Deep Research
Falcon
(snx41-deep-research-falcon.md)
Falcon
(snx41-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: snx41 / Sorting nexin-41 (UniProt O60107) in Schizosaccharomyces pombe (strain 972)
0) Identity verification (critical disambiguation)
The research target is the S. pombe gene snx41 (synonyms mug186, ORF SPBC14F5.11c) encoding Sorting nexin-41, annotated as a member of the sorting nexin (SNX) family with PX and BAR-superfamily domains (UniProt description provided by the user). In the literature retrieved here, “Snx41” is consistently used for a fungal SNX-BAR/PX-BAR sorting nexin that forms heterodimeric complexes with Snx4-family partners; this domain/family assignment matches the UniProt domain expectations (PX for PI-binding; BAR for membrane curvature scaffolding). (hanley2020sortingnexinsin pages 2-5, goyal2021vps501anovel pages 21-27)
A key limitation of the currently retrievable corpus is that most mechanistic/quantitative primary evidence explicitly concerns Saccharomyces cerevisiae Snx41 and the conserved Snx4-family SNX-BAR system, with only limited direct statements about S. pombe Snx41. Therefore, organism-specific claims about S. pombe Snx41 are restricted to (i) what is directly stated for S. pombe in the retrieved texts and (ii) conservative functional inference from highly conserved SNX-BAR mechanisms in fungi. (jr2017autophagicclearanceof pages 8-9, hanley2020sortingnexinsin pages 2-5)
1) Key concepts and current understanding
1.1 Sorting nexins, PX domains, and SNX-BAR architecture
Sorting nexins are a conserved family of membrane-associated trafficking proteins defined by a PX (Phox homology) domain that binds phosphoinositides (commonly PI3P-enriched endosomal membranes). SNX-BAR subfamily members contain a BAR domain (often with coiled-coils) that promotes dimerization and drives/senses membrane curvature, enabling membrane tubulation/scaffolding and protein–protein interactions. (hanley2020sortingnexinsin pages 2-5, goyal2021vps501anovel pages 21-27)
This architecture aligns with the user-provided UniProt domain annotations for S. pombe Snx41 (PX + BAR-superfamily). (hanley2020sortingnexinsin pages 2-5)
1.2 The Snx4-family SNX-BAR system in fungi (context for Snx41)
In yeast systems, Snx41 is described as part of a Snx4-family group (Snx4, Snx41, and Atg20/Snx42), where Snx4 forms alternative heterodimers (e.g., Snx4–Snx41 or Snx4–Atg20/Snx42) associated with distinct trafficking/autophagy roles. (goyal2021vps501anovel pages 21-27, jr2017autophagicclearanceof pages 8-9)
This places Snx41 at the intersection of endosomal recycling/retrograde trafficking and autophagy-linked membrane remodeling, which is consistent with SNX-BAR biology more broadly. (hanley2020sortingnexinsin pages 20-21, hanley2020sortingnexinsin pages 2-5)
2) Recent developments (prioritizing 2023–2024)
2.1 2023 advance: a SNX-BAR complex controls autophagosome “mouth” size and non-selective capture
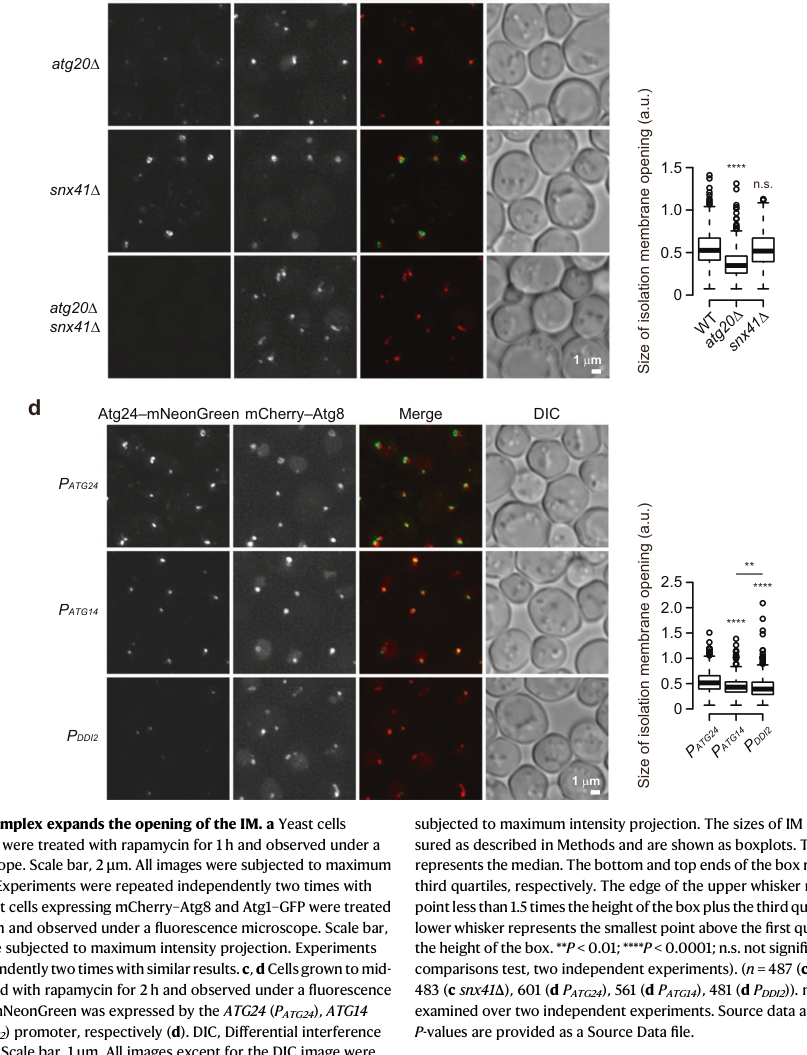
A major 2023 development is the discovery that a sorting nexin complex localizes to the opening edge of the isolation membrane (IM) (phagophore) and is critical for ensuring non-selective sequestration of cytoplasm during autophagosome biogenesis. In this work, the complex components are described as PX-BAR sorting nexins; Snx41 is specifically mentioned as a sorting nexin that can form a complex with Atg24 competitively with Atg20. (Kotani et al., Nature Communications, accepted 7 Sep 2023; published Sep 2023; https://doi.org/10.1038/s41467-023-41525-x). (kotani2023amechanismthat pages 1-2)
Mechanistically, the study proposes that the BAR-domain-containing sorting nexin complex stabilizes the highly curved IM edge, allowing the IM to expand with a large opening; without the complex, the IM opening becomes too small to allow entry of large assemblies. (kotani2023amechanismthat pages 1-2)
2.2 Quantitative size-selectivity: ~25 nm effective cutoff without the complex
Kotani et al. report that without the complex, the isolation membrane expands with a small opening that prevents entry of particles larger than ~25 nm, explicitly including ribosomes and proteasomes, even though autophagosomes of nearly normal size eventually form. (kotani2023amechanismthat pages 1-2)
The same paper supports size-threshold reasoning using engineered oligomeric particle reporters, estimating particle sizes around ~20 nm (Dps–GFP) and ~25 nm (RibH–GFP) to probe the size-dependence of autophagic capture. (kotani2023amechanismthat pages 2-3)
2.3 Localized action at the IM opening edge (visualized)
The localization of the sorting nexin complex to the IM opening edge is visualized as ring-like structures at the cup-shaped IM. This is central evidence that Snx41-family SNX-BARs can act at a specific autophagosome biogenesis subdomain rather than broadly across the IM surface. (kotani2023amechanismthat pages 1-2, kotani2023amechanismthat media 68a2f9a9)
2.4 2024 literature availability in this run
A 2024 review reference related to autophagy mechanisms was detected but was not obtainable within the current tool environment, limiting direct 2024 primary/review coverage in this report. (jr2017autophagicclearanceof pages 8-9)
3) Molecular function, pathways, and localization of Snx41 (what can be stated with evidence)
3.1 Molecular function (most supported): membrane-binding and membrane-shaping scaffold within a SNX-BAR complex
Based on conserved SNX-BAR principles, Snx41’s PX domain supports recruitment to PI3P-enriched membranes, while the BAR domain supports dimeric scaffolding/curvature stabilization—functions that are required to shape or stabilize membrane subdomains in trafficking and autophagy. (hanley2020sortingnexinsin pages 2-5, goyal2021vps501anovel pages 21-27)
In the 2023 study, Snx41 is explicitly described as a sorting nexin that can form a complex with Atg24 competitively with Atg20, and is observed at the IM opening edge, consistent with a role as a structural membrane adaptor/scaffold rather than an enzyme with a catalytic substrate. (kotani2023amechanismthat pages 1-2)
3.2 Pathway placement: autophagy (macroautophagy) and coupling to membrane morphogenesis
The 2023 study directly places the Atg24–Atg20/Snx41 sorting nexin system in autophagosome formation, acting at the opening edge of the IM to permit entry of large cytoplasmic structures in non-selective autophagy. (kotani2023amechanismthat pages 1-2, kotani2023amechanismthat pages 3-4)
Although these data are in budding yeast, the study explicitly frames sorting nexins (including this complex) as conserved machinery and highlights the broader possibility that related sorting nexins could play similar roles in other eukaryotes. (kotani2023amechanismthat pages 5-6)
3.3 Subcellular localization: endosome-associated puncta and IM-edge localization in autophagy
