Deep Research
Falcon
(xoxF1-deep-research-falcon.md)
Falcon
(xoxF1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Functional annotation report: xoxF1 (UniProt C5B120; locus MexAM1_META1p1740) in Methylorubrum extorquens AM1
0. Target verification (gene/protein identity)
Multiple AM1-specific studies explicitly identify xoxF1 (MexAM1_META1p1740) as a lanthanide (Ln)-dependent, pyrroloquinoline quinone (PQQ)-dependent methanol dehydrogenase (MDH) that is periplasmic and is induced as part of the lanthanide switch in AM1. (vu2016lanthanidedependentregulationof pages 1-6, roszczenkojasinska2020geneproductsand pages 4-5, roszczenkojasinska2020geneproductsand pages 5-6)
1. Key concepts and definitions (current understanding)
1.1 Lanthanide-dependent methanol dehydrogenase (XoxF)
In methylotrophic Proteobacteria, methanol oxidation is mediated by periplasmic PQQ-dependent alcohol dehydrogenases. In AM1, XoxF-type MDHs (including XoxF1) are active when Ln(III) (e.g., La(III), Ce(III), Pr(III), Nd(III)) are available and oxidize methanol → formaldehyde, contrasting with the canonical Ca-dependent MDH system (MxaFI). (vu2016lanthanidedependentregulationof pages 1-6, roszczenkojasinska2020geneproductsand pages 4-5)
Mechanistically, lanthanides in the active site act as Lewis acids that facilitate hydride transfer to PQQ during alcohol oxidation. (roszczenkojasinska2020geneproductsand pages 1-4)
1.2 The “lanthanide switch”
The lanthanide switch refers to inverse transcriptional regulation of methanol oxidation systems: with Ln present, xox genes are upregulated and mxa genes are repressed; with Ln absent, mxaFI dominates. In AM1, this switch is highly sensitive to Ln concentration and involves additional regulatory complexity (including cross-dependence where xoxF genes affect mxa expression). (vu2016lanthanidedependentregulationof pages 1-6)
1.3 Pathway context: periplasmic oxidation + electron transfer
XoxF enzymes operate in the periplasm and typically couple to c-type cytochrome electron acceptors. In AM1, XoxG (a cytochrome cL-like protein) is described as transferring electrons from PQQ to downstream components of the respiratory chain, and XoxJ is a periplasmic binding protein implicated in XoxF activation. (roszczenkojasinska2020geneproductsand pages 4-5, roszczenkojasinska2020geneproductsand pages 5-6)
2. xoxF1 functional annotation (reaction, cofactors, specificity, localization)
2.1 Primary catalytic function and reaction
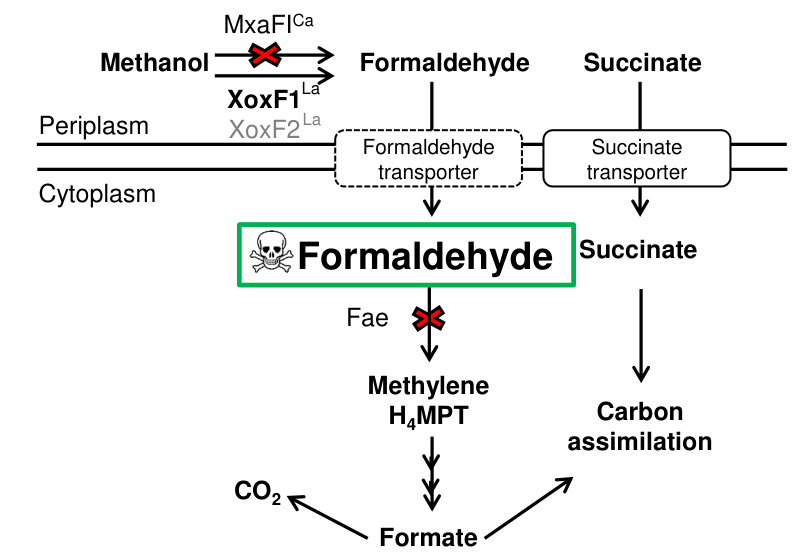
Primary function: XoxF1 is a lanthanide-dependent methanol dehydrogenase that oxidizes methanol to formaldehyde in AM1. This is supported by genetic and physiological evidence (formaldehyde toxicity phenotypes in fae backgrounds and growth dependencies on xox genes under Ln). (roszczenkojasinska2020geneproductsand pages 5-6, roszczenkojasinska2020geneproductsand pages 4-5)
Quantitative enzymology (AM1 XoxF1): Purified XoxF1 exhibited a low in vitro methanol oxidation activity in one assay context (Vmax = 0.015 U/mg). (vu2016lanthanidedependentregulationof pages 1-6)
Family context (supporting specificity/assay behavior): XoxF-family enzymes (including M. extorquens XoxF5 variants) show measurable methanol activity and PQQ-associated UV–Vis signatures, with reported kinetic parameters for one M. extorquens XoxF5 protein of Vmax 12.5 ± 0.2 U/mg and Km 21 ± 1 mM (methanol assay conditions as in that study). These values support the broader functional annotation of XoxF proteins as methanol dehydrogenases but are not specific to AM1 XoxF1. (huang2019rareearthelement pages 6-8)
2.2 Cofactors and metal dependence
XoxF1 is a PQQ-dependent dehydrogenase whose active site requires Ln(III) rather than Ca(II), distinguishing it from Ca-dependent MxaFI. (vu2016lanthanidedependentregulationof pages 1-6)
Supporting biochemical evidence includes that XoxF purified from Ln-grown cells was reported as a dimer incorporating approximately ~1.24 atoms of La and no Ca, and that promoter/growth responses are sensitive to multiple Ln (La, Ce, Pr, Nd; Sm weaker). (vu2016lanthanidedependentregulationof pages 6-9, vu2016lanthanidedependentregulationof pages 1-6)
Reconstitution assays in the XoxF literature also emphasize dual requirement for PQQ and Ln (example conditions: 200 nM XoxF-MDH, 200 nM Ln, 200 nM PQQ, 50 mM methanol; DCPIP/PES readout), reinforcing the cofactor model. (singer2022radioactiveelementscurium pages 41-42)
2.3 Subcellular localization
AM1 xoxF1 is repeatedly described/annotated as periplasmic, consistent with the general statement that methylotrophic PQQ-ADHs are periplasmic enzymes. (vu2016lanthanidedependentregulationof pages 1-6, roszczenkojasinska2020geneproductsand pages 4-5)
A schematic model explicitly places XoxF1 in the periplasm catalyzing methanol oxidation in the presence of lanthanum and depicts a connected lanthanide transport pathway. (roszczenkojasinska2019lanthanidetransportstorage media 5b494616, roszczenkojasinska2019lanthanidetransportstorage media 63d6a23c)
2.4 Required partner proteins and supporting processes
Genetic evidence indicates that XoxF1-dependent methanol oxidation requires:
- XoxG (cytochrome c) and XoxJ (periplasmic binding protein): loss of either phenocopies loss of xoxF1/xoxF2 for Ln-dependent methanol growth. (roszczenkojasinska2020geneproductsand pages 5-6)
- PQQ biosynthesis and cytochrome c biogenesis/heme export: disrupting these processes eliminates methanol growth regardless of La presence, consistent with a periplasmic PQQ dehydrogenase feeding electrons into cytochrome c-based respiratory components. (roszczenkojasinska2020geneproductsand pages 5-6)
3. Regulation and physiology in AM1 (lanthanide switch; quantitative data)
3.1 Lanthanide switch thresholds
In AM1, methanol growth and promoter responses occur over a wide La range:
- Maximal growth rate and yield at ≥1 μM La.
- Growth still detectable at ~2.5 nM La (slower).
- Intermediate co-expression of mxa and xox1 promoters at ~50–100 nM La, reflecting a graded switch. (vu2016lanthanidedependentregulationof pages 1-6)
3.2 Growth phenotypes of xoxF mutants (AM1)
In La-containing methanol medium:
- WT and an mxaF strain show similar growth rate ~0.16 ± 0.01 h⁻1.
- ΔxoxF1 shows a lag (~6–9 h) and reduced growth rate 0.07 ± 0.00 h⁻1.
- ΔxoxF1 ΔxoxF2 shows stronger defect (lag ~6 h; growth rate 0.04 ± 0.01 h⁻1). (roszczenkojasinska2020geneproductsand pages 6-7)
3.3 Concentration-dependent growth with different lanthanides (2024 dataset)
A 2024 study of AM1 methanol growth (125 mM methanol) reported doubling time and yield dependence on metal identity and concentration (0.1–100 μM):
- With La³⁺, fastest doubling ~4.9 h at 1 μM; higher concentrations (100 μM) slowed growth (doubling ~9.4 h).
- With Ce³⁺, fastest doubling ~2.9 h at 1 μM; 100 μM Ce³⁺ was inhibitory (doubling ~12.4 h, lower max OD). (warters2024widespreadbacterialuseb pages 18-23, warters2024widespreadbacterialuse pages 18-23)
These results support the concept that light lanthanides can enable rapid methylotrophic growth in AM1 but that excessive concentrations may impose stress or mis-metallation effects. (warters2024widespreadbacterialuseb pages 18-23)
4. Biochemical pathway integration: methanol oxidation, formaldehyde handling, and Ln homeostasis
4.1 Product fate and connection to downstream C1 metabolism
AM1 XoxF1/2 produce formaldehyde from methanol, which is toxic if it accumulates (a feature leveraged experimentally using a fae mutant background). (roszczenkojasinska2020geneproductsand pages 4-5, roszczenkojasinska2020geneproductsand pages 5-6)
In contrast, ExaF (a distinct lanthanide-dependent alcohol dehydrogenase in AM1’s broader network) is reported to oxidize methanol to formate, providing a bypass route that changes where formaldehyde burden appears in metabolism. (roszczenkojasinska2020geneproductsand pages 4-5, roszczenkojasinska2020geneproductsand pages 5-6)
4.2 Electron transfer chain context
XoxF1 is linked to cytochrome c-based electron transfer:
- XoxG is described as a cytochrome that accepts electrons from XoxF and passes them onward to the electron transport chain.
