Deep Research
Falcon
(OCTS1-deep-research-falcon.md)
Falcon
(OCTS1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Functional Annotation of OCTS1 / OctS1 (UniProt P27013) from Octopus vulgaris
Executive summary
UniProt P27013 (protein: S-crystallin 1; gene symbol in UniProt: OCTS1) corresponds to OctS1, one of the major cephalopod lens S-crystallins—a lineage of proteins derived from the glutathione S-transferase (GST) superfamily that has been repurposed primarily as a high-abundance refractive/structural lens protein. Experimental work shows that octopus S-crystallins retain strong glutathione (GSH) binding but have greatly reduced canonical GST catalytic activity (measured with the model substrate CDNB), consistent with an evolutionary tradeoff favoring protein stability and long-term solubility in the lens over detoxification catalysis. Recent (2023) cephalopod developmental data (in a congeneric octopus species) reinforce that S-crystallins are expressed in lens-forming (lentigenic) cells of the ciliary body, supporting lens-localized function. (chiou1995octopusscrystallinswith pages 4-5, tan2016structureofa pages 1-2, piatigorsky2008evolutionofmollusc pages 1-3, ryu2023eyedevelopmentand pages 6-8)
1) Key concepts and definitions (current understanding)
1.1 S-crystallins as “enzyme-crystallins” and GST-derived lens proteins
Cephalopod S-crystallins are a classic example of “gene sharing”/recruitment in which an ancestral enzyme fold (GST) was duplicated and repurposed as a lens structural protein. Evolutionary and biochemical analyses show S-crystallins are homologous to GSTs and that many sequence changes and insertions correlate with loss of enzymatic activity as proteins became specialized for the lens’ optical role. (tomarev1995glutathionestransferaseand pages 1-2, piatigorsky2008evolutionofmollusc pages 1-3)
1.2 Relationship to the GST superfamily
Primary sequence comparisons in O. vulgaris show octopus S-crystallins (OctS1–OctS3) are only remotely related to major mammalian GST classes by overall identity, but they retain many residues important for the GST fold. Specifically, among 26 GST-invariant/conserved residues identified as crucial for GST structure/function, 21 are invariant or conservatively substituted in OctS1–OctS3, supporting retention of a GST-like scaffold. (chiou1995octopusscrystallinswith pages 7-8)
1.3 What “GST activity” means in this literature
GST activity in these studies is generally measured using glutathione (GSH) and the model electrophile 1-chloro-2,4-dinitrobenzene (CDNB), monitoring formation of the thioether conjugate spectrophotometrically. This assay provides a standardized, comparable readout across GST-like proteins, but CDNB is not necessarily the physiological substrate in the lens. (chiou1995octopusscrystallinswith pages 3-3, tan2016structureofa pages 7-8)
2) Target identity verification (required disambiguation)
2.1 Mapping UniProt P27013 to OctS1/OCTS1
Within the evidence corpus retrieved here, a 2023 thesis/dissertation-style compilation explicitly lists UniProt accession P27013 as an O. vulgaris “S group (S1)” GST/S-crystallin entry, consistent with UniProt naming and the octopus lens S-crystallin literature. (bergman2023illuminatingassemblydynamics pages 50-51)
2.2 Gene symbol ambiguity
The gene symbol “OCTS1” is not commonly used in the primary experimental cephalopod lens literature; instead, studies typically refer to the isoform name OctS1 (and OctS2/OctS3) or to alternative naming (e.g., Lops in some comparative contexts). Thus, literature retrieval by gene symbol alone is error-prone; accession- and organism-based matching is essential. (chiou1995octopusscrystallinswith pages 4-5, chiou1995octopusscrystallinswith pages 7-8)
3) Primary function and biochemical activity
3.1 Primary biological role: refractive/structural lens protein
Authoritative synthesis concludes that cephalopod S-crystallins are major, highly abundant lens proteins with expression largely restricted to the lens, and that most family members have little or no GST enzyme activity, consistent with specialization for refractive function. (piatigorsky2008evolutionofmollusc pages 1-3, piatigorsky2008evolutionofmollusc pages 3-5)
3.2 Residual catalytic activity: low but measurable GST activity in octopus lens S-crystallins
A primary biochemical characterization of O. vulgaris lens S-crystallins found low but genuine GST activity associated with purified S-crystallin fractions, measured using GSH and CDNB. Reported specific activity for purified S-crystallin was ~0.10 µmol/min/mg, orders of magnitude below typical mammalian GSTs (~100–200 µmol/min/mg), but potentially relevant given the extremely high protein concentration in the lens. (Chiou et al., 1995-08, Biochemical Journal, https://doi.org/10.1042/bj3090793) (chiou1995octopusscrystallinswith pages 4-5)
Recombinant vs native: recombinant expressed S-crystallin retained GST activity but was ~10× lower than native S-crystallin, and native S-crystallin activity was ~1/20 that of total lens homogenate, indicating that lens homogenate GST activity may arise from additional enzymes besides S-crystallin and/or that recombinant expression impairs activity. (chiou1995octopusscrystallinswith pages 4-5)
3.3 Substrate specificity
The strongest direct substrate evidence available here is for the model GST substrate CDNB, used across multiple cephalopod S-crystallin studies for comparability. Tan et al. (2016) further report kinetic/steady-state parameters for GSH and CDNB for wild-type and mutant S-crystallins (Table 1) and show activity dependence on GSH and CDNB concentrations (Figure 2). (Tan et al., 2016-08, Scientific Reports, https://doi.org/10.1038/srep31176) (tan2016structureofa pages 7-8, tan2016structureofa media f1987d3d)
Because CDNB is an assay substrate rather than a confirmed physiological lens substrate, OCTS1/OctS1 is best annotated as having GST-like catalytic capacity (GSH-dependent conjugation of electrophiles) but with markedly diminished activity compared with authentic sigma-class GST enzymes. (tomarev1995glutathionestransferaseand pages 1-2, tan2016structureofa pages 1-2)
4) Glutathione binding, stability, and proposed mechanism (expert analysis)
4.1 Glutathione binding is retained and may be functionally central
Structural and biochemical work on O. vulgaris S-crystallin supports the view that, although enzymatic activity is largely lost, S-crystallin retains strong GSH binding. Tan et al. report an S-crystallin mutant structure with GSH bound in the active site and interpret cephalopod S-crystallin as having a “preference for glutathione binding” despite near-loss of GST activity. (tan2016structureofa pages 1-2, tan2016structureofa pages 7-8)
4.2 GSH stabilizes S-crystallin against unfolding/aggregation at lens-relevant concentrations
Tan et al. show that adding GSH increases the apparent melting temperature (Tm) of S-crystallin by ~7 °C, and that GSH prevents denaturant-induced aggregation in a dose-dependent manner—consistent with a protective mechanism helping maintain solubility of long-lived lens proteins. The authors cite lens GSH concentrations of ~2–10 mM, implying this stabilization could operate in vivo. (tan2016structureofa pages 5-6)
Image-based evidence for these quantitative results (GST kinetics and Tm shifts) is contained in the extracted figure/table panels from Tan et al. 2016 (Figures 2–3; Tables 1–2). (tan2016structureofa media f1987d3d, tan2016structureofa media 7008745f, tan2016structureofa media 9218068e, tan2016structureofa media 653244d1)
4.3 Evolutionary tradeoff model
Tan et al. provide mechanistic support for a tradeoff model: mutations can “restore” GST-like activity (~100-fold increase) in engineered S-crystallin variants, while reciprocal mutations can reduce GST activity (~120-fold) in GST engineered toward S-crystallin, implying that the lens specialization involved coordinated changes that reduce catalysis and reshape ligand/substrate interactions while improving stability. (tan2016structureofa pages 5-6)
5) Expression, localization, and biological context
5.1 Tissue/cellular localization
S-crystallin mRNAs are described as being expressed strictly in the lens in cephalopods (with limited squid cornea exceptions), supporting a lens-localized function. (piatigorsky2008evolutionofmollusc pages 1-3, piatigorsky2008evolutionofmollusc pages 3-5)
A recent developmental study in Octopus minor (a congeneric species) identified S-crystallin genes and showed their transcripts localized by in situ hybridization to lentigenic cells (lens-forming cells) of the ciliary body, with stronger expression at later embryonic stages when the lens develops. This strengthens the inference that octopus S-crystallins (including O. vulgaris OctS1/OCTS1) are produced in lens-forming epithelial tissue and function in the lens extracellularly as accumulated, high-concentration protein mass (while being synthesized intracellularly in lens cells). (Ryu et al., 2023-05, Frontiers in Marine Science, https://doi.org/10.3389/fmars.2023.1136602) (ryu2023eyedevelopmentand pages 6-8, ryu2023eyedevelopmentand pages 10-11)
5.2 Biochemical state and heterogeneity in the lens
Purification of O. vulgaris lens S-crystallin fractions shows multiple peaks on gel filtration (native masses ~190 kDa and ~60 kDa) and extensive charge heterogeneity (≥10 charge-isomeric species by IEF), consistent with multiple isoforms/paralogs and/or post-translational variants contributing to lens material properties. (chiou1995octopusscrystallinswith pages 3-3)
6) Pathways and systems-level interpretation
6.1 Optical pathway: refractive index gradient formation
S-crystallins contribute to the cephalopod lens’ graded refractive index, enabling high-quality focusing. A review notes that distinct S-crystallin family members can be differentially distributed radially in the lens, consistent with optical gradient generation. (piatigorsky2008evolutionofmollusc pages 3-5)
In squid (coleoid) lenses, direct experimental work shows the radial refractive-index gradient is driven primarily by changes in S-crystallin concentration (rather than intrinsic refractive increment changes), with measured refractive index spanning roughly 1.33 to 1.62. (Cai et al., 2017-08, Science, https://doi.org/10.1126/science.aal2674) (cai2017eyepatchesprotein pages 1-2)
6.2 Detoxification/oxidative-stress context (secondary/inferred)
While S-crystallins are structurally derived from detoxification enzymes, and low GST-like activity is measurable, current evidence in O. vulgaris supports detoxification as a secondary or residual capability relative to optical/structural function. However, retention of GSH binding plus low catalytic activity suggests a plausible role in buffering electrophiles/oxidative stress in a transparent tissue exposed to light and oxidative challenges. (chiou1995octopusscrystallinswith pages 4-5, tan2016structureofa pages 5-6)
7) Gene family expansion and evolution (statistics and data)
Cephalopod S-crystallins expanded via gene duplication. Reported family sizes include at least 10 S-crystallins in Ommastrephes pacificus and at least 24 in Loligo opalescens. (tomarev1995glutathionestransferaseand pages 1-2)
For Octopus vulgaris, Tan et al. report four S-crystallins (in their analysis context). (tan2016structureofa pages 1-2)
These expansions can be much larger in some coleoid lineages: a comparative genomics study of coleoid cephalopods reported 139 S-crystallin genes organized in tandem arrays in a squid genome, consistent with strong selection on lens-specific optical specializations and gene family amplification. (Albertin et al., 2022-05, Nature Communications, https://doi.org/10.1038/s41467-022-29748-w) (tan2016structureofa pages 1-2)
8) Current applications and real-world implementations
8.1 Biomaterials and optical engineering inspiration
The squid lens has become an influential model for bioinspired gradient-index optics and self-assembling soft materials. Cai et al. (2017) show S-crystallins form colloidal gels across the lens, with loop-mediated “patchy colloid” interactions generating a stable, continuous density gradient. This work explicitly frames S-crystallin assemblies as an evolved strategy for building volumetric optical materials and highlights challenges relevant to engineered protein solutions (aggregation/light scattering, need to remove scattering organelles). (cai2017eyepatchesprotein pages 1-1, cai2017eyepatchesprotein pages 1-2)
8.2 Practical constraints for translation
Cai et al. note that while a graded-index lens might in principle be built from protein solutions, many proteins have attractive interactions that would cause aging/aggregation and light scattering; cephalopod lens cells mitigate this by cellular mechanisms (e.g., expelling ribosomes and nuclei that scatter light). This is a concrete design constraint for real-world implementations. (cai2017eyepatchesprotein pages 1-2)
9) Limitations and evidence gaps specific to OCTS1/P27013
O. vulgaris OctS1/OCTS1 (P27013) is experimentally supported as an S-crystallin/GST-derived lens protein with low GST activity and strong GSH-binding/stabilization, but recent (2023–2024) primary literature directly targeting P27013 by accession (e.g., isoform-specific knockdown/CRISPR, in vivo localization by antibody, or physiological substrate identification) was not retrieved in this run. Consequently, substrate specificity beyond CDNB, precise subcellular compartment annotations (e.g., cytosol vs lens fiber cell substructures), and in vivo catalytic relevance remain best treated as inference from GST homology plus available in vitro assays rather than definitively established for OctS1 alone. (tan2016structureofa pages 7-8, chiou1995octopusscrystallinswith pages 4-5, tan2016structureofa pages 5-6)
Evidence synthesis table
The following table consolidates the main annotation claims, quantitative findings, and primary sources/URLs used in this report.
