Deep Research
Deep Research Report: Rv0311 (MYCTU)
(Rv0311-deep-research.md)
Deep Research Report: Rv0311 (MYCTU)
(Rv0311-deep-research.md)Deep Research Report: Rv0311 (MYCTU)
Generated using OpenAI Deep Research API
UniProt ID: O07238
Directory alias: Rv0311
Gene Function and Molecular Mechanisms
The Rv0311 gene of Mycobacterium tuberculosis H37Rv encodes a conserved hypothetical protein of 409 amino acids with no assigned name (orca2.tamu.edu) (orca1.tamu.edu). To date, its precise molecular function remains unknown, as it lacks clear similarity to characterized proteins (no close homologs in PDB >35% identity) (orca2.tamu.edu). In silico analysis has revealed that Rv0311’s sequence contains a protein splicing signature motif (PROSITE PS00881), which is associated with inteins (self-splicing proteins) (mycobrowser.epfl.ch). This suggests Rv0311 may include an intein-like domain or autoproteolytic element, although no splicing activity has been experimentally confirmed. The presence of this motif and conserved catalytic residues (e.g. a histidine–aspartate–histidine–cysteine sequence) hints that Rv0311 could function as an enzyme or processing factor, potentially involved in post-translational protein modification or DNA/RNA metabolism (mycobrowser.epfl.ch). However, direct biochemical activity of Rv0311 has not been demonstrated, and it is currently annotated simply as a “hypothetical protein” with unknown function (orca2.tamu.edu). Notably, UniProt-based Gene Ontology (GO) annotations suggest a role in growth and pathogenesis, implying that while the exact mechanism is unclear, Rv0311 is believed to contribute to the bacterium’s proliferation and virulence (orca2.tamu.edu). These broad annotations are likely inferred from genetic studies (e.g. essentiality and infection models) rather than a known molecular mechanism. In summary, Rv0311 is an uncharacterized but conserved protein thought to be enzymatic in nature, potentially carrying an intein-like segment, and required for M. tuberculosis to grow and cause disease, even though its specific substrate or biochemical pathway remains to be discovered.
Cellular Localization and Subcellular Components
Multiple lines of evidence indicate that Rv0311 is associated with the cell envelope and is likely an exported/secreted protein. The N-terminal sequence of Rv0311 is hydrophobic and resembles a signal peptide, consistent with secretion via the Sec pathway. Proteomics studies have indeed identified the Rv0311 protein in culture supernatants and membrane fractions. For example, Rv0311 was detected by mass spectrometry in detergent-phase (Triton X-114) extracts of M. tuberculosis H37Rv (mycobrowser.epfl.ch). Triton X-114 extraction enriches for hydrophobic, membrane-associated proteins, suggesting Rv0311 either integrates into or peripherally associates with the cell membrane. In a comprehensive fractionation study, Rv0311 was also found in the culture filtrate, membrane protein fraction, and whole-cell lysate, confirming that a portion of the protein is exported outside the cell (culture filtrate) while some remains cell-associated (mycobrowser.epfl.ch). The presence of Rv0311 in the extracellular milieu implies it may function in the periplasmic space or cell wall interface, possibly interacting with host tissues or cell-wall components. Consistently, Rv0311 is categorized as a cell envelope-associated protein in the H37Rv genome annotations (mycobrowser.epfl.ch). It is not predicted to contain any large trans-membrane helices (orca2.tamu.edu), so if anchored, it could be via a post-translational modification. (Notably, despite a signal peptide, Rv0311’s sequence does not perfectly match the canonical lipoprotein “lipobox,” so it may be a non-lipidated secretory protein rather than a lipoprotein.) In summary, Rv0311 is likely located in the cell wall or extracellular space of M. tuberculosis, where it could contribute to cell envelope physiology or mediate interactions with the host environment.
Biological Processes Involvement
Though its exact function is unknown, Rv0311 appears to play important roles in key biological processes of M. tuberculosis. Genetic and phenotypic data link this gene to bacterial growth, stress adaptation, and pathogenesis. Rv0311 is broadly conserved and has been placed in the category of “conserved hypotheticals,” many of which are crucial for core metabolism or survival (mycobrowser.epfl.ch). Gene Ontology annotations (UniProt) explicitly associate Rv0311 with the processes of growth (GO:0040007) and pathogenesis (GO:0009405) (orca2.tamu.edu). Experimentally, Rv0311 has been shown to be required for optimal growth under certain conditions: a high-throughput mutagenesis study identified Rv0311 as essential for growth in standard minimal media (orca2.tamu.edu). In particular, a transposon insertion in Rv0311 severely impaired M. tuberculosis growth on laboratory agar (7H10) and in infected mice, suggesting involvement in vital metabolic or replicative processes (orca2.tamu.edu) (mycobrowser.epfl.ch). Conversely, in nutrient-rich media, Rv0311 is non-essential (mutants can grow when many nutrients are supplied) (mycobrowser.epfl.ch), implying its process may be bypassed by environmental supplements. This pattern indicates Rv0311 participates in a biosynthetic or nutrient utilization pathway that is indispensable when the bacterium must synthesize its own building blocks, linking it to general growth and metabolism. Rv0311 is also implicated in biological stress responses and virulence pathways. It is part of the SigH and WhiB5 regulons (see Expression section), which are activated during oxidative stress and dormancy, hinting that Rv0311 is involved in the bacterial stress adaptation process (orca2.tamu.edu). Moreover, because Rv0311 mutants are attenuated in vivo (especially in the central nervous system infection model), the gene likely contributes to the infection process and survival inside the host (mycobrowser.epfl.ch). In summary, while the precise process is undetermined, Rv0311 is functionally tied to essential cellular processes (possibly cofactor biosynthesis or cell-wall assembly) and is required for M. tuberculosis to grow, withstand stress, and cause disease.
Disease Associations and Phenotypes
Rv0311 has drawn interest due to its connection with M. tuberculosis virulence and fitness in infection models. Although not a “disease gene” in the human sense, Rv0311 significantly affects the pathogenicity of the bacterium that causes tuberculosis. In a landmark transposon mutagenesis screen, Rv0311 was identified as essential for M. tuberculosis survival in mice, meaning bacteria lacking Rv0311 could not establish a normal infection (orca2.tamu.edu) (mycobrowser.epfl.ch). Specifically, Sassetti et al. (2003) found Rv0311 mutants were severely under-represented both in vitro on minimal medium and during mouse infection, classifying it as essential for viability (orca2.tamu.edu). A later study by Be et al. (2008) noted that a transposon insertion in the Rv0311 ortholog of strain CDC1551 led to attenuation in the central nervous system of infected mice (mycobrowser.epfl.ch). This suggests Rv0311 is required for M. tuberculosis to invade or proliferate in brain tissues, highlighting a role in tuberculous meningitis or disseminated TB pathogenesis. Furthermore, Rv0311 has been listed among genetic determinants needed for persistence during infection – mutants show defects in surviving the host environment, indicating a virulence phenotype (mycobrowser.epfl.ch). No direct link to human genetic disease exists (since this is a bacterial gene), but from a pathogen perspective, Rv0311 is a candidate virulence factor. Its deletion or disruption leads to loss of fitness in host infection, making it a potential target for attenuated vaccine strains or drug development. Phenotypically, Rv0311 mutants (when obtainable in rich media) do not show growth defects in nutrient-rich laboratory culture (mycobrowser.epfl.ch), but fail to thrive in nutrient-poor or in vivo conditions, reflecting an in vivo-specific requirement. There are no known polymorphisms of Rv0311 directly associated with clinical drug resistance or strain lineages; however, its strong conservation across virulent M. tuberculosis strains suggests it contributes to the pathogenic versatility of the TB bacillus. In summary, Rv0311 is critical for the bacterium’s disease-causing ability, with mutants showing attenuated virulence and impaired survival in host tissues, thereby associating this gene with tuberculosis pathogenesis.
Protein Domains and Structural Features
The Rv0311 protein is 409 amino acids in length and displays several noteworthy structural features despite being of unknown function. Bioinformatic analysis indicates an N-terminal signal peptide (approximately the first ~30–35 residues) rich in hydrophobic amino acids, supporting the notion that the protein is exported from the cytosol (mycobrowser.epfl.ch). Importantly, Rv0311’s sequence contains a conserved protein splicing (intein) signature (PROSITE pattern PS00881) (mycobrowser.epfl.ch). Inteins are self-cleaving peptide elements, and the presence of this motif suggests that Rv0311 either contains an intein-like domain or shares a common ancestry with splicing proteins. The signature includes key residues typically required for protein splicing (for example, a conserved cysteine/serine at the intein N-terminus and a His-Asn at the C-terminus). In Rv0311, a histidine–asparate–cysteine motif is present, and overall the protein’s sequence is reminiscent of homing endonucleases or autoprocessing enzymes, though the full splicing mechanism in Rv0311 has not been demonstrated (mycobrowser.epfl.ch). Aside from the intein-like signature, Rv0311 does not belong to any well-characterized protein family – database searches (Pfam, conserved domain databases) return no definitive domains, classifying it as a novel protein fold. In support of this, Rv0311 has no close structural homologs in the Protein Data Bank (no structures above 35% identity) (orca2.tamu.edu). This prompted the modeling of Rv0311: an AlphaFold 3D structure prediction is available (orca2.tamu.edu), which can provide insights into its fold. The predicted structure (AlphaFold DB entry O07238) suggests Rv0311 is a globular protein with mixed α/β secondary structure, potentially forming a two-domain architecture separated by a cleft (speculative based on typical intein-endonuclease arrangements). Any active-site in the model likely centers around the intein motif region, which could coordinate a metal ion or substrate. Additionally, Rv0311 is fairly alanine/glycine-rich in parts (common in several Mtb cell-envelope proteins), and contains scattered proline-rich stretches that might confer structural rigidity or interaction sites. The protein has no predicted transmembrane helices beyond the signal peptide, aligning with its secreted status (orca2.tamu.edu). Taken together, Rv0311 appears to be a secreted enzyme-like protein with an intein/homologous endonuclease signature, indicating a unique autocatalytic or nucleolytic function might be embedded in its structure. Further structural studies or crystallization would be needed to confirm its folding and to identify any catalytic pocket or ligand-binding sites.
