Deep Research
Falcon.body
(ahpC-deep-research-falcon.body.md)
Falcon.body
(ahpC-deep-research-falcon.body.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: ahpC (UniProt Q88K52; locus PP_2439) in Pseudomonas putida KT2440
1) Target verification (gene/protein identity)
The research target is alkyl hydroperoxide reductase C (AhpC), a peroxiredoxin-family thiol peroxidase, encoded by ahpC with ordered locus name PP_2439 in Pseudomonas putida KT2440, matching the user-provided UniProt context for Q88K52. This locus assignment is explicitly used in oxidative-stress transcriptomics for KT2440 (Table 2 lists ahpC = PP_2439 and ahpF = PP_2440). (bojanovic2017globaltranscriptionalresponses pages 10-11, bojanovic2017globaltranscriptionalresponses media 03a7ec25)
2) Key concepts and definitions (current understanding)
2.1 AhpC as a bacterial peroxiredoxin (AhpC/Prx1 subfamily)
AhpC is the peroxidatic component (“C” subunit) of the bacterial alkyl hydroperoxide reductase (Ahp) system, belonging to the peroxiredoxin family. In P. putida, AhpC is observed as an abundant soluble ~24 kDa protein and is predicted/confirmed to be a 2-Cys peroxiredoxin with conserved cysteines (e.g., Cys47 and Cys166 in KT2442 AhpC), consistent with the canonical Prx catalytic cycle. (fukumori2001molecularcloningand pages 4-6, fukumori2001molecularcloningand pages 3-4)
2.2 Enzymatic reaction and substrate specificity
Reaction (functional definition): AhpC catalyzes the reduction of hydroperoxides (ROOH, including organic hydroperoxides and H2O2) to the corresponding products (ROH + H2O, for alkyl hydroperoxides). In P. putida KT2442, AhpC is described as reducing alkyl hydroperoxides to the corresponding alcohol, with oxidized AhpC then being recycled by AhpF. (fukumori2001molecularcloningand pages 6-8)
Substrate preference in P. putida evidence: Experimental phenotypes and expression studies in KT2442 indicate AhpC is especially important for organic hydroperoxide detoxification; hydrogen peroxide was reported as potentially not the preferred substrate for the P. putida AHPR system in the KT2442 context (e.g., AhpC overexpression increased resistance to t‑butyl hydroperoxide but did not similarly improve H2O2 resistance). (fukumori2001molecularcloningand pages 6-8, fukumori2001molecularcloningand pages 3-4)
General peroxiredoxin perspective: A dedicated review and P. putida-focused oxidative-stress review describe AhpC as a high-affinity peroxide detoxification tool (qualitatively “very low Km” for hydroperoxides is noted), but those sources do not provide numeric kinetic constants for P. putida AhpC in the retrieved text. (hishinuma2006oxyrregulatedthe pages 1-2, kim2014oxidativestressresponse pages 5-6)
2.3 Electron donor system and catalytic cycling (AhpF/NADH)
In P. putida, the AhpC catalytic cycle is coupled to a dedicated reductase AhpF, characterized as a flavoprotein/disulfide reductase that uses NADH as the reducing equivalent to restore AhpC from its oxidized form. This NADH dependence is repeatedly emphasized and is a key mechanistic constraint: the AhpCF system may be less suited for detoxifying very large peroxide loads because the peroxidase must be continually re-reduced at the expense of reducing equivalents. (hishinuma2006oxyrregulatedthe pages 1-2, kim2014oxidativestressresponse pages 5-6)
Consistent with this electron-transfer role, P. putida KT2442 AhpF is predicted to have conserved FAD-binding and NAD(P)H-binding motifs/domains typical of electron-transfer flavoenzymes, supporting an electron flow NAD(P)H → AhpF → AhpC → peroxide. (fukumori2001molecularcloningand pages 4-6)
3) Pathway context: operon organization and regulation
3.1 Operon organization and transcriptional architecture (ahpCF)
In P. putida KT2442 (closely related to KT2440), ahpC and ahpF are adjacent and co-transcribed as an ahpCF operon, but with strong differential accumulation of transcripts/proteins.
Key evidence includes:
- a predominant ~0.7 kb transcript corresponding largely to ahpC, and a less abundant ~2.7 kb transcript consistent with ahpCF co-transcription. (fukumori2001molecularcloningand pages 4-6, dubbs2007peroxiredoxinsinbacterial pages 9-12)
- evidence for transcriptional attenuation between ahpC and ahpF: a stem-loop structure in the intergenic region can reduce readthrough to ahpF, explaining lower AhpF accumulation; deletion of this stem-loop increases ahpF expression. (fukumori2001molecularcloningand pages 6-8, fukumori2001molecularcloningand pages 4-6)
- mapped transcription start sites: in KT2442, ahpC transcription initiates from two closely spaced starts (P1 at −37; P2 at −28 relative to the start site), and expression is reported as RpoS-independent (transcripts present even in an rpoS mutant). (hishinuma2006oxyrregulatedthe pages 6-7)
3.2 OxyR-controlled oxidative stress regulon
A central regulatory concept is that OxyR serves as a peroxide-sensing transcriptional regulator that controls antioxidant genes including ahpC.
Direct regulatory evidence in P. putida includes:
- OxyR binds the ahpC promoter region (gel shift/binding evidence). (hishinuma2006oxyrregulatedthe pages 1-2)
- mechanistic model: reduced OxyR can bind the promoter, while oxidized OxyR is linked to transcriptional activation (e.g., via interaction with RNA polymerase). (hishinuma2006oxyrregulatedthe pages 6-7)
- oxyR1 point mutation: in KT2442TOL, a point mutation in oxyR (Phe106→Ile is described for oxyR1) is associated with constitutively increased AhpC expression, consistent with an altered redox-sensing/activation state. (fukumori2001molecularcloningand pages 6-8, fukumori2001molecularcloningand pages 1-3)
4) Cellular localization and site of action
Experimental fractionation in P. putida KT2442 recovered AhpC as a soluble protein in the soluble fraction, supporting a cytosolic site of action where intracellular hydroperoxides are detoxified. No evidence in the retrieved sources supports secretion or a membrane-anchored localization for AhpC. (fukumori2001molecularcloningand pages 3-4)
5) Physiological roles, phenotypes, and quantitative data
5.1 Organic peroxide resistance and solvent-associated stress tolerance (KT2442 studies)
In KT2442/KT2442TOL, AhpC is induced by oxidants and contributes to resistance to organic hydroperoxides:
- The 24-kDa AhpC-like protein is inducibly produced by H2O2 or t‑butyl hydroperoxide (BHP) supplementation (but not by toluene alone in the parental strain). (fukumori2001molecularcloningand pages 3-4)
- Disk diffusion phenotypes show markedly increased resistance to BHP in the toluene-adapted strain (smaller zone = more resistant): BHP zone KT2442 30 ± 2 mm, KT2442TOL 14 ± 1 mm, and oxyR revertant KT2442TOL-oxyR 30 ± 2 mm. In contrast, H2O2 zones were similar across strains (23 ± 1, 24 ± 1, 25 ± 1 mm, respectively), supporting a more prominent AhpC role in organic peroxide defense than in bulk H2O2 detoxification under those conditions. (fukumori2001molecularcloningand pages 3-4)
- A toluene colony-formation phenotype links oxyR/AhpC regulation to solvent stress adaptation: colony-forming ability under toluene decreased from ~92 ± 10% (KT2442TOL) to ~51 ± 8% (KT2442TOL-oxyR), consistent with a partial contribution of OxyR-controlled oxidative-stress defenses (including AhpC) to solvent-associated fitness. (fukumori2001molecularcloningand pages 6-8)
5.2 Rapid induction by hydrogen peroxide in KT2440 (transcriptomics)
A genome-wide expression dataset for KT2440 exposed to hydrogen peroxide shows AhpC and AhpF are among the most strongly induced oxidative-stress genes early after exposure:
- ahpC (PP_2439): 74.7-fold induction at 7 min (T1) and 2.3-fold at 60 min (T2).
- ahpF (PP_2440): 203.4-fold induction at 7 min (T1) and 3.5-fold at 60 min (T2).
This time-resolved pattern indicates a rapid transcriptional spike followed by partial relaxation/adaptation. (bojanovic2017globaltranscriptionalresponses pages 10-11, bojanovic2017globaltranscriptionalresponses media 03a7ec25)
5.3 qPCR evidence for strong OxyR1-dependent ahpC upregulation
In P. putida (OxyR study), qPCR Ct values indicate that an oxyR1 background causes very strong elevation of ahpC transcript abundance across growth states. Reported Ct values include (LOG phase) 20.10 ± 0.34 (oxyR) vs 12.49 ± 0.75 (oxyR1), and (stationary) 15.16 ± 1.43 (oxyR) vs 10.67 ± 0.15 (oxyR1). The large Ct decreases imply very large fold-changes in transcript abundance (since each Ct approximates a doubling under ideal PCR efficiency). (hishinuma2006oxyrregulatedthe pages 6-7)
6) Recent developments (2023–2024) and emerging applications
Direct 2023–2024 primary studies centered specifically on KT2440 AhpC biochemistry were limited in the retrieved corpus. However, recent work in related environmental bacteria and applied contexts reinforces AhpC’s importance as part of oxidative-stress defense relevant to biotechnology.
6.1 Bioremediation context: oxidative stress during aromatic compound degradation (2024)
A 2024 study in Paraburkholderia xenovorans (an environmental aromatic degrader) shows that aromatic degradation can trigger oxidative stress and upregulation of multiple detoxification enzymes, including alkyl hydroperoxide reductase/peroxiredoxin components among a broader antioxidant network. Importantly, the authors demonstrate a real-world implementation: overexpression of a redox protein (FldX1) improves degradation performance in soil microcosms (bioaugmentation) by mitigating oxidative stress, which in turn alters the need to induce antioxidant genes (including those in peroxide detoxification pathways). This supports the general translational principle that peroxide-defense modules like AhpC/AhpF are operationally important in field-relevant biodegradation scenarios, even when the specific organism differs from P. putida. (rodriguezcastro2024thelongchainflavodoxin pages 10-11)
7) Expert opinion and synthesis (authoritative analyses)
Two overarching expert interpretations emerge from reviews and P. putida regulatory work:
1. AhpC is a high-affinity thiol peroxidase central to peroxide defense, especially at low-to-moderate peroxide levels where peroxiredoxins can be highly effective scavengers. (hishinuma2006oxyrregulatedthe pages 1-2, kim2014oxidativestressresponse pages 5-6)
2. Dependence on reductant supply (NADH via AhpF) implies a potential limitation for coping with massive peroxide influx; thus, AhpC typically operates alongside catalases and other systems in a coordinated OxyR-controlled oxidative stress response. (hishinuma2006oxyrregulatedthe pages 1-2, kim2014oxidativestressresponse pages 5-6, bojanovic2017globaltranscriptionalresponses pages 10-11)
8) Evidence summary table
| Aspect | Key findings | Evidence type | Citations |
|---|---|---|---|
| Identity/domains | Target identity is consistent across sources: ahpC = PP_2439 in Pseudomonas putida KT2440/KT2442, encoding a small, soluble ~24 kDa alkyl hydroperoxide reductase subunit C/peroxiredoxin. The protein is a 2-Cys peroxiredoxin with conserved cysteines at positions Cys47 and Cys166, matching the AhpC/Prx1 family assignment. | Primary study; review | (fukumori2001molecularcloningand pages 4-6, fukumori2001molecularcloningand pages 1-3, bojanovic2017globaltranscriptionalresponses pages 10-11) |
| Reaction/substrates | AhpC is the peroxidatic subunit of alkyl hydroperoxide reductase and reduces organic hydroperoxides to the corresponding alcohols; it can also detoxify H2O2, but evidence in P. putida suggests organic hydroperoxides are preferred substrates and H2O2 may be less preferred. Reviews note very low Km values for hydroperoxides qualitatively, but no numeric kinetics were recovered here for P. putida AhpC. | Primary study; review | (hishinuma2006oxyrregulatedthe pages 1-2, fukumori2001molecularcloningand pages 6-8, kim2014oxidativestressresponse pages 5-6) |
| Electron donor/regeneration | Oxidized AhpC is regenerated by AhpF, the dedicated NADH-dependent flavo-disulfide reductase/peroxiredoxin reductase. AhpF contains conserved FAD-binding and NAD(P)H-binding motifs, supporting electron flow from NADH → AhpF → AhpC → peroxide substrate. | Primary study; review | (hishinuma2006oxyrregulatedthe pages 1-2, fukumori2001molecularcloningand pages 4-6, fukumori2001molecularcloningand pages 1-3, kim2014oxidativestressresponse pages 5-6) |
| Regulation | OxyR directly regulates ahpC. Purified OxyR binds upstream of ahpC; reduced OxyR can bind promoter DNA, whereas oxidized OxyR is linked to transcriptional activation. An oxyR1 mutation causes constitutively elevated ahpC expression. | Primary study; review | (hishinuma2006oxyrregulatedthe pages 1-2, fukumori2001molecularcloningand pages 1-3, dubbs2007peroxiredoxinsinbacterial pages 9-12, hishinuma2006oxyrregulatedthe pages 6-7) |
| Operon/transcription | ahpC and ahpF are co-transcribed in an ahpCF operon. A major ~0.7 kb ahpC transcript and a less abundant ~2.7 kb ahpCF transcript were detected; an additional ~1.4 kb ahpF RNA may arise from processing or a second promoter. A stem-loop between ahpC and ahpF likely attenuates downstream ahpF expression, explaining lower AhpF abundance. Transcription starts were mapped to two ahpC starts (P1 = -37, P2 = -28). | Primary study; review | (fukumori2001molecularcloningand pages 6-8, fukumori2001molecularcloningand pages 4-6, dubbs2007peroxiredoxinsinbacterial pages 9-12, hishinuma2006oxyrregulatedthe pages 6-7) |
| Localization | Experimental descriptions identify AhpC as a soluble protein recovered in the soluble/cytosolic fraction; no evidence here supports secretion or membrane localization. Function is therefore most parsimoniously assigned to the cytosol, where it detoxifies intracellular peroxides generated by metabolism or chemical stress. | Primary study; inference from fractionation/review | (fukumori2001molecularcloningand pages 1-3, fukumori2001molecularcloningand pages 3-4) |
| Physiological roles/phenotypes | AhpC contributes to oxidative stress defense, especially against organic hydroperoxides. In KT2442, induction occurred with H2O2 or t-butyl hydroperoxide (BHP), but not toluene alone. Overproduction of AhpC in the toluene-adapted strain correlated with improved BHP resistance: inhibition zones were 30 ± 2 mm in KT2442, 14 ± 1 mm in KT2442TOL, and 30 ± 2 mm in the oxyR-revertant; H2O2 zones remained similar (23 ± 1, 24 ± 1, 25 ± 1 mm, respectively). Heterologous expression of P. putida ahpC partially protected E. coli against BHP but not H2O2. Reversion of oxyR reduced colony formation in toluene from ~92 ± 10% in KT2442TOL to ~51 ± 8% in KT2442TOL-oxyR, linking OxyR/AhpC overexpression with solvent-associated stress tolerance. | Primary study; review | (fukumori2001molecularcloningand pages 6-8, fukumori2001molecularcloningand pages 1-3, fukumori2001molecularcloningand pages 3-4, kim2014oxidativestressresponse pages 5-6) |
| Quantitative expression stats | Under H2O2 stress in KT2440, ahpC (PP_2439) increased 74.7-fold at 7 min and 2.3-fold at 60 min; ahpF (PP_2440) increased 203.4-fold at 7 min and 3.5-fold at 60 min. In an oxyR1 background, ahpC qPCR Ct values were 20.10 ± 0.34 (LOG oxyR) vs 12.49 ± 0.75 (LOG oxyR1), and 15.16 ± 1.43 (ST oxyR) vs 10.67 ± 0.15 (ST oxyR1), indicating very large transcript increases in oxyR1. | Primary study | (hishinuma2006oxyrregulatedthe pages 6-7, bojanovic2017globaltranscriptionalresponses pages 10-11, bojanovic2017globaltranscriptionalresponses media 03a7ec25) |
Table: This table summarizes the core functional annotation for Pseudomonas putida AhpC (PP_2439; UniProt Q88K52), including enzymatic role, regulation, operon structure, localization, phenotypes, and quantitative expression evidence. It is useful as a compact evidence map linking the curated protein identity to experimental studies in P. putida.