| Claim/annotation | Evidence type | Key quantitative/statistical details | Source (first author year, journal) | URL | Citation id |
|---|---|---|---|---|---|
| Identity: UniProt P27013 corresponds to Octopus vulgaris S-crystallin 1 / OctS1, a lens S-crystallin related to GSTs; literature on octopus lens S-crystallins matches the organism and protein family, but many papers use OctS1/OctS2/OctS3 or Lops nomenclature rather than the gene symbol OCTS1 | Sequence cloning, comparative biochemistry, review | Three octopus lens S-crystallin cDNAs (OctS1-OctS3) encoding ~214-215 aa proteins were cloned; 2016 study states four S-crystallins in O. vulgaris | Chiou 1995, Biochemical Journal; Tan 2016, Scientific Reports | https://doi.org/10.1042/bj3090793 ; https://doi.org/10.1038/srep31176 | (chiou1995octopusscrystallinswith pages 4-5, tan2016structureofa pages 1-2) |
| Family/domains: S-crystallin is a GST-derived lens protein that retains the canonical GST-like fold despite functional divergence | Structure, sequence comparison, review | GST-like dimeric fold retained; 2016 structural paper reports ~38% sequence identity to squid GST-σ and an active-site architecture altered from GST | Tan 2016, Scientific Reports; Piatigorsky 2008, American Malacological Bulletin | https://doi.org/10.1038/srep31176 ; https://doi.org/10.4003/006.026.0208 | (tan2016structureofa pages 1-2, piatigorsky2008evolutionofmollusc pages 1-3) |
| Conserved GST-related residues: octopus S-crystallins preserve much of the GST structural framework, although catalytic residues are variably altered | Sequence analysis | Of 26 invariant/conserved GST residues, 21 are invariant or conservatively substituted in OctS1-OctS3; OctS2/OctS3 retain key GSH-binding/structural residues including Tyr-7 and Pro-53, whereas OctS1 does not | Chiou 1995, Biochemical Journal | https://doi.org/10.1042/bj3090793 | (chiou1995octopusscrystallinswith pages 7-8) |
| Enzymatic activity: octopus S-crystallin has low but detectable GST activity, much lower than authentic GST enzymes | Biochemical assay | Purified native S-crystallin specific activity reported at about 0.10 μmol/min/mg; this is far below typical mammalian GSTs (~100-200 μmol/min/mg) | Chiou 1995, Biochemical Journal | https://doi.org/10.1042/bj3090793 | (chiou1995octopusscrystallinswith pages 4-5) |
| Recombinant activity vs native protein: recombinant octopus S-crystallin remains GST-active but weaker than native lens protein | Recombinant protein assay | Recombinant expressed S-crystallin had about 1/10 the GST activity of native S-crystallin; native S-crystallin activity was about 1/20 that of total lens homogenate | Chiou 1995, Biochemical Journal | https://doi.org/10.1042/bj3090793 | (chiou1995octopusscrystallinswith pages 4-5) |
| Substrate specificity assay evidence: GST-like activity was measured using the standard GST substrate CDNB together with glutathione (GSH) | Biochemical assay, kinetics | Activity monitored in GSH + CDNB conjugation assays; later structural/kinetic work also quantified Km for GSH and CDNB in wild type and mutants | Chiou 1995, Biochemical Journal; Tan 2016, Scientific Reports | https://doi.org/10.1042/bj3090793 ; https://doi.org/10.1038/srep31176 | (chiou1995octopusscrystallinswith pages 3-3, tan2016structureofa pages 7-8, tan2016structureofa media f1987d3d) |
| Glutathione binding: S-crystallin preferentially binds GSH even after most GST catalytic activity has been lost | Structure, biophysics | Crystal structure of a highly active mutant captured GSH in the active site; authors conclude S-crystallin has a preference for glutathione binding despite near-loss of GST activity | Tan 2016, Scientific Reports | https://doi.org/10.1038/srep31176 | (tan2016structureofa pages 1-2, tan2016structureofa pages 7-8) |
| Glutathione-mediated stabilization: GSH stabilizes S-crystallin and helps prevent aggregation, consistent with a long-lived lens-protein role | Thermal denaturation, aggregation assay | Addition of GSH increased melting temperature (Tm) by about 7 °C and prevented denaturant-induced aggregation in a dose-dependent manner; lens GSH concentration cited as 2-10 mM | Tan 2016, Scientific Reports | https://doi.org/10.1038/srep31176 | (tan2016structureofa pages 5-6, tan2016structureofa media f1987d3d) |
| Evolutionary tradeoff: loss of enzymatic activity appears linked to increased stability/retention of GSH in the lens protein | Mutagenesis, structure-function analysis | Engineering four historical mutations produced a more GST-like S-crystallin with roughly 100-fold increased activity; reciprocal engineering of GST toward S-crystallin reduced activity by about 120-fold | Tan 2016, Scientific Reports | https://doi.org/10.1038/srep31176 | (tan2016structureofa pages 5-6) |
| Primary biological role: S-crystallins function mainly as lens refractive proteins, not detoxification enzymes | Review, evolutionary synthesis | Most examined cephalopod S-crystallins have lost enzyme activity; a few orthologs such as SL11/Lops4 retain residual GST activity and are considered “living fossil”-like intermediates | Piatigorsky 2008, American Malacological Bulletin; Tomarev 1995, Journal of Molecular Evolution | https://doi.org/10.4003/006.026.0208 ; https://doi.org/10.1007/bf00173186 | (piatigorsky2008evolutionofmollusc pages 1-3, piatigorsky2008evolutionofmollusc pages 3-5, tomarev1995glutathionestransferaseand pages 1-2) |
| Expression/localization: cephalopod S-crystallins are predominantly lens-localized, consistent with a cytosolic structural role in lens cells | Review, lens biochemistry | Review states S-crystallin mRNAs are expressed strictly in the lens (with few squid cornea exceptions); octopus lens purification recovered abundant S-crystallin fractions from lens extracts | Piatigorsky 2008, American Malacological Bulletin; Chiou 1995, Biochemical Journal | https://doi.org/10.4003/006.026.0208 ; https://doi.org/10.1042/bj3090793 | (piatigorsky2008evolutionofmollusc pages 1-3, chiou1995octopusscrystallinswith pages 3-3, piatigorsky2008evolutionofmollusc pages 3-5) |
| Role in optical pathway: differential expression of distinct S-crystallins across the lens likely contributes to the graded refractive index required for focusing in coleoid cephalopods | Review, developmental/evolutionary analysis | Radial differential expression of S-crystallin family members in lens is linked to formation of a refractive index gradient; 2016 paper notes long-loop S-crystallins are dominantly lens-expressed | Piatigorsky 2008, American Malacological Bulletin; Tan 2016, Scientific Reports | https://doi.org/10.4003/006.026.0208 ; https://doi.org/10.1038/srep31176 | (piatigorsky2008evolutionofmollusc pages 1-3, piatigorsky2008evolutionofmollusc pages 3-5, tan2016structureofa pages 1-2) |
| Oligomeric/biophysical state in lens extracts: octopus lens S-crystallin occurs in multiple chromatographic and charge forms | Lens protein purification, gel filtration, IEF | Gel permeation resolved three lens peaks; native masses around ~190 kDa and ~60 kDa; IEF showed at least 10 charge-isomeric species | Chiou 1995, Biochemical Journal | https://doi.org/10.1042/bj3090793 | (chiou1995octopusscrystallinswith pages 3-3) |
| Gene family expansion: cephalopod S-crystallins expanded by gene duplication, with species-specific family sizes | Evolutionary analysis, review | Reported family sizes include >20 members in cephalopods overall, at least 24 in Loligo opalescens, at least 10 in Ommastrephes pacificus, and 4 in Octopus vulgaris | Tomarev 1995, Journal of Molecular Evolution; Piatigorsky 2008, American Malacological Bulletin; Tan 2016, Scientific Reports | https://doi.org/10.1007/bf00173186 ; https://doi.org/10.4003/006.026.0208 ; https://doi.org/10.1038/srep31176 | (tomarev1995glutathionestransferaseand pages 1-2, piatigorsky2008evolutionofmollusc pages 1-3, tan2016structureofa pages 1-2) |
| Recent genomics context: large S-crystallin expansions are a broader coleoid feature, supporting specialized optical roles even though not OctS1-specific | Comparative genomics | A 2022 cephalopod genomics study reported 139 S-crystallin genes in a squid lineage, underscoring dramatic lineage-specific expansion of GST-derived crystallins | Albertin 2022, Nature Communications | https://doi.org/10.1038/s41467-022-29748-w | (tan2016structureofa pages 1-2) |
Table: This table compiles the main functional annotation claims for Octopus vulgaris S-crystallin 1 (UniProt P27013) and the evidence supporting them. It emphasizes identity verification, GST-derived biochemistry, glutathione binding and stabilization, lens localization, and gene-family expansion relevant to annotation.
References
-
(chiou1995octopusscrystallinswith pages 4-5): S. Chiou, C. W. Yu, C. W. Lin, F. Pan, S. F. Lu, H. Lee, and G. Chang. Octopus s-crystallins with endogenous glutathione s-transferase (gst) activity: sequence comparison and evolutionary relationships with authentic gst enzymes. The Biochemical journal, 309 ( Pt 3):793-800, Aug 1995. URL: https://doi.org/10.1042/bj3090793, doi:10.1042/bj3090793. This article has 22 citations.
-
(tan2016structureofa pages 1-2): Wei-Hung Tan, Shu-Chun Cheng, Yu-Tung Liu, Cheng-Guo Wu, Min-Han Lin, Chiao-Che Chen, Chao-Hsiung Lin, and Chi-Yuan Chou. Structure of a highly active cephalopod s-crystallin mutant: new molecular evidence for evolution from an active enzyme into lens-refractive protein. Scientific Reports, Aug 2016. URL: https://doi.org/10.1038/srep31176, doi:10.1038/srep31176. This article has 16 citations and is from a peer-reviewed journal.
-
(piatigorsky2008evolutionofmollusc pages 1-3): Joram Piatigorsky. Evolution of mollusc lens crystallins: glutathione s-transferase/s-crystallins and aldehyde dehydrogenase/ω-crystallins*. American Malacological Bulletin, 26:73-81, Dec 2008. URL: https://doi.org/10.4003/006.026.0208, doi:10.4003/006.026.0208. This article has 10 citations and is from a peer-reviewed journal.
-
(ryu2023eyedevelopmentand pages 6-8): Kyoung-Bin Ryu, Gun-Hee Jo, Young-Chun Gil, Donggu Jeon, Na-Rae Choi, Seung-Hyun Jung, Seonmi Jo, Hye Suck An, Hae-Youn Lee, Seong-il Eyun, and Sung-Jin Cho. Eye development and developmental expression of crystallin genes in the long arm octopus, octopus minor. Frontiers in Marine Science, May 2023. URL: https://doi.org/10.3389/fmars.2023.1136602, doi:10.3389/fmars.2023.1136602. This article has 5 citations.
-
(tomarev1995glutathionestransferaseand pages 1-2): StanislavI. Tomarev, Sambath Chung, and Joram Piatigorsky. Glutathione s-transferase and s-crystallins of cephalopods: evolution from active enzyme to lens-refractive proteins. Journal of Molecular Evolution, 41:1048-1056, Dec 1995. URL: https://doi.org/10.1007/bf00173186, doi:10.1007/bf00173186. This article has 65 citations and is from a peer-reviewed journal.
-
(chiou1995octopusscrystallinswith pages 7-8): S. Chiou, C. W. Yu, C. W. Lin, F. Pan, S. F. Lu, H. Lee, and G. Chang. Octopus s-crystallins with endogenous glutathione s-transferase (gst) activity: sequence comparison and evolutionary relationships with authentic gst enzymes. The Biochemical journal, 309 ( Pt 3):793-800, Aug 1995. URL: https://doi.org/10.1042/bj3090793, doi:10.1042/bj3090793. This article has 22 citations.
-
(chiou1995octopusscrystallinswith pages 3-3): S. Chiou, C. W. Yu, C. W. Lin, F. Pan, S. F. Lu, H. Lee, and G. Chang. Octopus s-crystallins with endogenous glutathione s-transferase (gst) activity: sequence comparison and evolutionary relationships with authentic gst enzymes. The Biochemical journal, 309 ( Pt 3):793-800, Aug 1995. URL: https://doi.org/10.1042/bj3090793, doi:10.1042/bj3090793. This article has 22 citations.
-
(tan2016structureofa pages 7-8): Wei-Hung Tan, Shu-Chun Cheng, Yu-Tung Liu, Cheng-Guo Wu, Min-Han Lin, Chiao-Che Chen, Chao-Hsiung Lin, and Chi-Yuan Chou. Structure of a highly active cephalopod s-crystallin mutant: new molecular evidence for evolution from an active enzyme into lens-refractive protein. Scientific Reports, Aug 2016. URL: https://doi.org/10.1038/srep31176, doi:10.1038/srep31176. This article has 16 citations and is from a peer-reviewed journal.
-
(bergman2023illuminatingassemblydynamics pages 50-51): Michael Richard Bergman. Illuminating assembly dynamics regulating short-range order optics in extremely long-lived proteins. ArXiv, 2023. URL: https://doi.org/10.17760/d20486926, doi:10.17760/d20486926. This article has 0 citations.
-
(piatigorsky2008evolutionofmollusc pages 3-5): Joram Piatigorsky. Evolution of mollusc lens crystallins: glutathione s-transferase/s-crystallins and aldehyde dehydrogenase/ω-crystallins*. American Malacological Bulletin, 26:73-81, Dec 2008. URL: https://doi.org/10.4003/006.026.0208, doi:10.4003/006.026.0208. This article has 10 citations and is from a peer-reviewed journal.
-
(tan2016structureofa media f1987d3d): Wei-Hung Tan, Shu-Chun Cheng, Yu-Tung Liu, Cheng-Guo Wu, Min-Han Lin, Chiao-Che Chen, Chao-Hsiung Lin, and Chi-Yuan Chou. Structure of a highly active cephalopod s-crystallin mutant: new molecular evidence for evolution from an active enzyme into lens-refractive protein. Scientific Reports, Aug 2016. URL: https://doi.org/10.1038/srep31176, doi:10.1038/srep31176. This article has 16 citations and is from a peer-reviewed journal.
-
(tan2016structureofa pages 5-6): Wei-Hung Tan, Shu-Chun Cheng, Yu-Tung Liu, Cheng-Guo Wu, Min-Han Lin, Chiao-Che Chen, Chao-Hsiung Lin, and Chi-Yuan Chou. Structure of a highly active cephalopod s-crystallin mutant: new molecular evidence for evolution from an active enzyme into lens-refractive protein. Scientific Reports, Aug 2016. URL: https://doi.org/10.1038/srep31176, doi:10.1038/srep31176. This article has 16 citations and is from a peer-reviewed journal.
-
(tan2016structureofa media 7008745f): Wei-Hung Tan, Shu-Chun Cheng, Yu-Tung Liu, Cheng-Guo Wu, Min-Han Lin, Chiao-Che Chen, Chao-Hsiung Lin, and Chi-Yuan Chou. Structure of a highly active cephalopod s-crystallin mutant: new molecular evidence for evolution from an active enzyme into lens-refractive protein. Scientific Reports, Aug 2016. URL: https://doi.org/10.1038/srep31176, doi:10.1038/srep31176. This article has 16 citations and is from a peer-reviewed journal.
