Deep Research
Falcon
(rpoH-deep-research-falcon.md)
Falcon
(rpoH-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: rpoH (UniProt Q7CCA6; locus PP_5108) in Pseudomonas putida KT2440 (PSEPK)
0) Identity verification (mandatory)
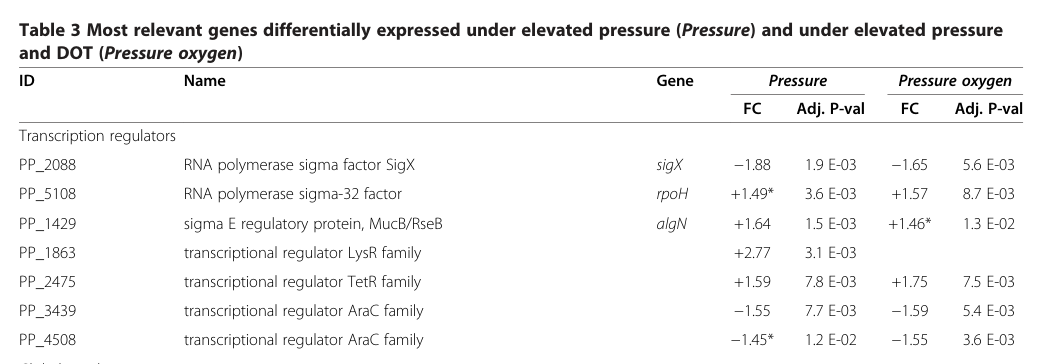
The literature gathered matches the UniProt target Q7CCA6 because it explicitly identifies PP_5108 in P. putida KT2440 as rpoH, annotated as the RNA polymerase sigma-32 factor (σ32/RpoH), which is the canonical heat-shock sigma factor in Gram-negative bacteria. In KT2440 pressure transcriptomics, PP_5108 is labeled “RNA polymerase sigma-32 factor” with gene name rpoH, confirming the locus-to-function mapping. (follonier2013newinsightson pages 5-6)
1) Key concepts and definitions (current understanding)
1.1 What RpoH (σ32) is
RpoH (σ32) is an alternative sigma factor in the σ70 family. Sigma factors are RNA polymerase (RNAP) specificity subunits that redirect RNAP to particular promoter classes. In Gram-negative bacteria, σ32 is the major transcriptional regulator of the heat-shock response (HSR), activating genes that restore protein homeostasis (proteostasis)—molecular chaperones and proteases that refold or degrade damaged proteins. (potvin2008sigmafactorsin pages 3-5, ito2014geneticandphenotypic pages 2-3)
In a widely cited Pseudomonas-focused review, RpoH is described as an σ70-like factor with substantial similarity to E. coli σ32 (noted as ~61% identity for the P. aeruginosa RpoH). It is responsible for the canonical heat-shock induction response and supports basal expression of multiple heat-shock proteins. (potvin2008sigmafactorsin pages 3-5)
1.2 σ70-family domain architecture relevant to RpoH function
σ70-family sigma factors share conserved regions that implement promoter recognition and RNAP interaction. In Pseudomonas, these conserved σ70-family regions are summarized as:
- Region/domain 2: most conserved; involved in core RNAP binding, DNA melting, and recognition of the −10 element.
- Region/domain 4: contains an HTH DNA-binding motif for recognition of the −35 element.