At the family level, SNX-BAR proteins are endosome-associated due to PI-binding and are often seen as punctate cytosolic/endosomal structures; in autophagy, the Atg24/Atg20/Snx41 system can relocalize to autophagosome formation sites and, specifically, the IM opening edge in the 2023 work. (hanley2020sortingnexinsin pages 2-5, kotani2023amechanismthat pages 1-2)
Image-based evidence (cropped figure regions retrieved) supports quantitative IM-opening measurements and IM-edge localization behavior in strains including snx41Δ, visually grounding the claim that perturbing Snx41-family components affects IM opening morphology/size metrics. (kotani2023amechanismthat media 68a2f9a9, kotani2023amechanismthat media 9f40f4b1, kotani2023amechanismthat media 9d2722bf)
4) Genetics/phenotypes and statistics (recent quantitative data)
4.1 IM opening-size measurements with sample sizes
Kotani et al. quantified IM opening-related size metrics using fluorescence microscopy and reported large sample sizes across genotypes, including n = 487 (WT), 483 (atg20Δ), and 483 (snx41Δ) IMs measured over two independent experiments (as indicated in the figure caption text captured in the retrieved material). (kotani2023amechanismthat pages 5-6, kotani2023amechanismthat media 68a2f9a9)
4.2 Viability under nitrogen starvation
Kotani et al. report that ATG24 knockout and double knockout of ATG20 and SNX41 “largely promoted cell death” under nitrogen starvation, whereas atg20Δ showed milder viability defects and snx41Δ showed little/no defect, consistent with partial redundancy of Atg20 and Snx41 in the functional complex. (kotani2023amechanismthat pages 5-6)
4.3 Electron microscopy evidence of failed ribosome capture
Electron microscopy in Kotani et al. indicates that in atg24Δ cells, ribosomes are present in the cytoplasm but autophagic bodies are strikingly absent from ribosomes, while wild-type autophagic bodies contain ribosomes at similar density to the cytoplasm—supporting a model of defective large-particle sequestration rather than a complete block in autophagosome formation. (kotani2023amechanismthat pages 1-2)
5) Expert opinions and authoritative synthesis
A 2020 expert review emphasizes that sorting nexins are a conserved thread connecting endosomal trafficking, autophagy, and protein homeostasis, and highlights the role of SNX-BAR complexes in these interconnected pathways. This review-level synthesis supports interpreting Snx41 primarily as a membrane adaptor that couples phosphoinositide binding (PX) with membrane remodeling (BAR) to influence trafficking/autophagy outcomes. (Hanley & Cooper, Cells, published Nov 2020; https://doi.org/10.3390/cells10010017). (hanley2020sortingnexinsin pages 2-5, hanley2020sortingnexinsin pages 20-21)
6) Current applications and real-world implementations
6.1 Functional annotation and pathway modeling in fungi
For S. pombe gene annotation efforts, Snx41’s value is as a mechanistic node linking PI3P-positive membranes to membrane curvature remodeling in pathways including endosomal recycling and autophagy-related membrane morphogenesis. The 2023 IM-edge mechanism provides a concrete, testable functional hypothesis (IM edge stabilization / opening-size regulation) that can be probed experimentally in other fungi, including S. pombe, using analogous fluorescent IM markers and cargo-size reporters. (kotani2023amechanismthat pages 1-2, kotani2023amechanismthat pages 3-4)
6.2 Imaging and quantitative morphometrics workflows
Kotani et al. explicitly rely on quantitative fluorescence imaging and IM-opening measurements with large n, representing a practical implementation of “autophagosome morphometrics” that can be ported to functional annotation pipelines (e.g., screening snx41 mutants for IM opening size and capture efficiency of large particles). (kotani2023amechanismthat pages 5-6, kotani2023amechanismthat media 68a2f9a9)
7) Evidence map (summary table)
The following table compiles the most directly supported claims and quantitative details relevant to Snx41-family SNX-BAR function (including Snx41) and highlights where evidence is strongest.
| Claim/finding | Evidence type (genetics, microscopy, EM, assay) | Key quantitative detail | Organism | Source (with year and URL) |
|---|---|---|---|---|
| snx41 corresponds to a fission-yeast sorting nexin family member with PX and BAR domains, matching the UniProt O60107 annotation and supporting classification as a PX-BAR sorting nexin | Domain/bioinformatic background; review synthesis | PX domains are generally ~100–130 aa and bind phosphoinositides; BAR domains mediate dimerization/membrane curvature sensing and remodeling | Schizosaccharomyces pombe (target) with family-level conservation across yeasts | Hanley & Cooper, 2020, Cells, https://doi.org/10.3390/cells10010017; Popelka et al., 2017, PNAS, https://doi.org/10.1073/pnas.1708367114 (hanley2020sortingnexinsin pages 2-5, popelka2017structureandfunction pages 1-2, popelka2017structureandfunction pages 2-3) |
| In yeast SNX-BAR systems, Snx41 belongs to the Snx4-family of sorting nexins that form distinct heterodimeric complexes with cargo/pathway specificity | Review/background from primary literature synthesis | Snx4 family comprises Snx4, Atg20/Snx42, and Snx41; distinct dimers have overlapping but nonidentical roles | Yeast (family-level, including fungi) | Hanley & Cooper, 2020, Cells, https://doi.org/10.3390/cells10010017 (hanley2020sortingnexinsin pages 20-21, goyal2021vps501anovel pages 21-27, jr2017autophagicclearanceof pages 8-9) |
| In the autophagy-edge model, Snx41 forms a complex with Atg24 competitively with Atg20, and this Atg24–Atg20/Snx41 complex localizes to the opening edge of the isolation membrane (IM) | Live-cell fluorescence microscopy | Snx41 localization to the IM opening edge is reported; Atg24/Atg20 show ring-like signal at the edge of cup-shaped IMs | Budding yeast (Saccharomyces cerevisiae) | Kotani et al., 2023, Nature Communications, https://doi.org/10.1038/s41467-023-41525-x (kotani2023amechanismthat pages 1-2) |
| The Atg24–Atg20/Snx41 sorting nexin complex is required to keep the IM opening sufficiently large for non-selective autophagic sequestration of large particles | Genetics, microscopy, EM, autophagy cargo assays | Without the complex, IMs expand with a small opening that excludes particles larger than about 25 nm, including ribosomes and proteasomes | Budding yeast (Saccharomyces cerevisiae) | Kotani et al., 2023, Nature Communications, https://doi.org/10.1038/s41467-023-41525-x (kotani2023amechanismthat pages 1-2, kotani2023amechanismthat pages 3-4) |
| snx41Δ alone causes milder defects than loss of ATG24 or atg20Δ snx41Δ double deletion, indicating partial redundancy with Atg20 in the Atg24 complex | Genetics; viability assays; microscopy-based IM measurements | ATG24 knockout and atg20Δ snx41Δ strongly reduce viability under nitrogen starvation; atg20Δ is milder and snx41Δ shows no or minimal viability defect alone | Budding yeast (Saccharomyces cerevisiae) | Kotani et al., 2023, Nature Communications, https://doi.org/10.1038/s41467-023-41525-x (kotani2023amechanismthat pages 5-6) |
