Deep Research
Deep Research on mxaI: The Small Subunit of Calcium-Dependent Methanol Dehydrogenase in Methylorubrum extorquens AM1
(mxaI-claude-deep-research.md)
Deep Research on mxaI: The Small Subunit of Calcium-Dependent Methanol Dehydrogenase in Methylorubrum extorquens AM1
(mxaI-claude-deep-research.md)Deep Research on mxaI: The Small Subunit of Calcium-Dependent Methanol Dehydrogenase in Methylorubrum extorquens AM1
Introduction and Historical Context
Methanol dehydrogenase (MDH) represents a critical enzyme in the metabolism of methylotrophic bacteria, catalyzing the first step in methylotrophic growth by oxidizing methanol to formaldehyde in the periplasm [PMID:21873495, "the first step in methylotrophic growth is the oxidation of methanol to formaldehyde in the periplasm by methanol dehydrogenase"]. In most methylotrophic organisms studied to date, this enzyme consists of two distinct protein subunits: MxaF (the large catalytic subunit) and MxaI (the small structural subunit) [PMID:21873495, "this enzyme consists of the MxaF and MxaI proteins, which make up the large and small subunits of this heterotetrameric enzyme"]. The MxaI protein, encoded by the mxaI gene (locus tag MexAM1_META1p4535 in M. extorquens AM1), has remained one of the more enigmatic components of this essential enzyme complex, with its precise functional role continuing to be a subject of investigation.
The canonical calcium-dependent MxaFI-type methanol dehydrogenase exhibits a heterotetrameric quaternary structure with α₂β₂ stoichiometry, consisting of two 66-kDa large subunits (MxaF) and two small 8.5-kDa subunits (MxaI). This heterotetrameric architecture distinguishes MxaFI-type MDHs from the more recently characterized XoxF-type lanthanide-dependent methanol dehydrogenases, which function as homodimers without a small subunit component. The presence of MxaI thus represents a unique structural feature of the calcium-dependent enzyme system.
Protein Structure and Molecular Characteristics
MxaI is an exceptionally small protein, comprising only approximately 69 amino acids with a theoretical molecular weight of approximately 7.77-8.5 kDa [Web search results, "the beta subunit has a length of 69 amino acids with a theoretical weight of 7.77 KDa" and "two small 8.5-kDa subunits (MxaI)"]. The slight variation in reported molecular weight likely reflects differences in measurement methods or post-translational modifications across different methylotrophic species. High-resolution crystallographic studies of methanol dehydrogenase from Methylobacterium extorquens at 1.94 Å resolution revealed the overall architecture of the α₂β₂ tetramer, though specific structural details of the beta subunit were not fully elaborated in the available abstracts [PMID:7735834, "The structure of the α2β2 tetramer of MDH from Methylobacterium extorquens has now been determined at 1.94 A with an R-factor of 19.85%"].
The most distinctive structural characteristic of MxaI is its intimate association with the MxaF large subunit. The highly basic small subunit tightly wraps around the large subunit, forming an essentially inseparable complex [Web search results, "The highly basic small subunit, which is not found in other quinoproteins, tightly wraps against the large subunit"]. This tight association has important biochemical implications: the beta subunit cannot be reversibly dissociated from the alpha subunit under normal experimental conditions. Notably, MxaI is not found in any other quinoprotein enzymes, making it a unique structural element specific to MxaFI-type methanol dehydrogenases.
Despite decades of structural and biochemical characterization, the precise function of MxaI remains unclear [Web search results, "The MxaI subunits tightly wrap against the MxaF subunits, but the function is unclear"]. Significantly, active MxaF methanol dehydrogenases lacking the beta subunit have been purified and shown to retain catalytic activity, suggesting that MxaI is not absolutely essential for the enzyme's catalytic mechanism per se [Web search results, "Active MxaF MDHs, thus lacking the β subunit, have been purified before, suggesting that this subunit is not always essential for enzyme activity per se"]. This finding has prompted speculation that MxaI may play roles in enzyme stability, proper folding, quaternary structure organization, or regulation rather than direct participation in catalysis.
Gene Organization and the mxa Operon
The mxaI gene is positioned within a large polycistronic operon that encodes the complete machinery for methanol oxidation and associated processes. Early genetic characterization of Methylobacterium extorquens identified the methanol oxidation genes as the mxaFJGIR(S)ACKLD cluster [PMID:8997703, "The region between mxaFJGI and five other methanol oxidation genes (mxaACKLD) was sequenced, revealing one open reading frame (mxaR) and a possible second open reading frame (mxaS)"]. More recent comprehensive analyses have extended this operon to mxaFJGIRSACKLDEHB, encompassing additional regulatory and maturation factors.
Within this operon, the mxaI gene is positioned immediately downstream of mxaG (encoding cytochrome c_L, the electron acceptor for MDH) and mxaJ (encoding the chaperone responsible for PQQ incorporation). The gene order mxaF-mxaJ-mxaG-mxaI reflects a logical organization in which the structural genes for the large and small subunits of MDH (mxaF and mxaI) flank the genes encoding their cognate chaperone (mxaJ) and electron acceptor (mxaG). This genetic architecture facilitates coordinated expression of all components required for functional methanol oxidation.
The genomic organization of the mxa operon supports the hypothesis that MxaI plays an important structural or regulatory role, as its gene has been maintained in close linkage with mxaF throughout the evolution of methylotrophic bacteria. If MxaI were dispensable or merely adventitious, one might expect greater variability in its presence or genomic location across different methylotrophs. Instead, the conservation of mxaI within the mxa operon suggests it provides selective advantage, possibly through enhanced enzyme stability, protection from proteolysis, or optimization of quaternary structure.
Assembly Process and the Role of MxaJ Chaperone
The biogenesis of functional MxaFI methanol dehydrogenase involves a sophisticated assembly pathway mediated by the MxaJ chaperone protein. Although numerous crystal structures of PQQ-dependent MDHs have been resolved over several decades, the molecular mechanism of enzyme assembly remained elusive until recent cryo-electron microscopy studies revealed the detailed process of cofactor incorporation. MxaJ functions as a dedicated chaperone responsible for PQQ incorporation into the nascent MxaF polypeptide [Web search results, "MxaJ is a chaperone responsible for PQQ incorporation"]. Genetic studies demonstrating that mxaJ deletion mutants contain cytochrome c_L but neither the MxaF nor MxaI subunits of methanol dehydrogenase provided early evidence that MxaJ is involved in processing or assembling the enzyme [PMID:8997703, "The mutant contained cytochrome cL (MxaG), but neither subunit of methanol dehydrogenase (MxaF and MxaI)"].
The assembly mechanism follows a strictly ordered pathway with three key steps. First, PQQ binds specifically to the MxaF/MxaJ complex with high affinity (K_D approximately 77 nM), rather than to MxaF alone or to the MxaF/MxaI complex [Web search results, "PQQ can only bind to the MxaF/MxaJ complex (with KD about 77 nM), rather than to the MxaF/MxaI complex or MxaF alone"]. This requirement for the chaperone in productive PQQ binding ensures proper cofactor incorporation before quaternary structure assembly. Second, the incorporation of PQQ into the MxaF/MxaJ complex facilitates the subsequent interaction between MxaI and MxaF, accelerating the release of MxaJ from MxaF and enabling formation of the PQQ-loaded MxaF/MxaI complex [Web search results, "The incorporation of PQQ into the MxaF/MxaJ complex facilitates the interaction between MxaI and MxaF, accelerating the release of MxaJ from MxaF and forming the PQQ-loaded MxaF/MxaI complex"]. Third, MxaJ dissociates from the complex, leaving the mature MxaFI heterotetramer with properly incorporated PQQ and calcium cofactors.