9) Key evidence figure/table
A cropped image of Table 2 reporting H2O2-induced fold changes for ahpC (PP_2439) and ahpF (PP_2440) at 7 min and 60 min is available for direct visual verification. (bojanovic2017globaltranscriptionalresponses media 03a7ec25)
10) Practical functional annotation (concise)
- Primary molecular function: thiol-dependent peroxidase reducing organic hydroperoxides (and H2O2 under some conditions) using the AhpF/NADH reducing system. (fukumori2001molecularcloningand pages 6-8, kim2014oxidativestressresponse pages 5-6)
- Likely cellular compartment: cytosol/soluble fraction. (fukumori2001molecularcloningand pages 3-4)
- Pathway/regulon: OxyR-controlled oxidative stress regulon; ahpCF operon with transcriptional attenuation influencing AhpF abundance. (fukumori2001molecularcloningand pages 4-6, dubbs2007peroxiredoxinsinbacterial pages 9-12, hishinuma2006oxyrregulatedthe pages 6-7)
Limitations of the current evidence set
While core function and regulation of P. putida AhpC are strongly supported by classic primary literature and authoritative reviews, the retrieved corpus contained few 2023–2024 studies directly focused on AhpC in P. putida KT2440 with detailed enzymology (e.g., Km/kcat for specific hydroperoxide substrates). Consequently, quantitative enzymatic kinetics for the specific Q88K52 protein are not reported here, and the “recent developments” section relies partly on 2024 work in a closely related environmental biodegradation context rather than P. putida KT2440 itself. (hishinuma2006oxyrregulatedthe pages 1-2, fukumori2001molecularcloningand pages 6-8, rodriguezcastro2024thelongchainflavodoxin pages 10-11)
References
-
(bojanovic2017globaltranscriptionalresponses pages 10-11): Klara Bojanovič, Isotta D'Arrigo, and Katherine S. Long. Global transcriptional responses to osmotic, oxidative, and imipenem stress conditions in pseudomonas putida. Applied and Environmental Microbiology, Apr 2017. URL: https://doi.org/10.1128/aem.03236-16, doi:10.1128/aem.03236-16. This article has 82 citations and is from a peer-reviewed journal.
-
(bojanovic2017globaltranscriptionalresponses media 03a7ec25): Klara Bojanovič, Isotta D'Arrigo, and Katherine S. Long. Global transcriptional responses to osmotic, oxidative, and imipenem stress conditions in pseudomonas putida. Applied and Environmental Microbiology, Apr 2017. URL: https://doi.org/10.1128/aem.03236-16, doi:10.1128/aem.03236-16. This article has 82 citations and is from a peer-reviewed journal.
-
(fukumori2001molecularcloningand pages 4-6): Fumiyasu Fukumori and Mitsuru Kishii. Molecular cloning and transcriptional analysis of the alkyl hydroperoxide reductase genes from pseudomonas putida kt2442. The Journal of general and applied microbiology, 47 5:269-277, Oct 2001. URL: https://doi.org/10.2323/jgam.47.269, doi:10.2323/jgam.47.269. This article has 21 citations.
-
(fukumori2001molecularcloningand pages 3-4): Fumiyasu Fukumori and Mitsuru Kishii. Molecular cloning and transcriptional analysis of the alkyl hydroperoxide reductase genes from pseudomonas putida kt2442. The Journal of general and applied microbiology, 47 5:269-277, Oct 2001. URL: https://doi.org/10.2323/jgam.47.269, doi:10.2323/jgam.47.269. This article has 21 citations.
-
(fukumori2001molecularcloningand pages 6-8): Fumiyasu Fukumori and Mitsuru Kishii. Molecular cloning and transcriptional analysis of the alkyl hydroperoxide reductase genes from pseudomonas putida kt2442. The Journal of general and applied microbiology, 47 5:269-277, Oct 2001. URL: https://doi.org/10.2323/jgam.47.269, doi:10.2323/jgam.47.269. This article has 21 citations.
-
(hishinuma2006oxyrregulatedthe pages 1-2): Sota Hishinuma, Masahiro Yuki, Makoto Fujimura, and Fumiyasu Fukumori. Oxyr regulated the expression of two major catalases, kata and katb, along with peroxiredoxin, ahpc in pseudomonas putida. Environmental microbiology, 8 12:2115-24, Dec 2006. URL: https://doi.org/10.1111/j.1462-2920.2006.01088.x, doi:10.1111/j.1462-2920.2006.01088.x. This article has 86 citations and is from a domain leading peer-reviewed journal.
-
(kim2014oxidativestressresponse pages 5-6): Jisun Kim and Woojun Park. Oxidative stress response in pseudomonas putida. Applied Microbiology and Biotechnology, 98:6933-6946, Jun 2014. URL: https://doi.org/10.1007/s00253-014-5883-4, doi:10.1007/s00253-014-5883-4. This article has 142 citations and is from a domain leading peer-reviewed journal.
-
(dubbs2007peroxiredoxinsinbacterial pages 9-12): James M. Dubbs and Skorn Mongkolsuk. Peroxiredoxins in bacterial antioxidant defense. Sub-cellular biochemistry, 44:143-93, Jan 2007. URL: https://doi.org/10.1007/978-1-4020-6051-9_7, doi:10.1007/978-1-4020-6051-9_7. This article has 133 citations.
-
(hishinuma2006oxyrregulatedthe pages 6-7): Sota Hishinuma, Masahiro Yuki, Makoto Fujimura, and Fumiyasu Fukumori. Oxyr regulated the expression of two major catalases, kata and katb, along with peroxiredoxin, ahpc in pseudomonas putida. Environmental microbiology, 8 12:2115-24, Dec 2006. URL: https://doi.org/10.1111/j.1462-2920.2006.01088.x, doi:10.1111/j.1462-2920.2006.01088.x. This article has 86 citations and is from a domain leading peer-reviewed journal.
-
(fukumori2001molecularcloningand pages 1-3): Fumiyasu Fukumori and Mitsuru Kishii. Molecular cloning and transcriptional analysis of the alkyl hydroperoxide reductase genes from pseudomonas putida kt2442. The Journal of general and applied microbiology, 47 5:269-277, Oct 2001. URL: https://doi.org/10.2323/jgam.47.269, doi:10.2323/jgam.47.269. This article has 21 citations.
-
(rodriguezcastro2024thelongchainflavodoxin pages 10-11): Laura Rodríguez-Castro, Roberto E. Durán, Valentina Méndez, Flavia Dorochesi, Daniela Zühlke, Katharina Riedel, and Michael Seeger. The long-chain flavodoxin fldx1 improves the biodegradation of 4-hydroxyphenylacetate and 3-hydroxyphenylacetate and counteracts the oxidative stress associated to aromatic catabolism in paraburkholderia xenovorans. Biological Research, Apr 2024. URL: https://doi.org/10.1186/s40659-024-00491-4, doi:10.1186/s40659-024-00491-4. This article has 6 citations and is from a peer-reviewed journal.
Falcon
(ahpC-deep-research-falcon.md)
Falcon
(ahpC-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: ahpC (UniProt Q88K52; locus PP_2439) in Pseudomonas putida KT2440
1) Target verification (gene/protein identity)
The research target is alkyl hydroperoxide reductase C (AhpC), a peroxiredoxin-family thiol peroxidase, encoded by ahpC with ordered locus name PP_2439 in Pseudomonas putida KT2440, matching the user-provided UniProt context for Q88K52. This locus assignment is explicitly used in oxidative-stress transcriptomics for KT2440 (Table 2 lists ahpC = PP_2439 and ahpF = PP_2440). (bojanovic2017globaltranscriptionalresponses pages 10-11, bojanovic2017globaltranscriptionalresponses media 03a7ec25)
2) Key concepts and definitions (current understanding)
2.1 AhpC as a bacterial peroxiredoxin (AhpC/Prx1 subfamily)
AhpC is the peroxidatic component (“C” subunit) of the bacterial alkyl hydroperoxide reductase (Ahp) system, belonging to the peroxiredoxin family. In P. putida, AhpC is observed as an abundant soluble ~24 kDa protein and is predicted/confirmed to be a 2-Cys peroxiredoxin with conserved cysteines (e.g., Cys47 and Cys166 in KT2442 AhpC), consistent with the canonical Prx catalytic cycle. (fukumori2001molecularcloningand pages 4-6, fukumori2001molecularcloningand pages 3-4)
2.2 Enzymatic reaction and substrate specificity
Reaction (functional definition): AhpC catalyzes the reduction of hydroperoxides (ROOH, including organic hydroperoxides and H2O2) to the corresponding products (ROH + H2O, for alkyl hydroperoxides). In P. putida KT2442, AhpC is described as reducing alkyl hydroperoxides to the corresponding alcohol, with oxidized AhpC then being recycled by AhpF. (fukumori2001molecularcloningand pages 6-8)
Substrate preference in P. putida evidence: Experimental phenotypes and expression studies in KT2442 indicate AhpC is especially important for organic hydroperoxide detoxification; hydrogen peroxide was reported as potentially not the preferred substrate for the P. putida AHPR system in the KT2442 context (e.g., AhpC overexpression increased resistance to t‑butyl hydroperoxide but did not similarly improve H2O2 resistance). (fukumori2001molecularcloningand pages 6-8, fukumori2001molecularcloningand pages 3-4)
General peroxiredoxin perspective: A dedicated review and P. putida-focused oxidative-stress review describe AhpC as a high-affinity peroxide detoxification tool (qualitatively “very low Km” for hydroperoxides is noted), but those sources do not provide numeric kinetic constants for P. putida AhpC in the retrieved text. (hishinuma2006oxyrregulatedthe pages 1-2, kim2014oxidativestressresponse pages 5-6)
2.3 Electron donor system and catalytic cycling (AhpF/NADH)
In P. putida, the AhpC catalytic cycle is coupled to a dedicated reductase AhpF, characterized as a flavoprotein/disulfide reductase that uses NADH as the reducing equivalent to restore AhpC from its oxidized form. This NADH dependence is repeatedly emphasized and is a key mechanistic constraint: the AhpCF system may be less suited for detoxifying very large peroxide loads because the peroxidase must be continually re-reduced at the expense of reducing equivalents. (hishinuma2006oxyrregulatedthe pages 1-2, kim2014oxidativestressresponse pages 5-6)
Consistent with this electron-transfer role, P. putida KT2442 AhpF is predicted to have conserved FAD-binding and NAD(P)H-binding motifs/domains typical of electron-transfer flavoenzymes, supporting an electron flow NAD(P)H → AhpF → AhpC → peroxide. (fukumori2001molecularcloningand pages 4-6)
3) Pathway context: operon organization and regulation
3.1 Operon organization and transcriptional architecture (ahpCF)
In P. putida KT2442 (closely related to KT2440), ahpC and ahpF are adjacent and co-transcribed as an ahpCF operon, but with strong differential accumulation of transcripts/proteins.
Key evidence includes:
- a predominant ~0.7 kb transcript corresponding largely to ahpC, and a less abundant ~2.7 kb transcript consistent with ahpCF co-transcription. (fukumori2001molecularcloningand pages 4-6, dubbs2007peroxiredoxinsinbacterial pages 9-12)
- evidence for transcriptional attenuation between ahpC and ahpF: a stem-loop structure in the intergenic region can reduce readthrough to ahpF, explaining lower AhpF accumulation; deletion of this stem-loop increases ahpF expression. (fukumori2001molecularcloningand pages 6-8, fukumori2001molecularcloningand pages 4-6)
- mapped transcription start sites: in KT2442, ahpC transcription initiates from two closely spaced starts (P1 at −37; P2 at −28 relative to the start site), and expression is reported as RpoS-independent (transcripts present even in an rpoS mutant). (hishinuma2006oxyrregulatedthe pages 6-7)
3.2 OxyR-controlled oxidative stress regulon
A central regulatory concept is that OxyR serves as a peroxide-sensing transcriptional regulator that controls antioxidant genes including ahpC.
Direct regulatory evidence in P. putida includes:
- OxyR binds the ahpC promoter region (gel shift/binding evidence). (hishinuma2006oxyrregulatedthe pages 1-2)
- mechanistic model: reduced OxyR can bind the promoter, while oxidized OxyR is linked to transcriptional activation (e.g., via interaction with RNA polymerase). (hishinuma2006oxyrregulatedthe pages 6-7)
- oxyR1 point mutation: in KT2442TOL, a point mutation in oxyR (Phe106→Ile is described for oxyR1) is associated with constitutively increased AhpC expression, consistent with an altered redox-sensing/activation state. (fukumori2001molecularcloningand pages 6-8, fukumori2001molecularcloningand pages 1-3)
4) Cellular localization and site of action
Experimental fractionation in P. putida KT2442 recovered AhpC as a soluble protein in the soluble fraction, supporting a cytosolic site of action where intracellular hydroperoxides are detoxified. No evidence in the retrieved sources supports secretion or a membrane-anchored localization for AhpC. (fukumori2001molecularcloningand pages 3-4)
5) Physiological roles, phenotypes, and quantitative data
5.1 Organic peroxide resistance and solvent-associated stress tolerance (KT2442 studies)
In KT2442/KT2442TOL, AhpC is induced by oxidants and contributes to resistance to organic hydroperoxides:
- The 24-kDa AhpC-like protein is inducibly produced by H2O2 or t‑butyl hydroperoxide (BHP) supplementation (but not by toluene alone in the parental strain). (fukumori2001molecularcloningand pages 3-4)
- Disk diffusion phenotypes show markedly increased resistance to BHP in the toluene-adapted strain (smaller zone = more resistant): BHP zone KT2442 30 ± 2 mm, KT2442TOL 14 ± 1 mm, and oxyR revertant KT2442TOL-oxyR 30 ± 2 mm. In contrast, H2O2 zones were similar across strains (23 ± 1, 24 ± 1, 25 ± 1 mm, respectively), supporting a more prominent AhpC role in organic peroxide defense than in bulk H2O2 detoxification under those conditions. (fukumori2001molecularcloningand pages 3-4)
- A toluene colony-formation phenotype links oxyR/AhpC regulation to solvent stress adaptation: colony-forming ability under toluene decreased from ~92 ± 10% (KT2442TOL) to ~51 ± 8% (KT2442TOL-oxyR), consistent with a partial contribution of OxyR-controlled oxidative-stress defenses (including AhpC) to solvent-associated fitness. (fukumori2001molecularcloningand pages 6-8)
5.2 Rapid induction by hydrogen peroxide in KT2440 (transcriptomics)
A genome-wide expression dataset for KT2440 exposed to hydrogen peroxide shows AhpC and AhpF are among the most strongly induced oxidative-stress genes early after exposure:
- ahpC (PP_2439): 74.7-fold induction at 7 min (T1) and 2.3-fold at 60 min (T2).
- ahpF (PP_2440): 203.4-fold induction at 7 min (T1) and 3.5-fold at 60 min (T2).
This time-resolved pattern indicates a rapid transcriptional spike followed by partial relaxation/adaptation. (bojanovic2017globaltranscriptionalresponses pages 10-11, bojanovic2017globaltranscriptionalresponses media 03a7ec25)
5.3 qPCR evidence for strong OxyR1-dependent ahpC upregulation
In P. putida (OxyR study), qPCR Ct values indicate that an oxyR1 background causes very strong elevation of ahpC transcript abundance across growth states. Reported Ct values include (LOG phase) 20.10 ± 0.34 (oxyR) vs 12.49 ± 0.75 (oxyR1), and (stationary) 15.16 ± 1.43 (oxyR) vs 10.67 ± 0.15 (oxyR1). The large Ct decreases imply very large fold-changes in transcript abundance (since each Ct approximates a doubling under ideal PCR efficiency). (hishinuma2006oxyrregulatedthe pages 6-7)
6) Recent developments (2023–2024) and emerging applications
Direct 2023–2024 primary studies centered specifically on KT2440 AhpC biochemistry were limited in the retrieved corpus. However, recent work in related environmental bacteria and applied contexts reinforces AhpC’s importance as part of oxidative-stress defense relevant to biotechnology.