These concepts support functional inference for Q7CCA6 (RpoH subfamily) in promoter recognition and transcription initiation. (potvin2008sigmafactorsin pages 3-5)
1.3 σ32 regulon size and promoter consensus (reference framework)
While regulon membership differs across organisms, a quantitative reference point for σ32 control comes from E. coli: σ32 is cited as controlling ~50 transcriptional units (~90 genes), emphasizing its broad role in proteostasis. (ito2014geneticandphenotypic pages 2-3)
A σ32 promoter consensus reported in the P. putida heat-shock genetics study (citing the E. coli consensus) is CTTGAA N13–17 CCCCATNT, and P. putida candidate σ32 promoter sequences were identified upstream of cbpA, hfq (PP4894), and secA (PP1345). (ito2014geneticandphenotypic pages 6-8)
2) Primary function and pathway placement in P. putida KT2440
2.1 Primary molecular function
RpoH is not an enzyme; its primary function is sequence-specific transcription initiation control. RpoH binds the RNAP core enzyme to form an alternative holoenzyme and drives transcription from heat-shock promoters, upregulating chaperones/proteases and related stress-repair functions. This is the central mechanistic meaning of “heat shock sigma factor.” (ito2014geneticandphenotypic pages 2-3, ito2014geneticandphenotypic pages 6-8)
2.2 Biological processes governed (HSR/proteostasis control)
In Pseudomonas putida, the heat-shock response is described as being controlled by σ32 (encoded by rpoH) and implemented by conserved chaperone/protease systems, notably DnaK/DnaJ/GrpE and GroEL/GroES chaperone machines and ATP-dependent proteases (including FtsH) that together establish a feedback-regulated proteostasis circuit. (ito2014geneticandphenotypic pages 1-2, ito2014geneticandphenotypic pages 2-3)
2.3 Cellular localization of RpoH action
RpoH functions where transcription occurs: it acts as part of the RNAP holoenzyme on the bacterial chromosome in the cytoplasm/nucleoid (i.e., it is not secreted and does not act in the periplasm). Its regulatory targets include cytosolic chaperones (DnaK/DnaJ/GrpE; GroEL/GroES) and proteases; regulation also involves the membrane-embedded protease FtsH, linking cytosolic σ32 levels to membrane-associated proteolysis. (ito2014geneticandphenotypic pages 1-2, ito2014geneticandphenotypic pages 2-3)
3) Regulation of RpoH/σ32 in P. putida: mechanisms and experimentally supported phenotypes
3.1 Negative feedback control by chaperones and proteolysis
A central current model is chaperone-mediated negative feedback: when proteins misfold under stress, chaperones are titrated away from σ32, freeing σ32 to activate heat-shock transcription; as chaperone levels rise, they re-bind/inactivate σ32 and promote its turnover.
In P. putida, σ32 activity/quantity is controlled by DnaK/DnaJ/GrpE and GroEL/GroES, and σ32 is degraded by a membrane ATP-dependent protease (implicated as FtsH) with chaperone assistance. (ito2014geneticandphenotypic pages 1-2, ito2014geneticandphenotypic pages 2-3)
3.2 Induction kinetics (quantitative)
σ32 induction after heat shock is described as transient, peaking at ~5–15 minutes, and in P. putida heat-shock experiments increased expression of heat-shock genes occurs within ~10 minutes and correlates with σ32 levels. Moreover, rpoH mRNA can continue increasing even as σ32 protein levels decline, indicating layered transcriptional/post-transcriptional regulation. (ito2014geneticandphenotypic pages 2-3, ito2014geneticandphenotypic pages 6-8)
3.3 Genetic evidence for the σ32-centered proteostasis circuit
While not direct rpoH knockouts, genetic perturbations in σ32 effector systems demonstrate the pathway’s biological necessity:
- A ΔclpB mutant in P. putida shows normal heat-shock protein synthesis (except ClpB) but is highly temperature sensitive and cannot disaggregate thermo-induced aggregates, demonstrating that σ32-driven proteostasis requires specific disaggregation capacity for survival after heat stress. (ito2014geneticandphenotypic pages 1-2)
- A ΔdnaJ mutant is temperature sensitive and accumulates more (especially high-molecular-weight) aggregates upon heat stress; CbpA can partially substitute for DnaJ, consistent with σ32-regulated tuning of DnaK co-chaperone capacity in P. putida. (ito2014geneticandphenotypic pages 1-2, ito2014geneticandphenotypic pages 2-3)
4) Stress contexts in KT2440 where rpoH/σ32 is implicated (omics + experimental evidence)
4.1 Elevated pressure / bioreactor-relevant conditions (KT2440)
Under elevated pressure conditions relevant to industrial-scale bioreactors, KT2440 microarrays show upregulation of rpoH (PP_5108) alongside canonical heat-shock genes. Specifically:
- rpoH (PP_5108): +1.49 fold-change (adjusted p = 3.6×10−3) under pressure; +1.57 (adjusted p = 8.7×10−3) under pressure + elevated oxygen.
