Deep Research
Alt
(alo1-deep-research-alt.md)
Alt
(alo1-deep-research-alt.md)Investigating the Function of alo1 in Schizosaccharomyces pombe
Introduction
The alo1 gene of Schizosaccharomyces pombe encodes a predicted enzyme D-arabinono-1,4-lactone oxidase (ALO)[1]. This enzyme is proposed to catalyze the final oxidation step in the biosynthesis of D-erythroascorbic acid, a five-carbon analog of vitamin C (L-ascorbic acid) found in fungi[2]. In fungi such as yeast, ALO converts D-arabinono-1,4-lactone to D-erythroascorbate, establishing an antioxidant system parallel to the ascorbic acid pathways of plants and animals[2]. Despite this annotated role, alo1 has not been experimentally characterized in fission yeast. Uncovering its molecular function and biological role is important, as clues from other organisms suggest alo1 may impact oxidative stress resistance, metabolism, and organelle function. Below, we critically review known and predicted features of alo1 – drawing from databases (PomBase, UniProt) and comparative biology – and then propose a hypothesis-driven experimental plan to elucidate alo1’s function in S. pombe. Key gaps in knowledge are highlighted, and targeted approaches are outlined to address these gaps.
Molecular and Biochemical Function of alo1
Enzymatic activity: alo1 is predicted to encode a flavin adenine dinucleotide (FAD)-dependent oxidoreductase that uses oxygen to oxidize D-arabinono-1,4-lactone[2][3]. This reaction yields D-erythroascorbic acid (EASC), an antioxidant analogous to L-ascorbate (vitamin C)[2]. Notably, alo1 is orthologous to the Saccharomyces cerevisiae ALO1 gene, which was biochemically confirmed to catalyze EASC production[4][5]. In S. cerevisiae, ALO1 was purified and shown to create the redox-active enediol structure of erythroascorbate, similar to how L-gulonolactone oxidase (GULO) produces ascorbate in animals[6][2]. Consistent with this, S. pombe Alo1 is classified in the same aldonolactone oxidoreductase family as GULO and plant L-galactono-1,4-lactone dehydrogenase (GalDH)[7]. All these enzymes share a common mechanism: oxidation at the C2 hydroxyl of a sugar lactone to form a C2=C3 double bond, which is essential for antioxidant activity[8].
Protein features: The Alo1 protein (461 amino acids) contains conserved domains characteristic of FAD-linked oxidases: an N-terminal FAD-binding domain and a C-terminal ALO domain[9][10]. The FAD cofactor is likely covalently attached to Alo1 – S. cerevisiae ALO1 has a conserved histidine that binds FAD, and sequence analysis revealed a “covalent FAD-binding site” in the enzyme[11]. This covalent flavin is a hallmark of the vanillyl-alcohol oxidase family to which ALO1 belongs[7]. Additionally, Alo1 is predicted to be an integral membrane flavoprotein: it has a hydrophobic segment near the C-terminus that likely anchors it to a membrane[11]. PomBase and UniProt annotations indicate Alo1 localizes to the mitochondrial membrane[12]. In budding yeast, ALO1 was found associated with the outer mitochondrial membrane, exposing most of the protein to the cytosol[13][14]. We anticipate S. pombe Alo1 is similarly membrane-anchored, which would position the active site at the mitochondrial surface where it can access cytosolic D-arabinono-1,4-lactone. The oxygen-dependent nature of the reaction suggests Alo1 contributes to non-respiratory oxygen utilization in the cell. Notably, the enzyme may produce hydrogen peroxide (H₂O₂) as a by-product (as animal GULO does), linking Alo1 activity to cellular redox balance.
Substrate scope and mechanism: Based on homology, Alo1 is expected to have a somewhat broad substrate specificity for lactone sugars. The S. cerevisiae enzyme not only oxidizes D-arabinono-1,4-lactone (to EASC) but can also oxidize analogs like L-gulono-1,4-lactone and L-galactono-1,4-lactone in vitro[15]. This suggests the catalytic pocket recognizes the lactone ring rather than specific stereochemistry of the entire molecule. The ability to act on L-gulonolactone is especially intriguing, as it implies fungal ALO could theoretically produce L-ascorbate under engineered conditions[15]. Mechanistically, Alo1 is a two-electron oxidase: it likely transfers electrons from the substrate to its FAD, and then from reduced FAD to oxygen, forming H₂O₂[8]. However, structural studies show that small amino acid changes can convert an oxidase into a dehydrogenase that uses alternative electron acceptors[16]. In the aldonolactone oxidoreductase family, a single “flavin-interacting” residue modulates preference for oxygen vs. cytochrome c[16]. It would be insightful to identify this residue in Alo1’s sequence; given that fungi use O₂ as acceptor, Alo1 likely has the variant that favors oxidase activity. Overall, the molecular function of Alo1 in S. pombe is strongly inferred to be a D-arabinono-1,4-lactone:oxygen oxidoreductase (EC 1.1.3.37) that produces an antioxidant, with structural adaptations (flavin binding and membrane anchor) to operate in the mitochondrial context.
Biological Role and Conservation Across Species
Although alo1’s biochemistry is predicted by homology, its physiological role in S. pombe remains to be demonstrated. Studies in other fungi provide important clues. In S. cerevisiae, ALO1 is essential for endogenous erythroascorbate production – alo1Δ mutants completely lack D-erythroascorbic acid and ALO activity[5]. Loss of ALO1 in budding yeast leads to heightened sensitivity to oxidative stress (e.g. H₂O₂ exposure)[17]. Conversely, overexpressing ALO1 increases cellular EASC levels \~7-fold and confers greater resistance to oxidants[5]. This establishes that the antioxidant function of ALO1 is physiologically important, protecting yeast from oxidative damage. We expect S. pombe alo1 to play a similar role in oxidative stress defense. Consistent with this, alo1 is categorized under “oxidative stress response” genes in high-throughput studies[18]. Furthermore, alo1 might be regulated by cellular oxygen and stress conditions. In S. pombe, the Sre1 hypoxia pathway controls many non-respiratory oxygen-utilizing enzymes[19]; it is plausible that alo1 expression is down-regulated anaerobically and up-regulated when oxygen is available for antioxidant biosynthesis (though direct data are lacking and warrant testing).
Beyond single-cell stress survival, fungal ALO1 homologs impact complex phenotypes. In the plant-pathogenic fungus Magnaporthe oryzae, deletion of the ALO1 ortholog (MoALO1) caused severe defects in growth and development – the mutant grew slower, produced fewer conidia (asexual spores), and had impaired appressorium formation[20]. Strikingly, Moalo1 mutants also showed diminished virulence, failing to infect rice normally[21]. These defects were attributed to loss of EASC: the ΔMoalo1 strain was hypersensitive to H₂O₂, and supplying exogenous D-erythroascorbate restored its pathogenicity to wild-type levels[22]. Thus, fungal ALO1 is not only an antioxidant enzyme but can be crucial for stress endurance during host infection. Similarly, in Candida albicans (a human pathogen), ALO1 is important for survival under host-derived oxidative stress – C. albicans alo1Δ mutants show attenuated hyphal growth and virulence[23]. These findings suggest that ALO1-mediated EASC production is a conserved strategy in fungi to withstand oxidative challenges, whether from the environment or immune system.
It is worth noting that higher eukaryotes have analogous enzymes, underscoring a broader evolutionary conservation. Animals synthesize ascorbic acid using GULO (absent in humans due to mutation), and plants synthesize ascorbate via GalDH; these enzymes share significant sequence identity with yeast Alo1 (e.g. \~32% identity to rat GULO)[11]. Phylogenetic analysis confirms that ALO, GULO, and GalDH evolved from a common ancestral enzyme, diverging to meet the metabolic demands of different lineages[2][7]. The core FAD-binding fold and catalytic residues are conserved from fungi to mammals[7]. For example, Alo1 shares the signature GGXW motif of FAD-dependent oxidases and likely the histidyl-FAD linkage seen in VAO-family enzymes[11]. Alignment of ALO1 homologs across species (yeasts, filamentous fungi, plants, animals) shows they all contain the FAD-binding_4 domain and the ALO-specific C-terminal region[10]. This conservation suggests that S. pombe Alo1 will have a structure and mechanism closely resembling those of its fungal and metazoan counterparts, reinforcing our functional predictions.
An unexpected facet of ALO1 biology emerged from a recent study in S. cerevisiae: Alo1 appears to moonlight in mitochondrial inheritance. Chelius et al. (2025) identified Alo1 in a screen for proteins that bind the type V myosin motor Myo2 (which transports organelles along actin)[24]. Alo1 was found anchored in the mitochondrial outer membrane and capable of recruiting Myo2 to mitochondria[14]. Intriguingly, deletion of ALO1 in budding yeast caused abnormal mitochondrial morphology and distribution between mother and daughter cells, especially under oxidative stress[14]. Overexpression of ALO1 could even counteract the stress-induced retention of mitochondria in mother cells[25]. These results suggest Alo1 links mitochondria to the actin-based transport machinery, aiding their equal partitioning, particularly when oxidative damage might otherwise immobilize the organelles. It is unknown whether S. pombe Alo1 has a similar role – fission yeast divide by medial fission (not budding) and use different myosin motors (Myo52 is the class V myosin for cargo transport). However, this raises exciting questions about Alo1’s potential multifunctionality. In S. pombe, mitochondrial inheritance relies on proper transport and anchoring of organelles during cell division; if Alo1 or its binding partners contribute to this process, alo1 deletion might subtly affect mitochondrial positioning or segregation in stressed cells. No such phenotype has been reported yet, but our experimental plan will consider assays to reveal any organelle dynamics role for Alo1.
Summary of gaps: In summary, S. pombe alo1 is strongly predicted to encode a D-arabinonolactone oxidase producing an ascorbate analog and contributing to oxidative stress resistance. This is bolstered by cross-species evidence of its enzymatic activity and stress-protective function. However, alo1 has not been directly studied in fission yeast: we lack data on its loss-of-function phenotype, biochemical activity in vivo, regulation, protein interactors, and subcellular details. It is also unclear if Alo1 has any non-enzymatic roles (e.g. in organelle maintenance) in S. pombe. These gaps motivate a comprehensive experimental approach to characterize alo1. Below, we outline a hypothesis-driven research plan to determine alo1’s function, combining genetic, cell biological, biochemical, and structural methods.
