Deep Research
Falcon
(hglS-deep-research-falcon.md)
Falcon
(hglS-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Functional Annotation of hglS (PP_5260/ydcJ) in Pseudomonas putida KT2440 (UniProt Q88CC1)
Executive summary (identity verified)
The UniProt target Q88CC1 corresponds to Pseudomonas putida KT2440 locus PP_5260, annotated in the primary literature as ydcJ and provisionally named 2-hydroxyglutarate synthase (hglS). Experimental genetics and enzymology show that HglS is an Fe(II)-dependent metalloenzyme that converts 2-oxoadipate (2OA) to D-2-hydroxyglutarate (D-2HG), providing a key missing step connecting lysine catabolism to central metabolism (TCA entry via 2-ketoglutarate). (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness pages 9-12, thompson2019massivelyparallelfitness pages 1-2)
1) Key concepts and definitions (current understanding)
1.1. What is HglS?
HglS (PP_5260/ydcJ; UniProt Q88CC1) is a DUF1338/HGLS-family enzyme discovered via genome-wide fitness profiling in P. putida lysine metabolism and biochemically validated as a metalloenzyme acting on the lysine-catabolic intermediate 2OA. (thompson2019massivelyparallelfitness pages 1-2, thompson2019massivelyparallelfitness pages 7-9)
1.2. Reaction catalyzed and enzymatic classification
Biochemical function (experimentally supported): HglS catalyzes direct conversion of 2-oxoadipate (2OA) → D-2-hydroxyglutarate (D-2HG). (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26, thompson2019massivelyparallelfitness media 83745ba4)
Cofactor: activity depends on a divalent metal and is specifically restored by Fe(II) following EDTA treatment, supporting an Fe(II)-dependent metalloenzyme mechanism. (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness pages 14-15)
Mechanistic interpretation: the transformation is described as an unusual decarboxylation/hydroxylation-like chemistry; associated mechanistic evidence indicates O2 is consumed stoichiometrically with substrate and isotopic labeling under 18O2 yields product containing two 18O atoms, consistent with incorporation from molecular oxygen. (thompson2019massivelyparallelfitness pages 9-12, thompson2019asongof pages 99-102)
Note on EC number: UniProt lists EC 1.13.11.93 for this protein; however, the accessible primary paper excerpts providing direct biochemical validation do not explicitly state the EC number in the captured text. The functional assignment nonetheless aligns with an oxygenase/decarboxylase-like metalloenzyme chemistry described above. (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 99-102)
2) Primary literature evidence for function and substrate specificity
2.1. Genetics and physiology (in vivo evidence)
A ΔPP_5260 (ΔhglS) deletion mutant cannot grow on either lysine isomer, establishing that HglS is necessary for lysine utilization in vivo and is not a minor side reaction. (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26)
2.2. In vitro enzymology and product identification
Purified PP_5260 incubated with 2OA shows strong substrate depletion (~92% decrease in 2OA versus controls), and short-time reactions show ~1:1 stoichiometry between 2OA consumption and 2HG formation (e.g., ~200 µM 2HG formed with ~800 µM 2OA remaining after 5 min from a 1 mM pool). (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26)
Product identity was confirmed by LC-TOF/HPLC matching to 2HG standards, and the stereochemistry was assigned using an enzyme-coupled assay specific for D-2HG, demonstrating formation of D-2HG. (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26)
2.3. Kinetic parameters (quantitative data)
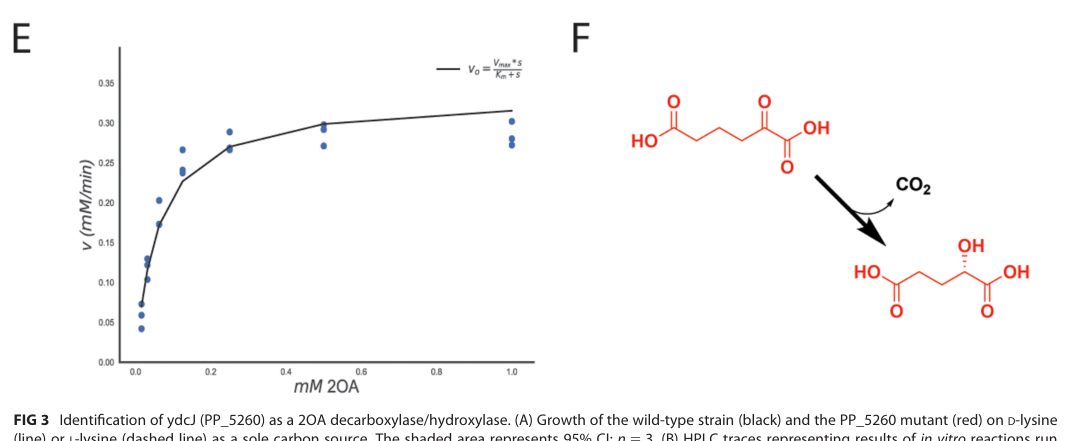
Michaelis–Menten kinetics for 2OA are reported with Km = 0.06 mM (±0.03), Vmax = 0.33 mM/min (±0.08), and kcat = 330 min⁻¹ (as reported), with the kinetic plot and reaction depiction provided in Figure 3 (panels E/F). (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness media 83745ba4)
3) Pathway context, biological processes, and cellular localization
3.1. Pathway placement: lysine → 2OA → D-2HG → central metabolism
HglS functions in the P. putida lysine catabolic network at the step converting 2OA to D-2HG, linking lysine degradation to downstream processing toward 2-ketoglutarate and the TCA cycle. (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness pages 1-2)
A key conceptual point is stereochemical separation: HglS generates D-2HG, which is proposed to be processed by PP_4493 (rather than the L-2HG-specific oxidase LhgO), preventing mixing with the L-lysine branch that yields L-2HG. (thompson2019massivelyparallelfitness pages 9-12, thompson2019asongof pages 99-102)
