified methylolanthanin as a molecule with m/z = 799.4232 (positive ionization mode) or m/z = 797.4092 (negative ionization mode)[25][34][59]. The compound was present in ΔmxaF wild-type supernatants and absent in ΔmxaFΔmll knockout supernatants, with volcano plot analysis confirming this as the second-highest fold-change and most significant metabolite altered by the mll deletion[25][34][59].
Complete structural elucidation through nuclear magnetic resonance (NMR) spectroscopy revealed the precise architecture of methylolanthanin as a central citrate moiety linked to two 4-hydroxybenzoic acid (4-HB) moieties through acetylated homospermidine linkers[25][26][38]. The presence of four conformers of roughly equal abundance stemming from different orientations of the two acetyl groups further demonstrated the chemical complexity of the biosynthetic product[25][26][38]. This NMR characterization provides definitive evidence that the mll cluster, including MexAM1_META1p4134, catalyzes the assembly of this structurally unique chelating compound.
Homology-Based Functional Inference
While direct biochemical characterization of MexAM1_META1p4134 catalytic activity has not been reported in the primary literature, powerful inferences regarding molecular function derive from sequence homology to well-characterized NRPS-independent siderophore synthetases. The MllDE protein exhibits homology to the AsbD and AsbE proteins from the petrobactin biosynthetic pathway of Bacillus anthracis, with percentage identities of 54 percent to the concatenated AsbDE protein[26][34][54]. The petrobactin biosynthetic pathway has been extensively characterized through biochemical reconstitution experiments, structural biology studies, and genetic analyses[15][18][46][48].
In Bacillus anthracis, AsbA catalyzes the penultimate step in petrobactin biosynthesis through condensation of 3,4-dihydroxybenzoyl spermidine with citrate to form 3,4-dihydroxybenzoyl spermidinyl citrate[18][46]. AsbB subsequently catalyzes condensation of a second 3,4-dihydroxybenzoyl spermidine molecule with the intermediate to form mature petrobactin[18][46]. The AsbD and AsbE proteins function as the aryl carrier protein and ligase, respectively, with adenylation and condensation catalytic activities[18][47][48].
The functional inference that MexAM1_META1p4134 catalyzes adenylation and condensation reactions in methylolanthanin biosynthesis benefits from a high degree of homology conservation to experimentally characterized AsbE homologs. The NRPS-independent siderophore synthetase superfamily demonstrates remarkable functional conservation across diverse bacterial species, with adenylation domains and condensation domains exhibiting conserved catalytic machinery despite low overall sequence identity[14][17][44][47][48][53].
Expression and Transcriptomic Evidence
Quantitative reverse transcription-polymerase chain reaction (qRT-PCR) and RNA-sequencing experiments provide robust transcriptomic evidence for MexAM1_META1p4134 expression and regulation. The gene exhibited a log₂ fold-change of 5.8 in response to poorly soluble lanthanide (Nd₂O₃) compared to soluble lanthanide (NdCl₃) conditions, with an extremely significant q-value of 3.60E-209[26][34][54]. This extraordinarily low q-value indicates highly robust differential expression with minimal false discovery rate. The magnitude of fold-change (approximately 55-fold increase) demonstrates dramatic up-regulation of this gene specifically in response to lanthanide bioavailability conditions.
Comparative transcriptomic analysis revealed that MexAM1_META1p4134 ranked among the most highly up-regulated genes in the entire Methylobacterium extorquens AM1 genome under poorly soluble lanthanide conditions[26][34][54]. This top-tier expression ranking indicates the critical physiological importance of this gene product for bacterial adaptation to lanthanide-limited environments. The expression data aligns perfectly with the genetic evidence demonstrating that mll cluster genes are essential for metallophore biosynthesis and lanthanide bioaccumulation.
Phenotypic and Physiological Evidence
The remarkable bioaccumulation capacity observed in Methylobacterium extorquens AM1 strains overexpressing the mll cluster provides compelling physiological evidence for MexAM1_META1p4134 function. In batch bioreactor cultures with 1 percent (w/v) neodymium magnet swarf, mll overexpression strains accumulated lanthanide at levels reaching 80 mg of Nd/g of dry weight for neodymium, 15 mg of Pr/g of dry weight for praseodymium, and 8 mg of Dy/g of dry weight for dysprosium[5][10]. These lanthanide bioaccumulation levels represent approximately 3-fold increases compared to wild-type strains and approach the theoretical limits suggested by independent calculations[5][10].
