Deep Research
Falcon
(atg2-deep-research-falcon.md)
Falcon
(atg2-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: Functional annotation of Schizosaccharomyces pombe Atg2 (mug36; UniProt O94649)
0) Target verification (mandatory)
The research target is Atg2 from fission yeast Schizosaccharomyces pombe strain 972, also known as mug36, reported in the fission-yeast autophagy literature as atg2Δ and associated with the fission-yeast autophagy factor set (e.g., in systematic genetic/localization work). (sun2013globalanalysisof pages 3-5, sun2013globalanalysisof pages 7-10, xu2022fissionyeastautophagy pages 5-7)
Important disambiguation: “Atg2/ATG2” is a conserved autophagy gene family across eukaryotes; mechanistic work in budding yeast and mammals is used below only as conserved background and is labeled as such, while S. pombe-specific claims are supported by S. pombe studies. (kotani2018theatg2atg18complex pages 1-2, gomezsanchez2018atg9establishesatg2dependent pages 21-21, duarte2023theorganizationand pages 5-7)
1) Key concepts and definitions (current understanding)
1.1 Macroautophagy, phagophore/PAS, and where Atg2 fits
Macroautophagy (“autophagy”) proceeds through assembly of a phagophore (isolation membrane) that expands and closes into a double-membrane autophagosome. In yeast, many autophagy factors concentrate at the PAS (phagophore assembly site). In fission yeast, Atg2 is treated as part of the core autophagosome biogenesis machinery and is recruited to the PAS in an Atg18-family-dependent manner (review synthesis). (xu2022fissionyeastautophagy pages 4-5, xu2022fissionyeastautophagy pages 5-7)
1.2 ATG2/Atg2 family: conserved functional model (lipid transfer + tethering)
A widely supported model (largely from budding yeast and mammalian systems) is that ATG2/Atg2 is a rod-like, bridge-like lipid transfer protein that operates at phagophore–ER membrane contact sites, providing lipid flux to support phagophore expansion; Atg2 also acts as a tether between membranes. (duarte2023theorganizationand pages 4-5, duarte2023theorganizationand pages 5-7, duarte2023theorganizationand pages 7-9)
A second core concept is cooperation between ATG2 and ATG9: ATG2 provides non-vesicular lipid transfer, while ATG9 functions as a lipid scramblase to equilibrate lipids across bilayer leaflets, enabling net membrane expansion. (duarte2023theorganizationand pages 13-14, choi2024emergingrolesof pages 7-8, duarte2023theorganizationand pages 4-5)
2) Organism-specific functional annotation: what is known in S. pombe
2.1 Primary biological role
In S. pombe, Atg2 is required for starvation-induced bulk autophagy as measured by the standard CFP-Atg8 processing assay: atg2Δ mutants fail to show CFP-Atg8 processing under nitrogen starvation, indicating blocked autophagy flux. (sun2013globalanalysisof pages 3-5)
2.2 Phenotypes and pathway position inferred from Atg8 dynamics
Live-cell imaging in S. pombe shows that atg2Δ mutants form Atg8 puncta that are abnormally numerous and long-lived, consistent with impaired progression/maturation of autophagic structures. (sun2013globalanalysisof pages 3-5)
These phenotypes position Atg2 as a factor required for normal phagophore/autophagosome biogenesis dynamics rather than merely for autophagosome-vacuole fusion. (sun2013globalanalysisof pages 3-5)
2.3 Localization: PAS/phagophore rim (direct S. pombe evidence)
A key advance for S. pombe Atg2 localization comes from high-resolution imaging/EM in 2023. In S. pombe, Atg2 is described as residing at the phagophore rim, a highly curved region (reported rim diameter ~15–17 nm), and Atg2 is among factors thought to localize to this rim. (wang2023aconservedmembrane pages 1-3)
Wang et al. (2023) directly support rim/tip localization in S. pombe using:
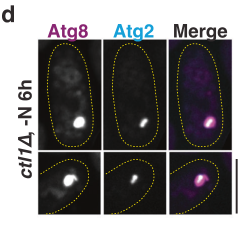
- Fluorescence microscopy: in ctl1Δ cells with enlarged phagophores, Atg2 localizes to a sub-region of the Atg8-positive structure (consistent with subdomain/rim restriction). (wang2023aconservedmembrane pages 8-10, wang2023aconservedmembrane media e70bb975)
- APEX2 EM labeling: Atg2-APEX2 electron-dense precipitate concentrates at the tips/rims of cup-shaped phagophores, whereas Atg5 shows more even distribution, supporting specific Atg2 enrichment at phagophore extremities. (wang2023aconservedmembrane pages 8-10, wang2023aconservedmembrane media e70bb975)
Quantitative localization statistic: Wang et al. quantify that ~80% of open phagophores exhibit rim labeling for Atg2 by APEX2 EM. (wang2023aconservedmembrane media e70bb975)
(Visual evidence is provided in cropped Figure 6 panels showing Atg2 subdomain localization and APEX2 EM rim labeling/quantification.) (wang2023aconservedmembrane media e70bb975, wang2023aconservedmembrane media 0cd67841, wang2023aconservedmembrane media fe55a58b)
2.4 Interacting/functional partners and trafficking roles (Atg9, Ctl1, Atg18)
Atg9 and Ctl1 recycling from the PAS depends on Atg2: In starved S. pombe atg2Δ (and atg1Δ) cells, Atg9 puncta almost completely overlap with Atg8 puncta, leading to the conclusion that Atg9 recycling from PAS requires Atg1 and Atg2. (sun2013globalanalysisof pages 7-10)
Similarly, Ctl1 becomes largely restricted to the PAS in starved atg2Δ cells, indicating Atg2 is also required for Ctl1 recycling/retrograde trafficking away from the PAS. (sun2013globalanalysisof pages 7-10)
Atg9–Ctl1 physical association: Atg9 and Ctl1 co-immunoprecipitate in S. pombe, supporting that these proteins operate in a shared module whose correct cycling/localization requires Atg2. (sun2013globalanalysisof pages 7-10)
Atg18-family linkage (review synthesis in S. pombe): The fission-yeast review states that Atg2 is recruited to the PAS, and that Atg18a promotes PAS targeting of Atg2 (noted as unpublished in the review). The review also states that deletion of atg1 or atg2 restricts Atg9 and Ctl1 to the PAS, reinforcing the primary-study findings. (xu2022fissionyeastautophagy pages 4-5, xu2022fissionyeastautophagy pages 5-7)
3) Mechanistic interpretation: what can be inferred from conserved ATG2 biology (clearly labeled as inference)
Although direct biochemical lipid-transfer measurements for S. pombe Atg2 were not found in the retrieved S. pombe texts, mechanistic work in other systems provides a coherent model consistent with S. pombe phenotypes and rim localization.