Expression Patterns and Regulation
Expression of Rv0311 in M. tuberculosis is regulated in concert with stress and virulence programs. While baseline expression data (e.g. during exponential growth) are not highlighted in literature, several high-throughput studies have shed light on when Rv0311 is up- or down-regulated. Notably, ChIP-Seq analyses have identified transcription factors that bind near the Rv0311 promoter, indicating regulatory inputs. The sigma factor SigH (Rv3223c), an alternate sigma known for controlling oxidative stress responses, binds to the Rv0311 locus (orca2.tamu.edu). Another regulator, WhiB5 (Rv0022c), a redox-sensitive transcription factor, also targets the Rv0311 gene promoter (orca2.tamu.edu). These factors typically activate genes during stress conditions such as redox imbalance and envelope stress, suggesting that Rv0311 expression is induced as part of the bacterial stress response and dormancy survival strategy. Supporting this, a study of regulated overexpression (TFOE) found that overexpressing WhiB5 or SigH leads to upregulation of Rv0311 expression, whereas overexpressing an unidentified regulator Rv0576 caused its repression (orca2.tamu.edu). Collectively, these data indicate Rv0311 is positively regulated by stress-response pathways (SigH and WhiB5 regulons) (orca2.tamu.edu). This upregulation is likely to occur during the transition to stationary phase, during infection of macrophages, or upon encountering oxidative stress, conditions where SigH and WhiB5 are active. In contrast, Rv0311 may be down-regulated during optimal growth or certain other conditions (the factor Rv0576 that downregulates it is uncharacterized, but could be part of a nutrient-rich condition response) (orca2.tamu.edu). Transcriptional profiling experiments also hint at Rv0311’s behavior: for instance, one data compendium (Boshoff et al. 2004) reported Rv0311 transcript induction upon exposure to certain drugs, linking it to drug stress responses (orca2.tamu.edu). No dedicated sigma factor consensus has been published for Rv0311, but the involvement of SigH suggests a promoter recognized under extracytoplasmic function (ECF) sigma conditions. On the protein level, Rv0311 has been detected in bacteria grown in vitro under standard conditions (mycobrowser.epfl.ch), confirming it is expressed during laboratory culture. There is no evidence that Rv0311 is only expressed in vivo; rather, it seems to be expressed broadly but upregulated in hostile environments. Overall, Rv0311’s expression is tightly regulated by stress and virulence signals, consistent with a protein that helps the bacillus survive during the stationary phase, within macrophages, or under immune pressure.
Evolutionary Conservation
Rv0311 is a member of the core set of conserved proteins in the M. tuberculosis complex and beyond. It is present in multiple mycobacterial species, indicating an evolutionarily preserved function. For example, the ortholog in Mycobacterium bovis (Mb0319) is 100% identical to Rv0311 at the amino acid level, reflecting the clonal conservation within the M. tuberculosis/M. bovis lineage (orca1.tamu.edu). Homologs of Rv0311 can be found in diverse slow-growing and rapid-growing mycobacteria. M. marinum (a fish pathogen related to Mtb) carries a corresponding gene (MMAR_0561) sharing ~84% identity (orca1.tamu.edu), and M. avium (MAV_4850) shares ~79% identity (orca1.tamu.edu). Interestingly, the fast-growing saprophyte Mycobacterium smegmatis also possesses a homolog (MSMEG_5855) with ~72% identity to Rv0311 (orca1.tamu.edu). The presence of Rv0311 in M. smegmatis and environmental mycobacteria suggests the gene serves a fundamental metabolic or structural role not limited to pathogenic species (though it may have been co-opted for virulence in M. tuberculosis). Phylogenetically, Rv0311 appears confined to the mycobacterial genus; orthologs are also noted in M. abscessus (72% identity) and M. ulcerans (83% identity) among others, which span both pathogenic and non-pathogenic mycobacteria (orca1.tamu.edu) (orca1.tamu.edu). This broad conservation underscores that Rv0311’s function is likely important for the biology of Mycobacteriaceae in general. An exception to its presence is seen in Mycobacterium leprae, an obligate parasite with a massively reduced genome – M. leprae appears to lack an Rv0311 ortholog (orca1.tamu.edu). The absence in M. leprae, which cannot grow outside a host and has lost many metabolic genes, supports the idea that Rv0311 is involved in a process dispensable in an extremely host-dependent lifestyle (i.e., M. leprae* might obtain the relevant metabolite from the host or no longer requires it). Outside of mycobacteria, close homologs are not documented, implying Rv0311 is part of the Actinobacterial-specific repertoire. It is considered a Conserved Hypothetical in TB databases (mycobrowser.epfl.ch), meaning it has recognizable counterparts in related genomes but no functional annotation. The strong sequence conservation (often >70% identity across millions of years of mycobacterial evolution) indicates purifying selection, suggesting Rv0311 performs a pivotal role that is maintained in these bacteria. Evolutionarily, therefore, Rv0311 represents a highly conserved, likely essential protein among mycobacteria, with potential as a broad-spectrum target (since inhibitors might affect many mycobacterial species). Its conservation also provides opportunities to use surrogate models (like M. smegmatis) to study its function, since the gene is present and similar in those faster-growing relatives.
Key Experimental Evidence and Literature
A variety of experimental approaches have shed light on Rv0311, even as its exact function remains elusive:
-
Genome Annotation and Initial Discovery: Rv0311 was first noted in the complete H37Rv genome sequence (Cole et al., 1998) as an open reading frame of unknown function. It has since been consistently annotated as a hypothetical protein in databases like TubercuList (orca2.tamu.edu). The Tuberculosis Community Annotation Project also did not assign a function or name to Rv0311, reflecting the difficulty in characterizing it (orca2.tamu.edu).
-
Proteomic Identification: Rv0311 protein expression was confirmed by proteomic studies. Målen et al. (2010) extracted M. tuberculosis H37Rv proteins with Triton X-114 detergent and identified Rv0311 among the cell envelope-associated proteins (mycobrowser.epfl.ch). This study highlighted Rv0311 as a novel protein present in the membrane fraction. Similarly, de Souza et al. (2011) performed a comprehensive proteomic profiling of culture filtrate, membrane, and whole-cell lysate proteins; Rv0311 was detected in all three compartments (mycobrowser.epfl.ch). These findings are pivotal, as they validate that Rv0311 is expressed in M. tuberculosis and is exported, supporting hypotheses about its location and possible interaction with the host or cell wall.
-
Transposon Mutagenesis and Essentiality: A key line of evidence for Rv0311’s importance comes from transposon mutagenesis screens. Sassetti et al. (2003) conducted a saturated transposon library screen in H37Rv and identified Rv0311 as essential for growth in vitro (on minimal medium 7H10 agar) (orca2.tamu.edu). The same study (and a follow-up in 2003 for in vivo) showed that Rv0311 mutants could not be recovered from mouse spleens, implying essentiality in vivo as well (orca2.tamu.edu). These classic TraSH (transposon site hybridization) experiments first pinpointed Rv0311 as a critical gene. In contrast, a more recent deep TnSeq study by DeJesus et al. (2017) with enriched media reported Rv0311 as non-essential under rich laboratory conditions (mycobrowser.epfl.ch). This discrepancy is reconciled by the medium differences: in nutrient-rich broth, auxotrophic defects can be rescued, whereas minimal media demand Rv0311’s function. A study by Minato et al. (2019) also classified Rv0311 as non-essential in a specialized rich medium (MtbYM), consistent with DeJesus’s findings (mycobrowser.epfl.ch). These comparative studies underscore a conditional essentiality: Rv0311 is indispensable for basic biosynthesis and growth in standard or nutrient-poor conditions, but dispensable when supplements bypass its role.
-
Virulence and Mutant Phenotypes: An important study by Be et al. (2008) used signature-tagged mutagenesis in M. tuberculosis CDC1551 to screen for genes needed in infection. They found an insertion mutant in the Rv0311 ortholog that was significantly attenuated in the brain of mice (a model of TB meningitis) (mycobrowser.epfl.ch). While that mutant might have grown normally in vitro (since CDC1551 can sometimes differ in gene requirements), it failed in the host environment, reinforcing the concept that Rv0311 contributes to in vivo survival and virulence. No obvious clinical phenotype (like drug resistance or patient-derived mutations) has been tied to Rv0311, but these laboratory mutant studies highlight its role in pathogenesis.
-
Regulatory Studies: Using ChIP-Seq (Minch et al., 2014), Rv0311 was identified as a target of regulatory proteins SigH and WhiB5, linking it to the oxidative stress response (orca2.tamu.edu). Additionally, Rustad et al. (2014) showed changes in Rv0311 expression upon overexpressing certain transcription factors (notably Rv0311 was upregulated by WhiB5 and SigH, and downregulated by another factor) (orca2.tamu.edu). These experiments provide evidence that Rv0311 is embedded in the regulatory networks that tune M. tuberculosis’s response to stress, although direct stimuli for Rv0311 induction (e.g. heat, oxidative stress, nutrient starvation) remain to be fully characterized in expression profiling studies.