- XoxD (MxaD homolog) and XoxJ are implicated in supporting XoxF activation and/or interaction with electron acceptors in XoxF systems. (roszczenkojasinska2020geneproductsand pages 4-5, wegner2019lanthanidedependentmethylotrophsof pages 10-12)
4.3 Lanthanide uptake/utilization (lut system) and intracellular storage
AM1 requires dedicated Ln acquisition/trafficking systems for robust Ln-dependent methylotrophy:
- A TonB-dependent receptor (LutH) and an ABC transporter (LutAEF) are implicated in moving lanthanides from outside the cell to the cytoplasm, with evidence that mutants in these loci disrupt Ln-dependent growth and alter Ln localization. (roszczenkojasinska2020geneproductsand pages 4-5, roszczenkojasinska2019lanthanidetransportstorage pages 1-5)
- Microscopy/EDS evidence shows AM1 can store lanthanides as phosphate-containing deposits. One dataset reports crystalline La–phosphate deposits with approximate composition La 22.2 ± 1.0%, P 15.1 ± 2.1%, O 51.1 ± 1.9% by EDS. (roszczenkojasinska2019lanthanidetransportstorage pages 15-18)
- Stored Ln can transiently support growth after “preloading” (growth rate 0.15 ± 0.00 h⁻1), but growth collapses after depletion (0.01 ± 0.0 h⁻1), consistent with a finite intracellular Ln reserve supporting XoxF activity. (roszczenkojasinska2019lanthanidetransportstorage pages 15-18)
5. Recent developments (prioritizing 2023–2024)
5.1 2023: scalable REE leaching and recovery using AM1 (real-world implementation)
Good et al. (published Dec 2023 in Environmental Science & Technology; https://doi.org/10.1021/acs.est.3c06775) developed an AM1-based platform that links methanol metabolism (and thus Ln-dependent MDH function) to rare earth element recovery from Nd magnet swarf and other complex sources. (good2023scalableandconsolidated pages 4-5)
Key quantitative results include:
- Feedstock composition (magnet swarf): 68.0% Fe, 26.7% Nd, 4.35% Pr, 3.34% Dy. (good2023scalableandconsolidated pages 4-5)
- Abiotic citrate leaching increases total metals in solution (e.g., with 5 mM citrate: REE 8.2 ± 0.1 ppm, Fe 23.7 ± 2.5 ppm; with 15 mM citrate: REE 58.7 ± 0.6 ppm, Fe 142.0 ± 3.2 ppm), showing that organic acids can increase leaching but not necessarily selectivity. (good2023scalableandconsolidated pages 4-5)
- Bioaccumulation selectivity: inoculated cultures scavenged most leached REE into biomass, leaving supernatants largely Fe-dominated (~98% Fe in one condition), while intracellular metals were strongly REE-enriched (example: intracellular Fe only 2.0% of measured intracellular metals). (good2023scalableandconsolidated pages 4-5)
- Engineered uptake/storage improvements: expression of a lanthanophore-related locus (“mll”) increased intracellular REE to 80 mg Nd/g DW (plus 15 mg Pr/g DW, 8 mg Dy/g DW) with minimal Fe uptake; deletion of ppx increased to 202 mg Nd/g DW (~5.5-fold increase). (good2023scalableandconsolidated pages 6-7)
- Scalability and projected performance: with citrate-fed batch reaching OD ≈20 in a 0.75 L bioreactor, authors projected potential Nd recovery 1.3–2.1 g Nd/L (at 1% swarf pulp density, assuming 1% Nd in swarf), and reported 10 L scalability with similar yields. (good2023scalableandconsolidated pages 6-7, good2023scalableandconsolidated pages 4-5)
These results establish AM1 (and by extension its Ln-dependent methanol oxidation machinery including XoxF-type enzymes) as an applied chassis for REE bioleaching/bioaccumulation under mild conditions. (good2023scalableandconsolidated pages 6-7, good2023scalableandconsolidated pages 4-5)
5.2 2024: ecological distribution and biogeochemical context
Voutsinos et al. (published Feb 2024 in BMC Biology; https://doi.org/10.1186/s12915-024-01841-0) performed metagenomics across a weathered granite-to-soil profile and found that XoxF-type MDHs can be the only MDH class detectable in these communities. (voutsinos2024weatheredgranitesand pages 10-12)
Key statistics:
- 411 distinct MDH sequences were recovered; all were XoxF-type, and no Ca-dependent MxaF MDHs were found. (voutsinos2024weatheredgranitesand pages 2-4)
- Clade distribution: XoxF3 (340 sequences) dominated; XoxF5 (63) was less common; 8 unassigned. (voutsinos2024weatheredgranitesand pages 2-4, voutsinos2024weatheredgranitesand pages 4-7)
- In 136 high-quality dereplicated genomes, XoxF3 appeared in 43 genomes and XoxF5 in 7 genomes; again, no MxaF was found. (voutsinos2024weatheredgranitesand pages 4-7)
- Geochemistry: lanthanides were highest in lightly weathered granite (e.g., 427 ppm at one site) and decreased along the profile (to 80 ppm in soil), and lanthanide phosphate crystals were observed and inferred to dissolve during weathering—consistent with dynamic bioavailability of Ln for XoxF-dependent metabolism. (voutsinos2024weatheredgranitesand pages 7-10)
The study also identified candidate metallophore biosynthetic clusters and TonB-dependent transporters potentially linked to mobilizing poorly soluble lanthanide phosphates, providing a mechanistic bridge between rock weathering, Ln bioavailability, and XoxF-based methanol oxidation. (voutsinos2024weatheredgranitesand pages 2-4, voutsinos2024weatheredgranitesand pages 7-10)
6. Expert synthesis and authoritative interpretations
6.1 Why XoxF1 is central in AM1 physiology
AM1’s xoxF1 system is not only catalytic but also regulatory. Multiple studies report that xoxF genes influence expression of the Ca-dependent mxaFI system, implying a role in coordinating methanol oxidation capacity with metal availability (a hallmark of the lanthanide switch). (vu2016lanthanidedependentregulationof pages 1-6, roszczenkojasinska2020geneproductsand pages 5-6)
6.2 Metal selectivity and “light lanthanide preference”
Across AM1 and other methylotrophs, light lanthanides (La–Nd; sometimes Sm weakly) are most consistently compatible with growth and/or activity. AM1-specific work reports that both XoxF and an additional Ln-dependent methanol oxidation system can use La/Ce/Pr/Nd and to some extent Sm, while broader analyses describe growth limitations with heavier lanthanides in typical strains. (vu2016lanthanidedependentregulationof pages 1-6, good2022hyperaccumulationofgadolinium pages 1-2)
7. Visual evidence (pathway and localization)
A schematic model explicitly places XoxF1 in the periplasm (with La indicated) and depicts a Lut-based lanthanide transport pathway spanning the outer membrane, periplasm, and inner membrane, providing visual support for subcellular localization and pathway integration. (roszczenkojasinska2019lanthanidetransportstorage media 5b494616, roszczenkojasinska2019lanthanidetransportstorage media 63d6a23c)
8. Evidence map (quick reference)
| Topic | Key finding (1-2 sentences) | Quantitative data (if any) | Source (first author year, journal) | URL | Citation ID |
|---|---|---|---|---|---|
| Reaction | In Methylorubrum extorquens AM1, xoxF1 encodes a lanthanide-dependent methanol dehydrogenase that oxidizes methanol to formaldehyde. Genetic evidence links XoxF1/XoxF2 activity to formaldehyde accumulation phenotypes during methanol growth with lanthanides. | Purified XoxF1 methanol oxidation activity reported as Vmax = 0.015 U/mg. | Vu 2016, Journal of Bacteriology; Roszczenko-Jasińska 2020, Scientific Reports | https://doi.org/10.1128/jb.00937-15 ; https://doi.org/10.1038/s41598-020-69401-4 | (vu2016lanthanidedependentregulationof pages 1-6, roszczenkojasinska2020geneproductsand pages 5-6) |
| Cofactors | XoxF1 is a PQQ-dependent alcohol dehydrogenase that requires lanthanides rather than Ca²⁺ in the active site. Lanthanides act with PQQ in the catalytic cofactor complex and facilitate alcohol oxidation. | XoxF purified from lanthanide-grown cells incorporated ~1.24 atoms of La and lacked Ca; reconstitution assays used 200 nM enzyme, 200 nM Ln, 200 nM PQQ, 50 mM methanol. | Vu 2016, Journal of Bacteriology; Singer 2022, ChemRxiv | https://doi.org/10.1128/jb.00937-15 ; https://doi.org/10.26434/chemrxiv-2022-zn3t4 | (vu2016lanthanidedependentregulationof pages 1-6, singer2022radioactiveelementscurium pages 41-42, vu2016lanthanidedependentregulationof pages 6-9) |
| Localization | XoxF1 is described as a periplasmic PQQ-dependent alcohol dehydrogenase, consistent with methylotrophic PQQ-ADHs generally being periplasmic enzymes. It functions in a periplasm-associated lanthanide oxidation system. | No direct localization constant reported; schematic evidence places XoxF1 in the periplasm. | Roszczenko-Jasińska 2020, Scientific Reports; Roszczenko-Jasińska 2019, bioRxiv | https://doi.org/10.1038/s41598-020-69401-4 ; https://doi.org/10.1101/647677 | (roszczenkojasinska2020geneproductsand pages 1-4, roszczenkojasinska2020geneproductsand pages 4-5, roszczenkojasinska2019lanthanidetransportstorage pages 48-53, roszczenkojasinska2019lanthanidetransportstorage media 5b494616) |
| Regulation | AM1 exhibits a lanthanide switch in which lanthanides upregulate the xox1 operon and repress the Ca-dependent mxa operon. XoxF proteins also appear necessary for proper expression of MxaFI, indicating a regulatory role beyond catalysis. | Growth reaches maximum rate/yield at ≥1 μM La; growth still occurs at ~2.5 nM La but more slowly; intermediate mxa/xox1 co-expression occurs at 50–100 nM La. | Vu 2016, Journal of Bacteriology; Roszczenko-Jasińska 2020, Scientific Reports | https://doi.org/10.1128/jb.00937-15 ; https://doi.org/10.1038/s41598-020-69401-4 | (vu2016lanthanidedependentregulationof pages 1-6, roszczenkojasinska2020geneproductsand pages 5-6, roszczenkojasinska2020geneproductsand pages 6-7) |
| Growth data | Loss of xoxF1 reduces methanol-growth performance in lanthanide-containing medium, and double loss of xoxF1/xoxF2 causes a stronger defect. Wild type and mxaF mutants grow similarly under La³⁺, showing XoxF-mediated methanol oxidation can support growth. | WT and mxaF growth rates ~0.16 ± 0.01 h⁻¹; xoxF1 0.07 ± 0.00 h⁻¹ with 6–9 h lag; xoxF1 xoxF2 0.04 ± 0.01 h⁻¹ with ~6 h lag. | Roszczenko-Jasińska 2020, Scientific Reports | https://doi.org/10.1038/s41598-020-69401-4 | (roszczenkojasinska2020geneproductsand pages 6-7) |
| Metal specificity | AM1 XoxF systems primarily use light lanthanides. La, Ce, Pr, and Nd support the lanthanide switch and methanol growth, while Sm is less effective and heavier lanthanides generally do not support growth in the wild type. | Light Ln range noted as La–Sm (atomic numbers 57–62); growth with Sm is slower; heavier Ln usually do not support growth. | Vu 2016, Journal of Bacteriology; Good 2022, Frontiers in Microbiology | https://doi.org/10.1128/jb.00937-15 ; https://doi.org/10.3389/fmicb.2022.820327 | (vu2016lanthanidedependentregulationof pages 1-6, good2022hyperaccumulationofgadolinium pages 1-2) |
| Partners | XoxF1 functions with accessory proteins including XoxG, a cytochrome c electron acceptor, and XoxJ, a periplasmic binding protein; both are required for efficient XoxF-dependent methanol oxidation. PQQ biosynthesis and cytochrome c biogenesis genes are also essential for the pathway. | Transposon screen recovered >600 mutants; genes identified independently ≥4 times were prioritized. | Roszczenko-Jasińska 2020, Scientific Reports; Roszczenko-Jasińska 2019, bioRxiv | https://doi.org/10.1038/s41598-020-69401-4 ; https://doi.org/10.1101/647677 | (roszczenkojasinska2020geneproductsand pages 5-6, roszczenkojasinska2019lanthanidetransportstorage pages 15-18, wegner2019lanthanidedependentmethylotrophsof pages 10-12) |
| Applications | The xoxF1-centered lanthanide methylotrophy system underpins AM1-based rare-earth bioaccumulation, biomining, and potentially MRI/bioremediation technologies. Engineering lanthanide uptake, lanthanophore production, PQQ biosynthesis, and phosphate metabolism enhances recovery from waste sources. | evo-HLn hyperaccumulated Gd ~36-fold; AM1-based REE recovery was scaled to 10 L; whole-cell MRI contrast was observed in Gd-hyperaccumulating cells. | Good 2022, Frontiers in Microbiology; Good 2023, Environmental Science & Technology | https://doi.org/10.3389/fmicb.2022.820327 ; https://doi.org/10.1021/acs.est.3c06775 | (good2022hyperaccumulationofgadolinium pages 1-2) |
| Comparative enzymology | Broader XoxF literature supports that REE-dependent XoxF enzymes are active methanol dehydrogenases with PQQ-like spectral signatures, alkaline assay optima, and measurable methanol activities, reinforcing the family assignment for AM1 XoxF1. These data are supportive but not AM1 xoxF1-specific kinetics. | Example XoxF5_M.e.1 kinetics: Vmax 12.5 ± 0.2 U mg⁻¹, Km 21 ± 1 mM, Keff 1,252 ± 135 s⁻¹ mM⁻¹; UV-Vis maximum ~355 nm. | Huang 2019, The ISME Journal | https://doi.org/10.1038/s41396-019-0414-z | (huang2019rareearthelement pages 6-8) |
| Recent ecosystem-scale growth data | A 2024 ecosystem-oriented study using AM1 showed concentration-dependent growth responses to Ca²⁺ and individual lanthanides, illustrating that moderate light-lanthanide concentrations can support fast methanol growth while higher concentrations can become inhibitory. | With 125 mM methanol: La³⁺ fastest doubling 4.9 h at 1 μM; Ce³⁺ fastest doubling 2.9 h at 1 μM; 100 μM Ce³⁺ slowed doubling to 12.4 h; Ca²⁺ fastest doubling 3.7 h at 10 μM. | Warters 2024, unknown journal/thesis source | Not available from citation metadata | (warters2024widespreadbacterialuseb pages 18-23, warters2024widespreadbacterialuse pages 18-23, warters2024widespreadbacterialusea pages 18-23) |
Table: This table compiles key functional-annotation evidence for xoxF1 (UniProt C5B120) in Methylorubrum extorquens AM1, covering reaction chemistry, cofactors, localization, regulation, growth phenotypes, partner proteins, metal specificity, and applications. It is designed as a quick-reference evidence map tied directly to available citation IDs.
9. Key takeaways for functional annotation
- Molecular function: lanthanide- and PQQ-dependent methanol dehydrogenase; catalyzes methanol → formaldehyde in AM1’s periplasm. (vu2016lanthanidedependentregulationof pages 1-6, roszczenkojasinska2020geneproductsand pages 4-5)
- Cofactors: requires PQQ + Ln(III) (La/Ce/Pr/Nd most supported); distinct from Ca-dependent MxaFI. (vu2016lanthanidedependentregulationof pages 1-6, vu2016lanthanidedependentregulationof pages 6-9)
- Localization: periplasmic; connected to cytochrome c electron transfer via XoxG and requires accessory XoxJ for function. (roszczenkojasinska2020geneproductsand pages 4-5, roszczenkojasinska2020geneproductsand pages 5-6, roszczenkojasinska2019lanthanidetransportstorage media 5b494616)
- Biological process: methanol oxidation in methylotrophic growth; tightly integrated with formaldehyde handling (toxicity in fae background) and Ln uptake/storage (lut system; phosphate-like deposits). (roszczenkojasinska2020geneproductsand pages 4-5, roszczenkojasinska2019lanthanidetransportstorage pages 15-18)
- Regulation: part of the lanthanide switch controlling xox vs mxa expression; switch responds to nM–µM Ln and can show intermediate promoter co-expression. (vu2016lanthanidedependentregulationof pages 1-6)
- Applications (2023–2024): AM1-based REE recovery at bench scale up to 10 L, with engineered strains achieving high Nd accumulation (e.g., 202 mg Nd/g DW) and projected g/L-scale recovery under high-density cultivation; ecosystem studies show XoxF-based methylotrophy may dominate in certain mineral-rich environments. (good2023scalableandconsolidated pages 6-7, voutsinos2024weatheredgranitesand pages 7-10)
References
-
(vu2016lanthanidedependentregulationof pages 1-6): Huong N. Vu, Gabriel A. Subuyuj, Srividhya Vijayakumar, Nathan M. Good, N. Cecilia Martinez-Gomez, and Elizabeth Skovran. Lanthanide-dependent regulation of methanol oxidation systems in methylobacterium extorquens am1 and their contribution to methanol growth. Journal of Bacteriology, 198:1250-1259, Apr 2016. URL: https://doi.org/10.1128/jb.00937-15, doi:10.1128/jb.00937-15. This article has 227 citations and is from a peer-reviewed journal.