This ordered assembly pathway has important implications for understanding MxaI's role. The observation that MxaI can only efficiently associate with MxaF after PQQ incorporation suggests that MxaI may recognize specific structural features of the PQQ-bound or calcium-coordinated active site. MxaI binding may "lock in" the properly configured active site, stabilizing the catalytically competent conformation and preventing cofactor loss. Alternatively, MxaI might facilitate the subsequent incorporation of the catalytically essential calcium ion, though this hypothesis requires experimental verification.
Cryo-electron microscopy studies have captured intermediate complexes formed by MxaJ and MxaF during MDH maturation, revealing the chaperone-mediated molecular mechanism of cofactor incorporation at atomic resolution [Web search results, "using cryo-electron microscopy, structures of intermediate complexes formed by MxaJ and MxaF during MDH maturation reveal a chaperone-mediated molecular mechanism of cofactor incorporation"]. These structural insights complement genetic and biochemical data, providing a comprehensive picture of how the MxaFI enzyme assembles. The sophisticated quality control mechanisms inherent in this assembly pathway—requiring both proper PQQ incorporation and chaperone release before MxaI can stably associate—ensure that only fully functional enzyme complexes are produced.
The Active Site and Catalytic Mechanism
The catalytic machinery of methanol dehydrogenase resides entirely within the large MxaF subunit. Each MxaF monomer contains one molecule of the prosthetic group pyrroloquinoline quinone (PQQ), which is coordinated to a catalytically essential calcium ion in the active site [Web search results, "The large subunit contains the active-site residues and the PQQ prosthetic group, which is coordinated to a calcium ion in the active site"]. The PQQ cofactor serves as the immediate electron acceptor during methanol oxidation, while the calcium ion plays critical roles in substrate binding, transition state stabilization, and maintaining the proper redox potential of PQQ.
The alpha-subunit of MDH exhibits an unusual eight-fold radial symmetry, with eight β-sheets stabilized by a novel tryptophan docking motif [PMID:7735834, "The α-subunit of MDH has an eight-fold radial symmetry, with its eight β-sheets stabilized by a novel tryptophan docking motif"]. The PQQ cofactor is held in place within this intricate protein scaffold by a coplanar tryptophan residue and by a novel disulfide ring formed between adjacent cysteine residues [PMID:7735834, "The PQQ in the active site is held in place by a coplanar tryptophan and by a novel disulphide ring formed between adjacent cysteines"]. These structural features ensure precise positioning of PQQ for optimal catalytic activity.
MxaI does not directly participate in substrate binding or catalysis. The small subunit does not contain any portion of the PQQ cofactor, the calcium ion, or the active site residues. Instead, each MxaF subunit houses one PQQ molecule and one calcium ion in its catalytic center, representing the catalytic component of the enzyme, while the MxaI subunits fulfill structural or regulatory functions [Web search results, "Each MxaF subunit houses one PQQ molecule and one calcium ion in its catalytic center and represents the catalytic component of the enzyme, while the MxaI subunits tightly wrap against the MxaF subunits"]. The heterotetrameric arrangement positions two active sites within each enzyme complex, potentially allowing for coordinated or cooperative catalysis, though evidence for such cooperativity remains limited.
MDH catalyzes the two-electron oxidation of methanol to formaldehyde according to the reaction: CH₃OH + PQQ → HCHO + PQQH₂. The electrons captured by PQQ during this reaction are subsequently transferred to the enzyme's physiological electron acceptor, cytochrome c_L, which serves as a dedicated one-electron carrier. The enzyme uses pyrroloquinoline quinone to sequentially transfer two electrons to cytochrome c_L, which then enters the electron transport chain [Web search results, "This enzyme uses pyrroloquinoline quinone (PQQ) to sequentially transfer two electrons to cytochrome cL, which enters the electron transport chain"]. This periplasmic electron transfer chain—methanol → MDH → cytochrome c_L → cytochrome c_H → oxidase → oxygen—represents a unique metabolic architecture that enables methylotrophs to derive energy from single-carbon compounds.
Electron Transfer and Interaction with Cytochrome c_L
Methanol dehydrogenase functions as the first component in an unusual periplasmic electron transport chain responsible for methanol oxidation in methylotrophic bacteria [Web search results, "It catalyses the first reaction in an unusual periplasmic electron transport chain responsible for oxidation of methanol to formaldehyde in methylotrophic bacteria"]. Following methanol oxidation and PQQ reduction within the MDH active site, electrons must be efficiently transferred to cytochrome c_L, an atypical c-type cytochrome that serves as the specific and dedicated electron acceptor for MDH [Web search results, "Electrons are transferred from MDH to cytochrome cL, an atypical cytochrome which serves as the specific electron acceptor for MDH"].
The interaction between MDH and cytochrome c_L has been extensively characterized using fluorescence spectroscopy and other biophysical techniques [PMID:7492322, "A fluorescence method is described for direct measurement of the interaction between methanol dehydrogenase (MDH) and its electron acceptor cytochrome cL"] and [PMID:1311606, "The interactions of methanol dehydrogenase (MDH, EC1.1.99.8) with its specific electron acceptor cytochrome cL has been investigated in Methylobacterium extorquens and Methylophilus methylotrophus"]. These studies have revealed that the initial docking of MDH and cytochrome c_L occurs through ionic interactions between lysine residues on the surface of MDH and carboxylate groups on the surface of cytochrome c_L [Web search results, "The initial 'docking' of MDH and cytochrome cL is by way of ionic interactions between lysyl residues on its surface and carboxylate groups on the surface of cytochrome cL"]. This electrostatic recognition mechanism ensures rapid and specific complex formation in the periplasmic space.
Once the MDH-cytochrome c_L complex is formed, electrons are passed one at a time from the reduced PQQH₂ to the heme group of cytochrome c_L [Web search results, "The electrons are passed one at a time to the cytochrome cL, which is bound by carboxylate residues to lysyl residues on MDH"]. Because PQQ undergoes a two-electron reduction during methanol oxidation while cytochrome c_L can accept only one electron at a time, the system must cycle through a semiquinone intermediate state (PQQ•⁻) before complete reoxidation to PQQ. This electron transfer mechanism requires precise geometric arrangement and optimal electronic coupling between the PQQ cofactor and the heme of cytochrome c_L.
The role of MxaI in modulating these electron transfer interactions remains uncertain. The small subunit might influence the electrostatic surface potential of the MDH complex, potentially affecting the rate or specificity of cytochrome c_L binding. Alternatively, MxaI might stabilize conformational states of MxaF that are optimal for electron transfer. The observation that MxaI wraps around MxaF suggests it could potentially influence protein dynamics and conformational flexibility, which often play important roles in intermolecular electron transfer processes. However, direct experimental evidence linking MxaI to electron transfer efficiency or specificity is currently lacking.