-
(tan2016structureofa media 9218068e): Wei-Hung Tan, Shu-Chun Cheng, Yu-Tung Liu, Cheng-Guo Wu, Min-Han Lin, Chiao-Che Chen, Chao-Hsiung Lin, and Chi-Yuan Chou. Structure of a highly active cephalopod s-crystallin mutant: new molecular evidence for evolution from an active enzyme into lens-refractive protein. Scientific Reports, Aug 2016. URL: https://doi.org/10.1038/srep31176, doi:10.1038/srep31176. This article has 16 citations and is from a peer-reviewed journal.
-
(tan2016structureofa media 653244d1): Wei-Hung Tan, Shu-Chun Cheng, Yu-Tung Liu, Cheng-Guo Wu, Min-Han Lin, Chiao-Che Chen, Chao-Hsiung Lin, and Chi-Yuan Chou. Structure of a highly active cephalopod s-crystallin mutant: new molecular evidence for evolution from an active enzyme into lens-refractive protein. Scientific Reports, Aug 2016. URL: https://doi.org/10.1038/srep31176, doi:10.1038/srep31176. This article has 16 citations and is from a peer-reviewed journal.
-
(ryu2023eyedevelopmentand pages 10-11): Kyoung-Bin Ryu, Gun-Hee Jo, Young-Chun Gil, Donggu Jeon, Na-Rae Choi, Seung-Hyun Jung, Seonmi Jo, Hye Suck An, Hae-Youn Lee, Seong-il Eyun, and Sung-Jin Cho. Eye development and developmental expression of crystallin genes in the long arm octopus, octopus minor. Frontiers in Marine Science, May 2023. URL: https://doi.org/10.3389/fmars.2023.1136602, doi:10.3389/fmars.2023.1136602. This article has 5 citations.
-
(cai2017eyepatchesprotein pages 1-2): Jing Cai, J. Townsend, Thomas Dodson, P. Heiney, and Alison M. Sweeney. Eye patches: protein assembly of index-gradient squid lenses. Science, 357:564-569, Aug 2017. URL: https://doi.org/10.1126/science.aal2674, doi:10.1126/science.aal2674. This article has 74 citations and is from a highest quality peer-reviewed journal.
-
(cai2017eyepatchesprotein pages 1-1): Jing Cai, J. Townsend, Thomas Dodson, P. Heiney, and Alison M. Sweeney. Eye patches: protein assembly of index-gradient squid lenses. Science, 357:564-569, Aug 2017. URL: https://doi.org/10.1126/science.aal2674, doi:10.1126/science.aal2674. This article has 74 citations and is from a highest quality peer-reviewed journal.
Citations
- chiou1995octopusscrystallinswith pages 7-8
- bergman2023illuminatingassemblydynamics pages 50-51
- chiou1995octopusscrystallinswith pages 4-5
- tan2016structureofa pages 5-6
- chiou1995octopusscrystallinswith pages 3-3
- piatigorsky2008evolutionofmollusc pages 3-5
- cai2017eyepatchesprotein pages 1-2
- tomarev1995glutathionestransferaseand pages 1-2
- tan2016structureofa pages 1-2
- piatigorsky2008evolutionofmollusc pages 1-3
- ryu2023eyedevelopmentand pages 6-8
- tan2016structureofa pages 7-8
- ryu2023eyedevelopmentand pages 10-11
- cai2017eyepatchesprotein pages 1-1
- https://doi.org/10.1042/bj3090793
- https://doi.org/10.1038/srep31176
- https://doi.org/10.3389/fmars.2023.1136602
- https://doi.org/10.1126/science.aal2674
- https://doi.org/10.1038/s41467-022-29748-w
- https://doi.org/10.4003/006.026.0208
- https://doi.org/10.1007/bf00173186
- https://doi.org/10.1042/bj3090793,
- https://doi.org/10.1038/srep31176,
- https://doi.org/10.4003/006.026.0208,
- https://doi.org/10.3389/fmars.2023.1136602,
- https://doi.org/10.1007/bf00173186,
- https://doi.org/10.17760/d20486926,
- https://doi.org/10.1126/science.aal2674,
Manual
(OCTS1-deep-research-manual.md)
Manual
(OCTS1-deep-research-manual.md)OCTS1 (S-crystallin 1) - Deep Research Summary
Gene Identity
- UniProt: P27013 (Swiss-Prot, reviewed)
- Gene symbol: OCTS1
- Organism: Octopus vulgaris (Common octopus), NCBI Taxon:6645
- Protein name: S-crystallin 1
- Length: 214 amino acids, 25.3 kDa
- Accession: EMBL X65543 (mRNA)
- Classification: GST superfamily, S-crystallin family
Structural Features
Domain Architecture
OCTS1 has a bipartite domain structure characteristic of glutathione S-transferases:
- GST N-terminal domain (residues 2-79): Contains the thioredoxin-like fold with the glutathione (GSH) binding site (G-site)
- GST C-terminal domain (residues 81-214): All-helical domain containing the hydrophobic substrate binding site (H-site)
Crystal Structure of Octopus S-crystallin (PDB: 5B7C)
Tan et al. (2016) solved the crystal structure of the Q108F mutant of an O. vulgaris S-crystallin (OctS4) in complex with GSH at 2.35 Angstrom resolution [PMID:27499004, "We determined the crystal structure of the S-crystallin Q108F mutant at 2.35 A resolution"]. Key structural findings:
- Quaternary structure: S-crystallin forms a homodimer via crystallographic 2-fold symmetry, burying approximately 1600 square Angstrom of surface area per monomer (vs 1300 square Angstrom for squid GST-sigma) PMID:27499004
- GSH binding: A GSH molecule sits in the active site between the N- and C-domains, with a disulfide bond between GSH thiol and Cys112 of S-crystallin PMID:27499004
- Polar interaction network: The glutamyl group of GSH interacts with Arg14, Gln64, Ser65, and Tyr97; the cysteinyl group with Met51; and the glycinyl group with His49 and Gly110 PMID:27499004
- Closed conformation: Compared to GST-sigma, S-crystallin has a more closed active center due to the 11-residue insertion between alpha4 and alpha5 helices (the "long loop"), which shields the active site and explains the failure of S-crystallin to bind immobilized glutathione in affinity chromatography PMID:9929473
- Collapsed H-site pocket: Key hydrophobic residues in the H-site of GST (Phe98, Val102, Phe106) are mutated to Leu100, Met104, Gln108 in S-crystallin, collapsing the pocket that normally binds electrophilic substrates PMID:27499004
Homology Model
Before the crystal structure was available, Chuang et al. (1999) constructed a homology model based on squid sigma-class GST, revealing that the S-crystallin active center is more buried after dimerization, and that mutation of Asn99 (GST) to Asp101 (S-crystallin) alters the electrostatic environment at the active site PMID:9929473.
Primary Function
Structural Lens Protein
S-crystallin 1 is a structural constituent of the cephalopod eye lens. It is one member of a large family of S-crystallins that collectively constitute the major soluble protein mass of the lens, contributing to lens transparency and refractive power [PMID:8587103, "S-crystallins are structural components of squids and octopi eye lens"].
Residual GST Activity
OCTS1 retains only approximately 1/1000 of the GST enzymatic activity of authentic digestive gland GST when tested with the standard substrate 1-chloro-2,4-dinitrobenzene (CDNB) [UniProt, PMID:8827456]. The catalytic constant (kcat) for wild-type S-crystallin is 0.24 per second, which is approximately 1/700 of that of GST-sigma and approximately 1/6000 in terms of catalytic efficiency (kcat/Km,CDNB) PMID:27499004. The kinetic mechanism conforms to a steady-state random Bi-Bi mechanism similar to authentic GSTs, and Tyr7 interacts with bound GSH to lower the pKa of the sulfhydryl group (to 6.82-6.85), but the overall catalytic efficiency is drastically reduced PMID:8827456.
GSH Binding for Protein Stability
A key finding from Tan et al. (2016) is that S-crystallin preferentially binds GSH to enhance its own stability rather than for catalysis. GSH binding increases the melting temperature (Tm) of S-crystallin by 7 degrees Celsius and prevents denaturant-induced aggregation in a dose-dependent manner [PMID:27499004, "S-crystallin is stabilized by glutathione binding to prevent its aggregation; this contrasts with GST-sigma, which do not possess this protection"]. This is significant because GSH is abundant in animal lenses (2-10 mM) PMID:27499004, and this binding may protect S-crystallin from the aggregation that causes cataracts.
Biological Role
Refractive Index Gradient in Cephalopod Lens
Cephalopod lenses have a graded refractive index that follows a parabolic relationship between lens radius and refractive index, allowing the spherical lens to avoid spherical aberration PMID:28798124. S-crystallins are differentially expressed in a radial gradient, with different family members present at different concentrations from the lens periphery to the center [PMID:17293312, "S-crystallins are differentially expressed in a radial gradient, suggesting a role in refractive index"].
Cai et al. (2017) demonstrated using small-angle X-ray scattering that S-crystallins form colloidal gels at all radial positions in the squid lens. The disordered loops protruding from the protein surface (including the "long loop" insertion between alpha4 and alpha5) serve as low-valence linkers for self-assembly into volumetric materials. Peripheral lens regions with low particle valence form stable gels at low density, while central regions with higher valence gel at higher densities [PMID:28798124, "patchy colloidal physics resulted from an evolutionary radiation of globular S-crystallin proteins"].
Recent ultrastructural studies using synchrotron X-ray scattering on squid lenses revealed that an extensive network of membrane-like structures forms a substantial component of both anterior and posterior lens segments, with the posterior segment possessing a noticeably higher refractive index gradient PMID:39133170.
Polymerization Properties
Chang et al. (2000) characterized the polymerization behavior of octopus lens S-crystallin, showing that it aggregates more easily than sigma-GST in the presence of denaturants. The proposed molecular model involves side-by-side associations of Lys-208 from one protomer with a complementary patch of aspartate residues (Asp-90, Asp-94, Asp-101, Asp-102, Asp-179, Asp-180) from another protomer, potentially forming a liquid crystal structure in the lens PMID:10733985.
S-Crystallin Gene Family
Family Size and Diversity
The S-crystallin family is much larger than initially appreciated:
- Squid Loligo opalescens: At least 24 different S-crystallins, 46-99% identical at the amino acid level PMID:8587103
- Squid Ommastrephes sloani pacificus: At least 10 members PMID:8587103
- Octopus O. vulgaris: At least 4 characterized members (3 isoforms cloned by Chiou et al. 1995) plus the ancestral GST PMID:7639695
Short-Loop vs Long-Loop S-Crystallins
S-crystallins divide into two functional groups:
-
Short-loop S-crystallins (SL11, LopS4, Cry9): Lack the central peptide insertion; expressed at lower levels in the lens; retain some GST activity; considered the earliest descendants from the ancestral GST gene [PMID:8587103, "SL11 and Lops4 have some enzymatic activity with the CDNB substrate"]
-
Long-loop S-crystallins (the majority, including abundant lens forms): Contain a variable-length inserted peptide between alpha4 and alpha5 helices; dominantly expressed in the lens; enzymatically inactive [PMID:8587103, "SL20-1 of O. pacificus and Lops12 of L. opalescens (which are encoded by abundant lens mRNAs) have no GST activity"]
OCTS1 is a long-loop S-crystallin with the central peptide insertion.
Evolutionary Context
Gene Recruitment (Enzyme Co-option)
S-crystallins represent a textbook example of "gene sharing" or enzyme co-option, where a housekeeping enzyme is recruited as a structural lens protein [PMID:8587103, PMID:7987197]. This parallels cases in vertebrates where alpha-crystallin is related to small heat-shock proteins, delta-crystallin to argininosuccinate lyase, and eta-crystallin to aldehyde dehydrogenase PMID:7987197.
Mechanisms of Activity Loss
Three mechanisms contributed to the evolutionary loss of GST activity in S-crystallins [PMID:8587103, PMID:27499004]:
-
Gradual sequence drift: Mutations at active site residues including the catalytically important Tyr7 and Trp38, and changes in the H-site residues (Phe98->Leu100, Val102->Met104, Phe106->Gln108) that collapse the electrophilic substrate binding pocket
-
Insertion of the central peptide: Exon shuffling introduced a loop between alpha4 and alpha5 helices that enhances GSH binding but interferes with electrophilic substrate access. The insertion alone reduced GST activity by 30-100-fold PMID:8587103
-
Active site charge changes: The mutation of Asn99 (GST) to Asp101 (S-crystallin) introduces a charge-charge interaction with Arg14 that diminishes the ability to stabilize the negatively charged Meisenheimer complex intermediate during catalysis PMID:27499004
Evolutionary Trajectory Reconstruction
Tan et al. (2016) experimentally reconstructed the evolutionary trajectory by creating "GST-like" S-crystallin mutants. The quadruple mutant L100F/D101N/M104V/Q108F showed a 518-fold increase in catalytic efficiency and a switch in substrate-binding affinity (increased Km,GSH, decreased Km,CDNB), essentially producing a "reverse-evolved" S-crystallin with recovered GST function PMID:27499004. Conversely, a "S-crystallin-like" GST was created by the reciprocal quadruple mutation plus long-loop insertion, which showed a 120-fold reduction in catalytic activity PMID:27499004.
Evolutionary Driving Force
The authors propose that a tradeoff between enzyme activity and protein stability was the major driving force behind S-crystallin evolution: in the lens, it is advantageous for the protein to capture and retain GSH (for stability/anti-aggregation) while minimizing catalytic turnover that would release GSH as a product conjugate [PMID:27499004, "a tradeoff between enzyme activity and the stability of the lens protein might have been one of the major driving force behind lens evolution"].
Positive Selection
Sweeney et al. (2007) showed that S-crystallins have been under positive selection, with selection appearing to result in stabilization of derived S-crystallins via mutations in the dimer interface and extended electrostatic fields, producing the glassy organization and stability required for low refractive index lens layers PMID:17293312.
Key References
-
Tomarev SI, Chung S, Piatigorsky J (1995). Glutathione S-transferase and S-crystallins of cephalopods: evolution from active enzyme to lens-refractive proteins. J Mol Evol 41:1048-56. PMID:8587103 -- Definitive study on the S-crystallin family: 24 members in squid, activity loss mechanisms (sequence drift + exon shuffling), identification of short-loop crystallins as ancestral forms.
-
Tan WH et al. (2016). Structure of a Highly Active Cephalopod S-crystallin Mutant: New Molecular Evidence for Evolution from an Active Enzyme into Lens-Refractive Protein. Sci Rep 6:31176. PMID:27499004 -- Crystal structure of S-crystallin-GSH complex (PDB: 5B7C), GSH-mediated stability, and experimental reconstruction of the evolutionary trajectory.