- Chaperone genes are co-induced: htpG +1.65/+1.92, groES +1.61/+1.77, groEL +1.78/+2.19, consistent with activation of a heat-shock-like proteostasis response. (follonier2013newinsightson pages 5-6, follonier2013newinsightson media 908730bc)
4.2 Aromatic solvent stress / proteotoxicity (KT2440)
In a KT2440 study examining toluene and related aromatics, induced proteins are described as belonging to the RpoH regulon, including GroES, GroEL, GrpE, Lon protease, DnaK, and IbpA, linking solvent exposure to heat-shock/proteostasis activation. (dominguezcuevas2006transcriptionaltradeoffbetween pages 7-8)
Quantitatively, large inductions were observed for multiple heat-shock/proteostasis genes upon aromatic exposure (selected examples): HslU 12.03-fold (o-xylene), DnaK 6.43-fold (o-xylene), HSP20 family protein 47.05-fold (toluene), and IbpA up to 6.46-fold (3MB), illustrating strong compound-specific activation of proteostasis pathways. (dominguezcuevas2006transcriptionaltradeoffbetween pages 7-8)
4.3 Aromatic bioproduct stress in a KT2440-background strain (2023)
A 2023 study in P. putida DOT-T1E (a solvent-tolerant strain, KT2440 background) exposed to aromatics (styrene/trans-cinnamic acid) reported induction of multiple chaperones and proteostasis-related proteins including GroEL, SurA, HtpG, and GroSL, with associated oxidative-stress defenses and efflux systems; some genes were overexpressed by up to 7.7-fold and ATP synthase genes induced 2.2–5.2-fold, indicating substantial physiological burden and engagement of stress repair systems relevant to industrial aromatic production. (garciafranco2023insightsintothe pages 9-10, garciafranco2023insightsintothe pages 10-11)
5) Applications and real-world implementations
5.1 Process robustness and temperature-based control
A review of Pseudomonas stress responses emphasizes that heat-shock systems (including σ32/RpoH control and RNA thermosensors) can be leveraged for industrial applications. It highlights ROSE-type RNA thermometers in heat-shock gene regulation; in P. putida, a ROSE element precedes ibpA (small heat-shock protein), illustrating temperature-responsive post-transcriptional control within the broader σ32-linked heat-shock network. (craig2021leveragingpseudomonasstress pages 7-9)
As a concrete process-relevant example, P. putida KT2440 heterologous rhamnolipid production increased by >60% when temperature was raised from 30°C to 37°C, supporting the concept that temperature shifts can be used as a practical control input interacting with heat-shock regulatory circuitry. (craig2021leveragingpseudomonasstress pages 7-9)
5.2 Industrially relevant stresses that intersect RpoH biology
The same review notes that industrial workflows expose bacteria to temperature extremes (e.g., drying and sterilization steps) and that survival relies on “chaperones, proteases, thermosensors, and alternative sigma factors.” This frames RpoH-centered proteostasis as directly relevant to strain engineering for robustness and to formulation processes (e.g., drying). (craig2021leveragingpseudomonasstress pages 5-6, craig2021leveragingpseudomonasstress pages 11-12)
6) Recent developments and latest research (prioritizing 2023–2024)
6.1 2024 systems biology: genome-wide mapping of RpoH binding and sigmulons
A 2024 PLOS Genetics study applied genome-wide binding assays and transcriptomic systems biology to quantify sigma-factor regulatory dynamics during heat shock (in Salmonella as a model). While not P. putida, it exemplifies the current best practice for defining sigma regulons (ChIP-based occupancy + RNA-seq integration + iModulon inference) and produces quantitative benchmarks:
- Binding sites under control conditions: RpoH 181, with +40 new and −8 lost after heat shock.
- RpoH sigmulon size: 151 genes (93 TUs) under control vs 163 genes (98 TUs) after heat shock; 43 RpoH sigmulon genes changed significantly in expression.
- Overlap with housekeeping sigma RpoD: ~28.7% (control) and ~24.8% (heat shock), indicating shared/competitive promoter use.