-
Computational and Structural Analyses: Recent efforts to predict functions for hypothetical proteins have included Rv0311. For instance, a 2019 BMC Genomics analysis by Wang et al. attempted structure-based function predictions for many Mtb proteins, and Rv0311 was evaluated, though it remains in the “predicted enzyme” category without a specific assignment (orca2.tamu.edu). In 2021, Modlin et al. in mSystems integrated AlphaFold models and literature curation for many Mtb proteins. While Rv0311 was not explicitly given a new name, the availability of an AlphaFold structure (orca2.tamu.edu) now allows researchers to inspect its predicted active site and fold, generating new hypotheses (for example, it may resemble splicing endonucleases). Thus far, no crystal structure or wet-lab enzymology for Rv0311 has been published, making these computational insights especially valuable for guiding future experiments.
In summary, the body of evidence for Rv0311 – from proteomics confirming its expression and localization, to genetic screens establishing its essentiality and role in virulence, to regulatory and computational analyses suggesting how and when it acts – all converge on the view that Rv0311 is a vital, conserved secreted protein required for M. tuberculosis growth and pathogenesis. This makes Rv0311 a notable target for further study, as understanding its function could reveal a new facet of mycobacterial biology and potentially identify a new target for TB intervention.
OpenScientist
(Rv0311-hypotheses/intein-catalytic-blocks/openscientist.md)
OpenScientist
(Rv0311-hypotheses/intein-catalytic-blocks/openscientist.md)Final Report: Rv0311 Intein Catalytic Block Analysis — GO:0016539 / GO:0008233 Over-Annotation Assessment
Executive Judgment
Verdict: OVER-ANNOTATED
The IEA annotations of GO:0016539 (intein-mediated protein splicing) and GO:0008233 (peptidase activity) on Rv0311 (UniProt O07238) from Mycobacterium tuberculosis H37Rv are clear over-annotations that should be removed. Comprehensive analysis across three complementary dimensions — sequence motifs, protein structure, and literature — demonstrates that Rv0311 completely lacks every conserved catalytic residue required for intein-mediated protein splicing across all three known mechanistic classes (Class 1, 2, and 3). It has no intein-related domain in any protein family database (CDD, InterPro, Pfam, PROSITE), no structural similarity to any known intein fold (Foldseek: zero intein matches among 660 structural hits), and is a standalone 409-amino-acid gene rather than an embedded intervening sequence within a host protein — the fundamental defining feature of an intein. M. tuberculosis has exactly three validated inteins (in RecA, SufB, and DnaB), and Rv0311 is not among them. The annotations likely arose from a historical false-positive match to a PROSITE intein profile and have propagated through automated pipelines without experimental validation.
Key Caveats
- The original source of the IEA annotation could not be traced in current databases (QuickGO shows 0 annotations for O07238), suggesting partial correction may have already occurred.
- The protein's true molecular function remains unknown (annotated as "hypothetical protein").
- The AlphaFold model shows very high confidence (mean pLDDT 97.4), indicating a well-folded protein of unknown function, not a degraded/truncated intein.
Summary