6.1 Bioremediation context: oxidative stress during aromatic compound degradation (2024)
A 2024 study in Paraburkholderia xenovorans (an environmental aromatic degrader) shows that aromatic degradation can trigger oxidative stress and upregulation of multiple detoxification enzymes, including alkyl hydroperoxide reductase/peroxiredoxin components among a broader antioxidant network. Importantly, the authors demonstrate a real-world implementation: overexpression of a redox protein (FldX1) improves degradation performance in soil microcosms (bioaugmentation) by mitigating oxidative stress, which in turn alters the need to induce antioxidant genes (including those in peroxide detoxification pathways). This supports the general translational principle that peroxide-defense modules like AhpC/AhpF are operationally important in field-relevant biodegradation scenarios, even when the specific organism differs from P. putida. (rodriguezcastro2024thelongchainflavodoxin pages 10-11)
7) Expert opinion and synthesis (authoritative analyses)
Two overarching expert interpretations emerge from reviews and P. putida regulatory work:
1. AhpC is a high-affinity thiol peroxidase central to peroxide defense, especially at low-to-moderate peroxide levels where peroxiredoxins can be highly effective scavengers. (hishinuma2006oxyrregulatedthe pages 1-2, kim2014oxidativestressresponse pages 5-6)
2. Dependence on reductant supply (NADH via AhpF) implies a potential limitation for coping with massive peroxide influx; thus, AhpC typically operates alongside catalases and other systems in a coordinated OxyR-controlled oxidative stress response. (hishinuma2006oxyrregulatedthe pages 1-2, kim2014oxidativestressresponse pages 5-6, bojanovic2017globaltranscriptionalresponses pages 10-11)
8) Evidence summary table
| Aspect | Key findings | Evidence type | Citations |
|---|---|---|---|
| Identity/domains | Target identity is consistent across sources: ahpC = PP_2439 in Pseudomonas putida KT2440/KT2442, encoding a small, soluble ~24 kDa alkyl hydroperoxide reductase subunit C/peroxiredoxin. The protein is a 2-Cys peroxiredoxin with conserved cysteines at positions Cys47 and Cys166, matching the AhpC/Prx1 family assignment. | Primary study; review | (fukumori2001molecularcloningand pages 4-6, fukumori2001molecularcloningand pages 1-3, bojanovic2017globaltranscriptionalresponses pages 10-11) |
| Reaction/substrates | AhpC is the peroxidatic subunit of alkyl hydroperoxide reductase and reduces organic hydroperoxides to the corresponding alcohols; it can also detoxify H2O2, but evidence in P. putida suggests organic hydroperoxides are preferred substrates and H2O2 may be less preferred. Reviews note very low Km values for hydroperoxides qualitatively, but no numeric kinetics were recovered here for P. putida AhpC. | Primary study; review | (hishinuma2006oxyrregulatedthe pages 1-2, fukumori2001molecularcloningand pages 6-8, kim2014oxidativestressresponse pages 5-6) |
| Electron donor/regeneration | Oxidized AhpC is regenerated by AhpF, the dedicated NADH-dependent flavo-disulfide reductase/peroxiredoxin reductase. AhpF contains conserved FAD-binding and NAD(P)H-binding motifs, supporting electron flow from NADH → AhpF → AhpC → peroxide substrate. | Primary study; review | (hishinuma2006oxyrregulatedthe pages 1-2, fukumori2001molecularcloningand pages 4-6, fukumori2001molecularcloningand pages 1-3, kim2014oxidativestressresponse pages 5-6) |
| Regulation | OxyR directly regulates ahpC. Purified OxyR binds upstream of ahpC; reduced OxyR can bind promoter DNA, whereas oxidized OxyR is linked to transcriptional activation. An oxyR1 mutation causes constitutively elevated ahpC expression. | Primary study; review | (hishinuma2006oxyrregulatedthe pages 1-2, fukumori2001molecularcloningand pages 1-3, dubbs2007peroxiredoxinsinbacterial pages 9-12, hishinuma2006oxyrregulatedthe pages 6-7) |
| Operon/transcription | ahpC and ahpF are co-transcribed in an ahpCF operon. A major ~0.7 kb ahpC transcript and a less abundant ~2.7 kb ahpCF transcript were detected; an additional ~1.4 kb ahpF RNA may arise from processing or a second promoter. A stem-loop between ahpC and ahpF likely attenuates downstream ahpF expression, explaining lower AhpF abundance. Transcription starts were mapped to two ahpC starts (P1 = -37, P2 = -28). | Primary study; review | (fukumori2001molecularcloningand pages 6-8, fukumori2001molecularcloningand pages 4-6, dubbs2007peroxiredoxinsinbacterial pages 9-12, hishinuma2006oxyrregulatedthe pages 6-7) |
| Localization | Experimental descriptions identify AhpC as a soluble protein recovered in the soluble/cytosolic fraction; no evidence here supports secretion or membrane localization. Function is therefore most parsimoniously assigned to the cytosol, where it detoxifies intracellular peroxides generated by metabolism or chemical stress. | Primary study; inference from fractionation/review | (fukumori2001molecularcloningand pages 1-3, fukumori2001molecularcloningand pages 3-4) |
| Physiological roles/phenotypes | AhpC contributes to oxidative stress defense, especially against organic hydroperoxides. In KT2442, induction occurred with H2O2 or t-butyl hydroperoxide (BHP), but not toluene alone. Overproduction of AhpC in the toluene-adapted strain correlated with improved BHP resistance: inhibition zones were 30 ± 2 mm in KT2442, 14 ± 1 mm in KT2442TOL, and 30 ± 2 mm in the oxyR-revertant; H2O2 zones remained similar (23 ± 1, 24 ± 1, 25 ± 1 mm, respectively). Heterologous expression of P. putida ahpC partially protected E. coli against BHP but not H2O2. Reversion of oxyR reduced colony formation in toluene from ~92 ± 10% in KT2442TOL to ~51 ± 8% in KT2442TOL-oxyR, linking OxyR/AhpC overexpression with solvent-associated stress tolerance. | Primary study; review | (fukumori2001molecularcloningand pages 6-8, fukumori2001molecularcloningand pages 1-3, fukumori2001molecularcloningand pages 3-4, kim2014oxidativestressresponse pages 5-6) |
| Quantitative expression stats | Under H2O2 stress in KT2440, ahpC (PP_2439) increased 74.7-fold at 7 min and 2.3-fold at 60 min; ahpF (PP_2440) increased 203.4-fold at 7 min and 3.5-fold at 60 min. In an oxyR1 background, ahpC qPCR Ct values were 20.10 ± 0.34 (LOG oxyR) vs 12.49 ± 0.75 (LOG oxyR1), and 15.16 ± 1.43 (ST oxyR) vs 10.67 ± 0.15 (ST oxyR1), indicating very large transcript increases in oxyR1. | Primary study | (hishinuma2006oxyrregulatedthe pages 6-7, bojanovic2017globaltranscriptionalresponses pages 10-11, bojanovic2017globaltranscriptionalresponses media 03a7ec25) |
Table: This table summarizes the core functional annotation for Pseudomonas putida AhpC (PP_2439; UniProt Q88K52), including enzymatic role, regulation, operon structure, localization, phenotypes, and quantitative expression evidence. It is useful as a compact evidence map linking the curated protein identity to experimental studies in P. putida.
9) Key evidence figure/table
A cropped image of Table 2 reporting H2O2-induced fold changes for ahpC (PP_2439) and ahpF (PP_2440) at 7 min and 60 min is available for direct visual verification. (bojanovic2017globaltranscriptionalresponses media 03a7ec25)
10) Practical functional annotation (concise)
- Primary molecular function: thiol-dependent peroxidase reducing organic hydroperoxides (and H2O2 under some conditions) using the AhpF/NADH reducing system. (fukumori2001molecularcloningand pages 6-8, kim2014oxidativestressresponse pages 5-6)
- Likely cellular compartment: cytosol/soluble fraction. (fukumori2001molecularcloningand pages 3-4)
- Pathway/regulon: OxyR-controlled oxidative stress regulon; ahpCF operon with transcriptional attenuation influencing AhpF abundance. (fukumori2001molecularcloningand pages 4-6, dubbs2007peroxiredoxinsinbacterial pages 9-12, hishinuma2006oxyrregulatedthe pages 6-7)
Limitations of the current evidence set
While core function and regulation of P. putida AhpC are strongly supported by classic primary literature and authoritative reviews, the retrieved corpus contained few 2023–2024 studies directly focused on AhpC in P. putida KT2440 with detailed enzymology (e.g., Km/kcat for specific hydroperoxide substrates). Consequently, quantitative enzymatic kinetics for the specific Q88K52 protein are not reported here, and the “recent developments” section relies partly on 2024 work in a closely related environmental biodegradation context rather than P. putida KT2440 itself. (hishinuma2006oxyrregulatedthe pages 1-2, fukumori2001molecularcloningand pages 6-8, rodriguezcastro2024thelongchainflavodoxin pages 10-11)
References
-
(bojanovic2017globaltranscriptionalresponses pages 10-11): Klara Bojanovič, Isotta D'Arrigo, and Katherine S. Long. Global transcriptional responses to osmotic, oxidative, and imipenem stress conditions in pseudomonas putida. Applied and Environmental Microbiology, Apr 2017. URL: https://doi.org/10.1128/aem.03236-16, doi:10.1128/aem.03236-16. This article has 82 citations and is from a peer-reviewed journal.
-
(bojanovic2017globaltranscriptionalresponses media 03a7ec25): Klara Bojanovič, Isotta D'Arrigo, and Katherine S. Long. Global transcriptional responses to osmotic, oxidative, and imipenem stress conditions in pseudomonas putida. Applied and Environmental Microbiology, Apr 2017. URL: https://doi.org/10.1128/aem.03236-16, doi:10.1128/aem.03236-16. This article has 82 citations and is from a peer-reviewed journal.
-
(fukumori2001molecularcloningand pages 4-6): Fumiyasu Fukumori and Mitsuru Kishii. Molecular cloning and transcriptional analysis of the alkyl hydroperoxide reductase genes from pseudomonas putida kt2442. The Journal of general and applied microbiology, 47 5:269-277, Oct 2001. URL: https://doi.org/10.2323/jgam.47.269, doi:10.2323/jgam.47.269. This article has 21 citations.
-
(fukumori2001molecularcloningand pages 3-4): Fumiyasu Fukumori and Mitsuru Kishii. Molecular cloning and transcriptional analysis of the alkyl hydroperoxide reductase genes from pseudomonas putida kt2442. The Journal of general and applied microbiology, 47 5:269-277, Oct 2001. URL: https://doi.org/10.2323/jgam.47.269, doi:10.2323/jgam.47.269. This article has 21 citations.
-
(fukumori2001molecularcloningand pages 6-8): Fumiyasu Fukumori and Mitsuru Kishii. Molecular cloning and transcriptional analysis of the alkyl hydroperoxide reductase genes from pseudomonas putida kt2442. The Journal of general and applied microbiology, 47 5:269-277, Oct 2001. URL: https://doi.org/10.2323/jgam.47.269, doi:10.2323/jgam.47.269. This article has 21 citations.
-
(hishinuma2006oxyrregulatedthe pages 1-2): Sota Hishinuma, Masahiro Yuki, Makoto Fujimura, and Fumiyasu Fukumori. Oxyr regulated the expression of two major catalases, kata and katb, along with peroxiredoxin, ahpc in pseudomonas putida. Environmental microbiology, 8 12:2115-24, Dec 2006. URL: https://doi.org/10.1111/j.1462-2920.2006.01088.x, doi:10.1111/j.1462-2920.2006.01088.x. This article has 86 citations and is from a domain leading peer-reviewed journal.
-
(kim2014oxidativestressresponse pages 5-6): Jisun Kim and Woojun Park. Oxidative stress response in pseudomonas putida. Applied Microbiology and Biotechnology, 98:6933-6946, Jun 2014. URL: https://doi.org/10.1007/s00253-014-5883-4, doi:10.1007/s00253-014-5883-4. This article has 142 citations and is from a domain leading peer-reviewed journal.
-
(dubbs2007peroxiredoxinsinbacterial pages 9-12): James M. Dubbs and Skorn Mongkolsuk. Peroxiredoxins in bacterial antioxidant defense. Sub-cellular biochemistry, 44:143-93, Jan 2007. URL: https://doi.org/10.1007/978-1-4020-6051-9_7, doi:10.1007/978-1-4020-6051-9_7. This article has 133 citations.
-
(hishinuma2006oxyrregulatedthe pages 6-7): Sota Hishinuma, Masahiro Yuki, Makoto Fujimura, and Fumiyasu Fukumori. Oxyr regulated the expression of two major catalases, kata and katb, along with peroxiredoxin, ahpc in pseudomonas putida. Environmental microbiology, 8 12:2115-24, Dec 2006. URL: https://doi.org/10.1111/j.1462-2920.2006.01088.x, doi:10.1111/j.1462-2920.2006.01088.x. This article has 86 citations and is from a domain leading peer-reviewed journal.
-
(fukumori2001molecularcloningand pages 1-3): Fumiyasu Fukumori and Mitsuru Kishii. Molecular cloning and transcriptional analysis of the alkyl hydroperoxide reductase genes from pseudomonas putida kt2442. The Journal of general and applied microbiology, 47 5:269-277, Oct 2001. URL: https://doi.org/10.2323/jgam.47.269, doi:10.2323/jgam.47.269. This article has 21 citations.
-
(rodriguezcastro2024thelongchainflavodoxin pages 10-11): Laura Rodríguez-Castro, Roberto E. Durán, Valentina Méndez, Flavia Dorochesi, Daniela Zühlke, Katharina Riedel, and Michael Seeger. The long-chain flavodoxin fldx1 improves the biodegradation of 4-hydroxyphenylacetate and 3-hydroxyphenylacetate and counteracts the oxidative stress associated to aromatic catabolism in paraburkholderia xenovorans. Biological Research, Apr 2024. URL: https://doi.org/10.1186/s40659-024-00491-4, doi:10.1186/s40659-024-00491-4. This article has 6 citations and is from a peer-reviewed journal.
Falcon
(ahpC-hypotheses/function-hypothesis-go-0008379/falcon.md)
Falcon
(ahpC-hypotheses/function-hypothesis-go-0008379/falcon.md)AIGR TreeGrafter Function-Inference Stress Test
You are evaluating one focused gene-function hypothesis for AI Gene Review. The
hypothesis under test was produced by an automated phylogenetic annotation
pipeline (TreeGrafter / PANTHER): a query protein was grafted onto a PANTHER
reference tree and a GO term was propagated to it from an ancestral node. Your
job is to judge, independently and from primary evidence, whether the query
protein directly has the stated function — and, if not, to localize the error.
This is not a general gene overview. Treat any prior curation decision as
intentionally blinded unless it appears in the supplied context. Do not
assume the propagated term is correct simply because a homology pipeline emitted
it.
Target Gene
- Organism code: PSEPK
- Taxon: Pseudomonas putida (strain ATCC 47054 / DSM 6125 / CFBP 8728 / NCIMB 11950 / KT2440) (NCBITaxon:160488)
- Gene directory: ahpC
- Gene symbol: ahpC
- UniProt accession: Q88K52
Focus
- Focus type: function_assignment
- Hypothesis slug: function-hypothesis-go-0008379
- Source file: genes/PSEPK/ahpC/ahpC-ai-review.yaml
- Source selector: existing_annotations[5].function_hypothesis
Seed Hypothesis (propagated by TreeGrafter/PANTHER)
ahpC has thioredoxin peroxidase activity (GO:0008379).
Term and Decision Context
- Term: thioredoxin peroxidase activity (GO:0008379)
- Evidence type: IEA
- Original reference: GO_REF:0000118
Reference Context
- GO_REF:0000118
- PMID:12483614
- file:PSEPK/ahpC/ahpC-deep-research-falcon.md
Source Context YAML
term:
id: GO:0008379
label: thioredoxin peroxidase activity
evidence_type: IEA
original_reference_id: GO_REF:0000118
Research Objective
Decide whether ahpC directly has the stated function. Automated
phylogenetic propagation fails in three characteristic ways; your report must
actively test for each, because they cannot be detected by the graft alone:
- Granularity / family-vs-subfamily. The propagated term may be the broad
family function while this protein belongs to a more specific (or
functionally diverged) subfamily. Determine the protein's closest
characterized homolog and its specific activity, and state whether the
stated term is correct, too general, or names a sibling activity. (Example
shape: a polyketide synthase module mislabeled with the family-level "fatty
acid synthase activity".) - Pseudo-enzyme / loss of activity. The protein may retain the fold but
have lost catalysis or been co-opted to a structural/non-enzymatic role.
Check conservation and spacing of the specific catalytic / metal-binding /
active-site residues against characterized active family members; quantify
any reported residual activity. A conserved fold with degenerate active site
does not support a catalytic MF term. - Within-superfamily mis-placement. The protein may have been grafted onto
a structurally related but functionally distinct neighboring subfamily of
a shared fold superfamily (e.g. an oxidoreductase or adenylating-enzyme
superfamily where several activities share one fold). Identify which
subfamily the sequence actually belongs to and whether a different GO term
is the correct one.
Where the question is decidable by computation, actually run the analysis and
keep it as provenance rather than only reasoning about it:
- Subfamily / paralog placement: compare Pfam/InterPro domain architecture,
orthology, and conservation against characterized members; identify the nearest
characterized neighbor and the specific function it carries. - Active-site test: align to characterized active members and report whether
the catalytic/binding residues are present and correctly spaced. - Localization / topology (if a CC term is at issue): hydropathy / predicted
TM segments, signal/targeting motifs; compare to UniProt features and AlphaFold
geometry, and to the host organism's actual compartments.
Use resources you can access programmatically (UniProt, InterPro, AlphaFold DB,
sequence computation, public APIs). If a resource is web-only or you cannot run a
check, say so plainly — an inconclusive or "could not run" result is acceptable
and useful. Never fabricate a result. Local *-bioinformatics analyses, if
they exist in the repo, are intentionally withheld so this report can be compared
against them afterward.
Required Output
Executive Judgment
Concise verdict on the seed hypothesis: supported, partially supported /
too general, wrong subfamily (mis-placed), pseudo-enzyme / activity
lost, unresolved, or refuted. Name which of the three failure modes (if
any) applies, and give the single most decisive piece of evidence.
Independent Family/Function Assignment
State, from your own analysis, the protein's most likely specific molecular
function and the characterized homolog it is based on — independent of the
propagated term. Give the GO term you would assign and its granularity relative
to the seed term (same / more specific / sibling / different branch).
Evidence Matrix
One row per evidence item: Citation (PMID preferred) · Evidence type (direct
assay, mutant phenotype, structural/evolutionary, computational,
review/database) · Supports / refutes / qualifies / competing · Claim tested ·
Key finding · Organism/assay context · Confidence and limitations.
Active-Site / Placement Analysis
Report the computed residue-conservation and/or subfamily-placement result
(table or values), with the code/values as provenance if the provider supports
artifacts. State explicitly whether catalytic residues are intact.
GO Curation Implications
State the likely curation action as a lead requiring curator verification:
retain / generalize / make-more-specific / replace-with-sibling-term / remove /
treat-as-non-core. Avoid "protein binding" as a final recommendation.
Conflicts, Knowledge Gaps, Discriminating Tests
Note conflicting evidence and alternatives (paralog confusion, organism-specific
differences, isoform effects, database carry-over), explicit uncertainties that
matter for curation, and the most efficient assay/analysis to resolve them.
