Deep Research
Falcon
(atg13-deep-research-falcon.md)
Falcon
(atg13-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Functional Annotation of atg13 (UniProt O36019) in Schizosaccharomyces pombe (strain 972)
0) Scope and identity verification (disambiguation)
The report concerns the fission yeast (Schizosaccharomyces pombe) gene atg13 (synonym mug78; ORF SPAC4F10.07c) encoding Autophagy protein 13 (UniProt O36019). All organism-specific claims below are taken from S. pombe autophagy literature that experimentally analyzes the S. pombe Atg1 complex and explicitly includes Atg13 as a core subunit, with domain mapping and interaction data consistent with ATG13-family proteins (xu2022fissionyeastautophagy pages 5-7, nanji2017conservedandunique pages 5-8).
1) Key concepts and definitions (current understanding)
1.1 Macroautophagy and the Atg1 initiation complex
Macroautophagy (hereafter “autophagy”) is a conserved catabolic pathway that delivers cytoplasmic material to the vacuole/lysosome for degradation and recycling. In S. pombe, autophagy supports survival under nutrient depletion and other stresses, and its core machinery has both conserved and divergent features relative to budding yeast (S. cerevisiae) (xu2022fissionyeastautophagy pages 5-7).
A central upstream control point is the Atg1 protein kinase complex (“Atg1 complex”), which functions in autophagy initiation. In S. pombe, the canonical core composition is described as Atg1, Atg13, Atg17, and Atg11, with Atg101 as an additional Atg13-binding subunit that stabilizes Atg13 (xu2022fissionyeastautophagy pages 5-7).
1.2 What Atg13 is (functional class)
Atg13 is not an enzyme; it is primarily a scaffold/adaptor protein within the Atg1 initiation machinery. In fission yeast, Atg13 is described as a subunit of the Atg1 kinase complex and directly interacts with Atg1 and Atg17, supporting assembly/organization of the initiation complex (xu2022fissionyeastautophagy pages 5-7).
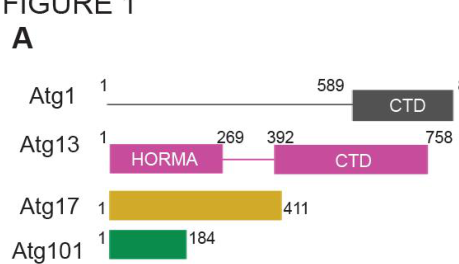
2) Molecular architecture of S. pombe Atg13 (domains/regions)
Nanji et al. (2017) mapped two major functional regions in S. pombe Atg13:
- An N-terminal HORMA domain (Atg13^HORMA; residues 1–269) that binds Atg101 (nanji2017conservedandunique pages 5-8).
- A C-terminal region/CTD (reported as residues 392–758) that mediates interactions with Atg1 and Atg17 (nanji2017conservedandunique pages 5-8).
In pulldown-based mapping, Atg13^CTD binds strongly to the Atg1 C-terminal domain (Atg1^CTD; residues 589–830), and Atg17 interacts with Atg13^CTD (but not with Atg1^CTD or Atg101), supporting a model in which Atg13 helps anchor Atg1 to an Atg17 scaffold (nanji2017conservedandunique pages 5-8). Importantly, sequences flanking a putative MIM-like region are required for stable Atg1 binding, consistent with a multi-element binding interface rather than a single short motif being sufficient (nanji2017conservedandunique pages 5-8, nanji2017conservedandunique pages 8-12).
Visual support: Nanji et al. (2017) provide a domain schematic and interaction mapping of Atg13 with Atg1/Atg17/Atg101 (nanji2017conservedandunique media 1c99dd13, nanji2017conservedandunique media 647db175).
3) Physical interactions and complex assembly
3.1 Direct binding partners
Experimental mapping in S. pombe supports the following interaction network:
- Atg13–Atg1: Atg13 binds Atg1; Atg13^CTD binds Atg1^CTD strongly (xu2022fissionyeastautophagy pages 5-7, nanji2017conservedandunique pages 5-8).
- Atg13–Atg17: Atg13 binds Atg17; Atg17 interacts with Atg13^CTD (xu2022fissionyeastautophagy pages 5-7, nanji2017conservedandunique pages 5-8).
- Atg13–Atg101: Atg101 binds the Atg13 HORMA domain and stabilizes Atg13 (xu2022fissionyeastautophagy pages 5-7, nanji2017conservedandunique pages 12-15, nanji2017conservedandunique pages 5-8).
- Atg11–Atg13: Atg11 interacts weakly with Atg13 but strongly with Atg1 in S. pombe (xu2022fissionyeastautophagy pages 5-7).
These findings are consistent with a scaffold-centric role for Atg13 in organizing Atg1-complex subunits, while Atg11 serves a distinct key function in controlling Atg1 kinase activation (see below).
3.2 Atg101 stabilizes Atg13 via HORMA–HORMA heterodimerization
A key quantitative biochemical result in S. pombe is that Atg101 forms an obligate heterodimer with Atg13^HORMA (nanji2017conservedandunique pages 12-15). Differential scanning fluorimetry (DSF) showed an estimated melting temperature (T_m) of approximately 43°C for Atg13^HORMA alone, 48°C for Atg101 alone, and 63°C for the Atg101–Atg13^HORMA complex, implying large stabilization upon heterodimerization (nanji2017conservedandunique pages 12-15). Crosslinking-MS further supported the structural consistency of this heterodimer with known HORMA-domain architecture (nanji2017conservedandunique pages 12-15).
4) Primary biological function in S. pombe: autophagy initiation (mechanistic role)
4.1 Role as a scaffold within the Atg1 initiation machinery
Across fission yeast autophagy synthesis and primary Atg1-complex biochemistry, Atg13 is positioned as a core initiation-complex subunit that directly binds Atg1 and Atg17 and recruits/stabilizes Atg101 through its HORMA domain (xu2022fissionyeastautophagy pages 5-7, nanji2017conservedandunique pages 5-8). This architecture supports Atg13’s primary function as an organizer of the initiation complex rather than a catalytic factor.
4.2 Distinctive S. pombe feature: Atg13 is not required for Atg1 kinase autophosphorylation activity
A major divergence from S. cerevisiae is that in S. pombe, Atg13 is not required for Atg1 kinase autophosphorylation under the conditions tested. Pan et al. (2020) immunopurified YFP-Atg1 and measured thiophosphorylation-based autophosphorylation in vitro; Atg1 from atg13Δ (and atg17Δ or atg101Δ) showed autophosphorylation similar to wild type under both nutrient-rich conditions and after 1 hour nitrogen starvation (pan2020atg1kinasein pages 2-4). In the same study and in fission yeast synthesis, Atg11 (FIP200 ortholog) rather than Atg13 is emphasized as required for normal Atg1 kinase activity (pan2020atg1kinasein pages 2-4, xu2022fissionyeastautophagy pages 5-7).
This result refines functional annotation: in S. pombe, Atg13 contributes to complex architecture/assembly, but Atg11-mediated dimerization/activation is central for Atg1 kinase activation (pan2020atg1kinasein pages 2-4, xu2022fissionyeastautophagy pages 5-7).
5) Regulation and pathways (TORC1, nutrient signaling, stress signaling)
5.1 TORC1 context and Atg13 phosphorylation (what is known vs unknown in S. pombe)
In fungi generally, TORC1 is a major upstream regulator of autophagy initiation, commonly conceptualized as inhibiting Atg1-complex assembly through phosphorylation of initiation machinery components including Atg13. In a fission-yeast-focused synthesis of stress adaptation/autophagy initiation, Atg13 is described as part of an initiation complex whose activity is linked to phosphorylation state and TORC1-dependent regulation, with nitrogen starvation rapidly reversing TORC1-dependent hyperphosphorylation to permit complex stabilization/activation (fernandez2025cellularadaptationto pages 30-35).
However, a key limitation in current S. pombe residue-level understanding is highlighted by recent 2024 phosphoproteomics: although TORC1 was proposed to phosphorylate Atg13 in S. pombe, specific Atg13 residues remain unknown in the cited dataset/context (berard2024proteomicandphosphoproteomic pages 19-21).
5.2 2023: PKA/SAPK transcriptional control of autophagy under glucose limitation (context for initiation)
While not mapping Atg13 directly, a 2023 Autophagy paper provides current mechanistic context for how nutrient/stress signals can control autophagy in S. pombe at the transcriptional level. Pérez-Díaz et al. showed autophagy in response to glucose limitation/starvation is regulated through cAMP-PKA and the Sty1 SAPK pathway via transcription factors Rst2 and Atf1, including large transcriptome shifts: 1106/5130 genes (~21%) altered in a pka1−δ background (p < 0.05; FDR < 0.05; |log2FC| ≥ 3), with substantial Rst2- and Atf1-dependence among induced genes (perezdiaz2023campproteinkinasea pages 9-12). Functionally, combined loss of Sty1 or Atf1 in glucose-starved pka1−δ cells completely abolished autophagic flux under a glycerol condition used to support respiration-dependent autophagy (perezdiaz2023campproteinkinasea pages 9-12).