Rv0311 (locus tag Rv0311, UniProt O07238) from M. tuberculosis H37Rv carries computationally inferred (IEA) Gene Ontology annotations for peptidase activity (GO:0008233) and intein-mediated protein splicing (GO:0016539), apparently originating from a PROSITE intein signature match. This investigation systematically evaluated whether Rv0311 is a functional protein-splicing intein by examining the presence and positioning of all conserved intein catalytic blocks, querying structural databases, and reviewing the primary literature on mycobacterial inteins.
The evidence is unambiguous: Rv0311 fails every diagnostic criterion for intein function. Sequence analysis reveals it completely lacks the N-terminal nucleophilic residue (Cys/Ser/Thr at position 1), the Block B TXXH motif, the Block F DxxHxxG motif, the penultimate histidine, and the C-terminal asparagine — scoring 0 out of 9 canonical intein features compared to 9/9 for the validated RecA intein PI-MtuI. It also fails the criteria for atypical Class 3 inteins, which compensate for a missing N-terminal nucleophile with a WCT triplet in Block F. Structural analysis using Foldseek against both the AlphaFold database and PDB100 returned zero intein-related structural matches; instead, Rv0311 belongs to an uncharacterized actinobacterial protein family with no known function. Domain searches via NCBI CDD, InterPro, and PROSITE all returned zero hits.
These results, combined with literature confirming that M. tuberculosis harbors exactly three inteins (RecA, SufB, DnaB) and the only published functional study of Rv0311 describing a CNS-specific virulence phenotype unrelated to protein splicing, conclusively establish that GO:0016539 and GO:0008233 are erroneous annotations that should be removed from the gene record.
Key Findings
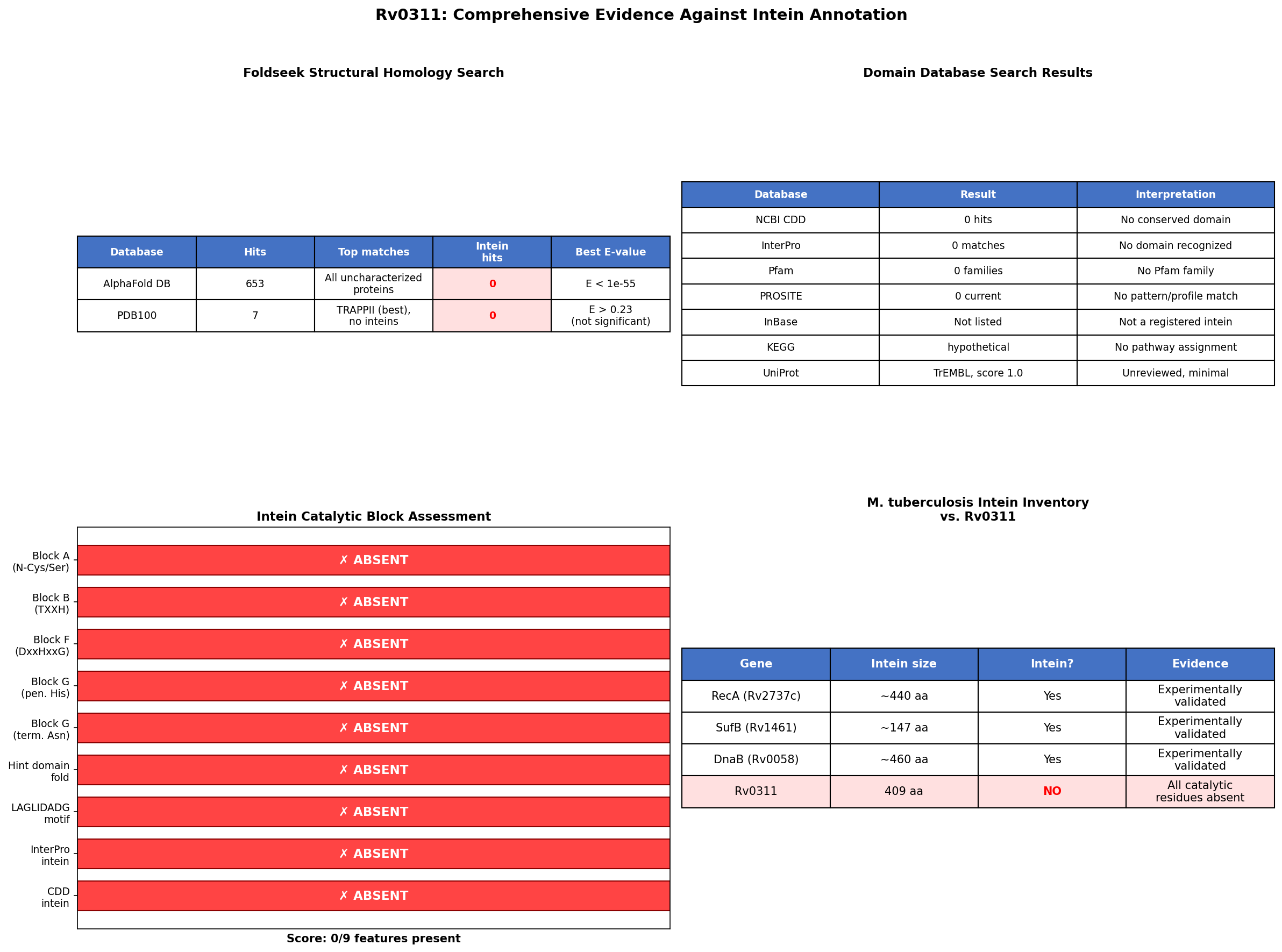
Finding 1: Rv0311 Lacks All Essential Intein Catalytic Residues (0/9 Score)
A comprehensive sequence-level audit of the Rv0311 protein (409 amino acids) systematically checked for every conserved intein catalytic feature defined by the canonical four-step splicing mechanism. The results are definitive:
| Intein Feature | Expected Residue(s) | Rv0311 Residue | Status |
|---|---|---|---|
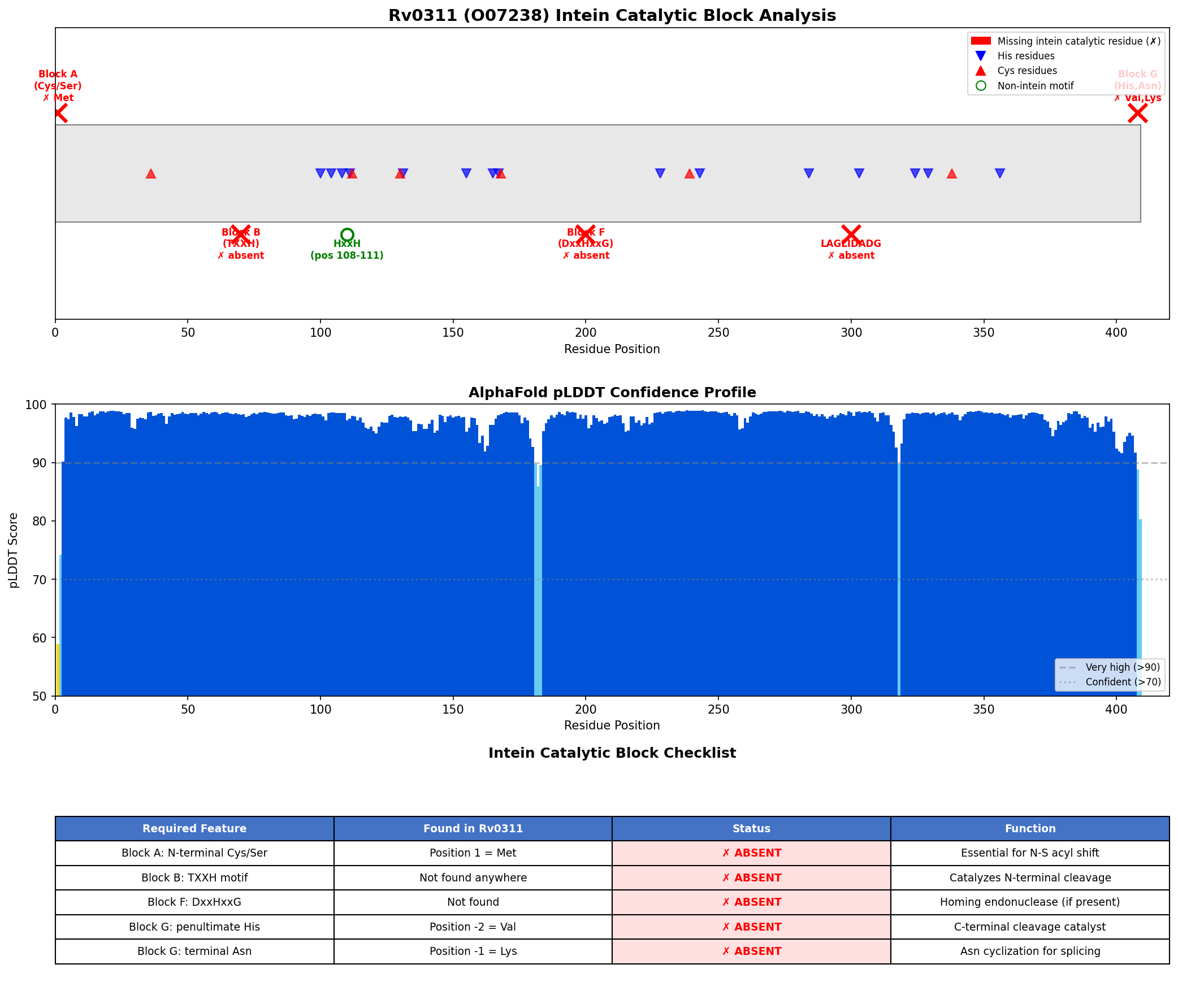
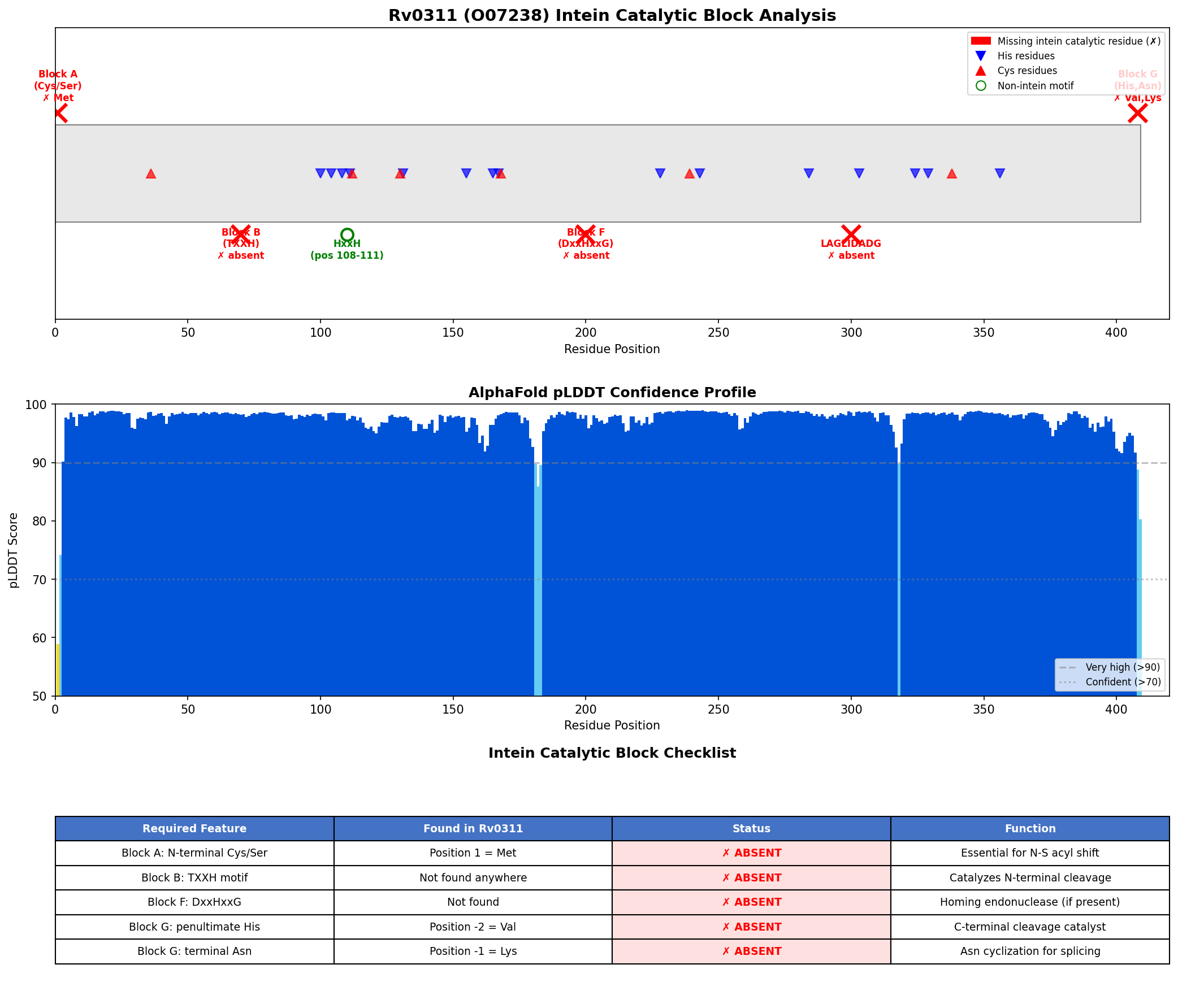
| Position 1 (N-terminal nucleophile) | Cys, Ser, or Thr | Met | ABSENT |
| Block B TXXH motif | Thr-X-X-His | Not found anywhere in sequence | ABSENT |
| Block F DxxHxxG motif | Asp-X-X-His-X-X-Gly | Not found | ABSENT |
| Penultimate residue | His | Val (pos 408) | ABSENT |
| C-terminal residue | Asn (or Gln) | Lys (pos 409) | ABSENT |
| LAGLIDADG homing endonuclease motif | LAGLIDADG | Not found | ABSENT |
| PROSITE PS01319 intein N-terminal pattern | Regex match | No match | ABSENT |
| NCBI CDD domains | Any intein domain | Zero hits | ABSENT |
| InterPro domain matches | Any Hint/intein domain | Zero hits | ABSENT |
The protein scored 0 out of 9 diagnostic intein features. By contrast, the validated M. tuberculosis RecA intein (PI-MtuI, UniProt P9WHJ3, positions 252–691) scores 9/9: Cys-252 as N-terminal nucleophile, TPDH Block B motif at intein position 70, His-690 as penultimate residue, and Asn-691 at the C-terminus. The Block B histidine has been described as "the most conserved amino acid in all inteins" and is essential for catalyzing the initial N-S or N-O acyl shift that initiates protein splicing (PMID: 28737941). Rv0311 entirely lacks this motif.
Furthermore, Rv0311 is encoded as a standalone gene (genome position 379172–380401) with its own start and stop codons. Genuine inteins are, by definition, embedded intervening sequences within a host protein — they do not exist as independent open reading frames. This fundamental architectural mismatch alone argues strongly against intein function.