3.1 Budding yeast Atg2 as ER–phagophore tether (conserved background)
In S. cerevisiae, Atg2 forms a complex with Atg18 and tethers pre-autophagosomal membranes to the ER, with distinct membrane-binding regions at the N- and C-termini; loss of Atg2–Atg18 blocks isolation membrane formation even when other Atg proteins accumulate at PAS. (kotani2018theatg2atg18complex pages 1-2)
3.2 Phagophore–ER contact site organization (2023 review)
A 2023 review on phagophore–ER membrane contact sites summarizes that ATG2 proteins are rod-like (~20 nm) bridge-like factors with lipid-handling features (hydrophobic groove/RBG modules) and that Atg2/ATG2 has membrane-binding sites at both ends—an N-terminal ER-associated lipid-transfer module and a C-terminal region involved in phagophore association and interaction with Atg9 and Atg18/WIPI proteins. (duarte2023theorganizationand pages 4-5, duarte2023theorganizationand pages 5-7, duarte2023theorganizationand pages 7-9)
3.3 ATG9–ATG2 cooperation and scramblase/bridge model (2023–2024 synthesis)
The same review emphasizes that ATG9/Atg9 is now strongly supported as a lipid scramblase and that ATG2 (and VPS13-family bridges) mediate lipid transfer to growing phagophores; together, these data support a model where ATG2 supplies lipids while ATG9 equilibrates lipids across leaflets to enable bilayer expansion. (duarte2023theorganizationand pages 13-14, duarte2023theorganizationand pages 4-5, duarte2023theorganizationand pages 1-2)
A 2024 review focused on ATG9/ATG9A similarly describes a cooperative model in which ATG9 and ATG2 function at ER–phagophore contact sites to drive phagophore expansion, while noting open questions such as how cells enforce directionality of lipid flow. (choi2024emergingrolesof pages 7-8)
4) Recent developments (prioritizing 2023–2024)
4.1 S. pombe (2023): high-resolution localization at the phagophore rim
Wang et al. (Aug 2023, Nature Communications) provide a strong S. pombe-specific advance by placing Atg2 at the phagophore rim/tips via APEX2 EM and by quantifying rim labeling in ~80% of open phagophores, giving high-confidence spatial context for function. (wang2023aconservedmembrane media e70bb975, wang2023aconservedmembrane pages 8-10)
4.2 Field-wide (2023): emphasis on phagophore–ER contact sites as functional units
Duarte & Reggiori (Jan 2023, Contact) highlight phagophore–ER membrane contact sites as organizational centers where ATG2, ATG9, and Atg18/WIPI accumulate and act, and summarize a shift from vesicle-fusion-only models toward a lipid-transfer/scrambling framework for membrane growth. (duarte2023theorganizationand pages 1-2, duarte2023theorganizationand pages 13-14)
4.3 Mammalian (2024): alternative lipid sources and new bridging factors
Wei et al. (Apr 2024, Cell Discovery) identify ANKFY1 as an ATG2A-binding FYVE-domain protein that binds PI3P and recruits/anchors ATG2A to PI3P-rich endosomal membranes; ANKFY1 approximately doubles ATG2A-mediated lipid transfer in an in vitro assay, and ANKFY1 depletion phenocopies ATG2A/B depletion (reduced autophagy flux, impaired autophagosome growth/closure). This expands the prevailing view that the ER is the only lipid donor by adding an endosomal lipid-transfer route to phagophores. (wei2024ankfy1bridgesatg2amediated pages 10-12, wei2024ankfy1bridgesatg2amediated pages 1-2, wei2024ankfy1bridgesatg2amediated pages 4-8)
5) Current applications and real-world implementations
5.1 S. pombe Atg2 as a model for conserved autophagy membrane growth
Because ATG2 family proteins are central to autophagosome biogenesis, S. pombe Atg2 is practically used as a genetically tractable model to interrogate conserved principles of membrane growth and organelle contact-site biology. The combination of strong loss-of-function phenotypes (blocked CFP-Atg8 processing) and high-resolution localization (phagophore rim) provides an experimentally robust system for mechanistic dissection. (sun2013globalanalysisof pages 3-5, wang2023aconservedmembrane media e70bb975)
5.2 Translational relevance: autophagy modulation in disease contexts (mechanistic bridge)
In mammals, ATG2-dependent lipid transfer and the ATG2–ATG9 partnership are active targets of mechanistic cell biology because autophagy influences neurodegeneration, infection biology, cancer, and metabolic disease; the 2024 ANKFY1 work illustrates that identifying new regulators of ATG2-mediated lipid flux can reveal new intervention points or biomarkers of autophagosome growth/closure. (wei2024ankfy1bridgesatg2amediated pages 10-12, wei2024ankfy1bridgesatg2amediated pages 4-8, choi2024emergingrolesof pages 7-8)
6) Expert opinions / authoritative synthesis (what experts emphasize)
Two authoritative recent reviews converge on the following expert consensus and open questions:
1) Consensus: phagophore growth is driven largely by lipid transfer at membrane contact sites, with ATG2/Atg2 as a key bridge-like lipid-transfer factor and ATG9 as a scramblase. (duarte2023theorganizationand pages 1-2, choi2024emergingrolesof pages 7-8)
2) Open questions: precise molecular coordination among ATG2, ATG9, and Atg18/WIPI (timing, directionality, and regulation of lipid flow) remains incompletely resolved. (duarte2023theorganizationand pages 1-2, choi2024emergingrolesof pages 7-8)
7) Key quantitative statistics and data (recent studies)
In S. pombe (Wang et al., 2023):
- ~80% of open phagophores show Atg2 rim labeling by APEX2 EM. (wang2023aconservedmembrane media e70bb975)
- Following nitrogen starvation, >80% of phagophores marked by Atg8 or Atg2 contain Rop1, consistent with early co-recruitment. (wang2023aconservedmembrane pages 8-10)
- Phagophore rim diameter reported as ~15–17 nm (provides scale for curvature at the Atg2-enriched rim). (wang2023aconservedmembrane pages 1-3)
In mammalian mechanistic reconstitution (Wei et al., 2024):
- ANKFY1 approximately doubles ATG2A-mediated lipid transfer in vitro in a FRET-based liposome assay; enhancement depends on PI3P and ANKFY1 FYVE domain integrity. (wei2024ankfy1bridgesatg2amediated pages 10-12)
8) Summary functional annotation for UniProt O94649 (Atg2/mug36) in S. pombe
Primary function (cell biological): Atg2 is an essential factor for starvation-induced macroautophagy in S. pombe and is required for normal phagophore/autophagosome biogenesis dynamics. (sun2013globalanalysisof pages 3-5)
Likely mechanistic role in S. pombe (supported by localization + conserved model): Atg2 localizes to the phagophore rim/tips, consistent with an organizational role at membrane contact sites that support phagophore expansion; its loss leads to accumulation of stalled Atg8 structures. (wang2023aconservedmembrane media e70bb975, sun2013globalanalysisof pages 3-5)
Localization: PAS/phagophore-associated, enriched at rims/tips of cup-shaped phagophores by APEX2 EM; subdomain localization on enlarged phagophores in ctl1Δ. (wang2023aconservedmembrane media e70bb975, wang2023aconservedmembrane pages 8-10)
Pathway/module membership: Functionally linked to the Atg9/Ctl1 trafficking/cycling module; Atg2 is required for recycling of Atg9 and Ctl1 away from PAS under starvation; Ctl1 and Atg9 physically associate. (sun2013globalanalysisof pages 7-10)
9) Evidence table
| Aspect (function/localization/partner/phenotype/quant data) | Key finding | Experimental basis (assay) | Organism/system | Citation (author year journal) + URL | Context ID |
|---|---|---|---|---|---|
| function | S. pombe-specific: Atg2 is required for starvation-induced autophagy; atg2Δ lacked CFP-Atg8 processing, consistent with blocked autophagy flux. | CFP-Atg8 processing assay under nitrogen starvation | Schizosaccharomyces pombe | Sun et al. 2013, PLoS Genetics — https://doi.org/10.1371/journal.pgen.1003715 | (sun2013globalanalysisof pages 3-5) |
| phenotype | S. pombe-specific: atg2Δ cells showed Atg8 puncta that were more numerous than wild type and long-lived, indicating defective progression/maturation of autophagic structures. | Time-lapse fluorescence microscopy of Atg8 puncta | Schizosaccharomyces pombe | Sun et al. 2013, PLoS Genetics — https://doi.org/10.1371/journal.pgen.1003715 | (sun2013globalanalysisof pages 3-5) |
| partner/localization | S. pombe-specific: Atg2 is required for retrograde trafficking of Atg9 from the PAS; in starved atg2Δ cells, Atg9 puncta almost completely overlapped with Atg8 puncta, indicating PAS retention. | Fluorescent protein co-localization microscopy (Atg9 with Atg8) in mutants | Schizosaccharomyces pombe | Sun et al. 2013, PLoS Genetics — https://doi.org/10.1371/journal.pgen.1003715 | (sun2013globalanalysisof pages 7-10) |
| partner/localization | S. pombe-specific: Ctl1 became largely restricted to the PAS in starved atg2Δ cells, supporting an Atg2-dependent recycling step for Ctl1 as well as Atg9. | Fluorescent localization of YFP-Ctl1 with Atg8 in mutants | Schizosaccharomyces pombe | Sun et al. 2013, PLoS Genetics — https://doi.org/10.1371/journal.pgen.1003715 | (sun2013globalanalysisof pages 7-10) |
| partner | S. pombe-specific: Ctl1 physically interacts with Atg9; together with the atg2Δ localization phenotype, this places Atg2 in the Atg9/Ctl1 trafficking module at the PAS. | Co-immunoprecipitation plus localization genetics | Schizosaccharomyces pombe | Sun et al. 2013, PLoS Genetics — https://doi.org/10.1371/journal.pgen.1003715 | (sun2013globalanalysisof pages 7-10) |
| localization | S. pombe-specific: Atg2 localizes to a sub-region of enlarged Atg8-positive phagophores in ctl1Δ cells, consistent with phagophore rim/subdomain localization rather than uniform distribution. | Dual-color fluorescence microscopy (Atg2-tdT with Atg8 marker) | Schizosaccharomyces pombe | Wang et al. 2023, Nature Communications — https://doi.org/10.1038/s41467-023-40530-4 | (wang2023aconservedmembrane pages 8-10, wang2023aconservedmembrane media e70bb975) |