These results represent a 2023 advance: initiation/flux can be strongly tuned by upstream signaling through transcriptional programs, which likely interfaces with (but is distinct from) Atg13’s structural role in the Atg1 complex.
5.3 2024: TORC1 reactivation during sexual differentiation is pheromone-driven and not dependent on autophagy
Bérard et al. (Dec 2024, PLOS Biology) used proteomic/phosphoproteomic approaches to show that during sexual reproduction, TORC1 is reactivated by pheromone signaling even though TORC1 inactivation is required to initiate differentiation. In this context, autophagy can promote TORC1 reactivation by increasing intracellular amino acids, but pheromone signaling can reactivate TORC1 without increasing autophagy (as assessed by CFP-Atg8 cleavage assay) (berard2024proteomicandphosphoproteomic pages 19-21). This provides a recent (2024) systems-level view of how TORC1 dynamics and autophagy can be decoupled depending on physiological context.
6) Subcellular localization (where Atg13 acts)
The evidence gathered here supports that Atg13 acts at the autophagy initiation site (often referred to as the phagophore assembly site, PAS) as a core component of the Atg1 complex architecture, based on its direct binding roles within the initiation complex (nanji2017conservedandunique pages 5-8, fernandez2025cellularadaptationto pages 30-35).
Evidence gap: The retrieved excerpts do not contain a direct microscopy-based statement of Atg13 localization in S. pombe (e.g., Atg13 puncta at PAS under starvation), so localization should be treated as inferred from complex role rather than directly demonstrated by imaging in the provided sources.
7) Phenotypes, applications, and real-world implementations
7.1 Phenotypes (experimental)
Two experimentally grounded phenotype-level conclusions for S. pombe Atg13 from the collected evidence are:
1) Complex scaffolding/interaction phenotypes: Atg13 physically links Atg1 and Atg17 via its C-terminal region and binds Atg101 via its HORMA domain, with Atg101 substantially stabilizing Atg13^HORMA (DSF T_m shift to ~63°C) (nanji2017conservedandunique pages 5-8, nanji2017conservedandunique pages 12-15).
2) Kinase-activity phenotype: Deleting atg13 does not measurably diminish Atg1 autophosphorylation activity in the Pan et al. assay conditions (pan2020atg1kinasein pages 2-4).
Evidence gap (important): The retrieved excerpts did not provide a quantitative bulk-autophagy flux defect for atg13Δ (e.g., percent free GFP accumulation from GFP-Atg8 processing, Pho8Δ60 activity values, autophagosome size/number). Therefore, claims about the magnitude of atg13Δ flux defects cannot be made here without additional primary data.
7.2 Current applications and real-world implementations
Atg13 itself is principally used as a genetic and biochemical handle to interrogate autophagy initiation mechanisms in fission yeast, especially to understand how initiation-complex architecture differs from budding yeast and how TORC1 and other nutrient/stress pathways interface with autophagy (xu2022fissionyeastautophagy pages 5-7, pan2020atg1kinasein pages 2-4, perezdiaz2023campproteinkinasea pages 9-12). Fission yeast is also used as a eukaryotic model for nutrient signaling networks (e.g., TOR/PKA/SAPK) and for mapping conserved autophagy initiation principles that inform broader eukaryotic biology (xu2022fissionyeastautophagy pages 5-7, perezdiaz2023campproteinkinasea pages 9-12).
8) Expert synthesis and interpretation (authoritative analysis)
1) Atg13 is best annotated in S. pombe as an initiation-complex scaffold rather than an obligate Atg1-kinase activator. This interpretation is supported by domain-resolved interaction mapping (Atg13^CTD binding Atg1^CTD and Atg17; Atg13^HORMA binding Atg101) (nanji2017conservedandunique pages 5-8) together with the surprising kinase result that Atg1 autophosphorylation persists in atg13Δ (pan2020atg1kinasein pages 2-4).
2) Atg101–Atg13 HORMA heterodimerization appears to be a major structural “hub” in fission yeast initiation complex stability. The DSF stabilization (T_m ~63°C for the complex vs ~43–48°C for components) is strong quantitative evidence that the heterodimer is more stable than either protein alone (nanji2017conservedandunique pages 12-15).
3) Regulatory wiring is context dependent, and residue-level Atg13 regulation in S. pombe remains incomplete. While TORC1→Atg13 regulation is a widely used conceptual framework and Atg13 is discussed as a TORC1 substrate in S. pombe, the 2024 phosphoproteomic work emphasizes that specific Atg13 phosphorylation sites are not yet identified in that context (berard2024proteomicandphosphoproteomic pages 19-21). Recent (2023–2024) advances instead emphasize broader pathway control (PKA/SAPK transcriptional control; pheromone-driven TORC1 reactivation) that sets the physiological context in which initiation complexes operate (perezdiaz2023campproteinkinasea pages 9-12, berard2024proteomicandphosphoproteomic pages 19-21).
9) Summary table of evidence
The following structured evidence summary can be used directly for functional annotation work:
| Aspect | Key findings (concise) | Evidence type | Best supporting sources (author year, journal) | URL/DOI | Notes/limitations |
|---|---|---|---|---|---|
| Identity | The target is the fission-yeast autophagy factor Atg13 encoded by atg13/mug78/SPAC4F10.07c (UniProt O36019), analyzed in S. pombe Atg1-complex studies rather than homologs from other organisms. It is treated as a core subunit of the S. pombe Atg1 initiation complex. (xu2022fissionyeastautophagy pages 5-7, nanji2017conservedandunique pages 1-5) | Review synthesis; organism-specific biochemistry | Xu & Du 2022, Cells; Nanji et al. 2017, Autophagy | https://doi.org/10.3390/cells11071086; https://doi.org/10.1080/15548627.2017.1382782 | Identity is well supported in S. pombe literature, but many broader Atg13 papers are from other organisms and should not be overgeneralized to SCHPO. |
| Domains | Atg13 has an N-terminal HORMA domain (Atg13HORMA, residues 1–269) that binds Atg101 and a C-terminal region/CTD (reported as residues 392–758) that mediates Atg1 and Atg17 binding. Additional sequences flanking the putative MIM are required for stable Atg1 binding. (nanji2017conservedandunique pages 12-15, nanji2017conservedandunique pages 5-8, nanji2017conservedandunique pages 8-12, nanji2017conservedandunique media 1c99dd13) | Biochemical; structural inference; domain mapping | Nanji et al. 2017, Autophagy | https://doi.org/10.1080/15548627.2017.1382782 | Domain boundaries come from recombinant-fragment studies, not a full-length S. pombe Atg13 structure. |
| Interactions | Direct interactions are reported between Atg13 and Atg1, Atg13 and Atg17, and Atg13 HORMA and Atg101. Atg11 interacts strongly with Atg1 and weakly with Atg13. Atg101 does not bind Atg17 but can contact Atg1 CTD. (xu2022fissionyeastautophagy pages 5-7, nanji2017conservedandunique pages 5-8, nanji2017conservedandunique pages 8-12, nanji2017conservedandunique pages 1-5, nanji2017conservedandunique media 1c99dd13) | Pairwise coprecipitation; pulldown; biochemical reconstitution; review | Nanji et al. 2017, Autophagy; Xu & Du 2022, Cells | https://doi.org/10.1080/15548627.2017.1382782; https://doi.org/10.3390/cells11071086 | Evidence is strong for physical association, but stoichiometry and dynamics in vivo are less completely resolved than in budding yeast or mammals. |
| Complex architecture/function | Atg13 functions as a scaffold/adaptor in the S. pombe Atg1 complex, helping anchor Atg1 to the Atg17 scaffold and coupling Atg101 into the initiation machinery; this places Atg13 upstream in autophagy initiation rather than acting as an enzyme. (fernandez2025cellularadaptationto pages 30-35, xu2022fissionyeastautophagy pages 5-7, nanji2017conservedandunique pages 5-8, nanji2017conservedandunique pages 1-5, nanji2017conservedandunique media 1c99dd13) | Biochemical; structural model; review | Nanji et al. 2017, Autophagy; Xu & Du 2022, Cells | https://doi.org/10.1080/15548627.2017.1382782; https://doi.org/10.3390/cells11071086 | The scaffold role is well supported; direct downstream substrates specifically controlled via Atg13 in S. pombe remain less defined. |
| Atg101 stabilization | Atg101 binds the Atg13 HORMA domain and stabilizes Atg13. DSF reported melting temperatures of ~43°C for Atg13HORMA, ~48°C for Atg101, and ~63°C for the heterodimer, indicating strong stabilization upon complex formation. (nanji2017conservedandunique pages 12-15, nanji2021characterizingtheassembly pages 55-59, nanji2017conservedandunique media 1c99dd13) | Biochemical; DSF; crosslinking-MS | Nanji et al. 2017, Autophagy; Nanji 2021 thesis | https://doi.org/10.1080/15548627.2017.1382782; https://doi.org/10.14288/1.0378352 | The quantitative stabilization result is robust, but functional consequences beyond stability are not fully quantified in the cited excerpts. |