| The Atg24 complex is specifically important for degradation of large cytoplasmic assemblies, whereas small soluble markers can still be degraded without it | Autophagy assays; immunoblot readouts | Small markers such as Pgk1–GFP and ALP remain degradable; larger assemblies around ~20–25 nm or more become increasingly dependent on the complex | Budding yeast (Saccharomyces cerevisiae) | Kotani et al., 2023, Nature Communications, https://doi.org/10.1038/s41467-023-41525-x (kotani2023amechanismthat pages 1-2, kotani2023amechanismthat pages 2-3) |
| Particle-size calibration experiments support a size threshold for Atg24-complex-dependent autophagy | Assay; particle engineering; immunoblot | Bacterial oligomer reporters estimated at ~20 nm (Dps–GFP) and ~25 nm (RibH–GFP) were used to define the entry threshold | Budding yeast (Saccharomyces cerevisiae) | Kotani et al., 2023, Nature Communications, https://doi.org/10.1038/s41467-023-41525-x (kotani2023amechanismthat pages 3-4, kotani2023amechanismthat pages 2-3) |
| Electron microscopy shows that in the absence of the Atg24 complex, autophagic bodies can form but are depleted of ribosomes, demonstrating failed capture of large complexes rather than complete autophagy arrest | Electron microscopy | In atg24Δ cells, autophagic bodies were present but were described as strikingly absent from ribosomes, unlike wild type | Budding yeast (Saccharomyces cerevisiae) | Kotani et al., 2023, Nature Communications, https://doi.org/10.1038/s41467-023-41525-x (kotani2023amechanismthat pages 1-2) |
| IM opening measurements directly show that snx41Δ alters opening-size behavior, although less strongly than more severe mutant combinations | Fluorescence microscopy; quantitative image analysis | Figure-associated boxplots report n = 487 WT, 483 atg20Δ, 483 snx41Δ IMs measured across two independent experiments | Budding yeast (Saccharomyces cerevisiae) | Kotani et al., 2023, Nature Communications, https://doi.org/10.1038/s41467-023-41525-x (kotani2023amechanismthat pages 5-6, kotani2023amechanismthat media 68a2f9a9) |
| Expert synthesis places Snx41-family proteins at the intersection of endosomal retrograde trafficking and autophagy/protein homeostasis, explaining why direct biochemical data for fission-yeast Snx41 remain limited but family-based functional inference is strong | Expert review/analysis | Review notes cytosolic Snx4-Atg20 and Snx4-Snx41 heterodimers relocalize during stress and participate in protein-homeostasis pathways | Yeast and broader eukaryotic sorting nexin field | Hanley & Cooper, 2020, Cells, https://doi.org/10.3390/cells10010017 (hanley2020sortingnexinsin pages 20-21, hanley2020sortingnexinsin pages 2-5) |
Table: This table summarizes evidence-supported findings relevant to Schizosaccharomyces pombe Snx41 and the Atg24–Atg20/Snx41 PX-BAR sorting nexin complex, emphasizing localization, autophagy function, and quantitative phenotypes. It is useful as a compact evidence map separating direct observations from family-level inference.
8) Conclusions specific to S. pombe Snx41 (O60107)
Schizosaccharomyces pombe Snx41 (O60107) is best annotated, based on domain/family alignment and conserved yeast SNX-BAR biology, as a PX-BAR sorting nexin functioning as a membrane adaptor/scaffold rather than an enzyme. Its most strongly evidenced mechanistic role (from recent high-quality primary literature in yeast SNX-BAR systems) is participation in a sorting-nexin complex that can act at autophagy membranes to control membrane edge stability/opening size and thereby influence size-dependent non-selective autophagic capture of large cytoplasmic assemblies. (hanley2020sortingnexinsin pages 2-5, kotani2023amechanismthat pages 1-2, kotani2023amechanismthat pages 3-4)
9) Key references (with dates and URLs)
- Kotani T. et al. “A mechanism that ensures non-selective cytoplasm degradation by autophagy.” Nature Communications (published Sep 2023; accepted 7 Sep 2023). https://doi.org/10.1038/s41467-023-41525-x (kotani2023amechanismthat pages 1-2)
- Hanley S.E., Cooper K.F. “Sorting Nexins in Protein Homeostasis.” Cells (published Nov 2020). https://doi.org/10.3390/cells10010017 (hanley2020sortingnexinsin pages 2-5, hanley2020sortingnexinsin pages 20-21)
- Popelka H. et al. “Structure and function of yeast Atg20, a sorting nexin that facilitates autophagy induction.” PNAS (published Nov 2017). https://doi.org/10.1073/pnas.1708367114 (popelka2017structureandfunction pages 1-2, popelka2017structureandfunction pages 2-3)
References
-
(hanley2020sortingnexinsin pages 2-5): Sarah E Hanley and Katrina F Cooper. Sorting nexins in protein homeostasis. Cells, Nov 2020. URL: https://doi.org/10.3390/cells10010017, doi:10.3390/cells10010017. This article has 88 citations.
-
(goyal2021vps501anovel pages 21-27): S Goyal. Vps501, a novel snx-bar protein involved in autophagy. Unknown journal, 2021.
-
(jr2017autophagicclearanceof pages 8-9): RJ Tomko Jr. Autophagic clearance of proteasomes in yeast requires the conserved sorting nexin snx4 antonia a. nemec, lauren a. howell, anna k. peterson, matthew a …. Unknown journal, 2017.
-
(hanley2020sortingnexinsin pages 20-21): Sarah E Hanley and Katrina F Cooper. Sorting nexins in protein homeostasis. Cells, Nov 2020. URL: https://doi.org/10.3390/cells10010017, doi:10.3390/cells10010017. This article has 88 citations.
-
(kotani2023amechanismthat pages 1-2): Tetsuya Kotani, Yuji Sakai, Hiromi Kirisako, Chika Kakuta, Soichiro Kakuta, Yoshinori Ohsumi, and Hitoshi Nakatogawa. A mechanism that ensures non-selective cytoplasm degradation by autophagy. Nature Communications, Sep 2023. URL: https://doi.org/10.1038/s41467-023-41525-x, doi:10.1038/s41467-023-41525-x. This article has 30 citations and is from a highest quality peer-reviewed journal.
-
(kotani2023amechanismthat pages 2-3): Tetsuya Kotani, Yuji Sakai, Hiromi Kirisako, Chika Kakuta, Soichiro Kakuta, Yoshinori Ohsumi, and Hitoshi Nakatogawa. A mechanism that ensures non-selective cytoplasm degradation by autophagy. Nature Communications, Sep 2023. URL: https://doi.org/10.1038/s41467-023-41525-x, doi:10.1038/s41467-023-41525-x. This article has 30 citations and is from a highest quality peer-reviewed journal.
-
(kotani2023amechanismthat media 68a2f9a9): Tetsuya Kotani, Yuji Sakai, Hiromi Kirisako, Chika Kakuta, Soichiro Kakuta, Yoshinori Ohsumi, and Hitoshi Nakatogawa. A mechanism that ensures non-selective cytoplasm degradation by autophagy. Nature Communications, Sep 2023. URL: https://doi.org/10.1038/s41467-023-41525-x, doi:10.1038/s41467-023-41525-x. This article has 30 citations and is from a highest quality peer-reviewed journal.
-
(kotani2023amechanismthat pages 3-4): Tetsuya Kotani, Yuji Sakai, Hiromi Kirisako, Chika Kakuta, Soichiro Kakuta, Yoshinori Ohsumi, and Hitoshi Nakatogawa. A mechanism that ensures non-selective cytoplasm degradation by autophagy. Nature Communications, Sep 2023. URL: https://doi.org/10.1038/s41467-023-41525-x, doi:10.1038/s41467-023-41525-x. This article has 30 citations and is from a highest quality peer-reviewed journal.