| localization | S. pombe-specific: APEX2 EM labeling placed Atg2 at the tips/rims of cup-shaped phagophores; unlike Atg5, labeling was not evenly distributed over the phagophore. | Correlative ultrastructure with APEX2 electron microscopy | Schizosaccharomyces pombe | Wang et al. 2023, Nature Communications — https://doi.org/10.1038/s41467-023-40530-4 | (wang2023aconservedmembrane pages 8-10, wang2023aconservedmembrane media e70bb975) |
| quant data | S. pombe-specific: In Wang et al., approximately 80% of open phagophores showed rim labeling for Atg2 by APEX2 EM quantification. | Quantified APEX2 EM localization | Schizosaccharomyces pombe | Wang et al. 2023, Nature Communications — https://doi.org/10.1038/s41467-023-40530-4 | (wang2023aconservedmembrane media e70bb975) |
| quant data | S. pombe-specific: Following nitrogen starvation, >80% of Atg8- or Atg2-marked phagophores contained Rop1, supporting early co-recruitment of a curvature factor with Atg2 at forming phagophores. | Live-cell fluorescence microscopy and colocalization quantification | Schizosaccharomyces pombe | Wang et al. 2023, Nature Communications — https://doi.org/10.1038/s41467-023-40530-4 | (wang2023aconservedmembrane pages 8-10) |
| localization/quant data | S. pombe-specific: Wang et al. discuss Atg2 at the highly curved phagophore rim; the reported yeast phagophore rim diameter is about 15–17 nm, consistent with Atg2 acting at extreme membrane curvature. | Ultrastructural interpretation integrated with localization data | Schizosaccharomyces pombe | Wang et al. 2023, Nature Communications — https://doi.org/10.1038/s41467-023-40530-4 | (wang2023aconservedmembrane pages 1-3) |
| localization/partner | S. pombe-specific review: Atg2 is recruited to the PAS, and Atg18a promotes PAS targeting of Atg2; deletion of atg1 or atg2 restricts Atg9 and Ctl1 localization to the PAS. | Review synthesis of fluorescence localization/genetic studies | Schizosaccharomyces pombe | Xu & Du 2022, Cells — https://doi.org/10.3390/cells11071086 | (xu2022fissionyeastautophagy pages 5-7, xu2022fissionyeastautophagy pages 7-8, xu2022fissionyeastautophagy pages 4-5) |
| function/localization | Non-S. pombe / inferred: Current consensus model is that ATG2/Atg2 is a rod-like lipid transfer protein and tether at phagophore-ER membrane contact sites, with an N-terminal ER-associated lipid-transfer module and C-terminal phagophore-binding region acting with Atg9 and Atg18/WIPI proteins. | Review of structural, biochemical, and cell-biological studies | Mainly budding yeast and mammalian systems; inferred to S. pombe by conservation | Duarte & Reggiori 2023, Contact — https://doi.org/10.1177/25152564231183898 | (duarte2023theorganizationand pages 4-5, duarte2023theorganizationand pages 5-7, duarte2023theorganizationand pages 7-9, duarte2023theorganizationand pages 1-2) |
| partner/function | Non-S. pombe / inferred: ATG2 works cooperatively with ATG9, which acts as a lipid scramblase; the combined model is that ATG2 transfers lipids into the phagophore while ATG9 equilibrates lipids across leaflets to support membrane expansion. | Review of recent cryo-EM, biochemical reconstitution, and genetics | Yeast and mammalian systems; inferred to S. pombe by conservation | Duarte & Reggiori 2023, Contact — https://doi.org/10.1177/25152564231183898; Choi et al. 2024, Autophagy — https://doi.org/10.1080/15548627.2024.2384349 | (duarte2023theorganizationand pages 13-14, duarte2023theorganizationand pages 4-5, choi2024emergingrolesof pages 7-8) |
| function/localization | Non-S. pombe / inferred: Budding-yeast Atg2-Atg18 tethers pre-autophagosomal membranes to the ER; Atg2 has membrane-binding regions at both ends, and loss of the complex blocks isolation membrane formation despite PAS accumulation of other Atg proteins. | Primary mechanistic study using mutagenesis, in vitro liposome tethering, PAS/ER localization assays | Saccharomyces cerevisiae; mechanistic inference for S. pombe homolog | Kotani et al. 2018, PNAS — https://doi.org/10.1073/pnas.1806727115 | (kotani2018theatg2atg18complex pages 1-2) |
| partner/localization | Non-S. pombe / inferred: Atg9 helps establish Atg2-dependent ER-phagophore contact sites and confines Atg2 to phagophore extremities; disrupting Atg2-Atg9 interaction causes abnormal Atg2/ER-contact distribution and severe autophagy defects. | Primary mechanistic study with Atg2 point mutants, localization analysis, and ultrastructure | Saccharomyces cerevisiae; mechanistic inference for S. pombe homolog | Gómez-Sánchez et al. 2018, Journal of Cell Biology — https://doi.org/10.1083/jcb.201710116 | (gomezsanchez2018atg9establishesatg2dependent pages 21-21) |
Table: This table compiles organism-specific evidence for Schizosaccharomyces pombe Atg2/mug36 and separates it from mechanistic inferences drawn from conserved Atg2 studies in other systems. It is useful for distinguishing direct annotation evidence from broader family-level functional models.
10) Key source list with URLs and publication dates
- Sun L-L et al. (2013-08). PLoS Genetics. “Global Analysis of Fission Yeast Mating Genes Reveals New Autophagy Factors.” https://doi.org/10.1371/journal.pgen.1003715 (sun2013globalanalysisof pages 3-5, sun2013globalanalysisof pages 7-10)
- Xu D-D, Du L-L. (2022-03). Cells. “Fission Yeast Autophagy Machinery.” https://doi.org/10.3390/cells11071086 (xu2022fissionyeastautophagy pages 5-7, xu2022fissionyeastautophagy pages 4-5)
- Wang N et al. (2023-08). Nature Communications. “A conserved membrane curvature-generating protein is crucial for autophagosome formation in fission yeast.” https://doi.org/10.1038/s41467-023-40530-4 (wang2023aconservedmembrane media e70bb975, wang2023aconservedmembrane pages 8-10, wang2023aconservedmembrane pages 1-3)
- Duarte PV, Reggiori F. (2023-01). Contact. “The Organization and Function of the Phagophore-ER Membrane Contact Sites.” https://doi.org/10.1177/25152564231183898 (duarte2023theorganizationand pages 1-2, duarte2023theorganizationand pages 5-7)
- Choi J et al. (2024-08). Autophagy. “Emerging roles of ATG9/ATG9A in autophagy: implications for cell and neurobiology.” https://doi.org/10.1080/15548627.2024.2384349 (choi2024emergingrolesof pages 7-8)
- Wei B et al. (2024-04). Cell Discovery. “ANKFY1 bridges ATG2A-mediated lipid transfer from endosomes to phagophores.” https://doi.org/10.1038/s41421-024-00659-y (wei2024ankfy1bridgesatg2amediated pages 10-12, wei2024ankfy1bridgesatg2amediated pages 4-8)
Limitations of the current evidence set
Direct biochemical demonstration of lipid-transfer activity for S. pombe Atg2 itself (e.g., purified SpAtg2 lipid-transfer kinetics) was not located in the retrieved S. pombe texts; the lipid-transfer/tether model is therefore presented as conserved mechanistic inference supported by authoritative reviews and non-S. pombe primary studies, while the core S. pombe phenotypes and localization are directly evidenced as above. (duarte2023theorganizationand pages 5-7, kotani2018theatg2atg18complex pages 1-2, wang2023aconservedmembrane media e70bb975)
References
-
(sun2013globalanalysisof pages 3-5): Ling-Ling Sun, Ming Li, Fang Suo, Xiao-Man Liu, En-Zhi Shen, Bing Yang, Meng-Qiu Dong, Wan-Zhong He, and Li-Lin Du. Global analysis of fission yeast mating genes reveals new autophagy factors. PLoS Genetics, 9:e1003715, Aug 2013. URL: https://doi.org/10.1371/journal.pgen.1003715, doi:10.1371/journal.pgen.1003715. This article has 123 citations and is from a domain leading peer-reviewed journal.
-
(sun2013globalanalysisof pages 7-10): Ling-Ling Sun, Ming Li, Fang Suo, Xiao-Man Liu, En-Zhi Shen, Bing Yang, Meng-Qiu Dong, Wan-Zhong He, and Li-Lin Du. Global analysis of fission yeast mating genes reveals new autophagy factors. PLoS Genetics, 9:e1003715, Aug 2013. URL: https://doi.org/10.1371/journal.pgen.1003715, doi:10.1371/journal.pgen.1003715. This article has 123 citations and is from a domain leading peer-reviewed journal.
-
(xu2022fissionyeastautophagy pages 5-7): Dan-Dan Xu and Li-Lin Du. Fission yeast autophagy machinery. Cells, 11:1086, Mar 2022. URL: https://doi.org/10.3390/cells11071086, doi:10.3390/cells11071086. This article has 29 citations.
-
(kotani2018theatg2atg18complex pages 1-2): Tetsuya Kotani, Hiromi Kirisako, Michiko Koizumi, Yoshinori Ohsumi, and Hitoshi Nakatogawa. The atg2-atg18 complex tethers pre-autophagosomal membranes to the endoplasmic reticulum for autophagosome formation. Proceedings of the National Academy of Sciences, 115:10363-10368, Sep 2018. URL: https://doi.org/10.1073/pnas.1806727115, doi:10.1073/pnas.1806727115. This article has 328 citations and is from a highest quality peer-reviewed journal.
-
(gomezsanchez2018atg9establishesatg2dependent pages 21-21): Rubén Gómez-Sánchez, Jaqueline Rose, Rodrigo Guimarães, Muriel Mari, Daniel Papinski, Ester Rieter, Willie J. Geerts, Ralph Hardenberg, Claudine Kraft, Christian Ungermann, and Fulvio Reggiori. Atg9 establishes atg2-dependent contact sites between the endoplasmic reticulum and phagophores. The Journal of Cell Biology, 217:2743-2763, May 2018. URL: https://doi.org/10.1083/jcb.201710116, doi:10.1083/jcb.201710116. This article has 319 citations.
-
(duarte2023theorganizationand pages 5-7): Prado Vargas Duarte and Fulvio Reggiori. The organization and function of the phagophore-er membrane contact sites. Contact, Jan 2023. URL: https://doi.org/10.1177/25152564231183898, doi:10.1177/25152564231183898. This article has 14 citations.
-
(xu2022fissionyeastautophagy pages 4-5): Dan-Dan Xu and Li-Lin Du. Fission yeast autophagy machinery. Cells, 11:1086, Mar 2022. URL: https://doi.org/10.3390/cells11071086, doi:10.3390/cells11071086. This article has 29 citations.
-
(duarte2023theorganizationand pages 4-5): Prado Vargas Duarte and Fulvio Reggiori. The organization and function of the phagophore-er membrane contact sites. Contact, Jan 2023. URL: https://doi.org/10.1177/25152564231183898, doi:10.1177/25152564231183898. This article has 14 citations.