These results provide up-to-date quantitative framing for σ32 network plasticity and sigma competition, which are likely relevant principles for Pseudomonas RpoH biology even if organism-specific maps differ. (park2024unveilingthenovel pages 5-7, park2024unveilingthenovel pages 4-5)
6.2 2024 population genomics: updated genomic context for KT2440 robustness
A 2024 FEMS Microbiology Reviews article quantifies stress-related gene inventories and regulatory capacity in the P. putida group, offering updated genomic context for KT2440 (noting its robustness). It reports that KT2440 encodes 10 universal stress proteins, 7 cold shock proteins, 5 heat shock proteins, and that ~10% of the genome is devoted to regulatory proteins, alongside extensive efflux capacity (e.g., 14 RND efflux pumps across four subfamilies). Although RpoH is not singled out in the excerpted passage, these data contextualize σ-factor–mediated stress regulation as embedded within a large regulatory repertoire supporting environmental and industrial resilience. (udaondo2024unravelingthegenomic pages 14-16)
7) Expert interpretation (authoritative synthesis)
Collectively, the evidence supports functional annotation of rpoH (Q7CCA6; PP_5108) in KT2440 as a σ70-family alternative sigma factor (σ32) that orchestrates a proteostasis-centered stress response. The strongest organism-specific signals are that KT2440/KT2442 quickly induces σ32-dependent programs upon heat upshift (minutes scale), and that diverse industrially relevant stresses that cause protein damage (aromatic solvents; elevated pressure/oxygen transfer conditions) activate rpoH-linked chaperone/protease expression, reflecting that these stresses converge on the same proteostasis bottleneck. (ito2014geneticandphenotypic pages 6-8, follonier2013newinsightson pages 5-6, dominguezcuevas2006transcriptionaltradeoffbetween pages 7-8)
8) Evidence summary table
| Claim / functional element | Evidence type | Key quantitative data | Stress condition / context | Source and URL |
|---|---|---|---|---|
| Identity: PP_5108 in P. putida KT2440 is rpoH, annotated as RNA polymerase sigma-32 factor (RpoH) | Omics annotation + organism-specific literature | Table entry explicitly maps PP_5108 = rpoH = RNA polymerase sigma-32 factor; functional studies in P. putida state rpoH encodes σ32 | KT2440 transcriptome under elevated pressure; KT2442 heat-shock genetics | Follonier et al. 2013 Microbial Cell Factories https://doi.org/10.1186/1475-2859-12-30; Ito et al. 2014 MicrobiologyOpen https://doi.org/10.1002/mbo3.217 (follonier2013newinsightson pages 5-6, ito2014geneticandphenotypic pages 1-2) |
| Induction kinetics: RpoH/σ32 is rapidly induced after heat shift and correlates with hsp induction | Experiment | σ32 induction is described as transient, peaking at ~5–15 min after heat shock; in P. putida, hsp gene expression increased within ~10 min and correlated with σ32 level | Heat-shock response in P. putida KT2442 | Ito et al. 2014 MicrobiologyOpen https://doi.org/10.1002/mbo3.217 (ito2014geneticandphenotypic pages 2-3, ito2014geneticandphenotypic pages 6-8) |
| Regulon size / promoter logic: RpoH directs RNAP to heat-shock promoters; canonical σ32 promoter motif reported | Experiment + comparative regulatory analysis | E. coli σ32 regulon cited as ~50 transcriptional units / ~90 genes; reported σ32 promoter consensus CTTGAA N13-17 CCCCATNT; candidate P. putida σ32-regulated genes include cbpA, hfq, secA | Heat-shock transcriptional regulation; inference used in P. putida promoter searches | Ito et al. 2014 MicrobiologyOpen https://doi.org/10.1002/mbo3.217 (ito2014geneticandphenotypic pages 6-8) |