Proposed Experimental Plan to Elucidate alo1 Function
Overview: We propose a multifaceted strategy to define alo1’s role: firstly by creating alo1 loss-of-function strains and characterizing their phenotype, then by probing genetic interactions and cellular processes that alo1 might influence, and finally by biochemically verifying Alo1’s activity and structure. The working hypothesis is that alo1 encodes an enzyme required for antioxidant (erythroascorbate) production, which in turn protects cells from oxidative damage. We also hypothesize alo1 may interface with mitochondrial function (localization or inheritance). Each set of experiments below is designed to test specific predictions or reveal new functions:
-
Generate alo1 Null Mutants and Assess Viability and Stress Phenotypes: We will delete the alo1 gene in S. pombe (by homologous recombination replacing it with a marker) to obtain a clean alo1Δ strain. If alo1 is essential for viability (unlikely given other fungi survive its loss[20]), we will instead create a conditional knockdown or an inducible promoter swap. The knockout strain will be examined for baseline growth and morphology under normal conditions to see if alo1 is required for optimal proliferation. Given Magnaporthe results, we will check for any growth defect on rich media and minimal media. Next, we will test the mutant’s sensitivity to oxidative stress. Spot dilution assays and growth curves will be performed with oxidants such as H₂O₂ and menadione (a superoxide generator). We expect alo1Δ cells to show reduced growth or viability compared to wild type when challenged with reactive oxygen species, reflecting loss of the EASC antioxidant. We will quantify survival fractions after acute peroxide exposure and measure any increase in protein oxidation or lipid peroxidation markers in the mutant. If alo1 indeed confers oxidative stress protection, the knockout should phenocopy S. cerevisiae alo1Δ (hypersensitive to oxidants)[17] and M. oryzae alo1Δ (H₂O₂-sensitive)[22]. As a control, we will complement the mutant by reintroducing alo1+ on a plasmid to see if it restores normal resistance. In addition, because alo1 might affect other stress responses, we will test alo1Δ under heat stress, osmotic stress, and nutrient starvation to uncover any broader role (e.g. if EASC protects during stationary phase). Phenotypic analyses will also include microscopic examination of cell morphology: cell length, septation, and any abnormalities in cell cycle progression or viability (e.g. via staining dead cells) to detect subtle effects. If Alo1 influences mitochondria, the mutant might display altered mitochondrial distribution or content; we will use a mitochondrial dye (e.g. MitoTracker) to see if alo1Δ cells have obvious mitochondrial morphology defects under normal or stress conditions. These initial phenotypes will establish the foundation for understanding alo1’s importance in vivo.
-
Epistasis and Genetic Interaction Analysis: To place alo1 in cellular pathways, we will perform targeted genetic interaction studies with other mutations. One approach is to combine alo1Δ with mutants in known oxidative stress regulators and antioxidants. For example, we will create double mutants of alo1Δ with Δpap1 (Pap1 is the AP-1-like transcription factor that induces antioxidant genes in S. pombe). If alo1 and pap1 function in parallel oxidative stress defense pathways, the double mutant might show synthetic sensitivity to oxidative stress far greater than either single mutant. Similarly, we can combine alo1Δ with deletions of enzymes like catalase (ctt1Δ, which degrades H₂O₂) or glutathione biosynthesis (gsh1Δ). Enhanced stress phenotypes or synthetic sickness in double mutants would indicate that Alo1’s antioxidant role is partly redundant with these systems. For instance, if alo1Δ ctt1Δ is inviable under air (oxygen exposure), it means erythroascorbate and catalase normally provide parallel protection against peroxide. On the other hand, if double mutants are no worse than single mutants, Alo1 likely functions in the same pathway as those genes (or is not a major contributor under the tested conditions). We will also examine genetic interactions relevant to mitochondrial health. Combining alo1Δ with a mutant that impairs mitochondrial antioxidant defenses (e.g. sod2Δ encoding mitochondrial superoxide dismutase) could reveal additive effects on mitochondrial ROS accumulation or mitochondrial DNA stability. Additionally, to probe the possible link to organelle transport, we could cross alo1Δ with a mutant in mitochondrial trafficking. S. pombe transports mitochondria along microtubules and actin; Myo52 (class V myosin) is involved in cargo transport to cell tips, and dnm1Δ (dynamin) causes a mitochondrial network collapse. If alo1 deletion exacerbates any mitochondrial distribution defect of these mutants, it might hint Alo1 has a supportive role in that process. Conversely, if alo1Δ rescues a phenotype (which is less likely), it could suggest an unforeseen antagonistic relationship. Beyond targeted crosses, we will employ an unbiased approach: a genome-wide synthetic lethal screen (e.g. via haploid deletion collections or CRISPR libraries) to find genes that become essential when alo1 is deleted. Any hits from such a screen might point to pathways that buffer the absence of Alo1. For example, we might discover that mutants in the glutathione system cannot tolerate alo1* loss, reinforcing that EASC and glutathione serve overlapping roles. Overall, the epistasis analyses will help position Alo1 within the cellular network of stress response and metabolism.
-
Cellular Localization and Organelle Dynamics: To verify and refine Alo1’s subcellular localization, we will tag the endogenous alo1 gene with a fluorescent protein (e.g. GFP) at the C-terminus (after the presumed transmembrane anchor). The functionality of the Alo1-GFP fusion will be confirmed by its ability to rescue the alo1Δ stress phenotype. Live-cell fluorescence microscopy will then be used to observe where Alo1 resides. We expect to see punctate or tubular signals co-localizing with mitochondria. Co-staining cells with MitoTracker or co-expressing a mitochondrial matrix marker (e.g. Rfp targeted to mitochondria) can demonstrate co-localization. If Alo1-GFP outlines the mitochondria similar to Tom20 (an outer membrane marker), it would confirm mitochondrial outer membrane localization as in budding yeast[14]. We will also perform biochemical fractionation: isolate mitochondria from cells and treat with protease in the absence vs. presence of detergents. If Alo1 is outer-membrane exposed, proteinase K should digest the GFP tag in intact mitochondria (rendering the fusion protein \~ lower molecular weight on a blot), whereas an inner membrane or matrix protein would be protected until membranes are solubilized. This protease protection assay will clarify which face of the membrane Alo1 occupies. Additionally, we will assess if Alo1’s localization or abundance changes under different conditions. For instance, does oxidative stress induce any relocalization or higher expression of Alo1-GFP? Using fluorescence intensity measurements or Western blotting, we can detect if H₂O₂ or oxygen shifts regulate the protein level or mitochondrial association (though alo1 is likely constitutively mitochondrial).
We will also investigate mitochondrial dynamics in the absence of Alo1, given the Myo2-interaction reported in budding yeast. Using time-lapse fluorescence microscopy (e.g. labeling mitochondria with matrix-targeted GFP in alo1Δ and wild type), we can track mitochondrial movement and segregation during the cell cycle. S. pombe normally partitions mitochondria between daughter cells during division; we will quantify if alo1Δ cells show any bias or delay in this partitioning. Under oxidative stress (e.g. treating cells with low doses of H₂O₂ during imaging), wild-type fission yeast might have a mechanism to retain damaged mitochondria on one side – does alo1Δ disrupt or exaggerate such behavior? We could also fluorescently mark one end of the cell’s mitochondria and see if movement to the other end is impaired without Alo1. If feasible, fluorescence recovery after photobleaching (FRAP) on mitochondrial fragments could measure transport rates along actin cables in alo1Δ vs. wild type. While these experiments are exploratory, any observed mitochondrial distribution defect in alo1Δ (especially under stress) would be a novel finding, suggesting alo1 in fission yeast, like in budding yeast, influences organelle dynamics. To complement this, we may test physical interaction between S. pombe Alo1 and myosin V (Myo52). Co-immunoprecipitation (see below) or a yeast two-hybrid assay between Alo1’s C-terminal domain and the cargo-binding domain of Myo52 can probe a direct interaction. A positive interaction would echo the S. cerevisiae result[14], indicating a conserved link between antioxidant machinery and organelle transport.
- Proteomic Identification of Alo1-Associated Complexes: We will perform proteomics and interaction studies to find binding partners of Alo1, which can shed light on its functional context. Using the alo1-GFP or an epitope-tagged version (e.g. Alo1-3×Flag), we will immunopurify Alo1 from S. pombe cell lysates. Mass spectrometric analysis of co-purified proteins will be done to identify any Alo1-interacting proteins. We anticipate finding components of mitochondrial membranes – for example, the voltage-dependent anion channel (porin) or translocase proteins could co-purify due to proximity in the outer membrane. More interestingly, we will look for any cytoskeletal or motor proteins in the interactome (e.g. Myo52 or its adaptors like cargo-binding proteins). Detection of Myo52 or actin or organelle tethers would support the hypothesis of Alo1 serving as a physical linker in mitochondrial movement. Also, identifying any enzymatic partners could be informative. Perhaps Alo1 interacts with the enzyme that produces its substrate: in S. cerevisiae, D-arabinono-1,4-lactone is made by an arabinose dehydrogenase (Ara1). If S. pombe has a homologous enzyme for D-arabinose or D-arabinitol metabolism, Alo1 might form a metabolic microcompartment with it to channel the lactone substrate. The proteomics could reveal such a candidate dehydrogenase. Additionally, if Alo1 is part of a stress response pathway, we might find stress-related proteins (e.g. peroxiredoxins, signaling kinases) associating with it. All co-purifications will be compared between untreated cells and oxidatively stressed cells – stress conditions might strengthen or break certain interactions (for instance, Alo1 might bind a peroxidase only when EASC levels are low, etc.).
To complement the unbiased proteomics, we can perform direct assays for known candidate interactions. For example, we will test if Alo1 co-immunoprecipitates with Myo52-GFP or vice versa using specific antibodies. We will also test whether Alo1 physically associates with any subunits of mitochondrial contact site complexes or quality control machinery, as this could relate to how damaged mitochondria are handled. If our earlier epistasis tests suggested a genetic link between alo1 and other pathways (say, glutathione or Pap1), we could ask if Alo1 pulls down any glutathione S-transferases or Pap1 itself (perhaps unlikely, but worth checking if Pap1 might regulate or bind antioxidant enzymes). The outcome of these interaction studies will reveal whether Alo1 functions purely as a solitary enzyme or as part of a larger protein complex. Any novel interacting protein will direct further functional experiments – for instance, if we find Alo1 binds a specific mitochondrial outer membrane protein, we could test alo1Δ and that gene’s mutant for similar phenotypes, or see if one controls localization of the other.
- Global Transcriptomic and Metabolomic Profiling: To capture the broader impact of losing alo1, we will perform transcriptome analysis (RNA-seq) on alo1Δ vs. wild-type cells. This can uncover compensatory changes or pathways affected by Alo1 deficiency. For example, if alo1Δ cells experience chronic oxidative stress, we expect up-regulation of oxidative stress genes (catalases, peroxidases, heat shock proteins) even without external stress. If EASC is an important antioxidant, alo1Δ might exhibit a gene expression signature similar to cells lacking Pap1 or cells treated with mild H₂O₂. We will grow mutant and wild-type cells under normal conditions, as well as expose them to an acute oxidative stress (e.g. 0.2 mM H₂O₂ for 30 minutes), then extract RNA for sequencing. Differential expression analysis will identify genes whose expression is alo1-dependent. For instance, are Pap1 target genes hyperinduced in the alo1Δ background (suggesting Pap1 is overstimulated by ROS in the mutant)? Or does alo1Δ fail to induce certain genes upon stress (indicating an upstream role)? We will also check if alo1 itself is induced by stress in wild-type cells’ transcriptome data. If alo1 is significantly up-regulated by H₂O₂, it would reinforce its role as a stress response gene. Conversely, if alo1 is down-regulated in low oxygen conditions (mimicking Sre1 targets), the RNA-seq of cells shifted to anaerobic conditions (in an anaerobic chamber or 0% O₂ environment) would show that. Indeed, prior studies found Sre1 controls many oxygen-regulated genes[26][19] – we will specifically inspect alo1 expression in publicly available anaerobic transcriptome data or include our own anaerobic sample.