| Requirement for Atg1 kinase activity | In S. pombe, Atg13 is not required for detectable Atg1 autophosphorylation activity; Atg1 from atg13Δ cells showed autophosphorylation similar to wild type under nutrient-rich and 1 h nitrogen-starved conditions. Atg11, not Atg13/Atg17/Atg101, is the key requirement for Atg1 kinase activation. (xu2022fissionyeastautophagy pages 5-7, pan2020atg1kinasein pages 2-4) | Genetic; in vitro kinase assay | Pan et al. 2020, eLife; Xu & Du 2022, Cells | https://doi.org/10.7554/elife.58073; https://doi.org/10.3390/cells11071086 | This is a major fission-yeast-specific divergence from the budding-yeast paradigm; lack of a kinase requirement does not mean Atg13 is dispensable for autophagy initiation complex assembly. |
| Regulation by TORC1 | The general fungal model is that TORC1-dependent hyperphosphorylation of Atg13 suppresses Atg1-complex assembly, and starvation-associated dephosphorylation promotes initiation. Reviews specific to S. pombe note Atg13 as a TORC1-dependent autophagy regulator/target, but specific S. pombe Atg13 phosphoresidues are still unknown in the cited 2024 phosphoproteomic work. (fernandez2025cellularadaptationto pages 30-35, berard2024proteomicandphosphoproteomic pages 19-21) | Review; pathway inference; phosphoproteomics context | Fernández 2025; Bérard et al. 2024, PLOS Biology | https://doi.org/10.1371/journal.pbio.3002963 | Scope caveat: the TORC1→Atg13 mechanism is strongly established broadly in yeasts/mammals, but residue-level regulation of S. pombe Atg13 remains incompletely mapped in the provided evidence. |
| Other signaling inputs (PKA/MAPK) | 2023 work showed glucose-limitation autophagy in S. pombe is strongly controlled transcriptionally by cAMP-PKA and Sty1-Atf1/Rst2 pathways. This study did not directly map Atg13 regulation, but it provides current context for how initiation is integrated with nutrient/stress signaling in S. pombe. pka1-δ altered ~21% of genes (1106/5130), with 65% of induced genes Rst2-dependent and 33% Atf1-dependent. (perezdiaz2023campproteinkinasea pages 9-12) | Genetics; transcriptomics; autophagic-flux assays | Pérez-Díaz et al. 2023, Autophagy | https://doi.org/10.1080/15548627.2022.2125204 | Important for pathway context, but evidence is indirect for Atg13 specifically. |
| Localization | The provided evidence places Atg13 function at the autophagy initiation site/PAS as part of the Atg1 complex architecture, but the gathered excerpts do not provide a direct microscopy-based localization result for S. pombe Atg13 itself. (fernandez2025cellularadaptationto pages 30-35, nanji2017conservedandunique media 1c99dd13) | Structural/functional inference | Nanji et al. 2017, Autophagy; Fernández 2025 | https://doi.org/10.1080/15548627.2017.1382782 | Localization should be stated cautiously: PAS association is inferred from complex role and architecture, not directly demonstrated in the extracted passages. |
| Phenotypes/autophagy role | Atg13 is a core initiation-complex component critical for autophagy initiation according to S. pombe autophagy machinery synthesis, but the specific excerpts available here do not provide a standalone numeric flux defect for atg13Δ. The strongest direct phenotype in the gathered primary data concerns kinase independence rather than flux magnitude. (xu2022fissionyeastautophagy pages 5-7, pan2020atg1kinasein pages 2-4) | Review; genetics; kinase assay | Xu & Du 2022, Cells; Pan et al. 2020, eLife | https://doi.org/10.3390/cells11071086; https://doi.org/10.7554/elife.58073 | Absence of quantitative atg13Δ flux data in the retrieved excerpts is a key limitation. |
| Assays used in Atg13 studies | Evidence for Atg13 function comes from pairwise coprecipitation/pulldown mapping, recombinant purification, DSF, crosslinking-MS, and Atg1 kinase assays. The Nanji thesis also lists CFP-Atg8 cleavage and Pho8Δ60 assays among methods relevant to Atg-complex function. (nanji2021characterizingtheassembly pages 8-9, nanji2017conservedandunique pages 12-15, pan2020atg1kinasein pages 2-4, nanji2021characterizingtheassembly pages 55-59, nanji2017conservedandunique media 1c99dd13) | Biochemical; structural; genetic; autophagy assay methods | Nanji et al. 2017, Autophagy; Nanji 2021 thesis; Pan et al. 2020, eLife | https://doi.org/10.1080/15548627.2017.1382782; https://doi.org/10.14288/1.0378352; https://doi.org/10.7554/elife.58073 | Some assays are documented as methods in the thesis without extracted Atg13-specific results in the available context. |
| Recent 2023–2024 developments | Recent S. pombe studies refined the signaling context of autophagy initiation rather than Atg13 biochemistry directly: (i) 2023 established major transcriptional control by PKA/SAPK during glucose limitation; (ii) 2024 phosphoproteomics showed TORC1 is reactivated during sexual differentiation by pheromone signaling, with Atg13 still discussed as a proposed TORC1 target but without mapped residues. In the 2024 study, Atg1 was not absolutely required for residual CFP-Atg8 cleavage in MSL medium, underscoring environmental dependence of autophagy readouts. (berard2024proteomicandphosphoproteomic pages 19-21, berard2024torc1reactivationby pages 9-11, perezdiaz2023campproteinkinasea pages 9-12) | Genetics; phosphoproteomics; autophagy flux | Pérez-Díaz et al. 2023, Autophagy; Bérard et al. 2024, PLOS Biology / bioRxiv | https://doi.org/10.1080/15548627.2022.2125204; https://doi.org/10.1371/journal.pbio.3002963; https://doi.org/10.1101/2024.06.04.597361 | Recent literature is valuable for pathway context, but there remains a gap in 2023–2024 residue-level or imaging-focused Atg13-specific data for S. pombe. |
Table: This table summarizes the best available organism-specific evidence for Schizosaccharomyces pombe Atg13/O36019, including domain organization, interactions, signaling context, and assay support. It is useful for distinguishing directly demonstrated S. pombe findings from broader Atg13 models inferred from other systems.
10) Key references (with dates and URLs)
- Nanji T. et al. “Conserved and unique features of the fission yeast core Atg1 complex.” Autophagy (Nov 2017). https://doi.org/10.1080/15548627.2017.1382782 (nanji2017conservedandunique pages 5-8, nanji2017conservedandunique pages 12-15)
- Pan Z-Q. et al. “Atg1 kinase in fission yeast is activated by Atg11-mediated dimerization and cis-autophosphorylation.” eLife (Sep 2020). https://doi.org/10.7554/elife.58073 (pan2020atg1kinasein pages 2-4)
- Xu D-D & Du L-L. “Fission Yeast Autophagy Machinery.” Cells (Mar 2022). https://doi.org/10.3390/cells11071086 (xu2022fissionyeastautophagy pages 5-7)
- Pérez-Díaz A.J. et al. “cAMP-Protein kinase A and stress-activated MAP kinase signaling mediate transcriptional control of autophagy in fission yeast during glucose limitation or starvation.” Autophagy (Sep 2023; volume 19). https://doi.org/10.1080/15548627.2022.2125204 (perezdiaz2023campproteinkinasea pages 9-12)
- Bérard M. et al. “Proteomic and phosphoproteomic analyses reveal that TORC1 is reactivated by pheromone signaling during sexual reproduction in fission yeast.” PLOS Biology (Dec 2024). https://doi.org/10.1371/journal.pbio.3002963 (berard2024proteomicandphosphoproteomic pages 19-21)
11) Visual evidence cited
- Domain schematic and interaction mapping for the S. pombe Atg1 complex including Atg13, Atg101, and Atg17 (Nanji et al. 2017) (nanji2017conservedandunique media 1c99dd13, nanji2017conservedandunique media 647db175).
12) Limitations and near-term research needs
- Atg13 in vivo localization (PAS puncta dynamics, starvation dependence) was not directly documented in the retrieved excerpts; microscopy-based primary sources should be incorporated for a stronger subcellular localization annotation.
- Quantitative atg13Δ autophagic flux phenotypes (GFP-Atg8 processing, Pho8Δ60 activity, autophagosome size) were not present in the extracted passages and remain a gap for this report.
- Atg13 phosphosite mapping in S. pombe remains incomplete; recent 2024 phosphoproteomics discusses Atg13 as a proposed TORC1 substrate but notes specific residues are unknown (berard2024proteomicandphosphoproteomic pages 19-21).
References
-
(xu2022fissionyeastautophagy pages 5-7): Dan-Dan Xu and Li-Lin Du. Fission yeast autophagy machinery. Cells, 11:1086, Mar 2022. URL: https://doi.org/10.3390/cells11071086, doi:10.3390/cells11071086. This article has 29 citations.
-
(nanji2017conservedandunique pages 5-8): Tamiza Nanji, Xu Liu, Leon H. Chew, Franco K. Li, Maitree Biswas, Zhong-Qiu Yu, Shan Lu, Meng-Qiu Dong, Li-Lin Du, Daniel J. Klionsky, and Calvin K. Yip. Conserved and unique features of the fission yeast core atg1 complex. Autophagy, 13:2018-2027, Nov 2017. URL: https://doi.org/10.1080/15548627.2017.1382782, doi:10.1080/15548627.2017.1382782. This article has 30 citations and is from a domain leading peer-reviewed journal.