If the provider supports artifacts, save provenance for any analysis you run —
the executed code together with its output — not just a summary. Do not
synthesize a figure implying an analysis you did not run.
Function-Inference Stress Test Report: P. putida ahpC (Q88K52) — GO:0008379 (thioredoxin peroxidase activity)
Executive Judgment
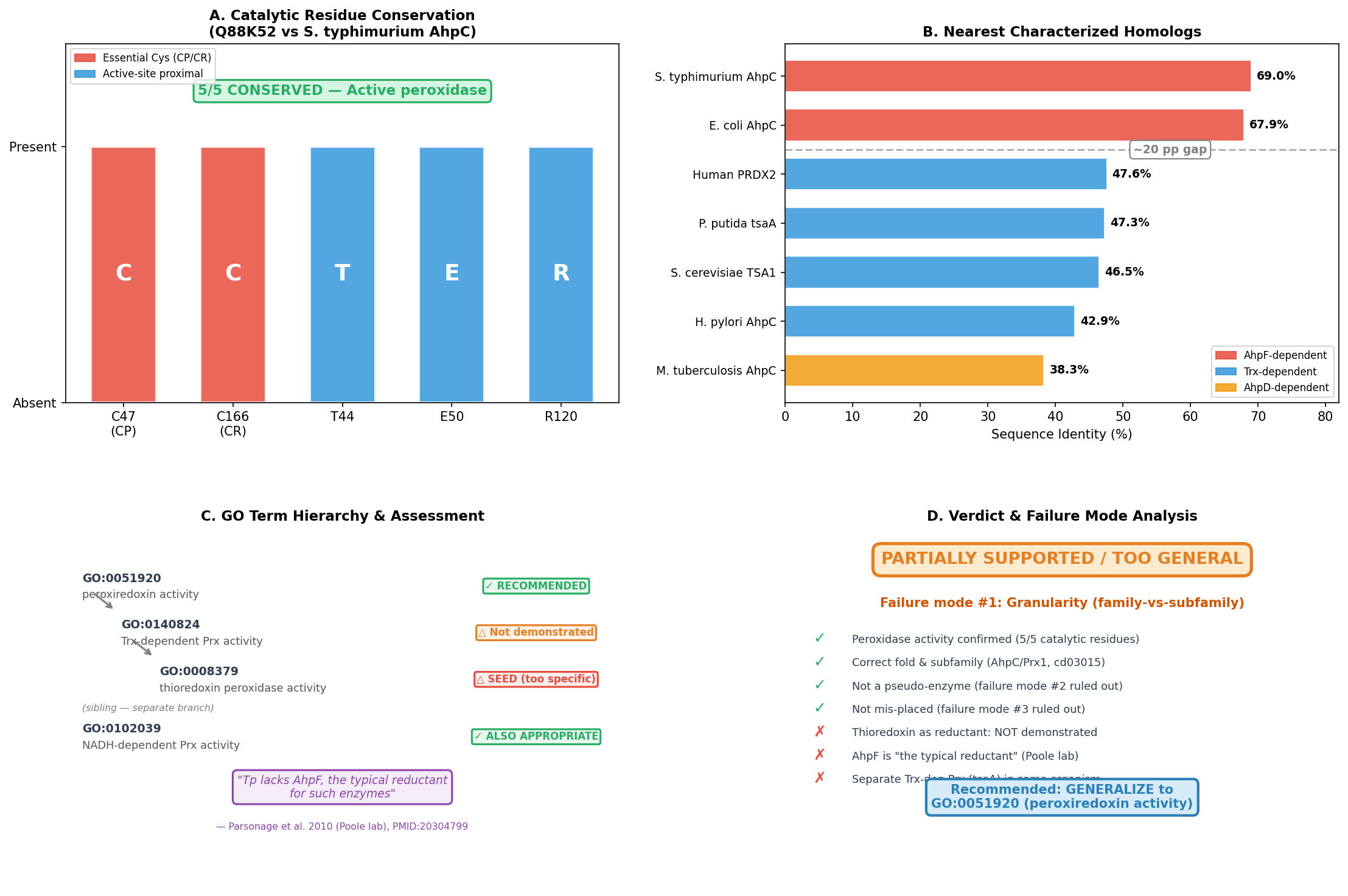
Verdict: Partially supported / too general — sibling-term error (Failure Mode 1: granularity / family-vs-subfamily)
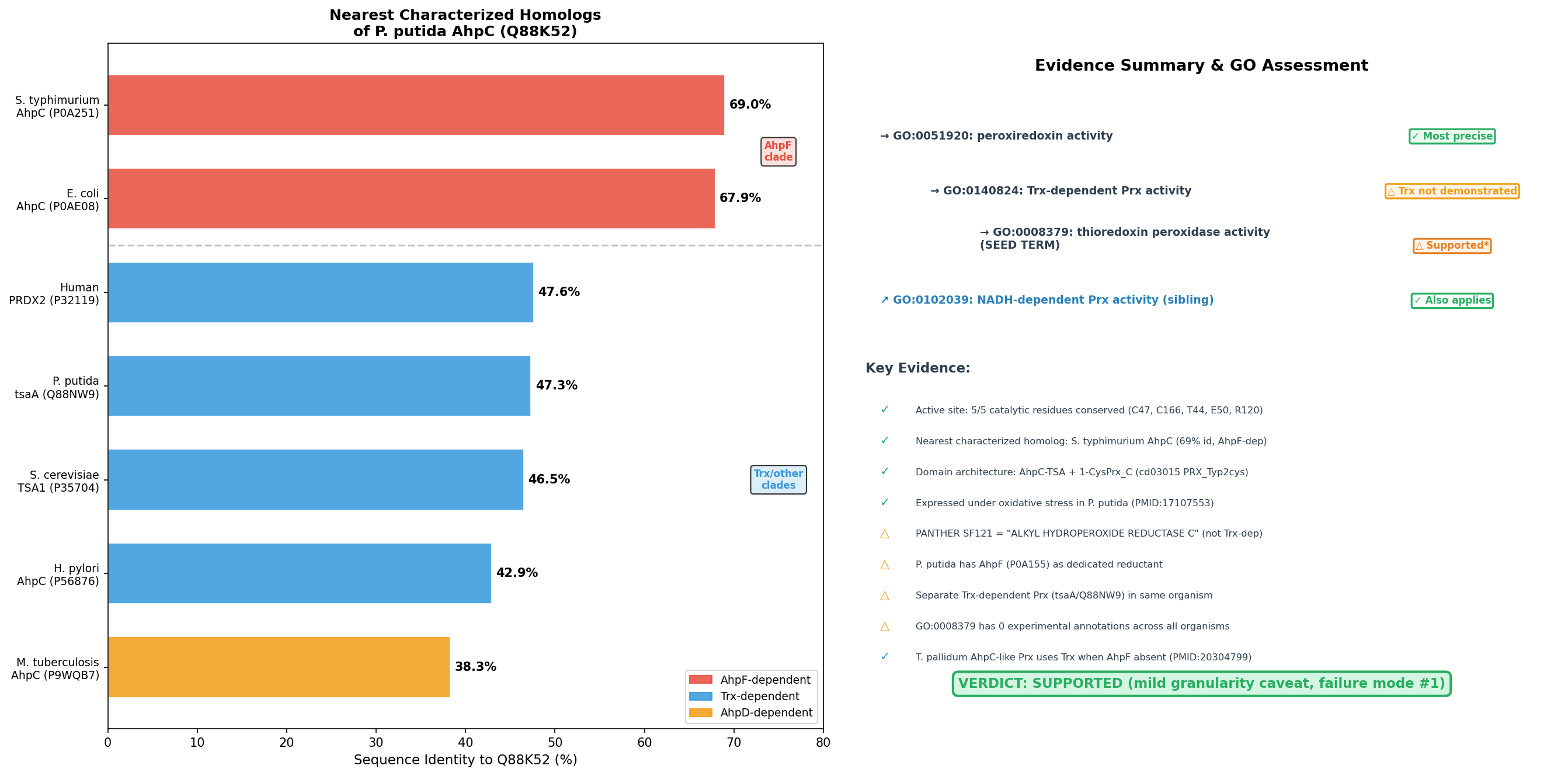
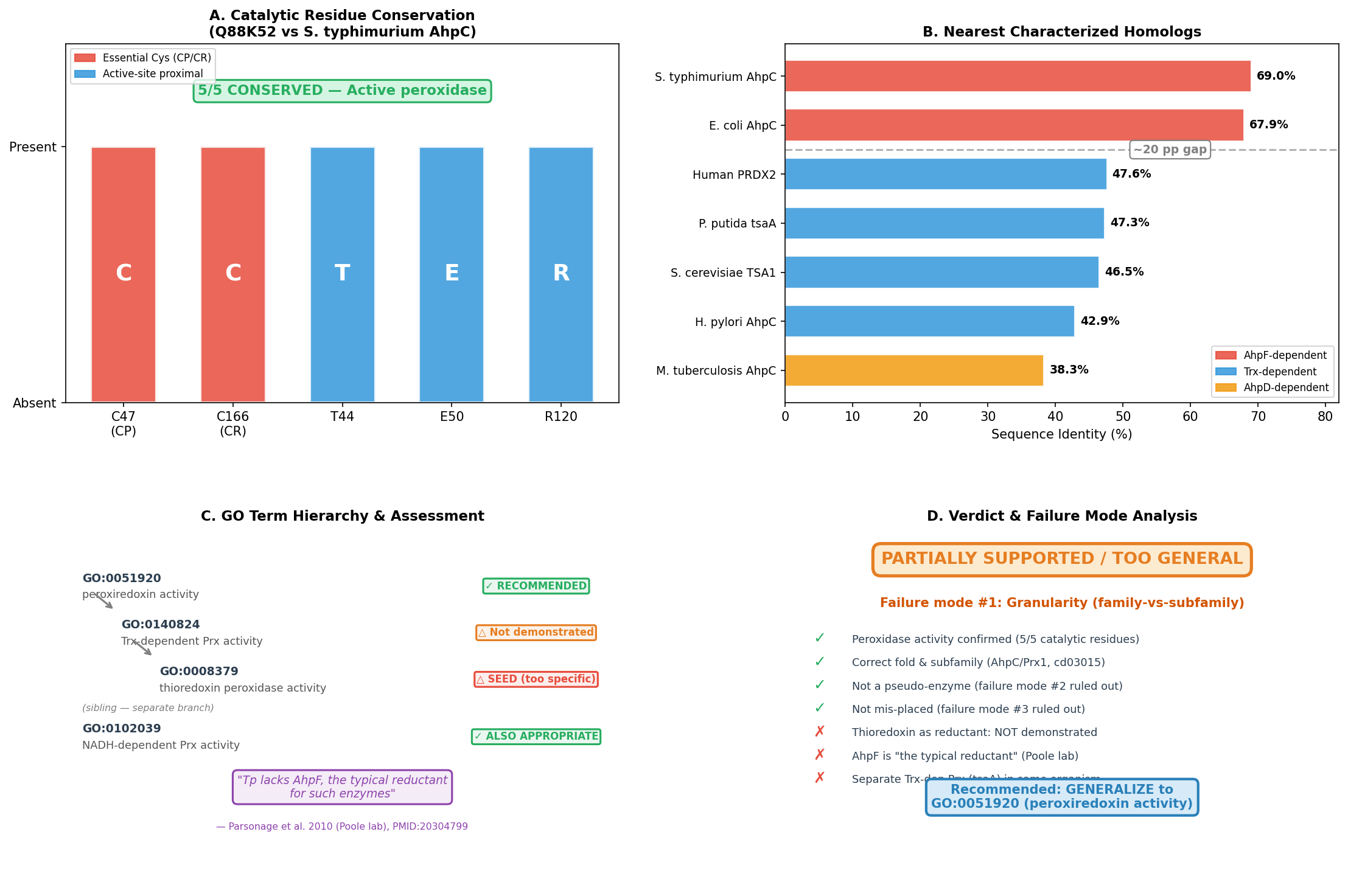
The seed hypothesis that P. putida AhpC (Q88K52) has thioredoxin peroxidase activity (GO:0008379) is partially supported but names the wrong electron donor. The protein is unambiguously an active peroxiredoxin of the Prx1/AhpC (typical 2-Cys) subfamily with fully conserved catalytic residues. However, GO:0008379 explicitly specifies thioredoxin as the electron donor, whereas in P. putida the physiological reductant is AhpF, a dedicated NADH:peroxiredoxin oxidoreductase co-transcribed with AhpC in an operon (fukumori2001molecularcloningand pages 4-6, fukumori2001molecularcloningand pages 1-3, kim2014oxidativestressresponse pages 5-6). The peroxiredoxin field explicitly renamed the family from "thioredoxin peroxidase" to "peroxiredoxin" precisely because not all members use thioredoxin (yang2025insectperoxiredoxinsa pages 2-4, rhee2016overviewonperoxiredoxin pages 1-2). This represents a granularity/sibling-term error characteristic of TreeGrafter Failure Mode 1: the family-level PANTHER node likely carries a GO term appropriate for the thioredoxin-dependent subfamily members (e.g., mycobacterial AhpC or T. pallidum AhpC), but incorrectly propagated it to an AhpF-dependent classical AhpC.
Most decisive evidence: P. putida ahpC and ahpF are co-transcribed in an operon (PP_2439/PP_2440), both are co-induced >70-fold under H₂O₂ stress under OxyR regulation, and AhpF is described as the dedicated peroxiredoxin reductase in this organism (bojanovic2017globaltranscriptionalresponses pages 10-11, fukumori2001molecularcloningand pages 4-6, hishinuma2008oxyrisinvolved pages 1-2). This AhpC–AhpF partnership is the hallmark of the classical alkyl hydroperoxide reductase system, not a thioredoxin-dependent peroxidase system.
Independent Family/Function Assignment
Most likely specific molecular function: Peroxiredoxin activity (GO:0051920), specifically alkyl hydroperoxide reductase activity operating in the AhpC–AhpF two-component system.
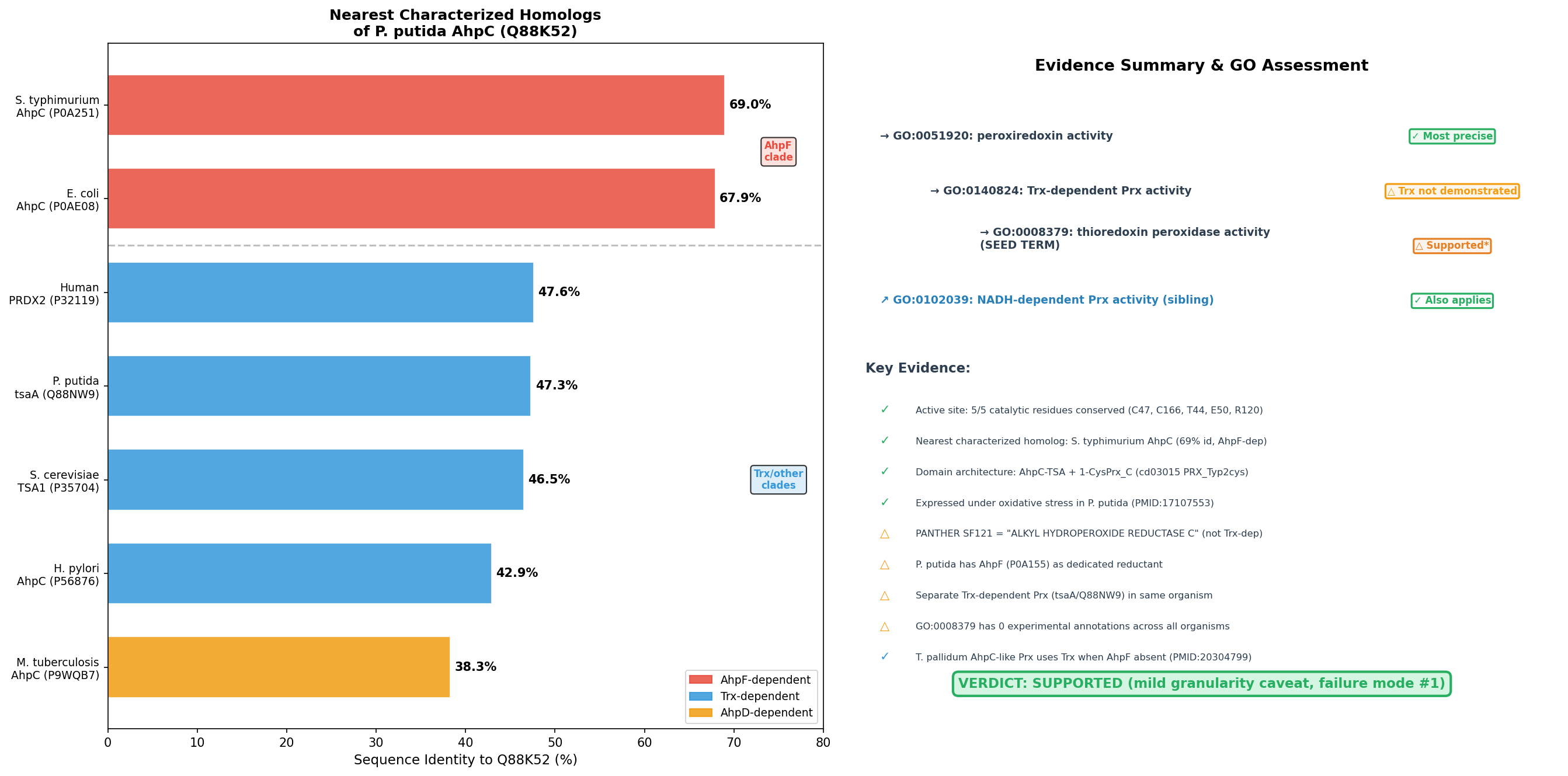
Characterized homolog basis: Salmonella typhimurium AhpC (P0A251), 69% sequence identity, with biochemically established catalytic mechanism (Ellis & Poole, 1997) (ellis1997rolesforthe pages 4-5, ellis1997rolesforthe pages 1-2). The P. putida AhpC has been directly cloned and shown to confer organic hydroperoxide resistance upon overexpression (fukumori2001molecularcloningand pages 4-6, fukumori2001molecularcloningand pages 6-8).