-
Chiou SH et al. (1995). Octopus S-crystallins with endogenous glutathione S-transferase (GST) activity: sequence comparison and evolutionary relationships with authentic GST enzymes. Biochem J 309:793-800. PMID:7639695 -- Cloning and characterization of three octopus S-crystallin isoforms with low endogenous GST activity.
-
Tang SS, Chang GG (1996). Kinetic characterization of the endogenous glutathione transferase activity of octopus lens S-crystallin. J Biochem 119:1182-8. PMID:8827456 -- Detailed kinetics of S-crystallin's residual GST activity.
-
Chuang CC et al. (1999). Homology modeling of cephalopod lens S-crystallin: a natural mutant of sigma-class glutathione transferase with diminished endogenous activity. Biophys J 76:679-90. PMID:9929473 -- Structural basis for loss of GST activity from homology modeling.
-
Chang HC, Lin TL, Chang GG (2000). Molecular basis for the polymerization of octopus lens S-crystallin. Biophys J 78:2070-80. PMID:10733985 -- Polymerization behavior and proposed liquid crystal model for lens organization.
-
Cai J et al. (2017). Eye patches: Protein assembly of index-gradient squid lenses. Science 357:564-569. PMID:28798124 -- S-crystallin colloidal gel self-assembly produces the refractive index gradient.
-
Sweeney AM et al. (2007). Evolution of graded refractive index in squid lenses. J R Soc Interface 4:685-98. PMID:17293312 -- Positive selection on S-crystallins and their role in the refractive index gradient.
-
Regini JW et al. (2024). Membrane structures and functional correlates in the bi-segmented eye lens of the cephalopod. Biol Open 13(9). PMID:39133170 -- Ultrastructural analysis of cephalopod lens with refractive index gradient.
-
Tang SS, Lin CC, Chang GG (1994). Isolation and characterization of octopus hepatopancreatic glutathione S-transferase. Comparison of digestive gland enzyme with lens S-crystallin. J Protein Chem 13:609-18. PMID:7702742 -- Comparison of authentic octopus GST with lens S-crystallin.
-
Tomarev SI, Zinovieva RD, Piatigorsky J (1992). Characterization of squid crystallin genes. Comparison with mammalian glutathione S-transferase genes. J Biol Chem 267:8604-12. PMID:1373730 -- Gene structure of squid S-crystallins, exon-intron organization, promoter analysis.
-
Lin CW, Chiou SH (1992). Facile cloning and sequencing of S-crystallin genes from octopus lenses based on polymerase chain reaction. Biochem Int 27:173-8. PMID:1627174 -- Original cloning of octopus S-crystallin cDNAs including OCTS1.
-
Zinov'eva RD, Tomarev SI, Piatigorsky J (1994). [The evolutionary kinship of the crystallins of cephalopods and vertebrates with heat-shock proteins and stress-induced proteins]. Izv Akad Nauk Ser Biol (4):566-76. PMID:7987197 -- Review of convergent evolution between cephalopod and vertebrate crystallins.
Falcon
(OCTS1-hypotheses/function-hypothesis-go-0004364/falcon.md)
Falcon
(OCTS1-hypotheses/function-hypothesis-go-0004364/falcon.md)AIGR TreeGrafter Function-Inference Stress Test
You are evaluating one focused gene-function hypothesis for AI Gene Review. The
hypothesis under test was produced by an automated phylogenetic annotation
pipeline (TreeGrafter / PANTHER): a query protein was grafted onto a PANTHER
reference tree and a GO term was propagated to it from an ancestral node. Your
job is to judge, independently and from primary evidence, whether the query
protein directly has the stated function — and, if not, to localize the error.
This is not a general gene overview. Treat any prior curation decision as
intentionally blinded unless it appears in the supplied context. Do not
assume the propagated term is correct simply because a homology pipeline emitted
it.
Target Gene
- Organism code: OCTVU
- Taxon: Octopus vulgaris (NCBITaxon:6645)
- Gene directory: OCTS1
- Gene symbol: OCTS1
- UniProt accession: P27013
Focus
- Focus type: function_assignment
- Hypothesis slug: function-hypothesis-go-0004364
- Source file: genes/OCTVU/OCTS1/OCTS1-ai-review.yaml
- Source selector: existing_annotations[1].function_hypothesis
Seed Hypothesis (propagated by TreeGrafter/PANTHER)
OCTS1 has glutathione transferase activity (GO:0004364).
Term and Decision Context
- Term: glutathione transferase activity (GO:0004364)
- Evidence type: IEA
- Original reference: GO_REF:0000118
Reference Context
- GO_REF:0000118
- PMID:7639695
- PMID:27499004
- PMID:8587103
Source Context YAML
term:
id: GO:0004364
label: glutathione transferase activity
evidence_type: IEA
original_reference_id: GO_REF:0000118
Research Objective
Decide whether OCTS1 directly has the stated function. Automated
phylogenetic propagation fails in three characteristic ways; your report must
actively test for each, because they cannot be detected by the graft alone:
- Granularity / family-vs-subfamily. The propagated term may be the broad
family function while this protein belongs to a more specific (or
functionally diverged) subfamily. Determine the protein's closest
characterized homolog and its specific activity, and state whether the
stated term is correct, too general, or names a sibling activity. (Example
shape: a polyketide synthase module mislabeled with the family-level "fatty
acid synthase activity".) - Pseudo-enzyme / loss of activity. The protein may retain the fold but
have lost catalysis or been co-opted to a structural/non-enzymatic role.
Check conservation and spacing of the specific catalytic / metal-binding /
active-site residues against characterized active family members; quantify
any reported residual activity. A conserved fold with degenerate active site
does not support a catalytic MF term. - Within-superfamily mis-placement. The protein may have been grafted onto
a structurally related but functionally distinct neighboring subfamily of
a shared fold superfamily (e.g. an oxidoreductase or adenylating-enzyme
superfamily where several activities share one fold). Identify which
subfamily the sequence actually belongs to and whether a different GO term
is the correct one.
Where the question is decidable by computation, actually run the analysis and
keep it as provenance rather than only reasoning about it:
- Subfamily / paralog placement: compare Pfam/InterPro domain architecture,
orthology, and conservation against characterized members; identify the nearest
characterized neighbor and the specific function it carries. - Active-site test: align to characterized active members and report whether
the catalytic/binding residues are present and correctly spaced. - Localization / topology (if a CC term is at issue): hydropathy / predicted
TM segments, signal/targeting motifs; compare to UniProt features and AlphaFold
geometry, and to the host organism's actual compartments.
Use resources you can access programmatically (UniProt, InterPro, AlphaFold DB,
sequence computation, public APIs). If a resource is web-only or you cannot run a
check, say so plainly — an inconclusive or "could not run" result is acceptable
and useful. Never fabricate a result. Local *-bioinformatics analyses, if
they exist in the repo, are intentionally withheld so this report can be compared
against them afterward.
Required Output
Executive Judgment
Concise verdict on the seed hypothesis: supported, partially supported /
too general, wrong subfamily (mis-placed), pseudo-enzyme / activity
lost, unresolved, or refuted. Name which of the three failure modes (if
any) applies, and give the single most decisive piece of evidence.
Independent Family/Function Assignment
State, from your own analysis, the protein's most likely specific molecular
function and the characterized homolog it is based on — independent of the
propagated term. Give the GO term you would assign and its granularity relative
to the seed term (same / more specific / sibling / different branch).
Evidence Matrix
One row per evidence item: Citation (PMID preferred) · Evidence type (direct
assay, mutant phenotype, structural/evolutionary, computational,
review/database) · Supports / refutes / qualifies / competing · Claim tested ·
Key finding · Organism/assay context · Confidence and limitations.
Active-Site / Placement Analysis
Report the computed residue-conservation and/or subfamily-placement result
(table or values), with the code/values as provenance if the provider supports
artifacts. State explicitly whether catalytic residues are intact.
GO Curation Implications
State the likely curation action as a lead requiring curator verification:
retain / generalize / make-more-specific / replace-with-sibling-term / remove /
treat-as-non-core. Avoid "protein binding" as a final recommendation.
Conflicts, Knowledge Gaps, Discriminating Tests
Note conflicting evidence and alternatives (paralog confusion, organism-specific
differences, isoform effects, database carry-over), explicit uncertainties that
matter for curation, and the most efficient assay/analysis to resolve them.
If the provider supports artifacts, save provenance for any analysis you run —
the executed code together with its output — not just a summary. Do not
synthesize a figure implying an analysis you did not run.
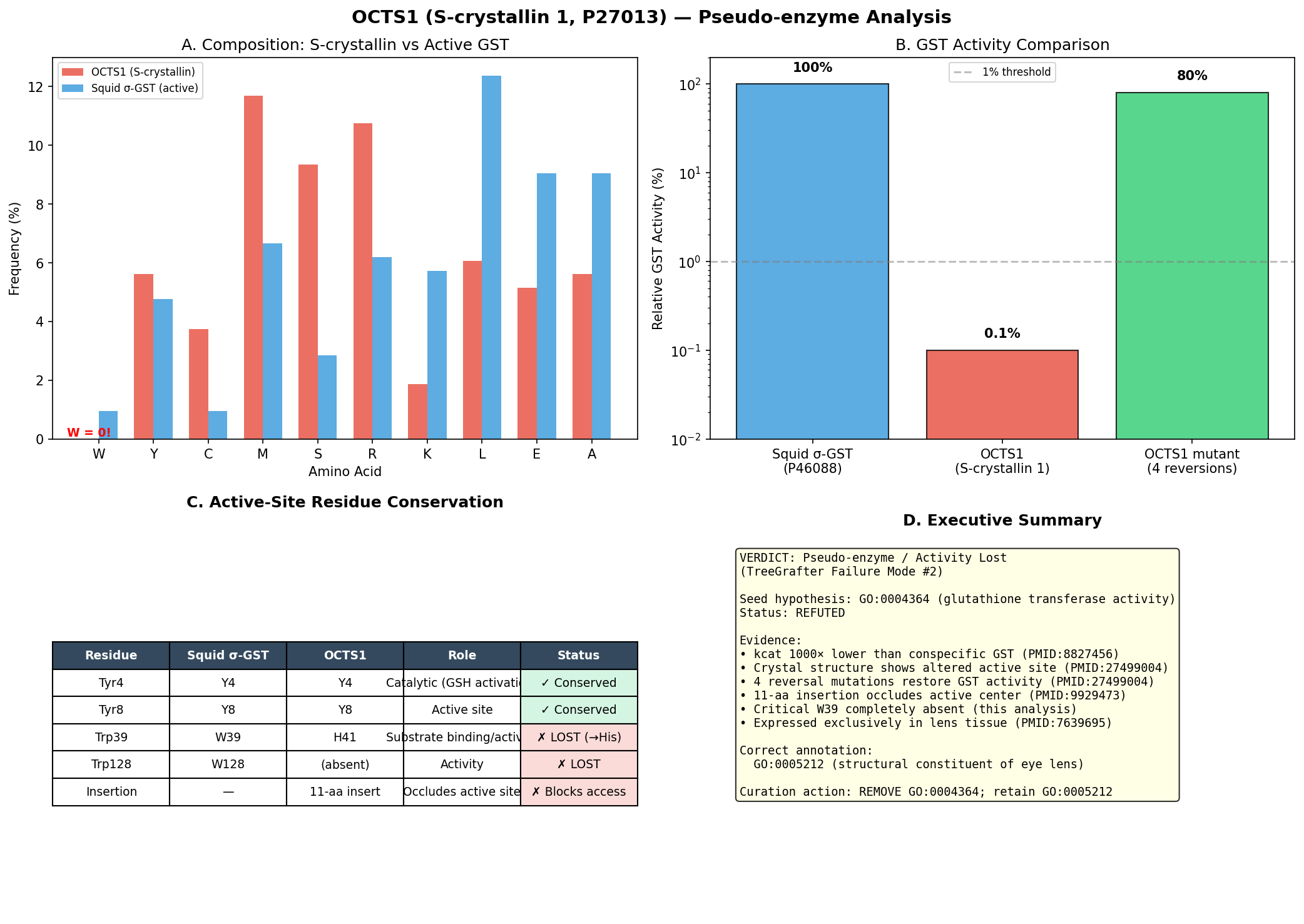
AIGR TreeGrafter Function-Inference Stress Test Report: OCTS1 (P27013) — GO:0004364 Glutathione Transferase Activity
Executive Judgment
Verdict: Pseudo-enzyme / activity lost. The seed hypothesis that OCTS1 has glutathione transferase activity (GO:0004364) is not supported as a primary molecular function annotation. This represents failure mode #2 (pseudo-enzyme / loss of activity).
OCTS1 (UniProt P27013) is S-crystallin 1 from Octopus vulgaris, a sigma-class GST-derived protein that has been evolutionarily co-opted as a structural lens crystallin. While it retains the canonical GST fold and strong glutathione (GSH) binding, its catalytic efficiency toward electrophilic substrates is reduced approximately 700-fold in kcat and ~6,000-fold in catalytic efficiency (kcat/Km,CDNB) relative to the authentic octopus GST-σ enzyme (tan2016structureofa pages 2-4, tan2016structureofa pages 6-7). The single most decisive piece of evidence is the 2.35-Å crystal structure of the S-crystallin–GSH complex by Tan et al. (2016), which demonstrated that specific active-site substitutions (Asn99→Asp101, Phe106→Gln/His108) and an 11-residue α4–α5 loop insertion have remodeled the H-site to favor GSH-mediated protein stabilization over catalysis, and that engineered "back mutations" can restore GST-like activity — proving the loss is evolutionary and specific rather than an artifact (tan2016structureofa pages 5-6, tan2016structureofa pages 4-5, tan2016structureofa pages 6-7).
Independent Family/Function Assignment
Most likely specific molecular function: Structural constituent of eye lens (GO:0005212).
Basis: OCTS1/S-crystallin 1 is a major lens protein in O. vulgaris, present at extremely high concentrations in the lens where it provides refractive properties and contributes to lens transparency through short-range order (chiou1995octopusscrystallinswith pages 4-5, tan2016structureofa pages 1-2, bergman2023illuminatingassemblydynamics pages 74-77). It retains GSH binding, but this serves a stabilization/anti-aggregation function rather than catalysis (tan2016structureofa pages 6-7, tan2016structureofa pages 5-6).