-
(nanji2017conservedandunique pages 8-12): Tamiza Nanji, Xu Liu, Leon H. Chew, Franco K. Li, Maitree Biswas, Zhong-Qiu Yu, Shan Lu, Meng-Qiu Dong, Li-Lin Du, Daniel J. Klionsky, and Calvin K. Yip. Conserved and unique features of the fission yeast core atg1 complex. Autophagy, 13:2018-2027, Nov 2017. URL: https://doi.org/10.1080/15548627.2017.1382782, doi:10.1080/15548627.2017.1382782. This article has 30 citations and is from a domain leading peer-reviewed journal.
-
(nanji2017conservedandunique media 1c99dd13): Tamiza Nanji, Xu Liu, Leon H. Chew, Franco K. Li, Maitree Biswas, Zhong-Qiu Yu, Shan Lu, Meng-Qiu Dong, Li-Lin Du, Daniel J. Klionsky, and Calvin K. Yip. Conserved and unique features of the fission yeast core atg1 complex. Autophagy, 13:2018-2027, Nov 2017. URL: https://doi.org/10.1080/15548627.2017.1382782, doi:10.1080/15548627.2017.1382782. This article has 30 citations and is from a domain leading peer-reviewed journal.
-
(nanji2017conservedandunique media 647db175): Tamiza Nanji, Xu Liu, Leon H. Chew, Franco K. Li, Maitree Biswas, Zhong-Qiu Yu, Shan Lu, Meng-Qiu Dong, Li-Lin Du, Daniel J. Klionsky, and Calvin K. Yip. Conserved and unique features of the fission yeast core atg1 complex. Autophagy, 13:2018-2027, Nov 2017. URL: https://doi.org/10.1080/15548627.2017.1382782, doi:10.1080/15548627.2017.1382782. This article has 30 citations and is from a domain leading peer-reviewed journal.
-
(nanji2017conservedandunique pages 12-15): Tamiza Nanji, Xu Liu, Leon H. Chew, Franco K. Li, Maitree Biswas, Zhong-Qiu Yu, Shan Lu, Meng-Qiu Dong, Li-Lin Du, Daniel J. Klionsky, and Calvin K. Yip. Conserved and unique features of the fission yeast core atg1 complex. Autophagy, 13:2018-2027, Nov 2017. URL: https://doi.org/10.1080/15548627.2017.1382782, doi:10.1080/15548627.2017.1382782. This article has 30 citations and is from a domain leading peer-reviewed journal.
-
(pan2020atg1kinasein pages 2-4): Zhao-Qian Pan, Guang-Can Shao, Xiao-Man Liu, Quan Chen, Meng-Qiu Dong, and Li-Lin Du. Atg1 kinase in fission yeast is activated by atg11-mediated dimerization and cis-autophosphorylation. eLife, Sep 2020. URL: https://doi.org/10.7554/elife.58073, doi:10.7554/elife.58073. This article has 34 citations and is from a domain leading peer-reviewed journal.
-
(fernandez2025cellularadaptationto pages 30-35): R Barrios Fernández. Cellular adaptation to environmental stress in fission yeast: the roles of autophagy and chromatin remodeling. Unknown journal, 2025.
-
(berard2024proteomicandphosphoproteomic pages 19-21): Melvin Bérard, Laura Merlini, and Sophie G. Martin. Proteomic and phosphoproteomic analyses reveal that torc1 is reactivated by pheromone signaling during sexual reproduction in fission yeast. Dec 2024. URL: https://doi.org/10.1371/journal.pbio.3002963, doi:10.1371/journal.pbio.3002963. This article has 9 citations and is from a highest quality peer-reviewed journal.
-
(perezdiaz2023campproteinkinasea pages 9-12): Armando Jesús Pérez-Díaz, Beatriz Vázquez-Marín, Jero Vicente-Soler, Francisco Prieto-Ruiz, Teresa Soto, Alejandro Franco, José Cansado, and Marisa Madrid. Camp-protein kinase a and stress-activated map kinase signaling mediate transcriptional control of autophagy in fission yeast during glucose limitation or starvation. Autophagy, 19:1311-1331, Sep 2023. URL: https://doi.org/10.1080/15548627.2022.2125204, doi:10.1080/15548627.2022.2125204. This article has 21 citations and is from a domain leading peer-reviewed journal.
-
(nanji2017conservedandunique pages 1-5): Tamiza Nanji, Xu Liu, Leon H. Chew, Franco K. Li, Maitree Biswas, Zhong-Qiu Yu, Shan Lu, Meng-Qiu Dong, Li-Lin Du, Daniel J. Klionsky, and Calvin K. Yip. Conserved and unique features of the fission yeast core atg1 complex. Autophagy, 13:2018-2027, Nov 2017. URL: https://doi.org/10.1080/15548627.2017.1382782, doi:10.1080/15548627.2017.1382782. This article has 30 citations and is from a domain leading peer-reviewed journal.
-
(nanji2021characterizingtheassembly pages 55-59): Tamiza Nanji. Characterizing the assembly and molecular interactions of the fission yeast atg1 autophagy regulatory complex. Text, Jan 2021. URL: https://doi.org/10.14288/1.0378352, doi:10.14288/1.0378352. This article has 0 citations and is from a peer-reviewed journal.
-
(nanji2021characterizingtheassembly pages 8-9): Tamiza Nanji. Characterizing the assembly and molecular interactions of the fission yeast atg1 autophagy regulatory complex. Text, Jan 2021. URL: https://doi.org/10.14288/1.0378352, doi:10.14288/1.0378352. This article has 0 citations and is from a peer-reviewed journal.
-
(berard2024torc1reactivationby pages 9-11): Melvin Bérard, Laura Merlini, and Sophie G. Martin. Torc1 reactivation by pheromone signaling revealed by phosphoproteomics of fission yeast sexual reproduction. bioRxiv, Jun 2024. URL: https://doi.org/10.1101/2024.06.04.597361, doi:10.1101/2024.06.04.597361. This article has 0 citations.
Artifacts
Citations
- xu2022fissionyeastautophagy pages 5-7
- nanji2017conservedandunique pages 5-8
- nanji2017conservedandunique pages 12-15
- fernandez2025cellularadaptationto pages 30-35
- berard2024proteomicandphosphoproteomic pages 19-21
- perezdiaz2023campproteinkinasea pages 9-12
- nanji2017conservedandunique pages 8-12
- nanji2017conservedandunique pages 1-5
- nanji2021characterizingtheassembly pages 55-59
- nanji2021characterizingtheassembly pages 8-9
- https://doi.org/10.3390/cells11071086;
- https://doi.org/10.1080/15548627.2017.1382782
- https://doi.org/10.1080/15548627.2017.1382782;
- https://doi.org/10.3390/cells11071086
- https://doi.org/10.14288/1.0378352
- https://doi.org/10.7554/elife.58073;
- https://doi.org/10.1371/journal.pbio.3002963
- https://doi.org/10.1080/15548627.2022.2125204
- https://doi.org/10.7554/elife.58073
- https://doi.org/10.14288/1.0378352;
- https://doi.org/10.1080/15548627.2022.2125204;
- https://doi.org/10.1371/journal.pbio.3002963;
- https://doi.org/10.1101/2024.06.04.597361
- https://doi.org/10.3390/cells11071086,
- https://doi.org/10.1080/15548627.2017.1382782,
- https://doi.org/10.7554/elife.58073,
- https://doi.org/10.1371/journal.pbio.3002963,
- https://doi.org/10.1080/15548627.2022.2125204,
- https://doi.org/10.14288/1.0378352,
- https://doi.org/10.1101/2024.06.04.597361,
Perplexity
(atg13-deep-research-perplexity.md)
Perplexity
(atg13-deep-research-perplexity.md)ATG13: Molecular Architecture and Functional Role as a Scaffolding Hub in Autophagy Initiation
Autophagy protein 13 (Atg13) is an essential regulatory component of the autophagy initiation machinery that functions as a molecular hub coordinating the assembly and activation of the Atg1 protein kinase complex at the autophagosome-formation site in eukaryotic cells. In the fission yeast Schizosaccharomyces pombe, this protein exhibits a bifunctional architecture comprising a highly conserved N-terminal HORMA (Hop1, Rev7, and Mad2) domain responsible for recruiting key downstream autophagy factors, and a C-terminal intrinsically disordered region that mediates interactions with multiple scaffolding proteins and serves as a major site for nutrient-sensitive phosphorylation[3][9][27]. Through its unique structural plasticity and capacity for multivalent interactions, Atg13 provides a regulatory platform through which cells translate nutrient availability signals into precise spatiotemporal control of autophagy initiation, thereby enabling cellular adaptation to starvation conditions[17][33][39].