Regulation of mxaI Expression and the Lanthanide Switch
Expression of the mxaI gene is coordinately regulated with other components of the mxa operon through a complex regulatory hierarchy involving two-component signal transduction systems. In Methylorubrum extorquens AM1, expression from the mxa promoter depends on a regulatory cascade involving the MxcQE and MxbDM two-component systems [PMID:21873495, "MxcE acts indirectly to activate expression of the mxbDM promoter" and "MxbM functions as a direct activator of the mxa promoter"]. The MxcE response regulator acts indirectly by activating expression of the mxbDM promoter, while the MxbM response regulator functions as a direct activator of the mxa promoter. Both response regulators are required for mxa operon expression [PMID:21873495, "Both the MxcE and MxbM response regulators are required for the expression of the mxa operon"].
A remarkable and unexpected finding revealed that XoxF, a lanthanide-dependent methanol dehydrogenase and structural homolog of MxaF, is essential for mxa operon expression [PMID:21873495, "XoxF is part of a complex regulatory cascade involving the 2-component systems MxcQE and MxbDM, which are required for the expression of the methanol dehydrogenase"]. When both xoxF homologs are deleted, cells cannot express mxa genes or grow on methanol, demonstrating that XoxF participates in the regulatory cascade controlling mxa expression. This discovery revealed the existence of a "lanthanide switch" mechanism in which the availability of lanthanide metals determines which methanol dehydrogenase system is expressed.
The lanthanide switch operates through a sophisticated metal-sensing mechanism. When lanthanides are available, the mxa operon (including mxaI) is downregulated and transcript levels of the xox1 operon genes are upregulated [Web search results, "When lanthanides (Ln) are available, the mxa operon is downregulated and transcript levels of the xox1 operon genes (xoxF1GJ) are upregulated"]. This reciprocal regulation allows M. extorquens to optimize its methanol oxidation system based on metal availability—using the lanthanide-dependent XoxF system when rare earth elements are present, and switching to the calcium-dependent MxaFI system when lanthanides are scarce. The MxbDM two-component system, along with the xoxF1 and xoxF2 genes themselves, has been shown to be required for operation of the lanthanide-switch [Web search results, "The MxbDM two-component system along with the xoxF1 and xoxF2 genes themselves have been shown to be required for operation of the Ln-switch"].
Comprehensive transcriptomic and proteomic analyses have revealed the global changes in gene expression associated with lanthanide homeostasis and methanol metabolism in M. extorquens AM1 [PMID:32704073, cited in web search results as "Lanthanide homeostasis study"]. These studies have identified numerous gene products and processes that contribute to the lanthanide switch, including metal transporters, regulatory proteins, and metabolic enzymes. The coordinate regulation of mxaI with other mxa operon genes ensures that the MxaFI enzyme system is expressed as a complete functional unit only under conditions when it provides selective advantage to the cell.
The biological rationale for the lanthanide switch relates to catalytic efficiency. Lanthanide-dependent XoxF enzymes exhibit superior catalytic properties compared to calcium-dependent MxaFI enzymes, with higher turnover rates and greater affinity for methanol. When lanthanides are available, cells gain competitive advantage by expressing the more efficient enzyme system. However, lanthanides are typically present at very low environmental concentrations, necessitating retention of the calcium-dependent MxaFI system as a metabolic backup. The regulatory complexity ensuring coordinated expression of mxaI with mxaF and the complete mxa operon machinery reflects the importance of this metabolic switch for bacterial fitness in diverse environments.
Comparative Aspects: MxaFI vs. XoxF Enzyme Systems
The discovery of lanthanide-dependent methanol dehydrogenases and the elucidation of the lanthanide switch mechanism have provided new perspectives on the role and significance of MxaI. Structurally, lanthanide-dependent XoxF-type methanol dehydrogenases function as homodimers without any small subunit component, contrasting sharply with the heterotetrameric α₂β₂ architecture of MxaFI-type enzymes. This fundamental difference in quaternary structure raises intriguing questions about why calcium-dependent methanol dehydrogenases require a small subunit while lanthanide-dependent enzymes do not.
Several hypotheses might explain this structural distinction. First, the nature of the metal cofactor might necessitate different structural scaffolding. Calcium ions are relatively large (ionic radius ~100 pm) and bind with different coordination geometries compared to lanthanide ions (ionic radii 86-103 pm for Ln³⁺ ions), potentially requiring different mechanisms for metal retention and active site stabilization. MxaI might provide additional structural constraints that optimize the active site geometry for calcium coordination while also preventing calcium loss from the enzyme.
Second, the evolutionary origins of the two enzyme types might account for their structural differences. XoxF-type enzymes are thought to represent the ancestral form of PQQ-dependent methanol dehydrogenases, while MxaFI-type enzymes may have evolved later as an adaptation to lanthanide scarcity in certain environments. The acquisition of MxaI during this evolutionary process might have been necessary to stabilize MxaF structure when using the less optimal calcium cofactor. The small subunit could compensate for reduced active site stability or altered protein dynamics resulting from calcium substitution for lanthanides.
Third, regulatory mechanisms might differ between the two enzyme systems in ways that necessitate the small subunit. MxaI might play a role in sensing or responding to cellular signals that regulate MxaFI activity post-translationally. For instance, MxaI could potentially serve as a binding site for regulatory proteins or small molecules that modulate enzyme activity in response to metabolic demands or environmental conditions. Such regulatory mechanisms might be less necessary for XoxF enzymes if their expression is primarily controlled at the transcriptional level through the lanthanide switch.
Fourth, enzyme stability and longevity in the periplasmic environment might differ between the two systems. The periplasm presents challenging conditions for protein stability, including variable pH, ionic strength fluctuations, and exposure to proteases. MxaI might enhance the proteolytic stability of MxaF, extend enzyme half-life, or facilitate proper folding in the periplasmic environment. XoxF enzymes, perhaps due to inherent structural differences or different exposure patterns, might not require such additional stabilization.
Experimental evidence supporting these hypotheses remains incomplete. The observation that active MxaF can be purified without MxaI suggests the small subunit is not absolutely required for catalytic activity, arguing against a direct catalytic role. However, measurements of enzyme stability, cofactor retention, and activity under various conditions comparing MxaFI with MxaF alone would help clarify MxaI's contribution to enzyme function. Similarly, domain-swapping experiments examining whether XoxF can functionally interact with MxaI, or whether MxaF can function effectively as a homodimer, would provide insights into the structural requirements for different metal cofactors.
Structural Biology and Crystallographic Studies
Multiple high-resolution crystal structures of methanol dehydrogenase from various methylotrophic organisms have been deposited in the Protein Data Bank, providing detailed atomic-level information about the MxaFI enzyme architecture. Notable structures include the methanol dehydrogenase from Methylobacterium extorquens at 1.94 Å resolution (PDB ID 1H4I) PMID:7735834, from Hyphomicrobium denitrificans (PDB ID 2D0V), from Paracoccus denitrificans (PDB ID 1LRW), and from Methylophilus W3A1 at high resolution (PDB ID 2AD7). These structures consistently reveal the α₂β₂ heterotetrameric architecture with cyclic C2 symmetry [Web search results, "The structures show global stoichiometry of hetero 4-mer - A2B2 with cyclic C2 symmetry"].
The high-resolution structure from M. extorquens revealed that the alpha-subunit adopts an unusual eight-blade β-propeller fold, with each blade containing antiparallel β-sheets. The eight-fold radial symmetry of the α-subunit is stabilized by a novel tryptophan docking motif, wherein tryptophan residues are precisely positioned to form aromatic-aromatic interactions between adjacent blades of the propeller PMID:7735834. At the center of this propeller structure lies the active site, with PQQ held in place by a coplanar tryptophan and a novel disulfide ring formed between adjacent cysteine residues.