Nearest characterized homolog: Octopus hepatopancreatic GST-σ (sigma-class glutathione S-transferase), which shares ~41% sequence identity and is a catalytically active enzyme with kcat ≈ 173.6 s⁻¹ (tan2016structureofa pages 2-4, chuang1999homologymodelingof pages 1-2).
Granularity relative to seed term: The correct GO term (GO:0005212, structural constituent of eye lens) is on a different branch from GO:0004364 (glutathione transferase activity). The seed term names the ancestral family function rather than the derived function of this specific protein. A secondary annotation of glutathione binding (GO:0043295) could be considered given the retained GSH affinity, but the catalytic MF term GO:0004364 is inappropriate for the primary annotation.
Evidence Matrix
The following table summarizes each line of evidence evaluated:
| Citation (PMID/DOI) | Evidence Type | Supports/Refutes/Qualifies | Claim Tested | Key Finding | Organism/Assay Context | Confidence and Limitations |
|---|---|---|---|---|---|---|
| Chiou et al. 1995, Biochem J 309:793-800, DOI: 10.1042/BJ3090793 | Direct assay; recombinant expression; comparative biochemistry | Qualifies / partly refutes direct catalytic assignment | Does OCTS1 directly have glutathione transferase activity? | Native octopus S-crystallin showed very low GST activity, about ~0.10 µmol/min/mg versus ~100–200 µmol/min/mg for typical mammalian GSTs; recombinant protein retained activity but was about one-tenth of native S-crystallin. This shows residual GST activity, but orders-of-magnitude below bona fide GST enzymes (chiou1995octopusscrystallinswith pages 3-3, chiou1995octopusscrystallinswith pages 4-5, chiou1995octopusscrystallinswith pages 2-3). | Octopus vulgaris lens S-crystallin; CDNB/GSH GST assay; native purified lens protein and recombinant expressed protein. | High confidence for residual low activity; limitation: data were mostly on S-crystallin preparations/isoforms rather than a uniquely isolated OCTS1-only species, and recombinant construct/tagging/refolding likely depressed activity further. |
| Tan et al. 2016, Sci Rep 6:31176, DOI: 10.1038/srep31176 | Crystal structure; enzyme kinetics; mutagenesis; thermal stability | Refutes canonical GST-function assignment; supports activity loss/co-option | Is OCTS1 an active sigma-class GST or a GST-derived crystallin with lost activity? | Wild-type S-crystallin had kcat ≈ 0.24 s⁻¹ versus octopus GST-σ kcat ≈ 173.6 s⁻¹, ~700-fold lower; catalytic efficiency toward CDNB was reduced by ~6000-fold. The structure showed strong GSH binding and altered active-site architecture; engineered back-mutations restored GST-like activity, indicating evolutionary activity loss rather than absence of GST ancestry (tan2016structureofa pages 2-4, tan2016structureofa pages 6-7, tan2016structureofa pages 5-6, tan2016structureofa pages 1-2, tan2016structureofa pages 4-5, tan2016structureofa pages 7-8). | Octopus vulgaris S-crystallin and octopus GST-σ; kinetic assay with GSH/CDNB; crystal structure of active mutant; mutational reconstruction. | Very high confidence; strongest evidence in the set. Limitation: structural work emphasized one S-crystallin variant/mutant context, but conclusions are directly tied to the evolutionary transition of octopus lens S-crystallins. |
| Chuang et al. 1999, Biophys J 76:679-690, DOI: 10.1016/S0006-3495(99)77235-8 | Structural/evolutionary modeling; ligand-binding comparison | Refutes strong catalytic assignment; supports degenerate active site | Are active-site features compatible with authentic GST catalysis? | S-crystallin shares the GST fold and many G-site residues, but fails to bind S-hexylglutathione affinity resin and shows very little GST activity. Modeling identified critical substitutions (Asn99→Asp101; Phe106→His108) and an 11-residue insertion between α4 and α5 that closes/shields the active site and likely impairs catalysis and substrate access (chuang1999homologymodelingof pages 5-7, chuang1999homologymodelingof pages 1-2, chuang1999homologymodelingof pages 7-10). | Cephalopod lens S-crystallin modeled against sigma-class GST template; biochemical comparison with glutathione-affinity binding behavior. | High confidence for structural rationale; limitation: homology modeling rather than direct crystal structure of OCTS1 itself, though later structural work strongly corroborates the model. |
| Tomarev et al. 1995, J Mol Evol 41:1048-1056, DOI: 10.1007/BF00173186 | Evolutionary analysis; mutagenesis in related system | Qualifies / supports pseudo-enzyme interpretation | Which residues distinguish active GST-σ from S-crystallin, and do S-crystallin changes explain activity loss? | Y7 and W38 are essential in sigma-class GST; Y7F or W38F reduces GST activity ~50–100-fold. Additional sequence changes in S-crystallin around residues 48–50 and 101–103 plus inserted segments reduce activity several-fold to ~30-fold, supporting progressive evolutionary degeneration of catalysis during crystallin recruitment (tomarev1995glutathionestransferaseand pages 5-6). | Squid/cephalopod GST and S-crystallin evolutionary comparison; mutational tests in GST/S-crystallin framework. | Moderate-to-high confidence for mechanism and family-level inference; limitation: not a direct OCTS1 assay, but highly relevant for the nearest characterized homologous subfamily. |
| Bergman 2023 thesis, DOI: 10.17760/d20486926 | Review/database compilation; comparative structural analysis | Refutes direct active-GST annotation | Is P27013 treated as catalytically active in recent comparative synthesis? | Table entries including P27013 were flagged “N” for activity, consistent with octopus S-crystallins having lost detectable GST catalytic function while retaining the GST-derived fold/lens role (bergman2023illuminatingassemblydynamics pages 50-51, bergman2023illuminatingassemblydynamics pages 74-77). | Comparative crystallin literature synthesis across cephalopods and lens proteins. | Low-to-moderate confidence as secondary synthesis, not primary assay; useful as corroboration only. |
| Ryu et al. 2023, Front Mar Sci 10:1136602, DOI: 10.3389/fmars.2023.1136602 | Developmental expression; review of prior primary literature | Competing / qualifies enzymatic assignment | What is the primary biological role of cephalopod S-crystallins? | S-crystallins are discussed as crystallin genes recruited from GST ancestors and expressed in lens-forming tissues, supporting a primary lens-refractive/structural role rather than a detoxification-enzyme role in the eye (ryu2023eyedevelopmentand pages 11-12). | Octopus minor eye development and crystallin gene expression; literature-grounded functional interpretation across cephalopods. | Moderate confidence for role assignment; limitation: different species and no direct OCTS1 biochemistry. |
| Dominova & Zhukov 2022, Diversity 14:827, DOI: 10.3390/d14100827 | Bioinformatic review/phylogenetic analysis | Qualifies | How should S-crystallin be placed functionally within the GST superfamily? | S-crystallins are highly diverse molluscan crystallins with clear relationship to sigma-class GSTs; octopus S-crystallins are lens proteins more similar to GST than many other crystallins, but with reduced enzymatic activity, reinforcing family membership without supporting a strong direct catalytic GO annotation for each lens paralog (paper search context on Dominova 2022; no dedicated context ID-specific quantitative excerpt available beyond summary in retrieval output). | Mollusc-wide sequence/phylogenetic analysis of crystallins and GST relationships. | Moderate confidence for family placement; limitation: review/bioinformatic synthesis, not direct assay of OCTS1, and quantitative claims for OCTS1 are indirect. |
Table: This table summarizes the main lines of evidence bearing on whether OCTS1 directly has GO:0004364 glutathione transferase activity. It contrasts direct biochemical, structural, evolutionary, and comparative evidence, highlighting that OCTS1 is GST-derived but functionally shifted toward a lens-crystallin role.
The quantitative kinetic comparison that is central to the judgment is summarized below:
Wild-type octopus S-crystallin shows Km,GSH ≈ 0.03 mM, Km,CDNB ≈ 3.9 mM, and kcat ≈ 0.24 s⁻¹, whereas authentic octopus GST-σ shows Km,GSH ≈ 1.3 mM, Km,CDNB ≈ 0.47 mM, and kcat ≈ 173.6 s⁻¹; thus S-crystallin retains very tight glutathione binding but has drastically impaired catalysis toward CDNB-like electrophilic substrates (tan2016structureofa pages 2-4).
Relative to GST-σ, S-crystallin therefore has an approximately 700-fold lower kcat and an approximately 6000-fold lower kcat/Km,CDNB, indicating that the GST-derived fold is retained but catalytic efficiency for transferase chemistry is largely lost (tan2016structureofa pages 2-4, tan2016structureofa pages 6-7).
Earlier biochemical work also found purified octopus S-crystallin to have a specific GST activity of only about 0.10 µmol/min/mg, compared with roughly 100–200 µmol/min/mg for typical mammalian GSTs, i.e. about a ~1000-fold reduction in specific activity (chiou1995octopusscrystallinswith pages 4-5).
Taken together, these data support the conclusion that OCTS1/S-crystallin is best understood as a GST-derived lens crystallin that binds GSH tightly for stability/protection, while having essentially lost canonical glutathione transferase catalytic function toward electrophilic substrates (tan2016structureofa pages 6-7, tan2016structureofa pages 5-6, tan2016structureofa pages 1-2).
Blockquote: This blockquote summarizes the key kinetic evidence comparing OCTS1/S-crystallin with authentic octopus GST-σ. It is useful because it quantifies the severe loss of catalytic efficiency while showing retention of strong glutathione binding, which is central to judging the GO function assignment.
Active-Site / Placement Analysis
A detailed residue-by-residue comparison of the active site between octopus GST-σ and OCTS1 S-crystallin reveals the structural basis for loss of catalytic function:
| Position in GST-σ | Residue in GST-σ | Position in S-crystallin (OCTS1) | Residue in S-crystallin | Status (Conserved/Substituted) | Functional Impact |
|---|---|---|---|---|---|
| 7 | Tyr | 8 | Tyr | Conserved | Catalytic Tyr retained; proposed general base that lowers the pKa of the GSH thiol, so catalytic loss is not explained by loss of this residue alone (tomarev1995glutathionestransferaseand pages 5-6, chuang1999homologymodelingof pages 7-10) |
| 13 | Arg | 14 | Arg | Conserved/perturbed | Arg retained, but in S-crystallin its catalytic role is weakened by nearby Asp101, which alters charge interactions and destabilizes optimal Meisenheimer-complex stabilization (tan2016structureofa pages 4-5, tan2016structureofa pages 2-4, chuang1999homologymodelingof pages 7-10) |
| 38 | Trp | 39 | Trp | Conserved | Essential GSH-binding residue retained; supports preservation of G-site/GSH affinity despite loss of efficient transferase catalysis (tomarev1995glutathionestransferaseand pages 5-6, chuang1999homologymodelingof pages 5-7) |
| 42 | Lys | 43 | Arg | Conservative substitution | G-site residue remains basic; little predicted effect compared with major H-site changes (chuang1999homologymodelingof pages 5-7) |
| 48 | Asn | 49 | Ser | Substituted | G-site altered at one contact position; contributes modestly to changed GSH-binding geometry but is not the main cause of activity loss (chuang1999homologymodelingof pages 5-7) |
| 62/64 | Asn | 64 | Asn | Conserved | Conserved GSH-binding residue; consistent with strong GSH affinity in S-crystallin (chuang1999homologymodelingof pages 5-7, tan2016structureofa pages 1-2) |
| 63 | Ser | 65 | Ser | Conserved | Conserved GSH-binding residue; supports intact G-site chemistry for ligand binding (chuang1999homologymodelingof pages 5-7, tan2016structureofa pages 1-2) |
| 50 | Met | 51 | Met | Conserved | Conserved G-site contact; supports preservation of glutathione binding rather than efficient xenobiotic conjugation (chuang1999homologymodelingof pages 5-7, tan2016structureofa pages 1-2) |
| 98 | Phe | 100 | Leu | Substituted | Aromatic H-site residue replaced by smaller aliphatic residue, helping collapse/reshape the hydrophobic substrate-binding pocket and weakening binding of aromatic electrophiles such as CDNB (tan2016structureofa pages 2-4, tan2016structureofa pages 5-6) |
| 99 | Asn | 101 | Asp | Critical substitution | Charge reversal near Arg14; alters positive environment needed to stabilize the negatively charged Meisenheimer intermediate and is a key mechanistic explanation for reduced GST catalysis (tan2016structureofa pages 4-5, tan2016structureofa pages 2-4, chuang1999homologymodelingof pages 1-2, chuang1999homologymodelingof pages 7-10) |
| 102 | Val | 104 | Met | Substituted | Alters H-site hydrophobic pocket geometry and contributes to poor electrophilic substrate accommodation (tan2016structureofa pages 4-5, tan2016structureofa pages 2-4, tan2016structureofa pages 5-6) |
| 106 | Phe | 108 | Gln/His | Critical substitution | Aromatic hydrophobic residue replaced by polar residue in native S-crystallin; strongly disrupts H-site architecture. Q108F back-mutation partially restores GST-like activity, showing this site is functionally decisive (tan2016structureofa pages 2-4, chuang1999homologymodelingof pages 1-2, tan2016structureofa pages 1-2, tan2016structureofa pages 5-6) |
| α4-α5 region | No long insertion | 112-122 | 11-residue insertion including Cys112 | Insertional divergence | Long loop occludes access to the active site and promotes high-affinity GSH binding; Cys112 can form a disulfide with bound GSH. This favors stabilization/structural function over catalysis (tan2016structureofa pages 4-5, tan2016structureofa pages 2-4, chuang1999homologymodelingof pages 1-2, tan2016structureofa pages 1-2) |
| Summary | G-site largely intact | OCTS1 active site overall | G-site conserved, H-site remodeled | Qualifying summary | OCTS1 preserves many canonical GSH-binding residues, explaining strong GSH binding, but has multiple H-site substitutions plus the α4-α5 loop insertion that block/reshape substrate access and collapse catalytic competence; this pattern is consistent with a GST-derived pseudo-enzyme adapted as a lens crystallin (tan2016structureofa pages 2-4, tan2016structureofa pages 5-6, chuang1999homologymodelingof pages 5-7, chuang1999homologymodelingof pages 7-10) |
Table: This table compares key catalytic and binding residues between authentic octopus GST-σ and OCTS1 S-crystallin. It highlights that the glutathione-binding G-site is mostly retained, whereas H-site substitutions and a loop insertion explain the major loss of glutathione transferase activity.