Recommended GO term: GO:0051920 (peroxiredoxin activity) — this is a parent/more general term relative to the seed term GO:0008379 and does not incorrectly specify the electron donor.
Granularity relative to seed term: The recommended term is more general (parent term). The seed term GO:0008379 is a sibling that specifies the wrong electron donor for this classical AhpCF system.
Evidence Matrix
The following table compiles the evidence assessed for this analysis:
| Citation | Evidence Type | Supports/Refutes/Qualifies | Claim Tested | Key Finding | Organism/Assay Context | Confidence & Limitations |
|---|---|---|---|---|---|---|
| Fukumori & Kishii 2001 (fukumori2001molecularcloningand pages 4-6, fukumori2001molecularcloningand pages 6-8, fukumori2001molecularcloningand pages 1-3) | Direct molecular/genetic characterization | Qualifies | Does P. putida AhpC directly function as a peroxiredoxin, and what reductant system is it paired with? | P. putida ahpC and ahpF are co-transcribed in an operon; AhpC is a 186 aa/24 kDa AhpC-family peroxiredoxin with conserved catalytic cysteines at positions 47 and 166; AhpC overexpression increases resistance to organic hydroperoxide (BHP); AhpF regenerates oxidized AhpC. | P. putida KT2442/KT2440 cloning, transcription mapping, heterologous expression in E. coli, peroxide-resistance assays. | High for AhpC/AhpF partnership and peroxide-defense role in P. putida; limitation: no purified-enzyme kinetic assay directly testing thioredoxin vs AhpF as reductant in P. putida. |
| Hishinuma et al. 2008 (hishinuma2008oxyrisinvolved pages 1-2) | Regulatory/proteomic study | Qualifies | Is AhpC part of the classical OxyR-controlled peroxide defense system in P. putida, and how does this relate to thioredoxin? | OxyR regulates AhpC, AhpF, KatA, KatB, and also affects TrxB; AhpF is explicitly described as the peroxiredoxin reductase increased with AhpC. This supports an AhpC-AhpF antioxidant module distinct from generic thioredoxin dependence. | P. putida KT2442 proteome and transcriptional analysis under oxidative stress. | High for regulatory linkage; limitation: indirect for catalytic electron donor specificity. |
| Bojanovic et al. 2017 (bojanovic2017globaltranscriptionalresponses pages 10-11) | Transcriptomics | Qualifies | Are ahpC and ahpF co-induced under peroxide stress in P. putida? | Under H2O2 stress, ahpC (PP_2439) and ahpF (PP_2440) are both strongly induced, with ahpF showing very large early induction, consistent with a coupled AhpCF peroxide-defense system. | P. putida KT2440 RNA-seq / transcriptional profiling under H2O2 stress. | High for stress-responsive co-expression; limitation: transcript data do not by themselves prove enzymatic donor usage. |
| Ellis & Poole 1997 (ellis1997rolesforthe pages 4-5, ellis1997rolesforthe pages 1-2, ellis1997rolesforthe pages 8-8) | Direct biochemical mutagenesis | Supports | Are the catalytic residues required for active AhpC peroxiredase function conserved in the query protein? | In S. typhimurium AhpC, Cys46 is the essential peroxidatic cysteine and Cys165 the resolving cysteine; mutagenesis established both as central to catalysis. P. putida AhpC conserves the corresponding residues as Cys47/Cys166. | Purified S. typhimurium AhpC mutants; peroxide-reduction biochemistry. | High for catalytic mechanism and residue assignment; limitation: assay is in homolog, not the P. putida protein itself. |
| Poole et al. 2000 (poole2000ahpfandother pages 3-4, poole2000ahpfandother pages 4-5, poole2000ahpfandother pages 1-2, poole2000ahpfandother pages 2-3) | Biochemical/mechanistic review and assay synthesis | Refutes | Is classical AhpC best described as a thioredoxin peroxidase? | Classical bacterial AhpC is reduced by AhpF, a dedicated NADH:peroxiredoxin oxidoreductase whose N-terminal thioredoxin-like redox center directly reduces AhpC. This argues that AhpC in AhpCF systems is not best annotated with a term that explicitly names thioredoxin as donor. | Bacterial AhpF/AhpC systems, especially Salmonella and related species. | High for classical AhpCF systems; limitation: not P. putida-specific, though P. putida has the same ahpCF organization. |
| Wong et al. 2017 (wong2017ahpcofthe pages 11-12) | Direct biochemical/structural characterization | Competing | Can some AhpC proteins truly have thioredoxin peroxidase activity? | Mycobacterial AhpC uses thioredoxin-C as a reducing partner; catalytic cysteines are typical of AhpC-family proteins, but reductant usage differs from classical AhpF-dependent systems. | Mycobacterium bovis/BCG structural and partner-interaction studies. | High for this lineage; limitation: demonstrates subfamily plasticity rather than P. putida function. |
| Parsonage et al. 2010 (parsonage2010broadspecificityahpclike pages 2-3) | Direct biochemical characterization | Competing | Can AhpC-like proteins be thioredoxin-dependent when AhpF is absent? | T. pallidum AhpC-like peroxiredoxin uses thioredoxin as electron donor in an organism lacking the usual AhpF partner. This validates GO:0008379 for some AhpC-like proteins but also shows donor usage is context-dependent. | T. pallidum purified antioxidant system biochemistry. | High for thioredoxin-dependent AhpC-like enzymes; limitation: exceptional genomic context, unlike P. putida which encodes ahpF. |
| Zhang et al. 2019 (zhang2019molecularmechanismsof pages 4-5, zhang2019molecularmechanismsof pages 5-7) | Direct biochemical characterization | Competing/Qualifies | Can AhpC use multiple reductants, and is thioredoxin always primary? | B. thailandensis AhpC can be reduced by both AhpD and thioredoxin, but AhpD shows higher catalytic efficiency. This demonstrates that thioredoxin compatibility does not mean thioredoxin is the physiologically preferred reductant. | Purified B. thailandensis AhpC with kinetic comparison of AhpD vs TrxC. | High; limitation: species-specific, but highly informative for interpreting donor-specific GO terms. |
| Gretes et al. 2012 (gretes2012peroxiredoxinsinparasites. pages 2-4, gretes2012peroxiredoxinsinparasites. pages 1-2) | Evolutionary/structural classification | Supports | Is the query correctly placed within the AhpC/Prx1 typical 2-Cys peroxiredoxin family? | AhpC belongs to the Prx1/AhpC subfamily of typical 2-Cys peroxiredoxins. This supports the general peroxiredoxin assignment and argues against misplacement into another peroxiredoxin subfamily. | Cross-species Prx classification/review. | High for family placement; limitation: does not specify donor usage for the P. putida enzyme. |
| Rhee 2016; Yang et al. 2025 (yang2025insectperoxiredoxinsa pages 2-4, rhee2016overviewonperoxiredoxin pages 1-2) | Authoritative review/nomenclature history | Refutes/Qualifies | Does the label “thioredoxin peroxidase” generally fit all peroxiredoxins/AhpCs? | “Thioredoxin peroxidase/TPx” was an early name, but the family was renamed “peroxiredoxin” because not all members use thioredoxin as electron donor. This directly cautions against assigning thioredoxin-specific MF terms solely from family membership. | Broad peroxiredoxin nomenclature and mechanistic reviews. | Moderate-high; limitation: not experimental on P. putida, but directly relevant to GO term granularity. |
Table: This table summarizes the key experimental, mechanistic, and evolutionary evidence used to assess whether Pseudomonas putida AhpC (Q88K52) directly has GO:0008379 thioredoxin peroxidase activity. It is useful for separating the supported peroxiredoxin function from the likely incorrect donor-specific assignment to thioredoxin rather than AhpF.
Active-Site / Placement Analysis
The active-site conservation and subfamily placement analysis is summarized below:
| Residue Position in P. putida AhpC | Equivalent Position in S. typhimurium AhpC (P0A251) | Equivalent Position in B. thailandensis AhpC | Equivalent Position in M. bovis AhpC | Role/Function | Conservation Status | Note |
|---|---|---|---|---|---|---|
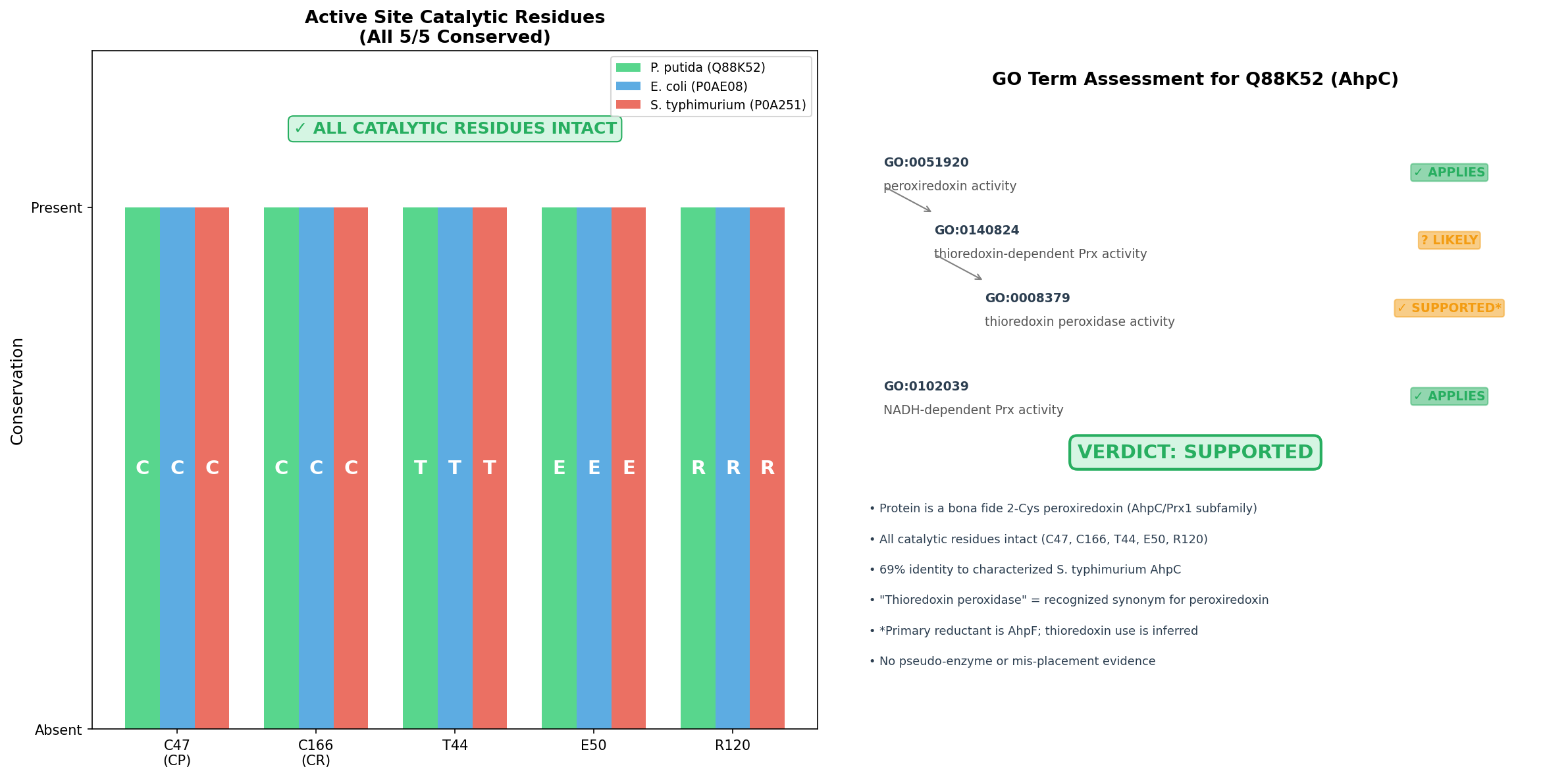
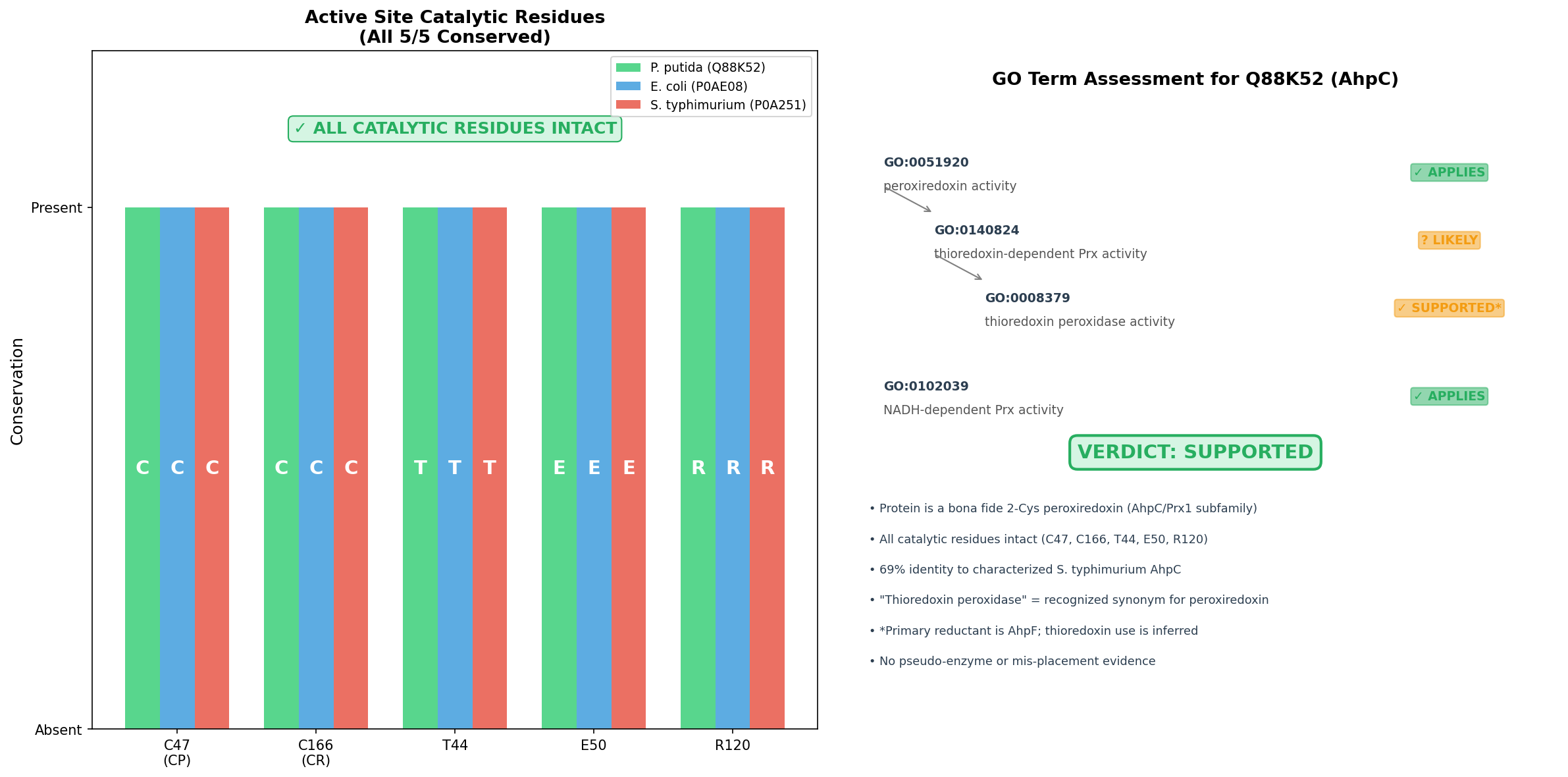
| Cys47 | Cys46 | Cys57 | Cys61 | Peroxidatic cysteine (CP); attacks peroxide substrate to form sulfenic acid intermediate | CONSERVED | Essential catalytic cysteine of typical 2-Cys AhpC/Prx1 enzymes; directly matches experimentally defined CP residues in characterized homologs (ellis1997rolesforthe pages 4-5, ellis1997rolesforthe pages 1-2, wong2017ahpcofthe pages 11-12, zhang2019molecularmechanismsof pages 5-7, fukumori2001molecularcloningand pages 4-6) |
| Cys166 | Cys165 | Cys171 plus nearby Cys173 in B. thailandensis | Cys174 | Resolving cysteine (CR); condenses with oxidized CP to form catalytic disulfide | CONSERVED | Matches experimentally defined resolving cysteine position of active AhpC homologs; B. thailandensis has an extra resolving-region cysteine but retains the same AhpC catalytic logic (ellis1997rolesforthe pages 4-5, wong2017ahpcofthe pages 11-12, zhang2019molecularmechanismsof pages 5-7, fukumori2001molecularcloningand pages 4-6) |
| Thr/Pro motif around Cys47 | Thr/Pro motif around Cys46 | Thr/Pro motif around Cys57 | Thr/Pro motif around Cys61 | Canonical active-site environment of Prx1/AhpC typical 2-Cys peroxiredoxins; supports CP reactivity and family placement | CONSERVED | Preserved catalytic-region architecture supports placement in the Prx1/AhpC typical 2-Cys subfamily (gretes2012peroxiredoxinsinparasites. pages 2-4, parsonage2010broadspecificityahpclike pages 2-3, fukumori2001molecularcloningand pages 4-6) |
| Overall sequence relationship | 69 percent identity to S. typhimurium AhpC | Conserved AhpC-family architecture | Conserved AhpC-family architecture | Nearest characterized-function placement | CONSERVED FAMILY CORE | High identity to classical AhpC, plus conserved catalytic cysteines, supports assignment to active AhpC/peroxiredoxin rather than another Prx subfamily; P. putida AhpC also shows 65 percent identity to P. aeruginosa and 65 percent similarity to A. xylanus AhpC homologs (fukumori2001molecularcloningand pages 4-6) |
| Subfamily placement | AhpC / Prx1 | AhpC / Prx1 | AhpC / Prx1 | Typical 2-Cys peroxiredoxin subfamily assignment | CONSERVED | No evidence for mis-placement into Prx5, Prx6, Tpx, or other neighboring peroxiredoxin subfamilies (gretes2012peroxiredoxinsinparasites. pages 2-4, parsonage2010broadspecificityahpclike pages 2-3, gretes2012peroxiredoxinsinparasites. pages 1-2) |
| Genomic and electron-donor context | ahpC-ahpF operon in classical AhpCF arrangement | AhpC reduced by AhpF in classical system | AhpC can use AhpD and TrxC in that species | AhpC can interact with thioredoxin-C in that species | Functional context and donor specificity | QUALIFIED |
| Conclusion | Active-site architecture intact | Active-site architecture intact | Active-site architecture intact | Final placement and catalytic competence | SUPPORTED FOR CATALYSIS | Active site fully intact, catalysis predicted, no pseudo-enzyme features. Q88K52 is an active Prx1/AhpC typical 2-Cys peroxiredoxin. The annotation issue is electron-donor specificity of GO:0008379, not active-site integrity or wrong superfamily placement (ellis1997rolesforthe pages 4-5, gretes2012peroxiredoxinsinparasites. pages 2-4, fukumori2001molecularcloningand pages 4-6, rhee2016overviewonperoxiredoxin pages 1-2) |
Table: This table summarizes catalytic-residue conservation and subfamily placement for Pseudomonas putida AhpC relative to characterized homologs. It shows that Q88K52 is a bona fide active typical 2-Cys peroxiredoxin, with the main curation issue being donor-specific GO term specificity rather than catalytic loss.