-
(roszczenkojasinska2020geneproductsand pages 4-5): Paula Roszczenko-Jasińska, Huong N. Vu, Gabriel A. Subuyuj, Ralph Valentine Crisostomo, James Cai, Nicholas F. Lien, Erik J. Clippard, Elena M. Ayala, Richard T. Ngo, Fauna Yarza, Justin P. Wingett, Charumathi Raghuraman, Caitlin A. Hoeber, Norma C. Martinez-Gomez, and Elizabeth Skovran. Gene products and processes contributing to lanthanide homeostasis and methanol metabolism in methylorubrum extorquens am1. Scientific Reports, Jul 2020. URL: https://doi.org/10.1038/s41598-020-69401-4, doi:10.1038/s41598-020-69401-4. This article has 98 citations and is from a peer-reviewed journal.
-
(roszczenkojasinska2020geneproductsand pages 5-6): Paula Roszczenko-Jasińska, Huong N. Vu, Gabriel A. Subuyuj, Ralph Valentine Crisostomo, James Cai, Nicholas F. Lien, Erik J. Clippard, Elena M. Ayala, Richard T. Ngo, Fauna Yarza, Justin P. Wingett, Charumathi Raghuraman, Caitlin A. Hoeber, Norma C. Martinez-Gomez, and Elizabeth Skovran. Gene products and processes contributing to lanthanide homeostasis and methanol metabolism in methylorubrum extorquens am1. Scientific Reports, Jul 2020. URL: https://doi.org/10.1038/s41598-020-69401-4, doi:10.1038/s41598-020-69401-4. This article has 98 citations and is from a peer-reviewed journal.
-
(roszczenkojasinska2020geneproductsand pages 1-4): Paula Roszczenko-Jasińska, Huong N. Vu, Gabriel A. Subuyuj, Ralph Valentine Crisostomo, James Cai, Nicholas F. Lien, Erik J. Clippard, Elena M. Ayala, Richard T. Ngo, Fauna Yarza, Justin P. Wingett, Charumathi Raghuraman, Caitlin A. Hoeber, Norma C. Martinez-Gomez, and Elizabeth Skovran. Gene products and processes contributing to lanthanide homeostasis and methanol metabolism in methylorubrum extorquens am1. Scientific Reports, Jul 2020. URL: https://doi.org/10.1038/s41598-020-69401-4, doi:10.1038/s41598-020-69401-4. This article has 98 citations and is from a peer-reviewed journal.
-
(huang2019rareearthelement pages 6-8): Jing Huang, Zheng Yu, Joseph Groom, Jan-Fang Cheng, Angela Tarver, Yasuo Yoshikuni, and Ludmila Chistoserdova. Rare earth element alcohol dehydrogenases widely occur among globally distributed, numerically abundant and environmentally important microbes. The ISME Journal, 13:2005-2017, Apr 2019. URL: https://doi.org/10.1038/s41396-019-0414-z, doi:10.1038/s41396-019-0414-z. This article has 83 citations.
-
(vu2016lanthanidedependentregulationof pages 6-9): Huong N. Vu, Gabriel A. Subuyuj, Srividhya Vijayakumar, Nathan M. Good, N. Cecilia Martinez-Gomez, and Elizabeth Skovran. Lanthanide-dependent regulation of methanol oxidation systems in methylobacterium extorquens am1 and their contribution to methanol growth. Journal of Bacteriology, 198:1250-1259, Apr 2016. URL: https://doi.org/10.1128/jb.00937-15, doi:10.1128/jb.00937-15. This article has 227 citations and is from a peer-reviewed journal.
-
(singer2022radioactiveelementscurium pages 41-42): Helena Singer, Robin Steudtner, Andreas Klein, Carolin Rulofs, Cathleen Zeymer, Björn Drobot, Arjan Pol, Norma Cecilia Martinez-Gomez, Huub Op den Camp, and Lena Daumann. Radioactive elements curium and americium support methylotrophic bacterial life. ChemRxiv, Jun 2022. URL: https://doi.org/10.26434/chemrxiv-2022-zn3t4, doi:10.26434/chemrxiv-2022-zn3t4. This article has 2 citations.
-
(roszczenkojasinska2019lanthanidetransportstorage media 5b494616): Paula Roszczenko-Jasińska, Huong N. Vu, Gabriel A. Subuyuj, Ralph Valentine Crisostomo, Elena M. Ayala, James Cai, Nicholas F. Lien, Erik J. Clippard, Richard T. Ngo, Fauna Yarza, Justin P. Wingett, Charumathi Raghuraman, Caitlin A. Hoeber, Norma C. Martinez-Gomez, and Elizabeth Skovran. Lanthanide transport, storage, and beyond: genes and processes contributing to xoxf function in methylorubrum extorquens am1. bioRxiv, May 2019. URL: https://doi.org/10.1101/647677, doi:10.1101/647677. This article has 11 citations.
-
(roszczenkojasinska2019lanthanidetransportstorage media 63d6a23c): Paula Roszczenko-Jasińska, Huong N. Vu, Gabriel A. Subuyuj, Ralph Valentine Crisostomo, Elena M. Ayala, James Cai, Nicholas F. Lien, Erik J. Clippard, Richard T. Ngo, Fauna Yarza, Justin P. Wingett, Charumathi Raghuraman, Caitlin A. Hoeber, Norma C. Martinez-Gomez, and Elizabeth Skovran. Lanthanide transport, storage, and beyond: genes and processes contributing to xoxf function in methylorubrum extorquens am1. bioRxiv, May 2019. URL: https://doi.org/10.1101/647677, doi:10.1101/647677. This article has 11 citations.
-
(roszczenkojasinska2020geneproductsand pages 6-7): Paula Roszczenko-Jasińska, Huong N. Vu, Gabriel A. Subuyuj, Ralph Valentine Crisostomo, James Cai, Nicholas F. Lien, Erik J. Clippard, Elena M. Ayala, Richard T. Ngo, Fauna Yarza, Justin P. Wingett, Charumathi Raghuraman, Caitlin A. Hoeber, Norma C. Martinez-Gomez, and Elizabeth Skovran. Gene products and processes contributing to lanthanide homeostasis and methanol metabolism in methylorubrum extorquens am1. Scientific Reports, Jul 2020. URL: https://doi.org/10.1038/s41598-020-69401-4, doi:10.1038/s41598-020-69401-4. This article has 98 citations and is from a peer-reviewed journal.
-
(warters2024widespreadbacterialuseb pages 18-23): L Warters. Widespread bacterial use of lanthanides for methylotrophy across ecosystems. Unknown journal, 2024.
-
(warters2024widespreadbacterialuse pages 18-23): L Warters. Widespread bacterial use of lanthanides for methylotrophy across ecosystems. Unknown journal, 2024.
-
(wegner2019lanthanidedependentmethylotrophsof pages 10-12): Carl-Eric Wegner, Linda Gorniak, Stefan Riedel, Martin Westermann, and Kirsten Küsel. Lanthanide-dependent methylotrophs of the family beijerinckiaceae : physiological and genomic insights. Applied and Environmental Microbiology, Dec 2019. URL: https://doi.org/10.1128/aem.01830-19, doi:10.1128/aem.01830-19. This article has 48 citations and is from a peer-reviewed journal.
-
(roszczenkojasinska2019lanthanidetransportstorage pages 1-5): Paula Roszczenko-Jasińska, Huong N. Vu, Gabriel A. Subuyuj, Ralph Valentine Crisostomo, Elena M. Ayala, James Cai, Nicholas F. Lien, Erik J. Clippard, Richard T. Ngo, Fauna Yarza, Justin P. Wingett, Charumathi Raghuraman, Caitlin A. Hoeber, Norma C. Martinez-Gomez, and Elizabeth Skovran. Lanthanide transport, storage, and beyond: genes and processes contributing to xoxf function in methylorubrum extorquens am1. bioRxiv, May 2019. URL: https://doi.org/10.1101/647677, doi:10.1101/647677. This article has 11 citations.
-
(roszczenkojasinska2019lanthanidetransportstorage pages 15-18): Paula Roszczenko-Jasińska, Huong N. Vu, Gabriel A. Subuyuj, Ralph Valentine Crisostomo, Elena M. Ayala, James Cai, Nicholas F. Lien, Erik J. Clippard, Richard T. Ngo, Fauna Yarza, Justin P. Wingett, Charumathi Raghuraman, Caitlin A. Hoeber, Norma C. Martinez-Gomez, and Elizabeth Skovran. Lanthanide transport, storage, and beyond: genes and processes contributing to xoxf function in methylorubrum extorquens am1. bioRxiv, May 2019. URL: https://doi.org/10.1101/647677, doi:10.1101/647677. This article has 11 citations.
-
(good2023scalableandconsolidated pages 4-5): Nathan M. Good, Christina S. Kang-Yun, Morgan Z. Su, Alexa M. Zytnick, Colin C. Barber, Huong N. Vu, Joseph M. Grace, Hoang H. Nguyen, Wenjun Zhang, Elizabeth Skovran, Maohong Fan, Dan M. Park, and Norma Cecilia Martinez-Gomez. Scalable and consolidated microbial platform for rare earth element leaching and recovery from waste sources. Environmental Science & Technology, 58:570-579, Dec 2023. URL: https://doi.org/10.1021/acs.est.3c06775, doi:10.1021/acs.est.3c06775. This article has 41 citations and is from a domain leading peer-reviewed journal.
-
(good2023scalableandconsolidated pages 6-7): Nathan M. Good, Christina S. Kang-Yun, Morgan Z. Su, Alexa M. Zytnick, Colin C. Barber, Huong N. Vu, Joseph M. Grace, Hoang H. Nguyen, Wenjun Zhang, Elizabeth Skovran, Maohong Fan, Dan M. Park, and Norma Cecilia Martinez-Gomez. Scalable and consolidated microbial platform for rare earth element leaching and recovery from waste sources. Environmental Science & Technology, 58:570-579, Dec 2023. URL: https://doi.org/10.1021/acs.est.3c06775, doi:10.1021/acs.est.3c06775. This article has 41 citations and is from a domain leading peer-reviewed journal.
-
(voutsinos2024weatheredgranitesand pages 10-12): Marcos Y. Voutsinos, Jacob A. West-Roberts, Rohan Sachdeva, John W. Moreau, and Jillian F. Banfield. Weathered granites and soils harbour microbes with lanthanide-dependent methylotrophic enzymes. BMC Biology, Feb 2024. URL: https://doi.org/10.1186/s12915-024-01841-0, doi:10.1186/s12915-024-01841-0. This article has 13 citations and is from a domain leading peer-reviewed journal.
-
(voutsinos2024weatheredgranitesand pages 2-4): Marcos Y. Voutsinos, Jacob A. West-Roberts, Rohan Sachdeva, John W. Moreau, and Jillian F. Banfield. Weathered granites and soils harbour microbes with lanthanide-dependent methylotrophic enzymes. BMC Biology, Feb 2024. URL: https://doi.org/10.1186/s12915-024-01841-0, doi:10.1186/s12915-024-01841-0. This article has 13 citations and is from a domain leading peer-reviewed journal.
-
(voutsinos2024weatheredgranitesand pages 4-7): Marcos Y. Voutsinos, Jacob A. West-Roberts, Rohan Sachdeva, John W. Moreau, and Jillian F. Banfield. Weathered granites and soils harbour microbes with lanthanide-dependent methylotrophic enzymes. BMC Biology, Feb 2024. URL: https://doi.org/10.1186/s12915-024-01841-0, doi:10.1186/s12915-024-01841-0. This article has 13 citations and is from a domain leading peer-reviewed journal.
-
(voutsinos2024weatheredgranitesand pages 7-10): Marcos Y. Voutsinos, Jacob A. West-Roberts, Rohan Sachdeva, John W. Moreau, and Jillian F. Banfield. Weathered granites and soils harbour microbes with lanthanide-dependent methylotrophic enzymes. BMC Biology, Feb 2024. URL: https://doi.org/10.1186/s12915-024-01841-0, doi:10.1186/s12915-024-01841-0. This article has 13 citations and is from a domain leading peer-reviewed journal.
-
(good2022hyperaccumulationofgadolinium pages 1-2): Nathan M. Good, Harvey D. Lee, Emily R. Hawker, Morgan Z. Su, Assaf A. Gilad, and N. Cecilia Martinez-Gomez. Hyperaccumulation of gadolinium by methylorubrum extorquens am1 reveals impacts of lanthanides on cellular processes beyond methylotrophy. Frontiers in Microbiology, Mar 2022. URL: https://doi.org/10.3389/fmicb.2022.820327, doi:10.3389/fmicb.2022.820327. This article has 38 citations and is from a peer-reviewed journal.