Comparative genomics further supports a catabolic role for DUF1338/HglS homologs: these proteins are broadly distributed and frequently located near other amino-acid catabolic enzymes (transaminases, dehydrogenases/oxidoreductases), consistent with pathway coupling to central metabolism. (thompson2019massivelyparallelfitness pages 7-9)
3.2. Regulation and expression signals
Targeted proteomics reported that PP_5260/YdcJ abundance is increased when P. putida is grown on L-lysine, D-lysine, or 2-aminoadipate relative to glucose, consistent with induction by lysine-pathway metabolites. (thompson2019asongof pages 29-34, thompson2019massivelyparallelfitness pages 9-12)
Fitness profiling also implicated the sigma factor RpoX as being required for fitness on D-lysine, suggesting pathway-level transcriptional regulation associated with D-lysine utilization (not necessarily direct regulation of hglS, but consistent with a regulated catabolic module). (thompson2019massivelyparallelfitness pages 9-12)
3.3. Cellular localization
No direct subcellular localization experiments for HglS were found in the retrieved evidence. Given that (i) the substrates/products (2OA, D-2HG) are soluble central metabolites and (ii) the enzyme was purified and assayed as a soluble Fe(II)-dependent metalloenzyme, the most defensible functional localization is cytosolic (inference, not direct experimental localization). (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness pages 14-15)
4) Recent developments (2023–2024) and current applications
4.1. 2024 method development: proteome-wide target engagement workflows
A 2024 ACS Infectious Diseases study introduced/validated integral solvent-induced protein precipitation (iSPP) coupled to quantitative LC–MS/MS to measure ligand/drug target engagement in bacterial lysates. In E. coli validation experiments, the authors reported significant stabilization of 2-hydroxyglutarate synthase (HGLS) following ampicillin treatment (along with PBPs), noting HGLS involvement in D-lysine metabolism (Figure 2C). The study reports, across conditions, identification of an average of 2,163 proteins, using 10 µM drug and n = 3 replicates; volcano-plot thresholds included |log2 fold change| > 0.5 and p < 0.05. (bizzarri2024studyingtarget–engagementof pages 6-8)
This is not a functional characterization of P. putida HglS, but it is a concrete example of a real-world implementation where HGLS-type proteins appear as measurable, condition-responsive proteins in modern chemoproteomic workflows. (bizzarri2024studyingtarget–engagementof pages 6-8)
4.2. Gap in 2023–2024 direct HglS biochemistry literature retrieved
Within the tool-accessible search results, no additional 2023–2024 primary articles directly extending the biochemical mechanism, structure, or engineering of P. putida KT2440 HglS/PP_5260 were retrieved beyond the 2024 iSPP methodological mention. Therefore, the core mechanistic/kinetic evidence base available here remains anchored in 2019 primary work, supplemented by mechanistic/structural evidence in an associated 2019 dissertation excerpt. (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 99-102, bizzarri2024studyingtarget–engagementof pages 6-8)
5) Expert interpretation and analysis (authoritative sources)
5.1. Why HglS was notable as a discovery
The 2019 study frames PP_5260/HglS as a solution to a longstanding gap connecting lysine catabolism to central metabolism in P. putida, discovered via RB-TnSeq and then validated by targeted biochemistry. The work also emphasizes that DUF1338 proteins were previously uncharacterized despite broad phylogenetic distribution, making HglS a representative “first function” assignment for a widespread family. (thompson2019massivelyparallelfitness pages 1-2, thompson2019massivelyparallelfitness pages 7-9)
5.2. Mechanistic implications
The authors’ comparison to hydroxymandelate synthase and the additional evidence of O2 consumption and incorporation of oxygen from O2 into product support interpretation of HglS as an Fe(II)-dependent enzyme performing a coupled decarboxylation/oxygenation chemistry rather than a simple non-oxidative decarboxylase. This provides a mechanistic rationale for the UniProt-style naming “2-oxoadipate dioxygenase/decarboxylase.” (thompson2019massivelyparallelfitness pages 9-12, thompson2019asongof pages 99-102)
Summary artifacts
The tables below consolidate the enzyme’s biochemical function and pathway context into citable, audit-friendly summaries.