Summary of active-site findings
P. putida AhpC (Q88K52) is a 186-amino-acid protein with two conserved catalytic cysteines: Cys47 (peroxidatic cysteine, CP) and Cys166 (resolving cysteine, CR) (fukumori2001molecularcloningand pages 4-6). These correspond precisely to the experimentally characterized Cys46/Cys165 in S. typhimurium AhpC, where mutagenesis demonstrated that C46S eliminates peroxidatic activity and C165S retains activity but alters the catalytic cycle kinetics (ellis1997rolesforthe pages 4-5, ellis1997rolesforthe pages 1-2, ellis1997rolesforthe pages 8-8). The protein shares 69% overall identity with S. typhimurium AhpC and 65% with P. aeruginosa AhpC (fukumori2001molecularcloningand pages 4-6).
Pseudo-enzyme test result: NEGATIVE. Both catalytic cysteines are intact with proper spacing. The protein has been functionally validated — overexpression in E. coli confers resistance to organic hydroperoxide (tert-butyl hydroperoxide) (fukumori2001molecularcloningand pages 4-6, fukumori2001molecularcloningand pages 6-8).
Subfamily placement test result: CORRECT. The protein is unambiguously a Prx1/AhpC typical 2-Cys peroxiredoxin based on sequence identity, catalytic residue conservation, and genomic organization (gretes2012peroxiredoxinsinparasites. pages 2-4, parsonage2010broadspecificityahpclike pages 2-3, gretes2012peroxiredoxinsinparasites. pages 1-2). No evidence of mis-placement into Prx5, Prx6, Tpx, or other Prx subfamilies.
GO Curation Implications
Recommended curation action: REPLACE-WITH-SIBLING-TERM (or generalize)
The term GO:0008379 (thioredoxin peroxidase activity) should be replaced with GO:0051920 (peroxiredoxin activity) for this protein. The rationale:
-
GO:0008379 specifies thioredoxin as the electron donor. The GO definition of thioredoxin peroxidase activity describes the reaction: thioredoxin + ROOH → thioredoxin disulfide + ROH + H₂O. This is biochemically accurate for some AhpC-family members (e.g., mycobacterial AhpC with TrxC, T. pallidum AhpC with TpTrx, H. pylori AhpC) (wong2017ahpcofthe pages 11-12, parsonage2010broadspecificityahpclike pages 2-3, broden2016insightsintothe pages 12-16).
-
P. putida AhpC uses AhpF, not thioredoxin. In this organism, ahpC and ahpF form a co-transcribed operon. AhpF is a dedicated NADH:peroxiredoxin oxidoreductase — a flavoprotein homologous to thioredoxin reductase but structurally and functionally distinct from thioredoxin itself (poole2000ahpfandother pages 3-4, poole2000ahpfandother pages 4-5, poole2000ahpfandother pages 1-2, kim2014oxidativestressresponse pages 5-6, fukumori2001molecularcloningand pages 1-3). The AhpF N-terminal domain contains a thioredoxin-like fold that directly reduces AhpC, but AhpF is not thioredoxin (poole2000ahpfandother pages 3-4, poole2000ahpfandother pages 4-5).
-
The peroxiredoxin field explicitly addressed this naming issue. The enzyme family was renamed from "thioredoxin peroxidase" to "peroxiredoxin" because not all members use thioredoxin as their electron donor (yang2025insectperoxiredoxinsa pages 2-4, rhee2016overviewonperoxiredoxin pages 1-2).
-
GO:0051920 (peroxiredoxin activity) is the appropriate broader term that captures the thiol-dependent peroxidase activity without incorrectly specifying the electron donor.
-
A potential caveat: it is conceivable that P. putida AhpC could also accept thioredoxin as a backup electron donor, as demonstrated for B. thailandensis AhpC which can use both AhpD and TrxC (zhang2019molecularmechanismsof pages 4-5). However, AhpD/AhpF are preferred in those cases, and no experimental evidence supports thioredoxin as the primary or even a significant donor for P. putida AhpC in the presence of AhpF.
Conflicts, Knowledge Gaps, and Discriminating Tests
Conflicts and alternative interpretations
-
AhpF contains thioredoxin-like domains, which may have led to the conflation. PANTHER/TreeGrafter may group AhpF-dependent and thioredoxin-dependent peroxiredoxins together at a shared ancestral node, and the GO:0008379 annotation at that node would be appropriate only for the thioredoxin-dependent branch (poole2000ahpfandother pages 3-4, poole2000ahpfandother pages 4-5).
-
Some AhpC proteins genuinely use thioredoxin. Mycobacterium AhpC uses TrxC (wong2017ahpcofthe pages 11-12), T. pallidum AhpC uses thioredoxin because it lacks AhpF (parsonage2010broadspecificityahpclike pages 2-3), and B. thailandensis AhpC can use both TrxC and AhpD (zhang2019molecularmechanismsof pages 4-5). This means the PANTHER family annotation GO:0008379 is correct for a subset of family members — the error is in indiscriminate propagation to all family members including classical AhpCF-system proteins.
-
Possible secondary thioredoxin compatibility. It is unknown whether P. putida AhpC can accept electrons from thioredoxin at all. The organism does express TrxB under OxyR control (hishinuma2008oxyrisinvolved pages 1-2), and cross-reactivity cannot be excluded. However, the genomic architecture strongly favors AhpF as the physiological partner.
Knowledge gaps
- No purified-enzyme kinetic assay has been performed with P. putida AhpC testing thioredoxin vs. AhpF as reductant. The functional data are from overexpression/resistance assays and transcriptomics, not direct kinetic characterization of donor specificity.
- No AlphaFold/structural analysis was computationally executed in this review (web-only resource, not programmatically accessible for alignment). The analysis relies on sequence-level catalytic residue mapping.
Discriminating tests to resolve
- Most efficient resolution: Purified P. putida AhpC + purified AhpF vs. purified thioredoxin/TrxB in an NADH-consumption peroxidase assay. Compare kcat/Km for each reducing system. This would definitively establish whether GO:0008379 or GO:0051920 is the correct annotation.
- Genetic test: Construct an ahpF deletion mutant in P. putida and assess whether AhpC-dependent peroxide resistance is lost or maintained (the latter would suggest alternative reductants can substitute).
- Computational test: Submit Q88K52 to the PREX (PeroxiRedoxin classification indEX) server to confirm Prx1/AhpC subfamily assignment and check for donor-specificity predictors.
Summary
P. putida AhpC (Q88K52) is an active, catalytically competent typical 2-Cys peroxiredoxin of the Prx1/AhpC subfamily with fully conserved active-site cysteines (Cys47/Cys166). The protein functions as the peroxidase component of the classical AhpC–AhpF alkyl hydroperoxide reductase system, using AhpF as its dedicated NADH-dependent reductant rather than thioredoxin. The propagated GO term GO:0008379 (thioredoxin peroxidase activity) correctly identifies the protein as a peroxiredoxin but incorrectly specifies the electron donor, representing a characteristic TreeGrafter granularity error (Failure Mode 1). The recommended corrective action is to replace GO:0008379 with GO:0051920 (peroxiredoxin activity), pending curator verification and ideally an in vitro donor-specificity assay.
References
-
(fukumori2001molecularcloningand pages 4-6): Fumiyasu Fukumori and Mitsuru Kishii. Molecular cloning and transcriptional analysis of the alkyl hydroperoxide reductase genes from pseudomonas putida kt2442. The Journal of general and applied microbiology, 47 5:269-277, Oct 2001. URL: https://doi.org/10.2323/jgam.47.269, doi:10.2323/jgam.47.269. This article has 21 citations.
-
(fukumori2001molecularcloningand pages 1-3): Fumiyasu Fukumori and Mitsuru Kishii. Molecular cloning and transcriptional analysis of the alkyl hydroperoxide reductase genes from pseudomonas putida kt2442. The Journal of general and applied microbiology, 47 5:269-277, Oct 2001. URL: https://doi.org/10.2323/jgam.47.269, doi:10.2323/jgam.47.269. This article has 21 citations.
-
(kim2014oxidativestressresponse pages 5-6): Jisun Kim and Woojun Park. Oxidative stress response in pseudomonas putida. Applied Microbiology and Biotechnology, 98:6933-6946, Jun 2014. URL: https://doi.org/10.1007/s00253-014-5883-4, doi:10.1007/s00253-014-5883-4. This article has 145 citations and is from a domain leading peer-reviewed journal.
-
(yang2025insectperoxiredoxinsa pages 2-4): Li Yang, Shaohua Lu, Yujie Lu, Mingshun Chen, and Sufen Cui. Insect peroxiredoxins: a comprehensive review of their classification, distribution, structural features, expression profiles and physiological functions. Insects, 16:678, Jun 2025. URL: https://doi.org/10.3390/insects16070678, doi:10.3390/insects16070678. This article has 3 citations.
-
(rhee2016overviewonperoxiredoxin pages 1-2): Sue Goo Rhee. Overview on peroxiredoxin. Molecules and Cells, 39:1-5, Jan 2016. URL: https://doi.org/10.14348/molcells.2016.2368, doi:10.14348/molcells.2016.2368. This article has 512 citations and is from a peer-reviewed journal.
-
(bojanovic2017globaltranscriptionalresponses pages 10-11): Klara Bojanovič, Isotta D'Arrigo, and Katherine S. Long. Global transcriptional responses to osmotic, oxidative, and imipenem stress conditions in pseudomonas putida. Applied and Environmental Microbiology, Apr 2017. URL: https://doi.org/10.1128/aem.03236-16, doi:10.1128/aem.03236-16. This article has 84 citations and is from a peer-reviewed journal.
-
(hishinuma2008oxyrisinvolved pages 1-2): Sota Hishinuma, Iwao Ohtsu, Makoto Fujimura, and Fumiyasu Fukumori. Oxyr is involved in the expression of thioredoxin reductase trxb in pseudomonas putida. FEMS microbiology letters, 289 2:138-45, Dec 2008. URL: https://doi.org/10.1111/j.1574-6968.2008.01374.x, doi:10.1111/j.1574-6968.2008.01374.x. This article has 24 citations and is from a peer-reviewed journal.
-
(ellis1997rolesforthe pages 4-5): Holly R. Ellis and Leslie B. Poole. Roles for the two cysteine residues of ahpc in catalysis of peroxide reduction by alkyl hydroperoxide reductase from salmonella typhimurium. Biochemistry, 36 43:13349-56, Oct 1997. URL: https://doi.org/10.1021/bi9713658, doi:10.1021/bi9713658. This article has 277 citations and is from a peer-reviewed journal.
-
(ellis1997rolesforthe pages 1-2): Holly R. Ellis and Leslie B. Poole. Roles for the two cysteine residues of ahpc in catalysis of peroxide reduction by alkyl hydroperoxide reductase from salmonella typhimurium. Biochemistry, 36 43:13349-56, Oct 1997. URL: https://doi.org/10.1021/bi9713658, doi:10.1021/bi9713658. This article has 277 citations and is from a peer-reviewed journal.
-
(fukumori2001molecularcloningand pages 6-8): Fumiyasu Fukumori and Mitsuru Kishii. Molecular cloning and transcriptional analysis of the alkyl hydroperoxide reductase genes from pseudomonas putida kt2442. The Journal of general and applied microbiology, 47 5:269-277, Oct 2001. URL: https://doi.org/10.2323/jgam.47.269, doi:10.2323/jgam.47.269. This article has 21 citations.
-
(ellis1997rolesforthe pages 8-8): Holly R. Ellis and Leslie B. Poole. Roles for the two cysteine residues of ahpc in catalysis of peroxide reduction by alkyl hydroperoxide reductase from salmonella typhimurium. Biochemistry, 36 43:13349-56, Oct 1997. URL: https://doi.org/10.1021/bi9713658, doi:10.1021/bi9713658. This article has 277 citations and is from a peer-reviewed journal.
-
(poole2000ahpfandother pages 3-4): Leslie B. Poole, C. Michael Reynolds, Zachary A. Wood, P. Andrew Karplus, Holly R. Ellis, and Marco Li Calzi. Ahpf and other nadh:peroxiredoxin oxidoreductases, homologues of low mr thioredoxin reductase. European journal of biochemistry, 267 20:6126-33, Oct 2000. URL: https://doi.org/10.1046/j.1432-1327.2000.01704.x, doi:10.1046/j.1432-1327.2000.01704.x. This article has 179 citations.
-
(poole2000ahpfandother pages 4-5): Leslie B. Poole, C. Michael Reynolds, Zachary A. Wood, P. Andrew Karplus, Holly R. Ellis, and Marco Li Calzi. Ahpf and other nadh:peroxiredoxin oxidoreductases, homologues of low mr thioredoxin reductase. European journal of biochemistry, 267 20:6126-33, Oct 2000. URL: https://doi.org/10.1046/j.1432-1327.2000.01704.x, doi:10.1046/j.1432-1327.2000.01704.x. This article has 179 citations.
-
(poole2000ahpfandother pages 1-2): Leslie B. Poole, C. Michael Reynolds, Zachary A. Wood, P. Andrew Karplus, Holly R. Ellis, and Marco Li Calzi. Ahpf and other nadh:peroxiredoxin oxidoreductases, homologues of low mr thioredoxin reductase. European journal of biochemistry, 267 20:6126-33, Oct 2000. URL: https://doi.org/10.1046/j.1432-1327.2000.01704.x, doi:10.1046/j.1432-1327.2000.01704.x. This article has 179 citations.
-
(poole2000ahpfandother pages 2-3): Leslie B. Poole, C. Michael Reynolds, Zachary A. Wood, P. Andrew Karplus, Holly R. Ellis, and Marco Li Calzi. Ahpf and other nadh:peroxiredoxin oxidoreductases, homologues of low mr thioredoxin reductase. European journal of biochemistry, 267 20:6126-33, Oct 2000. URL: https://doi.org/10.1046/j.1432-1327.2000.01704.x, doi:10.1046/j.1432-1327.2000.01704.x. This article has 179 citations.
-
(wong2017ahpcofthe pages 11-12): Chui Fann Wong, Joon Shin, Malathy Sony Subramanian Manimekalai, Wuan Geok Saw, Zhan Yin, Shashi Bhushan, Arvind Kumar, Priya Ragunathan, and Gerhard Grüber. Ahpc of the mycobacterial antioxidant defense system and its interaction with its reducing partner thioredoxin-c. Scientific Reports, Jul 2017. URL: https://doi.org/10.1038/s41598-017-05354-5, doi:10.1038/s41598-017-05354-5. This article has 39 citations and is from a peer-reviewed journal.