Detailed information about the MxaI beta subunit structure has been more difficult to extract from the available literature and crystal structures. The small subunit contains approximately 69 amino acids and wraps around the periphery of the large subunit. However, the precise secondary structure elements of MxaI, the specific contacts it makes with MxaF, and the structural basis for its tight association have not been comprehensively described in the available crystallographic literature. The small size of MxaI and its intimate association with MxaF may have complicated efforts to determine its structure independently or to describe its architecture separately from the large subunit.
The structure of MxaJ, the chaperone protein responsible for PQQ incorporation, has been determined and reveals a periplasmic binding protein-like architecture with unique secondary structural elements [PMID:28295618, "MxaJ structure reveals a periplasmic binding protein-like architecture with unique secondary structural elements"]. This structural information, combined with cryo-EM visualization of MxaJ-MxaF assembly intermediates, has provided insights into how the chaperone facilitates PQQ incorporation. Understanding the structural transitions that occur during the MxaJ-to-MxaI exchange on MxaF could further illuminate MxaI's functional role.
Biotechnological Implications and Applications
Methanol dehydrogenase enzymes, including both MxaFI and XoxF types, have attracted significant interest for biotechnological applications due to their ability to efficiently oxidize methanol and other short-chain alcohols. The development of methanol-based bioprocesses—in which methanol serves as an inexpensive carbon and energy source for microbial production of chemicals, fuels, and materials—depends critically on efficient methanol oxidation. Understanding the structure, assembly, and regulation of MxaFI enzymes is therefore relevant to efforts to engineer methylotrophic bacteria for industrial applications.
The role of MxaI in enzyme assembly and stability has practical implications for heterologous expression systems. Attempts to produce recombinant methanol dehydrogenase in non-native hosts for industrial applications must account for the complex assembly requirements, including PQQ biosynthesis or supplementation, proper calcium incorporation, MxaJ chaperone function, and MxaI co-expression. The observation that mxaJ deletion mutants fail to produce detectable MxaF or MxaI PMID:8997703 underscores the essential role of the chaperone in enzyme biogenesis and suggests that successful heterologous production requires expression of the complete assembly machinery.
The discovery of the lanthanide switch and the characterization of regulatory elements controlling mxa operon expression have enabled synthetic biology approaches to control methanol oxidation. By manipulating the regulatory circuits involving MxcQE, MxbDM, and XoxF, researchers can potentially engineer M. extorquens strains that constitutively express either MxaFI or XoxF regardless of lanthanide availability, or that express both systems simultaneously for increased metabolic capacity. Understanding how mxaI expression is coordinated with the rest of the mxa operon is crucial for such engineering efforts.
Directed evolution and rational design approaches aimed at improving methanol dehydrogenase properties—such as increasing catalytic efficiency, broadening substrate specificity, or enhancing stability—must consider the contributions of both MxaF and MxaI to overall enzyme function. If MxaI plays important roles in stability, folding, or quaternary structure organization, then beneficial mutations in MxaF might be dependent on specific MxaI variants, necessitating co-evolution or co-optimization of both subunits. Conversely, if MxaI serves primarily to compensate for suboptimal properties of calcium-coordinated MxaF, engineering efforts might focus on producing stabilized MxaF variants that eliminate the need for MxaI, potentially simplifying enzyme production and application.
The comparative biochemical properties of MxaFI and XoxF enzymes—including differences in substrate affinity, turnover rate, electron acceptor specificity, and stability—continue to be characterized in various methylotrophic organisms. These comparative studies provide insights into how metal cofactor identity influences enzyme function and may guide efforts to engineer enzymes with designer metal specificity or improved catalytic properties. The structural differences between heterotetrameric MxaFI and homodimeric XoxF represent an intriguing example of how evolution has solved the same biochemical problem (methanol oxidation) using distinct structural solutions depending on cofactor availability.
Outstanding Questions and Future Directions
Despite several decades of research on methanol dehydrogenase, significant questions about MxaI's function remain unresolved. The most fundamental question concerns the precise biochemical role of the small subunit. Why do calcium-dependent methanol dehydrogenases require a small subunit while lanthanide-dependent enzymes do not? What specific functions does MxaI provide to the enzyme complex? Addressing these questions will require careful comparative biochemical studies examining enzyme stability, cofactor retention, catalytic efficiency, and quaternary structure dynamics in the presence and absence of MxaI.
The structural basis for MxaI's tight association with MxaF deserves further investigation. High-resolution structural studies specifically focused on characterizing the MxaI subunit—its secondary structure, its interaction interface with MxaF, and conformational changes that might occur during catalysis—would provide molecular-level insights into its function. Advanced biophysical techniques including hydrogen-deuterium exchange mass spectrometry, cross-linking mass spectrometry, and molecular dynamics simulations could reveal dynamic aspects of MxaI structure and its influence on MxaF conformational flexibility.
The mechanism by which MxaI recognizes PQQ-loaded MxaF during enzyme assembly remains unclear. What structural features of PQQ-bound MxaF does MxaI recognize? Does calcium incorporation precede or follow MxaI binding? Does MxaI play any role in facilitating calcium coordination? Reconstitution experiments using purified components—MxaF, MxaI, MxaJ, PQQ, and calcium—could dissect the assembly pathway step by step and identify the minimal requirements for productive enzyme formation. Such experiments could also determine whether MxaI directly influences the kinetics or thermodynamics of PQQ or calcium binding.
The potential regulatory roles of MxaI warrant investigation. Does the small subunit influence enzyme activity in response to cellular signals? Might MxaI serve as a binding site for regulatory proteins, metabolites, or signaling molecules? Post-translational modifications of MxaI—such as phosphorylation, acetylation, or proteolysis—have not been systematically explored but could represent mechanisms for activity modulation. Proteomic studies examining MxaI modification states under different growth conditions or stress responses might reveal regulatory mechanisms.
The evolutionary history of MxaI and its relationship to the divergence of calcium-dependent and lanthanide-dependent methanol dehydrogenases would benefit from phylogenetic and comparative genomic analyses. When and how did the small subunit emerge during the evolution of methylotrophy? Are there evolutionary intermediates or related proteins that might shed light on MxaI's ancestral function? Examining the distribution of mxaI genes across diverse methylotrophic bacteria and archaea, and correlating this distribution with ecological factors such as lanthanide bioavailability, might reveal selective pressures that shaped the evolution of heterotetrameric enzyme architecture.
Finally, practical applications of methanol dehydrogenase knowledge to biotechnology and synthetic biology depend on comprehensive understanding of all enzyme components. Engineering methylotrophic organisms or heterologous systems for optimized methanol utilization requires consideration of MxaI expression, assembly, and function alongside other components of the methanol oxidation system. Whether MxaI represents an essential component that must be maintained and optimized, or an expendable evolutionary legacy that can be eliminated through protein engineering, remains an open question with significant practical implications.
Conclusion
The mxaI gene encodes the small subunit of calcium-dependent methanol dehydrogenase, a compact 69-amino acid protein that forms an integral structural component of the heterotetrameric α₂β₂ MxaFI enzyme. Despite its small size, MxaI appears to play important roles in enzyme assembly, as evidenced by its ordered incorporation following PQQ loading during the MxaJ-mediated assembly process. The small subunit wraps tightly around the catalytic MxaF large subunit, forming an essentially inseparable complex, yet paradoxically, active enzyme can be isolated without it, suggesting its role relates more to stability, optimization, or regulation than to catalysis per se.