-
(duarte2023theorganizationand pages 7-9): Prado Vargas Duarte and Fulvio Reggiori. The organization and function of the phagophore-er membrane contact sites. Contact, Jan 2023. URL: https://doi.org/10.1177/25152564231183898, doi:10.1177/25152564231183898. This article has 14 citations.
-
(duarte2023theorganizationand pages 13-14): Prado Vargas Duarte and Fulvio Reggiori. The organization and function of the phagophore-er membrane contact sites. Contact, Jan 2023. URL: https://doi.org/10.1177/25152564231183898, doi:10.1177/25152564231183898. This article has 14 citations.
-
(choi2024emergingrolesof pages 7-8): Jiyoung Choi, Haeun Jang, Zhao Xuan, and Daehun Park. Emerging roles of atg9/atg9a in autophagy: implications for cell and neurobiology. Autophagy, 20:2373-2387, Aug 2024. URL: https://doi.org/10.1080/15548627.2024.2384349, doi:10.1080/15548627.2024.2384349. This article has 25 citations and is from a domain leading peer-reviewed journal.
-
(wang2023aconservedmembrane pages 1-3): Ning Wang, Yoko Shibata, Joao A. Paulo, Steven P. Gygi, and Tom A. Rapoport. A conserved membrane curvature-generating protein is crucial for autophagosome formation in fission yeast. Nature Communications, Aug 2023. URL: https://doi.org/10.1038/s41467-023-40530-4, doi:10.1038/s41467-023-40530-4. This article has 24 citations and is from a highest quality peer-reviewed journal.
-
(wang2023aconservedmembrane pages 8-10): Ning Wang, Yoko Shibata, Joao A. Paulo, Steven P. Gygi, and Tom A. Rapoport. A conserved membrane curvature-generating protein is crucial for autophagosome formation in fission yeast. Nature Communications, Aug 2023. URL: https://doi.org/10.1038/s41467-023-40530-4, doi:10.1038/s41467-023-40530-4. This article has 24 citations and is from a highest quality peer-reviewed journal.
-
(wang2023aconservedmembrane media e70bb975): Ning Wang, Yoko Shibata, Joao A. Paulo, Steven P. Gygi, and Tom A. Rapoport. A conserved membrane curvature-generating protein is crucial for autophagosome formation in fission yeast. Nature Communications, Aug 2023. URL: https://doi.org/10.1038/s41467-023-40530-4, doi:10.1038/s41467-023-40530-4. This article has 24 citations and is from a highest quality peer-reviewed journal.
-
(wang2023aconservedmembrane media 0cd67841): Ning Wang, Yoko Shibata, Joao A. Paulo, Steven P. Gygi, and Tom A. Rapoport. A conserved membrane curvature-generating protein is crucial for autophagosome formation in fission yeast. Nature Communications, Aug 2023. URL: https://doi.org/10.1038/s41467-023-40530-4, doi:10.1038/s41467-023-40530-4. This article has 24 citations and is from a highest quality peer-reviewed journal.
-
(wang2023aconservedmembrane media fe55a58b): Ning Wang, Yoko Shibata, Joao A. Paulo, Steven P. Gygi, and Tom A. Rapoport. A conserved membrane curvature-generating protein is crucial for autophagosome formation in fission yeast. Nature Communications, Aug 2023. URL: https://doi.org/10.1038/s41467-023-40530-4, doi:10.1038/s41467-023-40530-4. This article has 24 citations and is from a highest quality peer-reviewed journal.
-
(duarte2023theorganizationand pages 1-2): Prado Vargas Duarte and Fulvio Reggiori. The organization and function of the phagophore-er membrane contact sites. Contact, Jan 2023. URL: https://doi.org/10.1177/25152564231183898, doi:10.1177/25152564231183898. This article has 14 citations.
-
(wei2024ankfy1bridgesatg2amediated pages 10-12): Bin Wei, Yuhui Fu, Xiuzhi Li, Fang Chen, Yiqing Zhang, Hanmo Chen, Mindan Tong, Linsen Li, Yi Pan, Shen Zhang, She Chen, Xiaoxia Liu, and Qing Zhong. Ankfy1 bridges atg2a-mediated lipid transfer from endosomes to phagophores. Cell Discovery, Apr 2024. URL: https://doi.org/10.1038/s41421-024-00659-y, doi:10.1038/s41421-024-00659-y. This article has 13 citations and is from a peer-reviewed journal.
-
(wei2024ankfy1bridgesatg2amediated pages 1-2): Bin Wei, Yuhui Fu, Xiuzhi Li, Fang Chen, Yiqing Zhang, Hanmo Chen, Mindan Tong, Linsen Li, Yi Pan, Shen Zhang, She Chen, Xiaoxia Liu, and Qing Zhong. Ankfy1 bridges atg2a-mediated lipid transfer from endosomes to phagophores. Cell Discovery, Apr 2024. URL: https://doi.org/10.1038/s41421-024-00659-y, doi:10.1038/s41421-024-00659-y. This article has 13 citations and is from a peer-reviewed journal.
-
(wei2024ankfy1bridgesatg2amediated pages 4-8): Bin Wei, Yuhui Fu, Xiuzhi Li, Fang Chen, Yiqing Zhang, Hanmo Chen, Mindan Tong, Linsen Li, Yi Pan, Shen Zhang, She Chen, Xiaoxia Liu, and Qing Zhong. Ankfy1 bridges atg2a-mediated lipid transfer from endosomes to phagophores. Cell Discovery, Apr 2024. URL: https://doi.org/10.1038/s41421-024-00659-y, doi:10.1038/s41421-024-00659-y. This article has 13 citations and is from a peer-reviewed journal.
-
(xu2022fissionyeastautophagy pages 7-8): Dan-Dan Xu and Li-Lin Du. Fission yeast autophagy machinery. Cells, 11:1086, Mar 2022. URL: https://doi.org/10.3390/cells11071086, doi:10.3390/cells11071086. This article has 29 citations.
Artifacts
Citations
- sun2013globalanalysisof pages 3-5
- wang2023aconservedmembrane pages 1-3
- sun2013globalanalysisof pages 7-10
- choi2024emergingrolesof pages 7-8
- wang2023aconservedmembrane pages 8-10
- xu2022fissionyeastautophagy pages 5-7
- duarte2023theorganizationand pages 5-7
- xu2022fissionyeastautophagy pages 4-5
- duarte2023theorganizationand pages 4-5
- duarte2023theorganizationand pages 7-9
- duarte2023theorganizationand pages 13-14
- duarte2023theorganizationand pages 1-2
- xu2022fissionyeastautophagy pages 7-8
- https://doi.org/10.1371/journal.pgen.1003715
- https://doi.org/10.1038/s41467-023-40530-4
- https://doi.org/10.3390/cells11071086
- https://doi.org/10.1177/25152564231183898
- https://doi.org/10.1177/25152564231183898;
- https://doi.org/10.1080/15548627.2024.2384349
- https://doi.org/10.1073/pnas.1806727115
- https://doi.org/10.1083/jcb.201710116
- https://doi.org/10.1038/s41421-024-00659-y
- https://doi.org/10.1371/journal.pgen.1003715,
- https://doi.org/10.3390/cells11071086,
- https://doi.org/10.1073/pnas.1806727115,
- https://doi.org/10.1083/jcb.201710116,
- https://doi.org/10.1177/25152564231183898,
- https://doi.org/10.1080/15548627.2024.2384349,
- https://doi.org/10.1038/s41467-023-40530-4,
- https://doi.org/10.1038/s41421-024-00659-y,
Perplexity
(atg2-deep-research-perplexity.md)
Perplexity
(atg2-deep-research-perplexity.md)Autophagy-Related Protein 2 (Atg2) in Schizosaccharomyces pombe: Structure, Function, and Mechanisms in Autophagosome Biogenesis
Summary of Key Findings
Autophagy-related protein 2 (Atg2, UniProt O94649) from the fission yeast Schizosaccharomyces pombe is a large multifunctional protein representing one of the most evolutionarily conserved yet previously enigmatic components of the autophagy machinery. Recent structural and biochemical advances have revealed that Atg2 functions as a novel nonvesicular phospholipid transfer protein that mediates de novo autophagosome formation through dual mechanisms: physically tethering the endoplasmic reticulum (ER) to the expanding phagophore membrane while simultaneously transferring lipids between these two membrane compartments[1][7]. The protein localizes exclusively to contact sites between the ER exit sites (ERES) and the edge of the isolation membrane (IM), where it operates in complex with the phosphoinositide-binding protein Atg18 and the lipid scramblase Atg9 to coordinate the massive lipid redistribution required for autophagosome expansion. This discovery has fundamentally changed our understanding of how cells build the double-membrane autophagosomes during autophagy, revealing that lipid transfer proteins can directly mediate de novo organelle biogenesis rather than relying solely on vesicular trafficking mechanisms.
Historical Discovery and Early Characterization of Atg2
Initial Identification and Genetic Basis
The atg2 gene was first identified in 1993 through a comprehensive screening of Saccharomyces cerevisiae autophagy-defective mutants that discovered thirteen autophagy genes, designated apg genes, which were later renamed to the atg nomenclature[1]. In S. pombe, the gene was identified through a large-scale screen for genes required for critical meiotic events and is also known by its alternative name mug36 (meiotically up-regulated gene 36), reflecting its initial discovery in the context of meiotic processes[2][21]. Eight years after the initial identification, in the early 2000s, the atg2 gene was formally cloned and sequenced, establishing that the protein product was essential for autophagosome formation during starvation-induced autophagy[1].