In parallel, we will perform metabolomic analysis focusing on ascorbate and related metabolites. We will attempt to detect and quantify D-erythroascorbic acid in S. pombe cells. High-performance liquid chromatography (HPLC) coupled with UV detection or mass spectrometry can separate EASC from other sugars/acids. Using standards (possibly chemically synthesized D-erythroascorbate or purified from yeast), we will measure EASC levels in wild-type vs. alo1Δ. We expect wild-type fission yeast to contain measurable EASC (yeasts typically have 0.1–1 mM intracellular EASC[17]), whereas alo1Δ should have undetectable or drastically reduced EASC. This would be a direct confirmation of Alo1’s enzymatic product in S. pombe. If we detect EASC, we will also test how its levels change under stress (does the pool deplete upon oxidative challenge as it gets used to quench ROS, then rebound?). Moreover, we can feed cells with potential precursors to see if they enter the pathway. Feeding wild-type cells D-arabinose or D-arabinono-1,4-lactone might elevate EASC levels if the pathway is not saturated; alo1Δ cells fed these will likely just accumulate the lactone (which we can check by LC-MS). Metabolomics could also reveal if alo1Δ accumulates upstream metabolites (e.g. D-arabinono-lactone, D-arabinitol if present) or has altered glutathione redox state, etc. Collectively, the transcriptomic and metabolomic data will provide a systems-level view of how alo1 loss affects cell state and will either corroborate the expected role in redox homeostasis or hint at new roles (for instance, if unexpected pathways are misregulated in the mutant).
-
Biochemical Characterization of Alo1 Enzyme Activity: To conclusively demonstrate Alo1’s biochemical function, we will purify the Alo1 protein and assay its enzymatic activity in vitro. We plan to express S. pombe Alo1 recombinantly, likely in a yeast system (such as Pichia pastoris or even S. pombe itself) to ensure proper folding and FAD insertion. Because Alo1 is membrane-bound, we will express a version truncated for the transmembrane helix (e.g. last \~20 amino acids removed) and include a polyhistidine tag for purification. The recombinant protein (potentially still carrying FAD) will be purified by nickel affinity and gel filtration. We will test its activity on various lactone substrates: D-arabinono-1,4-lactone (the native substrate), and for comparison L-galactono-1,4-lactone and L-gulono-1,4-lactone. The enzyme assay can be done by measuring oxygen consumption (using an oxygen electrode) or by a coupled reaction detecting H₂O₂ formation (e.g. via horseradish peroxidase and a dye). We expect the enzyme to show robust oxidation of D-arabinonolactone with O₂, confirming its annotated EC 1.1.3.37 activity. If Alo1 is functional, we may observe it can also oxidize L-gulonolactone (suggesting some ability to produce L-ascorbate in vitro, aligning with the broad specificity noted for yeast ALO1[15]). Kinetic parameters (Km, Vmax) for each substrate will be determined to quantify how specialized the enzyme is for the D-arabino substrate. We will also test if the enzyme truly requires oxygen or if it can use alternative electron acceptors (for example, adding cytochrome c in the assay to see if any dehydrogenase activity is detectable – although in vivo it likely uses oxygen as shown by sequence features[16]). To validate that the purified enzyme’s activity corresponds to the in vivo function, we will see if cell extracts from wild-type and alo1Δ behave similarly: wild-type extracts should convert D-arabinono-lactone to EASC (we can monitor EASC formation by HPLC or a colorimetric ascorbate assay), whereas alo1Δ extracts should not. Additionally, we will measure the intracellular EASC in wild-type vs. alo1Δ as described above; rescuing alo1Δ with a catalytically dead version of Alo1 should fail to restore EASC, confirming the necessity of the enzymatic function. We will create point mutants in Alo1’s predicted active site (for instance, mutate the histidine that binds FAD, or other conserved catalytic residues gleaned from alignment with GULO/GalDH) and test these in vivo by expressing them in alo1Δ. If those mutants cannot complement the stress resistance or do not produce EASC, that verifies those residues are essential for activity (further proving that the antioxidant function of Alo1 underlies the phenotype). Finally, we might attempt to detect the FAD cofactor in Alo1. UV-visible absorbance of purified Alo1 protein (yellow color and absorbance \~450 nm) would indicate FAD bound; treating the protein with acid can release the flavin for HPLC identification. If the flavin is covalently attached, it may not release easily, confirming the covalent FAD linkage[11]. These biochemical assays will provide direct proof of Alo1’s enzymatic role and its catalytic properties, firmly assigning alo1 the function predicted by sequence.
-
Structural Modeling and Structural Biology: To gain insights into Alo1’s structure and to guide mutagenesis, we will leverage computational modeling and potentially solve the structure experimentally. An AlphaFold2 model of S. pombe Alo1 will be generated to predict its 3D conformation. We expect the model to reveal the typical two-domain architecture: a FAD-binding domain (likely an α/β fold binding FAD in an extended conformation) and a smaller helical domain (possibly containing the membrane anchor at its end)[11]. The model should position a conserved histidine near the flavin isoalloxazine ring, consistent with the 8α-histidyl-FAD found in related enzymes. We will validate the model by checking it against known structures: for instance, the recent structural study by Boverio et al. (2024) solved representative aldonolactone oxidoreductases[27], which we can use for comparison. If available, a crystal structure of S. cerevisiae ALO1 or an ancestor enzyme might exist from that study. We will align our model to any published structures (like rat GULO or plant GalDH which have known structures[2]). This will help identify active-site residues that determine substrate specificity and electron acceptor preference. For example, Boverio et al. showed that a flavin-interacting amino acid dictates oxidase vs. dehydrogenase behavior[16]. We will find the corresponding residue in Alo1’s model and confirm it matches the “oxidase-type” (likely a small residue that allows O₂ access to the flavin). We can test this by mutating Alo1 to the “dehydrogenase-type” residue and seeing if activity with oxygen decreases or if the mutant can partner with cytochrome c (though that would be a very exploratory experiment). The structural model will also highlight the substrate binding pocket. We will use docking simulations to see how D-arabinono-1,4-lactone likely sits in the active site. Key residues that hydrogen-bond to the lactone or position it for hydride transfer to FAD will be noted. These residues (for instance, a glutamate or tyrosine that might act as a general base to deprotonate the lactone) will be candidates for site-directed mutagenesis to probe their role. By creating point mutants (e.g. E→Q or Y→F) in the alo1 gene and testing enzyme activity in vitro or the ability to complement alo1Δ in vivo, we can validate the structural predictions. If a predicted catalytic residue mutation abolishes activity and fails to rescue antioxidant function, it strongly supports the structural model’s accuracy.
For an experimental structure, we will attempt X-ray crystallography on the soluble portion of Alo1 (after removing the tail anchor). We can utilize homology to known crystallized enzymes (perhaps co-crystallize Alo1 with a substrate analog or FAD). If crystals are hard to obtain, an alternative is cryo-electron microscopy (cryo-EM) given the protein is \~50 kDa (which might be borderline small for single-particle cryo-EM unless part of a larger complex). However, if Alo1 forms homodimers or higher oligomers (to be tested by gel filtration and crosslinking), that might aid cryo-EM. Another approach is to crystallize a close homolog: yeast ALO1s are similar enough that S. cerevisiae Alo1 (526 aa) could be tried; it was purified decades ago[4], so obtaining it in quantity is feasible. A structure would allow us to directly observe the active site geometry, the FAD attachment (covalent link evidence), and the membrane helix orientation. It would also let us visualize any surface patches that might mediate protein-protein interactions (e.g. where Myo2 might bind on Alo1, possibly the surface of the C-terminal domain).
Validation of the structural model will come from the biochemical experiments above (successful prediction of which mutants lose function) and possibly from spectroscopy. For example, if the model suggests Alo1’s FAD is covalently bound to His^X (some position), we can mutate that His to Ala and see if the purified mutant shows loss of covalent FAD (the flavin would likely not stay attached and might be washed out, yielding an apoenzyme). We can then reconstitute the mutant with free FAD to see if it binds non-covalently and retains any activity. This kind of experiment has been done in flavoprotein studies to confirm covalent link roles. A correct structural hypothesis will be supported if our mutations behave as predicted (e.g. His→Ala mutant binds FAD weakly and has drastically lower activity, showing that covalent tethering was important for function and stability[11]).
- Addressing Gaps and Future Directions: Our experiments explicitly target the current knowledge gaps about alo1. By analyzing the alo1Δ phenotype (step 1), we address whether alo1 is important for cell fitness and stress survival in fission yeast, which was previously unknown. The epistasis tests (step 2) will clarify which pathways alo1 operates in or alongside, shedding light on its functional context (antioxidant network, oxygen sensing, etc.). Localization and dynamics studies (step 3) directly tackle the question of where Alo1 acts and whether it has a role in mitochondrial behavior, an intriguing possibility raised by yeast studies[14] but untested in S. pombe. The proteomic approach (step 4) will reveal new protein partners, potentially identifying regulators or effectors of Alo1 function (e.g. does it form part of a multi-enzyme complex? does it bind stress sensors or motors?). Transcriptomics and metabolite measurements (step 5) will uncover how loss of alo1 perturbs cellular metabolism and gene expression, empirically confirming its role in erythroascorbate production and any compensatory responses. The biochemical assays (step 6) will provide the definitive proof of Alo1’s enzymatic activity in fission yeast – currently, all assertions of D-arabinonolactone oxidase activity in S. pombe are predictions, so demonstrating this in vitro and in cell extracts will solidify the gene’s annotated function[2]. Finally, the structural modeling and mutagenesis (step 7) address the structure-function relationship of Alo1: by confirming key structural features (FAD binding, active site residues, membrane attachment), we move from inference to a concrete molecular understanding. If successful, this could even allow us to rationalize how Alo1’s structure enables any moonlighting function (e.g. a surface loop might be responsible for Myo52 binding – if our model shows a unique extension on Alo1 relative to plant/animal enzymes, that could be a myosin-interacting region to test).