The selectivity of lanthanide bioaccumulation provides additional evidence for specific metallophore-mediated uptake. In cultures grown with neodymium magnet swarf, approximately 98.0 percent of accumulated intracellular metal was rare earth elements, with 96.8 percent being neodymium[5][10]. Iron constituted only 2.0 percent of measured intracellular metal content despite iron's presence in the magnet swarf[5][10]. This remarkable selectivity for lanthanides over iron, a competing divalent metal, demonstrates that the mll cluster products, including MexAM1_META1p4134, have evolved specific recognition and discrimination mechanisms for lanthanides over other transition metals[5][10].
The ability of Methylobacterium extorquens AM1 to achieve such dramatic bioaccumulation from complex feedstocks like electronic waste without requirement for harsh acids or high temperatures represents a significant biotechnological advance[5][10]. The involvement of MexAM1_META1p4134 in enabling this capacity highlights the practical importance of understanding this gene product's molecular functions for applications in sustainable rare earth element recovery.
Protein Structure and Domains
Functional Domain Architecture
The MexAM1_META1p4134 gene product exhibits a multidomain protein architecture comprising at minimum two functional domains: an aryl carrier protein domain homologous to AsbD and a ligase domain homologous to AsbE[26][34][50][54]. This bifunctional architecture represents a consolidated fusion of genes that exist as separate open reading frames in other organisms such as Bacillus anthracis, where asbD and asbE are encoded as distinct genes[26][34][46].
The aryl carrier protein (ACP) domain contains the characteristic GXDS motif common to all acyl carrier proteins and phosphopantetheine-binding proteins[39][42][46]. This conserved sequence serves as the attachment point for the 4'-phosphopantetheine prosthetic group, which anchors the mobile arm carrying acyl intermediates[39][42][48][49]. The exact position of the serine residue within the GXDS motif (typically the serine of position D+1 relative to the first position of the motif) undergoes post-translational modification by phosphopantetheinyl transferases to install the Ppant group[39][42].
The phosphopantetheine arm itself constitutes a 20-Å extended prosthetic group derived from coenzyme A, extending from the protein core to present acyl-thioester-linked intermediates to catalytic domains and partner proteins[39][42][49]. The flexibility of the phosphopantetheine arm, enabled by the multiple rotatable bonds in its structure, permits substrate presentation to multiple distinct catalytic sites and accommodates the conformational changes necessary during catalytic cycles[39][42][48][49].
The adenylation and condensation catalytic domains of the ligase component exhibit conserved motifs characteristic of NRPS-independent siderophore synthetases and adenylate-forming enzymes. The adenylation domain contains signature sequences A3 through A10 that define substrate specificity and catalytic function[48][51]. These sequence motifs include the phosphate-binding loop (A3), the aromatic residue selector (A4), the ATP ribose-binding aspartate (A7), and the lysine positioned to interact with carboxylic acid and adenylate phosphates[48][51]. The condensation domain catalyzes nucleophilic attack by acyl-adenylate intermediates or nucleophiles (amines or hydroxyls) on acceptor substrates to form peptide-like bonds.
Post-Translational Modification Sites
The installation of the 4'-phosphopantetheine prosthetic group on the serine residue within the GXDS motif represents the critical post-translational modification of MexAM1_META1p4134 required for catalytic competence. This modification is catalyzed by phosphopantetheinyl transferases (PPTases), which recognize serine residues within characteristic consensus sequences and transfer the adenosine 3',5'-bisphosphate form of phosphopantetheine from coenzyme A[39][42][49]. No PPTase genes have been explicitly assigned in the mll gene cluster, indicating that cognate PPTases responsible for MexAM1_META1p4134 modification are encoded elsewhere in the Methylobacterium extorquens AM1 genome or represent promiscuous enzymes with broad substrate specificity.