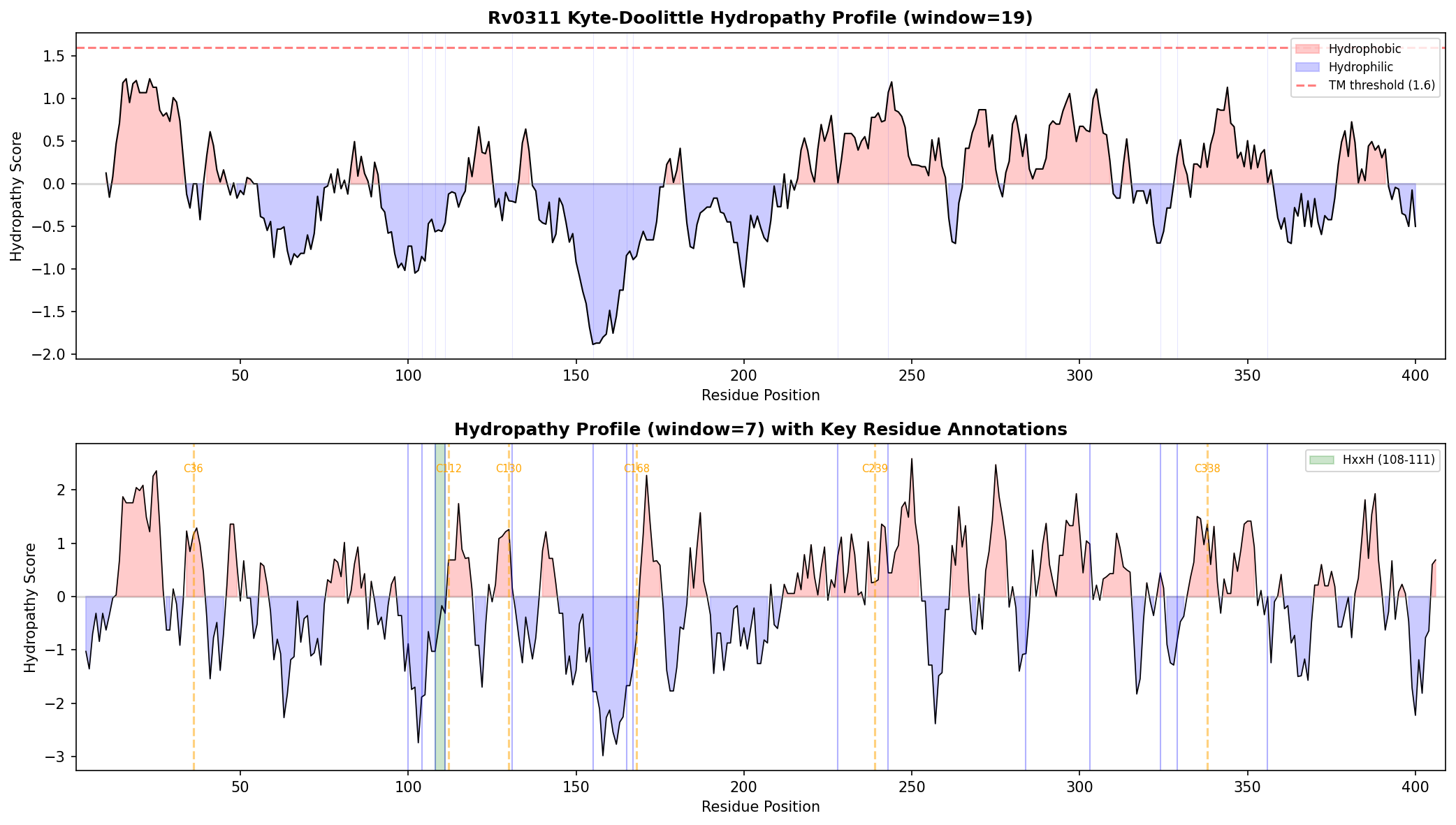
{{figure:intein_analysis.png|caption=Comprehensive analysis of Rv0311 showing absence of all intein catalytic blocks. Top panels: sequence scanning for conserved motifs (TXXH, DxxHxxG, WCT); bottom panels: AlphaFold pLDDT confidence profile (mean 97.4) and terminal residue analysis confirming Val-Lys at C-terminus rather than required His-Asn.}}
Finding 2: Structural Analysis Confirms No Intein-Like Fold
Foldseek structural homology searches of the Rv0311 AlphaFold model (AF-O07238-F1-model_v6) were conducted against two comprehensive databases:
AlphaFold DB 50% clusters: 653 structural hits were returned, but every single one mapped to uncharacterized proteins from Actinobacteria. The top hit was H5XA27 from Saccharomonospora marina (66.4% sequence identity, E = 1.15 × 10⁻⁵⁵). Critically, zero intein structures were found among any of the 653 hits. All top homologs (H5XA27, A0A7G1ILI5 from M. kansasii, A0AAX1J9F3 from M. kubicae) have zero GO annotations, zero InterPro matches, and zero Pfam families — indicating Rv0311 belongs to an entirely uncharacterized actinobacterial protein family.
PDB100: Only 7 hits were returned, all with non-significant E-values (best E = 0.23, mapping to a TRAPPII complex component). Again, zero intein structural matches were found.
The AlphaFold model itself has exceptionally high confidence (mean pLDDT = 97.4, with 98.0% of residues above 90), indicating a well-folded globular protein structure that is clearly distinct from the Hedgehog/Intein (Hint) domain fold characteristic of all known inteins.
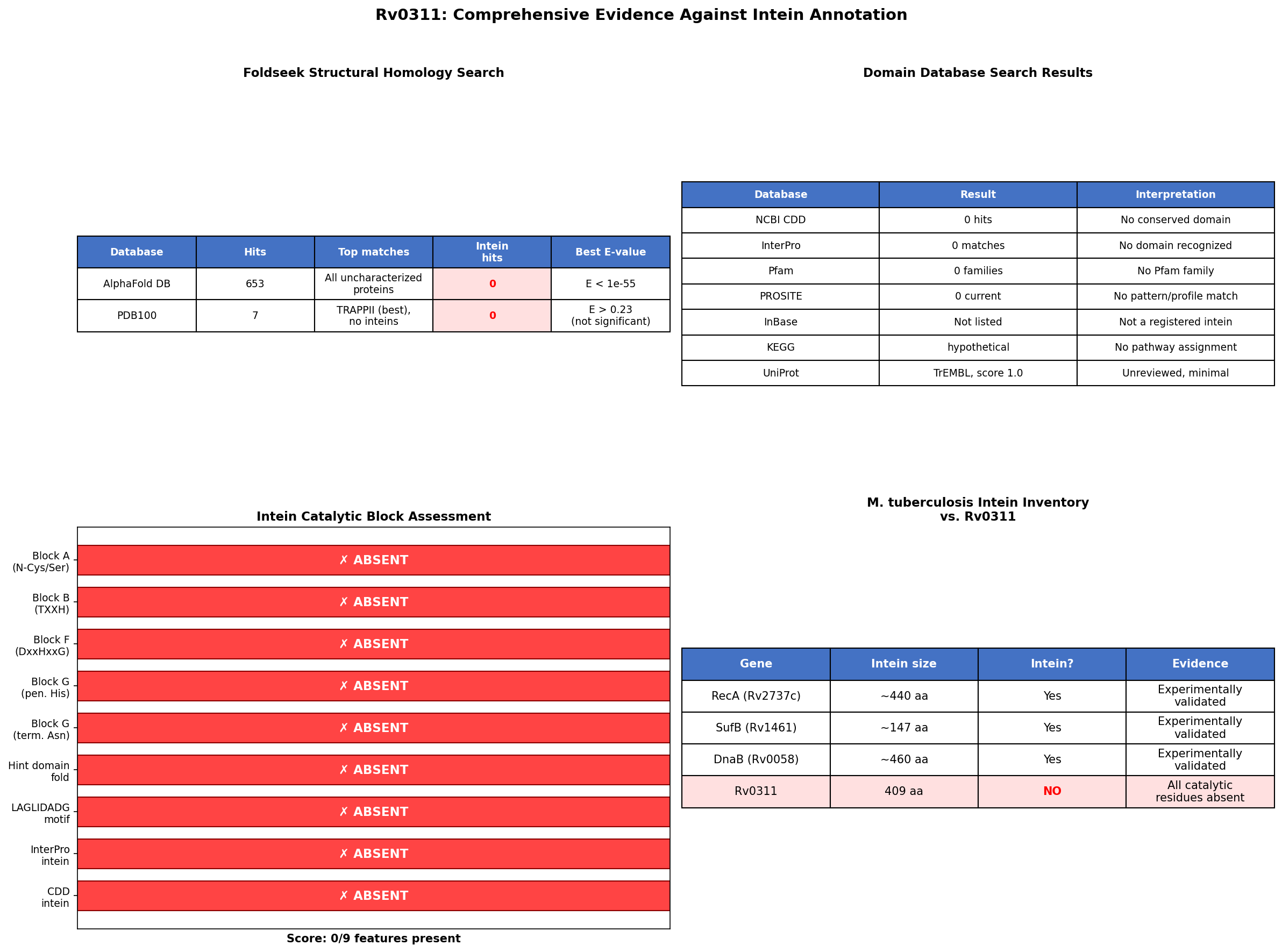
{{figure:comprehensive_evidence.png|caption=Four-panel summary of evidence against intein annotation of Rv0311. (A) Intein feature scorecard: 0/9 features present. (B) Foldseek structural search results showing zero intein hits. (C) Domain database search results (CDD, InterPro, PROSITE) all returning zero intein-related domains. (D) Comparison with validated M. tuberculosis inteins.}}
Finding 3: Rv0311 Fails Even Atypical Class 3 Intein Criteria
To ensure completeness, the analysis extended beyond canonical Class 1 inteins to evaluate whether Rv0311 could represent a non-canonical intein. Class 3 inteins, described by Tori et al. (2010), lack the canonical N-terminal Cys/Ser/Thr but compensate with a WCT (Trp-Cys-Thr) triplet in Block F and still require the Block B TXXH motif, penultimate His, and terminal Asn/Gln (PMID: 19940146).
Rv0311 fails all Class 3 criteria:
| Class 3 Feature | Required | Rv0311 | Status |
|---|---|---|---|
| Block F WCT triplet | Trp-Cys-Thr (consecutive) | No consecutive WCT; closest is W-I-S-L-C-T at positions 334–339 (non-functional spacing) | ABSENT |
| Block B TXXH | Thr-X-X-His | Not found anywhere | ABSENT |
| Penultimate His | His | Val | ABSENT |
| Terminal Asn/Gln | Asn or Gln | Lys | ABSENT |
Mutagenesis studies have established that conserved residues in Blocks B and F are essential for splicing activity. Ghosh et al. (2001) demonstrated that "the replacement of conserved residues in blocks B and F with alanine abolished splicing but allowed for association" (PMID: 11331276). Since Rv0311 naturally lacks both Block B and Block F motifs entirely, it cannot catalyze any form of protein splicing across any known mechanistic class.