| Mechanistic regulation: RpoH activity and abundance are controlled by chaperones and proteolysis | Review + experiment | DnaK/DnaJ/GrpE and GroEL/GroES bind/inactivate σ32; FtsH degrades σ32; P. putida dnaJ mutant is temperature sensitive and accumulates more protein aggregates, while clpB mutant is heat sensitive and defective in aggregate solubilization | Heat-shock quality-control circuit in cytoplasm | Ito et al. 2014 MicrobiologyOpen https://doi.org/10.1002/mbo3.217; Potvin et al. 2008 FEMS Microbiology Reviews https://doi.org/10.1111/j.1574-6976.2007.00092.x (ito2014geneticandphenotypic pages 1-2, potvin2008sigmafactorsin pages 3-5, ito2014geneticandphenotypic pages 2-3) |
| Protein family / domain context: RpoH is an alternative sigma-70 family sigma factor for heat-shock transcription | Review | RpoH shows 61% identity to E. coli σ32; σ70-family architecture includes conserved domain 2 (core RNAP binding, DNA melting, -10 recognition) and domain 4 (HTH, -35 recognition) | General bacterial transcription; relevant to Pseudomonas RpoH annotation | Potvin et al. 2008 FEMS Microbiology Reviews https://doi.org/10.1111/j.1574-6976.2007.00092.x (potvin2008sigmafactorsin pages 3-5) |
| Pressure-associated induction in KT2440: rpoH and canonical heat-shock genes are upregulated | Omics (microarray; supported by qRT-PCR for most tested genes) | Under elevated pressure: rpoH +1.49 (adj. P = 3.6E-03); with pressure + O2: +1.57 (adj. P = 8.7E-03). Associated genes: htpG +1.65 / +1.92, groES +1.61 / +1.77, groEL +1.78 / +2.19, grpE +1.33 | KT2440 under elevated pressure and elevated pressure + dissolved oxygen; bioprocess-relevant stress | Follonier et al. 2013 Microbial Cell Factories https://doi.org/10.1186/1475-2859-12-30 (follonier2013newinsightson pages 5-6, follonier2013newinsightson media 908730bc, follonier2013newinsightson media 43885585) |
| Solvent response link: aromatic solvent stress recruits the RpoH heat-shock program | Omics + experiment | Toluene/o-xylene exposure induced proteins assigned to the RpoH regulon, including GroES, GroEL, GrpE, Lon, DnaK, IbpA; activation of the TOL meta operon was reported to require the contribution of σ32 and/or σ38 host factors | KT2440 exposed to toluene/aromatic solvents; proteotoxic stress tradeoff with metabolism | Domínguez-Cuevas et al. 2006 Journal of Biological Chemistry https://doi.org/10.1074/jbc.m509848200 (dominguezcuevas2006transcriptionaltradeoffbetween pages 1-2) |
Table: This table compiles organism-specific evidence supporting the functional annotation of Pseudomonas putida KT2440/KT2442 rpoH (PP_5108) as the sigma-32 heat-shock sigma factor. It highlights identity, regulatory mechanism, kinetics, regulon features, and stress-responsive expression data most useful for gene-function annotation.
9) Key quantitative evidence highlights (selected)
- KT2440 elevated pressure: rpoH +1.49 FC (adj. p 3.6×10−3), groEL +1.78 FC, groES +1.61 FC, htpG +1.65 FC. (follonier2013newinsightson pages 5-6, follonier2013newinsightson media 908730bc)
- KT2440 aromatic exposure: HSP20 family protein 47.05-fold (toluene), HslU 12.03-fold (o-xylene), DnaK 6.43-fold (o-xylene), IbpA up to 6.46-fold (3MB). (dominguezcuevas2006transcriptionaltradeoffbetween pages 7-8)
- σ32 induction timing: peak ~5–15 min, hsp induction within ~10 min. (ito2014geneticandphenotypic pages 2-3, ito2014geneticandphenotypic pages 6-8)
- 2024 genome-wide sigma mapping (model organism): RpoH binding sites 181 baseline, +40/−8 remodeling; sigmulon 151→163 genes under heat shock. (park2024unveilingthenovel pages 5-7, park2024unveilingthenovel pages 4-5)
- 2024 KT2440-group genomics: 10 universal stress proteins, 7 cold shock proteins, 5 heat shock proteins, ~10% genome regulatory proteins, 14 RND efflux pumps. (udaondo2024unravelingthegenomic pages 14-16)