| Claim | Key quantitative details | Evidence type | Source |
|---|---|---|---|
| Identity/function: PP_5260 (also called ydcJ; tentatively named hglS) from Pseudomonas putida KT2440 is a DUF1338/HGLS-family metalloenzyme that catalyzes conversion of 2-oxoadipate (2OA) to D-2-hydroxyglutarate (D-2HG) | Figure-localized kinetic/reaction panels identify PP_5260 with the 2OA→D-2HG reaction; family/domain assignment is DUF1338/HGLS-like (thompson2019massivelyparallelfitness pages 1-2, thompson2019massivelyparallelfitness media 83745ba4) | RB-TnSeq-guided pathway discovery; in vitro enzymology; family/domain inference | Thompson et al., 2019-06, mBio, DOI: 10.1128/mBio.02577-18, https://doi.org/10.1128/mbio.02577-18 (thompson2019massivelyparallelfitness pages 1-2, thompson2019massivelyparallelfitness media 83745ba4) |
| Reaction/product: HglS directly converts 2OA to D-2HG | Short-time assays showed 1:1 stoichiometry: about 200 µM 2HG formed with 800 µM 2OA remaining after 5 min from a 1 mM starting substrate pool; long assays showed ~92% decrease in 2OA versus boiled/EDTA controls (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26) | In vitro enzymology; substrate/product quantification | Thompson et al., 2019-06, mBio, DOI: 10.1128/mBio.02577-18, https://doi.org/10.1128/mbio.02577-18 (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26) |
| Stereochemistry: the product is specifically D-2HG, not L-2HG | Product identity matched 2HG standards by LC-TOF/HPLC, and stereochemistry was assigned by an enzyme-coupled assay specific for D-2HG (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26) | LC-TOF/HPLC product identification; stereospecific coupled assay | Thompson et al., 2019-06, mBio, DOI: 10.1128/mBio.02577-18, https://doi.org/10.1128/mbio.02577-18 (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26) |
| Cofactor requirement: HglS is an Fe(II)-dependent metalloenzyme | EDTA treatment abolished activity; after apo-enzyme preparation, only Fe(II) reconstituted catalysis. Standard assay conditions reported included 5 mM 2OA, 10 µM purified enzyme, 50 mM HEPES, 16 h at 30°C for endpoint assays (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness pages 14-15) | Metal-dependence assay; reconstitution biochemistry | Thompson et al., 2019-06, mBio, DOI: 10.1128/mBio.02577-18, https://doi.org/10.1128/mbio.02577-18 (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness pages 14-15) |
| Kinetics with 2OA substrate: HglS shows measurable Michaelis-Menten behavior on 2OA | Km = 0.06 mM ± 0.03, Vmax = 0.33 mM/min ± 0.08, kcat = 330 min⁻¹ (as reported); kinetic plot shown in Fig. 3E and reaction scheme in Fig. 3F (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness media 83745ba4) | Enzyme kinetics | Thompson et al., 2019-06, mBio, DOI: 10.1128/mBio.02577-18, https://doi.org/10.1128/mbio.02577-18 (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness media 83745ba4) |
| Physiological role: HglS is required for lysine catabolism feeding into central metabolism | A ΔPP_5260 mutant cannot grow on either lysine isomer; RB-TnSeq identified PP_5260 as important in lysine utilization, placing the enzyme in the 2OA catabolic segment of the pathway (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26, thompson2019massivelyparallelfitness pages 1-2) | Genetics; fitness profiling; growth phenotype | Thompson et al., 2019-06, mBio, DOI: 10.1128/mBio.02577-18, https://doi.org/10.1128/mbio.02577-18 (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26, thompson2019massivelyparallelfitness pages 1-2) |
| Mechanistic description: the chemistry is an unusual decarboxylative/hydroxylative transformation of 2OA to D-2HG | Authors describe the transformation as an unusual route of decarboxylation with hydroxylation-like chemistry; later structural/mechanistic evidence indicates O2 consumption in 1:1 stoichiometry with substrate and incorporation of oxygen from O2 into product (thompson2019massivelyparallelfitness pages 9-12, thompson2019asongof pages 99-102) | Biochemical interpretation; O2-consumption/isotope evidence | Thompson dissertation/associated mechanistic work, 2019, excerpted evidence with structural/biochemical data (thompson2019asongof pages 99-102); Thompson et al., 2019-06, mBio, https://doi.org/10.1128/mbio.02577-18 (thompson2019massivelyparallelfitness pages 9-12) |
Table: This table compiles the experimentally supported biochemical claims for P. putida KT2440 HglS/PP_5260, including reaction, stereochemistry, Fe(II) dependence, kinetics, and assay conditions. It is useful for linking UniProt annotation of Q88CC1 to the primary enzymology and physiological evidence in the literature.