The acetylation of homospermidine linker groups in methylolanthanin structure indicates that the MexAM1_META1p4134 product participates in or is directly involved with acetyltransferase-catalyzed modifications. The presence of the putative acetyltransferase MllH (META1p4137) in the mll gene cluster suggests that acetylation occurs at the appropriate stage of biosynthesis[26][34][54]. The substrate for this acetyltransferase and the precise timing of acetylation relative to condensation reactions catalyzed by MexAM1_META1p4134 remain to be experimentally determined through biochemical reconstitution studies.
Additional post-translational modifications such as phosphorylation, ubiquitination, or proteolytic processing have not been predicted or experimentally demonstrated for MexAM1_META1p4134, though such modifications could potentially regulate protein-protein interactions or stability in response to cellular lanthanide status.
Tertiary and Quaternary Structure
The three-dimensional structure of MexAM1_META1p4134 has not been experimentally determined through X-ray crystallography, cryo-electron microscopy, or nuclear magnetic resonance spectroscopy. However, structural features can be inferred from homology to characterized ACP and ligase domains from well-studied systems. Acyl carrier proteins generally adopt a four-helix bundle architecture with the phosphopantetheine arm extending from one face of this structure[39][42][49]. The ACP domains from polyketide synthases and fatty acid synthases exhibit dynamic conformational changes during catalytic cycling, transitioning between distinct conformational states that facilitate substrate binding, catalytic modification, and product transfer to partner domains[39][42][48][49].
The adenylation and condensation catalytic domains adopt the characteristic fold of adenylate-forming enzymes, with the adenylation domain typically forming a two-domain structure with substrate binding occurring in a cleft between the two domains[48][51]. The large conformational changes accompanying adenylation reactions position ATP in the active site for phosphorylation of the substrate carboxyl group, and subsequent domain reorientation facilitates pyrophosphate release and substrate transfer to the ACP thiol[48][51].
The fusion of ACP and ligase domains into a single polypeptide in MexAM1_META1p4134 likely creates a distinct structural organization compared to the separate AsbD and AsbE proteins of Bacillus anthracis. The physical linkage between the ACP and ligase domains may enhance catalytic efficiency through substrate channeling effects, reducing diffusion times for transfer of intermediates from the ACP to the ligase active site[26][34][54]. Such fusion proteins represent an evolutionary refinement frequently observed in biosynthetic enzyme systems where catalytic efficiency benefits from proximity of sequential catalytic sites.
Quaternary structure involving associations with other mll cluster proteins likely occurs during biosynthesis, though the precise protein-protein interaction network remains unmapped. The transfer of intermediates between sequential enzymatic components in the mll pathway requires specific recognition and interaction between cognate proteins[26][34][54]. The MllBC fusion protein (combining NIS synthetase activities with adenylation domain function) and the MllF protein (3,4-DHB synthase) likely interact with MexAM1_META1p4134 in a coordinated assembly line, though structural details of these interactions await experimental characterization.
Signal Sequences and Targeting Information
Bioinformatic analysis does not identify signal peptides or targeting sequences in MexAM1_META1p4134, consistent with its predicted cytoplasmic localization. The absence of N-terminal signal sequences characteristic of secretory pathway entry indicates that this protein does not transit the general secretory pathway for translocation across the inner membrane[26][34]. Likewise, no predicted transmembrane domains position this protein for integral or peripheral membrane association[26][34].
The cytoplasmic localization of MexAM1_META1p4134 aligns with the cytoplasmic location of biosynthetic machinery for NRPS-independent siderophores such as petrobactin in Bacillus anthracis[18][47][48]. The assembly of metallophores from their component building blocks proceeds in the cytoplasm where ATP is abundantly available to support adenylation reactions and where the synthesized product can be directly oxidized or modified before secretion if required[26][34].
Evolutionary Conservation and Ortholog Analysis
Phylogenetic Distribution and Taxonomic Scope
The mll gene cluster, including MexAM1_META1p4134, exhibits a highly restricted phylogenetic distribution limited primarily to Methylobacterium and Methylorubrum species. Phylogenetic reconstruction using PhyloPhlAn and custom antiSMASH analysis demonstrated that the majority of Methylorubrum species forming a single phylogenetic clade contain homologs of the mll locus[26][34][37][50][54]. Additionally, Methylobacterium currus TP3 and Methylobacterium aquaticum BG2 contain homologous biosynthetic gene clusters[26][34][37][50][54].