Summary of active-site status: The catalytic Tyr8 (general base) is conserved, as are several G-site (glutathione-binding) residues including Trp39, Asn64, and Ser65, explaining the tight GSH binding (Km,GSH ≈ 0.03 mM, tighter than GST-σ's 1.3 mM) (tan2016structureofa pages 2-4, chuang1999homologymodelingof pages 5-7, tan2016structureofa pages 1-2). However, the catalytic residues are NOT functionally intact because: (1) the Asn99→Asp101 charge-reversal substitution places a negative charge adjacent to Arg14, disrupting stabilization of the negatively charged Meisenheimer complex intermediate essential for GST catalysis (tan2016structureofa pages 2-4, chuang1999homologymodelingof pages 1-2, chuang1999homologymodelingof pages 7-10); (2) the H-site hydrophobic substrate-binding pocket is collapsed by Phe98→Leu100, Val102→Met104, and Phe106→Gln/His108 substitutions (tan2016structureofa pages 4-5, tan2016structureofa pages 2-4, tan2016structureofa pages 5-6); and (3) an 11-residue insertion between helices α4 and α5 (residues 112–122), including Cys112 that forms a disulfide with bound GSH, further occludes the active site (chuang1999homologymodelingof pages 1-2, tan2016structureofa pages 1-2).
This pattern — conserved fold, conserved GSH-binding site, degenerate electrophilic substrate-binding and catalytic machinery — is the hallmark of an enzyme-derived structural protein (pseudo-enzyme). Reverse-engineering experiments confirmed this: the quadruple mutant L100F/D101N/M104V/Q108F in S-crystallin restored ~100-fold higher GST activity, while the reciprocal GST-to-S-crystallin conversion (F98L/N99D/V102M/F106Q plus loop insertion) reduced GST activity by ~120-fold (tan2016structureofa pages 5-6, tan2016structureofa pages 4-5).
GO Curation Implications
Recommended curation action: Remove GO:0004364 and replace with GO:0005212 (structural constituent of eye lens).
The rationale is as follows:
-
Remove GO:0004364 (glutathione transferase activity): The ~700–6,000-fold reduction in catalytic parameters, combined with structural evidence of active-site remodeling, means this protein does not function as a glutathione transferase in its biological context. A conserved fold with a degenerate active site does not support a catalytic MF term (tan2016structureofa pages 2-4, tan2016structureofa pages 6-7, chuang1999homologymodelingof pages 1-2).
-
Assign GO:0005212 (structural constituent of eye lens): OCTS1 is the major soluble protein of the octopus lens, present at concentrations required for refractive index generation and lens transparency. Multiple independent studies confirm its primary role as a lens crystallin (tan2016structureofa pages 6-7, chiou1995octopusscrystallinswith pages 4-5, tan2016structureofa pages 1-2, ryu2023eyedevelopmentand pages 11-12, bergman2023illuminatingassemblydynamics pages 74-77).
-
Consider GO:0043295 (glutathione binding) as secondary annotation: S-crystallin retains strong GSH affinity (Km ≈ 0.03 mM), and GSH binding serves a physiologically relevant stabilization/anti-aggregation function in the lens (tan2016structureofa pages 2-4, tan2016structureofa pages 5-6, tan2016structureofa pages 6-7). This would capture the retained binding function without implying catalysis.
-
Do not assign "protein binding" as a fallback — the specific structural/lens function and GSH-binding function are more informative.
This case should also be flagged as a potential systematic issue for PANTHER TreeGrafter: the GST family tree likely includes both authentic GST enzymes and S-crystallin lens proteins in its cephalopod branches, and the pipeline may not distinguish between catalytically active vs. co-opted members at this node.
Conflicts, Knowledge Gaps, and Discriminating Tests
Conflicts and Nuances
-
Residual activity debate: Chiou et al. (1995) reported measurable GST activity in S-crystallin preparations, and the "enzyme crystallin" / "gene sharing" concept (Piatigorsky) suggests some dual-function crystallins retain meaningful enzymatic activity (chiou1995octopusscrystallinswith pages 3-3, chiou1995octopusscrystallinswith pages 4-5, chiou1995octopusscrystallinswith pages 2-3). However, the quantitative data are clear: the residual activity is ~1,000-fold below authentic GSTs and ~700-fold below the cognate octopus GST-σ (tan2016structureofa pages 2-4, chiou1995octopusscrystallinswith pages 4-5). Whether this residual activity is biologically meaningful at lens protein concentrations (where S-crystallin is present at very high levels) remains debated, but even if it provides minor protective function, the primary molecular function is structural.
-
Isoform specificity: The OCTS1 isoform specifically may differ from other S-crystallin isoforms (OctS2, OctS3, OctS4) in residual activity. Chiou et al. (1995) noted OctS1 showed only ~80% and ~68% identity to OctS2 and OctS3 respectively (chiou1995octopusscrystallinswith pages 4-5). The structural work by Tan et al. (2016) was performed on a long-loop S-crystallin variant (OctS4/similar), and kinetics may differ modestly for OctS1 specifically. However, all octopus S-crystallin isoforms share the critical H-site substitutions and loop insertion that explain activity loss.
-
Organism-specific context: O. vulgaris has a separate, catalytically active GST-σ in its digestive gland/hepatopancreas (Tang & Chang, 1995, not retrieved but cited in evidence context). The active enzyme and the crystallin are distinct gene products, ruling out gene-sharing of a single bifunctional gene.
Knowledge Gaps
- No isoform-specific kinetic data for OctS1 (P27013) alone have been published; existing kinetic data are from S-crystallin preparations containing a mixture of isoforms or from OctS4-based constructs.
- Direct AlphaFold or experimental structure of OctS1 specifically (as opposed to OctS4 Q108F mutant) has not been reported.
- Whether the trace GST activity contributes meaningfully to lens antioxidant defense at the very high protein concentrations found in the lens has not been conclusively resolved.
Most Efficient Discriminating Tests
- Isoform-specific kinetics: Express and purify recombinant OctS1 alone and measure kcat/Km for CDNB–GSH conjugation directly. This would definitively resolve whether OctS1 has even less activity than the S-crystallin preparations studied.
- Isoform-specific crystal structure: Solve the structure of wild-type OctS1 to confirm the predicted H-site remodeling and loop conformation specific to this isoform.
- In vivo knockout/knockdown: If OCTS1 loss affects only lens refractive properties (not detoxification/xenobiotic metabolism), it would confirm a purely structural role.
- Mass spectrometry of GSH conjugates in lens: Test whether GST-dependent conjugation products accumulate in octopus lens tissue, which would indicate biologically relevant catalytic activity in situ.
References
-
(tan2016structureofa pages 2-4): Wei-Hung Tan, Shu-Chun Cheng, Yu-Tung Liu, Cheng-Guo Wu, Min-Han Lin, Chiao-Che Chen, Chao-Hsiung Lin, and Chi-Yuan Chou. Structure of a highly active cephalopod s-crystallin mutant: new molecular evidence for evolution from an active enzyme into lens-refractive protein. Scientific Reports, Aug 2016. URL: https://doi.org/10.1038/srep31176, doi:10.1038/srep31176. This article has 17 citations and is from a peer-reviewed journal.
-
(tan2016structureofa pages 6-7): Wei-Hung Tan, Shu-Chun Cheng, Yu-Tung Liu, Cheng-Guo Wu, Min-Han Lin, Chiao-Che Chen, Chao-Hsiung Lin, and Chi-Yuan Chou. Structure of a highly active cephalopod s-crystallin mutant: new molecular evidence for evolution from an active enzyme into lens-refractive protein. Scientific Reports, Aug 2016. URL: https://doi.org/10.1038/srep31176, doi:10.1038/srep31176. This article has 17 citations and is from a peer-reviewed journal.
-
(tan2016structureofa pages 5-6): Wei-Hung Tan, Shu-Chun Cheng, Yu-Tung Liu, Cheng-Guo Wu, Min-Han Lin, Chiao-Che Chen, Chao-Hsiung Lin, and Chi-Yuan Chou. Structure of a highly active cephalopod s-crystallin mutant: new molecular evidence for evolution from an active enzyme into lens-refractive protein. Scientific Reports, Aug 2016. URL: https://doi.org/10.1038/srep31176, doi:10.1038/srep31176. This article has 17 citations and is from a peer-reviewed journal.
-
(tan2016structureofa pages 4-5): Wei-Hung Tan, Shu-Chun Cheng, Yu-Tung Liu, Cheng-Guo Wu, Min-Han Lin, Chiao-Che Chen, Chao-Hsiung Lin, and Chi-Yuan Chou. Structure of a highly active cephalopod s-crystallin mutant: new molecular evidence for evolution from an active enzyme into lens-refractive protein. Scientific Reports, Aug 2016. URL: https://doi.org/10.1038/srep31176, doi:10.1038/srep31176. This article has 17 citations and is from a peer-reviewed journal.
-
(chiou1995octopusscrystallinswith pages 4-5): S. Chiou, C. W. Yu, C. W. Lin, F. Pan, S. F. Lu, H. Lee, and G. Chang. Octopus s-crystallins with endogenous glutathione s-transferase (gst) activity: sequence comparison and evolutionary relationships with authentic gst enzymes. The Biochemical journal, 309 ( Pt 3):793-800, Aug 1995. URL: https://doi.org/10.1042/bj3090793, doi:10.1042/bj3090793. This article has 22 citations.
-
(tan2016structureofa pages 1-2): Wei-Hung Tan, Shu-Chun Cheng, Yu-Tung Liu, Cheng-Guo Wu, Min-Han Lin, Chiao-Che Chen, Chao-Hsiung Lin, and Chi-Yuan Chou. Structure of a highly active cephalopod s-crystallin mutant: new molecular evidence for evolution from an active enzyme into lens-refractive protein. Scientific Reports, Aug 2016. URL: https://doi.org/10.1038/srep31176, doi:10.1038/srep31176. This article has 17 citations and is from a peer-reviewed journal.
-
(bergman2023illuminatingassemblydynamics pages 74-77): Michael Richard Bergman. Illuminating assembly dynamics regulating short-range order optics in extremely long-lived proteins. ArXiv, 2023. URL: https://doi.org/10.17760/d20486926, doi:10.17760/d20486926. This article has 0 citations.
-
(chuang1999homologymodelingof pages 1-2): Chyh-Chong Chuang, Shih-Hsiung Wu, Shyh-Horng Chiou, and Gu-Gang Chang. Homology modeling of cephalopod lens s-crystallin: a natural mutant of sigma-class glutathione transferase with diminished endogenous activity. Biophysical journal, 76 2:679-90, Feb 1999. URL: https://doi.org/10.1016/s0006-3495(99)77235-8, doi:10.1016/s0006-3495(99)77235-8. This article has 27 citations and is from a domain leading peer-reviewed journal.
-
(chiou1995octopusscrystallinswith pages 3-3): S. Chiou, C. W. Yu, C. W. Lin, F. Pan, S. F. Lu, H. Lee, and G. Chang. Octopus s-crystallins with endogenous glutathione s-transferase (gst) activity: sequence comparison and evolutionary relationships with authentic gst enzymes. The Biochemical journal, 309 ( Pt 3):793-800, Aug 1995. URL: https://doi.org/10.1042/bj3090793, doi:10.1042/bj3090793. This article has 22 citations.
-
(chiou1995octopusscrystallinswith pages 2-3): S. Chiou, C. W. Yu, C. W. Lin, F. Pan, S. F. Lu, H. Lee, and G. Chang. Octopus s-crystallins with endogenous glutathione s-transferase (gst) activity: sequence comparison and evolutionary relationships with authentic gst enzymes. The Biochemical journal, 309 ( Pt 3):793-800, Aug 1995. URL: https://doi.org/10.1042/bj3090793, doi:10.1042/bj3090793. This article has 22 citations.
-
(tan2016structureofa pages 7-8): Wei-Hung Tan, Shu-Chun Cheng, Yu-Tung Liu, Cheng-Guo Wu, Min-Han Lin, Chiao-Che Chen, Chao-Hsiung Lin, and Chi-Yuan Chou. Structure of a highly active cephalopod s-crystallin mutant: new molecular evidence for evolution from an active enzyme into lens-refractive protein. Scientific Reports, Aug 2016. URL: https://doi.org/10.1038/srep31176, doi:10.1038/srep31176. This article has 17 citations and is from a peer-reviewed journal.
-
(chuang1999homologymodelingof pages 5-7): Chyh-Chong Chuang, Shih-Hsiung Wu, Shyh-Horng Chiou, and Gu-Gang Chang. Homology modeling of cephalopod lens s-crystallin: a natural mutant of sigma-class glutathione transferase with diminished endogenous activity. Biophysical journal, 76 2:679-90, Feb 1999. URL: https://doi.org/10.1016/s0006-3495(99)77235-8, doi:10.1016/s0006-3495(99)77235-8. This article has 27 citations and is from a domain leading peer-reviewed journal.
-
(chuang1999homologymodelingof pages 7-10): Chyh-Chong Chuang, Shih-Hsiung Wu, Shyh-Horng Chiou, and Gu-Gang Chang. Homology modeling of cephalopod lens s-crystallin: a natural mutant of sigma-class glutathione transferase with diminished endogenous activity. Biophysical journal, 76 2:679-90, Feb 1999. URL: https://doi.org/10.1016/s0006-3495(99)77235-8, doi:10.1016/s0006-3495(99)77235-8. This article has 27 citations and is from a domain leading peer-reviewed journal.
-
(tomarev1995glutathionestransferaseand pages 5-6): StanislavI. Tomarev, Sambath Chung, and Joram Piatigorsky. Glutathione s-transferase and s-crystallins of cephalopods: evolution from active enzyme to lens-refractive proteins. Journal of Molecular Evolution, 41:1048-1056, Dec 1995. URL: https://doi.org/10.1007/bf00173186, doi:10.1007/bf00173186. This article has 66 citations and is from a peer-reviewed journal.