10) URLs and publication dates (most-cited sources used here)
- Ito et al. 2014-10. MicrobiologyOpen. “Genetic and phenotypic characterization of the heat shock response in Pseudomonas putida.” https://doi.org/10.1002/mbo3.217 (ito2014geneticandphenotypic pages 2-3, ito2014geneticandphenotypic pages 6-8)
- Follonier et al. 2013-03. Microbial Cell Factories. “New insights on the reorganization of gene transcription in P. putida KT2440 at elevated pressure.” https://doi.org/10.1186/1475-2859-12-30 (follonier2013newinsightson pages 5-6, follonier2013newinsightson media 908730bc)
- Domínguez-Cuevas et al. 2006-04. Journal of Biological Chemistry. “Transcriptional tradeoff between metabolic and stress-response programs in P. putida KT2440 exposed to toluene.” https://doi.org/10.1074/jbc.m509848200 (dominguezcuevas2006transcriptionaltradeoffbetween pages 7-8)
- García-Franco et al. 2023-02. Microbial Cell Factories. “Susceptibility of P. putida to industrial aromatic hydrocarbons.” https://doi.org/10.1186/s12934-023-02028-y (garciafranco2023insightsintothe pages 9-10)
- Park et al. 2024-10. PLOS Genetics. Sigma-factor systems biology mapping under heat shock. https://doi.org/10.1371/journal.pgen.1011464 (park2024unveilingthenovel pages 5-7, park2024unveilingthenovel pages 4-5)
- Udaondo et al. 2024-10. FEMS Microbiology Reviews. Population genomics of the P. putida group. https://doi.org/10.1093/femsre/fuae025 (udaondo2024unravelingthegenomic pages 14-16)
11) Notes on limitations (scope and organism specificity)
A complete, experimentally defined KT2440-specific RpoH regulon map (e.g., ChIP-seq/ChIP-exo of RpoH in KT2440) was not available in the retrieved corpus. Therefore, regulon membership is supported here by (i) direct KT2440 stress transcriptomics, (ii) KT2442 heat-shock genetics and promoter motif searches, and (iii) solvent-stress gene induction patterns explicitly labeled as RpoH regulon members by the authors. (ito2014geneticandphenotypic pages 6-8, dominguezcuevas2006transcriptionaltradeoffbetween pages 7-8)
References
-
(follonier2013newinsightson pages 5-6): Stéphanie Follonier, Isabel F Escapa, Pilar M Fonseca, Bernhard Henes, Sven Panke, Manfred Zinn, and María Auxiliadora Prieto. New insights on the reorganization of gene transcription in pseudomonas putida kt2440 at elevated pressure. Microbial Cell Factories, 12:30-30, Mar 2013. URL: https://doi.org/10.1186/1475-2859-12-30, doi:10.1186/1475-2859-12-30. This article has 46 citations and is from a peer-reviewed journal.
-
(potvin2008sigmafactorsin pages 3-5): Eric Potvin, François Sanschagrin, and Roger C. Levesque. Sigma factors in pseudomonas aeruginosa. FEMS microbiology reviews, 32 1:38-55, Jan 2008. URL: https://doi.org/10.1111/j.1574-6976.2007.00092.x, doi:10.1111/j.1574-6976.2007.00092.x. This article has 427 citations and is from a domain leading peer-reviewed journal.
-
(ito2014geneticandphenotypic pages 2-3): Fumihiro Ito, Takayuki Tamiya, Iwao Ohtsu, Makoto Fujimura, and Fumiyasu Fukumori. Genetic and phenotypic characterization of the heat shock response in pseudomonas putida. MicrobiologyOpen, 3:922-936, Oct 2014. URL: https://doi.org/10.1002/mbo3.217, doi:10.1002/mbo3.217. This article has 28 citations and is from a peer-reviewed journal.
-
(ito2014geneticandphenotypic pages 6-8): Fumihiro Ito, Takayuki Tamiya, Iwao Ohtsu, Makoto Fujimura, and Fumiyasu Fukumori. Genetic and phenotypic characterization of the heat shock response in pseudomonas putida. MicrobiologyOpen, 3:922-936, Oct 2014. URL: https://doi.org/10.1002/mbo3.217, doi:10.1002/mbo3.217. This article has 28 citations and is from a peer-reviewed journal.