The discovery of lanthanide-dependent XoxF methanol dehydrogenases, which function as homodimers without small subunits, has highlighted the unique nature of MxaI and raised intriguing questions about its evolutionary and functional significance. The coordinate regulation of mxaI with other mxa operon genes through the complex regulatory hierarchy involving MxcQE and MxbDM two-component systems, and its reciprocal expression pattern with xoxF genes in the lanthanide switch, underscore its integration into sophisticated metabolic regulation networks.
As methylotrophy gains increasing attention for biotechnological applications ranging from methanol-based bioprocesses to bioremediation and carbon capture, comprehensive understanding of all methanol dehydrogenase components becomes increasingly important. The MxaI small subunit, though long regarded as enigmatic and functionally unclear, represents an essential piece of the methylotrophic metabolism puzzle. Continued research combining structural biology, biochemistry, molecular genetics, and systems biology approaches promises to finally illuminate the precise functions of this small but intriguing protein and its contributions to one of the most important enzymes in single-carbon metabolism.
Falcon
(mxaI-deep-research-falcon.md)
Falcon
(mxaI-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Functional annotation of moxI / mxaI (UniProt P14775) in Methylorubrum extorquens strain AM1
0) Mandatory identity verification (to avoid gene-symbol ambiguity)
The target protein is UniProt P14775 from Methylorubrum extorquens AM1 (formerly Methylobacterium extorquens AM1), annotated as “Methanol dehydrogenase [cytochrome c] subunit 2” (small/beta subunit) encoded by moxI / mxaI.
Genome-based methylotrophy annotation for AM1 explicitly assigns mxaI as “methanol dehydrogenase small subunit,” with the previous name moxI and links it to the methanol oxidation (MOX) gene module (co-located with mxaF and mxaG) (Chistoserdova et al., 2003, published May 2003, https://doi.org/10.1128/JB.185.10.2980-2987.2003) (chistoserdova2003methylotrophyinmethylobacterium pages 2-3). This matches the UniProt-provided identity and organism context.
1) Key concepts, definitions, and current understanding
1.1 Canonical methanol oxidation in Gram-negative methylotrophs
In Methylorubrum/Methylobacterium methylotrophs, the canonical primary oxidation step is catalyzed by a periplasmic, pyrroloquinoline quinone (PQQ)-dependent methanol dehydrogenase (MDH) that oxidizes methanol → formaldehyde, producing reducing equivalents that enter the periplasmic electron transfer chain (schmidt2010functionalinvestigationof pages 10-14, schmidt2010functionalinvestigationof pages 14-17).
1.2 What MoxI/MxaI is
MxaI (MoxI) is the small (β) subunit of the canonical Ca2+/PQQ-dependent MDH system encoded in the mxa/mox (MOX) gene cluster in AM1 (chistoserdova2003methylotrophyinmethylobacterium pages 2-3). A recent synthesis source describes the canonical MxaFI MDH as an α2β2 heterotetramer consisting of two large catalytic MxaF (α) subunits and two small MxaI (β) subunits (xie2023molecularmechanismsofb pages 13-18, xie2023molecularmechanismsof pages 13-18).
1.3 The MxaFI respiratory electron transfer context (partners)
The periplasmic methanol oxidation chain is commonly described as: MDH → cytochrome cL → cytochrome cH → terminal oxidase → O2 (schmidt2010functionalinvestigationof pages 14-17). In AM1, mxaG is annotated as the cytochrome c component associated with MDH electron transfer (chistoserdova2003methylotrophyinmethylobacterium pages 2-3).
A key mechanistic point from experimental literature is that MDH–cytochrome cL interaction is primarily electrostatic and (in cross-linking/interaction studies summarized there) is associated mainly with the MxaF α-subunit rather than the β-subunit, implying MxaI is not the dominant cytochrome cL-contacting interface (schmidt2010functionalinvestigationof pages 14-17).
2) Functional role of MoxI/MxaI in AM1
2.1 Primary function in the cell
MoxI/MxaI’s primary functional assignment is as the structural/auxiliary small subunit required for the functional MxaFI periplasmic methanol dehydrogenase complex, enabling the organism’s canonical methanol oxidation capacity when Ca/PQQ MDH is expressed/assembled (chistoserdova2003methylotrophyinmethylobacterium pages 2-3, xie2023molecularmechanismsofb pages 13-18, xie2023molecularmechanismsof pages 13-18).
While the catalytic chemistry (EC 1.1.2.7) is attributed to the MDH holoenzyme, the pathway-level reaction catalyzed by the MxaFI system is clearly described as oxidation of methanol to formaldehyde in the periplasm in AM1-like systems (schmidt2010functionalinvestigationof pages 10-14, schmidt2010functionalinvestigationof pages 14-17). Substrate specificity at the complex level is methanol oxidation in this canonical pathway context (schmidt2010functionalinvestigationof pages 10-14).
2.2 Subcellular localization and processing
The canonical PQQ-dependent MDH system is described as periplasmic in Gram-negative methylotrophs including AM1 (schmidt2010functionalinvestigationof pages 10-14, schmidt2010functionalinvestigationof pages 14-17). In addition, PQQ biogenesis and MDH assembly are described such that cofactor/apoenzyme trafficking leads to periplasmic holoenzyme formation, consistent with periplasmic localization of the active enzyme complex containing MxaI (schmidt2010functionalinvestigationof pages 31-34).
Important evidence limitation: within the retrieved corpus, there is not a direct, gene-specific statement describing the MxaI signal peptide or explicit “precursor processing” details for AM1 MxaI itself. Therefore, MxaI’s periplasmic functional location is inferred from its role as a subunit of the periplasmic MxaFI complex rather than from a directly cited MxaI signal-peptide experiment in the retrieved texts (schmidt2010functionalinvestigationof pages 31-34, schmidt2010functionalinvestigationof pages 14-17).
3) Pathways and systems context
3.1 The methanol oxidation module (MOX)
AM1 genome synthesis places mxaI/moxI, mxaF/moxF, and mxaG/moxG together in the primary oxidation (MOX) module/cluster (chistoserdova2003methylotrophyinmethylobacterium pages 2-3). Multiple additional genes in the same broader mxa system are annotated as required for MDH function, including factors “essential for Ca2+ insertion into MDH” and PQQ synthesis genes (chistoserdova2003methylotrophyinmethylobacterium pages 2-3).
3.2 Lanthanide (REE) switch and implications for mxaI
A major development in methylotrophy is the recognition that expression of canonical Ca-dependent MxaFI and alternative lanthanide-dependent XoxF systems is inversely regulated depending on lanthanide availability (“lanthanide switch/REE switch”), with the presence of rare earth elements tending to suppress MxaFI-type expression and promote XoxF-type MDH expression (good2019contrastinginvitro pages 1-2, xie2023molecularmechanismsofb pages 13-18, xie2023molecularmechanismsof pages 13-18).
In AM1 specifically, transcriptomic results under lanthanum conditions are described as upregulation of xox1 and downregulation of mxa genes, consistent with this switch (Good et al., 2019; published 2019; https://doi.org/10.1038/s41598-019-41043-1) (good2019contrastinginvitro pages 1-2). This regulation affects the functional deployment of MxaI because MxaI is part of the MxaFI system that is downregulated under lanthanides.