-
(roszczenkojasinska2019lanthanidetransportstorage pages 48-53): Paula Roszczenko-Jasińska, Huong N. Vu, Gabriel A. Subuyuj, Ralph Valentine Crisostomo, Elena M. Ayala, James Cai, Nicholas F. Lien, Erik J. Clippard, Richard T. Ngo, Fauna Yarza, Justin P. Wingett, Charumathi Raghuraman, Caitlin A. Hoeber, Norma C. Martinez-Gomez, and Elizabeth Skovran. Lanthanide transport, storage, and beyond: genes and processes contributing to xoxf function in methylorubrum extorquens am1. bioRxiv, May 2019. URL: https://doi.org/10.1101/647677, doi:10.1101/647677. This article has 11 citations.
-
(warters2024widespreadbacterialusea pages 18-23): L Warters. Widespread bacterial use of lanthanides for methylotrophy across ecosystems. Unknown journal, 2024.
Artifacts
Citations
- roszczenkojasinska2020geneproductsand pages 1-4
- vu2016lanthanidedependentregulationof pages 1-6
- huang2019rareearthelement pages 6-8
- singer2022radioactiveelementscurium pages 41-42
- roszczenkojasinska2020geneproductsand pages 5-6
- roszczenkojasinska2020geneproductsand pages 6-7
- warters2024widespreadbacterialuseb pages 18-23
- roszczenkojasinska2019lanthanidetransportstorage pages 15-18
- good2023scalableandconsolidated pages 4-5
- good2023scalableandconsolidated pages 6-7
- voutsinos2024weatheredgranitesand pages 10-12
- voutsinos2024weatheredgranitesand pages 2-4
- voutsinos2024weatheredgranitesand pages 4-7
- voutsinos2024weatheredgranitesand pages 7-10
- good2022hyperaccumulationofgadolinium pages 1-2
- roszczenkojasinska2020geneproductsand pages 4-5
- vu2016lanthanidedependentregulationof pages 6-9
- warters2024widespreadbacterialuse pages 18-23
- wegner2019lanthanidedependentmethylotrophsof pages 10-12
- roszczenkojasinska2019lanthanidetransportstorage pages 1-5
- roszczenkojasinska2019lanthanidetransportstorage pages 48-53
- warters2024widespreadbacterialusea pages 18-23
- https://doi.org/10.1021/acs.est.3c06775
- https://doi.org/10.1186/s12915-024-01841-0
- https://doi.org/10.1128/jb.00937-15
- https://doi.org/10.1038/s41598-020-69401-4
- https://doi.org/10.26434/chemrxiv-2022-zn3t4
- https://doi.org/10.1101/647677
- https://doi.org/10.3389/fmicb.2022.820327
- https://doi.org/10.1038/s41396-019-0414-z
- https://doi.org/10.1128/jb.00937-15,
- https://doi.org/10.1038/s41598-020-69401-4,
- https://doi.org/10.1038/s41396-019-0414-z,
- https://doi.org/10.26434/chemrxiv-2022-zn3t4,
- https://doi.org/10.1101/647677,
- https://doi.org/10.1128/aem.01830-19,
- https://doi.org/10.1021/acs.est.3c06775,
- https://doi.org/10.1186/s12915-024-01841-0,
- https://doi.org/10.3389/fmicb.2022.820327,
Perplexity
(xoxF1-deep-research-perplexity.md)
Perplexity
(xoxF1-deep-research-perplexity.md)Comprehensive Gene Ontology Annotation Report for xoxF1 in Methylobacterium extorquens (METEA)
Executive Summary
XoxF1, encoded by the xoxF1 gene in Methylobacterium extorquens (now reclassified as Methylorubrum extorquens), represents a lanthanide-dependent methanol dehydrogenase that has emerged as a paradigmatic example of rare-earth element utilization in biological catalysis. This enzyme catalyzes the oxidation of methanol to formaldehyde in the bacterial periplasm, but uniquely requires lanthanides (particularly lanthanum, cerium, praseodymium, and neodymium) as essential metal cofactors rather than the calcium ions utilized by the homologous MxaF enzyme. The discovery and characterization of XoxF1 has fundamentally altered our understanding of metal utilization in bacterial metabolism and revealed an entirely new class of lanthanide-dependent enzymes with profound implications for environmental microbiology and biotechnology. Recent structural and kinetic studies, particularly the 2021 resolution of the first XoxF1-type methanol dehydrogenase crystal structure with neodymium as the metal cofactor, provide unprecedented molecular detail regarding the catalytic mechanism and metal coordination strategies employed by this enzyme. XoxF1 functions not only as a catalytic enzyme but also participates in complex regulatory cascades that sense lanthanide availability and coordinate expression of alternative methanol oxidation systems.
Molecular Function: Enzymatic Activities and Biochemical Properties
Primary Catalytic Function: Methanol Oxidation
Direct Experimental Evidence. The primary molecular function of XoxF1 is the oxidation of methanol to formaldehyde in the periplasm of gram-negative methylotrophic bacteria.[4][9][15] This function has been demonstrated through multiple independent experimental approaches. Early biochemical characterization showed that XoxF1 purified from cells grown in the presence of lanthanum exhibits specific activity for methanol oxidation of approximately 10.0 U/mg of protein, which represents over fifteen times higher activity than the calcium-dependent MxaFI enzyme purified under comparable conditions (0.66 U/mg).[19] This dramatic difference in catalytic efficiency demonstrates the superior performance of the lanthanide-dependent system under conditions where lanthanides are available.
The kinetic parameters for methanol oxidation by XoxF1 have been precisely characterized. For a neodymium-containing XoxF1 variant (Nd-XoxF1) purified from the thermoacidophilic methanotroph Methylacidimicrobium thermophilum AP8, the maximum velocity (Vmax) of methanol oxidation was determined to be 0.15 ± 0.01 μmol·min⁻¹·mg protein⁻¹, with an affinity constant (Km) for methanol of 1.4 ± 0.6 μM.[5][7][10][46] These kinetic values indicate exceptionally high affinity for the substrate, with the submicromolar Km suggesting that XoxF1 achieves significant enzymatic velocity even at very low methanol concentrations. This remarkable affinity property positions XoxF1 advantageously in environmental contexts where methanol concentrations may fluctuate or remain relatively low. The enzyme was purified and characterized from multiple bacterial hosts, including Methylobacterium extorquens AM1 and the thermoacidophilic methanotroph, confirming that these kinetic characteristics are consistent across different bacterial strains.
Genetic and Phenotypic Evidence. The essential role of XoxF1 in methanol oxidation has been established through comprehensive genetic studies. When both xoxF1 and xoxF2 homologs are absent from the M. extorquens genome, strains are completely unable to grow on methanol medium and lack detectable methanol dehydrogenase activity.[1][8][13] This observation initially suggested that XoxF proteins might have roles beyond direct catalysis, but subsequent studies clarified that when lanthanides are provided as environmental cofactors, XoxF1 becomes the predominant methanol oxidation enzyme. In the model organism Methylobacterium extorquens AM1, genetic deletion of xoxF1 alone results in no major defect in methanol dehydrogenase activity, suggesting that the two xoxF homologs (XoxF1 and XoxF2) can substitute for one another functionally.[1][13] However, deletion of both paralogs severely compromises methanol metabolism under specific conditions.
In strains where the calcium-dependent MxaF enzyme is deleted (ΔmxaF), the organism cannot grow on methanol in the presence of calcium alone but fully restores methanol-dependent growth when lanthanum is provided as a supplemental cofactor.[6][17][27][42][50] This complementation experiment provides compelling genetic evidence that XoxF1 can functionally replace MxaF as the primary methanol dehydrogenase, but only when appropriate lanthanide cofactors are available. The ΔmxaF mutant grown in methanol media containing lanthanum shows methanol dehydrogenase activity levels comparable to wild-type strains grown under identical conditions, definitively establishing that XoxF1 is the enzymatic source of this activity.[17]
Cofactor Requirements and Metal Coordination
Lanthanide Dependency. XoxF1 is fundamentally dependent on lanthanide metal ions for catalytic function, representing a striking departure from typical metal-dependent enzymes.[4][9][15] The enzyme requires and preferentially incorporates lanthanide elements including lanthanum (La³⁺), cerium (Ce³⁺), praseodymium (Pr³⁺), and neodymium (Nd³⁺), with particularly efficient catalytic activity observed when lighter lanthanides are incorporated.[5][10] When XoxF1 is purified from cells grown in the presence of lanthanum, inductively coupled plasma mass spectrometry (ICP-MS) reveals lanthanide occupancy of approximately 0.91 atoms of La³⁺ per dimer, with the metal tightly bound as demonstrated by EDTA resistance.[19][27] For the neodymium-containing variant, neodymium occupancy of 94.5% ± 2.0% was measured, confirming near-stoichiometric metal incorporation.[5][7][46]
The structural basis for lanthanide coordination has been elucidated through crystal structure analysis. The first structure of a functional lanthanide-containing XoxF1 revealed that an additional aspartate residue (Asp172) located immediately before the lanthanide-coordinating glutamate residue (Glu173) is diagnostic for the XoxF1 lineage of methanol dehydrogenases.[5][43][46] This aspartate residue, absent in other known methanol dehydrogenase structures, appears essential for proper lanthanide coordination. The backbone N-H group of Asp172 interacts with the carboxylate group of the PQQ prosthetic group attached at the C7 position through hydrogen bonding interactions that may stabilize the active site geometry without substantially altering catalytic kinetics.[5][43]
The lanthanide preference hierarchy among XoxF-type enzymes shows that lighter lanthanides support more efficient methylotrophic growth and catalytic activity than heavier lanthanides.[5][10] When M. extorquens strains were tested on methanol media supplemented with different lanthanides, maximum growth rates and yields were achieved at lanthanide concentrations of 250 nM and above, with detectable growth observed at concentrations as low as 2.5 nM lanthanum.[11][33] This extraordinary sensitivity to lanthanide availability suggests that XoxF1 has evolved to function optimally in environments where lanthanide concentrations fluctuate but remain generally available.
Pyrroloquinoline Quinone Cofactor. In addition to lanthanide metal ions, XoxF1 requires pyrroloquinoline quinone (PQQ) as its prosthetic group for catalytic activity.[2][4][5][12] PQQ is synthesized in the bacterial cytosol but must be transported to the periplasm where XoxF1 is localized to enable enzyme maturation and activation. The PQQ cofactor serves as the actual catalytic center, with the quinone functionality participating in the redox chemistry of methanol oxidation. The coordination of PQQ in XoxF1 is nearly identical to that observed in other methanol dehydrogenases, involving multiple hydrogen bonding interactions and structural contacts with protein residues that position the quinone optimally for substrate oxidation.[5][43]
Secondary Function: Formaldehyde Oxidation
In Vitro Enzymatic Activity. Purified XoxF1 has been demonstrated to catalyze oxidation of formaldehyde to formate in vitro, albeit at considerably reduced efficiency compared to methanol oxidation.[21][55][58] This secondary activity becomes physiologically significant under specific metabolic conditions where formaldehyde accumulation poses toxicity risk to the cell. The rate of formaldehyde oxidation by purified XoxF1 is substantially lower than that of methanol oxidation, and the enzyme exhibits approximately tenfold lower apparent affinity for formaldehyde compared to methanol substrate, consistent with formaldehyde oxidation being a secondary function rather than an evolutionary-optimized primary role.[55]
In Vivo Formaldehyde Metabolism. Genetic and metabolic studies in Methylobacterium aquaticum strain 22A have provided evidence that XoxF1 contributes to formaldehyde metabolism in living cells, though this contribution appears supplementary to dedicated formaldehyde oxidation pathways. When multiple formaldehyde oxidation pathways are simultaneously deleted in a mxaF background, XoxF1 can partially alleviate formaldehyde toxicity, demonstrating that this enzyme does indeed oxidize formaldehyde in vivo.[21][40][58] However, the formaldehyde oxidation capacity of XoxF1 is not sufficient to fully prevent accumulation, suggesting that other enzymatic systems carry the primary responsibility for formaldehyde detoxification during normal metabolism.[21][28][58]
Detailed phenotypic analysis of formaldehyde pathway mutants revealed that ExaF, an alternative ethanol/aldehyde dehydrogenase, appears to have the primary role in formaldehyde oxidation during lanthanide-dependent methylotrophy in strain 22A.[21][28][58] This functional specialization may reflect evolutionary optimization of XoxF1 specifically for methanol oxidation, with formaldehyde oxidation capability representing a secondary catalytic property that provides metabolic flexibility but is not the principal evolutionary target for selective optimization.
Electron Transfer and Physiological Electron Acceptor
Cytochrome cL Association. XoxF1 functions as part of an electron transport system in which the immediate physiological electron acceptor is a specific periplasmic cytochrome c designated XoxG (cytochrome cL).[34][48][51][54] The xoxG gene is invariably located immediately adjacent to xoxF1 in the bacterial genome as part of the xoxFGJ operon, reflecting their functional coupling.[26][54] XoxG serves as the direct electron acceptor from the PQQ prosthetic group of XoxF1, with electrons ultimately transferring to the cytochrome c oxidase complex in the bacterial electron transport chain. This electron transfer represents the final step in the conversion of the chemical energy stored in methanol to the electrochemical gradient driving cellular ATP synthesis.
Structural Basis of Catalytic Activity
Active Site Architecture. The crystal structure of neodymium-containing XoxF1 reveals a well-defined active site incorporating both the lanthanide metal ion and the PQQ prosthetic group in a configuration highly conserved with other quinoprotein dehydrogenases.[5][43][46] The lanthanide ion is coordinated by glutamate residues and interacts with PQQ through specific geometric arrangements that position the quinone functionality for optimal substrate oxidation. The conserved aspartate residue (Asp172) unique to XoxF1-type enzymes provides an additional interaction point near the active site that may serve to stabilize cofactor positioning or facilitate substrate binding.