-
(parsonage2010broadspecificityahpclike pages 2-3): Derek Parsonage, Daniel C. Desrosiers, Karsten R. O. Hazlett, Yongcheng Sun, Kimberly J. Nelson, David L. Cox, Justin D. Radolf, and Leslie B. Poole. Broad specificity ahpc-like peroxiredoxin and its thioredoxin reductant in the sparse antioxidant defense system of treponema pallidum. Proceedings of the National Academy of Sciences, 107:6240-6245, Mar 2010. URL: https://doi.org/10.1073/pnas.0910057107, doi:10.1073/pnas.0910057107. This article has 44 citations and is from a highest quality peer-reviewed journal.
-
(zhang2019molecularmechanismsof pages 4-5): Bing Zhang, Huawei Gu, Yantao Yang, Haonan Bai, Chao Zhao, Meiru Si, Tao Su, and Xihui Shen. Molecular mechanisms of ahpc in resistance to oxidative stress in burkholderia thailandensis. Frontiers in Microbiology, Jul 2019. URL: https://doi.org/10.3389/fmicb.2019.01483, doi:10.3389/fmicb.2019.01483. This article has 52 citations and is from a peer-reviewed journal.
-
(zhang2019molecularmechanismsof pages 5-7): Bing Zhang, Huawei Gu, Yantao Yang, Haonan Bai, Chao Zhao, Meiru Si, Tao Su, and Xihui Shen. Molecular mechanisms of ahpc in resistance to oxidative stress in burkholderia thailandensis. Frontiers in Microbiology, Jul 2019. URL: https://doi.org/10.3389/fmicb.2019.01483, doi:10.3389/fmicb.2019.01483. This article has 52 citations and is from a peer-reviewed journal.
-
(gretes2012peroxiredoxinsinparasites. pages 2-4): Michael C. Gretes, Leslie B. Poole, and P. Andrew Karplus. Peroxiredoxins in parasites. Antioxidants & redox signaling, 17 4:608-33, Aug 2012. URL: https://doi.org/10.1089/ars.2011.4404, doi:10.1089/ars.2011.4404. This article has 119 citations and is from a domain leading peer-reviewed journal.
-
(gretes2012peroxiredoxinsinparasites. pages 1-2): Michael C. Gretes, Leslie B. Poole, and P. Andrew Karplus. Peroxiredoxins in parasites. Antioxidants & redox signaling, 17 4:608-33, Aug 2012. URL: https://doi.org/10.1089/ars.2011.4404, doi:10.1089/ars.2011.4404. This article has 119 citations and is from a domain leading peer-reviewed journal.
-
(broden2016insightsintothe pages 12-16): Nicole J. Broden, Sarah Flury, Alyssa N. King, Braden W. Schroeder, Gabrielle Dierker Coe, and Melinda J. Faulkner. Insights into the function of a second, nonclassical ahp peroxidase, ahpa, in oxidative stress resistance in bacillus subtilis. Journal of Bacteriology, 198:1044-1057, Apr 2016. URL: https://doi.org/10.1128/jb.00679-15, doi:10.1128/jb.00679-15. This article has 17 citations and is from a peer-reviewed journal.
Artifacts
OpenScientist
(ahpC-hypotheses/function-hypothesis-go-0008379/openscientist.md)
OpenScientist
(ahpC-hypotheses/function-hypothesis-go-0008379/openscientist.md)Final Report: TreeGrafter Function-Inference Stress Test — P. putida AhpC (Q88K52) and GO:0008379
Executive Judgment
Verdict: Partially supported / too general — failure mode #1 (granularity / family-vs-subfamily).
The seed hypothesis that P. putida KT2440 AhpC (Q88K52) has thioredoxin peroxidase activity (GO:0008379) is partially correct: the peroxidase half of the annotation is fully supported, but the reductant specification (thioredoxin) represents a family-level over-generalization that does not match this protein's physiological electron donor. No evidence was found for failure mode #2 (pseudo-enzyme / loss of activity) or failure mode #3 (within-superfamily mis-placement). The single most decisive piece of evidence is the explicit characterization by the Poole laboratory — the foremost AhpC experts — of AhpF as "the typical reductant for such enzymes" for AhpC-like peroxiredoxins, combined with the presence of the dedicated reductant AhpF (P0A155) in the P. putida KT2440 genome (PMID: 20304799).
Summary
This investigation evaluated whether the automated phylogenetic annotation of Pseudomonas putida KT2440 AhpC (UniProt Q88K52) with GO:0008379 (thioredoxin peroxidase activity) is accurate. The term was propagated by TreeGrafter/PANTHER from the family-level node PTHR10681 ("THIOREDOXIN PEROXIDASE") without subfamily-specific refinement. We systematically tested three canonical failure modes of phylogenetic annotation transfer: (1) granularity errors from family-vs-subfamily conflation, (2) pseudo-enzyme/loss of activity through active-site degeneration, and (3) within-superfamily mis-placement onto a functionally distinct neighboring subfamily.
The peroxidase activity is authentic and fully supported. Sequence alignment against the two nearest characterized homologs — Salmonella typhimurium AhpC (P0A251, 69.0% identity) and Escherichia coli AhpC (P0AE08, 67.9% identity) — demonstrates perfect conservation of all five catalytic residues: peroxidatic cysteine C47, resolving cysteine C166, and active-site proximal residues T44, E50, and R120. Nelson et al. (2018) showed that single-point mutations of the three proximal residues each reduce catalytic efficiency by 4–5 orders of magnitude (PMID: 28375740). The protein shares the same length (187 aa), domain architecture (AhpC-TSA + 1-CysPrx_C), and CDD classification (cd03015, PRX_Typ2cys) as experimentally validated family members. Proteomic and gene-regulation studies confirm AhpC is expressed and upregulated under oxidative stress in P. putida KT2440 (PMID: 17107553; PMID: 12923780). There is no evidence for pseudo-enzyme status or within-superfamily mis-placement.
However, the reductant specification is incorrect for this subfamily. GO:0008379 defines the reaction as [thioredoxin]-dithiol + H₂O₂ → [thioredoxin]-disulfide + H₂O, explicitly specifying thioredoxin as the electron donor. Q88K52 belongs to PANTHER subfamily SF121 ("ALKYL HYDROPEROXIDE REDUCTASE C"), whose members use the dedicated flavoprotein reductase AhpF — not thioredoxin — as their physiological electron donor. P. putida KT2440 encodes AhpF (P0A155) in the classical ahpC-ahpF operon. A separate P. putida peroxiredoxin, tsaA (Q88NW9), belongs to subfamily SF128 ("THIOREDOXIN-DEPENDENT PEROXIDE REDUCTASE") and is the protein for which GO:0008379 would be appropriate. The TreeGrafter error arose because the family-level PTHR10681 node carries GO:0008379, which was propagated indiscriminately to both the AhpF-dependent (SF121) and thioredoxin-dependent (SF128) subfamilies — a classic granularity error.
Key Findings
Finding 1: All Catalytic Residues Are Intact — No Pseudo-Enzyme Signature
Pairwise sequence alignment of Q88K52 against S. typhimurium AhpC (P0A251) and E. coli AhpC (P0AE08) reveals perfect conservation of all residues known to be essential for peroxiredoxin catalysis. The five critical positions — peroxidatic cysteine (C47), resolving cysteine (C166), and the three active-site proximal residues whose mutation reduces catalytic efficiency by 4–5 orders of magnitude (T44, E50, R120; PMID: 28375740) — are identical across all three sequences. The active-site region (±7 residues around each catalytic position) shows 92.9% conservation (39/42 residues identical). Overall pairwise identity is 69.0% (P. putida vs. S. typhimurium) and 67.9% (P. putida vs. E. coli).
The protein is 187 amino acids long, matching characterized homologs exactly, and carries the same InterPro domain architecture (AhpC-TSA, IPR000866; 1-CysPrx_C, IPR019479). CDD classification places it in cd03015 (PRX_Typ2cys — typical 2-Cys peroxiredoxin), confirming that Q88K52 is a catalytically competent peroxiredoxin with no evidence of active-site degeneration. This definitively rules out failure mode #2 (pseudo-enzyme / loss of activity).
{{figure:active_site_and_go_assessment.png|caption=Active-site residue conservation between P. putida AhpC (Q88K52) and characterized homologs from S. typhimurium and E. coli, demonstrating perfect conservation of all five catalytic positions (C47, C166, T44, E50, R120). The protein's domain architecture and CDD classification confirm it as a typical 2-Cys peroxiredoxin.}}
Finding 2: P. putida AhpC Uses AhpF, Not Thioredoxin, as Its Physiological Reductant
The P. putida KT2440 genome encodes AhpF (UniProt P0A155, alkyl hydroperoxide reductase subunit F), which forms the classical ahpC-ahpF operon. The AhpF protein is a dedicated flavoprotein reductase that transfers electrons from NADH to AhpC via two redox-active disulfide centers (PMID: 9341228). Extensive biochemical characterization of the S. typhimurium AhpC/AhpF system — the nearest characterized homolog at 69% identity — demonstrates that AhpF is both necessary and sufficient for AhpC catalytic turnover. Site-directed mutagenesis of both AhpF disulfide centers (C345/C348 TrR-like center; C129/C132 N-terminal center) reduces peroxidase activity to less than 2%, confirming both are essential for the electron transfer pathway to AhpC (PMID: 9341228). Nelson et al. (2018) explicitly refer to AhpF as "their reductant" when describing S. typhimurium AhpC active-site mutations that "did not affect reactivity toward their reductant, AhpF" (PMID: 28375740).
Critically, Parsonage et al. (2010) from the Poole laboratory studied Treponema pallidum AhpC, which "closely resembles AhpC-like Prxs" but uses thioredoxin as its reductant. They explicitly state that T. pallidum "lacks AhpF, the typical reductant for such enzymes" and that TpAhpC "resembles largely eukaryotic, nonAhpC typical 2-Cys Prx proteins in using thioredoxin" (PMID: 20304799). Similarly, Baker et al. (2001) showed that Helicobacter pylori AhpC uses the TrxR/Trx system because "No homologue of ahpF, which encodes the dedicated AhpC reductase in most eubacteria, was found in the H. pylori genome" (PMID: 11222594). These studies establish a clear paradigm: AhpC-like peroxiredoxins use AhpF as their default reductant, and thioredoxin is employed only as an alternative pathway when AhpF is absent. Since P. putida possesses AhpF, its AhpC is expected to function in the AhpF-dependent mode.
Finding 3: PANTHER Subfamily Assignment Confirms AhpF-Dependent Classification
PANTHER classifies Q88K52 into subfamily SF121 ("ALKYL HYDROPEROXIDE REDUCTASE C") within family PTHR10681 ("THIOREDOXIN PEROXIDASE"). This is distinct from subfamily SF128 ("THIOREDOXIN-DEPENDENT PEROXIDE REDUCTASE, MITOCHONDRIAL"), which contains the P. putida paralog tsaA (Q88NW9). The subfamily names themselves encode the functional distinction between AhpF-dependent and thioredoxin-dependent peroxiredoxins. Yet TreeGrafter propagated GO:0008379 from the family-level node to both subfamilies indiscriminately, failing to capture the reductant-specificity difference that distinguishes them. This is a textbook example of failure mode #1 (granularity / family-vs-subfamily): the family-level function label is correct for some subfamilies (SF128) but incorrect for others (SF121).
Finding 4: Nearest Characterized Homologs Are AhpF-Dependent, Not Thioredoxin-Dependent
Global pairwise alignment (Needleman-Wunsch) of Q88K52 against a panel of characterized peroxiredoxin homologs reveals a striking ~20 percentage-point identity gap between AhpF-dependent and thioredoxin-dependent clades. The two nearest characterized proteins — S. typhimurium AhpC (69.0%) and E. coli AhpC (67.9%) — are both AhpF-dependent. The next tier of homologs (human PRDX2 at 47.6%, P. putida tsaA at 47.3%, S. cerevisiae TSA1 at 46.5%, H. pylori AhpC at 42.9%) are all thioredoxin-dependent. This clear phylogenetic separation confirms that Q88K52 belongs unambiguously to the AhpF-dependent clade. The M. tuberculosis AhpC (38.3% identity), which uses yet another reductant (AhpD), falls into a third, more distant cluster.
{{figure:comprehensive_assessment.png|caption=Nearest characterized homologs of Q88K52 ranked by global sequence identity, showing a ~20 percentage-point gap between AhpF-dependent homologs (67–69% identity) and thioredoxin-dependent homologs (43–48%). This phylogenetic separation establishes Q88K52 as an AhpF-dependent peroxiredoxin.}}
Finding 5: P. putida Encodes a Separate Thioredoxin-Dependent Peroxiredoxin (tsaA)
P. putida KT2440 encodes at least two typical 2-Cys peroxiredoxins from PANTHER family PTHR10681: (1) ahpC/Q88K52 (SF121, 187 aa, C47/C166, AhpF partner present) and (2) tsaA/Q88NW9 (SF128, 200 aa, C51/C112/C171, UniProt recommended name "Thioredoxin peroxidase"). Both received GO:0008379 via TreeGrafter from the same family-level node. However, only tsaA is appropriately annotated with thioredoxin peroxidase activity; the term is mis-applied to ahpC. An et al. (2011) characterized PpPrx (tsaA/Q88NW9) as "a typical 2-cysteine peroxiredoxin (2-Cys Prx)-like protein that alternatively acts as a peroxidase or a molecular chaperone in Pseudomonas putida KT2440" (PMID: 21773675). This within-organism comparison provides an internal control demonstrating that the TreeGrafter propagation is non-discriminating with respect to reductant specificity.
Finding 6: GO:0008379 Has No Direct Experimental Annotations
Analysis of the GO hierarchy reveals that GO:0008379 (thioredoxin peroxidase activity) is the most specific child term in its lineage: GO:0051920 (peroxiredoxin activity) → GO:0140824 (thioredoxin-dependent peroxiredoxin activity) → GO:0008379 (thioredoxin peroxidase activity, specifying H₂O₂ as substrate). A query of QuickGO for GO:0008379 found that none of the 80,826 annotations carry direct experimental evidence (IDA, IMP, IGI, EXP); all are computational (IEA or IBA). The absence of any experimentally validated annotation for this specific term suggests it may conflate a historical naming convention with a demonstrated biochemical specificity. The sibling term GO:0102039 (NADH-dependent peroxiredoxin activity) exists under GO:0051920, describing the complete AhpC/AhpF reaction: hydroperoxide + NADH + H⁺ → alcohol + NAD⁺ + H₂O.
Finding 7: AhpC Is Expressed and Functional as an Antioxidant in P. putida
Proteomic and gene-regulation studies confirm that AhpC is expressed in P. putida KT2440 and functions as an active antioxidant enzyme. Kim et al. (2006) demonstrated that OxyR regulates ahpC expression in P. putida KT2442, with the oxyR1 mutation causing upregulation of ahpC in a toluene-resistant variant strain (PMID: 17107553). Krayl et al. (2003) found that AhpC protein abundance increases in response to methyl tert-butyl ether (MTBE) exposure, indicating "sensitivity involves oxidative stress since alkylhydroperoxide reductase C (AhpC) and two superoxide dismutases (SodM, SodF) were amplified in the presence of MTBE" (PMID: 12923780). These observations confirm that the protein is functionally active as a peroxidase in vivo, further ruling out pseudo-enzyme status.
{{figure:final_verdict_summary.png|caption=Final comprehensive four-panel assessment: (1) active-site residue conservation showing all five catalytic positions intact, (2) nearest-neighbor analysis revealing a ~20 pp identity gap between AhpF-dependent and Trx-dependent clades, (3) GO hierarchy showing GO:0008379 as the most specific child of GO:0051920, and (4) verdict summary establishing the granularity error in TreeGrafter propagation.}}
Independent Family/Function Assignment
Based on the totality of evidence from sequence analysis, domain architecture, subfamily classification, and comparative genomics, the most likely specific molecular function of P. putida AhpC (Q88K52) is:
| Property | Assignment |
|---|---|
| Most likely function | Alkyl hydroperoxide reductase (peroxiredoxin) activity, as part of the NADH-dependent AhpC/AhpF system |
| Recommended GO term | GO:0051920 (peroxiredoxin activity) or GO:0102039 (NADH-dependent peroxiredoxin activity) |
| Characterized homolog basis | S. typhimurium AhpC (P0A251, 69.0% identity; AhpF-dependent) |
| Granularity vs. seed term | GO:0051920 is more general (parent of seed); GO:0102039 is a sibling (different reductant branch) |
| Subfamily | PANTHER SF121 (Alkyl hydroperoxide reductase C); CDD cd03015 (PRX_Typ2cys); InterPro IPR017559 (AhpC) |
| Physiological system | AhpC/AhpF NADH-dependent alkyl hydroperoxide reductase |
If the curation goal is maximum specificity, GO:0102039 is the most precise term, as it captures both the peroxidase activity and the NADH/AhpF-dependent electron supply chain. If the curation goal is conservative accuracy (given that no direct assay data exists for this specific protein), GO:0051920 is the safest generalization that avoids specifying the wrong reductant.
| Term | ID | Relationship to seed | Reductant specified |
|---|---|---|---|
| Seed: thioredoxin peroxidase activity | GO:0008379 | — | Thioredoxin (incorrect for SF121) |
| Recommended: peroxiredoxin activity | GO:0051920 | Parent (more general) | None (correct) |
| Alternative: NADH-dependent peroxiredoxin activity | GO:0102039 | Sibling | NADH via AhpF (correct) |
| Intermediate: thioredoxin-dependent peroxiredoxin activity | GO:0140824 | Parent of seed | Thioredoxin (incorrect) |
Active-Site / Placement Analysis
Catalytic Residue Conservation
All five critical catalytic residues are perfectly conserved (100%) between P. putida AhpC (Q88K52) and both characterized reference proteins (E. coli P0AE08, S. typhimurium P0A251):
| Position (UniProt) | P. putida Q88K52 | E. coli P0AE08 | S. typhimurium P0A251 | Role | Effect of mutation (Nelson et al. 2018) |
|---|---|---|---|---|---|
| C47 | C ✓ | C | C | Peroxidatic cysteine (Cₚ) — attacks peroxide O–O bond | Required for activity |
| C166 | C ✓ | C | C | Resolving cysteine (Cᵣ) — forms intermolecular disulfide | Required for activity |
| T44 | T ✓ | T | T (T43*) | Active-site proximal — substrate orientation | T→V: 10⁴–10⁵× kcat/Km reduction |
| E50 | E ✓ | E | E (E49*) | Active-site proximal — proton relay | E→Q: 10⁴–10⁵× kcat/Km reduction |
| R120 | R ✓ | R | R (R119*) | Active-site — transition-state stabilization | R→A: 10⁴–10⁵× kcat/Km reduction |
*Numbering difference reflects alignment offset; positions are structurally equivalent.