Early Functional Recognition
Even before detailed molecular characterization, researchers recognized that Atg2 possessed unusual properties as a peripheral membrane protein with high affinity for membranes[1]. The protein's behavior during autophagy was distinctive: it accumulated at the preautophagosomal structure (PAS, also called the phagophore assembly site) and localized specifically to nascent autophagosomal structures during their formation and expansion. When the atg2 gene was deleted in yeast strains, autophagy was severely impaired, resulting in accumulation of autophagic cargo and defective autophagosome formation. However, the precise molecular mechanism by which Atg2 contributed to autophagy remained mysterious for decades, earning it a reputation as one of the most poorly understood proteins in the autophagy field[1][7].
Structural Organization and Molecular Architecture
Full-Length Protein Architecture
Atg2 proteins are exceptionally large molecules, with Schizosaccharomyces pombe Atg2 comprising 1,646 amino acid residues, and mammalian ATG2A reaching 1,938 residues[2]. Despite these large sizes, the protein maintains a characteristic elongated, rod-like morphology that extends approximately 20 nanometers in length with a width of roughly 30 angstroms[8]. This extended structure is fundamentally different from the compact, globular folds typical of most proteins, instead resembling a long bridge or tunnel-like apparatus.
Conserved Domains and Functional Modules
At the molecular level, Atg2 proteins are organized into distinct functional domains with remarkable conservation across eukaryotes. The most highly conserved region resides at the N-terminus, where a domain designated Chorein_N (named after its similarity to domains in the protein Chorein, also called VPS13) contains the primary lipid transfer module[1][8][9]. This Chorein_N domain has been crystallized from Schizosaccharomyces pombe Atg2 (designated SpAtg2^NR^) at 2.7 angstrom resolution, revealing a globular structure comprising a helical region and a twisted β-sheet that fold together to create a large hydrophobic cavity[1][50]. When phosphatidylethanolamine (PE), a major cellular glycerophospholipid, is mixed with the SpAtg2^NR^ domain in crystallization conditions, a complex forms in which the PE molecule binds with its acyl chains buried within the hydrophobic cavity while the phosphate head group remains exposed to the aqueous solvent[1][50].
The central region of Atg2 contains five repeat structures primarily composed of β-strands rich in hydrophobic residues, predicted to form β-jellyroll-like folds that create successive hydrophobic grooves or pathways[1][50]. These repeating structures are proposed to form a continuous hydrophobic channel extending through the rod-shaped molecule. At the C-terminal end, Atg2 proteins contain a specialized domain designated ATG2_CAD (a cysteine-alanine-aspartic acid triad region) that appears to form one of the membrane-binding surfaces in complex with the cofactor Atg18[1][7][28].
Two distinct membrane-binding regions have been identified through truncation analysis: one at the N-terminus involving amino acids 2-21, and another at the C-terminus spanning residues 1,347-1,592 in S. cerevisiae Atg2[1][7]. While each binding site alone is sufficient for binding to membranes, both sites are required for the membrane-tethering function, indicating that both tips of the elongated rod simultaneously engage with separate membrane bilayers[1][7][38].
Amphipathic Helix Localization Region
In mammalian ATG2A and ATG2B proteins, and similarly conserved in other organisms, a region designated the C-terminal localization region (CLR) spanning amino acids 1,723-1,829 in ATG2A contains amphipathic α-helices that mediate both autophagosomal membrane association and localization to lipid droplets[16][49]. This region is distinct from the primary lipid transfer module but contributes importantly to proper subcellular targeting of the protein.
Biochemical Functions: Lipid Transfer and Membrane Tethering
Discovery of Lipid Transfer Activity
The recognition that Atg2 functions as a lipid transfer protein emerged from parallel structural and biochemical investigations conducted primarily between 2018 and 2020[1][7][8][9]. Purified mammalian ATG2A protein was shown to bind to liposomes in a phosphoinositide-independent manner, in contrast to other autophagy proteins such as WIPI4 that strictly require high percentages of phosphatidylinositol-3-phosphate (PI3P) for liposome binding[1][8]. This broader binding capability suggested a different molecular basis for Atg2 function.
When researchers performed classic biochemistry experiments mixing purified ATG2A with donor and acceptor liposomes containing fluorescently labeled lipids, they observed transfer of multiple lipid species between the liposomes, including phosphatidylethanolamine (PE), phosphatidylserine (PS), and phosphatidylinositol (PI)[8][9]. Critically, each individual ATG2A molecule can bind and transfer approximately twenty glycerophospholipid molecules simultaneously, distinguishing it from classic lipid transfer proteins that typically transfer one lipid molecule at a time[9]. This high-capacity binding is achieved through the extended hydrophobic cavity running the length of the elongated rod-shaped molecule.
Lipid Transfer Kinetics and Rate Parameters
Quantitative analysis of ATG2A-mediated lipid transfer in in vitro assays revealed transfer rates of approximately 0.017 fluorescent-lipid molecules per protein per second in standard fluorescence-based assays[54]. When accounting for non-fluorescent lipid transfer, which occurs in parallel, the overall transfer rate could reach approximately 50-fold higher, estimating overall transfer rates of potentially 8-10 lipid molecules per protein per second[10][54]. However, even with these optimistic corrections, researchers recognized that the calculated transfer rates appeared insufficient to account for the tens of millions of lipids required to build an entire autophagosome in the observed timeframe of 5-10 minutes[10][54].
Membrane Tethering Activity
In parallel with lipid transfer characterization, biochemical studies revealed that purified ATG2A exhibits robust membrane-tethering (MT) activity[1][7][8]. When ATG2A was incubated with small unilamellar vesicles (SUVs), dynamic light scattering experiments demonstrated a dramatic shift in the size distribution toward larger vesicles, indicating that ATG2A was bridging and clustering SUVs together[11][37][40]. The two tips of the elongated ATG2A molecule act independently to bind separate membranes, spacing them approximately 10-20 nanometers apart—a distance characteristic of organelle contact sites observed throughout the cell[11][40].
The membrane-tethering activity of ATG2A demonstrated preference for membranes containing lipids with lipid-packing defects, such as those rich in phosphatidylethanolamine (PE) or those with high membrane curvature[37][40][51]. Small unilamellar vesicles with high curvature were efficiently tethered by ATG2A, whereas large unilamelular vesicles (LUVs) with lower curvature were not tethered despite maintaining interaction with the protein[40]. This selectivity for high-curvature membranes proved functionally relevant, as both the expanding IM edge and the ERES naturally assume high-curvature geometries in cells[1][50].
Substrate Specificity and Lipid Preference
While ATG2A can transfer multiple glycerophospholipid species, detailed studies examining lipid transfer efficiency revealed important substrate preferences[8][37][40][54]. Transfer occurred most efficiently for lipids with packing defects introduced by small head groups (such as PE) or for negatively charged lipids including phosphatidylserine (PS) and phosphatidylinositol (PI)[37][40][54]. The transfer efficiency depended strongly on both the lipid composition and the presentation geometry of the donor and acceptor membranes, with tethered membranes showing dramatically enhanced transfer rates compared to untethered membranes[37][40].
Notably, PI3P-enriched membranes (such as the nascent phagophore) served as preferred acceptor membranes for ATG2A-mediated lipid transfer, while the phosphoinositide-rich ER provided the donor compartment[8][40][54]. This directional preference, while not absolute, suggests that the phagophore-ER membrane contact sites and the local lipid compositions naturally favor the directionality of lipid flow required for autophagosome expansion[8][40][54].
Protein Interactions and Complex Assembly
The Atg2-Atg18 Complex
Atg2 functions as part of a larger molecular complex with the phosphoinositide-binding protein Atg18 (called WIPI1, WIPI2, or WIPI4 in mammalian cells)[1][7][15][26]. The Atg18 protein belongs to the PROPPIN (phosphoinositide-interacting protein) family and comprises seven WD40 repeats that fold into a characteristic β-propeller structure[26][38][53]. The Atg2-Atg18 complex was first recognized as a functional unit through genetic and cell biological studies, and the physical interaction was subsequently confirmed through biochemical approaches[26].
Recent structural studies revealed that Atg18 binding occurs near the C-terminal region of Atg2, with the 7AB loop of Atg18 serving as a key binding interface[26][29]. The crystal structure of Schizosaccharomyces pombe Atg18 determined at 2.8 angstrom resolution demonstrated that the 7AB loop located on the β-propeller structure provides a critical contact point for Atg2 interaction[26][29]. Disruption of this specific loop by deletion resulted in severely impaired Atg2 binding and loss of Atg2 localization to the phagophore assembly site, demonstrating the functional importance of this interaction[26][29].
Atg18 itself contains two phosphoinositide-binding sites: Site 1 located on blade 5 shows higher affinity for PI(3,5)P₂, while Site 2 on blade 6 exhibits stronger binding to PI(3)P[26][38][53]. The 6CD loop of Atg18 is particularly flexible and prone to forming an amphipathic helix that can insert into lipid bilayers, with this region being critical for the membrane scission activity attributed to the Atg2-Atg18 complex[26][53].