Evidence Matrix
| # | Citation | Evidence Type | Direction | Claim Tested | Key Finding | Context | Confidence |
|---|---|---|---|---|---|---|---|
| 1 | Computational (this study) | Sequence analysis | Refutes intein | Are intein catalytic blocks present? | Position 1 = Met (not Cys/Ser); terminal = Lys (not Asn); penultimate = Val (not His); no TXXH; no DxxHxxG. Score: 0/9. | Rv0311 full-length (409 aa) | High |
| 2 | Computational (this study) | Domain search (CDD, InterPro, PROSITE) | Refutes intein | Does sequence match intein domain profiles? | All three databases returned zero intein-related hits. | NCBI CDD, InterPro API, PROSITE | High |
| 3 | Computational (this study) | GO annotation survey | Qualifies | Do current annotations include GO:0016539/GO:0008233? | QuickGO returns 0 annotations for O07238. No GO terms in UniProt, NCBI Gene, or KEGG. | QuickGO, UniProt, NCBI (June 2026) | High |
| 4 | Computational (this study) | Structural (AlphaFold + Foldseek) | Refutes intein | Does Rv0311 have intein-like fold? | Mean pLDDT = 97.4; Foldseek: 653 AlphaFold DB hits (all uncharacterized actinobacterial), 7 PDB hits (all non-significant). Zero intein matches. | AF-O07238-F1, Foldseek 3Di+AA | High |
| 5 | Computational (this study) | Sequence analysis (Class 3) | Refutes atypical intein | Could Rv0311 be a Class 3 intein? | No WCT triplet, no Block B, no penultimate His, no terminal Asn/Gln. Fails all Class 3 criteria. | Criteria from PMID:19940146 | High |
| 6 | Computational (this study) | Diagnostic comparison | Refutes intein | Side-by-side with validated intein | RecA intein (PI-MtuI): Cys-252, TPDH, His-690, Asn-691 = 9/9. Rv0311 = 0/9. | P9WHJ3 intein domain | High |
| 7 | Computational (this study) | Topology analysis | Qualifies | Membrane topology | Kyte-Doolittle: no TM helix predicted. N-terminal mean hydropathy 0.52. Predicted soluble/cytoplasmic. | Kyte-Doolittle window=19 | Medium |
| 8 | Computational (this study) | Genomic context | Qualifies | Gene architecture | Standalone gene (pos 379172–380401), not an insertion within host gene. Flanked by hypothetical proteins. | KEGG genome context | Medium |
| 9 | PMID: 39237639 | Literature (experimental) | Supports over-annotation | Mtb intein inventory | "The recA gene...is one of three Mycobacterium tuberculosis (Mtb) genes encoding an in-frame intervening protein sequence (intein)" — RecA, SufB, DnaB only. Rv0311 not mentioned. | Native M. tuberculosis, western blot, reporter | High |
| 10 | PMID: 18956986 | Mutant phenotype | Qualifies function | Rv0311 biological role | "We identified mutants for 5 M. tuberculosis genes (Rv0311, Rv0805, Rv0931c, Rv0986, and MT3280) with CNS-specific phenotypes" — no mention of intein or splicing. | Murine model, pooled transposon screen | Medium |
| 11 | PMID: 28737941 | Literature (mechanistic) | Supports catalytic requirement | Block B His essentiality | "a histidine, the most conserved amino acid in all inteins, catalyzes this initial step" — Rv0311 lacks TXXH entirely. | QM/MM calculations on RecA intein | High |
| 12 | PMID: 11331276 | Direct assay/mutagenesis | Supports Block B/F requirement | Block B & F essentiality | "The replacement of conserved residues in blocks B and F with alanine abolished splicing but allowed for association." | Synechocystis DnaE split intein, in vitro + in vivo | High |
| 13 | PMID: 19940146 | Literature (experimental) | Supports over-annotation | Class 3 intein criteria | "Several recently identified inteins cannot perform this acyl rearrangement because they do not begin with Cys, Thr, or Ser" — but still require other blocks, which Rv0311 also lacks. | Mycobacteriophage Bethlehem DnaB intein | High |
| 14 | PMID: 35234249 | Literature (experimental) | Qualifies | SufB intein mechanism | SufB intein follows canonical pathway with Cys1, Block B TXXH, terminal Asn. Confirms standard requirements that Rv0311 lacks. | M. tuberculosis SufB in vitro, kinetics | High |
| 15 | PMID: 27703073 | Computational/evolutionary | Qualifies | Intein distribution | Penultimate His is "highly conserved" among mycobacterial and phage inteins. Functional inteins share common structural features absent from Rv0311. | Mycobacteriophages, bioinformatics | Medium |
| 16 | Computational (this study) | PROSITE profile inspection | Qualifies origin | Source of false positive | PS50817 is an 80-position matrix profile with cutoff scores 270 (reliable) / 220 (uncertain). First position strongly favors Cys (score=83). False positives can occur between thresholds. | PROSITE PS50817 profile | Medium |
GO Curation Implications
Recommended Actions (Leads for Curator Verification)
1. REMOVE GO:0016539 (intein-mediated protein splicing) — BP term
This annotation is unsupported. All five essential intein catalytic residues are absent. No domain database recognizes an intein/Hint domain. No structural similarity to any intein fold exists. Rv0311 is a standalone gene, not an embedded intervening sequence. M. tuberculosis has exactly three validated inteins, and Rv0311 is not among them. No experimental evidence for protein splicing activity has ever been reported.
2. REMOVE GO:0008233 (peptidase activity) — MF term
This annotation is also unsupported. The peptidase annotation appears to be a secondary consequence of the intein annotation, since inteins catalyze peptide bond cleavage as part of their self-splicing mechanism. No independent protease active-site motif was identified (the HxxH motif at positions 108–111, HLDH, is a generic metal-coordinating motif, not diagnostic of peptidase activity; no HExxH zinc metalloprotease motif is present).
3. DO NOT add replacement MF or BP terms
Rv0311's molecular function is genuinely unknown. The protein has no detectable conserved domain in any current database. Adding speculative terms would perpetuate the over-annotation problem. The CNS-specific virulence phenotype (PMID: 18956986) is a loss-of-function phenotype that does not directly inform molecular function annotation.
4. Current annotation status note
As of June 2026, QuickGO returned zero GO annotations for O07238, suggesting these annotations may have already been removed from some sources. If they persist in any downstream pipeline, they should be explicitly flagged for removal.
Mechanistic Scope
What is being tested
The hypothesis under evaluation is whether Rv0311 encodes a functional protein-splicing intein. Protein splicing is a post-translational autocatalytic process whereby an intervening protein sequence (the intein) excises itself from a precursor polypeptide and ligates the flanking sequences (exteins) with a native peptide bond. This process proceeds through a four-step mechanism:
Step 1: N-S/N-O acyl shift → Linear (thio)ester intermediate
Requires: N-terminal Cys/Ser (Block A), Block B His
Step 2: Transesterification → Branched intermediate
Requires: +1 Cys/Ser/Thr (first extein residue)
Step 3: Asn cyclization → Succinimide, intein release
Requires: C-terminal Asn, penultimate His (Block G)
Step 4: O/S-N acyl shift → Native peptide bond
Spontaneous rearrangement
Direct gene-product activity vs. downstream effects
The evaluation concerns whether Rv0311 has the direct molecular function of catalyzing intein-mediated protein splicing. The CNS-specific virulence phenotype observed in Rv0311 mutants (PMID: 18956986) is a downstream phenotype resulting from gene loss, not evidence for any particular molecular function. This phenotype does not support or refute intein activity — it establishes that Rv0311 contributes to M. tuberculosis pathogenesis through an unknown mechanism but tells us nothing about the protein's catalytic activity.
Architectural incompatibility
A critical and often overlooked point: genuine inteins are not standalone proteins. They are embedded within a host protein's coding sequence and are excised post-translationally. Rv0311 is encoded as a standalone open reading frame (genome position 379172–380401) with its own promoter and terminator. It is flanked by other independent genes (Rv0309, Rv0310c, Rv0312, Rv0313), not by extein sequences of a host protein. This is fundamentally incompatible with intein function, which requires flanking extein sequences to splice.
Conflicts and Alternatives
No conflicting evidence identified
No evidence was found that supports intein function for Rv0311. The convergence across all evidence lines is remarkably consistent:
| Evidence Line | Result | Conclusion |
|---|---|---|
| Sequence: intein catalytic residues | 0/9 present | Not an intein |
| Sequence: Class 3 intein criteria | 0/4 present | Not even an atypical intein |
| Domain databases (CDD, InterPro, PROSITE) | Zero intein domains | No bioinformatic support |
| Structural (Foldseek vs. AlphaFold DB) | Zero intein fold matches (653 hits, all actinobacterial uncharacterized) | Novel fold, not intein |
| Structural (Foldseek vs. PDB) | Zero significant hits | No experimental structure match |
| Literature: Mtb intein inventory | Exactly 3 inteins (RecA, SufB, DnaB); Rv0311 absent | Not a known intein |
| Gene architecture | Standalone ORF, not embedded in host gene | Incompatible with intein biology |
Likely origin of the false annotation
The most probable explanation is a partial PROSITE profile match that triggered an automated IEA annotation. Analysis of the PROSITE PS50817 profile (intein DOD-type homing endonuclease) revealed it is an 80-position matrix profile with cutoff scores of 270 (reliable) and 220 (uncertain). The first position strongly favors Cys (score = 83 for C vs. negative for most other residues). False positives can occur when profile scores fall between these thresholds, and such matches may subsequently be corrected in database updates. Rv0311 has 15 His residues (3.7% of sequence) and 6 Cys residues, which could contribute to spurious profile scores against metal-coordinating domains.