-
(ito2014geneticandphenotypic pages 1-2): Fumihiro Ito, Takayuki Tamiya, Iwao Ohtsu, Makoto Fujimura, and Fumiyasu Fukumori. Genetic and phenotypic characterization of the heat shock response in pseudomonas putida. MicrobiologyOpen, 3:922-936, Oct 2014. URL: https://doi.org/10.1002/mbo3.217, doi:10.1002/mbo3.217. This article has 28 citations and is from a peer-reviewed journal.
-
(follonier2013newinsightson media 908730bc): Stéphanie Follonier, Isabel F Escapa, Pilar M Fonseca, Bernhard Henes, Sven Panke, Manfred Zinn, and María Auxiliadora Prieto. New insights on the reorganization of gene transcription in pseudomonas putida kt2440 at elevated pressure. Microbial Cell Factories, 12:30-30, Mar 2013. URL: https://doi.org/10.1186/1475-2859-12-30, doi:10.1186/1475-2859-12-30. This article has 46 citations and is from a peer-reviewed journal.
-
(dominguezcuevas2006transcriptionaltradeoffbetween pages 7-8): Patricia Domínguez-Cuevas, José-Eduardo González-Pastor, Silvia Marqués, Juan-Luis Ramos, and Víctor de Lorenzo. Transcriptional tradeoff between metabolic and stress-response programs in pseudomonas putida kt2440 cells exposed to toluene*. Journal of Biological Chemistry, 281:11981-11991, Apr 2006. URL: https://doi.org/10.1074/jbc.m509848200, doi:10.1074/jbc.m509848200. This article has 269 citations and is from a domain leading peer-reviewed journal.
-
(garciafranco2023insightsintothe pages 9-10): Ana García-Franco, Patricia Godoy, Estrella Duque, and Juan Luis Ramos. Insights into the susceptibility of pseudomonas putida to industrially relevant aromatic hydrocarbons that it can synthesize from sugars. Microbial Cell Factories, Feb 2023. URL: https://doi.org/10.1186/s12934-023-02028-y, doi:10.1186/s12934-023-02028-y. This article has 15 citations and is from a peer-reviewed journal.
-
(garciafranco2023insightsintothe pages 10-11): Ana García-Franco, Patricia Godoy, Estrella Duque, and Juan Luis Ramos. Insights into the susceptibility of pseudomonas putida to industrially relevant aromatic hydrocarbons that it can synthesize from sugars. Microbial Cell Factories, Feb 2023. URL: https://doi.org/10.1186/s12934-023-02028-y, doi:10.1186/s12934-023-02028-y. This article has 15 citations and is from a peer-reviewed journal.
-
(craig2021leveragingpseudomonasstress pages 7-9): Kelly Craig, Brant R. Johnson, and Amy Grunden. Leveraging pseudomonas stress response mechanisms for industrial applications. Frontiers in Microbiology, May 2021. URL: https://doi.org/10.3389/fmicb.2021.660134, doi:10.3389/fmicb.2021.660134. This article has 67 citations and is from a peer-reviewed journal.
-
(craig2021leveragingpseudomonasstress pages 5-6): Kelly Craig, Brant R. Johnson, and Amy Grunden. Leveraging pseudomonas stress response mechanisms for industrial applications. Frontiers in Microbiology, May 2021. URL: https://doi.org/10.3389/fmicb.2021.660134, doi:10.3389/fmicb.2021.660134. This article has 67 citations and is from a peer-reviewed journal.
-
(craig2021leveragingpseudomonasstress pages 11-12): Kelly Craig, Brant R. Johnson, and Amy Grunden. Leveraging pseudomonas stress response mechanisms for industrial applications. Frontiers in Microbiology, May 2021. URL: https://doi.org/10.3389/fmicb.2021.660134, doi:10.3389/fmicb.2021.660134. This article has 67 citations and is from a peer-reviewed journal.
-
(park2024unveilingthenovel pages 5-7): Joon Young Park, Minchang Jang, Sang-Mok Lee, Jihoon Woo, Eun-Jin Lee, and Donghyuk Kim. Unveiling the novel regulatory roles of rpod-family sigma factors in salmonella typhimurium heat shock response through systems biology approaches. Oct 2024. URL: https://doi.org/10.1371/journal.pgen.1011464, doi:10.1371/journal.pgen.1011464. This article has 12 citations and is from a domain leading peer-reviewed journal.