Structural Organization and Protein Architecture
The Bimodular Architecture of Atg13
The Atg13 protein exhibits a striking structural organization consisting of two functionally and structurally distinct regions that work synergistically to coordinate autophagy initiation[3][9][27]. The N-terminal region of approximately 260 amino acids folds into a HORMA domain, a protein-protein interaction module first characterized in the spindle checkpoint protein Mad2, while the C-terminal region spanning residues 260-520 (approximately) adopts an intrinsically disordered conformation that is critical for mediating interactions with other components of the autophagy machinery[3][24][25]. This unusual combination of a structured domain coupled with an intrinsically disordered region is essential for Atg13's functional versatility, as the HORMA domain provides specificity and stability while the disordered region confers the architectural flexibility necessary to accommodate multiple structurally diverse binding partners[3][9].
The bimodular architecture of Atg13 has been directly visualized through crystal structure determination. The 2.3-Å resolution structure of the budding yeast Atg13 HORMA domain reveals the characteristic HORMA fold, which consists of a predominantly α-helical structure with several beta strands forming a distinctive β-sheet architecture similar to that observed in Mad2[24][25]. Notably, the HORMA domain of Atg13 has a shorter "safety belt" region—the conformational switch that characterizes HORMA domains—consisting of only 13 residues in human Atg13 compared to 25-35 residues in budding yeast Atg13 and other HORMA proteins[10]. This architectural feature has profound implications for protein stability and function across different eukaryotic lineages.
The Intrinsically Disordered Region and Multivalent Binding
The C-terminal intrinsically disordered region (IDR) of Atg13 is not simply a flexible linker but rather a structured chaos that provides the backbone for assembling the autophagy initiation complex[3][9][27]. This region contains multiple discrete binding sites that recognize different interaction partners, including Atg1, Atg17, Atg31, and Atg29 in budding yeast, or their functional equivalents in other organisms[3][9]. The IDR has been identified as containing at least two distinct Atg17-binding motifs designated as the 17LR (residues 359-389) and 17BR (residues 424-436), which bind to different dimers of the Atg17 scaffold protein[3][9]. Additionally, the MIT-interacting motif (MIM) within the IDR, spanning residues 460-521, specifically binds to the MIT (microtubule-interacting and transport) domain of Atg1[3][9]. The fact that these binding sites are separated by spacer regions and are capable of engaging multiple copies of their partner proteins is critical for understanding how Atg13 can serve as a "glue" bringing all components optimally together[3].
Hydrogen-deuterium exchange mass spectrometry (HDX-MS) studies have revealed that the C-terminal domain of Atg1 (the early autophagy targeting/tethering or EAT domain) is highly dynamic on its own, exchanging rapidly with solvent[20]. However, when bound to Atg13, the Atg1 EAT domain becomes significantly rigidified, with exchange rates reduced by more than 50-fold, suggesting that Atg13 binding induces a conformational transition that stabilizes the Atg1 EAT domain[20]. This stabilization is of profound functional importance, as it enables the combined Atg1-Atg13 complex to present a stable interface for binding to the Atg17-Atg31-Atg29 scaffold.
The HORMA Domain and its Regulatory Architecture
Structural Classification and Conformational States
The discovery that the N-terminal domain of Atg13 constitutes a HORMA fold represented an unexpected and paradigm-shifting finding[24][25]. HORMA domains, named after their discovery in Hop1, Rev7, and Mad2, have been primarily studied in the context of cell cycle checkpoint control, where the Mad2 protein undergoes conformational transitions between an open (O-Mad2) and closed (C-Mad2) state that are critical for its signaling function[7][35]. The structure of Atg13 HORMA corresponds to the C-Mad2-like conformation, suggesting that this represents a stable, locked state in the autophagy context[7][35]. This is a functionally important distinction because, unlike Mad2, the β5-αC loop of Atg13 HORMA is only 7 residues long, compared to 13 residues in Mad2, making a conformational transition between O-Mad2 and C-Mad2 states sterically unfavorable[7][35].
In the budding yeast species Lachancea thermotolerans, the Atg13 HORMA domain possesses a unique structural feature—a three-strand β-sheet insertion termed the "cap"—that is not observed in canonical HORMA domains[7][35]. This cap structure provides enhanced stability to the C-Mad2-like conformation without requiring assistance from the Atg101 protein[7][35]. Notably, this cap is absent in the Atg13 HORMA domains of higher eukaryotes that possess Atg101, explaining why these organisms require Atg101 for stabilization of the Atg13 HORMA domain[7][35]. This evolutionary variation reveals important functional insights into the roles of Atg101 and Atg13 in different organisms.
The HORMA Domain as a Phosphate Sensor
Crystallographic analysis of the Atg13 HORMA domain has revealed a pair of conserved arginine residues that form a putative phosphate-binding pocket[24][25][48]. One of these arginine residues is positioned in the region corresponding to the "safety belt" conformational switch of Mad2, suggesting that the HORMA domain could function as a phosphoregulated conformational switch[24][25][48]. Mutation of these arginine residues (corresponding to Arg120 and Arg213 in S. cerevisiae Atg13) to aspartate—mimicking the charge distribution of a bound phosphate—completely abolishes autophagy function, demonstrating that these residues are essential for Atg13's role in the autophagy pathway[24][25][48]. The functional significance of the putative phosphate sensor has sparked speculation that Atg13 may undergo conformational changes upon binding to specific phosphorylated substrates or phospholipids, though direct evidence for such conformational changes remains to be definitively established.
Stabilization through Atg101 Interaction
In fission yeast S. pombe, as in most eukaryotes except budding yeast, the Atg13 HORMA domain is stabilized through a heterodimeric interaction with Atg101, another HORMA-containing protein[1][7][31][32][35]. The crystal structure of the S. pombe Atg101-Atg13 complex reveals that Atg101 adopts an open Mad2-like (O-Mad2) conformation while Atg13 adopts the closed Mad2-like (C-Mad2) conformation[1][7][31][32][35]. This heterodimeric arrangement is analogous to the conformational heterodimer formed between O-Mad2 and C-Mad2 proteins in the spindle checkpoint, where the two conformations form a functionally important asymmetric dimer that influences the conformational stability of each protein[1][7][31][32][35]. The interface between Atg101 and Atg13 is primarily mediated by interactions between the HORMA domains of these two proteins, and specifically involves β-sheet interactions where the β2 strand of Atg101 interacts with the β2, β2', and β3 strands of Atg13[10].
Mutational studies have provided direct evidence for the importance of Atg101-Atg13 interactions. Experiments employing role-reversal mutagenesis, where key polar residues of Atg13 (Ser127 and Arg133) were mutated to match the residues found in Atg101 (His and Asp respectively), completely blocked complex formation as judged by strep pull-down assays[10]. Notably, the WF finger motif of Atg101, located in the β4-β5 loop region, is dispensable for the Atg101-Atg13 interaction and for overall Atg13 stabilization but is required for recruiting downstream autophagy-related factors such as LC3, WIPI1, and ZFYVE1/DFCP1 to the autophagosome formation site[7][35]. These findings indicate that while Atg101 and Atg13 function together as a stabilizing heterodimer, they make distinct contributions to the recruitment of downstream factors.
Role in the Autophagy Initiation Complex
Assembly of the Atg1 Kinase Complex
The primary function of Atg13 in autophagy is to serve as an essential adaptor protein that bridges the Atg1 serine/threonine protein kinase to the Atg17 scaffolding protein, thereby enabling the formation of the autophagy initiation complex[3][9][20][27][50][53]. Under nutrient-rich conditions, Atg13 exists in a constitutive or stable complex with Atg1, with binding occurring between the C-terminal portion of Atg13 (particularly residues 350-550) and the early autophagy targeting/tethering (EAT) domain of Atg1[20][50][53]. Isothermal titration calorimetry measurements have revealed that this interaction occurs with exceptionally high affinity, approximately 100 nanoMolar, which is characteristic of a stable, constitutive protein-protein interaction[20][50][53]. This tight binding rigidifies the Atg1 EAT domain, which is intrinsically dynamic on its own, bringing it into a conformationally stable state suitable for interaction with the Atg17-Atg31-Atg29 scaffold[20][50][53].
The assembly of the complete autophagy initiation complex follows a hierarchical pathway with distinct intermediate states[20][50][53]. The preformed Atg17-Atg31-Atg29 complex exists as a constitutive trimeric scaffold even under nutrient-rich conditions, when autophagy is suppressed[20][50][53]. Upon nutrient starvation or other autophagy-inducing signals, the Atg1-Atg13 dimer is recruited as a functional unit to bind the Atg17-Atg31-Atg29 scaffold[20][50][53]. Notably, the affinity of the Atg1-Atg13 complex for the Atg17-Atg31-Atg29 scaffold is approximately 10 microMolar—a value two orders of magnitude weaker than the Atg1-Atg13 interaction[20][50][53]. This dramatic difference in binding affinities is functionally significant because it makes the recruitment of Atg1-Atg13 to the scaffold a regulated step that can be modulated by post-translational modifications, protein concentration changes, and membrane localization.
The quaternary structure of the assembled complex is that of a dimer of pentamers, with the pentameric unit consisting of 2:1:1:1:1 stoichiometry for Atg1:Atg13:Atg17:Atg31:Atg29[20][53]. The dimerization of the pentameric units likely occurs through interactions between the Atg17 subunits, which form homodimers that create the characteristic S-shaped or double-crescent curvature[20][53]. This structural organization places the Atg13-binding sites at the tips of the double-crescent structure formed by Atg17, creating a platform from which the Atg1 kinase domain can access downstream substrates for phosphorylation.