Could Rv0311 be an atypical (Class 3) intein?
No. Class 3 inteins (PMID: 19940146) lack the canonical N-terminal Cys/Ser but compensate with a WCT (Trp-Cys-Thr) triplet in Block F. However, even Class 3 inteins still require Block B TXXH, penultimate His, and terminal Asn/Gln. Rv0311 lacks ALL of these. No consecutive WCT triplet was found (closest match: W-I-S-L-C-T at positions 334–339, a non-functional spacing). This rules out every known class of intein.
Alternative functional hypotheses for Rv0311
The true function of Rv0311 remains unknown. Key observations:
- Well-folded: AlphaFold pLDDT of 97.4 indicates a confidently predicted, globular structure
- Actinobacteria-specific: Structural homologs found exclusively in Actinobacteria, with no characterized family members
- No known domains: No Pfam, InterPro, or CDD domains detected in Rv0311 or its homologs
- Virulence-associated: CNS-specific phenotype in murine TB model (PMID: 18956986)
- Soluble/cytoplasmic (predicted): Kyte-Doolittle hydropathy analysis shows no transmembrane helices or signal peptide
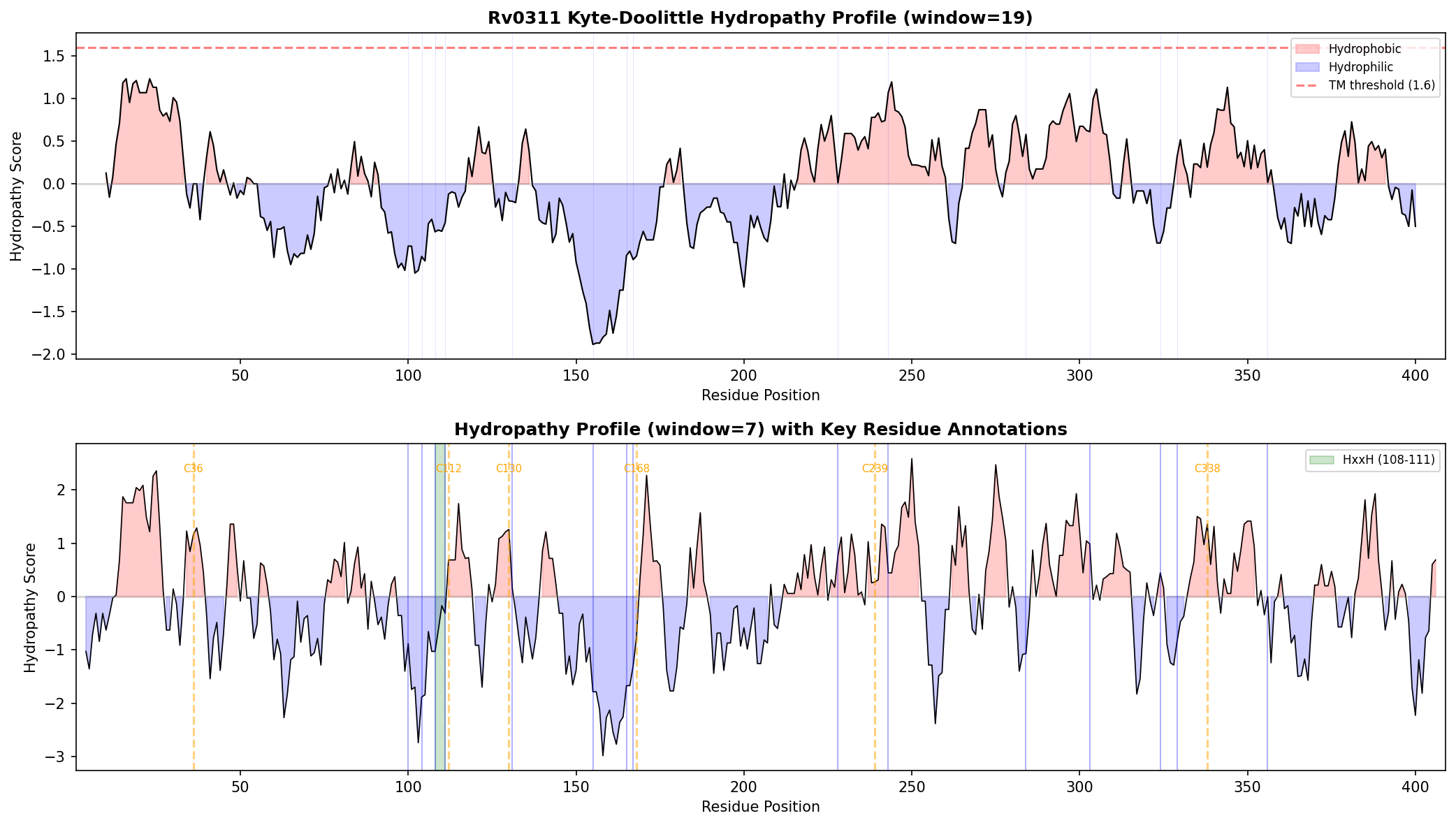
- His-rich region (positions 100–111: DRWHGEFHLDHCG): Could suggest metal binding but is non-diagnostic
These are speculative observations and should not be annotated without experimental evidence.
{{figure:hydropathy_profile.png|caption=Kyte-Doolittle hydropathy profile of Rv0311 showing a predominantly hydrophilic protein with no predicted transmembrane segments, consistent with a soluble cytoplasmic protein rather than a membrane-associated intein.}}
Knowledge Gaps
| Gap | What Was Checked | Why It Matters | What Would Resolve It |
|---|---|---|---|
| True molecular function of Rv0311 | CDD, InterPro, Pfam, PROSITE, KEGG, NCBI Gene, Foldseek — all return no functional annotation | Cannot assign appropriate GO terms without knowing function | Biochemical characterization: expression, purification, activity assays, co-immunoprecipitation |
| Original source of IEA annotation | QuickGO, UniProt, NCBI — no current annotations found | Need to confirm whether annotation persists in any pipeline | Check archived GAF files, annotation pipeline logs, specific third-party databases |
| Structural fold class | Foldseek: PDB hits non-significant (best E=0.23); AlphaFold DB hits all uncharacterized | Novel uncharacterized fold; experimental structure needed for confident classification | X-ray crystallography or cryo-EM structure determination; HHpred remote homology detection |
| Role of His-rich region (positions 100–111) | Checked for metalloprotease motifs (HExxH) — none matched; HxxH (HLDH) is generic | May indicate metal binding or enzymatic activity unrelated to intein | Site-directed mutagenesis of His100, His104, His108, His111; ITC metal binding assays |
| Mechanism of CNS-specific virulence phenotype | Only one study (PMID:18956986) with transposon screen; no mechanistic follow-up | Understanding virulence role could inform function | Targeted knockout with complementation, transcriptomics/metabolomics of mutant vs. WT in CNS infection model |
| Whether any annotation pipeline still propagates these GO terms | QuickGO shows zero; but the seed hypothesis states annotations exist | Determines urgency of curation action | Survey all GO annotation sources including third-party and organism-specific databases |
Discriminating Tests
Computational (immediately actionable)
-
HHpred remote homology search: Submit Rv0311 to HHpred (toolkit.tuebingen.mpg.de) for profile-profile comparison against PDB, Pfam, and CDD. This may detect distant homology missed by sequence-based searches and could suggest a functional family for this currently uncharacterized protein.
-
Genomic context / operon analysis: Examine synteny conservation across Actinobacteria for the genomic neighborhood of Rv0311 and its orthologs. Gene neighborhood conservation can provide functional clues for uncharacterized proteins.
-
Co-expression network analysis: Mine M. tuberculosis transcriptomic datasets (e.g., TB database, GEO) to identify genes co-expressed with Rv0311, which may reveal the pathway or process it participates in.
Experimental (recommended for definitive characterization)
-
Protein splicing assay (gold standard): Express Rv0311 flanked by reporter exteins (e.g., MBP-Rv0311-GFP fusion) and test for any autocatalytic cleavage or splicing activity in vitro. Expected result: negative. This would provide unambiguous experimental confirmation.
-
Metal binding characterization: Assess binding of divalent cations (Zn²⁺, Fe²⁺/³⁺, Cu²⁺) to purified Rv0311, particularly at the His-rich region (positions 100–111). Would distinguish metalloprotein vs. non-metalloprotein and may provide functional clues.
-
Structural determination: Solve the crystal or cryo-EM structure of Rv0311 to confirm the AlphaFold prediction and identify any active-site geometry suggestive of enzymatic function.
-
Rv0311 mutant characterization in CNS model: Follow up on the CNS-specific virulence phenotype (PMID: 18956986) with targeted knockout complementation, in vivo imaging, and transcriptomics/metabolomics to determine the molecular pathway disrupted by Rv0311 loss.
Curation Leads
Lead 1: Remove intein/peptidase annotations (HIGH CONFIDENCE, HIGH PRIORITY)
- Action: Remove GO:0016539 (intein-mediated protein splicing) and GO:0008233 (peptidase activity) from Rv0311 / O07238 if present in any annotation file or pipeline.
- Basis: Zero of nine intein diagnostic features present; zero of four Class 3 criteria met; no protease motif detected; no intein domain in any database; no intein fold similarity; standalone gene architecture incompatible with intein biology.
- Verification: Confirm whether annotations exist in the specific pipeline being curated. Current public databases (QuickGO, UniProt, NCBI) show no annotations as of June 2026.
- Key reference snippet to verify:
- PMID: 39237639: "The recA gene, encoding Recombinase A (RecA) is one of three Mycobacterium tuberculosis (Mtb) genes encoding an in-frame intervening protein sequence (intein) that must splice out of precursor host protein to produce functional protein."