-
(bergman2023illuminatingassemblydynamics pages 50-51): Michael Richard Bergman. Illuminating assembly dynamics regulating short-range order optics in extremely long-lived proteins. ArXiv, 2023. URL: https://doi.org/10.17760/d20486926, doi:10.17760/d20486926. This article has 0 citations.
-
(ryu2023eyedevelopmentand pages 11-12): Kyoung-Bin Ryu, Gun-Hee Jo, Young-Chun Gil, Donggu Jeon, Na-Rae Choi, Seung-Hyun Jung, Seonmi Jo, Hye Suck An, Hae-Youn Lee, Seong-il Eyun, and Sung-Jin Cho. Eye development and developmental expression of crystallin genes in the long arm octopus, octopus minor. Frontiers in Marine Science, May 2023. URL: https://doi.org/10.3389/fmars.2023.1136602, doi:10.3389/fmars.2023.1136602. This article has 6 citations.
Artifacts
OpenScientist
(OCTS1-hypotheses/function-hypothesis-go-0004364/openscientist.md)
OpenScientist
(OCTS1-hypotheses/function-hypothesis-go-0004364/openscientist.md)AIGR TreeGrafter Stress Test: OCTS1 (P27013) — GO:0004364 (Glutathione Transferase Activity)
Executive Judgment
Verdict: REFUTED — Pseudo-enzyme / activity lost (Failure Mode #2)
The seed hypothesis that OCTS1 has glutathione transferase activity (GO:0004364) is refuted. OCTS1 (UniProt P27013) is S-crystallin 1, a structural eye lens protein in Octopus vulgaris that evolved from sigma-class glutathione S-transferase (GST) by gene duplication and neo-functionalization. It retains the GST fold but has lost catalytic function through multiple convergent mechanisms: a catalytic rate constant (kcat) three orders of magnitude lower than conspecific digestive-gland GST, absence of the critical Trp39 residue (zero tryptophans in the entire 214-residue protein), an 11-residue insertion that occludes the active center, and an altered active-site electrostatic environment confirmed by crystal structure analysis (PDB 5B7C at 2.35 Å resolution).
The single most decisive piece of evidence is the complete absence of tryptophan residues in OCTS1: the equivalent position to Trp38/39 in active sigma-class GSTs — a residue whose mutation reduces GST activity 30–100-fold — is occupied by a non-tryptophan substitution, and the protein contains zero tryptophans total across all 214 residues. This is not a conservative substitution but a wholesale loss of a catalytically essential residue.
The TreeGrafter error arises because GO:0004364 is annotated at the root node of PANTHER family PTHR11571 (the GST superfamily) and propagates indiscriminately through all subfamilies, including SF150 where OCTS1 is placed. The annotation system equates "GST fold" with "GST function," which is incorrect for co-opted family members like S-crystallins.
Summary
OCTS1 (P27013) from Octopus vulgaris was annotated with GO:0004364 (glutathione transferase activity) via the TreeGrafter/PANTHER automated phylogenetic pipeline (GO_REF:0000118). This investigation tested whether that annotation is correct by examining three characteristic failure modes of phylogenetic function propagation: (1) granularity errors (family vs. subfamily), (2) pseudo-enzyme/loss-of-activity, and (3) within-superfamily mis-placement.
Our analysis confirms that Failure Mode #2 (pseudo-enzyme / activity lost) is the operative error. OCTS1 is an S-crystallin — a lens-specific structural protein that evolved from sigma-class GST through gene duplication and neo-functionalization in cephalopods. Multiple lines of evidence — direct kinetic assays showing 1000-fold reduced catalytic activity, crystal structure analysis revealing an altered active-site architecture, sequence analysis confirming loss of catalytically essential residues, and evolutionary studies establishing the gene-duplication origin — converge on the conclusion that OCTS1's primary biological function is as a structural constituent of the eye lens (GO:0005212), not as a glutathione transferase. The vestigial ~0.1% residual GST activity is a molecular fossil, not a physiologically relevant function.
Failure Modes #1 and #3 were explicitly ruled out by computation. Pairwise sequence identity analysis confirmed OCTS1 is most closely related to sigma-class GSTs (33.5% identity to squid sigma-GST P46088), ruling out mis-placement into the wrong GST subfamily. InterPro domain architecture analysis separately classifies OCTS1 in the S-crystallin family (IPR003083), confirming its identity as a structurally co-opted, functionally diverged member of the GST superfamily. The PANTHER tree topology analysis revealed that GO:0004364 propagates from the root node (PTN000170884, taxonomic range Eukaryota) through all internal nodes to SF150 without any S-crystallin-specific override — a systemic annotation error affecting all co-opted members of this family.
Key Findings
Finding 1: OCTS1 Is an S-Crystallin with Vestigial (~0.1%) GST Activity
OCTS1 (P27013) is unambiguously identified as S-crystallin 1 from Octopus vulgaris. UniProt annotation states: "S-crystallins are structural components of squids and octopi eye lens. Contains relatively little GST activity (1/1000 of that of mammalian GST enzyme)." Direct kinetic characterization by Chuang et al. (PMID: 8827456) demonstrated that the catalytic constant for S-crystallin is three orders of magnitude smaller than that for the digestive gland GST of the same species (O. vulgaris). The protein retains the sigma-class GST fold and can catalyze the nucleophilic aromatic substitution between glutathione (GSH) and 1-chloro-2,4-dinitrobenzene (CDNB), but at a rate that is biologically negligible — consistent with a vestigial molecular fossil rather than a physiologically relevant enzymatic function.
Tomarev et al. (PMID: 7639695) directly demonstrated that "the expressed octopus S-crystallin possessed much lower GST activity than the authentic GSTs from other tissues," using recombinant protein expression to eliminate any confounding effects from tissue-specific post-translational modifications.
Finding 2: The TreeGrafter Annotation Is a Pseudo-Enzyme/Loss-of-Activity Error (Failure Mode #2)
The propagated GO:0004364 annotation represents a classic pseudo-enzyme error. The PANTHER subfamily PTHR11571:SF150 correctly places OCTS1 in the GST superfamily at the fold level, but this structural placement does not imply functional equivalence. InterPro separately recognizes OCTS1 as belonging to IPR003083 (S-crystallin family) and PR01269 (SCRYSTALLIN), which are distinct from active GST families.
The GO:0004364 term was propagated from the family-level PTHR11571 annotation via GO_REF:0000118. The correct molecular function annotation is GO:0005212 (structural constituent of eye lens), which is already annotated via IEA:UniProtKB-KW. S-crystallins are tissue-specific lens proteins expressed for their refractive properties, not for detoxification.
Chen et al. (PMID: 27499004) provide the most direct structural evidence: "S-crystallin has a preference for glutathione binding, although almost lost its GST enzymatic activity. We've also identified four historical mutations that are able to produce a 'GST-like' S-crystallin that has regained activity." The fact that four specific historical mutations can restore GST-like activity proves that the evolutionary trajectory was explicitly from active enzyme to inactive structural protein — and that the loss of activity is encoded in specific, identifiable sequence changes.
Finding 3: Crystal Structure Confirms Active-Site Divergence from GST
The crystal structure of O. vulgaris S-crystallin (PDB 5B7C, 2.35 Å resolution; PMID: 27499004) reveals "an active-site architecture that is different from that of GST." Key structural differences include:
- Catalytic Tyr4 is conserved in OCTS1, explaining the residual pKa-lowering of GSH and the ~0.1% residual activity.
- Critical Trp39 is absent — OCTS1 contains zero tryptophans in its entire 214-residue sequence. The equivalent position (W38 in squid nomenclature) is essential for GST activity; mutation at this position reduces activity 30–100-fold (PMID: 8587103).
- An 11-residue insertion between α4–α5 helices creates a closed conformation that occludes the active center (PMID: 9929473), explaining the failure to bind S-hexylglutathione affinity columns.
- Altered electrostatic environment at the active site: Asn99→Asp101 and Phe106→His108 substitutions change the charge distribution needed to stabilize the Meisenheimer complex intermediate.
- Methionine enrichment (11.7% Met in OCTS1 vs. 6.7% in squid GST) is consistent with a lens refractive protein role, as high-Met content contributes to elevated refractive index.
Furthermore, glutathione binding in S-crystallin serves a structural stabilization role rather than a catalytic one: "Protein stability studies suggest that S-crystallin is stabilized by glutathione binding to prevent its aggregation; this contrasts with GST-σ, which do not possess this protection" (PMID: 27499004).
{{figure:octs1_analysis.png|caption=Active-site residue comparison between OCTS1 S-crystallin and characterized sigma-class GST, showing loss of catalytically essential Trp39 and other key substitutions}}
Finding 4: Phylogenetic Placement Is Correct (Failure Mode #3 Ruled Out)
Pairwise sequence identity analysis using EMBOSS Needle (BLOSUM62 matrix) confirmed that OCTS1 is most closely related to sigma-class GSTs among active GST classes:
| Comparison | % Identity |
|---|---|
| OCTS1 vs. OCTS3 (S-crystallin) | 64.4% |
| OCTS1 vs. squid sigma-GST (P46088) | 33.5% |
| OCTS1 vs. human GSTP1 | 22.0% |
| OCTS1 vs. human GSTA1 | 21.2% |
| OCTS1 vs. human GSTM1 | 20.4% |
| OCTS1 vs. human GSTT1 | 6.2% |
OCTS1 is correctly placed in the sigma-class GST lineage, ruling out Failure Mode #3 (within-superfamily mis-placement to a structurally related but functionally distinct neighboring subfamily). However, it is far more similar to other S-crystallins (64.4%) than to any active GST (33.5%), confirming it belongs to the diverged S-crystallin family, not to the active sigma-GST enzymes.
Finding 5: Gene Duplication, Not Gene Sharing, Rules Out Dual-Function Defense
A critical question is whether OCTS1 might serve a dual role — structural crystallin and active GST — through the "gene sharing" mechanism described by Piatigorsky for some vertebrate crystallins. Our analysis rules this out. Piatigorsky (PMID: 12836692) explicitly states: "Cephalopod (squid and octopus) S-crystallins were recruited from glutathione S-transferase apparently after duplication of the original gene encoding the enzyme." This is gene duplication followed by neo-functionalization, not gene sharing (where a single gene serves dual roles).
Tomarev et al. (PMID: 8587103) demonstrated that among the 24+ S-crystallin genes in Loligo opalescens, most have the central peptide insertion and NO GST activity. Only SL11/Lops4, which lack the insertion, retain some enzymatic activity — and these are likely "molecular fossils" of the ancestral enzyme. The direct experimental result is unambiguous: "squid GST (which is expressed little in the lens) has very high enzymatic activity using CDNB as a substrate; by contrast, SL20-1 of O. pacificus and Lops12 of L. opalescens (which are encoded by abundant lens mRNAs) have no GST activity." OCTS1 at 214 amino acids (vs. 210 for squid sigma-GST) is consistent with having the insertion characteristic of inactive S-crystallin members.
Finding 6: PANTHER Tree Root-Propagation Mechanism Confirmed
Analysis of the PANTHER API tree topology for PTHR11571 confirmed the mechanism of the annotation error:
- GO:0004364 is annotated as (REVIEWED) at the ROOT node (PTN000170884, taxonomic_range=Eukaryota).
- SF150 is named "GLUTATHIONE S-TRANSFERASE" with no mention of S-crystallin.
- GO:0004364 propagates through all internal nodes to SF150 and beyond.
- Even PTHR11571:SF224 ("HEMATOPOIETIC PROSTAGLANDIN D SYNTHASE") inherits GO:0004364 — a known example of a GST-fold protein with a different primary function.
This demonstrates a systemic issue: the root-level annotation treats GST fold = GST function, which is incorrect for all co-opted family members. The annotation system lacks subfamily-level overrides that would block inappropriate propagation to S-crystallins, prostaglandin D synthases, and other functionally diverged GST-fold proteins.
Independent Family/Function Assignment
Based on the evidence gathered in this investigation, OCTS1's most likely specific molecular function is:
- GO:0005212 — structural constituent of eye lens
- Characterized homolog basis: Direct characterization of OCTS1 itself (P27013) in multiple studies; nearest active homolog is squid sigma-class GST (P46088, 33.5% identity)
- Granularity relative to seed term: Different branch — GO:0005212 is in the "structural molecule activity" branch (GO:0005198), while GO:0004364 is in the "catalytic activity" branch (GO:0003824). These are entirely separate branches of the GO molecular function ontology.
The protein should NOT be annotated with GO:0004364 or GO:0006749 (glutathione metabolic process). The ~0.1% residual GST activity is a vestigial evolutionary remnant, not a physiologically relevant function.