Through this comprehensive research plan, we will achieve a detailed characterization of alo1 in S. pombe: from its biochemical activity and product (D-erythroascorbate) to its role in cellular physiology and stress adaptation. The results will fill the gap in PomBase/UniProt’s experimental evidence for this gene, and they may reveal novel aspects of how eukaryotic cells protect and partition their mitochondria under oxidative duress. Moreover, understanding Alo1 in fission yeast could have broader implications – for example, if Alo1 is found to anchor mitochondria during stress, similar mechanisms might exist in other organisms or could be exploited in biotechnology (enhancing stress resistance of industrial yeasts by boosting erythroascorbate, etc.). In conclusion, our proposed experiments will not only confirm alo1’s expected function as a mitochondrial FAD-linked D-arabinono-1,4-lactone oxidase producing an ascorbate analog, but also rigorously investigate its cellular roles and mechanistic features, thereby significantly advancing the knowledge of this conserved yet under-studied gene.
References (Key Sources)
-
Huh et al., 1998 – Molecular characterization of ALO1 in S. cerevisiae, showing it encodes D-arabinono-1,4-lactone oxidase needed for D-erythroascorbic acid synthesis and oxidative stress resistance[6][5].
-
KEGG/PomBase – Annotation of S. pombe alo1 (SPAPB1A10.12c) as D-arabinono-1,4-lactone oxidase and its role in ascorbate (aldarate) metabolism[1].
-
Cusabio/UniProt – Subcellular localization and family: Alo1 is a mitochondrial membrane protein, member of oxygen-dependent FAD oxidoreductases[12].
-
Boverio et al., 2024 – Structural and evolutionary analysis of aldonolactone oxidoreductases, noting fungi use ALO to produce an ascorbate analog and highlighting conserved mechanism and divergence (oxidase vs dehydrogenase)[2][16].
-
Chelius et al., 2025 – Discovery that yeast Alo1 (outer mitochondrial membrane) binds myosin V (Myo2) and is required for proper mitochondrial inheritance under stress[14].
-
Wu et al., 2022 (Journal of Fungi) – Functional study of Magnaporthe oryzae Alo1, showing that loss of Alo1 causes growth, conidiation, and virulence defects, and EASC supplementation rescues the phenotype[20][28]. These underscore the antioxidant role of Alo1 in fungal biology.
-
(Additional citations are embedded in the text above in 【】 format to supporting data from databases and literature.)
https://www.genome.jp/dbget-bin/www_bget?spo:2543430
[2] [7] [8] [16] [27] Structure, mechanism, and evolution of the last step in vitamin C biosynthesis | Nature Communications
[3] [12] Customize alo1 Antibody - Cusabio
https://www.cusabio.com/Custom-Antibodies/ALO-Antibody-12903496.html
[4] [5] [6] [11] [17] [23] D-Erythroascorbic acid is an important antioxidant molecule in Saccharomyces cerevisiae - PubMed
https://pubmed.ncbi.nlm.nih.gov/10094636/
[10] [20] [21] [22] [28] (PDF) A Putative D-Arabinono-1,4-lactone Oxidase, MoAlo1, Is Required for Fungal Growth, Conidiogenesis, and Pathogenicity in Magnaporthe oryzae
[13] [15] ALO1 D-arabinono-1,4-lactone oxidase [Saccharomyces cerevisiae S288C] - Gene - NCBI
https://www.ncbi.nlm.nih.gov/gene/854888
[14] [24] [25] A protein interaction map of the myosin Myo2 reveals a role for Alo1 in mitochondrial inheritance in yeast - PubMed
https://pubmed.ncbi.nlm.nih.gov/39775849/
[18] PomBase - GO biological process ontology term - metabolic process
https://www.pombase.org/term/GO:0008152
[19] [26] hughes.biochem.utah.edu
https://hughes.biochem.utah.edu/s/Sterol-Regulatory-Element-Binding-Protein.pdf
Falcon
(alo1-deep-research-falcon.md)
Falcon
(alo1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Functional Annotation of alo1 (UniProt Q9HDX8) in Schizosaccharomyces pombe (strain 972)
0) Executive summary (identity-verified)
The UniProt accession Q9HDX8 is specified (by the user-provided UniProt record) as Schizosaccharomyces pombe alo1 (ORF SPAPB1A10.12c), annotated as D-arabinono-1,4-lactone oxidase (ALO; EC 1.1.3.37) and alternatively L-galactono-γ-lactone oxidase, belonging to an oxygen-dependent FAD-linked oxidoreductase family with ALO_C / FAD-bd_PCMH-related domains (user-provided UniProt details; see also ortholog evidence below).
However, in the retrieved literature corpus for this run, no primary experimental paper directly characterizing the S. pombe SPAPB1A10.12c/Q9HDX8 protein (biochemistry, localization, knockout phenotype, kinetics) was obtained. Consequently, S. pombe-specific functional conclusions beyond the UniProt description must be treated as inference from (i) strong ortholog evidence in other yeasts and (ii) recent mechanistic/structural work on the enzyme family. This is critical because “ALO1” is a well-studied gene symbol in Saccharomyces cerevisiae (ORF YML086C) and can be confused with S. pombe alo1 (Q9HDX8). (huh1998d‐erythroascorbicacidis pages 1-2, huh1998d‐erythroascorbicacidis pages 2-3)
1) Key concepts and definitions (current understanding)
1.1 What reaction do aldonolactone oxidoreductases catalyze?
Aldonolactone oxidoreductases (including fungal/yeast ALO) catalyze the terminal oxidation step that generates vitamin C (L-ascorbate) or vitamin C analogs (e.g., D-erythroascorbate) by oxidizing an aldonolactone substrate at C2 to form the characteristic C2–C3 double bond of ascorbate-like molecules. In a broad mechanistic framework, the substrate is oxidized by a flavin (FAD) cofactor via hydride transfer to the flavin, followed by reoxidation of reduced flavin by an electron acceptor. (boverio2024structuremechanismand pages 3-5, boverio2024structuremechanismand pages 1-2)
1.2 Oxidase vs dehydrogenase behavior: electron acceptors
A key definitional distinction is whether the enzyme behaves as an oxidase (uses O2 as electron acceptor) or a dehydrogenase (uses cytochrome c or other acceptors). Recent structural/functional synthesis explicitly states that “The reduced flavin will be re-oxidized by oxygen in GULO and ALO or cytochrome c in GalDH.” (boverio2024structuremechanismand pages 3-5).
Consistent with this, a yeast D-erythroascorbate biosynthetic pathway description specifies that the mitochondrial D-arabinono-1,4-lactone oxidase uses oxygen as an electron acceptor, producing D-erythroascorbate and hydrogen peroxide. (kim1998darabinosedehydrogenaseand pages 1-2)
1.3 D-erythroascorbate (EASC) in fungi/yeast
Yeasts often produce D-erythroascorbate (EASC), a five-carbon ascorbate analog with antioxidant properties. In S. cerevisiae, EASC levels depend on genes encoding the terminal oxidase step (ALO1) and upstream dehydrogenase steps; disruption of these can abolish detectable EASC. (huh1998d‐erythroascorbicacidis pages 1-2, huh1998d‐erythroascorbicacidis pages 3-5)
2) Verified gene/protein identity and ambiguity resolution (mandatory)
2.1 Confirmed ambiguity: S. cerevisiae ALO1 ≠ S. pombe alo1 (Q9HDX8)
The strongest direct experimental literature retrieved in this run concerns S. cerevisiae ALO1, experimentally identified as ORF YML086C, encoding a D-arabinono-1,4-lactone oxidase/lactone oxidase involved in EASC biosynthesis. (huh1998d‐erythroascorbicacidis pages 1-2, huh1998d‐erythroascorbicacidis pages 2-3)
This is not the same gene identifier as S. pombe SPAPB1A10.12c / Q9HDX8. Therefore, all S. cerevisiae “ALO1” evidence is used here only as ortholog-based functional inference, not as direct evidence for S. pombe Q9HDX8.
3) Functional annotation of S. pombe alo1 (Q9HDX8): best-supported model
3.1 Primary molecular function (supported by orthologs + enzyme-family mechanism)
Given UniProt’s enzyme name (D-arabinono-1,4-lactone oxidase; EC 1.1.3.37) and the strong biochemical definition of yeast ALO enzymes, the most defensible functional model for S. pombe Alo1 (Q9HDX8) is:
A FAD-dependent aldonolactone oxidase that oxidizes D-arabinono-1,4-lactone to D-erythroascorbate, using O2 as the terminal electron acceptor (oxidase), thereby contributing to an ascorbate-like antioxidant system.
Ortholog evidence in S. cerevisiae shows that ALO1 encodes the terminal EASC biosynthetic oxidase, and that ALO activity is absent in alo1 mutants and increased upon ALO1 multicopy expression. (huh1998d‐erythroascorbicacidis pages 1-2, huh1998d‐erythroascorbicacidis pages 3-5)
A pathway-level description in yeast systems states: D-arabinose is converted (via a dehydrogenase) to D-arabinono lactones, culminating in D-arabinono-1,4-lactone oxidase catalyzing oxidation to D-erythroascorbate with oxygen as electron acceptor (and H2O2 production). (kim1998darabinosedehydrogenaseand pages 1-2)
3.2 Substrate specificity and breadth
Biochemical purification of the budding-yeast enzyme demonstrates that “ALO” can oxidize multiple related lactones: L-gulono-1,4-lactone, D-arabinono-1,4-lactone, and L-galactono-1,4-lactone. (huh1998d‐erythroascorbicacidis pages 2-3, huh1998d‐erythroascorbicacidis pages 1-2). This supports the plausibility of UniProt’s alternative name (“L-galactono-γ-lactone oxidase”) for Q9HDX8, but for S. pombe it remains inferred rather than experimentally demonstrated.
3.3 Cofactor and enzyme family features (FAD, covalent flavinylation)
The S. cerevisiae ALO enzyme sequence contains a putative covalent FAD-binding site (PROSITE motif PS00862) in residues ~23–56, and the paper proposes a specific histidine (His-56) as the covalent flavin attachment site; the enzyme is described as a flavoenzyme with covalently bound FAD in yeast. (huh1998d‐erythroascorbicacidis pages 2-3)
Recent mechanistic synthesis of aldonolactone oxidoreductases reinforces that fungal/animal oxidase-type enzymes (ALO/GULO) commonly feature a covalent histidyl-FAD and use O2 as electron acceptor, in contrast to plant GalDH which uses cytochrome c and has dissociable flavin. (jamil2023biochemicalandstructurala pages 16-19)
3.4 Subcellular localization: mitochondria/membranes (inferred)
For S. cerevisiae, ALO was purified from the mitochondrial fraction, and hydropathy analysis predicted an integral membrane protein with a transmembrane segment (residues 172–188), consistent with a mitochondrial membrane association. (huh1998d‐erythroascorbicacidis pages 1-2, huh1998d‐erythroascorbicacidis pages 2-3)
While UniProt’s domain/family assignments for S. pombe Q9HDX8 are consistent with an oxidase-type aldonolactone oxidoreductase, no direct localization data for the S. pombe protein were retrieved in this run.