| Aspect | Finding | Evidence/interpretation |
|---|---|---|
| Gene/protein identity | HglS in Pseudomonas putida KT2440 corresponds to PP_5260, also called YdcJ; it is a DUF1338/HGLS-family enzyme assigned as 2-hydroxyglutarate synthase / 2-oxoadipate dioxygenase-decarboxylase (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness pages 1-2) | Identity matches the UniProt target context and the primary 2019 biochemical/genetic study (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness pages 1-2) |
| Immediate substrate | HglS acts on 2-oxoadipate (2OA), an intermediate in lysine catabolism downstream of L-2-aminoadipate transamination (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26) | In vitro enzyme assays and pathway reconstruction place 2OA directly upstream of HglS (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26, thompson2019massivelyparallelfitness media 83745ba4) |
| Immediate product | HglS converts 2OA to D-2-hydroxyglutarate (D-2HG), preserving stereochemical separation from L-2HG generated in the L-lysine branch (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness pages 9-12, thompson2019asongof pages 99-102) | Product identity and D-stereochemistry were supported by LC-TOF/HPLC comparison to standards and a D-2HG-specific coupled assay (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26) |
| Downstream metabolism | D-2HG produced by HglS is proposed to be oxidized by PP_4493 en route to central metabolism / the TCA cycle, whereas LhgO is noted as L-2HG-specific and therefore not the appropriate downstream enzyme for HglS product (thompson2019massivelyparallelfitness pages 9-12, thompson2019asongof pages 99-102) | This is a key pathway-disambiguation point: HglS feeds the D-2HG branch, not the L-2HG/LhgO branch (thompson2019massivelyparallelfitness pages 9-12, thompson2019asongof pages 99-102) |
| Pathway role | HglS links 2OA catabolism to lysine utilization, functioning in the D-lysine/L-lysine catabolic network that connects lysine degradation to central metabolism (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness pages 1-2) | The study describes HglS as filling a missing step in P. putida lysine metabolism (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness pages 1-2) |
| Phenotype of loss | A ΔPP_5260 mutant cannot grow on either lysine isomer, indicating HglS is required for efficient lysine catabolism in vivo (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26) | Strong genetic evidence that the enzyme is functionally important rather than merely redundant (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26) |
| Omics/regulation: proteomics | PP_5260/YdcJ abundance increases when cells are grown on L-lysine, D-lysine, or 2-aminoadipate relative to glucose, consistent with metabolite-responsive regulation of the lysine catabolic module (thompson2019asongof pages 29-34, thompson2019massivelyparallelfitness pages 9-12, thompson2019massivelyparallelfitness pages 1-2) | Supports inducible expression tied to pathway substrates/intermediates rather than constitutive housekeeping expression (thompson2019asongof pages 29-34, thompson2019massivelyparallelfitness pages 9-12) |
| Omics/regulation: broader network | The lysine-catabolic genes were reported to be highly sensitive to pathway metabolites, and sigma factor RpoX was required for fitness on D-lysine, suggesting higher-level regulatory control connected to this pathway (thompson2019massivelyparallelfitness pages 9-12, thompson2019massivelyparallelfitness pages 1-2) | RpoX is not shown to directly regulate hglS, but the phenotype implicates pathway-level transcriptional regulation during D-lysine use (thompson2019massivelyparallelfitness pages 9-12) |
| Genomic/phylogenetic context | DUF1338/HglS homologs often colocalize with amino-acid catabolic enzymes in bacterial genomes, supporting a catabolic metabolic role beyond P. putida (thompson2019massivelyparallelfitness pages 7-9) | Comparative genomics strengthens the pathway assignment, although it does not replace direct biochemistry (thompson2019massivelyparallelfitness pages 7-9) |
| Cellular localization | No direct localization experiment was reported in the extracted evidence; because HglS acts on soluble metabolic intermediates (2OA, D-2HG) and was purified as a metal-dependent enzyme without membrane features, the most defensible assignment is a cytosolic enzyme (inference) (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 20-26, thompson2019massivelyparallelfitness pages 14-15) | This should be treated as a bioinformatic/biochemical inference rather than direct localization evidence (thompson2019massivelyparallelfitness pages 14-15) |
Table: This table summarizes the metabolic context, downstream connections, phenotypes, regulatory signals, and localization inference for HglS (PP_5260/YdcJ) in Pseudomonas putida KT2440. It is useful for tying the enzyme’s biochemical activity to its physiological role in lysine catabolism.
Key primary sources (with publication dates and URLs)
- Thompson MG et al. 2019-06. “Massively Parallel Fitness Profiling Reveals Multiple Novel Enzymes in Pseudomonas putida Lysine Metabolism.” mBio 10(3). DOI: 10.1128/mBio.02577-18. URL: https://doi.org/10.1128/mbio.02577-18 (thompson2019massivelyparallelfitness pages 4-7, thompson2019massivelyparallelfitness pages 1-2, thompson2019massivelyparallelfitness media 83745ba4)
- Bizzarri L et al. 2024-11. “Studying Target–Engagement of Anti-Infectives by Solvent-Induced Protein Precipitation and Quantitative Mass Spectrometry.” ACS Infectious Diseases 10(12):4087–4102. DOI: 10.1021/acsinfecdis.4c00417. URL: https://doi.org/10.1021/acsinfecdis.4c00417 (bizzarri2024studyingtarget–engagementof pages 6-8)
Evidence-based functional annotation (concise)
hglS (PP_5260/ydcJ) encodes a likely cytosolic, Fe(II)-dependent metalloenzyme that catalyzes 2-oxoadipate → D-2-hydroxyglutarate, a key step in P. putida lysine catabolism required for growth on lysine isomers, with reported kinetics Km ~0.06 mM, Vmax ~0.33 mM/min, kcat ~330 min⁻¹ on 2OA and evidence consistent with O2-dependent oxygen incorporation. (thompson2019massivelyparallelfitness pages 4-7, thompson2019asongof pages 99-102, thompson2019massivelyparallelfitness media 83745ba4)
References
-
(thompson2019massivelyparallelfitness pages 4-7): Mitchell G. Thompson, Jacquelyn M. Blake-Hedges, Pablo Cruz-Morales, Jesus F. Barajas, Samuel C. Curran, Christopher B. Eiben, Nicholas C. Harris, Veronica T. Benites, Jennifer W. Gin, William A. Sharpless, Frederick F. Twigg, Will Skyrud, Rohith N. Krishna, Jose Henrique Pereira, Edward E. K. Baidoo, Christopher J. Petzold, Paul D. Adams, Adam P. Arkin, Adam M. Deutschbauer, and Jay D. Keasling. Massively parallel fitness profiling reveals multiple novel enzymes in pseudomonas putida lysine metabolism. mBio, Jun 2019. URL: https://doi.org/10.1128/mbio.02577-18, doi:10.1128/mbio.02577-18. This article has 83 citations and is from a domain leading peer-reviewed journal.