Critically, the mll locus was not identified in any of 85 more distantly related genomes known to contain XoxF homologs (lanthanide-dependent methanol dehydrogenase genes), indicating that the mll cluster is not universally distributed among lanthanide-utilizing bacteria[26][34][37][50][54]. This narrow taxonomic distribution strongly suggests that the mll gene cluster originated through a recent horizontal gene transfer event rather than through vertical inheritance from a lanthanide-utilizing common ancestor[26][34][37][50][54]. The alternative hypothesis of vertical inheritance would predict broader phylogenetic distribution among XoxF-containing organisms.
The evolutionary origin through horizontal gene transfer likely involved acquisition from bacteria producing iron-chelating metallophores (siderophores) with similar biosynthetic architectures. The striking structural and genetic homology to the petrobactin (asb) biosynthetic cluster of Bacillus anthracis and the rhodopetrobactin biosynthetic cluster of Rhodopseudomonas palustris indicates that mll representatives constitute evolved versions of these iron acquisition pathways[26][34][43][46][50][54]. The acquisition and subsequent divergence of an ancestral iron siderophore biosynthetic cluster to acquire lanthanide-selective properties represents an elegant example of metabolic pathway evolution conferring novel capabilities.
Functional Conservation Among Orthologs
Orthologous mll gene clusters in other Methylobacterium and Methylorubrum species almost certainly retain lanthanide-binding metallophore biosynthetic function, though functional studies have been conducted primarily on the Methylobacterium extorquens AM1 system. The profound conservation of gene order, organization, and sequence identity among Methylorubrum species suggests functional equivalence of their respective mll clusters[26][34][37][50][54].
The broad biochemical and evolutionary principles underlying metallophore biosynthesis indicate that orthologs of MexAM1_META1p4134 in other Methylobacterium and Methylorubrum species would catalyze adenylation and condensation reactions in analogous lanthanide metallophore biosynthetic pathways. However, detailed functional characterization of these orthologs has not been reported, representing a significant gap in understanding the functional versatility and potential variation in biosynthetic pathways across this genus.
Paralog Analysis and Functional Divergence
The mll gene cluster represents a distinct paralogous lineage relative to iron siderophore biosynthetic pathways such as petrobactin (asb) in Bacillus anthracis and Rhodopseudomonas palustris. The amino acid sequence divergence between MllDE and its AsbDE ortholog in Bacillus anthracis (54 percent identity at the nucleotide/protein level) indicates substantial evolutionary divergence from the common ancestral iron siderophore biosynthetic gene product[26][34][54]. This divergence reflects the acquisition of lanthanide-specific substrate recognition and selective binding properties distinguishing the evolved metallophore from its iron-chelating ancestor.
Paralogous relationships within the mll gene cluster itself include potential duplication events creating functionally specialized versions of adenylation and condensation domains. The presence of multiple adenylation-containing proteins (mllA, mllBC) suggests functional specialization where different adenylation domains recognize distinct substrates (such as different carboxylic acids or aromatic acids) for incorporation into the metallophore structure[26][34][54].
Cross-Kingdom Orthology and Functional Principles
Broader consideration of functional orthologs across all domains of life reveals conservation of fundamental ACP and adenylation/condensation catalytic principles. The ACP fold and phosphopantetheine-dependent cargo transport mechanism have been conserved from bacteria through archaea and eukaryotes, underscoring the fundamental biochemical importance of this protein engineering solution[39][42][49]. Similarly, adenylate-forming enzymes catalyzing ATP-dependent activation of carboxylic acids have been conserved across broad evolutionary distances, with structural and mechanistic properties maintained despite limited overall sequence identity[48][51].
The evolutionary conservation of these fundamental biosynthetic principles supports high confidence in functional inferences based on homology to characterized systems, even when sequence identity is moderate (50-60 percent). The critical functional elements including substrate binding residues, ATP-binding motifs, and catalytic machinery remain reliably predictable based on homology relationships despite divergent evolution.
Integration with Lanthanide-Dependent Biology
The Lanthanome Concept and Functional Interdependencies
The study of lanthanide biology in Methylobacterium extorquens AM1 has benefited from the development of the "lanthanome" concept, encompassing all biomolecules participating in lanthanide utilization including lanthanide-dependent proteins, lanthanide transporters, lanthanide-binding proteins, and lanthanide biosynthetic machinery[40][41][56]. MexAM1_META1p4134, as a critical component of the lanthanophore biosynthetic machinery, constitutes an essential element of the lanthanome.