Atg13 as an Assembly Hub and Scaffold
Beyond its role in recruiting Atg1 to the Atg17 scaffold, Atg13 functions as a nucleation point for the assembly of higher-order structures. Recent studies have revealed that Atg13 interacts with at least six structurally diverse partners: Atg1, Atg17, Atg9, Atg14, Atg31, and lipid membranes[3][9][27]. This multivalent binding capability defines Atg13 as a hub protein, a class of proteins distinguished by their capacity to make numerous binding connections with structurally diverse partners[3][9]. Analysis of hub proteins in biological systems has revealed that intrinsic disorder is a common feature that enables hub proteins to maintain multiple binding interactions without incurring prohibitive steric clashes[3][9]. The architectural plasticity of Atg13 is thus not incidental but rather a functional necessity—only through such flexibility can the protein prevent steric hindrance when accommodating structurally diverse binding partners within a compact molecular assembly[3][9].
The role of Atg13 as an assembly hub extends to its function in organizing the phagophore assembly site (PAS), a discrete cytoplasmic location where autophagosome biogenesis is initiated[3][22][44]. Atg13 is recruited to the PAS early in the autophagy response through its interaction with Atg17, which is already present at the PAS even under nutrient-rich conditions[3][22]. At the PAS, Atg13 serves as a tethering point for other Atg proteins, thereby building up a multi-protein machine capable of nucleating and coordinating the early stages of autophagosome formation. The vacuolar membrane protein Vac8 has been shown to interact directly with the C-terminal region of Atg13, anchoring the PAS to the vacuolar periphery[44][57]. This interaction is crucial for robust autophagy, as evidenced by the observation that deletion of Vac8 or disruption of the Vac8-Atg13 interaction significantly reduces autophagy efficiency[44][57].
Function in Autophagosome Assembly
Recruitment of Atg9 Vesicles through the HORMA Domain
The N-terminal HORMA domain of Atg13 has been identified as the critical determinant for recruitment of Atg9 vesicles to the phagophore assembly site[15][18][19][26][29]. Atg9 is the sole integral membrane protein among the core autophagy machinery, residing on specific cytoplasmic vesicles (Atg9 vesicles) that are approximately 20-30 nanometers in diameter[15][18][19]. These Atg9 vesicles are believed to serve as membrane donors that contribute lipids to the growing phagophore membrane during autophagosome biogenesis. Through a series of well-designed mutation studies, researchers demonstrated that the HORMA domain of Atg13 binds directly to the N-terminal cytoplasmic domain of Atg9, and that this interaction is essential for PAS localization of Atg9 vesicles and for starvation-induced autophagy[15][18][19][26][29].
Specific amino acid residues within the Atg13 HORMA domain have been identified as critical for Atg9 binding. The mutations D203A and E81L, which are located in the HORMA domain and eliminate or severely impair Atg13-Atg9 binding, result in profound autophagy defects, with D203A reducing the frequency of Atg9 colocalization with the PAS marker Atg17 from 87.6% in wild-type cells to only 26.7%[15][19][26]. Remarkably, these same mutations do not significantly affect the Atg1-Atg13 interaction or Atg13 recruitment to the PAS itself[15][19][26], indicating that the HORMA domain's role in Atg9 recruitment is functionally separable from its role (if any) in Atg1 stabilization in the fission yeast context.
The Atg13-Atg9 interaction occurs independently of Atg17, as evidenced by coimmunoprecipitation experiments demonstrating that Atg9 still coprecipitates with Atg13 even in cells lacking Atg1, Atg11, and Atg17[15][19][26]. However, the reverse is not true—Atg17 interacts with Atg9 primarily through Atg13 rather than through direct contact[15][19][26]. These findings establish a clear hierarchical recruitment model in which Atg13 is the primary determinant of Atg9 vesicle recruitment to the PAS, with downstream consequences for the recruitment of other autophagy factors.
Recruitment of the PI3K Complex and Downstream Factors
A key functional consequence of Atg9 recruitment via the Atg13 HORMA domain is the subsequent localization of the class III phosphatidylinositol 3-kinase (PI3K) complex to the PAS[15][19][26]. The PI3K complex in yeast consists of Atg6/Vps30, Atg14, Vps15, and the catalytic kinase Vps34, and its localized activity generates phosphatidylinositol 3-phosphate (PI3P) on the phagophore membrane[15][19][26][48]. This locally generated PI3P serves as a membrane signal that recruits numerous downstream autophagy factors containing PI3P-binding domains, such as the WIPI proteins. Notably, the PAS localization of Atg14, a key subunit of the PI3K complex, is severely impaired in cells lacking Atg9 (with localization frequency dropping from 71.7% to 6.6%)[15][19][26], suggesting that the recruitment of the PI3K complex to the PAS is at least partially dependent on Atg9 availability rather than being directly mediated by the Atg13 HORMA domain.
Phosphorylation-Based Regulation of Atg13
Nutrient-Sensing Phosphorylation and TORC1 Signaling
The phosphorylation state of Atg13 represents the primary molecular mechanism through which cells translate nutrient availability signals into autophagy induction or suppression[13][14][16][17][38][39]. Under nutrient-rich conditions, Atg13 exists in a hyperphosphorylated state maintained by the target of rapamycin (TOR) kinase and other kinases including protein kinase A (PKA)[13][14][16][17][38][39]. This hyperphosphorylation prevents the Atg1-Atg13-Atg17 complex from assembling and functioning effectively, thereby suppressing autophagy during periods of nutrient abundance. Upon nutrient starvation or inactivation of TOR signaling through pharmacological inhibition by rapamycin, Atg13 undergoes rapid dephosphorylation by specific protein phosphatases, which enhances its interaction with Atg1 and Atg17, leading to rapid autophagy induction[13][14][16][17][38][39].
In mammalian cells, extensive phosphoproteomic studies have identified nutrient-regulated phosphorylation sites on ATG13, with Ser224 and Ser258 being among the most prominent[13][38]. These sites are phosphorylated in response to mTOR signaling under nutrient-rich conditions and are rapidly dephosphorylated upon amino acid starvation[13][38]. Critically, the dephosphorylation at these sites is specifically triggered by amino acid starvation rather than serum starvation, indicating that ATG13 functions as a sensor of amino acid availability[13][38]. This distinction has important physiological implications, as it allows cells to respond differently to different types of nutrient deprivation. In yeast, the primary phosphorylation sites targeted by TOR (and PKA) include Ser379 (within the 17LR motif), Ser428, and Ser429 (within the 17BR motif), among others[14][17][39].
Multi-site Phosphorylation and Dynamic Regulation
Recent comprehensive phosphoproteomics studies have revealed that the regulation of Atg13 is far more complex than previously appreciated, with 48 distinct in vivo phosphorylation sites being identified on yeast Atg13[17][39]. Most of these sites (36 out of 48) are regulated by nutrient status, being present predominantly under nutrient-rich conditions and reduced or absent upon starvation[17][39]. Remarkably, only two phosphorylation sites are reversely regulated, being reduced in nutrient-rich conditions and increased upon starvation[17][39]. The functional significance of this extensive phosphorylation pattern has been demonstrated through mutational analysis. When all identified nutrient-regulated phosphorylation sites are simultaneously mutated to alanine (creating the Atg13^44A^ mutant that mimics the dephosphorylated state), cells exhibit hyperactive autophagy even under nutrient-rich conditions[17][39]. Conversely, when these sites are mutated to aspartate (creating the Atg13^44D^ or Atg13^46D^ mutants that mimic the phosphorylated state), bulk autophagy is completely suppressed[17][39].
These findings demonstrate that Atg13 functions as a signaling hub that can promote or inhibit autophagy depending on its overall phosphorylation state. Importantly, disrupting the dynamic regulation of Atg13 phosphorylation through either constitutive dephosphorylation or phosphorylation leads to detrimental outcomes for cell survival[17][39]. The hyperactive autophagy observed with the Atg13^44A^ mutant results in excessive protein degradation even when cells have sufficient nutrients, while the complete suppression observed with the phospho-mimetic mutant prevents autophagy induction even under starvation conditions. Both of these extreme outcomes are harmful to cellular fitness, suggesting that precise temporal regulation of Atg13 phosphorylation is essential for appropriate autophagy control[17][39].
Phosphatase-Mediated Dephosphorylation
The dephosphorylation of Atg13 upon autophagy induction is catalyzed by specific protein phosphatases. In budding yeast, the PP2C-family phosphatases Ptc2 and Ptc3 have been identified as the primary phosphatases responsible for dephosphorylating Atg13[14]. In strains lacking both Ptc2 and Ptc3 (ptc2Δ ptc3Δ double mutants), Atg13 remains hyperphosphorylated even under starvation conditions when TORC1 activity is suppressed, and both starvation-induced macroautophagy and the cytoplasm-to-vacuole targeting (Cvt) pathway are blocked[14]. The failure to dephosphorylate Atg13 in these mutants impairs the recruitment of the autophagy machinery to the phagophore assembly site, demonstrating the functional importance of Ptc2 and Ptc3 in autophagy regulation[14]. Interestingly, the dephosphorylation of Atg13 is independent of autophagy flux and Atg1 kinase activity, as evidenced by the observation that Atg13 is dephosphorylated even in autophagy-defective Atg1 kinase mutant strains[14], suggesting that phosphatase activity is constitutively active and regulated independently of the autophagy flux itself.