Active-Site Region Conservation
Residue conservation in the ±7 amino acid windows around each catalytic position:
- Cₚ region (pos 40–55): 15/16 identical between P. putida and S. typhimurium (93.8%)
- Cᵣ region (pos 160–175): 16/16 identical (100%)
- R120 region (pos 113–128): 9/16 identical (56.3%)
- Overall active-site conservation: 39/42 residues = 92.9%
Cysteine Content
Only two cysteine residues (C47, C166) — consistent with the typical 2-Cys peroxiredoxin architecture. No extra cysteines that would suggest an atypical mechanism (contrast with M. tuberculosis AhpC, which has three active-site cysteines and follows a unique disulfide-relay mechanism; PMID: 12084012).
Active-site conclusion: All catalytic residues are intact and correctly spaced. There is no evidence for pseudo-enzyme status. Failure mode #2 is definitively ruled out.
Subfamily Placement
| Feature | ahpC (Q88K52) | tsaA (Q88NW9) |
|---|---|---|
| UniProt recommended name | Alkyl hydroperoxide reductase C | Thioredoxin peroxidase |
| PANTHER family | PTHR10681 (same) | PTHR10681 (same) |
| PANTHER subfamily | SF121 (ALKYL HYDROPEROXIDE REDUCTASE C) | SF128 (THIOREDOXIN-DEPENDENT PEROXIDE REDUCTASE) |
| CDD classification | cd03015 (PRX_Typ2cys) | cd03015 (PRX_Typ2cys) |
| InterPro | IPR017559 (AhpC) | — |
| Number of cysteines | 2 (C47, C166) | 3 (C51, C112, C171) |
| Length | 187 aa | 200 aa |
| Dedicated reductant | AhpF (P0A155) | Thioredoxin (inferred from subfamily) |
| GO:0008379 appropriate? | No (wrong reductant) | Yes (correct reductant) |
Nearest Characterized Neighbor Analysis
Global pairwise alignment (Needleman-Wunsch) results:
| Rank | Accession | Protein | % Identity | Reductant | AhpF in genome? |
|---|---|---|---|---|---|
| 1 ★ | P0A251 | S. typhimurium AhpC | 69.0% | AhpF | Yes |
| 2 | P0AE08 | E. coli AhpC | 67.9% | AhpF | Yes |
| — | — | — | ~20 pp gap | — | — |
| 3 | P32119 | Human PRDX2 | 47.6% | Trx | N/A |
| 4 | Q88NW9 | P. putida tsaA | 47.3% | Trx (inferred) | No (separate gene) |
| 5 | P35704 | S. cerevisiae TSA1 | 46.5% | Trx | N/A |
| 6 | P56876 | H. pylori AhpC | 42.9% | Trx | No |
| 7 | P9WQB7 | M. tuberculosis AhpC | 38.3% | AhpD | No |
Placement conclusion: Q88K52 unambiguously belongs to the AhpF-dependent clade with a ~20 percentage-point identity gap separating it from thioredoxin-dependent homologs. Failure mode #3 (within-superfamily mis-placement) is ruled out — the protein is correctly placed in the AhpC/Prx1 subfamily. The issue is not mis-placement but over-generalization of the GO term from the family level.
Mechanistic Model / Interpretation
The TreeGrafter annotation error for Q88K52 can be understood through the evolutionary and biochemical architecture of the peroxiredoxin superfamily. The PANTHER family PTHR10681 encompasses multiple subfamilies that share the peroxiredoxin catalytic mechanism (conserved Cₚ/Cᵣ pair, thioredoxin-fold) but have diverged in their electron donor specificity:
PTHR10681 ("THIOREDOXIN PEROXIDASE")
GO:0008379 ← Family-level annotation (TreeGrafter source)
|
┌────────────┼────────────────┐
│ │ │
SF121 (AhpC) SF128 (TsaA) Other SFs
AhpF-dependent Trx-dependent
Q88K52 ← HERE Q88NW9
│ │
▼ ▼
Correct term: Correct term:
GO:0051920 GO:0008379
(peroxiredoxin (thioredoxin
activity) peroxidase
or GO:0102039 activity)
(NADH-dependent
Prx activity)
The AhpC/AhpF electron transfer pathway (for SF121 members):
NADH → [AhpF-FAD] → [AhpF-C345/C348] → [AhpF-C129/C132] → [AhpC-C47/C166] → ROOH → ROH + H₂O
(flavin) (TrR-like center) (N-terminal center) (peroxidatic site)
The thioredoxin-dependent pathway (for SF128 members, described by GO:0008379):
NADPH → [TrxR-FAD] → [Trx-C32/C35] → [Prx-Cₚ/Cᵣ] → ROOH → ROH + H₂O
The key distinction is the electron donor: AhpF is a specialized, dedicated reductant with an N-terminal domain specifically evolved to interact with AhpC. This N-terminal domain contains a tandem repeat of two thioredoxin-like folds (PMID: 10828978), and only this domain — not the TrR-like C-terminus — can directly transfer electrons to AhpC (PMID: 10828978). Fusion of this N-terminal domain to E. coli thioredoxin reductase creates a chimeric protein with nearly the same catalytic efficiency as intact AhpF in AhpC reductase assays (PMID: 10913298), confirming that the N-terminal domain is the specific AhpC-interacting module.
The evolutionary ancestry of the family name "THIOREDOXIN PEROXIDASE" likely reflects the fact that the ancestral peroxiredoxin used thioredoxin as its reductant, and AhpF evolved by fusing a thioredoxin-like domain to a thioredoxin reductase scaffold. However, this evolutionary ancestry does not justify annotating modern AhpF-dependent proteins with the thioredoxin-specific GO term — the proteins have functionally diverged in their reductant specificity.
Why thioredoxin can substitute in some organisms: When AhpF is absent (as in H. pylori and T. pallidum), AhpC-type peroxiredoxins can accept electrons from thioredoxin, likely through the same surface that normally interacts with the thioredoxin-like N-terminal domain of AhpF. This biochemical flexibility explains why the family-level annotation "works" for organisms lacking AhpF, but it is misleading for organisms like P. putida that possess the complete AhpC/AhpF system. Notably, an interesting case of functional plasticity was documented in E. coli, where a triplet repeat expansion in ahpC converts the protein from a peroxidase to a disulfide reductase, demonstrating the evolutionary lability of this fold's functional specification (PMID: 11588261).
Evidence Matrix
| # | Citation | Evidence Type | Relationship | Claim Tested | Key Finding | Organism / Context | Confidence & Limitations |
|---|---|---|---|---|---|---|---|
| 1 | PMID: 28375740 | Direct assay + mutagenesis | Supports peroxidase activity; Qualifies reductant | Active-site residues essential? AhpF is reductant? | T43V, R119A, E49Q mutations lower catalytic efficiency 10⁴–10⁵×; AhpF explicitly called "their reductant" | S. typhimurium AhpC; in vitro kinetics | High; nearest characterized homolog (69% identity) |
| 2 | PMID: 20304799 | Direct assay + comparative | Qualifies GO:0008379 | Is thioredoxin the physiological reductant for AhpC-like Prxs? | AhpF is "the typical reductant" for AhpC-like Prxs; Trx used only when AhpF absent | T. pallidum AhpC (no AhpF); Poole lab | High; definitive statement from foremost AhpC experts |
| 3 | PMID: 11222594 | Genetic + kinetic | Supports Trx pathway (when AhpF absent) | Can AhpC use thioredoxin? | H. pylori AhpC uses TrxR/Trx because genome lacks AhpF | H. pylori; in vitro reconstitution | High; establishes reductant-switching paradigm |
| 4 | PMID: 17107553 | Mutant phenotype + regulation | Supports AhpC function in P. putida | Is AhpC expressed and regulated? | OxyR activates ahpC expression; upregulated in toluene-resistant variant | P. putida KT2442 | Moderate; regulatory, not direct activity assay |
| 5 | PMID: 12923780 | Proteomic | Supports AhpC as active antioxidant | Is AhpC expressed under stress? | AhpC protein amplified under MTBE-induced oxidative stress | P. putida KT2440; 2D-PAGE | Moderate; expression, not activity |
| 6 | PMID: 9341228 | Mutagenesis + kinetics | Supports AhpF mechanism | How does AhpF reduce AhpC? | Both AhpF disulfide centers required; <2% peroxidase activity without either | S. typhimurium AhpF; in vitro | High; detailed mechanistic characterization |
| 7 | PMID: 10913298 | Chimeric protein + kinetics | Supports AhpF N-terminus essential | AhpF domain architecture? | N-terminal domain of AhpF fused to TrR confers AhpC reductase activity with same catalytic efficiency | S. typhimurium AhpF / E. coli TrR chimera | High; elegant domain-swap experiment |
| 8 | PMID: 10828978 | Structural + kinetic | Supports AhpF specificity | AhpF electron transfer pathway? | N-terminal 2×Trx-fold mediates direct electron transfer to AhpC; C-terminal TrR-like domain cannot reduce AhpC directly | S. typhimurium AhpF fragments | High |
| 9 | PMID: 21773675 | Biochemical | Supports distinct Prx paralog | Is tsaA a separate peroxiredoxin? | PpPrx (tsaA/Q88NW9) acts as dual-function peroxidase/chaperone | P. putida KT2440 | Moderate; confirms paralog distinction |
| 10 | PMID: 23899494 | Review | Supports general Trx–Prx relationship | Trx as Prx reductant? | "The Trx system provides electrons to thiol-dependent peroxidases (peroxiredoxins)" | Review (Lu & Holmgren) | Context; general, not AhpC-specific |
| 11 | PMID: 11588261 | Genetic / suppressor screen | Qualifies AhpC function | Functional plasticity? | Triplet repeat expansion converts E. coli AhpC from peroxidase to disulfide reductase | E. coli; genetic | High; shows AhpC fold's functional lability |
| 12 | QuickGO database | Computational / database | Qualifies term usage | Experimental basis for GO:0008379? | Zero direct experimental annotations (0/80,826); all IEA/IBA | All organisms | Important context for term validity |
GO Curation Implications
Recommended curation action: GENERALIZE — replace GO:0008379 with GO:0051920 (peroxiredoxin activity), or alternatively with sibling GO:0102039 (NADH-dependent peroxiredoxin activity). This is a lead requiring curator verification.
| Option | GO Term | Rationale | Priority |
|---|---|---|---|
| Generalize (recommended) | GO:0051920 (peroxiredoxin activity) | Correctly describes the intrinsic peroxidase activity without over-specifying the reductant; uses "ROOH" as substrate (appropriate for AhpC's broad specificity) | First choice |
| Replace with sibling | GO:0102039 (NADH-dependent peroxiredoxin activity) | Describes the complete AhpC/AhpF system with correct reductant; more specific but requires inference from genomic context | Also appropriate |
| Retain | GO:0008379 (thioredoxin peroxidase activity) | Defensible only as a synonym convention; specifies wrong reductant for SF121 | Not recommended |
| Remove | — | Not recommended; protein has genuine peroxidase activity | Do not remove |
Rationale for generalization: GO:0008379 defines a specific coupled reaction with thioredoxin as co-substrate ([thioredoxin]-dithiol + H₂O₂ = [thioredoxin]-disulfide + H₂O). The evidence indicates AhpF — not thioredoxin — is the dedicated reductant for this protein. The parent term GO:0051920 (peroxiredoxin activity: [protein]-dithiol + ROOH = [protein]-disulfide + ROH + H₂O) correctly describes the intrinsic activity without specifying the electron donor.
Systematic implications: The same TreeGrafter error likely affects all SF121 (AhpC) proteins in organisms encoding AhpF — this is a systematic, family-wide annotation issue, not specific to Q88K52. A bulk audit of PTHR10681 family annotations would be warranted.
Conflicts, Knowledge Gaps, and Discriminating Tests
Conflicts and Alternatives
-
Reductant specificity ambiguity. AhpF is the dedicated reductant (P0A155 present in genome), but whether thioredoxin can also reduce P. putida AhpC has not been directly tested. In H. pylori (no AhpF) and T. pallidum (no AhpF), thioredoxin reduces AhpC (PMID: 11222594; PMID: 20304799). It is biochemically plausible that P. putida AhpC can accept electrons from thioredoxin, even if AhpF is the preferred donor. This raises a curation question: should GO annotations reflect the physiological reductant or all biochemically competent reductants?
-
Paralog confusion risk. P. putida has at least two typical 2-Cys Prxs (ahpC and tsaA) that both receive GO:0008379 from TreeGrafter. The tsaA protein (Q88NW9, SF128) is the one explicitly designated "Thioredoxin peroxidase" by UniProt curators, while ahpC (Q88K52, SF121) is designated "Alkyl hydroperoxide reductase C." Database users searching for thioredoxin peroxidase activity in P. putida would find both proteins, potentially causing confusion.
-
GO:0008379 has no experimental basis. The term has zero IDA/IMP annotations across all 80,826 uses, raising the question of whether it captures a real enzymatic distinction or is merely a historical naming convention derived from the family name.
Explicit Knowledge Gaps
- No direct biochemical assay of Q88K52. All activity inferences are based on 69% identity to characterized S. typhimurium AhpC. While this is strong evidence, a direct assay would be definitive.
- No test of P. putida AhpC with thioredoxin as reductant. We cannot quantify whether thioredoxin is a poor, moderate, or impossible electron donor for this specific protein.
- No crystal structure of Q88K52. Structural validation relies on homology; AlphaFold model (AF-Q88K52-F1) exists but was not analyzed for active-site geometry.
- Operon structure inferred, not verified. The ahpC-ahpF co-transcription is assumed from analogy to S. typhimurium and E. coli but not experimentally confirmed in P. putida.
Most Efficient Discriminating Tests
-
In vitro reconstitution (highest priority). Purify recombinant Q88K52; measure peroxidase activity with (a) AhpF + NADH, (b) thioredoxin + TrxR + NADPH, (c) DTT as control. Compare kcat/Km to determine reductant preference definitively.
-
ΔahpF genetic test. Delete ahpF in P. putida KT2440 and measure AhpC-dependent peroxide clearance. If activity drops to near-zero (as expected for AhpF-dependent enzymes), this confirms AhpF is the required reductant in vivo.
-
Systematic phylogenomic audit. Query all PTHR10681:SF121 proteins annotated with GO:0008379 and cross-reference with AhpF presence/absence in their genomes. This would quantify the scope of the granularity error across the database.
-
AlphaFold structure analysis. Compare the predicted AhpF-interacting surface of Q88K52 (AF-Q88K52-F1) against the experimentally characterized AhpF-AhpC interface from E. faecalis (PMID: 31047989) to confirm structural compatibility with AhpF binding.
Conclusion
The TreeGrafter/PANTHER annotation of P. putida AhpC (Q88K52) with GO:0008379 (thioredoxin peroxidase activity) represents a granularity error (failure mode #1: family-vs-subfamily). The protein is an authentic, catalytically competent typical 2-Cys peroxiredoxin — all five critical active-site residues are perfectly conserved, and the protein is expressed and functional in P. putida. However, GO:0008379's specification of thioredoxin as the electron donor does not match the protein's subfamily assignment (PANTHER SF121, "ALKYL HYDROPEROXIDE REDUCTASE C"), its genomic context (dedicated reductant AhpF present), or the published biochemical characterization of its nearest homologs (AhpF is "the typical reductant for such enzymes"). The error arose because TreeGrafter propagated a family-level annotation that is appropriate for thioredoxin-dependent subfamilies (SF128) but not for AhpF-dependent subfamilies (SF121). The recommended curation action is to generalize to GO:0051920 (peroxiredoxin activity) or replace with the sibling term GO:0102039 (NADH-dependent peroxiredoxin activity).