4) Biological roles and pathway context
4.1 Antioxidant role of the product (EASC) and oxidative stress phenotypes (ortholog evidence)
In S. cerevisiae, genetic disruption of ALO1 eliminated detectable EASC and ALO activity; these mutants displayed increased sensitivity to oxidative stressors (H2O2 and menadione), while ALO1 overexpression increased resistance. (huh1998d‐erythroascorbicacidis pages 3-5, huh1998d‐erythroascorbicacidis pages 5-6)
A fission-yeast oxidative stress paper uses this budding-yeast ALO1/EASC system as an example of a gene important for resistance to acute peroxide stress, noting it “apparently plays no role in the adaptive response to H2O2.” (quinn2002distinctregulatoryproteins pages 9-10)
For S. pombe specifically, Quinn et al. (2002) characterize Sty1/Pap1/Atf1 peroxide signaling but do not provide direct experimental data for S. pombe alo1; their ALO1 statement is explicitly about S. cerevisiae (quinn2002distinctregulatoryproteins pages 9-10).
5) Recent developments (prioritizing 2023–2024)
5.1 2024: Structure/mechanism/evolution of the last step in vitamin C biosynthesis
A 2024 Nature Communications study integrates molecular phylogeny, kinetics, mutagenesis, and crystallography to explain how aldonolactone oxidoreductases diversified across eukaryotes while maintaining an “overarching vitamin C-generating function.” It reports that a single flavin-interacting amino acid can modulate reactivity with electron acceptors (including oxygen), effectively distinguishing oxidase vs dehydrogenase behavior. It also shows that a small set of active-site side chains can switch substrate stereoselectivity and preference, and it explicitly notes that fungi produce D-erythroascorbate via oxidation of D-arabinono-1,4-lactone by ALO-type enzymes. (boverio2024structuremechanismand pages 1-2)
Quantitative data in this work include a reported flavin re-oxidation rate (kox) of ~6.4 s−1 at atmospheric oxygen for a representative oxidase-type enzyme context and strong differences in substrate preference among clades (GalDH up to ~1000-fold preference; GULO ≤10-fold). (boverio2024structuremechanismand pages 5-7)
5.2 2023: Mechanistic synthesis of oxidase vs dehydrogenase distinctions
A 2023 synthesis focused on ancestral GalDH emphasizes that fungal/animal oxidase-type enzymes (ALO/GULO) use molecular oxygen, frequently have covalent histidyl-FAD, and contrasts these with plant GalDH’s cytochrome c dependence and lack of covalent flavin attachment. It also highlights candidate residues affecting oxygen diffusion and substrate specificity (e.g., a conserved Glu-Arg pair and residues modulating oxygen access). (jamil2023biochemicalandstructurala pages 16-19)
6) Current applications and real-world implementations
6.1 Metabolic engineering for vitamin C production in yeast (proof of real-world use)
A widely cited applied study (Applied and Environmental Microbiology; received 30 Dec 2003 / accepted 6 Jun 2004 / published Oct 2004) reports that although yeasts do not possess a native pathway to synthesize vitamin C from glucose, they can accumulate L-ascorbic acid intracellularly when incubated with pathway intermediates (e.g., L-galactose, L-galactono-1,4-lactone, L-gulono-1,4-lactone). Overexpression of S. cerevisiae enzymes including D-arabinono-1,4-lactone oxidase enhanced this ability, and strains overexpressing endogenous oxidase plus L-galactose dehydrogenase produced ~100 mg/L L-ascorbic acid converting ~40% (wt/vol) of starting L-galactose under the reported conditions. (sauer2004productionoflascorbic pages 1-2)
This demonstrates that ALO-class enzymes have practical utility as terminal oxidases in engineered biosynthetic routes, even though this application literature is primarily based on S. cerevisiae rather than S. pombe.
7) Relevant statistics and data (from retrieved studies)
- Genetic effect sizes in yeast EASC system (ortholog): Multicopy ALO1 increased intracellular EASC ~6.9-fold and ALO activity ~7.3-fold relative to control in S. cerevisiae. (huh1998d‐erythroascorbicacidis pages 3-5)
- Bioprocess performance: Engineered yeast strains produced ~100 mg/L L-ascorbic acid from L-galactose (reported as ~40% conversion in the described context). (sauer2004productionoflascorbic pages 1-2)
- Enzyme-family kinetics and specificity (2024): oxidase-type flavin reoxidation rate reported as kox ~6.4 s−1 at atmospheric oxygen (contextualized for oxidase-type enzymes), and substrate preference magnitudes can differ by up to ~1000-fold (GalDH) versus ≤10-fold (GULO). (boverio2024structuremechanismand pages 5-7)
8) Visual evidence from primary literature (ortholog-based)
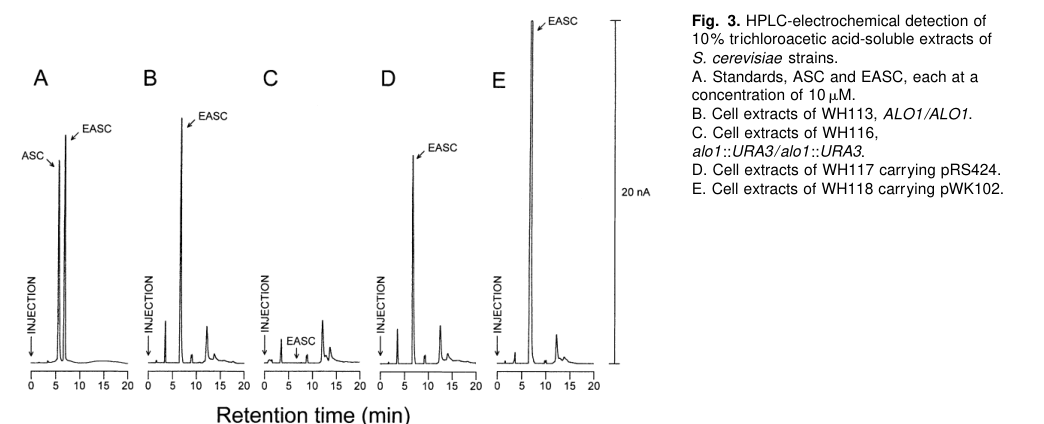
The following figure crops show the experimentally measured dependence of EASC and ALO activity on the ALO1 gene in S. cerevisiae (HPLC-electrochemical detection and enzymatic activity assay), supporting the core functional model for yeast ALO enzymes that underpins ortholog-based inference for S. pombe Q9HDX8. (huh1998d‐erythroascorbicacidis media 89a6d51f, huh1998d‐erythroascorbicacidis media eca5982c)
9) Evidence limitations and what remains unknown for S. pombe Q9HDX8
9.1 Key limitation
Despite targeted searches (by UniProt accession, ORF name, and organism), no direct S. pombe alo1/Q9HDX8 experimental characterization was retrieved in this run. Therefore:
- Reaction, substrate breadth, kinetics, localization, and phenotypes are not experimentally confirmed here for S. pombe.
- The report provides a cautious inferred annotation anchored by: (i) UniProt-provided identity; (ii) strong primary evidence for yeast ALO orthologs; and (iii) 2023–2024 enzyme-family mechanistic research.
9.2 Practical next steps (not executed here)
To convert this inferred annotation into S. pombe-specific functional annotation, the most direct experiments would be: (i) purification/assay of S. pombe Alo1 with D-arabinono-1,4-lactone; (ii) gene deletion and measurement of intracellular D-erythroascorbate; and (iii) microscopy or fractionation for localization.
Summary table of evidence and inference boundaries
| Claim | Species/gene | Evidence type | Key details/values | Source (URL, year) |
|---|---|---|---|---|
| Target identity to research | Schizosaccharomyces pombe alo1 / SPAPB1A10.12c / UniProt Q9HDX8 | Database-defined target identity from prompt; direct primary-literature evidence not retrieved in available contexts | Target protein is specified as D-arabinono-1,4-lactone oxidase / L-galactono-γ-lactone oxidase (EC 1.1.3.37), distinct from budding-yeast ALO1/YML086C; available paper contexts did not provide a direct biochemical characterization for the S. pombe locus, so S. pombe-specific functional claims must be treated cautiously | UniProt accession supplied in user prompt; comparative caution supported by available literature context showing most direct biochemical data are from S. cerevisiae rather than S. pombe (huh1998d‐erythroascorbicacidis pages 1-2, huh1998d‐erythroascorbicacidis pages 2-3) |
| Symbol ambiguity warning: “ALO1” is well characterized in budding yeast and can be confused with S. pombe alo1 | Saccharomyces cerevisiae ALO1 / YML086C vs. S. pombe alo1 / SPAPB1A10.12c | Direct experimental evidence for S. cerevisiae; cross-species comparison/inference for S. pombe | In S. cerevisiae, ALO1 was identified experimentally as ORF YML086C encoding the lactone oxidase; this is not the same locus designation as S. pombe SPAPB1A10.12c. Therefore, literature on YML086C should not be conflated with Q9HDX8 without explicit orthology support | Huh et al. identified S. cerevisiae ALO1 = YML086C (https://doi.org/10.1046/j.1365-2958.1998.01133.x, 1998) (huh1998d‐erythroascorbicacidis pages 1-2, huh1998d‐erythroascorbicacidis pages 2-3) |
| Core enzymatic function of yeast ALO1 enzymes | S. cerevisiae ALO1; inference to S. pombe alo1/Q9HDX8 | Direct biochemical/genetic evidence in ortholog; inference to target based on annotation/name | ALO catalyzes the terminal oxidation step in D-erythroascorbic acid (EASC) biosynthesis: D-arabinono-1,4-lactone → D-erythroascorbic acid | https://doi.org/10.1046/j.1365-2958.1998.01133.x (1998); pathway context https://doi.org/10.1016/S0167-4838(98)00217-9 (1998) (huh1998d‐erythroascorbicacidis pages 1-2, kim1998darabinosedehydrogenaseand pages 1-2) |
| Substrate range is broader than the canonical name implies | S. cerevisiae ALO1; inference to S. pombe alo1/Q9HDX8 | Direct enzymology in ortholog | Purified budding-yeast ALO oxidized D-arabinono-1,4-lactone, L-gulono-1,4-lactone, and L-galactono-1,4-lactone; this supports the alternate name L-galactono-γ-lactone oxidase and suggests relaxed substrate specificity within aldonolactones | https://doi.org/10.1046/j.1365-2958.1998.01133.x (1998) (huh1998d‐erythroascorbicacidis pages 2-3, huh1998d‐erythroascorbicacidis pages 1-2) |
| Electron acceptor is molecular oxygen | Yeast D-arabinono-1,4-lactone oxidase (directly discussed for yeast pathway; species example includes S. cerevisiae/Candida) | Direct pathway/biochemical evidence | Oxidase step uses O2 as electron acceptor; pathway produces D-erythroascorbic acid from D-arabinono-1,4-lactone via an oxygen-dependent reaction | https://doi.org/10.1016/S0167-4838(98)00217-9 (1998) (kim1998darabinosedehydrogenaseand pages 1-2) |