-
(kotani2023amechanismthat pages 5-6): Tetsuya Kotani, Yuji Sakai, Hiromi Kirisako, Chika Kakuta, Soichiro Kakuta, Yoshinori Ohsumi, and Hitoshi Nakatogawa. A mechanism that ensures non-selective cytoplasm degradation by autophagy. Nature Communications, Sep 2023. URL: https://doi.org/10.1038/s41467-023-41525-x, doi:10.1038/s41467-023-41525-x. This article has 30 citations and is from a highest quality peer-reviewed journal.
-
(kotani2023amechanismthat media 9f40f4b1): Tetsuya Kotani, Yuji Sakai, Hiromi Kirisako, Chika Kakuta, Soichiro Kakuta, Yoshinori Ohsumi, and Hitoshi Nakatogawa. A mechanism that ensures non-selective cytoplasm degradation by autophagy. Nature Communications, Sep 2023. URL: https://doi.org/10.1038/s41467-023-41525-x, doi:10.1038/s41467-023-41525-x. This article has 30 citations and is from a highest quality peer-reviewed journal.
-
(kotani2023amechanismthat media 9d2722bf): Tetsuya Kotani, Yuji Sakai, Hiromi Kirisako, Chika Kakuta, Soichiro Kakuta, Yoshinori Ohsumi, and Hitoshi Nakatogawa. A mechanism that ensures non-selective cytoplasm degradation by autophagy. Nature Communications, Sep 2023. URL: https://doi.org/10.1038/s41467-023-41525-x, doi:10.1038/s41467-023-41525-x. This article has 30 citations and is from a highest quality peer-reviewed journal.
-
(popelka2017structureandfunction pages 1-2): Hana Popelka, Alejandro Damasio, Jenny E. Hinshaw, Daniel J. Klionsky, and Michael J. Ragusa. Structure and function of yeast atg20, a sorting nexin that facilitates autophagy induction. Proceedings of the National Academy of Sciences, 114:E10112-E10121, Nov 2017. URL: https://doi.org/10.1073/pnas.1708367114, doi:10.1073/pnas.1708367114. This article has 52 citations and is from a highest quality peer-reviewed journal.
-
(popelka2017structureandfunction pages 2-3): Hana Popelka, Alejandro Damasio, Jenny E. Hinshaw, Daniel J. Klionsky, and Michael J. Ragusa. Structure and function of yeast atg20, a sorting nexin that facilitates autophagy induction. Proceedings of the National Academy of Sciences, 114:E10112-E10121, Nov 2017. URL: https://doi.org/10.1073/pnas.1708367114, doi:10.1073/pnas.1708367114. This article has 52 citations and is from a highest quality peer-reviewed journal.
Artifacts
Citations
- hanley2020sortingnexinsin pages 2-5
- kotani2023amechanismthat pages 1-2
- kotani2023amechanismthat pages 2-3
- jr2017autophagicclearanceof pages 8-9
- kotani2023amechanismthat pages 5-6
- hanley2020sortingnexinsin pages 20-21
- kotani2023amechanismthat pages 3-4
- popelka2017structureandfunction pages 1-2
- popelka2017structureandfunction pages 2-3
- https://doi.org/10.1038/s41467-023-41525-x
- https://doi.org/10.3390/cells10010017
- https://doi.org/10.3390/cells10010017;
- https://doi.org/10.1073/pnas.1708367114
- https://doi.org/10.3390/cells10010017,
- https://doi.org/10.1038/s41467-023-41525-x,
- https://doi.org/10.1073/pnas.1708367114,
Perplexity
(snx41-deep-research-perplexity.md)
Perplexity
(snx41-deep-research-perplexity.md)Sorting Nexin-41 (SNX41) in Schizosaccharomyces pombe: Structure, Function, and Role in Cellular Trafficking
Sorting nexin-41 (SNX41), encoded by the snx41 gene (also known as mug186) in the fission yeast Schizosaccharomyces pombe, represents a critical component of the endosomal trafficking machinery that mediates retrograde transport pathways and selective autophagy processes. This protein belongs to the highly conserved sorting nexin family, characterized by the presence of a phosphoinositide 3-phosphate (PI3P)-binding phox homology (PX) domain and a membrane-bending Bin-Amphiphysin-Rvs (BAR) domain, which enable it to engage with biological membranes and facilitate cargo sorting and transport[8][11][37]. SNX41 functions as a core component of heterodimeric complexes, particularly within the SNX4 subfamily that includes Snx4/Atg24 and Atg20/Snx42, where it participates in both retromer-independent retrograde trafficking pathways and selective autophagy processes including pexophagy[12][31]. The protein localizes to endosomal compartments enriched in PI3P and the phagophore assembly site (PAS) in yeast cells, enabling it to coordinate the retrieval of specific integral membrane proteins and lipids from the endosomal system and facilitate their recycling back to the trans-Golgi network[13][36]. Through its essential roles in protein sorting, membrane remodeling, and autophagy regulation, SNX41 contributes to maintaining cellular homeostasis and represents an important node in the endocytic-autophagic axis that is conserved across eukaryotic organisms and implicated in diverse physiological processes ranging from nutrient starvation responses to fungal pathogenesis.
Molecular Architecture and Structural Features of Sorting Nexin-41
Domain Organization and Phylogenetic Conservation
The sorting nexin-41 protein exhibits a modular domain architecture characteristic of the SNX4 subfamily of sorting nexins, with specific structural elements that confer its unique functions in membrane trafficking and autophagy. The protein is defined by the presence of a canonical phox homology (PX) domain, a conserved ~100-130 amino acid motif that was first identified in the NADPH phagocyte oxidase complex and has subsequently been recognized as the defining feature of the entire sorting nexin protein family[8][15][37]. This PX domain occupies a central position within the SNX41 sequence and serves as the primary determinant of membrane localization through its high-affinity binding to phosphatidylinositol 3-phosphate (PI3P), a phosphorylated lipid found enriched on endosomal and pre-autophagosomal membranes[18][40]. The crystal structure of PX domains reveals a characteristic structural fold consisting of three antiparallel β-strands (β1-β3) followed by three α-helices (α1-α3) that form a loop structure essential for PI3P recognition[37][42]. In addition to its PX domain, SNX41 contains a Bin-Amphiphysin-Rvs (BAR) domain at its C-terminus, though this domain may be weakly homologous to the typical BAR domain architecture found in other SNX-BAR family members[2][21]. The BAR domain is characterized by a curved, crescent-shaped dimeric structure that enables direct interaction with biological membranes, and in the context of SNX proteins, typically works in concert with the PX domain to generate membrane curvature and facilitate the formation of tubular carriers during retrograde trafficking[38][41]. The specific arrangement and properties of these domains in SNX41 reflects a hybrid architecture that combines structured domains with regions of intrinsic disorder, similar to related SNX4 subfamily members, enabling dynamic regulation and multiple protein-protein interactions[6][49].
Phylogenetic analysis reveals that sorting nexin-41 belongs to a well-characterized protein family that is highly conserved across eukaryotic organisms, with homologues identified in budding yeast (Saccharomyces cerevisiae), filamentous fungi, mammalian cells, and other model organisms[1][11][14][26]. The fission yeast SNX41 protein shows sequence similarity to both the budding yeast Snx41 and Snx42/Atg20 proteins, suggesting that the S. pombe orthologue may have combined functional roles of these two distinct yeast sorting nexins[2][19]. This evolutionary relationship is particularly significant because it indicates that SNX41 from fission yeast possesses functional domains that allow it to perform dual functions in both protein sorting (typically associated with Snx41) and selective autophagy processes including pexophagy (typically associated with Atg20/Snx42)[2][19]. The conservation of SNX41 across diverse eukaryotic lineages underscores the fundamental importance of this protein in cellular traffic control and suggests that the core mechanisms of endosomal sorting and autophagy mediated by SNX41 have remained largely unchanged throughout eukaryotic evolution[26][37]. This phylogenetic conservation provides strong support for the functional relevance of findings from model organisms like S. pombe to broader eukaryotic cell biology.