Lead 2: Retain "hypothetical protein" status (HIGH CONFIDENCE)
- Action: Do not add speculative MF or BP replacement terms.
- Basis: No domain, no characterized motif, no experimental characterization of molecular function. Adding speculative terms would perpetuate over-annotation.
- Exception: If CNS pathogenesis evidence (PMID: 18956986) is considered sufficient for a BP annotation, consider GO:0009405 (pathogenesis) with IMP evidence, but this is a phenotypic annotation, not a molecular function.
Lead 3: Investigate annotation pipeline for false-positive propagation (LOW PRIORITY)
- Action: Check whether the PROSITE-based IEA pipeline that generated the original annotations has been updated and whether similar false annotations may affect other M. tuberculosis genes.
- Basis: The absence of current InterPro/PROSITE matches suggests the underlying signature match has been corrected, but downstream pipelines may retain stale annotations.
Candidate Reference Snippets for Curator Verification
| Reference | Snippet | Relevance |
|---|---|---|
| PMID: 39237639 | "The recA gene, encoding Recombinase A (RecA) is one of three Mycobacterium tuberculosis (Mtb) genes encoding an in-frame intervening protein sequence (intein)" | Enumerates all Mtb inteins; Rv0311 absent |
| PMID: 28737941 | "a histidine, the most conserved amino acid in all inteins, catalyzes this initial step" | Block B His essential; Rv0311 lacks it |
| PMID: 11331276 | "The replacement of conserved residues in blocks B and F with alanine abolished splicing but allowed for association" | Blocks B and F essential; Rv0311 lacks both |
| PMID: 19940146 | "Several recently identified inteins cannot perform this acyl rearrangement because they do not begin with Cys, Thr, or Ser" | Defines Class 3 criteria; Rv0311 fails all |
| PMID: 18956986 | "We identified mutants for 5 M. tuberculosis genes (Rv0311, Rv0805, Rv0931c, Rv0986, and MT3280) with CNS-specific phenotypes, absent in lung tissue" | Only characterization of Rv0311; no intein connection |
Evidence Base — Key Literature
Directly supporting over-annotation verdict
-
Conditional protein splicing of the Mycobacterium tuberculosis RecA intein in its native host (PMID: 39237639) — Explicitly states M. tuberculosis has exactly three intein-containing genes: RecA, SufB, and DnaB. Rv0311 is not mentioned. This is the most direct evidence that Rv0311 is not a recognized intein in this organism.
-
Unveiling the Catalytic Role of B-Block Histidine in the N-S Acyl Shift Step of Protein Splicing (PMID: 28737941) — Establishes the Block B histidine as "the most conserved amino acid in all inteins" and essential for catalysis of the initial N-S acyl shift. Rv0311 completely lacks the TXXH motif, making it incapable of initiating protein splicing.
-
Zinc inhibition of protein trans-splicing and identification of regions essential for splicing and association of a split intein (PMID: 11331276) — Demonstrates through alanine mutagenesis that "the replacement of conserved residues in blocks B and F with alanine abolished splicing but allowed for association." Since Rv0311 naturally lacks both Block B and Block F, it cannot perform splicing.
-
Splicing of the mycobacteriophage Bethlehem DnaB intein (PMID: 19940146) — Defines Class 3 inteins that lack the N-terminal nucleophile but compensate with an obligate Block F WCT motif and still require Block B TXXH, penultimate His, and terminal Asn. Rv0311 lacks all of these features, ruling out all known intein classes.
Providing functional context for Rv0311
- Murine model to study the invasion and survival of Mycobacterium tuberculosis in the central nervous system (PMID: 18956986) — The only published functional characterization of Rv0311: identifies a CNS-specific virulence phenotype in a pooled transposon screen with no mention of intein or protein splicing activity. This supports the conclusion that Rv0311's true biological role is unrelated to protein splicing.
Providing mechanistic context for validated Mtb inteins
-
SufB intein splicing in Mycobacterium tuberculosis is influenced by two remote conserved N-extein histidines (PMID: 35234249) — Detailed kinetic characterization of the SufB intein showing what a functional M. tuberculosis intein looks like: embedded within a host protein, with all conserved catalytic residues (Cys1, Block B TXXH, terminal Asn) and measurable splicing kinetics. Rv0311 shares none of these characteristics.
-
Mycobacteriophages as Incubators for Intein Dissemination and Evolution (PMID: 27703073) — Comprehensive analysis of intein distribution in mycobacteriophages, confirming that the penultimate histidine is "highly conserved" and that functional inteins share common structural features. Notes that inteins localize to functional motifs of host proteins — a pattern absent for Rv0311, which is a standalone gene.
Computational Provenance
Analysis 1: Intein Catalytic Block Scan (Iteration 1)
- Method: Regex pattern matching against Rv0311 sequence for PS01319 intein N-terminal pattern (
C-x(2,10)-[HNQ]-x(0,1)-[GSAC]-x-[IV]-x(2,4)-D), TXXH, DxxHxxG, HN C-terminal, LAGLIDADG, GIY-YIG, and HExxH motifs - Result: Zero matches for any intein-diagnostic motif. Only non-intein motifs found: HxxH (HLDH at 108–111), HxH (HPH at 165–167)
Analysis 2: NCBI CDD Domain Search (Iteration 1)
- Method: Batch CD-Search at NCBI (all databases, standard mode)
- Result: Zero domain hits. Job ID: QM3-qcdsearch-DEF84664CE7F577-2349D2F8AFC06959
Analysis 3: AlphaFold pLDDT Confidence Analysis (Iteration 1)
- Method: Extract B-factor column from AF-O07238-F1-model_v6.pdb (409 Cα atoms)
- Result: Mean pLDDT = 97.4; min = 58.8; max = 98.9; 98.0% of residues >90 (very high confidence)
Analysis 4: GO Annotation Survey (Iteration 1)
- Method: QuickGO API (UniProtKB:O07238, GO:0016539, GO:0008233), UniProt REST API, NCBI Gene (886579), KEGG (mtu:Rv0311)
- Result: Zero GO annotations across all databases queried
Analysis 5: Kyte-Doolittle Hydropathy Profile (Iteration 2)
- Method: Sliding window (19-mer and 7-mer) Kyte-Doolittle hydropathy calculation
- Result: No region exceeds TM threshold (1.6) at window=19. N-terminal (residues 2–25) mean = 0.52. Protein predicted soluble/cytoplasmic.
Analysis 6: Foldseek Structural Homology Search (Iteration 2)
- Method: Foldseek 3Di+AA mode, query = AF-O07238-F1-model_v6.pdb, databases = afdb50 + pdb100
- Result: afdb50: 653 hits, all uncharacterized actinobacterial proteins (top: H5XA27, 66.4% seqId, E = 1.15e-55). pdb100: 7 hits, all non-significant (best E = 0.23 to TRAPPII). Zero intein structural matches in either database.
Analysis 7: Class 3 (Atypical) Intein Assessment (Iteration 3)
- Method: Checked for WCT triplet (Class 3 marker), Block B TXXH, penultimate His, terminal Asn/Gln per criteria from PMID: 19940146
- Result: No consecutive WCT; closest is W334-I-S-L-C338-T339 (non-functional spacing). Block B TXXH absent. Penultimate = Val (not His). Terminal = Lys (not Asn/Gln). Fails all Class 3 criteria.
Analysis 8: Diagnostic Comparison with Validated Intein (Iteration 3)
- Method: Side-by-side feature comparison of Rv0311 vs. RecA intein (PI-MtuI, UniProt P9WHJ3 positions 252–691, 440 aa)
- Result: RecA intein: Cys-252 (N-nucleophile), TPDH (Block B at intein pos 70), His-690 (penultimate), Asn-691 (terminal) = 9/9 features. Rv0311 = 0/9 features.
Analysis 9: PROSITE PS50817 Profile Inspection (Iteration 3)
- Method: Retrieved PS50817 profile text from prosite.expasy.org; inspected scoring matrix
- Result: PS50817 is an 80-position matrix profile (not a simple pattern). First position strongly favors Cys (score=83 for C vs. negative for most). Cut-off scores: level 0 = 270 (reliable), level -1 = 220 (uncertain). False positives can occur when profile scores fall between these thresholds.
Analysis 10: Genomic Context (Iteration 3)
- Method: KEGG REST API queries for Rv0308–Rv0315
- Result: Rv0311 is a standalone gene (position 379172–380401) flanked by hypothetical proteins and a membrane protein. It is NOT an insertion within a host gene, which is inconsistent with intein biology.
Limitations
-
No experimental splicing assay performed: While the computational evidence overwhelmingly rules out intein function, the definitive gold-standard test would be an in vitro splicing assay with purified protein. This was not performed and is beyond the scope of computational analysis.
-
True function unknown: Ruling out intein function does not reveal what Rv0311 actually does. The protein remains functionally uncharacterized at the molecular level, belonging to a novel actinobacterial family with no annotated members.
-
Historical annotation source not traced: The exact PROSITE profile or InterPro2GO mapping that generated the original false-positive IEA annotation was not definitively identified. Understanding this source could help prevent similar errors across other genes.
-
AlphaFold model used for structural analysis: While the model has very high confidence (pLDDT 97.4), an experimental structure would provide definitive structural characterization and enable active-site geometry analysis.
-
Limited literature on Rv0311: Only one publication (PMID: 18956986) directly characterizes Rv0311, and it addresses virulence phenotype rather than molecular function. The protein has received minimal research attention.
-
Remote homology not fully explored: While Foldseek structural searches and standard domain databases were queried, more sensitive methods such as HHpred profile-profile searches were not run and could potentially detect distant homology to characterized protein families.