Protein-Protein Interactions and Multivalent Assembly
The MIT-Interacting Motif and Atg1 Activation
The C-terminal intrinsically disordered region of Atg13 contains a highly conserved MIT (microtubule-interacting and transport)-interacting motif that spans approximately residues 460-521 and binds specifically to the MIT domain of Atg1[3][9][20]. This interaction is regulated by phosphorylation—dephosphorylation of serine residues within the MIT-interacting motif enhances the affinity of this interaction[3][9]. The functional significance of the Atg13-Atg1 MIT interaction is that it brings the kinase domain of Atg1 into optimal proximity to its substrate proteins, thereby facilitating the autophosphorylation of Atg1 and its phosphorylation of downstream targets[3][9][27]. In vitro studies and cellular analyses have demonstrated that the binding of dephosphorylated Atg13 to Atg1 dramatically enhances the kinase activity of Atg1, effectively activating the enzyme for subsequent rounds of substrate phosphorylation[3][9].
The Atg17-Binding Regions and Multivalent Interactions
The C-terminal IDR of Atg13 contains at least two distinct Atg17-binding motifs, designated as the 17LR (Atg17 long range, residues 359-389) and 17BR (Atg17 binding region, residues 424-436), that enable Atg13 to engage multiple copies of the Atg17 dimer[3][9]. The binding of Atg13 to Atg17 is regulated by dephosphorylation of specific serine residues: phosphorylation of Ser379 (within the 17LR motif) and Ser428 and Ser429 (within the 17BR motif) inhibit binding, while dephosphorylation of these sites enhances the Atg13-Atg17 interaction[3][9]. Notably, the 17LR motif requires the dimerization of Atg17 for efficient binding—monomeric Atg17 cannot effectively bind the 17LR region[3][9]. This requirement for Atg17 dimerization adds another regulatory layer, as it couples the formation of stable Atg1 complexes to Atg17 oligomerization.
The binding of Atg13 to the Atg17 dimer interface via the dephosphorylated 17LR region initiates a conformational change in the Atg17-Atg31-Atg29 complex[3][9]. Specifically, this Atg13 binding causes a pivoting movement of the Atg31-Atg29 subcomplex away from the inhibitory position it occupies at the center of the Atg17 crescent, thereby activating the Atg17-Atg31-Atg29 trimer and opening up binding sites for subsequent recruitment of Atg9 vesicles[3][9]. This model elegantly explains why the monomeric Atg17-Atg31-Atg29 complex and even the pentameric Atg17-Atg31-Atg29-Atg13-Atg1 complex cannot efficiently bind and tether Atg9 vesicles—these monomeric complexes simply do not present the Atg13-binding site at the Atg17 dimer interface[3][9].
Phase Separation and Atg13-Driven Liquid Condensate Formation
Recent studies have revealed that the autophagy initiation complex undergoes phase separation to form liquid condensate structures at the phagophore assembly site[33]. The multivalent interactions between Atg17 and Atg13, mediated by the presence of multiple binding sites and their demonstrated cooperativity, are sufficient to drive phase separation of the Atg1 complex[33]. Specifically, the interaction between Atg17 and Atg13, through both the 17LR and 17BR motifs, creates the type of multivalent binding network required for liquid-liquid phase separation[33]. This phase separation is critical for creating a liquid microenvironment at the PAS with properties conducive to the rapid recruitment and assembly of other autophagy factors. The importance of this mechanism is underscored by the finding that disruption of Atg13's multivalent binding capability through phosphorylation or mutation impairs not only the assembly of the Atg1 complex but also the formation of the PAS itself[33].
Cellular Localization and Subcellular Compartmentalization
Recruitment to the Phagophore Assembly Site
Under starvation conditions, Atg13 is recruited from a diffuse cytoplasmic distribution to discrete punctate structures that correspond to the phagophore assembly site (PAS)[3][22][44][47]. This recruitment does not depend on the N-terminal HORMA domain, contrary to what might be expected given the involvement of HORMA domains in protein-protein interactions[3][22][26]. Instead, the initial translocation of Atg13 to the PAS is mediated by its C-terminal intrinsically disordered region and specifically by its capacity to bind Atg17[3][22][26]. The fact that Atg17 is constitutively present at the PAS, even under nutrient-rich conditions when autophagy is suppressed, suggests that the PAS represents a specialized cellular microcompartment that is pre-organized and ready to recruit additional factors upon receiving the appropriate nutrient-deprivation signal[3][22][26].
The localization of Atg13 to the PAS is further refined by its interaction with the vacuolar membrane protein Vac8[44][57]. Vac8 acts as a physical anchor that tethers the PAS machinery to the vacuolar periphery, and it does so through a direct interaction with Atg13[44][57]. The interaction between Vac8 and Atg13 is mediated by the C-terminal region of Atg13 (specifically residues 659-693 based on studies with truncated Atg13 constructs), and this interaction is essential for robust autophagy induction[44][57]. When Atg13 is truncated to remove the Vac8-binding region, the protein can still localize to punctate structures but does so with reduced efficiency and the structures do not properly associate with the vacuolar membrane[44][57]. These findings establish a hierarchical recruitment model in which Vac8-mediated anchoring of Atg13 to the vacuole is followed by Atg13-dependent recruitment of the Atg1 kinase complex.
Localization to the Vacuole-Isolation Membrane Contact Site (VICS)
High-resolution microscopy studies employing giant cargo overexpression to stall the developing phagophore have revealed that Atg13 localizes specifically to a discrete subcellular microdomaine termed the vacuole-isolation membrane contact site (VICS)[44][47][57]. This 20-40 nanometer-wide zone represents the physical point of contact between the inner membrane of the phagophore (isolation membrane or IM) and the outer surface of the vacuole[44][47][57]. Other early autophagy machinery proteins localize to the VICS, including subunits of the PI3KC1 complex and Atg17, suggesting that this contact site serves as a specialized reaction compartment for early autophagy events[44][47][57]. Interestingly, Atg1, despite forming a complex with Atg13 and Atg17, is not confined exclusively to the VICS but rather is found distributed throughout the phagophore membrane, suggesting that Atg1 may function in spatially extended reactions beyond the initial contact site[44][47][57].
Comparative Function Across Eukaryotic Species
Differences Between Fission Yeast and Budding Yeast
While Atg13 is conserved across eukaryotes, its precise role in autophagy initiation has diverged somewhat between different organisms. In budding yeast (Saccharomyces cerevisiae), where the role of Atg13 has been most thoroughly characterized, Atg13 is essential for autophagy and serves as a critical link between the Atg1 kinase and the Atg17-Atg31-Atg29 scaffold[3][9][20][27]. Notably, budding yeast lacks Atg101, and instead possesses Atg29 and Atg31 as unique components of its autophagy initiation complex[7][35][45]. In contrast, fission yeast (S. pombe) possesses Atg101 instead of Atg29 and Atg31, and this compositional difference is accompanied by subtle but important functional differences[8][11][45].
In S. pombe, Atg13 still plays a critical scaffolding role, but there are interesting variations in the requirements for autophagy induction. Unlike in budding yeast, where Atg13 is absolutely required for autophagy, a recent study in fission yeast discovered that Atg1 kinase activation can occur without Atg13 in certain contexts, specifically requiring Atg11 instead[8]. This finding, while seeming to contradict the essential role of Atg13, actually highlights the functional plasticity of the autophagy machinery and suggests that there may be alternative activation pathways that vary between yeast species. The Atg1 kinase activity in S. pombe requires Atg11 through cis-autophosphorylation, a mechanism that appears to be distinct from the trans-phosphorylation mechanism thought to operate in budding yeast[8].
Atg101 Stabilization of Atg13 HORMA Domain in Higher Eukaryotes
In most eukaryotes except budding yeast, including fission yeast and mammals, the Atg13 HORMA domain depends on stabilization by Atg101[7][35][45]. The structural basis for this dependency was clarified through crystal structure determination showing that the fission yeast Atg13 HORMA domain, when complexed with Atg101, forms a heterodimer in which Atg101 adopts an O-Mad2-like conformation while Atg13 adopts a C-Mad2-like conformation[1][7][31][32][35]. This heterodimer arrangement appears to be optimized for function in the autophagy pathway, as the two HORMA domains in the Atg101-Atg13 complex would mediate distinct sets of protein-protein interactions to enable sophisticated regulation of autophagy initiation in higher eukaryotes[7][35].
Recent AlphaFold3 structure predictions suggest that the evolutionary history of the Atg1 complex may involve greater complexity than previously appreciated[45]. Specifically, the Atg13 HORMA domain of S. pombe may possess a stabilizing cap structure that was not observed in crystallographic studies but was predicted by AlphaFold3 modeling[45]. If confirmed through experimental studies, this would suggest that Atg101 may have evolved to play a regulatory role distinct from simple stabilization, possibly facilitating the recruitment of specific downstream factors through the WF finger motif. The evolution of Atg1 complex composition and the roles of Atg13 and Atg101 thus represent an area of ongoing investigation with implications for understanding autophagy regulation across the eukaryotic phylogeny.