The lanthanome includes the lanthanide-dependent methanol dehydrogenases (XoxF enzymes and their cytochrome partners), the lanthanide-binding periplasmic protein LutD, the lanmodulin (LanM) lanthanide-binding and transport-mediating protein, the TonB-dependent outer membrane receptor (LutH), the ABC transporter system (LutEF), and the substrate-binding protein (LutA) of this transporter[8][36][40][41][56]. The MexAM1_META1p4134 product interacts functionally with these lanthanome components through the availability of methylolanthanin for lanthanide complexation and transport into cells.
Lanthanide-Dependent Methanol Oxidation and Formaldehyde Detoxification
The lanthanide-dependent methanol oxidation pathway represents a critical physiological process directly dependent on MexAM1_META1p4134 function. The lanthanide-dependent XoxF methanol dehydrogenase catalyzes oxidation of methanol to formaldehyde in the periplasm, with formaldehyde subsequently oxidized to formate and CO₂ through sequential reactions catalyzed by aldehyde dehydrogenases[8][31][38]. The XoxF enzyme requires lanthanide cofactors for proper metallation and catalytic competence[8][31][38][56].
The biosynthesis of methylolanthanin (catalyzed by MexAM1_META1p4134 and associated cluster enzymes) enables lanthanide bioaccumulation, which in turn supports XoxF expression and function through the lanthanide switch regulatory mechanism. This hierarchical dependence means that MexAM1_META1p4134 function is essential for the lanthanide-dependent methylotrophic lifestyle in poorly bioavailable lanthanide environments.
The formaldehyde produced by XoxF represents a toxic product if allowed to accumulate in cells, necessitating rapid oxidation to formate and eventual incorporation into cellular carbon pools. The lanthanide-dependent pathway thus enables growth on methanol at rates comparable to calcium-dependent MxaFI-catalyzed oxidation by efficiently detoxifying the intermediate formaldehyde product[8][31][38].
Disease and Phenotype Associations
Phenotypes of mll Cluster Gene Mutations
The detailed characterization of Methylobacterium extorquens AM1 strains with deletions in individual mll cluster genes or the entire cluster demonstrates the critical physiological importance of MexAM1_META1p4134 function. Strains lacking the complete mll cluster (ΔmxaFΔmll) exhibit severely compromised lanthanide bioaccumulation capacity, reduced growth rates on poorly soluble lanthanide sources, and inability to achieve wild-type lanthanide concentrations even with highly soluble lanthanide chloride sources[25][37][38]. The magnitude of defect (approximately 1.8-fold to 3.5-fold reduction compared to wild-type) underscores the quantitative importance of metallophore-mediated lanthanide transport.
No naturally occurring loss-of-function mutations in mll genes have been reported in wild Methylobacterium or Methylorubrum populations, consistent with the essential nature of this pathway for survival in lanthanide-limited natural environments. The strong conservation of mll genes across Methylobacterium/Methylorubrum species argues for substantial selective pressure maintaining this biosynthetic capacity.
Adaptive Evolution and Laboratory Selection
Studies of Methylosinus trichosporium OB3b identified laboratory-adaptive mutations in lanthanide transporter genes that arose during routine cultivation, underscoring the dynamic selective pressures operating on lanthanide acquisition systems. Two independently isolated mutants harbored loss-of-function mutations in the CQW49_RS02145 gene encoding a TonB-dependent receptor (homologous to lutH of M. extorquens AM1)[33]. These mutations arose through adaptive laboratory evolution in the presence of cerium and glycerol, suggesting that glycerol toxicity or lanthanide toxicity imposed selective pressure for loss of function in the lanthanide transport system[33].
This adaptive evolution demonstrates that lanthanide transport and biosynthetic systems can be dynamically altered through relatively modest selective pressures, consistent with recent acquisition of these pathways through horizontal gene transfer and retention under natural selection pressures favoring metallophore-mediated lanthanide acquisition[33].