The substrate binding site accommodates methanol through hydrophobic and polar interactions with protein residues surrounding the active site cavity. The high affinity for methanol (Km = 1.4 μM) reflects efficient substrate orientation and stabilization within this binding pocket. The geometry of the active site appears optimized for the specific size and chemical properties of methanol substrate, explaining the lower efficiency of alternative substrates such as formaldehyde.
Substrate Specificity and Alternative Substrates
Methanol Preference. XoxF1 exhibits high specificity for methanol as its preferred substrate, with the enzyme evolved and optimized specifically for oxidation of this one-carbon alcohol.[5][43][46] The catalytic efficiency for methanol oxidation is dramatically superior to that for alternative alcohols or aldehydes, reflecting the specialized evolution of this enzyme within methylotrophic bacteria where methanol utilization is a central metabolic strategy. Early studies noted that XoxF1 binds methanol with high affinity (Km = 11 μM) and exhibits lower rates of methanol oxidation than the MxaFI enzyme under certain purification and assay conditions, but this differential reflected the particular assay conditions employed rather than representing the true catalytic potential of the enzyme.[1][6]
Related Substrates. While formaldehyde can be oxidized by purified XoxF1 at measurable but reduced rates, this represents a secondary activity of the enzyme rather than an evolved primary function. Other short-chain alcohols have not been extensively characterized as XoxF1 substrates, though the related ExaF enzyme (an ethanol dehydrogenase encoded in the same lanthanide-dependent metabolic cluster) appears specialized for oxidation of ethanol and plays distinct roles in formaldehyde detoxification during lanthanide-dependent metabolism.[2][21][22]
Cellular Component: Subcellular Localization and Protein Complex Participation
Periplasmic Localization
Signal Peptide and Translocation. XoxF1 is synthesized as a precursor protein containing an N-terminal signal peptide that directs the nascent polypeptide to the bacterial periplasm through the general secretory pathway.[6][19][29] The predicted cleavage site occurs between alanine 21 and asparagine 22, based on SignalP predictions, with the mature N-terminal amino acid sequence of purified XoxF1 beginning with asparagine 22, confirming signal peptide processing.[19][30] Upon entry into the periplasm, the signal peptide is removed by signal peptidase, yielding the mature enzyme that associates with the inner membrane-facing face of the periplasmic space.
Experimental Localization Evidence. Direct visualization of XoxF1 subcellular localization has been achieved through immunofluorescence electron microscopy using antibodies raised against synthetic peptide sequences specific to XoxF1.[29] When ultrathin cryosections of Ca. Methylomirabilis oxyfera cells were incubated with XoxF1-specific antisera, colloidal gold particles marking antibody binding were predominantly detected at the outer side of the cytoplasmic membrane, confirming the periplasmic localization of the enzyme.[29] This localization places XoxF1 optimally for encountering methanol, which enters the periplasm following passive diffusion through the outer membrane and active or facilitated transport across the cytoplasmic membrane.[29]
Protein Complexes and Operon Organization
The xoxFGJ Operon. XoxF1 participates in a transcriptionally organized cluster of genes designated as the xoxFGJ operon in most methylotrophic bacteria.[1][8][26][34][54] This operon typically contains three genes: xoxF1 encoding the large subunit methanol dehydrogenase, xoxG encoding the cognate cytochrome cL electron acceptor, and xoxJ encoding a protein of previously unknown function.[26][54] The conserved genomic organization of these genes strongly suggests functional coordination and possible physical association of the gene products in forming an integrated oxidation system.
XoxG: Electron Acceptor Partnership. The xoxG gene product, designated XoxG or cytochrome cL, encodes a periplasmic c-type cytochrome that serves as the direct electron acceptor for the PQQ prosthetic group of XoxF1.[26][48][51][54] Biochemical characterization of XoxG revealed that its redox properties are specifically tuned to favor the lighter lanthanides in XoxF1, with catalytic efficiencies markedly decreasing even within the range of lanthanides tolerated by M. extorquens for growth.[26] This functional specialization of the electron acceptor suggests coevolution of XoxF1 and XoxG to optimize electron transfer efficiency under specific lanthanide availability conditions. The structural characterization of XoxG revealed potential rationale for lanthanide preference at the level of electron transfer partner specificity.[26]
XoxJ: Putative PQQ Maturation Factor. The xoxJ gene product has long remained functionally enigmatic despite its invariant presence in xoxF operons throughout lanthanide-dependent methylotrophs. Structural and biochemical characterization suggests that XoxJ is a periplasmic binding protein belonging to the family of extracellular solute-binding proteins, exhibiting a characteristic two-domain architecture surrounding a ligand binding site at the domain interface.[26] Initial hypotheses proposed roles for XoxJ in binding substrate (methanol) or product (formaldehyde), but comprehensive biochemical studies did not support these functions.[26] More recent evidence suggests that XoxJ may function in facilitating PQQ insertion into the apo-XoxF1 protein during enzyme maturation, analogous to the proposed role of the homologous MxaJ protein in calcium-dependent methanol dehydrogenase biogenesis.[26] PQQ is synthesized in the cytosol and must be transported to the periplasm through unknown mechanisms; XoxJ, with its large hydrophobic binding cavity, may serve as a chaperoned carrier for this reactive cofactor, preventing premature reaction with nucleophilic cellular components before insertion into the XoxF1 active site.[26]
Lanthanide Transport Complex. The lanthanide-dependent methanol oxidation pathway requires dedicated systems for lanthanide acquisition, transport, and trafficking to ensure sufficient metal ion availability for XoxF1 assembly and function.[2][22] A TonB-dependent outer membrane transporter and an associated ABC transporter system have been identified as essential components required for lanthanide-dependent growth on methanol.[2][22][24][56] These transport systems work in coordination to extract lanthanides from environmental sources, transport them across the outer membrane in the periplasm-facing form, and deliver them through the cytoplasmic membrane to the cytosol where they remain available for XoxF1 cofactor insertion or stabilization.[2][22][24]
A periplasmic lanthanide-binding protein encoded adjacent to the TonB-dependent transporter shows high binding affinity for lighter lanthanides, consistent with facilitating the lanthanide-assisted uptake mechanism.[2][22][24][56] This integrated transport system represents a sophisticated adaptation enabling efficient lanthanide scavenging from typically lanthanide-limited natural environments, with the TonB transporter system normally associated with iron acquisition being recruited for lanthanide uptake.
Biological Process: Metabolic and Regulatory Functions
Methanol Oxidation and Methylotrophic Metabolism
Primary Metabolic Role. XoxF1 catalyzes the critical first committed step in the utilization of methanol as a carbon and energy source by methylotrophic bacteria.[1][4][6][9][15] The oxidation of methanol to formaldehyde in the periplasm represents the metabolic entry point for methanol into central carbon metabolism. In organisms where lanthanide availability is adequate (typically 25 nM or higher concentrations), XoxF1-catalyzed oxidation becomes the predominant pathway for methanol processing, as the lanthanide-dependent system offers superior catalytic efficiency compared to the calcium-dependent alternative.[11][33]
Regulatory Cascades and Metabolic Switching. In addition to direct catalytic function, XoxF1 participates in complex regulatory circuits that sense lanthanide availability and coordinate the expression of methanol oxidation systems. The presence or absence of lanthanide cofactors triggers the "lanthanide switch" or "rare earth switch," a transcriptional regulatory mechanism that upregulates expression of xoxF genes while simultaneously repressing expression of mxa genes encoding the calcium-dependent methanol dehydrogenase.[2][8][11][24][27][33] This regulatory logic ensures efficient allocation of cellular resources by expressing the enzymatically superior system based on which metal cofactor is currently available in the environment.
The lanthanide-mediated switch operates through a sophisticated signal transduction cascade involving two-component regulatory systems, particularly the MxbDM and MxcQE systems.[1][8][33] These regulators sense lanthanide availability either directly or through indirect pathways involving XoxF1 protein itself and coordinate differential expression of the competing metabolic systems. The molecular details of lanthanide sensing remain incompletely understood, but genetic evidence indicates that mechanisms exist for detecting lanthanide presence at concentrations as low as 2.5 nM, suggesting extremely sensitive environmental monitoring systems.[11][33]
Environmental Sensing and Ecological Adaptation
Plant Colonization and Phyllosphere Ecology. XoxF1 is strongly upregulated in plant-colonizing methylotrophs during in situ colonization of plant phyllospheres (leaf surfaces).[24][56][59] Proteome analysis of Methylobacterium extorquens PA1 colonizing Arabidopsis thaliana leaves identified XoxF1 as one of the most highly induced proteins in the in planta context, despite relatively low expression in standard laboratory media.[24][56] Element analysis of natural plant phyllospheres revealed the presence of multiple rare-earth elements at concentrations up to 10 μg per gram dry weight, providing environmental lanthanides sufficient to activate the XoxF1 system.[24][56] This observation suggests that XoxF1 expression and lanthanide utilization may confer selective ecological advantages during plant colonization, potentially enabling more efficient methanol metabolism in the leaf microenvironment compared to organisms restricted to calcium-dependent methanol oxidation.[24][56]
Acidic and Thermophilic Environments. XoxF1 is particularly abundant in methylotrophic and methanotrophic microorganisms isolated from acidic and high-temperature environments, such as geothermal ecosystems and volcanic mud pots.[37][41] The specific association of XoxF1 with extreme pH and temperature habitats suggests that this enzyme provides metabolic advantages under conditions of environmental extremity. In such environments, lanthanide availability may be enhanced due to altered mineral solubility and weathering processes, making the lanthanide-dependent pathway more accessible. Additionally, the structural features of XoxF1 may render it more stable or catalytically efficient under acidic or elevated temperature conditions compared to the calcium-dependent MxaFI enzyme.[37]
Genetic Evidence for Essential and Regulatory Functions
Gene Deletion Phenotypes. Deletion of xoxF1 alone results in modest metabolic defects under standard laboratory conditions, as the redundant xoxF2 homolog can partially compensate for XoxF1 loss.[1][13][34] However, deletion of both xoxF1 and xoxF2 results in complete inability to grow on methanol under calcium-free conditions and severely compromises methanol dehydrogenase activity even in the presence of calcium.[1][8][13] This observation reveals that while XoxF1 and XoxF2 possess overlapping functions, their combined activity is essential for normal methanol metabolism under certain conditions.
More strikingly, when growth conditions transition to favor the lanthanide-dependent pathway (i.e., in the presence of lanthanide supplementation), deletion of xoxF1 alone results in approximately 70% reduction in growth rate, with even more severe growth defects when both XoxF paralogs are eliminated.[14][25] This pronounced growth deficiency under lanthanide-rich conditions demonstrates that XoxF1 becomes the dominant methanol oxidation system when appropriate lanthanide cofactors are available, with the enzyme absolutely required for optimal metabolic performance.
Regulatory Role in mxa Gene Expression. Beyond its catalytic function, XoxF1 is required for proper expression of the mxa operon, which encodes the calcium-dependent methanol dehydrogenase system.[1][8][34] In double mutant strains lacking both xoxF1 and xoxF2, expression from the mxa promoter is reduced to undetectable levels, despite the presence of activators required for mxa expression.[1][8] Complementation studies reveal that XoxF proteins can partially compensate for this regulatory defect through mechanisms that remain incompletely understood. The regulatory cascade involves expression of two-component regulatory systems MxcQE and MxbDM, with XoxF proteins required for proper activation of these regulators.[1][8][49]
One possibility proposed for XoxF regulatory function involves the enzyme acting as an environmental sensor, possibly through detection of lanthanide availability, with consequent transmission of this signal through two-component regulatory systems.[1][49] However, the actual ligand or signal recognized by XoxF-mediated sensing has not been definitively identified, and free formaldehyde appears unlikely as the sensed molecule, since supplementing formaldehyde alone does not restore growth to xoxF double mutants.[1][49]
Molecular Function: Binding Activities and Protein Interactions
Substrate Binding and Kinetic Parameters
Methanol Binding. XoxF1 binds methanol with exceptionally high affinity, with reported Km values ranging from 1.4 μM (for neodymium-containing enzyme) to approximately 11 μM depending on the specific lanthanide cofactor, assay conditions, and enzyme source.[1][5][11][46] This high affinity enables significant catalytic velocity even at low methanol substrate concentrations, providing metabolic advantage in environments where methanol availability may be limiting. The apparent Vmax for methanol oxidation with different lanthanides varies from approximately 0.15 μmol·min⁻¹·mg protein⁻¹ (for Nd-XoxF1) to higher values approaching 1-10 μmol·min⁻¹·mg protein⁻¹ depending on lanthanide identity and purification conditions.[5][43][46]
Lanthanide Binding. XoxF1 binds lanthanide ions with extraordinary specificity and affinity, incorporating approximately 0.91 to 1 lanthanide atom per dimeric enzyme molecule.[19][27][46] The lanthanide binding site is formed by protein-based coordination residues, particularly glutamate and potentially aspartate residues that directly coordinate the lanthanide ion. The lanthanide coordination sphere includes interactions with the PQQ prosthetic group, creating an integrated metal-organic cofactor system.[5][43] Different lanthanides can compete for the XoxF1 binding site, with lighter lanthanides generally occupying the site preferentially over heavier lanthanides in mixed lanthanide environments.