4) Recent developments (prioritizing 2023–2024 sources where available)
4.1 2023 synthesis: MDH composition and REE switch
A 2023 synthesis source reiterates the canonical architecture that MxaFI MDH comprises two MxaF catalytic subunits and two MxaI small subunits and summarizes the REE switch regulatory logic that suppresses MxaF-type and promotes XoxF-type MDHs under lanthanides (xie2023molecularmechanismsofb pages 13-18, xie2023molecularmechanismsof pages 13-18). While not AM1-primary experimental work, it reflects current consensus framing of the system.
4.2 2024 systems view: methylotroph core gene sets (context)
A 2024 pangenomic analysis of type II methylotrophs provides broader context that methylotroph genomes commonly contain periplasmic and cytoplasmic gene complements (including MDH subunits), supporting the view that periplasmic primary oxidation is a conserved feature (Samanta et al., 2024; published Jun 2024; https://doi.org/10.1128/msystems.00248-24) (samanta2024fromgenometo pages referenced in search results; no direct mxaI-specific statement was extracted in current evidence set).
Note on 2023–2024 AM1-primary literature: In the retrieved documents, direct 2023–2024 primary experimental studies focused specifically on AM1 mxaI (rather than on the broader MDH systems) were limited; thus, some gene-specific details remain supported primarily by foundational AM1 literature and broader recent syntheses.
5) Quantitative data and recent statistics
5.1 Methanol induces canonical MDH system
For AM1, methanol growth is associated with strong induction of the canonical MDH system at the transcription/activity level: mxaF transcription is 5–10× higher on methanol than on succinate, and overall MDH activity is ~6× induced during growth on methanol (Schmidt et al., 2010; published Aug 2010; https://doi.org/10.1099/mic.0.038570-0) (schmidt2010functionalinvestigationof pages 31-34). While this is reported for mxaF/MDH activity rather than mxaI alone, it reflects system-level induction of the MxaFI complex in which MxaI participates.
5.2 Lanthanum improves growth rate and shifts expression away from mxa genes
Addition of exogenous lanthanum is reported to increase methanol growth rate by ~9–12%, and transcriptomics is summarized as showing downregulation of mxa genes with lanthanum methanol growth (Good et al., 2019; https://doi.org/10.1038/s41598-019-41043-1) (good2019contrastinginvitro pages 1-2). Because mxaI is within the mxa cluster, it is expected to follow this switch behavior at the operon/module level.
5.3 Alternative Ln-dependent MDH kinetics (context for when MxaI system is suppressed)
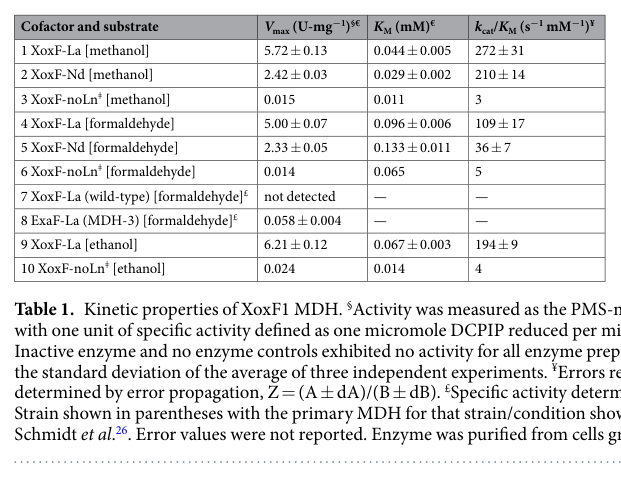
Good et al. (2019) provides kinetic parameters for XoxF1 purified with lanthanides and assayed by DCPIP reduction. The extracted Table 1 (image evidence) reports quantitative values for Vmax, KM, and catalytic efficiency for methanol/formaldehyde/ethanol substrates for La- and Nd-containing enzyme (good2019contrastinginvitro media 8ffe6924, good2019contrastinginvitro media bb7d04f0). This does not measure MxaI directly, but it quantifies the alternative periplasmic oxidation system that becomes dominant under lanthanides and thereby provides a quantitative “replacement context” for MxaI-containing MxaFI under the lanthanide switch.
6) Current applications and real-world implementations
A direct real-world application demonstrated in AM1 involves lanthanide-dependent methylotrophy and lanthanide acquisition/handling machinery (which functionally interfaces with periplasmic alcohol dehydrogenase systems and the MxaFI/XoxF regulatory swap). A variant of AM1 (“evo-HLn”) was shown to grow on methanol using gadolinium sources including the strong chelator Gd-DTPA, with reported similar growth rates for Gd-DTPA vs GdCl3, suggesting potential application in gadolinium recycling/remediation (Good et al., 2022; published Mar 2022; https://doi.org/10.3389/fmicb.2022.820327) (good2022hyperaccumulationofgadolinium pages 9-11).
The same study reports that whole-cell MRI scans showed a statistically significant reduction in T1 relaxation time for cells grown with Gd and that evo-HLn cells grown with Gd displayed T1 values >3-fold less than wild type without lanthanides, illustrating a potential route toward bio-based MRI contrast agents based on intracellular lanthanide accumulation (good2022hyperaccumulationofgadolinium pages 9-11). Although this application is more directly tied to Ln-dependent systems than specifically to MxaI, it reflects real deployment of M. extorquens methylotrophy-associated metal handling and periplasmic alcohol oxidation networks.
7) Expert opinion / authoritative analysis (what the field emphasizes)
Two authoritative themes emerge from the AM1 and broader methylotrophy literature:
- Periplasmic primary oxidation as a respiratory-entry point: canonical MDH systems are framed as periplasmic quinoproteins that feed electrons through dedicated cytochromes to terminal oxidases (schmidt2010functionalinvestigationof pages 14-17, schmidt2010functionalinvestigationof pages 10-14).
- Metal-driven rewiring (lanthanide switch): lanthanide availability reshapes expression from Ca-dependent MxaFI (and thus its β subunit MxaI) toward lanthanide-dependent XoxF systems, with measurable growth and transcriptomic consequences (good2019contrastinginvitro pages 1-2, xie2023molecularmechanismsofb pages 13-18, xie2023molecularmechanismsof pages 13-18).
8) Evidence gaps and recommended next steps (strictly scoped to this protein)
Within the retrieved sources, MxaI-specific experimental details are limited for:
- Signal peptide / precursor processing for AM1 MxaI itself (no direct statement located) (schmidt2010functionalinvestigationof pages 31-34).
- Direct mxaI knockout phenotype in AM1 (no direct statement located).
- MxaI-specific kinetic/biochemical role beyond its defined placement as β-subunit in the α2β2 complex.
Given UniProt’s “precursor” flag and the general periplasmic MDH localization, targeted follow-up should include primary papers on MxaFI maturation/assembly in AM1 and/or direct proteomic N-terminus mapping of MxaI; these were not available in the present retrieved set.