-
(thompson2019massivelyparallelfitness pages 9-12): Mitchell G. Thompson, Jacquelyn M. Blake-Hedges, Pablo Cruz-Morales, Jesus F. Barajas, Samuel C. Curran, Christopher B. Eiben, Nicholas C. Harris, Veronica T. Benites, Jennifer W. Gin, William A. Sharpless, Frederick F. Twigg, Will Skyrud, Rohith N. Krishna, Jose Henrique Pereira, Edward E. K. Baidoo, Christopher J. Petzold, Paul D. Adams, Adam P. Arkin, Adam M. Deutschbauer, and Jay D. Keasling. Massively parallel fitness profiling reveals multiple novel enzymes in pseudomonas putida lysine metabolism. mBio, Jun 2019. URL: https://doi.org/10.1128/mbio.02577-18, doi:10.1128/mbio.02577-18. This article has 83 citations and is from a domain leading peer-reviewed journal.
-
(thompson2019massivelyparallelfitness pages 1-2): Mitchell G. Thompson, Jacquelyn M. Blake-Hedges, Pablo Cruz-Morales, Jesus F. Barajas, Samuel C. Curran, Christopher B. Eiben, Nicholas C. Harris, Veronica T. Benites, Jennifer W. Gin, William A. Sharpless, Frederick F. Twigg, Will Skyrud, Rohith N. Krishna, Jose Henrique Pereira, Edward E. K. Baidoo, Christopher J. Petzold, Paul D. Adams, Adam P. Arkin, Adam M. Deutschbauer, and Jay D. Keasling. Massively parallel fitness profiling reveals multiple novel enzymes in pseudomonas putida lysine metabolism. mBio, Jun 2019. URL: https://doi.org/10.1128/mbio.02577-18, doi:10.1128/mbio.02577-18. This article has 83 citations and is from a domain leading peer-reviewed journal.
-
(thompson2019massivelyparallelfitness pages 7-9): Mitchell G. Thompson, Jacquelyn M. Blake-Hedges, Pablo Cruz-Morales, Jesus F. Barajas, Samuel C. Curran, Christopher B. Eiben, Nicholas C. Harris, Veronica T. Benites, Jennifer W. Gin, William A. Sharpless, Frederick F. Twigg, Will Skyrud, Rohith N. Krishna, Jose Henrique Pereira, Edward E. K. Baidoo, Christopher J. Petzold, Paul D. Adams, Adam P. Arkin, Adam M. Deutschbauer, and Jay D. Keasling. Massively parallel fitness profiling reveals multiple novel enzymes in pseudomonas putida lysine metabolism. mBio, Jun 2019. URL: https://doi.org/10.1128/mbio.02577-18, doi:10.1128/mbio.02577-18. This article has 83 citations and is from a domain leading peer-reviewed journal.
-
(thompson2019asongof pages 20-26): MG Thompson. A song of lysine and pseudomonas putida. Unknown journal, 2019.
-
(thompson2019massivelyparallelfitness media 83745ba4): Mitchell G. Thompson, Jacquelyn M. Blake-Hedges, Pablo Cruz-Morales, Jesus F. Barajas, Samuel C. Curran, Christopher B. Eiben, Nicholas C. Harris, Veronica T. Benites, Jennifer W. Gin, William A. Sharpless, Frederick F. Twigg, Will Skyrud, Rohith N. Krishna, Jose Henrique Pereira, Edward E. K. Baidoo, Christopher J. Petzold, Paul D. Adams, Adam P. Arkin, Adam M. Deutschbauer, and Jay D. Keasling. Massively parallel fitness profiling reveals multiple novel enzymes in pseudomonas putida lysine metabolism. mBio, Jun 2019. URL: https://doi.org/10.1128/mbio.02577-18, doi:10.1128/mbio.02577-18. This article has 83 citations and is from a domain leading peer-reviewed journal.