-
(park2024unveilingthenovel pages 4-5): Joon Young Park, Minchang Jang, Sang-Mok Lee, Jihoon Woo, Eun-Jin Lee, and Donghyuk Kim. Unveiling the novel regulatory roles of rpod-family sigma factors in salmonella typhimurium heat shock response through systems biology approaches. Oct 2024. URL: https://doi.org/10.1371/journal.pgen.1011464, doi:10.1371/journal.pgen.1011464. This article has 12 citations and is from a domain leading peer-reviewed journal.
-
(udaondo2024unravelingthegenomic pages 14-16): Zulema Udaondo, Juan-Luis Ramos, and Kaleb Z. Abram. Unraveling the genomic diversity of the pseudomonas putida group: exploring taxonomy, core pangenome, and antibiotic resistance mechanisms. FEMS Microbiology Reviews, Oct 2024. URL: https://doi.org/10.1093/femsre/fuae025, doi:10.1093/femsre/fuae025. This article has 13 citations and is from a domain leading peer-reviewed journal.
-
(follonier2013newinsightson media 43885585): Stéphanie Follonier, Isabel F Escapa, Pilar M Fonseca, Bernhard Henes, Sven Panke, Manfred Zinn, and María Auxiliadora Prieto. New insights on the reorganization of gene transcription in pseudomonas putida kt2440 at elevated pressure. Microbial Cell Factories, 12:30-30, Mar 2013. URL: https://doi.org/10.1186/1475-2859-12-30, doi:10.1186/1475-2859-12-30. This article has 46 citations and is from a peer-reviewed journal.
-
(dominguezcuevas2006transcriptionaltradeoffbetween pages 1-2): Patricia Domínguez-Cuevas, José-Eduardo González-Pastor, Silvia Marqués, Juan-Luis Ramos, and Víctor de Lorenzo. Transcriptional tradeoff between metabolic and stress-response programs in pseudomonas putida kt2440 cells exposed to toluene*. Journal of Biological Chemistry, 281:11981-11991, Apr 2006. URL: https://doi.org/10.1074/jbc.m509848200, doi:10.1074/jbc.m509848200. This article has 269 citations and is from a domain leading peer-reviewed journal.
Artifacts
Citations
- follonier2013newinsightson pages 5-6
- potvin2008sigmafactorsin pages 3-5
- ito2014geneticandphenotypic pages 2-3
- ito2014geneticandphenotypic pages 6-8
- ito2014geneticandphenotypic pages 1-2
- dominguezcuevas2006transcriptionaltradeoffbetween pages 7-8
- craig2021leveragingpseudomonasstress pages 7-9
- udaondo2024unravelingthegenomic pages 14-16
- dominguezcuevas2006transcriptionaltradeoffbetween pages 1-2
- garciafranco2023insightsintothe pages 9-10
- garciafranco2023insightsintothe pages 10-11
- craig2021leveragingpseudomonasstress pages 5-6
- craig2021leveragingpseudomonasstress pages 11-12
- park2024unveilingthenovel pages 5-7
- park2024unveilingthenovel pages 4-5
- https://doi.org/10.1186/1475-2859-12-30;
- https://doi.org/10.1002/mbo3.217
- https://doi.org/10.1002/mbo3.217;
- https://doi.org/10.1111/j.1574-6976.2007.00092.x
- https://doi.org/10.1186/1475-2859-12-30
- https://doi.org/10.1074/jbc.m509848200
- https://doi.org/10.1186/s12934-023-02028-y
- https://doi.org/10.1371/journal.pgen.1011464
- https://doi.org/10.1093/femsre/fuae025
- https://doi.org/10.1186/1475-2859-12-30,
- https://doi.org/10.1111/j.1574-6976.2007.00092.x,
- https://doi.org/10.1002/mbo3.217,
- https://doi.org/10.1074/jbc.m509848200,
- https://doi.org/10.1186/s12934-023-02028-y,
- https://doi.org/10.3389/fmicb.2021.660134,
- https://doi.org/10.1371/journal.pgen.1011464,
- https://doi.org/10.1093/femsre/fuae025,