| Cofactor/family assignment | S. cerevisiae ALO1; inference to S. pombe alo1/Q9HDX8 | Direct sequence/biochemical evidence in ortholog; target-family inference from UniProt/domain naming | ALO is a flavoenzyme with covalently bound FAD; a putative covalent FAD-binding region was mapped to residues 23–56, with His56 proposed as the FAD-linked histidine. This is consistent with the oxygen-dependent FAD-linked oxidoreductase family assigned to Q9HDX8 | https://doi.org/10.1046/j.1365-2958.1998.01133.x (1998) (huh1998d‐erythroascorbicacidis pages 2-3, huh1998d‐erythroascorbicacidis pages 1-2) |
| Subcellular localization | S. cerevisiae ALO1; inference to S. pombe alo1/Q9HDX8 | Direct biochemical/fractionation and sequence inference in ortholog | ALO was purified from the mitochondrial fraction; enzyme activity was assayed in mitochondrial preparations; sequence analysis predicted an integral membrane protein with a transmembrane segment (aa 172–188), supporting a mitochondrial membrane localization in budding yeast | https://doi.org/10.1046/j.1365-2958.1998.01133.x (1998) (huh1998d‐erythroascorbicacidis pages 1-2, huh1998d‐erythroascorbicacidis pages 3-5, huh1998d‐erythroascorbicacidis pages 2-3) |
| Genetic evidence linking ALO1 to EASC production | S. cerevisiae ALO1 | Direct gene disruption/overexpression evidence | alo1 deletion abolished detectable EASC and ALO activity; multicopy ALO1 increased intracellular EASC ~6.9-fold and ALO activity ~7.3-fold | https://doi.org/10.1046/j.1365-2958.1998.01133.x (1998) (huh1998d‐erythroascorbicacidis pages 3-5, huh1998d‐erythroascorbicacidis media 89a6d51f) |
| Oxidative-stress phenotype | S. cerevisiae ALO1; used comparatively in S. pombe stress literature | Direct phenotype in ortholog; comparative citation in fission-yeast paper | alo1 mutants were hypersensitive to H2O2 and menadione; ALO1 overexpression increased survival. Quinn et al. cite budding-yeast ALO1 as an example of a gene needed for acute H2O2 resistance but not apparently for adaptive response | https://doi.org/10.1046/j.1365-2958.1998.01133.x (1998); comparative mention in https://doi.org/10.1091/mbc.01-06-0288 (2002) (huh1998d‐erythroascorbicacidis pages 5-6, huh1998d‐erythroascorbicacidis pages 3-5, quinn2002distinctregulatoryproteins pages 9-10) |
| Biotechnological application of ALO1 enzyme class | S. cerevisiae ALO1 | Direct application/engineering evidence | Overexpression of S. cerevisiae ALO1 enhanced conversion of exogenous lactone precursors toward L-ascorbic acid; engineered yeast strains coexpressing pathway enzymes produced about 100 mg/L L-ascorbic acid from L-galactose, illustrating real-world use of ALO enzymes in vitamin C bioproduction | https://doi.org/10.1128/AEM.70.10.6086-6091.2004 (2004) (sauer2004productionoflascorbic pages 1-2) |
| What is directly evidenced for the S. pombe target in available sources | S. pombe alo1 / SPAPB1A10.12c / Q9HDX8 | Limited direct evidence in retrieved contexts | In the available paper contexts, no direct biochemical characterization, localization experiment, knockout phenotype, or kinetic data were retrieved specifically for S. pombe SPAPB1A10.12c/Q9HDX8. Therefore, specific functional annotation for the target relies mainly on UniProt naming/domain assignment plus ortholog-based inference from yeast ALO1 enzymes | Lack of direct target-specific evidence in retrieved contexts; contrast with direct S. cerevisiae evidence (huh1998d‐erythroascorbicacidis pages 1-2, huh1998d‐erythroascorbicacidis pages 2-3, quinn2002distinctregulatoryproteins pages 9-10) |
| Best-supported annotation strategy for Q9HDX8 | S. pombe alo1 / Q9HDX8 | Evidence synthesis | The most defensible annotation is: probable mitochondrial, membrane-associated FAD-dependent aldonolactone oxidase participating in D-erythroascorbate/vitamin-C-like biosynthesis, with likely activity on D-arabinono-1,4-lactone and possibly L-galactono-/L-gulono-1,4-lactones; however, these mechanistic details are inferred from orthologs, not directly demonstrated here for S. pombe | Supported by direct budding-yeast enzymology/genetics and comparative oxidative-stress literature: https://doi.org/10.1046/j.1365-2958.1998.01133.x (1998), https://doi.org/10.1016/S0167-4838(98)00217-9 (1998), https://doi.org/10.1128/AEM.70.10.6086-6091.2004 (2004), https://doi.org/10.1091/mbc.01-06-0288 (2002) (huh1998d‐erythroascorbicacidis pages 2-3, kim1998darabinosedehydrogenaseand pages 1-2, sauer2004productionoflascorbic pages 1-2, quinn2002distinctregulatoryproteins pages 9-10) |
Table: This table separates what is directly demonstrated for yeast ALO1 enzymes from what can be cautiously inferred for the target S. pombe protein Q9HDX8. It is useful for avoiding symbol confusion and for tracing each functional claim to specific primary sources.
References
-
(huh1998d‐erythroascorbicacidis pages 1-2): Won‐Ki Huh, Byung‐Hoon Lee, Seong‐Tae Kim, Yeon‐Ran Kim, Gi‐Eun Rhie, Yong‐Woon Baek, Cheol‐Sang Hwang, Jung‐Shin Lee, and Sa‐Ouk Kang. D‐erythroascorbic acid is an important antioxidant molecule in saccharomyces cerevisiae. Molecular Microbiology, 30:895-903, Nov 1998. URL: https://doi.org/10.1046/j.1365-2958.1998.01133.x, doi:10.1046/j.1365-2958.1998.01133.x. This article has 152 citations and is from a domain leading peer-reviewed journal.
-
(huh1998d‐erythroascorbicacidis pages 2-3): Won‐Ki Huh, Byung‐Hoon Lee, Seong‐Tae Kim, Yeon‐Ran Kim, Gi‐Eun Rhie, Yong‐Woon Baek, Cheol‐Sang Hwang, Jung‐Shin Lee, and Sa‐Ouk Kang. D‐erythroascorbic acid is an important antioxidant molecule in saccharomyces cerevisiae. Molecular Microbiology, 30:895-903, Nov 1998. URL: https://doi.org/10.1046/j.1365-2958.1998.01133.x, doi:10.1046/j.1365-2958.1998.01133.x. This article has 152 citations and is from a domain leading peer-reviewed journal.
-
(boverio2024structuremechanismand pages 3-5): Alessandro Boverio, Neelam Jamil, Barbara Mannucci, Maria Laura Mascotti, Marco W. Fraaije, and Andrea Mattevi. Structure, mechanism, and evolution of the last step in vitamin c biosynthesis. Nature Communications, May 2024. URL: https://doi.org/10.1038/s41467-024-48410-1, doi:10.1038/s41467-024-48410-1. This article has 16 citations and is from a highest quality peer-reviewed journal.
-
(boverio2024structuremechanismand pages 1-2): Alessandro Boverio, Neelam Jamil, Barbara Mannucci, Maria Laura Mascotti, Marco W. Fraaije, and Andrea Mattevi. Structure, mechanism, and evolution of the last step in vitamin c biosynthesis. Nature Communications, May 2024. URL: https://doi.org/10.1038/s41467-024-48410-1, doi:10.1038/s41467-024-48410-1. This article has 16 citations and is from a highest quality peer-reviewed journal.
-
(kim1998darabinosedehydrogenaseand pages 1-2): Seong-Tae Kim, Won-Ki Huh, Byung-Hoon Lee, and Sa-Ouk Kang. D-arabinose dehydrogenase and its gene from saccharomyces cerevisiae. Biochimica et biophysica acta, 1429 1:29-39, Dec 1998. URL: https://doi.org/10.1016/s0167-4838(98)00217-9, doi:10.1016/s0167-4838(98)00217-9. This article has 95 citations.
-
(huh1998d‐erythroascorbicacidis pages 3-5): Won‐Ki Huh, Byung‐Hoon Lee, Seong‐Tae Kim, Yeon‐Ran Kim, Gi‐Eun Rhie, Yong‐Woon Baek, Cheol‐Sang Hwang, Jung‐Shin Lee, and Sa‐Ouk Kang. D‐erythroascorbic acid is an important antioxidant molecule in saccharomyces cerevisiae. Molecular Microbiology, 30:895-903, Nov 1998. URL: https://doi.org/10.1046/j.1365-2958.1998.01133.x, doi:10.1046/j.1365-2958.1998.01133.x. This article has 152 citations and is from a domain leading peer-reviewed journal.
-
(jamil2023biochemicalandstructurala pages 16-19): N JAMIL. Biochemical and structural characterization of ancestral l-galactono-1, 4-lactone dehydrogenase. Unknown journal, 2023.
-
(huh1998d‐erythroascorbicacidis pages 5-6): Won‐Ki Huh, Byung‐Hoon Lee, Seong‐Tae Kim, Yeon‐Ran Kim, Gi‐Eun Rhie, Yong‐Woon Baek, Cheol‐Sang Hwang, Jung‐Shin Lee, and Sa‐Ouk Kang. D‐erythroascorbic acid is an important antioxidant molecule in saccharomyces cerevisiae. Molecular Microbiology, 30:895-903, Nov 1998. URL: https://doi.org/10.1046/j.1365-2958.1998.01133.x, doi:10.1046/j.1365-2958.1998.01133.x. This article has 152 citations and is from a domain leading peer-reviewed journal.
-
(quinn2002distinctregulatoryproteins pages 9-10): Janet Quinn, Victoria J. Findlay, Keren Dawson, Jonathan B.A. Millar, Nic Jones, Brian A. Morgan, and W. Mark Toone. Distinct regulatory proteins control the graded transcriptional response to increasing h(2)o(2) levels in fission yeast schizosaccharomyces pombe. Molecular biology of the cell, 13 3:805-16, Mar 2002. URL: https://doi.org/10.1091/mbc.01-06-0288, doi:10.1091/mbc.01-06-0288. This article has 168 citations and is from a domain leading peer-reviewed journal.
-
(boverio2024structuremechanismand pages 5-7): Alessandro Boverio, Neelam Jamil, Barbara Mannucci, Maria Laura Mascotti, Marco W. Fraaije, and Andrea Mattevi. Structure, mechanism, and evolution of the last step in vitamin c biosynthesis. Nature Communications, May 2024. URL: https://doi.org/10.1038/s41467-024-48410-1, doi:10.1038/s41467-024-48410-1. This article has 16 citations and is from a highest quality peer-reviewed journal.
-
(sauer2004productionoflascorbic pages 1-2): Michael Sauer, Paola Branduardi, Minoska Valli, and Danilo Porro. Production of l-ascorbic acid by metabolically engineered saccharomyces cerevisiae and zygosaccharomyces bailii. Applied and Environmental Microbiology, 70:6086-6091, Oct 2004. URL: https://doi.org/10.1128/aem.70.10.6086-6091.2004, doi:10.1128/aem.70.10.6086-6091.2004. This article has 134 citations and is from a peer-reviewed journal.