Interaction with Atg9 and Lipid Scramblase Coupling
Emerging research has revealed that Atg2 physically interacts with Atg9, the transmembrane lipid scramblase protein embedded in autophagosomal and vesicular membranes[10][20][39]. The structural basis for this interaction involves alignment of the hydrophobic groove of ATG2A with the internal channel of ATG9A, suggesting direct hand-off of lipids between the transfer and scramblase activities[39]. This coupling represents an elegant solution to the challenge of asymmetric lipid distribution: ATG2 transfers lipids primarily to the outer leaflet of the nascent phagophore, while ATG9 scramblase activity equilibrates lipids between the inner and outer leaflets[20][39].
ATG9A forms a stable trimeric complex with a solvated central pore connected laterally to the cytosol through cavities within each protomer[20]. Molecular dynamics simulations and structural analysis suggest that the central pore of ATG9A can open laterally to accommodate lipid headgroups, enabling lipids to flip between membrane leaflets in an energy-independent manner[20]. The interaction between ATG2A and ATG9A creates a heterotetrameric complex that couples lipid transfer with lipid scrambling for bidirectional lipid distribution[39].
Interaction with Atg1 Complex and Signaling
Recent studies discovered that the Atg2-Atg18 complex interacts with the Atg1 protein kinase complex, involving the C-terminal regions of Atg2[15][18]. This interaction appears to organize the spatial architecture of autophagosome nucleation sites and may coordinate signaling and lipid delivery processes during early autophagosome formation.
Interaction with WIPI4 and PI3P Recognition
In mammalian cells, the WIPI4 protein (the mammalian ortholog of Atg18) associates with ATG2A to facilitate binding to PI3P-enriched phagophore membranes[8][11][40]. The ATG2A-WIPI4 complex exhibits asymmetric membrane binding capabilities through its two tips: the CAD tip binds PI3P-containing membranes via WIPI4, while the N tip can bind PI3P-free membranes such as the ER[11][40]. This asymmetric tethering geometry is particularly important, as it allows the complex to simultaneously recognize the distinct lipid compositions of the phagophore and ER compartments[11][40].
Gabarap Interaction and Autophagosome Closure
Critical recent findings revealed that ATG2A contains a highly conserved LC3 interaction region (LIR) motif at amino acids 1,362-1,365 in ATG2A and 1,491-1,494 in ATG2B[16][55]. This LIR allows direct interaction with the Atg8/LC3/GABARAP family of proteins, which are ubiquitin-like proteins conjugated to phosphatidylethanolamine on autophagosomal membranes[55]. The ATG2A-GABARAP interaction proved essential for autophagosome closure rather than phagophore expansion, with mutations disrupting this interaction resulting in accumulation of unsealed phagophores[55]. Strikingly, this closure function is independent of the ATG2A-WIPI4 interaction required for lipid transfer and membrane tethering[55], demonstrating that Atg2 functions in two sequential, mechanistically independent steps of autophagosome biogenesis.
Subcellular Localization and Membrane Contact Site Assembly
Phagophore-ER Membrane Contact Sites (PERCS)
Schizosaccharomyces pombe Atg2, like its mammalian and budding yeast counterparts, localizes exclusively to contact sites between the ER and the edge of the expanding phagophore, designated phagophore-ER contact sites (PERCS) or more generally as phagophore-ER membrane contact sites (PERMCSs)[7][13][16]. These contact sites represent specialized regions where the two membranes are held in close apposition at distances of 10-20 nanometers, a geometry ideal for nonvesicular lipid transfer[7][13][16].
The localization of Atg2 to PERCS depends critically on several upstream factors[13]. The ER-exit site (ERES) component Sec13 and the TRAPPIII complex work cooperatively with Atg2 to establish and maintain these membrane contact sites[13]. Bimolecular fluorescence complementation (BiFC) studies demonstrated that Atg2 physically associates with Sec13, suggesting direct interaction between the autophagy machinery and ERES components[13]. Deletion of the trs85 gene, which encodes a TRAPPIII complex component, reduced the BiFC signal between Sec13 and Atg2, indicating that TRAPPIII contributes to organizing the ERES localization at phagophore-ER contact sites[13].
Temporal Dynamics of Atg2 Localization
Live-cell imaging studies in fission yeast revealed that newly formed PAS structures localize between the ER and the vacuole in most cases, with Atg2 appearing at the PAS immediately upon its formation[13]. This intimate spatial relationship between nascent phagophores and the ER suggests that the ER-phagophore contact site is not a late consequence of autophagosome formation but rather a primary determinant of autophagosome nucleation and early expansion[13]. At the PAS, Atg2-GFP forms punctate structures or small dots representing individual or clusters of autophagosome assembly sites[27].
Phosphoinositide-Dependent Recruitment
The recruitment of Atg2 to autophagosomal structures depends on the generation of PI3P by the phosphatidylinositol 3-kinase (PI3K) complexes localized at the PAS[1][33]. Treatment of cells with wortmannin, a pharmacological inhibitor of class III PI3K, prevented the recruitment of Atg2/ATG2A to forming autophagosomes, while the upstream Atg1/ULK complex could still form[7][33]. This hierarchical organization indicates that ATG2 recruitment occurs downstream of PI3K activation and PI3P generation on the nascent phagophore[7][33].
The PI3P effector WIPI proteins (particularly WIPI4 in mammalian cells) are crucial for targeting ATG2A to the phagophore[8]. Overexpression of inactive WIPI4 mutants unable to bind PI3P prevented normal ATG2A recruitment, while providing excess wild-type WIPI4 enhanced ATG2A localization to autophagosomal structures[8].
Atg2 in Autophagosome Formation and Expansion
Role in Phagophore Elongation and Growth
The primary function of Atg2 in autophagy is to facilitate the dramatic expansion of the nascent isolation membrane or phagophore from its initial nucleation as a small crescent or cup-like structure into a large double-membrane autophagosome capable of enclosing cytoplasmic cargo. During the expansion phase, the phagophore grows from a few hundred nanometers in diameter to eventually reach sizes of 500-1500 nanometers depending on the cargo being engulfed[1][7][8].
In atg2 deletion mutants of budding yeast, autophagosomes fail to expand properly and remain as small, immature structures, with cargo remaining protease-accessible rather than protected within sealed autophagosomes[1][45]. Similarly, in mammalian cells with simultaneous knockout of ATG2A and ATG2B, unclosed autophagic structures accumulate at the phagophore assembly sites, with the phagophore capable of forming but unable to expand and seal[33]. These observations directly implicate Atg2 in the critical expansion step rather than in the initial nucleation of the phagophore.
Lipid Supply from ER to Phagophore
The model emerging from multiple lines of evidence indicates that Atg2 mediates the physical supply of lipids from the ER to the growing phagophore membrane. In 2017, researchers demonstrated that the movement of lipophilic dyes from the ER to IM precursors was completely blocked by deletion of Atg2, even when other early autophagy factors were present[1][7]. This direct evidence for Atg2-mediated lipid movement between the ER and phagophore provided the crucial experimental support for the lipid transfer model.
The proposed mechanism involves Atg2-mediated extraction of lipids from the curved membranes of the ER-exit sites and subsequent delivery to the expanding edge of the phagophore where they serve as building blocks for membrane expansion[1][7][8]. Given that autophagosomes must grow by incorporating millions of lipid molecules within minutes, the high-capacity lipid binding of ATG2 (approximately twenty lipids per molecule) represents a significant advantage over single-lipid-transfer proteins in achieving the necessary flux[9].
Cooperation with Vps13 Proteins
Recent comprehensive studies revealed that the lipid transfer protein Vps13 cooperates with Atg2 at phagophore rims to ensure sufficient lipid delivery for autophagosome biogenesis[10]. Vps13 belongs to the same family of Chorein_N-domain proteins as Atg2 and localizes to the rims of approximately 90% of enlarged phagophores in wild-type cells[10]. Notably, Vps13 localization to forming autophagosomes does not require Atg2 or other core autophagy factors, suggesting parallel but independent recruitment of these two lipid transfer proteins[10].
The kinetic parameters revealed that Atg2-mediated phospholipid transfer (PLT) alone is not rate-limiting for autophagosome biogenesis[10]. Instead, the parallel action of Atg2 and Vps13 working at PERCS ensures sufficient lipid transfer rates to accommodate the rapid membrane expansion observed in living cells[10]. In the absence of Vps13, the number of Atg2 molecules at PERCS determines both the duration of autophagosome formation and the final size of forming autophagosomes, with an apparent in vivo transfer rate of approximately 200 phospholipids per Atg2 molecule per second[10].
Mechanisms of Autophagosome Closure
ATG2-GABARAP Interaction in Closure
Beyond its role in phagophore expansion through lipid transfer, Atg2 participates in a distinct process: the closure or sealing of the autophagosomal membrane[45][55]. The identification of a conserved LIR motif in ATG2A and ATG2B revealed direct interactions between ATG2 proteins and the Atg8/LC3/GABARAP family of autophagosomal proteins[55]. The ATG2A-GABARAP interaction appeared to function independently of the ATG2A-WIPI4 interaction required for lipid transfer[55].
In cells expressing ATG2A variants with mutations in the LIR motif (designated ATG2A-mLIR), the autophagy pathway exhibited severe defects in autophagosome closure despite retained ability to interact with WIPI4 and perform lipid transfer[55]. These mutant cells accumulated large numbers of unsealed, open autophagosomal structures containing Atg8/LC3 and other autophagy proteins but unable to form sealed compartments[55]. This separation-of-function analysis demonstrated that the ATG2-GABARAP interaction represents a mechanistically distinct closure pathway independent of lipid transfer activity[55].