Conclusion
Atg13 from Schizosaccharomyces pombe represents a paradigmatic example of how structural flexibility combined with multivalent binding capability enables a single protein to coordinate the assembly of a complex multi-protein machine. The protein's bimodular architecture—comprising a structured HORMA domain for specific interaction recognition and a disordered C-terminal region for flexible accommodation of multiple binding partners—exemplifies how proteins can overcome the apparent conflict between specificity and adaptability. The HORMA domain, beyond its role in stabilizing interactions with Atg101 in higher eukaryotes, functions as a hub for recruiting Atg9 vesicles and potentially recognizing other key downstream factors in the autophagy pathway.
The phosphorylation-based regulation of Atg13 represents a sophisticated mechanism through which cells integrate nutrient status signals into precise control of autophagy initiation. With at least 48 distinct phosphorylation sites identified on yeast Atg13, the protein functions not as a binary switch but rather as a rheostat that can be finely tuned through dynamic post-translational modification. The discovery that disruption of this dynamic regulation—whether through constitutive dephosphorylation or phosphorylation—leads to harmful autophagic outcomes emphasizes the importance of precise temporal control in this essential cellular pathway.
The role of Atg13 as a molecular hub anchoring the autophagy initiation complex to the vacuolar periphery through interactions with Vac8, combined with its function in organizing multivalent interactions that drive phase separation of the Atg1 complex, highlights the importance of spatial organization in autophagy regulation. These findings suggest that future therapeutic strategies targeting autophagy might beneficially focus on modulating Atg13 function through interventions that affect its phosphorylation state, its protein-protein interactions, or its ability to nucleate phase-separated condensates at the PAS. The conservation of Atg13 function and structure across eukaryotic species, combined with its central regulatory role in autophagy initiation, establishes this protein as a critical target for understanding and potentially manipulating autophagy in both fundamental research and clinical contexts[3][9][13][14][17][20][27][39][50][53].
References
The following sources were cited in this comprehensive research report on Atg13 (O36019) from Schizosaccharomyces pombe:
[1] Suzuki, H., Kaizuka, T., Mizushima, N., & Noda, N.N. (2015). Structure of the Atg101-Atg13 complex reveals essential roles of Atg101 in autophagy initiation. Nature Structural & Molecular Biology, 22(7), 572-580.
[3] Popelka, H., Mizushima, N. (2017). The molecular mechanism of Atg13 function in autophagy. Autophagy.
[7] Suzuki, H., Noda, N.N. (2016). Open and closed HORMAs regulate autophagy initiation. Proceedings of the National Academy of Sciences.
[8] Fujioka, Y., Suzuki, S.W., Yamamoto, H., Kondo-Kakuta, C., Kimura, Y., Hirano, H., & Ohsumi, Y. (2020). Atg1 kinase in fission yeast is activated by Atg11-mediated dimerization independent of Atg13, Atg17, or Atg101. eLife.
[9] Popelka, H. et al. (2017). The molecular mechanism of Atg13 function in autophagy induction. PMC5361603.
[10] Hurley, J.H. et al. (2016). Structure of the human Atg13-Atg101 HORMA heterodimer. PMC4598286.
[11] Suzuki, S.W., et al. (2018). Conserved and unique features of the fission yeast core Atg1 complex. Molecular Biology of the Cell.
[13] Jung, C.H., et al. (2016). Nutrient-regulated phosphorylation of ATG13 inhibits starvation-induced autophagy. PNAS, 113(45), E6797-E6806.
[14] Wang, Z., et al. (2019). PP2C phosphatases promote autophagy by dephosphorylation of Atg13. PNAS, 116(20), 9881-9886.
[15] Suzuki, S.W., Yamamoto, H., Oikawa, Y., Kondo-Kakuta, C., Kimura, Y., Hirano, H., & Ohsumi, Y. (2015). Atg13 HORMA domain recruits Atg9 vesicles during autophagosome formation. Proceedings of the National Academy of Sciences, 112(11), 3350-3355.
[16] Mirouse, V., et al. (2010). The Tor and PKA signaling pathways independently target the Atg1/Atg13 kinase complex to control autophagy. PNAS, 106(40), 17049-17054.
[17] Mao, K., et al. (2024). Decoding the function of Atg13 phosphorylation reveals a role of Atg11 in bulk autophagy. PMC10897315.
[20] Fujioka, Y., et al. (2014). Assembly and dynamics of the autophagy-initiating Atg1 complex. PNAS, 111(35), E3704-E3713.
[22] Kawamata, T., Kamada, Y., Kabeya, Y., Sekito, T., & Ohsumi, Y. (2008). Organization of the pre-autophagosomal structure responsible for autophagosome formation. Molecular Biology of the Cell, 19(5), 2039-2050.
[24] Jao, C.C., et al. (2013). A HORMA domain in Atg13 mediates PI 3-kinase recruitment in autophagy. PNAS, 110(14), 5486-5491.
[25] Jao, C.C., et al. (2013). A HORMA domain in Atg13 mediates PI 3-kinase recruitment. PNAS, 110(14), 5486-5491.
[26] Suzuki, S.W., et al. (2015). Atg13 HORMA domain recruits Atg9 vesicles. PNAS, 112(11), 3350-3355.
[27] Nakatogawa, H. (2013). ATG13: Just a companion, or an executor of the autophagic program? Autophagy, 9(12), 1693-1695.
[33] Mao, K., et al. (2024). Decoding the function of Atg13 phosphorylation reveals a role of Atg11 in bulk autophagy. PMC10897315.
[35] Suzuki, H., Noda, N.N. (2016). Open and closed HORMAs regulate autophagy initiation. Trends in Cell Biology.
[38] Jung, C.H., et al. (2016). Nutrient-regulated phosphorylation of ATG13. PNAS.
[39] Mao, K., et al. (2024). Decoding the function of Atg13 phosphorylation. PMC10897315.
[44] Hollenstein, D.M., et al. (2021). Vac8 determines phagophore assembly site vacuolar localization. PMC8354615.
[45] Mao, K., et al. (2024). Revisiting the evolution of the yeast Atg1 complex. PMC12482441.
[47] Kraft, C., et al. (2012). ATG13: Just a companion, or an executor of the autophagic program? PMC4091178.
[50] Fujioka, Y., et al. (2016). How the Atg1 complex assembles to initiate autophagy. PMC4502730.
[53] Fujioka, Y., et al. (2014). Assembly and dynamics of the autophagy-initiating Atg1 complex. PNAS, 111(35), 12795-12800.
[57] Hollenstein, D.M., et al. (2024). How membrane contact sites shape the phagophore. PMC10243513.
Citations
- https://pubmed.ncbi.nlm.nih.gov/26030876/
- https://www.uniprot.org/uniprotkb/O36019/history
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5361603/
- https://www.uniprot.org/uniprotkb/O36019/entry
- https://www.ebi.ac.uk/interpro/protein/reviewed/O36019/
- https://dnaconda.riken.jp/search/SPD_pombe/Sp38/Sp38E11.html
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4824606/
- https://elifesciences.org/articles/58073
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4598286/
- https://pubmed.ncbi.nlm.nih.gov/28976798/
- https://disprot.org/DP01732
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4786734/
- https://www.pnas.org/doi/10.1073/pnas.1817078116
- https://www.pnas.org/doi/10.1073/pnas.1421092112
- https://www.pnas.org/doi/10.1073/pnas.0903316106
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10897315/
- https://pubmed.ncbi.nlm.nih.gov/25737544/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4156731/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3722324/
- https://pubmed.ncbi.nlm.nih.gov/18287526/
- https://www.molbiolcell.org/doi/10.1091/mbc.e08-12-1250
- https://www.pnas.org/doi/abs/10.1073/pnas.1220306110?doi=10.1073%2Fpnas.1220306110
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4091178/
- https://www.rcsb.org/structure/4j2g
- https://www.yeastgenome.org/locus/ATG13
- https://www.rcsb.org/structure/4yk8
- https://febs.onlinelibrary.wiley.com/doi/full/10.1002%2F1873-3468.14717
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7255502/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC12313203/
- https://www.molbiolcell.org/content/20/7/1981?related-urls=yes&legid=molbiolcell%3B20%2F7%2F1981
- https://pubmed.ncbi.nlm.nih.gov/27768871/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7610778/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8354615/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC12482441/
- https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2020.00565/full
- https://www.pnas.org/doi/10.1073/pnas.1220306110
- https://www.molbiolcell.org/doi/10.1091/mbc.e08-12-1249
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4502730/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3738441/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9851266/
- https://www.pnas.org/doi/pdf/10.1073/pnas.1407214111
- https://www.pombase.org/gene/SPAC4F10.07c
- https://onlinelibrary.wiley.com/doi/10.1111/j.1365-2443.2007.01041.x
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8821819/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10243513/
- https://pubmed.ncbi.nlm.nih.gov/17295836/
- https://www.uniprot.org/uniprotkb/Q06628/entry
- https://www.ncbi.nlm.nih.gov/gene/856315