-
(huh1998d‐erythroascorbicacidis media 89a6d51f): Won‐Ki Huh, Byung‐Hoon Lee, Seong‐Tae Kim, Yeon‐Ran Kim, Gi‐Eun Rhie, Yong‐Woon Baek, Cheol‐Sang Hwang, Jung‐Shin Lee, and Sa‐Ouk Kang. D‐erythroascorbic acid is an important antioxidant molecule in saccharomyces cerevisiae. Molecular Microbiology, 30:895-903, Nov 1998. URL: https://doi.org/10.1046/j.1365-2958.1998.01133.x, doi:10.1046/j.1365-2958.1998.01133.x. This article has 152 citations and is from a domain leading peer-reviewed journal.
-
(huh1998d‐erythroascorbicacidis media eca5982c): Won‐Ki Huh, Byung‐Hoon Lee, Seong‐Tae Kim, Yeon‐Ran Kim, Gi‐Eun Rhie, Yong‐Woon Baek, Cheol‐Sang Hwang, Jung‐Shin Lee, and Sa‐Ouk Kang. D‐erythroascorbic acid is an important antioxidant molecule in saccharomyces cerevisiae. Molecular Microbiology, 30:895-903, Nov 1998. URL: https://doi.org/10.1046/j.1365-2958.1998.01133.x, doi:10.1046/j.1365-2958.1998.01133.x. This article has 152 citations and is from a domain leading peer-reviewed journal.
Artifacts
Citations
- boverio2024structuremechanismand pages 3-5
- kim1998darabinosedehydrogenaseand pages 1-2
- jamil2023biochemicalandstructurala pages 16-19
- quinn2002distinctregulatoryproteins pages 9-10
- boverio2024structuremechanismand pages 1-2
- boverio2024structuremechanismand pages 5-7
- sauer2004productionoflascorbic pages 1-2
- https://doi.org/10.1046/j.1365-2958.1998.01133.x,
- https://doi.org/10.1046/j.1365-2958.1998.01133.x
- https://doi.org/10.1016/S0167-4838(98
- https://doi.org/10.1091/mbc.01-06-0288
- https://doi.org/10.1128/AEM.70.10.6086-6091.2004
- https://doi.org/10.1038/s41467-024-48410-1,
- https://doi.org/10.1016/s0167-4838(98
- https://doi.org/10.1091/mbc.01-06-0288,
- https://doi.org/10.1128/aem.70.10.6086-6091.2004,
Deep Research Report: alo1 (pombe)
(alo1-deep-research.md)
Deep Research Report: alo1 (pombe)
(alo1-deep-research.md)Deep Research Report: alo1 (pombe)
Generated using OpenAI Deep Research API
alo1 (Schizosaccharomyces pombe) – Comprehensive Gene Analysis
Gene Function and Molecular Mechanism
The alo1 gene of Schizosaccharomyces pombe encodes a flavin-dependent oxidoreductase known as D-arabinono-1,4-lactone oxidase (thebiogrid.org). This enzyme catalyzes the oxidation of D-arabinono-1,4-lactone to dehydro-D-arabinono-1,4-lactone, using oxygen as an electron acceptor and producing hydrogen peroxide as a byproduct (ctdbase.org). In practice, alo1’s activity is required for the biosynthesis of D-erythroascorbic acid, a five-carbon analog of vitamin C (L-ascorbate) that serves as an antioxidant in fungi (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov). Consistent with this role, alo1 performs the terminal step of the D-erythroascorbic acid pathway in yeast (GO:0003885: D-arabinono-1,4-lactone oxidase activity) (pubmed.ncbi.nlm.nih.gov). The molecular mechanism involves an FAD cofactor mediating electron transfer to oxygen; notably, the enzyme carries a covalently bound FAD, indicating a tightly associated prosthetic group typical for this oxidase family (pubmed.ncbi.nlm.nih.gov). Together, these features establish alo1 as a key enzymatic defender against oxidative damage, analogous to L-gulonolactone oxidase in vitamin C-synthesizing animals (pubmed.ncbi.nlm.nih.gov) (humans lack the functional GULO enzyme, explaining our dietary requirement for vitamin C).
Cellular Localization and Subcellular Components
Evidence suggests that Alo1 is associated with mitochondrial or other organelle membranes. The S. pombe enzyme is predicted to contain a hydrophobic region that may serve as a transmembrane anchor (pubmed.ncbi.nlm.nih.gov). Similarly, the S. cerevisiae Alo1 ortholog was purified from the mitochondrial fraction of cells (pubmed.ncbi.nlm.nih.gov), hinting that the enzyme localizes to mitochondria or mitochondria-associated membranes. The presence of a putative transmembrane segment at the N-terminus could target or tether Alo1 to a membrane compartment (pubmed.ncbi.nlm.nih.gov). By analogy to the mammalian vitamin C synthesis enzyme (gulonolactone oxidase, which is anchored to the endoplasmic reticulum membrane), Alo1 may reside on the mitochondrial membrane (possibly the inner membrane) where it can access its substrate in the organelle. In summary, Alo1 is likely a membrane-associated mitochondrial enzyme (GO:0005739: mitochondrion; GO:0016020: membrane), though the precise organelle compartment in fission yeast awaits experimental confirmation.
Biological Processes and Pathways
alo1 is involved in antioxidant metabolism and stress response. Its enzymatic product, D-erythroascorbic acid, is an important cellular antioxidant in yeast (pubmed.ncbi.nlm.nih.gov). Accordingly, alo1 contributes to the response to oxidative stress (GO:0006979) by maintaining intracellular antioxidant levels. S. cerevisiae cells lacking ALO1 cannot synthesize D-erythroascorbic acid and show heightened sensitivity to oxidants (pubmed.ncbi.nlm.nih.gov). Conversely, ALO1 overexpression boosts D-erythroascorbate levels ~7-fold and confers greater resistance to oxidative damage (pubmed.ncbi.nlm.nih.gov). These findings indicate that Alo1 activity is part of the cell’s defense against reactive oxygen species, working in parallel with glutathione and other antioxidant systems. In Candida albicans (a pathogenic yeast), the Alo1 homolog is similarly required for coping with oxidative stress during filamentous growth (pubmed.ncbi.nlm.nih.gov), underscoring a conserved role in stress protection. Beyond general stress response, alo1-driven production of D-erythroascorbate may support longevity or stationary-phase survival in yeast, as higher antioxidant capacity can mitigate age-related oxidative damage (a hypothesis consistent with antioxidant-associated lifespan extension in various organisms (pmc.ncbi.nlm.nih.gov)). Additionally, Alo1 might intersect with metabolic pathways for sugar acids; for example, it acts after the pentose phosphate pathway supplies precursors for D-arabinono-1,4-lactone (pmc.ncbi.nlm.nih.gov). While not classically viewed as a central metabolic enzyme, alo1 defines a specialized biosynthetic process – the erythroascorbate biosynthetic process – that is crucial under stress conditions.
Disease Associations and Phenotypes
There are no direct human diseases caused by S. pombe alo1, since fission yeast is a unicellular model organism. However, knowledge of alo1 and its orthologs provides insights into pathogenicity and nutritional requirements in other species. In Candida albicans, deletion of the ALO1 gene (required for D-erythroascorbate production) leads to reduced hyphal growth and virulence (pubmed.ncbi.nlm.nih.gov). This suggests that fungal pathogens rely on this enzyme to withstand host-derived oxidative stress, making ALO1 a potential antifungal target. In plant pathogenic fungi like Magnaporthe oryzae, an alo1 homolog (Moalo1) is required for normal fungal growth, spore development (conidiogenesis), and pathogenicity on rice (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). These disease-context studies imply that loss of alo1 function impairs the organism’s ability to handle oxidative challenges during infection. By extension, the human dependency on dietary vitamin C is a notable evolutionary “disease” association: humans carry a nonfunctional GULO (gulonolactone oxidase) gene (a distant relative of alo1) and thus develop scurvy without sufficient vitamin C intake (pubmed.ncbi.nlm.nih.gov). While scurvy itself is not linked to fungal alo1, this human condition highlights the physiological importance of the enzymatic activity that alo1 orthologs perform. In lab strains of S. pombe, an alo1∆ (deletion) mutant is viable but expected to display phenotypes such as sensitivity to hydrogen peroxide or superoxide-generating drugs (by analogy to S. cerevisiae alo1 mutants (pubmed.ncbi.nlm.nih.gov)). Indeed, S. cerevisiae alo1 mutants are hypersensitive to oxidative stress, and overexpression alleviates such stress (pubmed.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). No gross developmental defects have been reported in alo1-null fission yeast under normal conditions, consistent with it being conditionally important (i.e. important for stress resistance rather than growth in rich media). Future studies on S. pombe alo1 mutants (for example, testing survival after peroxide exposure) would further illuminate its phenotypic impacts.
Protein Domains and Structural Features
Alo1 is a flavoprotein belonging to the oxygen-dependent FAD-linked oxidoreductase family (string-db.org). The protein sequence (~461 amino acids in S. pombe) contains two characteristic domains: an N-terminal FAD-binding domain and a C-terminal ALO domain (pmc.ncbi.nlm.nih.gov). The FAD-binding region (Pfam PF01565, sometimes called FAD_binding_4) binds the flavin adenine dinucleotide cofactor that is integral to the enzyme’s redox activity (pmc.ncbi.nlm.nih.gov). The ALO domain (Pfam PF04030) is named after arabinono-1,4-lactone oxidase and likely forms the substrate-binding and catalytic site specific for sugar lactones (pmc.ncbi.nlm.nih.gov). Notably, Alo1 is predicted to covalently attach FAD via a conserved histidyl residue – a feature it shares with its homologs in other species (pubmed.ncbi.nlm.nih.gov). This covalent FAD linkage is known from the rat L-gulonolactone oxidase structure and is inferred for yeast Alo1 based on sequence motifs (pubmed.ncbi.nlm.nih.gov). The holoprotein is thus a covalent flavoprotein, which can enhance stability of the cofactor and enzyme.