Structural Basis for Phosphoinositide Recognition
The molecular basis for PI3P recognition by SNX41 resides in the specific structural arrangement of residues within the PX domain that form a binding pocket optimized for interaction with the phosphorylated inositol headgroup of the phosphatidylinositol lipid[15][18][40]. The binding site is positioned at the junction between the β3 strand, α1 helix, and a proline-rich loop that connects the first and second α-helices, a region that is conserved among high-affinity PI3P-binding PX domains[15][37]. Key basic residues positioned within this binding pocket interact electrostatically with the negatively charged phosphate groups of PI3P, while an aromatic residue typically stacks against the inositol ring to stabilize the lipid binding interaction[15][40]. The interaction between the PX domain and PI3P is not merely a simple lock-and-key recognition event but rather involves conformational changes in the protein structure that increase membrane affinity upon lipid binding, creating a cooperative binding mechanism[15][18]. This PI3P-dependent membrane targeting mechanism is particularly significant for SNX41 function because it ensures that the protein localizes specifically to endosomal compartments where PI3P accumulates, as well as to forming autophagosomes and other autophagy-related structures where PI3P is generated by the Class III phosphatidylinositol 3-kinase (PI3K) complex[6][13][51]. The specificity of PX domain-lipid interactions can vary substantially across the sorting nexin family, with some PX domains exhibiting high-affinity binding to PI3P (dissociation constants of ~2-3 μM) while others show much lower affinity (>100 μM)[15]. The SNX41 protein appears to belong to the intermediate-affinity class of PI3P-binding proteins, enabling dynamic association and dissociation from membranes in response to changes in cellular conditions[13][36].
Functional Roles in Retrograde Trafficking Pathways
The SNX4-SNX41 Heterodimer and Atg27 Sorting
A crucial functional role for SNX41 has been elucidated through its assembly into heterodimeric complexes with other SNX4 subfamily members, particularly through its interaction with Snx4/Atg24, where the two proteins form a Snx4-Snx41 heterodimer that functions as a distinct molecular entity with cargo-specific sorting capacity[12][31][34]. Live cell imaging studies combined with biochemical analyses have revealed that this Snx4-Snx41 heterodimer coats endosome-derived tubular carriers and mediates retrograde transport of specific cargo proteins from post-Golgi endosomes back to the trans-Golgi network (TGN)[12][31]. The primary identified cargo of the Snx4-Snx41 pathway is Atg27, an integral membrane protein implicated in selective autophagy that cycles between the Golgi apparatus, endosomes, and autophagy-related compartments[12][16][31]. The functional specificity of the Snx4-Snx41 complex for Atg27 sorting appears to be conferred by the unique Snx41 subunit, with the Snx4 component serving as a common core that can partner with different SNX-BAR proteins (Snx41 or Atg20) to create distinct cargo-sorting complexes with different specificities[12][31][34]. The mechanistic basis for SNX41-dependent cargo recognition of Atg27 likely involves direct protein-protein interactions between SNX41 and specific sorting motifs within the Atg27 cytoplasmic tail, similar to well-characterized sorting nexin-cargo interactions in other systems[12][42]. Once Atg27 is recognized and recruited into Snx4-Snx41-coated tubules at the endosome, these carriers undergo fission to generate transport vesicles that traffic through the secretory pathway, ultimately delivering Atg27 to the TGN where it is then transported via the adaptor protein complex-3 (AP3) pathway to the vacuolar membrane[12][31]. The maintenance of proper Atg27 localization through SNX41-mediated sorting is important for sustaining the cellular pool of this autophagy protein and ensuring that adequate amounts of Atg27 are available at various compartments where it functions to facilitate autophagosome formation and cargo selection[16][31].
The Snx4-Snx41 pathway operates concurrently with other retrograde transport systems on individual maturing endosomes, as evidenced by live cell imaging studies showing that Snx4-Snx41-coated tubules coexist with retromer-coated carriers and other SNX-BAR complexes at the same endosomal structures[31][34]. This organization suggests that endosomes function as sorting stations where multiple SNX-BAR complexes simultaneously work to package different cargo molecules into distinct transport carriers heading to different destinations[31]. The fission of Snx4-Snx41-coated tubules from the endosomal membrane appears to involve the dynamin-related GTPase Vps1 in yeast, which was previously characterized for its role in promoting fission of retromer-coated carriers[31][34]. The coordinated actions of multiple SNX-BAR coat proteins on a single endosome likely reflect an ancient mechanism for ensuring efficient segregation of different cargo molecules and their delivery to appropriate destination compartments, a sorting principle that has been conserved from yeast through mammals[31][34]. This multiplexing of retrograde trafficking pathways on individual endosomes enables cells to maintain multiple regulatory checkpoints and achieve remarkable specificity in cargo delivery despite the biochemical complexity of endosomal sorting.
SNX41 in Distinct Retrograde Sorting Mechanisms
In addition to its role in the Snx4-Snx41 complex, SNX41 participates in other retrograde sorting mechanisms that mediate the retrieval of specific membrane proteins from post-Golgi endosomes back to the Golgi apparatus, representing a general cellular strategy for preventing cargo proteins from being degraded in the vacuole[13][36][53]. The sorting nexin SNX41, along with Snx4 and Snx42/Atg20, is specifically required for the retrieval of the exocytic v-SNARE protein Snc1p from post-Golgi endosomes, with genetic and biochemical studies demonstrating that deletion of snx41 results in mislocalization of Snc1 to the vacuole[13][36]. The requirement for SNX41 in Snc1 sorting is functionally distinct from its role in Atg27 retrieval, as the cargo specificity determinants for Snc1 differ from those required for Atg27 trafficking, indicating that SNX41 participates in multiple, cargo-specific retrograde pathways[12][13][31]. While the Snx4-Snx41 heterodimer mediates Atg27 sorting, the Snc1 SNARE protein appears to be primarily sorted by the Snx4-Atg20 (Snx4-Snx42) heterodimer, though SNX41 can participate in this pathway as well, suggesting some functional redundancy or cooperativity among SNX4 subfamily members[12][13][31]. The cross-linking of Snc1 directly to Snx4 demonstrates that SNX proteins can form specific protein-protein interactions with cargo molecules, and mutations in Snc1 that impair Snx4 binding (such as W86R) also prevent efficient Snc1 sorting, confirming the functional relevance of these interactions[13][36]. This cargo-selective binding model suggests that SNX41 similarly engages in direct interactions with Atg27 and potentially other cargo proteins, with the specificity being determined by the unique features of each SNX-BAR subunit[12][31].
Different sorting nexins operate in distinct endosomal compartments, reflecting a division of labor in retrograde trafficking that ensures specificity and efficiency of the sorting process[13][36][42]. While SNX41, Snx4, and Snx42 operate in post-Golgi endosomes (PGEs) in the early stages of endosomal maturation, other sorting nexins including Grd19p and retromer-associated SNX-BARs function at late/pre-vacuolar endosomes (PVEs), indicating a compartmental specialization of retrograde trafficking machinery[13][36]. This compartmentalization reflects a temporal aspect of cargo sorting, where different subsets of endosomal proteins are retrieved at distinct stages of the endosomal maturation pathway, allowing for multiple levels of quality control and trafficking decisions[13][36]. The SNX41-dependent retrieval pathways are generally non-essential for cell survival under normal laboratory growth conditions, suggesting that these proteins mediate regulatory trafficking functions that become critically important under specific physiological conditions or in specialized cell types[12][31]. However, in organisms with complex developmental programs such as fungi, the importance of SNX41-mediated sorting becomes apparent in contexts such as fungal conidiation and pathogenesis, as discussed in subsequent sections.