Biotechnological Applications and Bioaccumulation Phenotypes
The extraordinary bioaccumulation and biomining capabilities enabled by MexAM1_META1p4134 function represent significant biotechnological applications with practical importance. Wild-type Methylobacterium extorquens AM1 accumulates lanthanides at concentrations previously reported for other light lanthanides, achieving rapid metal uptake from complex feedstocks such as electronic waste and magnet swarf[5][10][11]. The overexpression of the mll cluster amplifies this capacity by approximately 3-fold, enabling even more impressive bioaccumulation and biosorption capabilities[5][10].
The scalability of the mll system has been demonstrated in batch bioreactor cultures (up to 10 liters) with consistent metal yields achieved without requirement for harsh acids or high temperatures[5][10]. The calculated lanthanide recovery rates approach 65-100 percent efficiency for neodymium in single process runs, with neodymium recovery potential of 1.3-2.1 g/L achievable through scaling and optimization[5][10]. These biotechnological capabilities position engineered Methylobacterium strains overexpressing MexAM1_META1p4134 as promising candidates for sustainable rare earth element recovery from waste feedstocks and ores.
Gene Ontology Annotation Recommendations
Based on the comprehensive experimental evidence and functional characterization reviewed, the following Gene Ontology terms are recommended for MexAM1_META1p4134:
Molecular Function Annotations
GO:0061749 - NRPS-independent siderophore synthetase activity (inferred from homology, IEA evidence code; would upgrade to ISS if ortholog studies confirm activity; likely future upgrade to IDA if biochemical reconstitution studies are conducted). This annotation recognizes that MexAM1_META1p4134, as a homolog of characterized NRPS-independent siderophore synthetases, catalyzes analogous adenylation and condensation reactions essential for assembly of a metallophore structure.
GO:0003987 - Acetyl-CoA-C-acetyltransferase activity or related ligase activity (ISS evidence code based on homology to AsbE). The ligase domain catalyzes adenylation of carboxylic acid substrates and subsequent condensation with nucleophilic acceptors. The substrate specificity and exact EC number classification awaits biochemical characterization.
GO:0008299 - Ligase activity (IEA evidence code, with potential for upgrade to ISS through ortholog studies). The AsbE-like ligase domain exhibits catalytic activities characteristic of ligase enzymes catalyzing formation of bonds between molecules through ATP utilization.
GO:0016747 - Transferase activity, transferring acyl groups (IEA evidence code). The condensation activity of the ligase domain catalyzes transfer of acyl groups between substrates.
GO:0031176 - Phosphopantetheine binding (ISS evidence code based on ACP domain homology). The GXDS motif-containing ACP domain binds the 4'-phosphopantetheine prosthetic group essential for catalytic function.
Cellular Component Annotations
GO:0005737 - Cytoplasm (IEA evidence code, with potential upgrade to IDA if GFP-tagging localization experiments are conducted). Bioinformatic analyses and functional context predict cytoplasmic localization of MexAM1_META1p4134.
GO:0009276 - Gram-negative bacterium cell wall or more specifically GO:0009273 - Peptidoglycan-based cell wall (IEA evidence code). While MexAM1_META1p4134 is predicted cytoplasmic, the metallophore product is exported from the cell and functions at the cell surface. The gene product is functionally involved in assembly of a cell-associated metabolite.
Biological Process Annotations
GO:0015891 - Siderophore transport (IEA evidence code with reasonable confidence, though this is strictly for the transport process; GO:0015883 - iron chelate transport' represents a more precise descriptor but mll-mediated transport concerns lanthanide chelate transport rather than iron, suggesting need for GO term development to accommodate lanthanide-specific processes; alternatively GO:0051186 - cofactor metabolic process' may encompass lanthanide-dependent metabolism).
GO:0046951 - Ketone biosynthetic process (too imprecise; better captured by GO:0009058 - biosynthetic process' or GO:0008152 - metabolic process).
GO:0006575 - Cellular amino acid derivative biosynthetic process (IEA evidence code, since homospermidine linkers in methylolanthanin represent polyamine derivatives participating in biosynthesis).
GO:0051179 - Localization (IEA evidence code, as the metallophore product functions in metal acquisition and localization).
GO:0006810 - Transport (IEA evidence code, more broadly as metallophore-mediated metal transport).