9) Summary table of evidence
The following table consolidates the functional annotation claims for MoxI/MxaI with sources, dates, and URLs.
| Annotation aspect | Key claim | Evidence type | Source with year, DOI/URL | Citation context ID(s) |
|---|---|---|---|---|
| Identity | In Methylorubrum extorquens AM1, mxaI is the methanol dehydrogenase small subunit and is synonymous with moxI; it is located in the methanol oxidation (MOX/mxa) cluster with mxaF and mxaG. | Genomics/review | Chistoserdova et al., 2003, J. Bacteriol. DOI: 10.1128/JB.185.10.2980-2987.2003, https://doi.org/10.1128/JB.185.10.2980-2987.2003 | (chistoserdova2003methylotrophyinmethylobacterium pages 2-3) |
| Complex role | MxaI is the small β subunit of the canonical MxaFI methanol dehydrogenase, which is described as an α2β2 heterotetramer containing two catalytic MxaF subunits and two MxaI subunits. | Review/primary | Schmidt et al., 2010, Microbiology DOI: 10.1099/mic.0.038570-0, https://doi.org/10.1099/mic.0.038570-0; Xie, 2023, URL not available in provided context | (schmidt2010functionalinvestigationof pages 14-17, xie2023molecularmechanismsofb pages 13-18, xie2023molecularmechanismsof pages 13-18) |
| Localization | The canonical PQQ-dependent MDH system of M. extorquens AM1 is periplasmic; therefore MxaI functions as part of a periplasm-localized MxaFI enzyme complex. | Primary/review | Schmidt et al., 2010, Microbiology DOI: 10.1099/mic.0.038570-0, https://doi.org/10.1099/mic.0.038570-0; Xie, 2023, URL not available in provided context | (schmidt2010functionalinvestigationof pages 14-17, schmidt2010functionalinvestigationof pages 10-14, schmidt2010functionalinvestigationof pages 31-34, xie2023molecularmechanismsofb pages 13-18, xie2023molecularmechanismsof pages 13-18) |
| Localization/processing | Direct experimental evidence for the MxaI-specific signal peptide or precursor processing was not identified in the provided contexts; however, the holoenzyme/cofactor assembly and enzyme function are described in the periplasm. | Evidence gap based on available primary/review sources | Schmidt et al., 2010, Microbiology DOI: 10.1099/mic.0.038570-0, https://doi.org/10.1099/mic.0.038570-0 | (schmidt2010functionalinvestigationof pages 37-39, schmidt2010functionalinvestigationof pages 31-34) |
| Pathway role | MxaFI catalyzes the oxidation of methanol to formaldehyde with PQQ as prosthetic group, releasing reducing equivalents into the methanol oxidation respiratory chain. MxaI contributes structurally to this enzyme complex rather than forming the catalytic active site itself. | Primary/review | Schmidt et al., 2010, Microbiology DOI: 10.1099/mic.0.038570-0, https://doi.org/10.1099/mic.0.038570-0; Good et al., 2019, Sci. Rep. DOI: 10.1038/s41598-019-41043-1, https://doi.org/10.1038/s41598-019-41043-1 | (schmidt2010functionalinvestigationof pages 14-17, schmidt2010functionalinvestigationof pages 10-14, good2019contrastinginvitro pages 1-2) |
| Electron acceptor | The physiological electron acceptor for MxaFI is cytochrome cL, encoded by mxaG; electrons then pass to cytochrome c(H) and terminal oxidase. Interaction has been described mainly with the MxaF α-subunit, with little evidence for direct β-subunit contact. | Primary/genomics | Schmidt et al., 2010, Microbiology DOI: 10.1099/mic.0.038570-0, https://doi.org/10.1099/mic.0.038570-0; Chistoserdova et al., 2003, J. Bacteriol. DOI: 10.1128/JB.185.10.2980-2987.2003, https://doi.org/10.1128/JB.185.10.2980-2987.2003 | (schmidt2010functionalinvestigationof pages 14-17, schmidt2010functionalinvestigationof pages 10-14, chistoserdova2003methylotrophyinmethylobacterium pages 2-3) |
| Maturation/cofactor requirements | Functional MxaFI requires PQQ and multiple mxa/mox maturation factors associated with Ca2+ insertion (e.g., mxaA, mxaC, mxaK, mxaL, mxaD). These are encoded in the methanol oxidation gene clusters of AM1. | Genomics/review | Chistoserdova et al., 2003, J. Bacteriol. DOI: 10.1128/JB.185.10.2980-2987.2003, https://doi.org/10.1128/JB.185.10.2980-2987.2003; Schmidt et al., 2010, Microbiology DOI: 10.1099/mic.0.038570-0, https://doi.org/10.1099/mic.0.038570-0 | (chistoserdova2003methylotrophyinmethylobacterium pages 2-3, schmidt2010functionalinvestigationof pages 31-34) |
| Regulation | Under the lanthanide (REE/Ln) switch, expression of canonical MxaFI-type MDH is suppressed while XoxF-type MDH is promoted; in AM1 this applies to the MxaFI system that includes MxaI. | Review/primary | Good et al., 2019, Sci. Rep. DOI: 10.1038/s41598-019-41043-1, https://doi.org/10.1038/s41598-019-41043-1; Xie, 2023, URL not available in provided context | (xie2023molecularmechanismsofb pages 13-18, xie2023molecularmechanismsof pages 13-18, good2019contrastinginvitro pages 1-2) |
| Quantitative data | In methanol + lanthanum conditions, M. extorquens AM1 showed 9–12% higher growth rate than without exogenous lanthanum, while transcriptomics showed upregulation of xox1 and downregulation of mxa genes, consistent with the Ln-switch. | Primary | Good et al., 2019, Sci. Rep. DOI: 10.1038/s41598-019-41043-1, https://doi.org/10.1038/s41598-019-41043-1 | (good2019contrastinginvitro pages 1-2) |
| Quantitative data | During growth on methanol, mxaF transcription is reported as 5- to 10-fold higher than in succinate-grown cells, and overall MDH activity is ~6-fold induced on methanol; this supports substrate-responsive induction of the MxaFI system containing MxaI. | Primary | Schmidt et al., 2010, Microbiology DOI: 10.1099/mic.0.038570-0, https://doi.org/10.1099/mic.0.038570-0 | (schmidt2010functionalinvestigationof pages 31-34) |
| Annotation confidence | The strongest AM1-specific evidence supports annotating P14775/MoxI-MxaI as the small periplasmic β subunit of the Ca/PQQ-dependent MxaFI methanol dehydrogenase in the methanol oxidation pathway; however, MxaI-specific biochemical and processing data remain limited in the provided literature. | Synthesis from available evidence | Supported by the sources above | (chistoserdova2003methylotrophyinmethylobacterium pages 2-3, schmidt2010functionalinvestigationof pages 14-17, schmidt2010functionalinvestigationof pages 10-14, xie2023molecularmechanismsofb pages 13-18, xie2023molecularmechanismsof pages 13-18, schmidt2010functionalinvestigationof pages 31-34, good2019contrastinginvitro pages 1-2) |
Table: This table summarizes literature-supported functional annotation evidence for MoxI/MxaI (UniProt P14775) in Methylorubrum extorquens AM1, including identity, role in the MxaFI complex, localization, pathway placement, electron transfer context, regulation, and quantitative observations. It also highlights where direct evidence is limited for this specific small subunit.