SNX41 in Selective Autophagy and Pexophagy
Pexophagy-Related Functions and Dual Roles of SNX41
Beyond its well-characterized roles in retrograde trafficking, SNX41 participates in selective autophagy pathways, particularly in the degradation of peroxisomes through a process termed pexophagy (peroxisomal autophagy), where damaged or excess peroxisomes are selectively engulfed by autophagosomes and delivered to the vacuole for degradation[2][19][27][30]. The involvement of SNX41 in pexophagy has been demonstrated through studies in both budding yeast and the rice blast fungus Magnaporthe oryzae, where SNX41-deletion mutants show defective pexophagy as evidenced by accumulation of intact peroxisomes within cells that normally undergo efficient autophagy under nutrient starvation conditions[2][19][30]. The molecular basis for SNX41 participation in pexophagy appears to involve its ability to recognize and tether peroxisomes that are destined for autophagic degradation, potentially through direct interaction with peroxisomal membrane proteins or through cargo selection activities similar to those mediated by canonical autophagy adaptors[2][19][30]. In the case of Magnaporthe oryzae, the fungal SNX41 orthologue (MoSnx41) exhibits sequence similarity to both budding yeast Snx41 and Snx42/Atg20, and complementation studies have demonstrated that MoSnx41 alone can functionally substitute for both yeast sorting nexins in different assays, indicating a functional consolidation of multiple sorting nexin roles in a single protein[2][19]. This consolidated functionality of fungal SNX41 proteins reflects an evolutionary divergence from budding yeast, where Snx41 and Snx42/Atg20 have maintained distinct, non-overlapping functions[2][19][30].
The pexophagy pathway represents an important cellular quality control mechanism that allows cells to regulate peroxisome abundance in response to changing metabolic demands and to eliminate damaged or senescent peroxisomes that may accumulate damaging oxidative byproducts[2][19][30]. Peroxisomes are organelles that catalyze numerous metabolic pathways including fatty acid oxidation and hydrogen peroxide metabolism, generating reactive oxygen species as byproducts that can damage cellular components if peroxisomes become dysfunctional[2][19][30]. The degradation of peroxisomes via pexophagy can be induced by nutrient starvation, particularly in response to glucose availability, creating a dynamic equilibrium between peroxisome biogenesis and autophagy-mediated degradation[2][19]. The molecular recognition of peroxisomes as selective autophagy substrates appears to depend on specific peroxisomal membrane proteins, with Pex14, a peroxisomal membrane protein, playing a particularly important role in pexophagy targeting[2][30]. Studies in Magnaporthe have revealed that the N-terminal cytosolic domain of Pex14 is essential for pexophagy, suggesting that this region serves as a recognition motif for autophagy machinery or SNX41-containing complexes[2][30]. The spatial organization and precise mechanisms by which SNX41 coordinates with core autophagy machinery (including Atg proteins) to facilitate selective autophagy of peroxisomes remains incompletely understood, but likely involves SNX41 functioning in a manner analogous to other selective autophagy receptors by recruiting cargo to assembling phagophores[2][30][47].
Integration with Broader Autophagy Networks
SNX41 functions within the broader context of cellular autophagy responses, participating in various selective autophagy pathways beyond pexophagy, including potential roles in the cytoplasm-to-vacuole targeting (Cvt) pathway and possibly mitophagy (selective autophagy of mitochondria)[2][27][49][50]. The Cvt pathway is a constitutive selective autophagy mechanism that targets hydrolytic enzymes such as the protease precursor aminopeptidase I (prApe1) to the vacuole, where they are processed and activated to become part of the vacuolar hydrolase complement[27][49][50]. SNX41 and its partner proteins are implicated in the Cvt pathway through their interactions with core autophagy proteins such as Atg1, Atg11, and Atg17, which form the autophagy initiation complex that nucleates phagophore formation at the phagophore assembly site[6][49]. The specific contribution of SNX41 to Cvt pathway cargo recruitment and autophagosome formation appears to involve its function in maintaining proper localization of autophagy proteins at the PAS, potentially through its PI3P-binding PX domain that localizes it to PI3P-enriched pre-autophagosomal membranes[6][49]. In addition to Cvt pathway components, SNX41 interacts with Atg11, a scaffold protein required for selective autophagy, and this interaction may be mediated by specific protein-protein interaction domains within SNX41[6][49]. The precise mechanistic role of SNX41 in facilitating cargo engagement with the autophagy machinery at the PAS remains to be fully elucidated, but likely involves either direct recruitment of cargo proteins or remodeling of pre-autophagosomal membranes to facilitate cargo sequestration[2][6][49].
The role of SNX41 in nonselective autophagy (bulk autophagy in response to nutrient starvation) appears to be distinct from its role in selective autophagy pathways, as some studies have reported that SNX41 is not strictly required for efficient bulk autophagy under standard laboratory conditions[2][9][25]. However, recent work has revealed that SNX4 family sorting nexins, including SNX41, can contribute to nonselective autophagy through indirect mechanisms involving lipid trafficking and maintenance of membrane fusion competence[9][25]. Specifically, SNX4-Atg20 and potentially SNX4-SNX41 complexes mediate export of phosphatidylserine (PS) from the endosome and vacuole through retrograde trafficking, which maintains proper lipid composition of the endolysosomal system membranes required for autophagosome-vacuole fusion[9][25]. This mechanism reveals an important connection between retrograde trafficking functions and autophagy, where the same molecular machinery (SNX-BAR complexes) that sorts specific proteins also regulates the lipid composition of trafficking compartments to support autophagy[9][25]. The discovery of this indirect role for SNX41-containing complexes in nonselective autophagy expands our understanding of how retrograde trafficking and autophagy are coordinately regulated to maintain cellular homeostasis[9][25].
SNX41 Localization and Membrane Association
Subcellular Distribution and PI3P-Dependent Targeting
The subcellular localization of SNX41 is primarily determined by its PI3P-binding PX domain, which directs the protein to endosomal and autophagy-related membrane compartments where this phosphoinositide accumulates[50][53]. In wild-type fission yeast cells expressing SNX41-GFP fusion proteins under native regulation, the protein exhibits a punctate distribution pattern characteristic of endosomal localization, with individual foci representing distinct endosomal compartments or tubular carriers derived from endosomes[50][53]. The localization of SNX41 to these membrane structures is largely dependent on the PX domain, as demonstrated by studies in which targeted deletion of the PX domain (generating SNX41Δ215-627 lacking the N-terminal PX region) results in predominantly cytoplasmic distribution of the protein[50]. This cytoplasmic localization of PX-deficient SNX41 correlates with loss of function in SNX41-dependent processes, confirming that membrane association through PX domain-mediated PI3P binding is essential for SNX41 biological activity[50]. The endosomal localization of SNX41 is partially overlapping with the distribution of other SNX4 subfamily members and retromer components, consistent with the model that multiple SNX-BAR complexes coexist on individual endosomes[50][53]. SNX41 additionally localizes to punctate structures that associate with or are adjacent to autophagosomes, as determined by colocalization with the autophagy marker Atg8 (visualized as RFP-Atg8 in immunofluorescence studies)[50]. This localization to pre-autophagosomal and autophagosomal structures is consistent with SNX41's role in selective autophagy processes and supports a model in which SNX41 functions directly at the sites of autophagosome formation to facilitate cargo selection and sequestration[50].
The localization of SNX41 appears to be independent of the canonical autophagy pathway in certain contexts, as the punctate distribution of SNX41-GFP persists in autophagy-defective mutants, suggesting that SNX41's endosomal localization is primarily determined by PI3P availability on endosomes rather than by direct recruitment to autophagosomes by autophagy machinery[50]. However, the localization of SNX41 to the PAS (phagophore assembly site, the site of autophagosome nucleation) appears to depend on Class III PI3K-mediated generation of PI3P at this site, as disruption of PI3K activity impairs SNX41 recruitment to the PAS[50][51]. This dependency on PI3K activity reveals a key regulatory mechanism: the localization of SNX41 (and by extension, its autophagy functions) is dynamically regulated by the activity of PI3K, which increases PI3P production when autophagy is initiated[6][49][50][51]. The perinuclear localization of SNX41 during starvation-induced autophagy has been observed in yeast, suggesting that nuclear cargos (such as nuclear proteins destined for autophagy or autophagy regulators) may be transported in SNX41-containing carriers from the nucleus to the vacuole[52]. This perinuclear recruitment may represent a specialized SNX41 function distinct from its roles in general endosomal sorting, potentially involving nuclear envelope-derived membranes that accumulate PI3P during autophagy induction[52].