Molecular Mechanism of Closure
The precise molecular mechanism by which the ATG2-GABARAP interaction promotes autophagosome closure remains under investigation, but several possibilities have been proposed[45][55]. One hypothesis suggests that multimerization of the LC3/GABARAP proteins on the phagophore membrane, mediated through their interactions with Atg2, generates sufficient mechanical force or stabilization to promote membrane fusion and sealing[45][55]. The LC3/GABARAP proteins possess the ability to interact with both lipids and proteins, including other LC3/GABARAP molecules, suggesting capacity for forming higher-order assemblies[55].
Another possibility involves the spatial organization of membrane scission machinery at the site of the Atg2-GABARAP interaction[45]. The ESCRT complexes and other scission factors known to be required for autophagosome closure might be brought into proximity through interactions with ATG2 and its associated proteins[45].
Biological Processes and Selective Autophagy
Role in Bulk, Non-Selective Autophagy
Schizosaccharomyces pombe Atg2 is required for starvation-induced bulk autophagy, the non-selective degradation of cytoplasmic components in response to nutrient starvation[3][6][25]. During nitrogen starvation, Atg2-GFP rapidly accumulates at autophagosomal structures, and deletion of atg2 severely impairs the autophagic response to nitrogen starvation[25].
Role in Selective Organelle Autophagy
Beyond bulk autophagy, Atg2 is particularly important for selective autophagy of organelles, especially the autophagy of endoplasmic reticulum (ER) and mitochondrial compartments (termed ER-autophagy and mitophagy, respectively)[3][6]. Studies in fission yeast demonstrated that during nitrogen starvation, both ER-GFP and mitochondrial markers (mito-mCherry) are delivered into vacuoles in an Atg5-dependent manner, and this organelle autophagy is severely compromised in atg20, atg24, and related mutants[3]. While not all selective autophagy pathways show equivalent dependence on Atg2, the protein appears critical for efficient selective degradation of large organellar structures.
Pexophagy and Peroxisome Degradation
In plant cells, Atg2 participates in pexophagy, the selective autophagy of peroxisomes[14][17]. Forward genetic screens in Arabidopsis thaliana identified atg2 mutations as suppressors of lon2 mutant phenotypes, with lon2 atg2 double mutants displaying rescued peroxisomal morphology and function[14]. The LON2 protease catalyzes matrix protein turnover within peroxisomes, and when LON2 is absent, peroxisomes accumulate abnormal proteins and are targeted for pexophagic degradation through Atg2-dependent mechanisms[14][17].
Evidence suggests that hydrogen peroxide and oxidative stress signals peroxisomal dysfunction and trigger selective autophagic degradation through Atg2-dependent mechanisms[17]. The selective autophagy of peroxisomes represents a quality control mechanism maintaining functional peroxisomal populations and preventing accumulation of organelles with compromised redox balance[14][17].
Evolutionary Conservation and Comparative Analysis
Conservation Across Eukaryotic Kingdoms
The structure and function of Atg2 have proven remarkably conserved across eukaryotic organisms from simple yeasts to plants and animals. The budding yeast Saccharomyces cerevisiae Atg2 shares fundamental structural organization with Schizosaccharomyces pombe Atg2, mammalian ATG2A and ATG2B, and plant ATG2 proteins[1][7][9]. The core lipid transfer domain (Chorein_N) is particularly well-conserved, with structural studies demonstrating essentially identical architecture in yeast and mammalian proteins[1][8][9].
Mammalian cells express two ATG2 paralogs, ATG2A and ATG2B, that arose from gene duplication and show approximately 44.5% amino acid sequence identity to each other but only 15-16% identity to the budding yeast protein[33]. Despite these sequence divergences, the proteins maintain functional redundancy, with both being capable of rescue of autophagy defects in cells lacking either protein[33]. The dual redundancy of mammalian ATG2 proteins suggests that the lipid transfer and membrane tethering functions are sufficiently important to warrant multiple genetic copies in higher organisms[33].
Chorein_N Domain Family
Atg2 belongs to a protein family united by their possession of the Chorein_N lipid transfer domain, which also includes the Vps13 family proteins (VPS13 in yeast and VPS13A, VPS13B, VPS13C, VPS13D in mammals)[12][31][34][56]. The N-terminal fragments of both Atg2 and Vps13 proteins sharing the Chorein_N domain show remarkable structural similarity despite low sequence conservation, with both folding into elongated molecules featuring large hydrophobic cavities[12][31][34][56]. The VPS13 proteins represent very large polypeptides (exceeding 3,000 amino acids in some cases) organized around repetitive β-groove (RBG) domains forming extended hydrophobic tunnels[56].
The shared architecture of the Chorein_N lipid transfer domain suggests that this represents an ancient lipid transfer module that was acquired early in eukaryotic evolution and subsequently deployed in multiple contexts involving membrane expansion and organellar biogenesis[12][31][34][56]. The discovery that both Atg2 and Vps13 function as high-capacity, bulk lipid transfer proteins distinguishes them from the previously characterized lipid transfer protein families, which typically transfer single lipids in a shuttle-like manner[12][31][34].
Structural Models of Lipid Transfer Mechanism
Bridge Model
The bridge model proposes that Atg2 stably tethers the phagophore and ER membranes through simultaneous binding of both membrane-binding tips, while lipids translocate through the internal cavity extending from one tip to the other[54]. In this model, lipids occupy the hydrophobic cavity and travel along the extended hydrophobic groove as if sliding through a tunnel, with their acyl chains remaining buried in the hydrophobic microenvironment while their polar head groups remain exposed to the cytosolic solution[1][50][54]. This mechanism would position Atg2 as a stable bridge mediating lipid transfer without requiring the protein itself to shuttle between membranes[54].
Ferry Model
An alternative ferry model suggests that the N-terminal lipid transfer domain of Atg2 can function independently as a lipid shuttle, with the N-terminal fragment cycling between the ER and phagophore while the C-terminal regions anchor the complex to the phagophore through WIPI interactions[54]. This model is supported by observations that N-terminal fragments of ATG2A comprising only amino acids 1-345 can mediate lipid transfer in vitro and fully rescue autophagy defects in cells[9][36]. In the ferry model, the anchored C-terminal regions hold the complex at the contact site while the N-terminal lipid transfer domain dynamically interacts with both membranes[54].
Recent evidence suggests that both models may operate: a stable bridge architecture for efficient bulk lipid transfer combined with dynamic shuttling of the N-terminal domain to extract lipids from the ER and insert them into the phagophore[54].
Hydrophobic Groove as Lipid Pathway
The structural evidence supports a model in which the continuous hydrophobic cavity or groove extending through the Atg2 rod acts as a dedicated pathway for lipid translocation[1][50]. The Chorein_N N-terminal domain, the five internal repeats, and the ATG2_CAD region collectively form this pathway, with lipid acyl chains sliding along the hydrophobic walls while the polar head groups remain solvated[1][50]. This contrasts with vesicular trafficking, where lipids would be packaged into membrane-enclosed compartments for transport, instead representing direct intermembrane transfer through a protein-mediated channel[1][50].
Recent Discoveries and Emerging Model of Autophagosome Biogenesis
Integration with Atg9 Scramblase Function
One of the most significant recent advances involved recognizing that Atg2 lipid transfer activity must be coupled with Atg9 lipid scramblase activity for effective autophagosome expansion[10][20][23][39]. While Atg2 can transfer lipids from the ER to the phagophore, it primarily inserts them into the outer leaflet of the phagophore membrane[20]. This would create severe lipid asymmetry incompatible with the bilayer geometry required for membrane expansion[20]. Atg9, which possesses lipid scramblase activity allowing energy-independent lipid flipping between membrane leaflets, solves this problem by equilibrating lipid distribution[20][23][39].
The physical interaction between Atg2 and Atg9 at the phagophore rim creates a minimal system for coordinated lipid transfer and scrambling[10][20][23][39]. Cryo-EM structural models suggest that the opening of Atg2's hydrophobic groove aligns with the internal channel of the Atg9 trimeric complex, allowing direct hand-off of lipids[39]. This coupling ensures that as Atg2 delivers lipids to the outer leaflet, Atg9 simultaneously equilibrates them between leaflets, preventing the osmotic stress and membrane disruption that would result from excessive outer leaflet expansion[20][39].
TRAPPIII Complex Coordination
Recent studies in yeast revealed that the TRAPPIII tethering complex participates in establishing and organizing the phagophore-ER membrane contact sites where Atg2 operates[13]. TRAPPIII is a guanine nucleotide exchange factor (GEF) that activates the small GTPase Ypt1, and its recruitment to PERCS by interaction with Atg9 and other factors organizes the tethering machinery[13]. The interaction between TRAPPIII and Atg2 appears cooperative, with both complexes working to maintain the appropriate spacing and organization of phagophore-ER contact sites[13].
Non-Rate-Limiting Nature of Atg2 Activity
A surprising finding emerging from quantitative live-cell imaging studies is that Atg2-mediated lipid transfer is not rate-limiting for autophagosome biogenesis in wild-type cells[10]. Instead, the parallel action of multiple lipid transfer proteins including Vps13 ensures that sufficient phospholipid delivery occurs to support the observed rates of autophagosome expansion[10]. The number of Atg2 molecules at individual phagophores correlates with the final size of the forming autophagosome, suggesting that Atg2 abundance becomes rate-limiting only in specialized circumstances or in the absence of other lipid transfer proteins[10].