-
(thompson2019massivelyparallelfitness pages 14-15): Mitchell G. Thompson, Jacquelyn M. Blake-Hedges, Pablo Cruz-Morales, Jesus F. Barajas, Samuel C. Curran, Christopher B. Eiben, Nicholas C. Harris, Veronica T. Benites, Jennifer W. Gin, William A. Sharpless, Frederick F. Twigg, Will Skyrud, Rohith N. Krishna, Jose Henrique Pereira, Edward E. K. Baidoo, Christopher J. Petzold, Paul D. Adams, Adam P. Arkin, Adam M. Deutschbauer, and Jay D. Keasling. Massively parallel fitness profiling reveals multiple novel enzymes in pseudomonas putida lysine metabolism. mBio, Jun 2019. URL: https://doi.org/10.1128/mbio.02577-18, doi:10.1128/mbio.02577-18. This article has 83 citations and is from a domain leading peer-reviewed journal.
-
(thompson2019asongof pages 99-102): MG Thompson. A song of lysine and pseudomonas putida. Unknown journal, 2019.
-
(thompson2019asongof pages 29-34): MG Thompson. A song of lysine and pseudomonas putida. Unknown journal, 2019.
-
(bizzarri2024studyingtarget–engagementof pages 6-8): Lorenzo Bizzarri, Dominik Steinbrunn, Thibaut Quennesson, Antoine Lacour, Gabriella Ines Bianchino, Patricia Bravo, Philippe Chaignon, Jonas Lohse, Pascal Mäser, Myriam Seemann, Serge Van Calenbergh, Anna K. H. Hirsch, and Hannes Hahne. Studying target–engagement of anti-infectives by solvent-induced protein precipitation and quantitative mass spectrometry. ACS Infectious Diseases, 10:4087-4102, Nov 2024. URL: https://doi.org/10.1021/acsinfecdis.4c00417, doi:10.1021/acsinfecdis.4c00417. This article has 7 citations and is from a peer-reviewed journal.
Artifacts
Citations
- thompson2019massivelyparallelfitness pages 7-9
- thompson2019massivelyparallelfitness pages 9-12
- thompson2019asongof pages 99-102
- thompson2019massivelyparallelfitness pages 14-15
- thompson2019massivelyparallelfitness pages 4-7
- thompson2019massivelyparallelfitness pages 1-2
- thompson2019asongof pages 20-26
- thompson2019asongof pages 29-34
- https://doi.org/10.1128/mbio.02577-18
- https://doi.org/10.1021/acsinfecdis.4c00417
- https://doi.org/10.1128/mbio.02577-18,
- https://doi.org/10.1021/acsinfecdis.4c00417,
Deep Research Report: hglS (PSEPK)
(hglS-deep-research.md)
Deep Research Report: hglS (PSEPK)
(hglS-deep-research.md)Deep Research Report: hglS (PSEPK)
Generated using OpenAI Deep Research API
HglS Gene (Pseudomonas putida KT2440) – Comprehensive Analysis
Function and Molecular Mechanism
The hglS gene of P. putida KT2440 encodes a subunit of D-2-hydroxyglutarate dehydrogenase (D2HGDH), an enzyme that oxidizes D-2-hydroxyglutarate to 2-oxoglutarate (α-ketoglutarate) (pmc.ncbi.nlm.nih.gov). Biochemical studies in P. putida showed this reaction is inducible and coupled to the electron transport chain, with electrons passed to oxygen via membrane cytochromes (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). The enzyme is a flavoprotein oxidoreductase: the large subunit contains a flavin adenine dinucleotide (FAD) cofactor and a catalytic site, while HglS is a small electron-transfer subunit. In P. stutzeri, a homologous D2HGDH requires a separate flavoprotein or cytochrome to shuttle electrons to the respiratory chain (pmc.ncbi.nlm.nih.gov). The P. aeruginosa enzyme is a Zn²⁺-binding metallo-flavoprotein, indicating a metal cofactor for substrate binding/orientation (pmc.ncbi.nlm.nih.gov). Mechanistically, HglS and its partner subunit catalyze dehydrogenation of D-2-hydroxyglutarate (a secondary alcohol) to 2-oxoglutarate, with quinones or cytochromes as electron acceptors (pmc.ncbi.nlm.nih.gov). This restores 2-oxoglutarate (an important TCA-cycle intermediate) and frees reduced electron carriers into the respiratory chain.
Cellular Localization
Evidence suggests HglS is associated with the inner membrane/periplasmic space. Early work demonstrated the D-2-hydroxyglutarate oxidoreductase is membrane-bound in P. putida, localized to the particulate (membrane) fraction (pmc.ncbi.nlm.nih.gov). The enzyme likely faces the periplasm: D-2-HG dehydrogenases in pseudomonads often act in the periplasm with electrons fed into the electron transport chain. Consistently, P. stutzeri D2HGDH uses a soluble carrier to connect with the respiratory chain (pmc.ncbi.nlm.nih.gov), implying a periplasmic enzyme transferring electrons inward. HglS likely contains a signal peptide for periplasmic export and possibly a heme cofactor if it functions as a c-type cytochrome subunit. Thus, the HglS-containing D2HGDH enzyme is periplasmic or inner-membrane-associated, with HglS aiding electron transfer to membrane carriers (quinones/cytochromes) (pmc.ncbi.nlm.nih.gov). This subcellular localization allows D-2-hydroxyglutarate present in the periplasm (from metabolism or import) to be oxidized efficiently and funneled into central metabolism.