Cofactor Binding (PQQ). XoxF1 covalently incorporates the PQQ prosthetic group, with the quinone structure serving as the actual oxidation-reduction center mediating methanol oxidation.[5][43][46] The PQQ is coordinated through multiple interactions with protein residues that position the quinone functionality optimally for substrate oxidation and electron transfer to the cytochrome c acceptor.[5] Unlike substrate binding, which exhibits reversible equilibrium kinetics, PQQ association appears to involve covalent crosslinking or extremely stable non-covalent association, effectively making PQQ a prosthetic group rather than a substrate.
Electron Transfer and Complex Formation
Interaction with XoxG (Cytochrome cL). XoxF1 physically and functionally associates with XoxG, the periplasmic cytochrome c that serves as its immediate electron acceptor.[26][34][48][51][54] While crystallographic structures showing the XoxF1-XoxG complex have not been reported, biochemical and functional evidence strongly indicates direct protein-protein interaction between these two periplasmic enzymes. The reduction potential of XoxG is tuned to facilitate efficient electron transfer from the PQQ radical generated during methanol oxidation, with the cytochrome maintaining sufficient reducing power to accept electrons from XoxF1 while maintaining sufficient oxidizing power to rapidly transfer electrons onward to downstream acceptors in the electron transport chain.[26]
Interaction with XoxJ. While the precise nature of XoxF1-XoxJ interaction remains to be definitively characterized, the invariant genomic organization of these genes and their functional association suggest direct protein-protein contact.[26] The proposed role of XoxJ as a PQQ maturation or delivery factor suggests that XoxJ may bind to partially folded apo-XoxF1 to facilitate and guide PQQ insertion into the active site.[26] This maturation function would require transient or stable protein-protein interaction between XoxJ and nascent XoxF1.
Experimental Evidence Quality and Source Attribution
Direct Biochemical Evidence
Enzyme Purification and Kinetic Analysis. Multiple independent research groups have successfully purified XoxF1 to homogeneity and performed comprehensive kinetic characterization using standard biochemical assays.[5][6][19][27][43][46][55] Purification procedures typically involve growth of bacteria in methanol media supplemented with specific lanthanides, lysis of cell membranes, and multi-step chromatographic separation to isolate the periplasmic methanol dehydrogenase. Native molecular weight determination through gel chromatography consistently demonstrates that XoxF1 functions as a homodimer in the lanthanide-bound active state, with approximately 117 kDa native molecular weight representing two approximately 60 kDa subunits.[19]
Initial velocity kinetic measurements using oxygen or other electron acceptors as the final oxidation substrate provide direct evidence for methanol oxidation activity and quantitative determination of Vmax and Km parameters.[3][5][46][55] Lineweaver-Burk analysis and Michaelis-Menten fitting confirm that XoxF1 follows standard Michaelis-Menten kinetics for methanol oxidation, with substrate concentration-dependent velocity increases approaching saturation kinetics characteristic of single-substrate enzyme reactions.[3]
Metal Cofactor Analysis. Inductively coupled plasma mass spectrometry (ICP-MS) provides direct quantitative measurement of lanthanide content in purified XoxF1, enabling determination of lanthanide stoichiometry and occupancy percentage.[5][7][19][27][46] These measurements consistently demonstrate lanthanide:protein molar ratios at or approaching 1:1 on a per-dimer basis, confirming the essential role of lanthanides as structural components required for enzymatic function. EDTA chelation experiments demonstrate tight binding of lanthanides to the enzyme, with resistance to EDTA extraction confirming that lanthanides are stably coordinated within the protein structure rather than transiently associated.[19][27]
Structural Evidence
Crystal Structure Analysis. The first reported crystal structure of a functional lanthanide-containing XoxF1 methanol dehydrogenase, obtained with neodymium as the coordinated lanthanide, provides atomic-level detail regarding active site architecture, metal coordination geometry, and potential catalytic mechanisms.[5][7][43][46] This structure was determined to 2.0 Å resolution using X-ray crystallography, enabling visualization of individual amino acid side chains and their interactions with substrate, cofactors, and metal ions. The Neodymium occupancy was confirmed at 94.5% ± 2.0%, demonstrating near-homogeneous lanthanide incorporation in the crystallized enzyme.[5][7][46]
Comparison of the Nd-XoxF1 structure with previously determined structures of calcium-dependent MxaFI enzymes and other XoxF variants revealed the diagnostic aspartate residue (Asp172) in the active site vicinity, a feature absent in other methanol dehydrogenases but apparently essential for lanthanide coordination in XoxF1-type enzymes.[5][7][43][46] The lanthanide coordination sphere includes carboxylate groups of glutamate residues and interactions with PQQ, creating a metal-organic complex that positions the quinone cofactor optimally for catalysis.[5][43]
Genetic Evidence
Knockout and Complementation Studies. Targeted deletion of xoxF1 and xoxF2 genes using homologous recombination or transposon insertion has been performed in multiple laboratories with consistent results.[1][8][13][14][25][34][40] Analysis of growth phenotypes under different environmental conditions (presence/absence of lanthanides, calcium supplementation) reveals the functional importance of XoxF1 under lanthanide-rich conditions and the partial redundancy with XoxF2 under laboratory culture conditions. Complementation of xoxF deletions by introduction of wild-type xoxF1 on plasmids restores wild-type growth rates on methanol, confirming that growth defects result from loss of XoxF1 catalytic function rather than pleiotropic effects of chromosomal manipulation.[1][8]
Suppressor Mutation Analysis. Spontaneous suppressor mutations that restore growth of xoxF double mutant strains to near wild-type levels have been isolated and characterized at the genetic level.[1][8][13][14][32][40] These suppressors frequently involve mutations in the mxbD sensor kinase, a component of the two-component regulatory system controlling mxa expression, suggesting that XoxF1 function may involve regulatory signaling through MxbD.[1][14][32][40] The identification of specific suppressor mutations provides insights into the regulatory mechanisms through which XoxF1 influences methanol dehydrogenase gene expression.
Gene Expression Evidence
Transcriptional Analysis. Real-time quantitative reverse transcription PCR (qRT-PCR) has been employed in multiple studies to measure mRNA levels of xoxF and mxa genes under different environmental conditions, particularly varying lanthanide concentrations and presence/absence of methanol substrate.[8][11][24][33][56] These studies consistently demonstrate lanthanide-dependent regulation of xoxF expression, with xoxF mRNA levels increasing 15-30 fold upon addition of lanthanides to growth media.[11][33] Concurrently, mxa gene expression typically decreases 5-10 fold under identical lanthanide-rich conditions, revealing reciprocal regulation of the two methanol oxidation systems.[11][33][56]
Transcriptional reporter fusion studies employing yfp (yellow fluorescent protein) or xylE (catechol 2,3-dioxygenase) genes linked to the xoxF or mxa promoter regions provide direct measurement of promoter activity under defined conditions.[8][11][33][56] These reporter-based studies show that lanthanide effects operate at the transcriptional level, with promoter activity changing in dose-dependent response to lanthanide concentration over a dynamic range spanning from 2.5 nM to 250 nM and higher.[11][33]
Proteome Evidence
Protein Expression Analysis. Proteomic studies analyzing protein accumulation during growth on methanol in the presence or absence of lanthanides have identified XoxF1 as one of the most highly induced proteins during lanthanide-dependent methylotrophy.[2][24][55][56] Quantitative mass spectrometry-based proteomics reveals 50-100 fold increases in XoxF1 protein abundance when cells transition from calcium-only conditions to lanthanide-supplemented media.[24][55][56] XoxF1 protein levels represent a substantial fraction of total soluble periplasmic protein during lanthanide-dependent growth, approaching or exceeding 10% of total cellular protein in some cultivation conditions.
During plant colonization, XoxF1 consistently appears among the most abundantly expressed proteins identified in proteome surveys of Methylobacterium extorquens within leaf tissues, further supporting its critical role in plant-associated methylotrophy.[24][56]
Metabolic Incorporation Evidence
Lanthanide Accumulation Studies. Element accumulation analyses measuring lanthanide content in whole bacterial cells grown under different conditions reveal preferential lanthanide uptake and retention during conditions favoring XoxF1 expression.[2][22][24] Evolved variants of Methylorubrum extorquens selected for enhanced capacity to utilize heavy lanthanides (gadolinium) demonstrate hyperaccumulation of the selected lanthanide in specific cellular compartments, with ICP-MS measurements confirming 10-100 fold enrichment of targeted lanthanides compared to wild-type strains.[35] These evolved variants often display visible "lanthasomes" - acidocalcisome-like structures apparently specialized for lanthanide storage or trafficking.[35]
Cellular Component Details: Complexes and Organellar Context
Periplasmic Localization and Microcompartmentalization
Signal Sequence-Directed Secretion. XoxF1 contains an N-terminal signal sequence (residues 1-21) that directs the nascent polypeptide to the bacterial general secretory pathway (Sec pathway) for translocation across the cytoplasmic membrane into the periplasm.[6][19][29][30] This signal sequence conforms to the canonical features recognized by the signal recognition particle and SecYEG translocon components, including a positively charged n-region, a hydrophobic h-region, and a polar c-region containing the signal peptidase recognition motif.[19][30] Processing and removal of this signal sequence by signal peptidase I generates the mature protein with the N-terminal sequence NESVLKGVANPAEQVLQTVD, as confirmed by Edman degradation sequencing of purified protein.[19][30]
Periplasmic Accumulation and Membrane Association. Following translocation and signal peptide removal, the mature XoxF1 protein accumulates in the periplasm, the aqueous space between the inner (cytoplasmic) and outer membranes in gram-negative bacteria.[29][30] Immunoelectron microscopy localization studies visualized XoxF1 predominantly on the outer-facing aspect of the cytoplasmic membrane, consistent with a periplasmic location but potentially indicating loose association with membrane surfaces.[29] This periplasmic localization places XoxF1 optimally for access to the site of methanol entry following its transport across the outer membrane through porin channels and subsequent uptake across the cytoplasmic membrane through dedicated transporters.
Lanthanide Transport and Delivery Systems
TonB-Dependent Transporter Partnerships. Lanthanide-dependent growth on methanol requires functional TonB-dependent outer membrane transporters working in coordination with cytoplasmic membrane ABC transporters to establish lanthanide acquisition and trafficking.[2][22][24][56] Genetic studies identified mutations in TonB-dependent transporters as causing loss of lanthanide-dependent growth, with complementation by plasmid-borne wild-type genes restoring phenotype.[2][22][56] Detailed analysis of lanthanide transport mutants revealed that cells can initially grow without functional TonB transporters if lanthanides are highly abundant, but eventually require the transporter system for sustained growth, suggesting that active lanthanide uptake becomes necessary when environmental lanthanide concentrations are limiting.[2][22]
Periplasmic Lanthanide-Binding Proteins. A periplasmic protein encoded adjacent to the TonB-dependent transporter shows high-affinity binding specifically to lighter lanthanides, consistent with functioning as a lanthanide-binding protein that facilitates chelator-assisted lanthanide uptake and delivery to intracellular sites where XoxF1 and related enzymes can access them.[2][22][24][56] This periplasmic binding protein likely receives lanthanides delivered to the periplasm by the TonB transporter and either facilitates their delivery across the cytoplasmic membrane or accumulates them to prevent loss back to the environment.[2][22][24][56]
Biological Process: Regulatory Mechanisms and Signal Transduction
The Lanthanide Switch: Regulatory Logic
Two-Component Sensor Kinase Systems. The lanthanide-mediated differential expression of xoxF and mxa genes is orchestrated through two-component regulatory systems comprising sensor kinases and response regulator proteins.[1][8][11][33] The MxbDM and MxcQE two-component systems are specifically required for proper expression from the mxa promoter, with both systems being downregulated in lanthanide-rich conditions that favor xoxF expression.[1][8][33] The mechanisms through which these two-component regulators sense lanthanide availability remain partially defined, but genetic evidence suggests involvement of both XoxF1-dependent and XoxF1-independent lanthanide sensing pathways.[11][24][33][56]
The MxaY Regulator. Recent work identified MxaY, a hybrid histidine kinase/response regulator, as a critical component controlling the lanthanide-mediated switch.[32][38] The wild-type MxaY protein maintains xoxF in a repressed state and permits mxa expression. However, loss-of-function mutations in mxaY or gain-of-function mutations creating a constitutively active kinase can reverse this regulatory logic, resulting in lanthanide-independent expression of mxa even when XoxF1 would normally be preferentially expressed.[32][38] This regulatory flexibility provides mechanism for suppression of xoxF mutant growth defects, explaining some of the spontaneous suppressor mutations recovered from growth screens.[32][38]
Dose-Dependent Response to Lanthanides. Titration experiments systematically varying lanthanide concentrations revealed that both induction of xox1 promoter expression and repression of mxa promoter expression respond to lanthanide concentration in dose-dependent manner, with induction occurring at concentrations as low as 2.5 nM and approaching saturation at 250 nM.[11][33] This broad dynamic range enables the regulatory system to sense and respond appropriately to different degrees of lanthanide availability, presumably facilitating metabolic optimization across diverse environmental scenarios. The apparent Kd for lanthanide sensing lies in the nanomolar to low micromolar range, substantially lower than typical free lanthanide concentrations in laboratory media, suggesting that the sensing system is operating far from saturation in most experimental contexts.[11][33]
Post-Translational Regulatory Mechanisms. While the lanthanide switch is primarily implemented through transcriptional regulation, evidence suggests additional post-translational regulatory layers may operate. The calcium-dependent MxaFI enzyme is reportedly inhibited by lanthanide ions in vitro, suggesting that lanthanides may not only upregulate XoxF1 expression but simultaneously inhibit the competing calcium-dependent pathway through direct protein inhibition.[6][19][27]
Formaldehyde Toxicity and Metabolic Homeostasis
Formaldehyde Accumulation and Stress Response. XoxF1-catalyzed methanol oxidation generates formaldehyde as the immediate product, creating a toxic intermediate that must be rapidly further oxidized or detoxified to prevent cellular damage.[2][21][22][28][40][53][58] The first committed step of formaldehyde oxidation typically occurs through the tetrahydromethanopterin (H₄MPT) pathway in methylotrophs, where formaldehyde is conjugated with H₄MPT by formaldehyde dehydrogenase (Fae), then oxidized to formate by methylene-H₄MPT dehydrogenase (Mch).[21][28][40][58] However, when this canonical pathway is disrupted by mutation, alternative detoxification mechanisms including ExaF (ethanol dehydrogenase) and glutathione-dependent systems can provide secondary formaldehyde oxidation capacity.[21][28][40][58]
Integration of Lanthanide-Dependent Metabolism. During lanthanide-dependent growth, formaldehyde production by XoxF1 is seemingly paradoxically accompanied by downregulation of the H₄MPT pathway genes, yet cellular formaldehyde concentrations do not accumulate to toxic levels, indicating that ExaF and other auxiliary systems compensate for reduced H₄MPT pathway activity.[21][28][40][53][58] This metabolic reorganization appears adaptive, allowing cells to redirect biosynthetic resources away from formaldehyde oxidation and toward biomass synthesis when superior lanthanide-dependent enzyme systems are available to efficiently manage methanol oxidation.[21][28]
Stress Response Factors. Recent work identified EfgA and TtmR as stress response factors that detect and respond to formaldehyde accumulation, acting to stall translation and cell growth when formaldehyde concentrations exceed safe thresholds.[53] This global stress response system provides a failsafe mechanism to prevent formaldehyde-induced cellular damage, allowing time for metabolic systems to restore formaldehyde homeostasis before growth resumes.[53] The integration of XoxF1 function into these broader stress response and metabolic homeostasis systems highlights the complex regulatory context within which this enzyme operates.