Biological Significance and Disease Implications
Essential Role in Cellular Homeostasis
The essential requirement for Atg2 in autophagosome formation makes this protein critical for cellular homeostasis. Autophagy participates in degradation of damaged organelles, misfolded proteins, and pathogens, processes necessary for cellular health and survival during nutrient stress[1][7][8]. The inability to complete autophagosome formation in atg2 mutants results in accumulation of autophagic cargo and defective clearance of damaged cellular components[1][33].
Senescence and Cell Death
In plant cells, disruption of Atg2 function through mutagenesis results in accelerated senescence, exhibiting phenotypes characteristic of premature aging including accumulation of cellular damage and oxidative stress[44]. The silencing of GmATG2 in soybean results in high levels of ubiquitinated protein accumulation and enhanced accumulation of polyubiquitinated protein aggregates, indicating that impaired autophagy leads to proteostasis failure[44]. These findings suggest that maintaining functional Atg2-mediated autophagy is essential for long-term cellular viability and the prevention of age-associated pathologies.
Pathogenic Potential and Cancer Biology
Recent investigations have noted that ATG9B, a less well-characterized mammalian homolog that can functionally cooperate with ATG2A, has been implicated in tumorigenesis, particularly in hepatocellular carcinoma development[23]. While detailed mechanisms remain to be elucidated, the connection between autophagy dysfunction and cancer suggests that alterations in Atg2/ATG2 functions could contribute to malignant transformation[23].
Comparative Analysis with Budding Yeast and Mammals
Fission Yeast Specific Features
Schizosaccharomyces pombe Atg2 exhibits several features that distinguish it from better-characterized mammalian and budding yeast proteins. The fission yeast autophagy machinery shows important differences in the composition and organization of the Atg1 complex, with fission yeast requiring Atg11 for starvation-induced bulk autophagy, whereas budding yeast Atg11 is dispensable for this process[25]. These organismal differences suggest that the role and regulation of Atg2 may be somewhat specialized in fission yeast, though the core lipid transfer function is conserved.
Mammalian Complexity and Redundancy
Mammals express both ATG2A and ATG2B, with these proteins showing redundant but also potentially specialized functions[33][39]. Both proteins can independently support autophagy, but combined deletion is required to completely block autophagy, indicating functional overlap[33]. The existence of two ATG2 paralogs in mammals potentially allows tissue-specific or temporally-regulated autophagy responses through selective expression of each isoform[33][39].
Remaining Questions and Future Research Directions
Open Questions Regarding Lipid Transfer Kinetics
Despite recent advances, several fundamental questions remain regarding the quantitative aspects of Atg2-mediated lipid transfer in living cells[54]. While in vitro lipid transfer rates have been measured, integrating these values with the known timecourse of autophagosome expansion and the total lipid content suggests that either additional mechanisms accelerate transfer, other lipid transfer proteins make essential contributions (as demonstrated for Vps13), or the in vitro assay conditions differ significantly from the cellular environment in ways that affect transfer rates[10][54].
Mechanistic Basis of Lipid Selectivity
The substrate selectivity of Atg2 and whether different lipid species are transferred at differential rates remains an area for investigation[54]. While bulk lipid transfer is a primary function of the Chorein_N proteins, whether specific lipid compositions or species are preferentially transferred to support particular aspects of autophagosome formation (such as maintaining specific phosphoinositide distributions or enriching particular lipid species) requires further study[54].
Structural Details of Full-Length Protein
Although the N-terminal lipid transfer domain and other regions of Atg2 have been structurally characterized, the complete three-dimensional structure of full-length Atg2 remains to be determined at atomic resolution[1][7][50]. The organization and connectivity of the five internal repeat domains, their precise role in forming the continuous hydrophobic cavity, and how these regions interact with binding partners remain subjects for future structural investigations[1][7].
Conclusion
Autophagy-related protein 2 (Atg2) in Schizosaccharomyces pombe represents a remarkable example of how molecular characterization of conserved cellular machinery continues to reveal previously unsuspected mechanisms of cellular organization and membrane biology[1][7]. What was once recognized simply as an essential but enigmatic protein required for autophagosome formation has emerged as a sophisticated, multifunctional molecular device mediating both stable membrane tethering and high-capacity lipid transfer between organellar compartments[1][7][8]. The discovery that Atg2 functions as a lipid transfer protein has revolutionized our understanding of de novo autophagosome formation, providing a molecular mechanism for how cells can rapidly construct new membrane compartments through direct intermembrane lipid transport rather than relying exclusively on vesicular trafficking[1][7][8][9].
The structure of the rod-shaped Atg2 molecule with its two membrane-binding tips and extensive hydrophobic cavity has proven ideally suited to serve as a bridge simultaneously tethering the ER and phagophore while channeling lipids between these membrane systems[1][7][8]. The biochemical capacity of individual Atg2 molecules to bind and transfer approximately twenty lipid molecules at a time, combined with the parallel action of related lipid transfer proteins such as Vps13, provides sufficient phospholipid flux to support the documented rates of autophagosome expansion observed in living cells[9][10].
The recognition that Atg2 participates in both phagophore expansion through lipid transfer and in autophagosome closure through distinct interactions with GABARAP proteins has unveiled the sequential, mechanistically distinct steps of autophagosome biogenesis[45][55]. The physical coupling of Atg2 lipid transfer activity with Atg9 lipid scramblase function has provided insights into how cells manage the complex challenge of generating the vast asymmetric lipid bilayers characteristic of autophagosomes[10][20][23][39].
The evolutionary conservation of Atg2 function across eukaryotes from simple yeasts to plants and animals, combined with the shared architecture of the Chorein_N lipid transfer domain found in Vps13 and other proteins, suggests that this represents a fundamental mechanism of eukaryotic membrane biology. The participation of Atg2 in not only bulk autophagy but also selective autophagy of organelles such as mitochondria, ER, and peroxisomes indicates that this protein's lipid transfer function addresses a general cellular requirement for rapid membrane expansion during diverse biological processes[1][3][6][14][17].
Future investigations into the atomic-resolution structure of full-length Atg2, the molecular details of lipid selectivity, and the quantitative kinetics of lipid transfer in cellular contexts promise to further illuminate this essential aspect of eukaryotic cell biology. The continued characterization of Atg2 and related lipid transfer proteins contributes not only to fundamental understanding of autophagy and cellular homeostasis but also holds potential for therapeutic applications targeting autophagy dysregulation in diseases ranging from neurodegeneration to cancer.
Citations
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6511744/
- https://www.uniprot.org/citations/16303567
- https://journals.biologists.com/jcs/article/129/22/4289/55695/Atg20-and-Atg24-family-proteins-promote-organelle
- https://www.uniprot.org/uniprotkb/O94649/entry
- https://string-db.org/network/284812.Q96VG1
- https://pubmed.ncbi.nlm.nih.gov/27737912/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6625793/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6548141/
- https://rupress.org/jcb/article/222/7/e202211039/214069/Parallel-phospholipid-transfer-by-Vps13-and-Atg2
- https://www.pnas.org/doi/10.1073/pnas.1811874115
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7885529/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC12618252/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3877801/
- https://www.molbiolcell.org/doi/10.1091/mbc.E25-06-0273
- https://journals.sagepub.com/doi/10.1177/25152564231183898
- https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2014.00139/full
- https://pubmed.ncbi.nlm.nih.gov/41563375/?fc=None&ff=20260121191715&v=2.18.0.post22+67771e2
- https://pubmed.ncbi.nlm.nih.gov/19200887/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7718406/
- https://www.pombase.org/gene/SPBC31E1.01c
- http://reactome.org/content/detail/R-HSA-5676229
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10936676/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8997447/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11073433/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5712628/
- https://www.uniprot.org/uniprotkb/O42651/entry
- https://www.semanticscholar.org/paper/The-crystal-structure-of-Atg18-reveals-a-new-site-Lei-Tang/d5711eb98b0310b56874e3719861d74040718c1b
- https://febs.onlinelibrary.wiley.com/doi/10.1002/1873-3468.14741
- https://pubmed.ncbi.nlm.nih.gov/34783437/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1805099/
- https://www.molbiolcell.org/doi/abs/10.1091/mbc.e11-09-0785
- https://febs.onlinelibrary.wiley.com/doi/10.1111/febs.16280
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4590589/
- https://rupress.org/jcb/article/218/6/1787/61839/ATG2-transports-lipids-to-promote-autophagosome
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3442503/
- https://pubmed.ncbi.nlm.nih.gov/36347259/
- https://elifesciences.org/articles/45777
- https://www.yeastgenome.org/locus/S000005186
- https://pdbj.org/emnavi/quick.php?id=emdb-15604
- https://pombase.org/gene/SPBC31E1.01c
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8584260/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8616111/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3362037/
- https://febs.onlinelibrary.wiley.com/doi/10.1002/1873-3468.14720
- https://pubmed.ncbi.nlm.nih.gov/29113029/
- https://febs.onlinelibrary.wiley.com/doi/10.1002/1873-3468.12901
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6844533/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7054675/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10683392/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7681076/
- https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2020.00565/full
- https://journals.sagepub.com/doi/abs/10.1177/25152564231211976
- https://onlinelibrary.wiley.com/doi/10.1111/cas.15112