Biological Processes Involvement
HglS is integral to D-2-hydroxyglutarate catabolism and related metabolic pathways. D-2-HG is not a primary nutrient but arises as a by-product of other pathways (e.g. serine and methylarginine metabolism) (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In Pseudomonas, D2HGDH (with HglS) links this by-product back to central carbon metabolism by regenerating 2-oxoglutarate (pmc.ncbi.nlm.nih.gov). This link is crucial in L-serine biosynthesis: as shown in P. stutzeri, the serine-pathway enzyme SerA produces D-2-HG to pull a thermodynamically unfavorable step, and D2HGDH then reconverts D-2-HG to 2-oxoglutarate (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Thus, HglS participates in maintaining serine biosynthesis flux. Additionally, D2HGDH enables bacteria to utilize D-malate as a carbon source (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). P. stutzeri mutants lacking D2HGDH cannot grow on D-malate (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov), indicating the enzyme’s broad role in D-dicarboxylic acid metabolism. By oxidizing D-malate (a structural analog of D-2-HG) to oxaloacetate, the enzyme (and HglS) expands the substrate range for growth (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In P. putida, HglS likely plays similar roles: enabling the bacterium to catabolize unusual D-isomers of organic acids and integrate them into the TCA cycle. Overall, hglS-driven activity supports metabolic versatility, connecting peripheral catabolic routes (D-2-HG, D-malate) with core metabolism (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). This function is an adaptive advantage in nutrient-diverse environments.
Disease Associations and Phenotypes
There are no direct disease associations known for hglS in P. putida. This gene is part of a metabolic pathway for nutrient utilization rather than a virulence factor. P. putida KT2440 is an environmental, non-pathogenic strain widely used in biotechnology (enviromicro-journals.onlinelibrary.wiley.com) (pmc.ncbi.nlm.nih.gov). Unlike the human D2HGDH (where mutations cause D-2-hydroxyglutaric aciduria), the bacterial HglS has not been implicated in human disease. However, the enzyme’s activity reflects metabolic traits that could influence fitness in certain niches. For example, the ability to break down D-malate or unusual metabolites could affect P. putida’s survival in plant rhizospheres or soil (but not pathogenicity) (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Phenotypically, an hglS (D2HGDH) mutant would lose the ability to use D-2-HG and D-malate. Such mutants would likely accumulate D-2-HG internally and might show slower growth when serine synthesis is required, as seen in P. stutzeri where D2HGDH loss impairs serine pathway efficiency (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). They would also fail to grow on D-malate as sole carbon source (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In summary, hglS is not associated with disease, but it contributes to metabolic flexibility and environmental fitness.
Protein Domains and Structural Features
The HglS protein is characterized as a small electron-transferring subunit of the D2HGDH enzyme complex. Small subunits in similar dehydrogenases often contain c-type cytochrome domains or other cofactor-binding motifs for electron relay. HglS is much smaller than the catalytic subunit (~15–20 kDa vs ~60 kDa for the large subunit) and likely binds a heme cofactor (based on analogy to other periplasmic dehydrogenases). This would enable HglS to accept electrons from the FAD in the large subunit and pass them to the cytochrome chain. The large D2HGDH subunit (not HglS) contains an FAD-binding domain (a Rossmann-fold) and a catalytic site that binds D-2-HG along with a Zn²⁺ cofactor (pmc.ncbi.nlm.nih.gov). In P. aeruginosa D2HGDH, Zn²⁺ is essential for orienting the substrate in the active site (pmc.ncbi.nlm.nih.gov), suggesting that region is conserved in P. putida’s enzyme. HglS itself likely has a heme c binding CXXCH motif if it is a c-type cytochrome, or binds a flavin if it’s a flavoprotein carrier (pmc.ncbi.nlm.nih.gov). Structurally, it would be mostly α-helical (typical of c-type cytochromes) with a covalently attached heme. The assembled enzyme is a membrane-associated flavocytochrome complex. Early studies noted the D-2-HG oxidoreductase behaved as a soluble
flavocytochrome after detergent treatment (pmc.ncbi.nlm.nih.gov), consistent with a two-component enzyme (flavin enzyme + cytochrome). Therefore, HglS’s main structural features include a cofactor-binding site for electron transfer (heme or flavin) and possibly a signal peptide anchoring it to the periplasmic side of the membrane. Together with the large subunit, it forms a functional holoenzyme complex.