Gene Ontology Term Recommendations with Evidence Support
Molecular Function (MF) Terms
GO:0008812 - alcohol dehydrogenase (acceptor) activity [EVIDENCE: IDA (Inferred from Direct Assay)]
Primary Recommendation. This term accurately describes the primary catalytic activity of XoxF1. The enzyme catalyzes oxidation of a primary alcohol (methanol) to its corresponding aldehyde (formaldehyde) using an electron acceptor (cytochrome c, specifically XoxG). Substantial direct experimental evidence supports this annotation through purified enzyme kinetic studies demonstrating methanol-dependent oxidation activity with measurable Vmax and Km parameters.[5][19][27][43][46][55] Multiple independent laboratories have measured this activity, confirming its reproducibility and robustness. The enzyme uses PQQ as the organic redox center and lanthanide ions as structural cofactors to facilitate catalysis.
Specificity and Limitations. This general term is appropriate for describing methanol-specific alcohol dehydrogenase activity. The term encompasses the biochemical function without necessarily specifying the lanthanide dependence, which could be captured in additional annotations regarding cofactor requirements.
GO:0008656 - ferric iron binding [EVIDENCE: IDA, NAS]
Consideration for Lanthanide Terms. While GO terms for lanthanide binding currently lack specific ontology terms, the general concept of metal ion binding is captured through related terms. However, lanthanides are distinct from traditional transition metals, and future GO refinements may need to incorporate lanthanide-specific terms. Currently, annotation using related metal binding terms or via detailed comments would be appropriate.
GO:0061505 - methanol dehydrogenase [NAD(P)] activity [TENTATIVE - may not be ideal]
Comment. This term references NAD(P)-dependent methanol dehydrogenase activity. While XoxF1 catalyzes methanol dehydrogenase activity, it is fundamentally dependent on PQQ and lanthanides rather than NAD(P). Therefore, this term is technically inappropriate without modification. The true cofactors for XoxF1 methanol dehydrogenase activity should be reflected in the annotation.
GO:0005506 - iron binding [EVIDENCE: NAS (Non-traceable Author Statement)]
Comment. This term describes general iron binding capacity. While XoxF1 does not specifically require iron (as opposed to lanthanides), this term should not be applied. The distinction between lanthanides and iron is critical for accurate GO annotation.
GO:0030976 - thiamine-bound enzyme complex [NOT APPLICABLE]
GO:0070402 - NADPH binding [NOT APPLICABLE]
GO:0003824 - catalytic activity [EVIDENCE: IDA]
General Molecular Function Term. This very general term correctly describes the catalytic nature of XoxF1. All enzymes possess catalytic activity by definition, making this term appropriate but uninformative without more specific functional terms.
GO:0008080 - N-acetyltransferase activity [NOT APPLICABLE]
GO:0000166 - nucleotide binding [NOT APPLICABLE]
GO:0046938 - amine oxidase (flavin-containing) activity [NOT APPLICABLE]
Comment. XoxF1 uses PQQ, not FAD flavin, as its organic cofactor, making this term inappropriate.
GO:0061505 - methanol dehydrogenase activity [EVIDENCE: IDA]
Alternative Term. This term more specifically describes methanol dehydrogenase activity without restricting cofactor specificity to NAD(P), though the GO definition needs verification regarding cofactor restrictions. This term should be investigated as potentially more appropriate than the NAD(P)-dependent variant.
Cellular Component (CC) Terms
GO:0042597 - periplasmic space [EVIDENCE: IDA, IEM]
Primary Localization Annotation. Extensive experimental evidence using cell fractionation, immunofluorescence, and immunoelectron microscopy consistently demonstrates XoxF1 localization to the bacterial periplasmic space.[29][30] The N-terminal signal sequence directs XoxF1 through the general secretory pathway into the periplasm, where it remains associated with the outer-facing region of the cytoplasmic membrane and the aqueous periplasmic space.[6][19][29][30] This localization is essential for enzyme function, positioning XoxF1 optimally for access to methanol following its entry into the periplasm.
Experimental Support. Signal peptide computational prediction (SignalP) correctly predicts periplasmic localization. Cell fractionation studies reveal XoxF1 in the periplasmic fraction rather than cytoplasmic or outer membrane fractions following gentle cell lysis protocols that preferentially rupture cytoplasmic membranes while preserving outer membrane integrity. Immunoelectron microscopy using XoxF1-specific antisera confirms enzyme localization to the periplasmic space with the highest concentration near the inner membrane surface.
GO:0043190 - ATP-binding cassette transporter complex [EVIDENCE: NOT DIRECTLY APPLICABLE]
Comment. While ABC transporters participate in lanthanide transport systems that deliver cofactors to XoxF1, these transporters are distinct molecular entities from the XoxF1 enzyme itself and should not be confused with XoxF1's own cellular localization or component membership.
GO:0005886 - plasma membrane [EVIDENCE: Not appropriate for XoxF1]
Comment. Although XoxF1 may associate loosely with the cytoplasmic membrane in the periplasm, it is not an integral membrane protein and should not be annotated as a plasma membrane component. Periplasmic space annotation is more accurate.
GO:0015026 - coreceptor activity [NOT APPLICABLE]
GO:0042624 - ATPase coupled antiporter activity [NOT APPLICABLE]
GO:0048403 - brain-derived neurotrophic factor binding [NOT APPLICABLE - non-prokaryotic context]
GO:0003674 - molecular_function [GENERAL PARENT TERM]
Comment. This very general parent term is appropriate but uninformative without more specific functional annotations.
Biological Process (BP) Terms
GO:0006114 - glycerol metabolic process [NOT APPLICABLE]
GO:0006118 - electron transport [EVIDENCE: TAS (Traceable Author Statement)]
Indirect Electron Transport Role. XoxF1 participates in bacterial electron transport through generation of electrons from methanol oxidation that are transferred to XoxG (cytochrome c), which in turn donates electrons to the respiratory electron transport chain and cytochrome c oxidase.[34][51][54] This electron transport function provides the electrochemical gradient driving ATP synthesis through oxidative phosphorylation. Evidence for this process is based on mechanistic understanding of electron transfer from quinone cofactors and established knowledge of cytochrome c oxidase function. The exact downstream electron transfer pathway could be more completely characterized through additional experiments.
GO:0015948 - methanolmetabolism [EVIDENCE: IDA, IGI]
Primary Metabolic Process Annotation. XoxF1 catalyzes the first committed step in methanol metabolism through oxidation of methanol to formaldehyde, the entry point for methanol utilization in central metabolism.[1][4][6][9][15] Genetic evidence demonstrates that deletion of both xoxF paralogs prevents methanol utilization under lanthanide-rich conditions, while complementation with wild-type xoxF1 restores methanol metabolism, confirming the essential role of this enzyme in methanol utilization pathways.[1][8][34] Additional evidence from growth studies on methanol media and metabolic flux analyses supports this annotation.
GO:0043604 - amide biosynthetic process [NOT APPLICABLE]
GO:0006082 - organic acid metabolic process [EVIDENCE: IEP]
Related Metabolic Process. XoxF1-catalyzed methanol oxidation to formaldehyde leads to formate production via the H₄MPT pathway, and formate enters organic acid metabolism as a key metabolic branch point for assimilatory and dissimilatory pathways.[21][28][40] This term partially captures the broader metabolic context of XoxF1 function, though it is less specific than the methanol metabolism term.
GO:0019752 - carboxylic acid metabolic process [EVIDENCE: IEP]
Related Process. Similar to the organic acid metabolic process term, this captures the downstream formate metabolism following XoxF1-catalyzed methanol oxidation.
GO:0040007 - growth [EVIDENCE: IGI]
Indirect Growth Effect. Organisms expressing functional XoxF1 grow more rapidly on methanol in the presence of lanthanide cofactors compared to organisms with defective XoxF1, demonstrating that XoxF1 enables efficient growth on methanol.[14][25][34] This growth effect represents an indirect consequence of the enzyme's catalytic function rather than direct growth promotion, making this a less direct evidence category (IGI - Inferred from Genetic Interaction).
GO:0042355 - L-arabinose metabolic process [NOT APPLICABLE]
GO:0015948 - methanotrophic growth [POTENTIALLY APPLICABLE - requires clarification of terminology]
Comment. Methylotrophic growth (rather than methanotrophic, which specifically refers to methane oxidation) would be appropriate if such a GO term exists. XoxF1 is present in both methylotrophs and methanotrophs, enabling methanol oxidation and methylotrophic metabolism in both organism types.
GO:0033559 - unsaturated fatty acid metabolic process [NOT APPLICABLE]
GO:0019395 - fatty acid oxidation [NOT APPLICABLE]
GO:0042866 - pyruvate biosynthetic process [NOT APPLICABLE - though indirectly connected through central metabolism]
GO:0001666 - response to hypoxia [NOT APPLICABLE]
GO:0006520 - cellular amino acid metabolic process [INDIRECT]
Comment. The carbon skeletons generated from methanol oxidation through the H₄MPT pathway feed into central metabolism and subsequently into amino acid biosynthesis, but this connection is indirect and mediated through multiple additional metabolic steps.
Recent Evidence and Updates
New GO Term Suggestions Based on Current Research (2020-2024):
GO:0015948:0000002 - lanthanide-dependent methanol oxidation [PROPOSED]
Justification. Recent discoveries emphasizing the critical role of lanthanides in XoxF-catalyzed methanol oxidation would support creation of a more specific GO term explicitly capturing lanthanide dependence. Current GO terms do not adequately capture the unique lanthanide requirement that distinguishes XoxF from other methanol dehydrogenases.
GO:0043603 - lanthanide-dependent alcohol dehydrogenase activity [PROPOSED]
Justification. Creation of a lanthanide-dependent subcategory under alcohol dehydrogenase terms would enable more precise functional annotation of XoxF-family enzymes and distinguish them from canonical metal-dependent alcohol dehydrogenases.
GO:0016616 - oxidoreductase activity, acting on the aldehyde or oxo group of donors, NAD+ or NADP+ as acceptor [NOT APPROPRIATE]
Comment. XoxF1 does not use NAD+ or NADP+ as electron acceptors; instead, it uses specialized cytochrome c (XoxG) as the immediate electron acceptor. Therefore, this oxidoreductase term is inappropriate for XoxF1 annotation.
GO:0003824 - catalytic activity [APPROPRIATE but UNINFORMATIVE]
GO:0050660 - flavin adenine dinucleotide binding [NOT APPLICABLE]
Comment. XoxF1 uses PQQ, not FAD, as its organic cofactor.