Key references (with publication dates and URLs)
- Chistoserdova L. et al. Journal of Bacteriology (May 2003). “Methylotrophy in Methylobacterium extorquens AM1 from a Genomic Point of View.” https://doi.org/10.1128/JB.185.10.2980-2987.2003 (chistoserdova2003methylotrophyinmethylobacterium pages 2-3)
- Schmidt S. et al. Microbiology (Aug 2010). “Functional investigation of methanol dehydrogenase-like protein XoxF in Methylobacterium extorquens AM1.” https://doi.org/10.1099/mic.0.038570-0 (schmidt2010functionalinvestigationof pages 14-17, schmidt2010functionalinvestigationof pages 31-34)
- Good N.M. et al. Scientific Reports (2019). “Contrasting in vitro and in vivo methanol oxidation activities of lanthanide-dependent alcohol dehydrogenases XoxF1 and ExaF from Methylobacterium extorquens AM1.” https://doi.org/10.1038/s41598-019-41043-1 (good2019contrastinginvitro pages 1-2, good2019contrastinginvitro media 8ffe6924)
- Good N.M. et al. Frontiers in Microbiology (Mar 2022). “Hyperaccumulation of Gadolinium by Methylorubrum extorquens AM1…” https://doi.org/10.3389/fmicb.2022.820327 (good2022hyperaccumulationofgadolinium pages 9-11)
Image-based evidence used
- Extracted kinetic and expression tables from Good et al. (2019), including Table 1 (kinetics) and Table 2 (lanthanide switch expression summary) (good2019contrastinginvitro media 8ffe6924, good2019contrastinginvitro media bb7d04f0).
References
-
(chistoserdova2003methylotrophyinmethylobacterium pages 2-3): Ludmila Chistoserdova, Sung-Wei Chen, Alla Lapidus, and Mary E. Lidstrom. Methylotrophy in methylobacterium extorquens am1 from a genomic point of view. Journal of Bacteriology, 185:2980-2987, May 2003. URL: https://doi.org/10.1128/jb.185.10.2980-2987.2003, doi:10.1128/jb.185.10.2980-2987.2003. This article has 401 citations and is from a peer-reviewed journal.
-
(schmidt2010functionalinvestigationof pages 10-14): Sabrina Schmidt, Philipp Christen, Patrick Kiefer, and Julia A. Vorholt. Functional investigation of methanol dehydrogenase-like protein xoxf in methylobacterium extorquens am1. Microbiology, 156 Pt 8:2575-86, Aug 2010. URL: https://doi.org/10.1099/mic.0.038570-0, doi:10.1099/mic.0.038570-0. This article has 141 citations and is from a peer-reviewed journal.
-
(schmidt2010functionalinvestigationof pages 14-17): Sabrina Schmidt, Philipp Christen, Patrick Kiefer, and Julia A. Vorholt. Functional investigation of methanol dehydrogenase-like protein xoxf in methylobacterium extorquens am1. Microbiology, 156 Pt 8:2575-86, Aug 2010. URL: https://doi.org/10.1099/mic.0.038570-0, doi:10.1099/mic.0.038570-0. This article has 141 citations and is from a peer-reviewed journal.
-
(xie2023molecularmechanismsofb pages 13-18): R Xie. Molecular mechanisms of rare earth element utilization by methane-oxidizing bacteria and protease-producing bacteria. Unknown journal, 2023.
-
(xie2023molecularmechanismsof pages 13-18): R Xie. Molecular mechanisms of rare earth element utilization by methane-oxidizing bacteria and protease-producing bacteria. Unknown journal, 2023.
-
(schmidt2010functionalinvestigationof pages 31-34): Sabrina Schmidt, Philipp Christen, Patrick Kiefer, and Julia A. Vorholt. Functional investigation of methanol dehydrogenase-like protein xoxf in methylobacterium extorquens am1. Microbiology, 156 Pt 8:2575-86, Aug 2010. URL: https://doi.org/10.1099/mic.0.038570-0, doi:10.1099/mic.0.038570-0. This article has 141 citations and is from a peer-reviewed journal.
-
(good2019contrastinginvitro pages 1-2): Nathan M. Good, Riley S. Moore, Carly J. Suriano, and N. Cecilia Martinez-Gomez. Contrasting in vitro and in vivo methanol oxidation activities of lanthanide-dependent alcohol dehydrogenases xoxf1 and exaf from methylobacterium extorquens am1. Scientific Reports, Mar 2019. URL: https://doi.org/10.1038/s41598-019-41043-1, doi:10.1038/s41598-019-41043-1. This article has 91 citations and is from a peer-reviewed journal.
-
(good2019contrastinginvitro media 8ffe6924): Nathan M. Good, Riley S. Moore, Carly J. Suriano, and N. Cecilia Martinez-Gomez. Contrasting in vitro and in vivo methanol oxidation activities of lanthanide-dependent alcohol dehydrogenases xoxf1 and exaf from methylobacterium extorquens am1. Scientific Reports, Mar 2019. URL: https://doi.org/10.1038/s41598-019-41043-1, doi:10.1038/s41598-019-41043-1. This article has 91 citations and is from a peer-reviewed journal.
-
(good2019contrastinginvitro media bb7d04f0): Nathan M. Good, Riley S. Moore, Carly J. Suriano, and N. Cecilia Martinez-Gomez. Contrasting in vitro and in vivo methanol oxidation activities of lanthanide-dependent alcohol dehydrogenases xoxf1 and exaf from methylobacterium extorquens am1. Scientific Reports, Mar 2019. URL: https://doi.org/10.1038/s41598-019-41043-1, doi:10.1038/s41598-019-41043-1. This article has 91 citations and is from a peer-reviewed journal.
-
(good2022hyperaccumulationofgadolinium pages 9-11): Nathan M. Good, Harvey D. Lee, Emily R. Hawker, Morgan Z. Su, Assaf A. Gilad, and N. Cecilia Martinez-Gomez. Hyperaccumulation of gadolinium by methylorubrum extorquens am1 reveals impacts of lanthanides on cellular processes beyond methylotrophy. Frontiers in Microbiology, Mar 2022. URL: https://doi.org/10.3389/fmicb.2022.820327, doi:10.3389/fmicb.2022.820327. This article has 38 citations and is from a peer-reviewed journal.
-
(schmidt2010functionalinvestigationof pages 37-39): Sabrina Schmidt, Philipp Christen, Patrick Kiefer, and Julia A. Vorholt. Functional investigation of methanol dehydrogenase-like protein xoxf in methylobacterium extorquens am1. Microbiology, 156 Pt 8:2575-86, Aug 2010. URL: https://doi.org/10.1099/mic.0.038570-0, doi:10.1099/mic.0.038570-0. This article has 141 citations and is from a peer-reviewed journal.
Artifacts
Citations
- chistoserdova2003methylotrophyinmethylobacterium pages 2-3
- schmidt2010functionalinvestigationof pages 14-17
- schmidt2010functionalinvestigationof pages 10-14
- schmidt2010functionalinvestigationof pages 31-34
- good2019contrastinginvitro pages 1-2
- good2022hyperaccumulationofgadolinium pages 9-11
- xie2023molecularmechanismsofb pages 13-18
- xie2023molecularmechanismsof pages 13-18
- schmidt2010functionalinvestigationof pages 37-39
- cytochrome c
- https://doi.org/10.1128/JB.185.10.2980-2987.2003
- https://doi.org/10.1038/s41598-019-41043-1
- https://doi.org/10.1128/msystems.00248-24
- https://doi.org/10.1099/mic.0.038570-0
- https://doi.org/10.3389/fmicb.2022.820327
- https://doi.org/10.1099/mic.0.038570-0;
- https://doi.org/10.1128/JB.185.10.2980-2987.2003;
- https://doi.org/10.1038/s41598-019-41043-1;
- https://doi.org/10.1128/jb.185.10.2980-2987.2003,
- https://doi.org/10.1099/mic.0.038570-0,
- https://doi.org/10.1038/s41598-019-41043-1,
- https://doi.org/10.3389/fmicb.2022.820327,