Regulation of SNX41 Localization and Activity
The localization and activity of SNX41 is regulated by post-translational modifications and protein-protein interactions that modulate its membrane binding capacity and cargo recognition specificity. SNX41 protein levels and localization can be affected by interactions with partner proteins, as deletion of Snx4 (the binding partner of Snx41 in yeast) results in loss of endosomal localization of Snx41, indicating that Snx4 stabilizes Snx41 association with membranes or recruits Snx41 to endosomes[12][55]. This dependency on partner proteins for localization suggests that SNX41 does not efficiently associate with endosomal PI3P on its own but requires stabilization through oligomerization with other SNX-BAR proteins, a principle that may underlie the requirement for SNX4 as a common core component of multiple heterodimeric complexes[12][55]. The BAR domain of SNX41, while potentially weakly conserved compared to canonical BAR domains, likely still contributes to protein-protein interactions and membrane binding, as even subtle disruptions to BAR domain structure can impair SNX41 function[2][21]. Post-translational modifications such as phosphorylation and acetylation, well-characterized for the related Atg20 protein, may also regulate SNX41 activity by modulating its conformation and protein-protein interaction capacity, though specific SNX41 modifications have not been extensively characterized[6][49]. The recruitment of SNX41 to specific endosomal subdomains may also be regulated by the distribution of its cognate cargo proteins, creating a feedback loop where cargo abundance influences SNX41 localization and activity[12][31].
Interaction Partners and Molecular Networks
Heterodimeric Complex Formation with SNX4 and Other SNX-BAR Proteins
SNX41 functions primarily as a component of heterodimeric complexes rather than as a solitary protein, with the yeast SNX4 protein representing the primary obligate binding partner that enables SNX41 endosomal localization and autophagy-related functions[12][31][34][55]. The Snx4-Snx41 heterodimer forms through interactions between the BAR domains of the two proteins, which dimerize to create the characteristic crescent-shaped architecture that enables membrane curvature sensing and induction[12][31][34]. This heterodimer assembly appears to be highly specific, as biochemical studies demonstrate that SNX41 does not form homodimers and shows minimal interaction with distantly related SNX-BAR family members[12][31]. The formation of distinct Snx4 heterodimers with different SNX-BAR partners (Snx41 versus Atg20/Snx42) represents a molecular strategy for generating multiple functional specificity despite employing a common structural scaffold, with the unique properties of Snx41 versus Snx42 determining which specific cargo molecules are recognized and sorted by each dimer[12][31]. The assembly of these heterodimeric complexes appears to be constitutive and relatively stable on endosomal membranes, suggesting that once assembled, the Snx4-Snx41 dimer remains intact as a functional unit as it transits through various trafficking compartments[12][31][34]. Cross-linking studies demonstrate specific protein-protein interaction surfaces between Snx4 and Snx41, with particular importance for the N-terminal region of Snx4 in mediating these interactions[12][31].
In organisms with more complex sorting nexin gene complements, such as mammals, SNX4 forms heterodimers with multiple related SNX-BAR proteins (SNX1/SNX2 and SNX5/SNX6 in mammals), creating a modular system for combinatorial sorting and trafficking regulation[37][42][58]. In contrast, the S. pombe genome contains fewer SNX genes than the mammalian genome, with the SNX41 protein assuming some of the functional roles of multiple mammalian SNX-BARs[37][42]. The SNX4-SNX41 complex in S. pombe functionally corresponds to mammalian SNX4-SNX7/SNX30 complexes that regulate autophagy through control of ATG9 trafficking[37][58][60]. The specific interaction surfaces and dimer stability of Snx4-Snx41 may differ from those of other SNX4 heterodimers, contributing to the specific cargo recognition and trafficking patterns mediated by this complex[12][31]. The requirement for Snx4 as a common subunit in multiple heterodimeric complexes suggests that the unique N-terminal region of Snx4 likely mediates initial recruitment to endosomal membranes and subsequent assembly with different partner proteins depending on cellular conditions and cargo availability[12][31][34].
Interactions with Autophagy Machinery and Cargo Proteins
SNX41 physically and functionally interacts with core autophagy proteins, particularly those involved in autophagy initiation and selective cargo recruitment, creating molecular bridges between endosomal sorting and autophagosome formation machinery[6][49][50]. The interaction between SNX41 and Atg11, a scaffold protein central to selective autophagy, has been proposed based on the requirement for SNX4 family proteins in Cvt pathway function and evidence that sorting nexins associate with Atg1 kinase complex components[6][49]. This SNX41-Atg11 interaction may serve to recruit SNX41-containing endosomal carriers to the PAS or to coordinate cargo selection between endosomal sorting and autophagosome formation[6][49]. SNX41 also interacts directly or indirectly with Atg9, an integral membrane protein required for autophagosome formation that cycles through various cellular compartments including the Golgi apparatus, endosomes, and autophagosomes[12][31][34]. The precise molecular basis for SNX41 interaction with Atg9 remains to be fully characterized, but likely involves either direct protein-protein interactions or indirect coupling through shared localization on endosomal membranes[12][31][34]. The importance of Atg27 as a primary cargo of Snx4-Snx41 retrograde trafficking has already been discussed, but it is notable that Atg27 itself is an integral membrane protein implicated in selective autophagy, suggesting a functional link between SNX41-mediated Atg27 sorting and the autophagy pathway[12][31].
In the fungal pathogen Magnaporthe oryzae, SNX41 physically interacts with Oxp1 (5-oxoprolinase), an enzyme involved in the gamma-glutamyl cycle that regenerates glutamate for continued glutathione synthesis[50]. This SNX41-Oxp1 interaction has been demonstrated through affinity purification and co-immunoprecipitation, and disruption of the interaction in snx41 deletion mutants results in mislocalization of Oxp1 to the vacuole rather than its normal cytoplasmic distribution[50]. The SNX41-dependent sorting and retrieval of Oxp1 from the vacuole suggests that SNX41 participates in retrograde pathways distinct from those involving Atg27, indicating functional specialization for different cargo molecules[50]. The importance of Oxp1 for fungal conidiation and the concomitant requirement for SNX41 in these processes suggests that SNX41-mediated sorting of metabolic enzymes involved in glutathione biosynthesis is critical for developmental programs requiring high antioxidant production[50]. This discovery expands the known cargo repertoire of SNX41 beyond canonical autophagy and endosomal trafficking to include metabolic regulation, highlighting the multiple roles that this protein plays in coordinating cellular physiology.
Regulatory Interactions and Protein Quality Control
SNX41 participates in protein quality control pathways by mediating the trafficking of proteins destined for degradation or reuse, thereby regulating the cellular abundance of specific proteins and protein complexes. The interaction between SNX4-SNX41 complexes and potential ubiquitinated cargo proteins has been proposed based on analogy with other sorting nexin cargo recognition mechanisms, where ubiquitin modifications on cargo proteins can serve as recognition signals[37][42][52]. SNX41 likely works in concert with ubiquitin-binding adaptor proteins and ESCRT (Endosomal Sorting Complex Required for Transport) machinery to distinguish between cargo molecules destined for recycling through retrograde pathways versus those destined for vacuolar degradation[37][42][52]. The regulation of SNX41 activity through interactions with regulatory proteins such as kinases and acetyltransferases may provide another layer of quality control, ensuring that SNX41 is active only when specific cellular conditions warrant retrograde trafficking and selective autophagy[6][49]. The coordination of SNX41 with other membrane trafficking machinery such as Vps1 (dynamin-related GTPase) for tubule fission suggests that SNX41 function is subject to regulatory inputs from cytoskeletal and energetic status of the cell[31][34].