Structurally, Alo1 likely folds into a two-domain architecture common to many flavin oxidases: a Rossmann-like fold for FAD binding and a more specialized domain for substrate recognition. Comparative sequence analysis shows ~32% identity between S. cerevisiae Alo1 and rat gulonolactone oxidase, and ~21% identity to plant L-galactono-1,4-lactone dehydrogenase (pubmed.ncbi.nlm.nih.gov). These homologies suggest a conserved overall structure and mechanism among these enzymes despite differences in substrate specificity. Alo1 also has a hydrophobic segment near its N-terminus that may form a single-pass transmembrane helix (pubmed.ncbi.nlm.nih.gov). This segment is relatively short and could function either as a membrane anchor or a targeting signal for insertion into organelle membranes. Some analyses have called it a “putative transmembrane segment” (pubmed.ncbi.nlm.nih.gov), though one yeast proteome resource did not predict any long transmembrane domain in the mature protein (www.ymdb.ca). It is possible that this region acts as a signal peptide that directs Alo1 to the mitochondrion and is cleaved upon import, meaning the active enzyme in the matrix is soluble (consistent with the lack of a long membrane span in the processed form). Overall, the key structural features of Alo1 include the FAD-binding site, the active-site ALO domain, and an N-terminal extension for subcellular targeting. No full 3D structure of Alo1 from fission yeast has been reported to date, but the conserved domains and sequence similarity to known enzymes provide a strong model for its structure and function.
Expression Patterns and Regulation
Little specific data has been published on alo1 transcriptional regulation in S. pombe. The gene’s mRNA is constitutively present under normal growth conditions (as inferred from its discovery as an ORF in the genome (pubmed.ncbi.nlm.nih.gov)), suggesting a baseline requirement for the enzyme to maintain antioxidant levels. In S. cerevisiae, ALO1 is a single-copy gene expressed to produce a ~1.8 kb mRNA (pubmed.ncbi.nlm.nih.gov). Expression of ALO1 in budding yeast does not appear to be highly induced by ordinary stress; instead, regulation may occur at the level of enzyme activity or cofactor availability. However, there is evidence that under extreme oxidative or metal stress, cells may modulate pathways involving Alo1: for instance, overexpression of ALO1 (above normal levels) was deliberately shown to increase oxidative stress tolerance (pubmed.ncbi.nlm.nih.gov), implying that increased expression or gene dosage can be beneficial during stress. It is plausible that S. pombe upregulates alo1 under oxidative stress via stress-responsive transcription factors. The Pap1 and Atf1 pathways in fission yeast activate many antioxidant genes in response to H2O2; whether alo1 is a direct target remains to be confirmed. No canonical stress-responsive elements (such as Sty1-Atf1 binding sites) have been reported in the alo1 promoter, but given the enzyme’s role, a modest induction during oxidative stress is likely. In Candida albicans, ALO1 expression correlates with the yeast-to-hypha transition and is critical for survival in host-like conditions (pubmed.ncbi.nlm.nih.gov), hinting at context-dependent regulation in that organism.
From a biotechnological perspective, alo1 has drawn interest: its expression in other systems has been manipulated for experimental purposes. For example, S. cerevisiae ALO1 has been overexpressed in bacteria to produce D-erythroascorbate in vitro (pmc.ncbi.nlm.nih.gov), and introduced into plants to bolster stress resistance (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). These studies often use strong promoters to drive ALO1 expression, underscoring that higher Alo1 levels directly increase antioxidant output. In the natural context of S. pombe, the gene is likely expressed at a level sufficient for routine antioxidant defense, with potential upregulation when cells face heightened ROS. High-throughput transcriptome data (e.g., RNA-seq) under stress conditions could shed more light – future analyses may reveal if alo1 mRNA rises under oxidative or stationary-phase stress. In summary, alo1 appears to be a constitutively expressed housekeeping gene involved in redox homeostasis, with the capacity to enhance stress tolerance when expressed at higher levels.
Evolutionary Conservation
The function of alo1 is highly conserved across different kingdoms of life, reflecting the universal need for ascorbate or ascorbate-analogs in oxidative stress management. Orthologs of S. pombe Alo1 are found in many fungi, plants, and animals (pmc.ncbi.nlm.nih.gov). Sequence analysis confirms that the ALO domain and FAD-binding domain of Alo1 are present in diverse eukaryotes including budding yeast (S. cerevisiae ALO1), pathogenic yeasts (C. albicans), filamentous fungi (Neurospora crassa, Fusarium oxysporum, Magnaporthe oryzae), higher plants (e.g. Brassica oleracea), and mammals (pmc.ncbi.nlm.nih.gov). In mammals, the closest functional counterparts are enzymes like L-gulonolactone oxidase (GULO) in rats and mice, which synthesizes true vitamin C. The S. cerevisiae Alo1 shares ~32% identity with rat GULO and significant similarity with plant L-galactono-1,4-lactone dehydrogenase, an enzyme in plant vitamin C biosynthesis (pubmed.ncbi.nlm.nih.gov). This moderate sequence identity, combined with conserved domain architecture, indicates a common evolutionary origin for these FAD-dependent lactone oxidases. Interestingly, while most mammals (except primates, guinea pigs, etc.) retain GULO for vitamin C production, fungi and yeasts have evolved the 5-carbon analog pathway with Alo1 – a convergence on the same biochemical solution (producing an antioxidant) via slightly different substrates. The presence of alo1 homologs in virtually all fungi examined suggests that D-erythroascorbic acid biosynthesis is a widespread fungal strategy for oxidative stress protection (pmc.ncbi.nlm.nih.gov). Even non-pathogenic yeasts like Schizosaccharomyces and Saccharomyces use this pathway, highlighting its fundamental role in cell physiology. In plants, although the main vitamin C pathway uses a different enzyme (L-galactono-1,4-lactone dehydrogenase in mitochondria), some plants and algae also contain genes with ALO/FAD_binding domains, possibly reflecting ancestral genes or parallel functions (pmc.ncbi.nlm.nih.gov).
From an evolutionary perspective, alo1 and its orthologs form a sub-family within the broader vanillyl-alcohol oxidase flavoprotein family (string-db.org). Phylogenetic analyses (such as those in M. oryzae Alo1 studies) show that fungal Alo1 enzymes cluster together, with plant and animal enzymes branching nearby, consistent with species phylogeny (pmc.ncbi.nlm.nih.gov). This suggests that the enzyme’s function was present in a common ancestor prior to the divergence of fungi, plants, and metazoans. The human genome still contains a GULOP pseudogene, the remnants of the gulonolactone oxidase gene that our primate ancestors lost (www.ncbi.nlm.nih.gov). That loss, and our subsequent dependence on dietary ascorbate, underscores how critical – yet sometimes dispensable – this pathway can be: some organisms dropped it (e.g., primates in vitamin C-rich diets), whereas most others retained it for survival. In summary, alo1 is part of an evolutionarily conserved network of enzymes dedicated to ascorbate-like molecule production, spanning yeast to mammals, with conservation of key domains and catalytic mechanisms across ~1.5 billion years of evolution.
Key Experimental Evidence and Literature
-
Discovery and Enzymatic Function (Yeast): S. cerevisiae ALO1 was first identified as a gene encoding D-arabinono-1,4-lactone oxidase, purified from yeast mitochondria (pubmed.ncbi.nlm.nih.gov). Huh et al. (1998) demonstrated that this enzyme produces D-erythroascorbic acid and that alo1∆ mutants completely lack D-erythroascorbate, confirming Alo1’s role in its biosynthesis (pubmed.ncbi.nlm.nih.gov). They also showed that alo1∆ cells have no detectable D-arabinono-1,4-lactone oxidase activity, whereas overexpression of ALO1 increases enzyme activity >7-fold (pubmed.ncbi.nlm.nih.gov). This study established D-erythroascorbic acid as an important antioxidant in yeast, with Alo1 being indispensable for its production (pubmed.ncbi.nlm.nih.gov).
-
Stress Tolerance Phenotypes: The Huh et al. 1998 study further reported that alo1 mutants are hypersensitive to oxidative stress, and extra copies of ALO1 render cells more resistant (pubmed.ncbi.nlm.nih.gov). Subsequent work by Kim et al. and others demonstrated that providing yeast with D-erythroascorbic acid or overexpressing ALO1 can mitigate damage from oxidants like hydrogen peroxide or the superoxide-generating agent paraquat (pmc.ncbi.nlm.nih.gov). Min et al. (2015) even showed that expressing yeast ALO1 in tobacco plants enhanced their tolerance to oxidative stress and aluminum toxicity (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov), illustrating the conserved protective effect of the enzyme across kingdoms.
-
Candida albicans Virulence: Huh et al. (2001) cloned the C. albicans ALO1 and found that mutants lacking it could not produce D-erythroascorbate and had attenuated hyphal growth and virulence (pubmed.ncbi.nlm.nih.gov). This key evidence linked Alo1 activity to pathogenic fitness, likely by compromising the fungus’s ability to neutralize reactive oxygen from the host immune response.
-
Magnaporthe oryzae Pathogenicity: Wu et al. (2022) characterized M. oryzae MoAlo1, confirming it has the same ALO and FAD_binding_4 domains as yeast Alo1 (pmc.ncbi.nlm.nih.gov). Deletion of Moalo1 caused severe defects in fungal growth, spore formation, and plant infection ability (pmc.ncbi.nlm.nih.gov). They also showed MoAlo1 mutant is more sensitive to external H2O2】, reinforcing that the enzyme’s role in oxidative stress resistance is critical for disease causation (pmc.ncbi.nlm.nih.gov).
-
Biochemical Properties: The yeast Alo1 protein (526 aa in S. cerevisiae) was noted to have a covalently bound FAD and a predicted single transmembrane segment (pubmed.ncbi.nlm.nih.gov). The covalent flavin was inferred from sequence motifs and later confirmed in related enzymes, indicating a common biochemical feature. Alignment of Alo1 with rat and plant enzymes provided early evidence of its evolutionary relatedness to known vitamin C biosynthetic enzymes (pubmed.ncbi.nlm.nih.gov). These comparative studies form the basis for current GO annotations of Alo1’s molecular function (oxidoreductase activity) and biological process (ascorbate analog biosynthesis and stress response).
-
Genetic Interaction and Omics: Thus far, alo1 has not featured prominently in S. pombe large-scale mutant screens for DNA damage or cell cycle (likely because its loss yields subtle phenotypes unless oxidative stress is applied). However, Alo1 does have a small network of protein interactions (e.g., reported physical or genetic interactions with four partners (thebiogrid.org) (string-db.org), one being a 6-phosphogluconolactonase in the pentose phosphate pathway). This hints at a possible metabolic linkage: the pentose phosphate pathway (which generates NADPH and pentoses) could supply precursors or reducing power for erythroascorbate synthesis, aligning with Alo1’s role in redox balance. No direct involvement of alo1 in cell cycle or meiosis was found in a fission yeast meiotic gene screen, indicating its functions are specific to metabolic stress contexts (pmc.ncbi.nlm.nih.gov).
In conclusion, the accumulated experimental evidence paints alo1 as a metabolic stress-response gene. It encodes an enzyme that is biochemically and functionally conserved, playing a crucial role in the biosynthesis of an antioxidant molecule. Curating Alo1 in Gene Ontology terms, one would capture its oxidoreductase activity (acting on the CH-OH group of donors, oxygen as acceptor), its involvement in ascorbate analog biosynthetic process and oxidative stress response, and its association with the mitochondrial/membrane compartment (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov). The literature cited above provides strong evidence for each of these facets, ensuring that GO annotations for alo1 are well supported by experimental findings.