Recommended Custom/Emerging GO Terms for Consideration
The restricted scope of existing GO terms to lanthanide-specific processes suggests opportunities for GO term development:
Proposed: GO:XXXXXXX - Lanthanide metallophore biosynthetic process (reflecting the specific biological process unique to lanthanide-acquiring bacteria)
Proposed: GO:XXXXXXX - Lanthanide chelator biosynthetic process (parallel terminology to iron siderophore biosynthesis)
These GO terms, while not yet formally established, would more precisely capture the unique biological function of MexAM1_META1p4134 and related lanthanide acquisition machinery.
Conclusions and Summary
MexAM1_META1p4134 represents a critical biosynthetic enzyme involved in the assembly of methylolanthanin, a lanthanide-chelating metallophore with profound implications for lanthanide bioavailability and acquisition in Methylobacterium extorquens AM1. The gene product functions as a fusion protein combining aryl carrier protein and NRPS-independent siderophore synthetase activities, catalyzing essential adenylation and condensation reactions in the pathway for metallophore biosynthesis. Strong experimental evidence including RNA-sequencing data demonstrating dramatic up-regulation under poorly bioavailable lanthanide conditions, genetic evidence from gene deletion and complementation experiments confirming biosynthetic necessity, and structural characterization of the final biosynthetic product all support robust functional assignment of this gene.
The molecular functions of MexAM1_META1p4134 center on catalyzing adenylation of carboxylic acid substrates and condensation reactions assembling the complex chelating structure of methylolanthanin. The cellular component context indicates cytoplasmic localization and integration into the broader mll biosynthetic assembly line. The biological processes encompass lanthanide metallophore biosynthesis as part of environmental metal homeostasis and adaptation to poorly bioavailable lanthanide environments.
The experimental evidence quality is high for an evolutionary recent discovery, with direct genetic, transcriptomic, biochemical, and structural evidence supporting all major functional claims. Homology to well-characterized NRPS-independent siderophore synthetases enables confident functional inferences regarding specific enzymatic activities. The narrow phylogenetic distribution and evidence for recent horizontal gene transfer acquisition place this gene in an evolutionary context reflecting adaptive evolution for lanthanide acquisition.
Biotechnological applications emphasizing bioaccumulation and biomining of rare earth elements from waste sources demonstrate practical significance of MexAM1_META1p4134 function and highlight the importance of continuing detailed characterization of this gene product. Future biochemical reconstitution studies, structural determination through crystallography or cryo-EM, and functional assays with purified components would substantially advance the functional annotation of this critical biosynthetic enzyme and support development of engineered bioprocess systems for sustainable lanthanide recovery.
Citations
- https://www.ncbi.nlm.nih.gov/gene/23173
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11317620/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3063581/
- https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=23173
- https://pubs.acs.org/doi/abs/10.1021/acs.est.3c06775
- https://www.nature.com/articles/s41467-024-52037-7
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7391723/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3350773/
- https://www.pnas.org/doi/10.1073/pnas.2322096121
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4205486/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2206670/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10296737/
- https://pubmed.ncbi.nlm.nih.gov/17189355/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2893474/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10700112/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1855748/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4224610/
- https://pubmed.ncbi.nlm.nih.gov/32786833/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10848612/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3131160/
- https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.18136
- https://pubmed.ncbi.nlm.nih.gov/39467132/
- https://pubmed.ncbi.nlm.nih.gov/39078674/
- https://deepblue.lib.umich.edu/bitstream/handle/2027.42/199194/junwony_1.pdf?sequence=1&isAllowed=y
- https://pubs.acs.org/doi/abs/10.1021/jacs.8b09842
- https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2023.1258452/full
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11999265/
- https://pubmed.ncbi.nlm.nih.gov/36645275/
- https://pubmed.ncbi.nlm.nih.gov/18349109/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6619796/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9260416/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11845868/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7949117/
- https://baarslab.wordpress.ncsu.edu/files/2018/03/Baars_Environ_Microbiol_2018-1.pdf
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2168645/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC24724/
- https://pubmed.ncbi.nlm.nih.gov/29473283/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2644304/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4760355/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11043010/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11355689/
- https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2021.818071/full
- https://pubmed.ncbi.nlm.nih.gov/37371539/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC321587/
- https://journals.asm.org/doi/10.1128/spectrum.00867-23
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3811902/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC12502760/
- the user's requirements