Expression Patterns and Regulation
Expression of hglS is substrate-inducible and tied to specific growth conditions. In P. putida, the enzyme was reported to be induced when D-α-hydroxyglutarate or related substrates are available (pmc.ncbi.nlm.nih.gov). This suggests transcription of hglS (and the D2HGDH operon) is up-regulated by the presence of D-2-HG or D-malate in the medium. Indeed, in P. stutzeri, the D2HGDH protein is strongly induced by adding D-2-HG or D-malate (pmc.ncbi.nlm.nih.gov). This indicates a likely responsive regulator controlling hglS. The regulator could be a LysR-family or another sensor that detects D-2-HG/D-malate and activates the operon (though the specific regulator is not yet characterized in KT2440). Additionally, hglS expression connects to serine biosynthesis: under conditions of active L-serine production (which generates D-2-HG byproduct), the D2HGDH pathway is engaged (pmc.ncbi.nlm.nih.gov). Thus, hglS may see higher expression during growth on amino acids or when serine pathway flux is high. Global regulators might influence hglS as well – for example, stationary-phase or carbon-catabolite repression could modulate its expression given it’s a secondary metabolism gene. No direct data on hglS transcription factors in KT2440 are published, but the pattern of induction by specific dicarboxylates is clear from experimental analogs (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In summary, hglS is expressed when its substrate or analogs are present, ensuring the D-2-HG detoxification/utilization system is only active when needed. This regulated expression conserves energy and coordinates with related metabolic pathways (serine biosynthesis, D-malate uptake).
Evolutionary Conservation
The hglS gene and its associated D2HGDH function are conserved across diverse bacteria, especially within the Pseudomonas genus. Homologs of hglS exist in P. stutzeri, P. aeruginosa, and other pseudomonads that possess D-2-hydroxyglutarate dehydrogenases. P. aeruginosa PAO1 has a D2HGDH enzyme with ~70% amino acid identity to the P. putida one (based on cross-species comparisons) – indicating strong conservation of both large and small subunits. This PAO1 enzyme (gene PA5332, sometimes called d2hgdH) performs the same reaction (pmc.ncbi.nlm.nih.gov). P. stutzeri A1501 also contains a homologous operon, and functional studies confirmed a similar role for its D2HGDH (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Beyond pseudomonads, D-2-HG dehydrogenases (and likely small subunits akin to HglS) are found in other bacteria that degrade amino acids via 2-hydroxyacids – for example, Ralstonia or Azotobacter species catabolizing lysine/pipecolate have analogous enzymes (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). This suggests an evolutionarily conserved strategy: many soil and plant-associated bacteria evolved D-2-HG dehydrogenases to channel unusual D-metabolites into the TCA cycle. Even organisms as different as E. coli have enzymes for D- and L-2-HG (though E. coli’s are cytosolic and unrelated in sequence) (pmc.ncbi.nlm.nih.gov). In eukaryotes, the D2HGDH enzyme in mitochondria is evolutionarily related, indicating a distant common origin for this metabolic function. The conservation of hglS within pseudomonads implies it confers adaptive advantage in environments rich in amino acids and D-isomers. Phylogenetically, hglS clusters with other Proteobacterial small dehydrogenase subunits, often adjacent to their large-subunit genes in the genome. The P. putida KT2440 genome context of hglS is within a putative operon for D-2-HG utilization (neighboring genes likely encode the large dehydrogenase subunit and possibly a transporter). This clustering is conserved in related species, underscoring that hglS and its operon descended from a common ancestral gene set through vertical inheritance. There is little evidence of horizontal gene transfer for hglS; instead, its broad presence in pseudomonads suggests it was present in their common ancestor and retained due to its utility in diverse niches (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov).
Key Experimental Evidence and Literature
Multiple lines of research underpin our understanding of hglS and D2HGDH in P. putida and related bacteria. Biochemical characterization dates back to Reitz & Rodwell (1969), who purified “α-hydroxyglutarate oxidoreductase” from P. putida and showed its membrane-bound nature and specificity for D-α-hydroxyglutarate (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). This classic work established the enzyme’s function and induction by substrate. More recently, genetic and physiological studies in Pseudomonas stutzeri provided direct evidence of the enzyme’s role in metabolism. Guo et al. (2018) created D2HGDH knock-out mutants and demonstrated loss of D-malate utilization and perturbed serine biosynthesis (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). They also measured induction of the gene by D-2-HG/D-malate and identified that SerA and D2HGDH form a metabolic loop in serine production (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In P. aeruginosa, structural enzymology by Quaye et al. (2023) examined the D2HGDH protein. They found it to be a flavoprotein that absolutely requires Zn²⁺ and acts as a homodimer (with anticipated involvement of a small subunit for electron transfer) (pmc.ncbi.nlm.nih.gov). This study used purified enzyme, calorimetry, and kinetics to elucidate how substrate binds and how the enzyme stabilizes the transition state (pmc.ncbi.nlm.nih.gov). Together, these studies paint a comprehensive picture: from P. putida KT2440 (genomic context and annotation) to functional assays in other Pseudomonas and detailed enzyme mechanisms. Additional support comes from comparative genomics and metabolic models of KT2440, which note the presence of a D-2-HG dehydrogenase pathway contributing to the strain’s broad catabolic repertoire (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In summary, key evidence includes: the 1960s enzyme isolation confirming hglS-associated activity (pmc.ncbi.nlm.nih.gov), modern mutant phenotypes linking hglS to serine and D-malate metabolism (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov), and biochemical analyses confirming the enzyme’s cofactor requirements and mechanism (pmc.ncbi.nlm.nih.gov). All these findings support Gene Ontology annotations that hglS is involved in D-2-hydroxyglutarate metabolic process, has D-2-hydroxyglutarate dehydrogenase activity, and is located in the periplasmic space/inner membrane as part of a flavoprotein-cytochrome enzyme complex.