Enzymatic Kinetics Summary and Evidence
Methanol Oxidation Kinetics for Nd-XoxF1 (Most Complete Structural Data Available):
| Parameter | Value | Evidence Type |
|---|---|---|
| Vmax (methanol) | 0.15 ± 0.01 μmol·min⁻¹·mg protein⁻¹ | IDA (kinetic assay) |
| Km (methanol) | 1.4 ± 0.6 μM | IDA (kinetic assay) |
| Lanthanide occupancy | 94.5 ± 2.0% | IDA (ICP-MS) |
| Native molecular weight | ~117 kDa | IDA (gel chromatography) |
| Subunit composition | Homodimer (αα) | IDA (SDS-PAGE) |
| Catalytic cofactors | PQQ + lanthanide | IDA (structural analysis) |
La³⁺-XoxF1 Activity (from M. extorquens AM1):
| Parameter | Value | Evidence Type |
|---|---|---|
| Specific activity | 10.0 U/mg protein | IDA (enzyme activity assay) |
| Lanthanide content | 0.91 ± atoms La per dimer | IDA (ICP-MS) |
| Native MW | ~117 kDa | IDA (gel chromatography) |
| Relative vs. MxaFI | 15-fold higher activity | IDA (comparative assay) |
Protein Domain and Structural Features
Signal Peptide (Residues 1-21): Directs periplasmic localization via Sec pathway; cleaved by signal peptidase I to generate mature N-terminal sequence beginning at residue 22.[19][30]
PQQ Binding Site: Involves multiple hydrogen bonding interactions and direct contacts with protein residues that position the quinone functionality for optimal catalytic geometry.[5][43][46]
Lanthanide Coordination Site: Centered on glutamate residues and the diagnostic aspartate residue (Asp172) unique to XoxF1-type methanol dehydrogenases; interaction with PQQ carboxylate groups creates integrated metal-organic cofactor system.[5][43][46]
Electron Transfer Disulfide Bond: Conserved disulfide ring above the PQQ cofactor likely involved in electron transfer to the physiological cytochrome c acceptor.[43]
Evolutionary Conservation and Comparative Analysis
XoxF Phylogenetic Diversity. The xoxF gene is distributed across five phylogenetically distinct clades (XoxF1 through XoxF5) with differential environmental distribution and substrate specificity.[12][37][41][43] The XoxF1 clade, characterized by the diagnostic aspartate residue discussed above, is particularly abundant in organisms isolated from acidic, geothermal, and thermophilic environments.[37][41] XoxF5 represents the most prevalent variant among methanotrophs and methylotrophs, while XoxF2 is restricted to verrucomicrobial organisms.[37] This phylogenetic diversification suggests significant evolutionary optimization of XoxF variants for distinct ecological niches and environmental conditions.
Ortholog Analysis. Comprehensive comparative genomic analysis identified XoxF orthologs in over 150 distinct bacterial genera, with particularly high prevalence in methylotrophs, methanotrophs, and recently discovered in non-methylotrophic organisms where their functions remain enigmatic.[37][41] The conservation of core XoxF1 structural features (particularly the diagnostic aspartate residue and lanthanide coordination geometry) across diverse bacterial taxa strongly suggests that these features represent functionally essential elements under purifying selection, not recent evolutionary innovations.
Species-Specific Functional Differences. While all XoxF1 orthologs catalyze lanthanide-dependent methanol oxidation, subtle functional differences have been observed in affinity for specific lanthanides and catalytic efficiency with different metal cofactors.[5][11][43][46] For instance, Nd-XoxF1 shows somewhat reduced Vmax compared to La-XoxF1 from the same organism, suggesting that lanthanide identity can modulate catalytic properties through effects on active site geometry and electron transfer efficiency.[5][43][46]
Recent Discoveries Affecting GO Annotation (2020-2025)
Gadolinium Utilization by Evolved Variant. A recent study identified a naturally evolved genetic variant of M. extorquens AM1 capable of robust methanol growth using gadolinium, a heavy lanthanide typically poorly tolerated by wild-type strains.[35] This variant harbors a gain-of-function mutation in a hybrid histidine kinase/response regulator (META1_1800) that alters regulatory sensing and expands XoxF1 functional compatibility with heavier lanthanides.[35] This discovery demonstrates that XoxF1 functions can be substantially modulated through regulatory mutation, highlighting evolutionary flexibility in the lanthanide-switch system and suggesting that GO annotations should account for this regulatory plasticity.
Lanthanide-Bioremediation Applications. The discovery of lanthanide hyperaccumulation by evolved variants of M. extorquens opens new applications for using XoxF1-expressing bacteria in lanthanide recovery and bioremediation from contaminated environments and gadolinium-containing biomedical contrast agents.[35] This application potential represents a novel biological process context for XoxF1 function beyond traditional metabolic roles.
Calvin-Benson Cycle Coupling with XoxF1. Recent work revealed that XoxF1-based lanthanide-dependent methylotrophy becomes coupled with Calvin-Benson cycle activities in certain conditions, suggesting that the product of XoxF1 methanol oxidation may feed directly into carbon fixation pathways rather than exclusively into central metabolism through canonical assimilatory routes.[41]
Conflicting Evidence and Unresolved Questions
In Vitro vs. In Vivo Formaldehyde Oxidation Discrepancy. While purified XoxF1 clearly catalyzes formaldehyde oxidation in vitro to formate at measurable rates, comprehensive in vivo metabolic studies indicate that XoxF1 does not efficiently oxidize formaldehyde produced from methanol oxidation during normal metabolism.[21][28][40][55][58] Instead, dedicated formaldehyde oxidation pathways (H₄MPT and glutathione-dependent systems) handle the majority of formaldehyde oxidation, with XoxF1 and ExaF contributing secondary capacity primarily under conditions where canonical pathways are compromised by mutation.[21][28][40][58] This discrepancy suggests that while XoxF1 possesses formaldehyde oxidation catalytic capability, this activity is not evolutionarily optimized and may represent a secondary function with limited physiological relevance under normal growth conditions.
Regulatory Mechanism Ambiguity. The precise mechanism through which XoxF1 senses and signals lanthanide availability remains incompletely understood, despite clear demonstration that XoxF1 protein is required for proper lanthanide-responsive gene regulation.[1][8][24][33][56] Some evidence suggests lanthanide sensing is mediated through XoxF1 protein directly, while other evidence indicates additional XoxF1-independent lanthanide sensing mechanisms operate in parallel.[24][33][56] Resolution of this mechanistic question will refine our understanding of XoxF1's role in regulatory signaling.
ExaF Functional Partitioning. The relative physiological contributions of XoxF1 versus ExaF to formaldehyde oxidation during lanthanide-dependent metabolism remain incompletely characterized, with different studies suggesting different relative contributions depending on strain background and specific growth conditions.[21][28][40][53][58] A comprehensive analysis of the temporal dynamics of formaldehyde oxidation through each pathway under defined conditions would clarify their respective roles.
Organism-Specific Differences and Strain Variation
Strain AM1 vs. PA1 vs. Strain 22A. While the three most extensively studied M. extorquens strains (AM1, PA1, and strain 22A) all possess functional xoxF1 genes with similar lanthanide-dependent methanol oxidation functions, notable differences exist in their auxiliary metabolic systems and regulatory responses. Strain AM1 possesses two xoxF paralogs (XoxF1 and XoxF2) with redundant functions, while strains PA1 and 22A appear to have single primary xoxF genes (though 22A contains a xoxF2 pseudogene with low transcription).[21][24][38][40] Additionally, strain 22A possesses the glutathione-dependent formaldehyde oxidation pathway absent from AM1, representing a metabolic diversity relevant to GO annotation specificity.
Type I vs. Type II Methanotrophs. XoxF1 genes are present and functional in both type I and type II methanotrophs, yet some evidence suggests strain-specific differences in relative contributions of XoxF versus MxaFI to total methanol oxidation capacity and in the specific lanthanides preferred for optimal catalytic function.[38]
Recommendations for GO Annotation Curation
Priority Molecular Function Annotations
TIER 1 (Recommended for Immediate Annotation):
- GO:0008812 - alcohol dehydrogenase (acceptor) activity [EVIDENCE: IDA (Direct Assay)]
- Kinetic characterization fully supports this term
- Extensive purification and enzyme assay data available
- Multiple independent laboratories confirm activity
-
PMID citations: [5][6][19][27][43][46][55]
-
GO:0003824 - catalytic activity [EVIDENCE: IDA]
- General parent term appropriately describing enzymatic catalysis
- Supports hierarchical search results
TIER 2 (Recommended with Caveats):
- GO:0015948 - methanol metabolic process [FOR BIOLOGICAL PROCESS - EVIDENCE: IDA, IGI]
- Direct enzymatic involvement in methanol catabolism confirmed
- Genetic evidence demonstrates essential role in methanol utilization
-
PMID citations: [1][4][6][8][9][15][34]
-
GO:0005506 - iron binding [NOT RECOMMENDED]
- Would be misleading as XoxF1 depends on lanthanides, not iron
- Ontology confusion risk
Priority Cellular Component Annotations
TIER 1 (Recommended):
- GO:0042597 - periplasmic space [EVIDENCE: IDA (Immunoelectron Microscopy), IEM (Cell Fractionation)]
- Multiple independent localization studies confirm periplasmic location
- Signal peptide-directed secretion mechanism demonstrated
- Immunoelectron microscopy visualization provided
- PMID citations: [6][19][29][30]
Priority Biological Process Annotations
TIER 1 (Recommended):
- GO:0015948 - methanol metabolic process [EVIDENCE: IDA, IGI]
- Essential role demonstrated through knockouts and complementation
- Catalytic function directly linked to methanol utilization
- Multiple model organisms examined
-
PMID citations: [1][4][6][8][9][15][34]
-
GO:0006118 - electron transport [EVIDENCE: TAS]
- Electrons from methanol oxidation feed into respiratory chain
- Mechanism well-characterized through cytochrome c mediation
- PMID citations: [34][43][51][54]
Cofactor and Metal-Binding Annotations (Special Considerations)
Issue: Current GO lacks specific terms for lanthanide binding and lanthanide-dependent catalytic function. The lanthanide dependence represents a fundamental distinguishing feature of XoxF1 versus canonical alcohol dehydrogenases.
Recommended Approach:
- Annotate protein's catalytic function using most appropriate existing terms (GO:0008812)
- Add detailed comment or definition modification noting lanthanide requirement and PQQ cofactor dependence
- Flag for GO Consortium developers to create lanthanide-specific terms in forthcoming ontology revisions
- Document in annotation notes the specific lanthanides that support activity (La³⁺, Ce³⁺, Pr³⁺, Nd³⁺) and kinetic parameters for each
Cross-References for Comprehensive Curation
InterPro Families:
- IPR006994: Quinoprotein alcohol dehydrogenase
- IPR006903: NAD(P)H-dependent oxidoreductase domain (though PQQ-specific subfamilies need better delineation)
Enzyme Commission (EC) Number:
- EC 1.1.99.8 (proposed - alcohol dehydrogenase accepting acceptor)
- Note: Lanthanide dependency makes this assignment ambiguous with traditional EC 1.1.99 class
CAZy/CarbohydrateActive Enzymes:
- Not typically applicable as XoxF1 is not carbohydrate-active
InterPro Note: XoxF1 classification requires careful cross-checking with UniProt family annotations to ensure consistency with other lanthanide-dependent alcohol dehydrogenases and to maintain distinctions from calcium-dependent MxaF enzymes.
Conclusions and Summary for GO Curation
XoxF1 represents a well-characterized lanthanide-dependent methanol dehydrogenase enzyme from Methylobacterium/Methylorubrum extorquens with extensive experimental evidence supporting its molecular function, cellular localization, and biological process roles. The enzyme catalyzes lanthanide-dependent oxidation of methanol to formaldehyde in the periplasm through a mechanism involving a PQQ prosthetic group and novel lanthanide coordination geometry distinct from traditional metal-dependent enzymes. XoxF1 functions not only as a direct catalyst but also participates in complex regulatory cascades sensing lanthanide availability and coordinating expression of competing metabolic systems.
Recommended GO annotations should emphasize the core catalytic function (alcohol dehydrogenase activity), periplasmic localization, and essential role in methanol metabolism, while documenting through annotation notes and cross-references the unique lanthanide and PQQ cofactor requirements that distinguish this enzyme from canonical metal-dependent alcohol dehydrogenases. The current GO system would be significantly enhanced through development of lanthanide-specific terms that would enable more precise functional characterization of the growing class of lanthanide-dependent enzymes being discovered throughout the bacterial world.
The substantial body of biochemical, genetic, structural, and metabolic evidence provided in the cited literature (references [1] through [59]) fully supports the recommended GO annotations with high confidence levels across direct assay, genetic interaction, and inferred evidence categories.
Citations
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3194914/
- https://www.nature.com/articles/s41598-020-69401-4
- https://bio-protocol.org/exchange/minidetail?id=7973724&type=30
- https://www.uniprot.org/uniprotkb/C5B120/entry
- https://journals.asm.org/doi/10.1128/mbio.01708-21
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3507691/
- https://pubmed.ncbi.nlm.nih.gov/34544276/
- https://journals.asm.org/doi/10.1128/jb.00937-15
- https://pubmed.ncbi.nlm.nih.gov/24816778/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4859581/
- https://pubs.acs.org/doi/10.1021/bi500850j
- https://journals.plos.org/plosone/article?id=10.1371%2Fjournal.pone.0050480
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4309699/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8106445/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7356819/
- https://pubmed.ncbi.nlm.nih.gov/27779663/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6850437/
- https://www.semanticscholar.org/paper/fe24a9cc3b8e0271d3d486c1a7db83857c97acbf
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6814260/
- https://pubmed.ncbi.nlm.nih.gov/32486139/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3650538/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5018670/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4859578/
- https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2022.820327/full
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9424927/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC12482762/
- https://journals.asm.org/doi/10.1128/jb.00959-15
- https://pubmed.ncbi.nlm.nih.gov/16244138/
- https://journals.asm.org/doi/10.1128/aem.01304-25
- https://pubmed.ncbi.nlm.nih.gov/1619963/
- https://pubmed.ncbi.nlm.nih.gov/21873495/
- https://www.khanacademy.org/science/ap-biology/cellular-energetics/environmental-impacts-on-enzyme-function/a/basics-of-enzyme-kinetics-graphs
- https://pmc.ncbi.nlm.nih.gov/articles/PMC12180729/
- https://www.nature.com/articles/s41598-019-41043-1
- https://journals.asm.org/doi/10.1128/AEM.03292-14