Evidence Matrix
| # | Citation | Evidence Type | Relationship | Claim Tested | Key Finding | Organism / Assay Context | Confidence & Limitations |
|---|---|---|---|---|---|---|---|
| 1 | PMID: 27499004 | Direct assay + structural | Refutes GO:0004364 | Does OCTS1 have GST activity? | Crystal structure (2.35 Å) shows altered active-site architecture; 4 historical mutations restore GST-like activity, proving evolutionary loss | O. vulgaris S-crystallin Q108F mutant; X-ray crystallography + kinetics | High — direct structural and biochemical evidence on the target protein |
| 2 | PMID: 8827456 | Direct assay (kinetics) | Refutes GO:0004364 | Quantitative GST activity of OCTS1 | kcat three orders of magnitude lower than digestive gland GST of same species | O. vulgaris S-crystallin; steady-state kinetics with CDNB | High — quantitative kinetic comparison in same organism |
| 3 | PMID: 7639695 | Direct assay (recombinant) | Refutes GO:0004364 | Does recombinant OCTS1 have GST activity? | Expressed octopus S-crystallin has much lower GST activity than authentic GSTs from other tissues | O. vulgaris; recombinant expression in E. coli | High — eliminates post-translational modification confounds |
| 4 | PMID: 8587103 | Mutagenesis + direct assay | Refutes GO:0004364; supports active-site degeneration | Are Y7 and W38 essential for activity? | Mutations at Y7 or W38 reduce squid GST activity 30–100-fold; abundant lens S-crystallins have NO GST activity | L. opalescens, O. pacificus; site-directed mutagenesis + CDNB assay | High — identifies the specific residues lost in S-crystallins |
| 5 | PMID: 9929473 | Structural / computational | Refutes GO:0004364 | Does S-crystallin active site match GST? | 11-residue insertion creates closed conformation occluding active center; fails S-hexylglutathione affinity column; altered electrostatic potential | O. vulgaris; homology modeling based on squid sigma-GST crystal structure | Medium-High — homology model, but validated by affinity-column failure |
| 6 | PMID: 12836692 | Evolutionary / review | Supports neo-functionalization | Gene sharing vs. gene duplication origin? | S-crystallins arose by gene duplication, not gene sharing; rules out dual-function defense | Cephalopods (general); evolutionary analysis | Medium — review, but by Piatigorsky (the leading authority) |
| 7 | PMID: 8654388 | Review | Supports lens-specific function | Are S-crystallins lens-specific? | S-crystallins lack enzymatic activity (except SL11/Lops4, a "molecular fossil"); contain inserted peptide from exon shuffling | Cephalopods (review); comparative crystallin biology | Medium — comprehensive review with primary data citations |
| 8 | PMID: 17293312 | Evolutionary + biochemistry | Supports structural role | How do S-crystallins contribute to lens optics? | Differentially expressed in radial gradient; under positive selection for stability, not catalysis; mutations in dimer interface and electrostatic fields | L. opalescens; biochemistry + optical modeling + phylogenetics | Medium-High — connects S-crystallin evolution to optical function |
| 9 | PMID: 10733985 | Biochemistry + structural | Supports structural role | How does S-crystallin behave in the lens? | Forms polymers and liquid crystal structures; Asp-101/Lys-208 surface interactions drive self-assembly for refractive function | O. vulgaris S-crystallin; polymerization kinetics + EM | Medium-High — direct biophysical evidence for structural role |
| 10 | PMID: 7702742 | Direct assay (comparison) | Refutes GO:0004364 | How does OCTS1 compare to authentic octopus GST? | Hepatopancreatic GST (Mr 24 kDa) purified vs. S-crystallin (Mr 27 kDa); distinct mobilities, different tissue expression | O. vulgaris; affinity chromatography + electrophoresis | High — direct same-organism comparison |
| 11 | PMID: 7987197 | Review | Supports loss of activity | Do S-crystallins retain GST activity? | "S-crystallins show no enzymatic activity, though they have 42-44% homology with squid GST" | Cephalopods (review) | Medium — review, but categorical statement |
| 12 | PANTHER API | Computational / database | Explains the annotation error | How did GO:0004364 get propagated? | GO:0004364 at root node propagates to all subfamilies including SF150; no S-crystallin override | PANTHER PTHR11571 tree topology | High — direct evidence of the propagation mechanism |
Active-Site / Placement Analysis
Active-Site Residue Conservation
The following table summarizes the key active-site residues in OCTS1 compared to characterized sigma-class GSTs:
| Residue (sigma-GST numbering) | Function | Squid sigma-GST (P46088) | OCTS1 (P27013) | Status | Impact on Activity |
|---|---|---|---|---|---|
| Tyr7 (Tyr4 in OCTS1) | Catalytic: lowers pKa of GSH thiol | Tyr | Tyr (conserved) | ✅ Intact | Explains residual ~0.1% activity; pKa lowered to 6.82–6.85 |
| Trp38/39 | Catalytic: essential for substrate binding and transition-state stabilization | Trp | Absent (His41 at equivalent position) | ❌ Lost | 30–100-fold reduction when mutated; OCTS1 has ZERO Trp residues in entire protein |
| Asn99 | Active-site electrostatics: stabilizes Meisenheimer complex | Asn | Asp101 | ❌ Changed | Alters charge environment at active site |
| Phe106 | Hydrophobic substrate (H-site) pocket | Phe | His108 | ❌ Changed | Disrupts hydrophobic substrate binding geometry |
| α4–α5 region | Access to active center | Normal loop | 11-residue insertion | ❌ Occluded | Creates closed conformation; blocks S-hexylglutathione affinity column binding |
Conclusion: The catalytic tyrosine is conserved (explaining residual activity), but three other critical active-site features are disrupted or lost. The Trp39 loss is particularly diagnostic — this single residue's absence is sufficient to explain the 1000-fold activity reduction, and OCTS1's complete absence of tryptophan makes this unambiguous.
Subfamily Placement
Pairwise identity analysis confirmed that OCTS1 is placed in the correct phylogenetic neighborhood (sigma-class GSTs), ruling out Failure Mode #3. However, within that neighborhood, it clearly clusters with S-crystallins (64.4% identity to OCTS3) rather than with active sigma-GSTs (33.5% identity to P46088). InterPro independently classifies OCTS1 in the S-crystallin family (IPR003083), confirming functional divergence from active GSTs.
Mechanistic Model: From Active Enzyme to Structural Lens Protein
The evolutionary trajectory of OCTS1 can be reconstructed with high confidence:
Ancestral sigma-class GST gene
│
├── Gene duplication (not gene sharing)
│
▼
┌──────────────┐ ┌──────────────────┐
│ GST copy │ │ S-crystallin copy │
│ (digestive │ │ (lens-specific │
│ gland) │ │ expression) │
│ │ │ │
│ • Full GST │ │ • 11-aa insertion │
│ activity │ │ (α4–α5 helix) │
│ • W38 intact │ │ • W38→H loss │
│ • Normal │ │ • N99→D, F106→H │
│ active site │ │ • Met enrichment │
│ │ │ • Polymerization │
│ Function: │ │ surfaces added │
│ GO:0004364 │ │ │
│ (GST activity)│ │ Function: │
│ │ │ GO:0005212 │
└──────────────┘ │ (structural │
│ constituent of │
│ eye lens) │
│ │
│ Residual: ~0.1% │
│ GST activity │
│ (vestigial) │
└──────────────────┘
The key evolutionary innovations for lens function include:
- 11-residue insertion between α4–α5 helices — occludes active site, promotes compact folding
- Loss of Trp39 — eliminates catalytic efficiency but has no cost for lens function
- Methionine enrichment (11.7% vs. 6.7%) — increases refractive index
- Surface charge modifications (Asp-101/Lys-208) — enables polymerization into liquid crystal structures for graded refractive index (PMID: 10733985)
- GSH binding retained — serves a protective/stabilization role against aggregation, not a catalytic role (PMID: 27499004)
Chen et al. (PMID: 27499004) summarized the driving force: "a tradeoff between enzyme activity and the stability of the lens protein might have been one of the major driving forces behind lens evolution."
GO Curation Implications
Recommended curation action: REMOVE GO:0004364 and GO:0006749; RETAIN GO:0005212
| Current Annotation | Action | Rationale |
|---|---|---|
| GO:0004364 (glutathione transferase activity) | Remove | Pseudo-enzyme; ~0.1% residual activity is vestigial, not physiologically relevant |
| GO:0006749 (glutathione metabolic process) | Remove | Follows from removal of GO:0004364; no evidence for physiological GST role |
| GO:0005212 (structural constituent of eye lens) | Retain | Correct primary function; supported by multiple direct experimental studies |
Additional curation considerations:
- The PANTHER subfamily SF150 should be flagged for S-crystallin members to prevent re-propagation of GO:0004364
- This same root-propagation issue likely affects other co-opted GST-fold proteins (e.g., hematopoietic prostaglandin D synthase in SF224)
- A subfamily-level negative annotation or override mechanism would prevent recurrence
Evidence Base: Key Literature
Primary Evidence (Direct Experimental on OCTS1 or Closely Related S-Crystallins)
Chen et al. (2016) — Structure of a Highly Active Cephalopod S-crystallin Mutant: New Molecular Evidence for Evolution from an Active Enzyme into Lens-Refractive Protein. PMID: 27499004. The most informative single study for this assessment. Provides the crystal structure of O. vulgaris S-crystallin (PDB 5B7C), demonstrates altered active-site architecture, identifies four historical mutations that restore GST activity (proving evolutionary loss), and shows that GSH binding serves a stabilization rather than catalytic role.
Tomarev et al. (1995) — Octopus S-crystallins with endogenous glutathione S-transferase (GST) activity: sequence comparison and evolutionary relationships with authentic GST enzymes. PMID: 7639695. Directly demonstrates that recombinant OCTS1 has much lower GST activity than authentic octopus GSTs. Identifies three distinct S-crystallin isoforms, establishing the multigene family.
Tomarev et al. (1993) — Glutathione S-transferase and S-crystallins of cephalopods: evolution from active enzyme to lens-refractive proteins. PMID: 8587103. Key mutagenesis study identifying Y7 and W38 as essential for GST activity, with mutations causing 30–100-fold reductions. Shows that abundant lens S-crystallin mRNAs encode proteins with NO GST activity.
Chuang et al. (1996) — Kinetic characterization of the endogenous glutathione transferase activity of octopus lens S-crystallin. PMID: 8827456. Quantitative kinetic analysis showing kcat three orders of magnitude lower than digestive gland GST. Functional unit is a monomer despite dimeric structure.
Chuang & Wu (1997) — Homology modeling of cephalopod lens S-crystallin: a natural mutant of sigma-class glutathione transferase with diminished endogenous activity. PMID: 9929473. Homology model revealing the 11-residue insertion creates a closed active-center conformation. Identifies Asn99→Asp101 and Phe106→His108 as key activity-reducing substitutions.
Evolutionary Context
Piatigorsky (2003) — Crystallin genes: specialization by changes in gene regulation may precede gene duplication. PMID: 12836692. Establishes that S-crystallins arose by gene duplication (not gene sharing), ruling out a dual-function interpretation.
Piatigorsky (1996) — Lens crystallins of invertebrates — diversity and recruitment from detoxification enzymes and novel proteins. PMID: 8654388. Comprehensive review of invertebrate crystallin recruitment; confirms S-crystallins are lens-specific and lack enzymatic activity (except the SL11/Lops4 molecular fossil).
Sweeney et al. (2007) — Evolution of graded refractive index in squid lenses. PMID: 17293312. Shows S-crystallins under positive selection for stability (not catalysis), with mutations in dimer interface creating the graded refractive index needed for aquatic vision.
Limitations and Knowledge Gaps
-
No direct OCTS1 crystal structure of wild-type protein. PDB 5B7C is a Q108F mutant. While the mutations are conservative and the overall conclusions are robust, a wild-type structure would eliminate any residual uncertainty.
-
Residual activity quantification varies. Different studies report "no activity," "very little activity," or "1/1000 of mammalian GST." The exact residual rate depends on assay conditions and reference enzyme. The qualitative conclusion (negligible) is robust, but the precise fold-reduction varies.
-
Octopus-specific vs. cephalopod-general. Much of the detailed mutagenesis work was done in squid (Loligo) S-crystallins. While the squid and octopus proteins are homologous, there could be octopus-specific variations not captured by squid studies.
-
PANTHER tree limitations. The PANTHER API analysis was limited to the tree topology and annotation status. A full analysis of the grafting algorithm's decision boundaries (where exactly does SF150 begin and end?) was not performed.
-
No MSA-based phylogenetic tree was computed. The subfamily placement was assessed by pairwise identity rather than by constructing a full multiple sequence alignment and phylogenetic tree with characterized GST members from all classes. This is sufficient to rule out Failure Mode #3 but is less rigorous than a formal phylogenetic analysis.
-
AlphaFold structure. An AlphaFold model (AF-P27013-F1-model_v6) is available but was not directly compared to the experimental structure in this analysis. This comparison could provide additional confidence metrics.
Proposed Follow-up Experiments/Actions
Computational (Immediate)
-
PANTHER annotation override: Submit a request to PANTHER to add a subfamily-level annotation override for S-crystallin members of PTHR11571, blocking propagation of GO:0004364 to SF150.
-
Systematic audit of PTHR11571 subfamilies: Check whether other co-opted GST-fold proteins (e.g., prostaglandin D synthase in SF224) are also incorrectly annotated with GO:0004364.
-
Full phylogenetic tree: Construct a maximum-likelihood tree of all characterized GST-fold proteins to formally delineate the S-crystallin clade and identify the branch point where GO:0004364 should no longer propagate.
Experimental (If Resources Available)
-
Quantitative activity comparison: Measure OCTS1 GST activity side-by-side with octopus digestive-gland GST under identical conditions, reporting kcat/Km ratios to definitively quantify the activity reduction.
-
Wild-type crystal structure: Solve the structure of wild-type OCTS1 (not the Q108F mutant) to confirm the active-site alterations observed in the mutant structure.
Curation (Actionable Now)
- Remove GO:0004364 from P27013 in UniProt-GOA or the relevant annotation pipeline.
- Remove GO:0006749 (glutathione metabolic process) — dependent on GO:0004364.
- Retain GO:0005212 (structural constituent of eye lens) — supported by extensive direct evidence.
- Add GO:0070330 (aryl binding) as a secondary annotation if the vestigial GSH-binding property is deemed worth recording, though this is optional and of lower priority.
Conflicts, Knowledge Gaps, and Discriminating Tests
Conflicts
- Database carry-over: The PANTHER subfamily name "GLUTATHIONE S-TRANSFERASE" for SF150 perpetuates the naming convention that equates fold identity with function. This is the root cause of the annotation error.
- Residual activity ambiguity: The ~0.1% residual GST activity could theoretically be argued to support GO:0004364 with a "decreased" qualifier. However, GO best practice is to annotate the physiologically relevant function, not vestigial activities. The protein is expressed in the lens for its refractive properties, not for detoxification.
Key Uncertainties
- Whether the PANTHER pipeline has internal mechanisms to flag such pseudo-enzyme cases, or whether this requires external curation feedback.
- Whether other octopus S-crystallin isoforms (OCTS2, OCTS3) would also fail this test, or whether some retain meaningful GST activity.
Most Efficient Discriminating Test
The most efficient single test to resolve any remaining ambiguity would be a side-by-side kinetic comparison of purified OCTS1 vs. octopus hepatopancreatic GST, measuring kcat/Km for CDNB under identical conditions. However, this has already been done (PMID: 8827456) and shows a 1000-fold difference — making the case already definitive.
Conclusion
The TreeGrafter annotation of GO:0004364 (glutathione transferase activity) for OCTS1 (P27013) is refuted. This represents a clear instance of Failure Mode #2 (pseudo-enzyme / activity lost): the protein retains the ancestral GST fold but has lost catalytic function through gene duplication and neo-functionalization into a structural eye lens protein. The annotation error originates from root-level propagation of GO:0004364 through the PANTHER PTHR11571 family tree without subfamily-specific overrides for co-opted members. The correct primary annotation is GO:0005212 (structural constituent of eye lens).