Biological Significance in Cellular Processes
Role in Sporulation and Developmental Transitions in S. pombe
The biological significance of SNX41 in Schizosaccharomyces pombe becomes particularly evident in the context of sporulation, the developmental transition from vegetative growth to sexual reproduction through meiosis and ascospore formation[1][43][46]. The snx41 gene was originally identified in S. pombe as mug186 (meiotically up-regulated gene 186), indicating elevated expression during meiosis and sporulation, which directly implicates SNX41 in the developmental program controlling spore formation[1][7][43]. During sporulation, S. pombe undergoes dramatic morphological and biochemical changes, including the formation of the forespore membrane (FSM) that encapsulates the developing ascospores, requiring extensive membrane trafficking and reorganization of cellular structures[43][46][47][48]. The remodeling of membrane systems during sporulation likely depends critically on retrograde trafficking pathways mediated by SNX41, as these pathways ensure that membrane proteins required for spore development are properly localized and maintained in appropriate cellular compartments[43][46][47][48]. The upregulation of SNX41 during meiosis suggests that SNX41-dependent trafficking becomes especially important during the developmental switch from mitotic growth to meiotic development, potentially to recycle proteins involved in mitotic cell cycle control and to promote new pathways required for sexual differentiation[43][46]. The meiotically-restricted expression pattern of SNX41 indicates that this protein is regulated at the transcriptional or post-transcriptional level in response to nutrient starvation signals that trigger sporulation, reflecting a sophisticated developmental program that coordinates SNX41 function with other aspects of the meiotic and sporulation programs[43][46].
The role of endocytic trafficking in sporulation has been further illuminated by studies showing that mutations in endocytic machinery, including components of the actin cytoskeleton required for endocytosis, severely impair sporulation[47][48]. Specific syntaxin (SNARE) proteins such as Psy1, which undergo dynamic endocytic recycling during sporulation, are critical for proper development, and their recycling is mediated in part by SNX41-dependent retrograde trafficking[47]. The relocation of Psy1 from the plasma membrane to the nascent forespore membrane during meiosis II requires both endocytosis to remove Psy1 from the cell surface and subsequent recycling back to the developing spore, processes that are likely coordinated by SNX41-containing trafficking carriers[47]. The severe defects in sporulation observed in endocytic mutants suggest that the trafficking of membrane proteins through SNX41-dependent routes is essential for establishing the unique membrane environment required for spore development[47][48]. This developmental context provides compelling evidence that SNX41's roles in membrane trafficking and autophagy are not merely housekeeping functions but are actively integrated into developmental programs controlling cell fate transitions.
Implications for Fungal Pathogenicity
The functional importance of SNX41 extends beyond laboratory model organism biology to fungal pathogenicity, as demonstrated by studies in the rice blast fungus Magnaporthe oryzae, where SNX41 homologues are essential for fungal virulence[2][19][30]. In M. oryzae, the SNX41 orthologue (MoSnx41) is required for conidiation (asexual spore formation), the developmental stage at which the fungus disperses and infects host plants[2][19][30][50]. The conidiation defect in snx41 deletion mutants of M. oryzae reflects a role for SNX41 in supporting the metabolic and cellular remodeling required for spore development, potentially through its functions in retrograde trafficking and the gamma-glutamyl cycle/glutathione biosynthesis pathway[50]. The importance of SNX41 for conidiation in M. oryzae parallels its meiotically-restricted expression and presumed developmental functions in S. pombe, suggesting that SNX41 has evolutionarily conserved roles in fungal sporulation and spore development across diverse fungal taxa[2][19][30][50]. Beyond conidiation, SNX41 is required for full pathogenicity of M. oryzae during infection of host plants, indicating that even beyond spore development, SNX41-dependent functions contribute to the ability of the fungus to survive and proliferate within host tissues[50]. The SNX41-dependent sorting of Oxp1 and the consequent maintenance of glutathione biosynthesis capacity appears to be particularly important for maintaining the redox balance required for both spore development and pathogenic colonization of the host[50].
Studies in Magnaporthe have additionally revealed that pexophagy, dependent on SNX41 function, occurs during appressorial development and function, the specialized infection structure that M. oryzae uses to breach the plant cell wall[2][19][30]. The pexophagy observed during appressorial development may support the high-energy demands of appressorium formation and function, where peroxisomes may be remodeled to support enhanced metabolic activity or to reduce cellular oxidative stress[2][19][30]. While genetic studies have shown that pexophagy itself is not absolutely essential for conidiation or pathogenesis (through complementation studies with mutations that disrupt pexophagy but not protein sorting functions of SNX41), the fact that pexophagy is naturally induced during these developmental stages suggests an important biological role even if redundant or backup mechanisms can substitute when pexophagy is specifically ablated[2][30]. The discovery of SNX41's role in fungal pathogenicity has important implications for understanding fungal-plant interactions and potentially for developing novel antifungal strategies targeting this conserved protein.
Molecular Mechanisms and Structural Dynamics
Membrane Remodeling and Tubule Formation
A critical molecular function of SNX41 involves its participation in the physical remodeling of biological membranes to generate the curved, tubular structures required for retrograde trafficking and selective autophagy. The BAR domain of SNX41, when assembled into heterodimeric complexes with Snx4, creates a membrane-binding surface with positive curvature that can induce or stabilize curved membrane topologies[21][38][41][49]. The BAR domain superfamily of proteins are known to generate membrane curvature through both scaffolding mechanisms (where the BAR domain acts as a molecular "spine" that supports membrane bending) and wedging mechanisms (where amphipathic helices at the domain periphery penetrate the lipid bilayer to generate curvature)[38][41]. The relative contribution of scaffolding versus wedging to SNX41-mediated membrane curvature is not fully established, but likely involves both mechanisms working in concert[38][41]. The generation of membrane curvature by Snx4-SNX41 dimers helps facilitate the transition from the relatively flat endosomal membrane to the highly curved tubular carriers that pinch off and transport cargo to the Golgi apparatus, a transition that is energetically unfavorable without BAR domain proteins[38][41]. The stabilization of curved membrane intermediates by SNX41-containing complexes can also recruit additional proteins that promote membrane fission, including dynamins and other GTPases, creating a functional platform for coordinated membrane deformation and carrier biogenesis[31][34][38][41].
The molecular dynamics of Snx4-SNX41 complex formation and assembly on membranes involve stepwise processes of protein recruitment, oligomerization, and cargo loading that are coordinated with membrane topology changes[31][34][38][41]. The initial recruitment of Snx4-SNX41 to endosomal membranes is driven by the PI3P-binding activity of the PX domains, which recognize and bind to PI3P-containing membrane microdomains[13][31][36]. Following initial recruitment through PX domain interactions, the BAR domains of the heterodimer interact with the lipid bilayer in a manner that is thought to stabilize membrane curvature, converting the initially planar or gently curved endosomal membrane into a highly curved tubular structure[31][34][38][41]. As cargo loading occurs through recognition of specific sorting motifs in cargo proteins, the oligomerization of multiple Snx4-SNX41 dimers may enhance membrane curvature by cooperative interactions between multiple BAR domain dimers, creating a lattice-like coat structure similar to COPII and clathrin coats[31][34][38][41]. The subsequent recruitment of fission machinery such as Vps1 leads to scission of the tubular carrier from the endosomal membrane, releasing a transport intermediate that carries sorted cargo toward its destination[31][34].
PI3K and PtdIns(3)P Signaling in SNX41 Function
The function of SNX41 is intimately connected to phosphoinositide signaling, particularly the generation and turnover of phosphatidylinositol 3-phosphate (PtdIns(3)P or PI3P) on endosomal and autophagy-related membranes, through the activities of Class III PI3K enzymes such as Vps34[13][51][54]. The Class III PI3K complex, which includes the catalytic subunit Vps34 and regulatory components such as Vps15 and Atg14, generates PI3P at the PAS during autophagy initiation and at endosomes during constitutive endosomal trafficking[13][51]. The PI3P generated by Vps34 serves as a lipid recognition signal that recruits PI3P-binding proteins including SNX41 to these membrane compartments, thereby coupling the activity of the PI3K complex to the localization and function of SNX41[13][51][54]. The requirement for PtdIns(3)P-mediated signaling in pexophagy has been demonstrated by genetic studies showing that deletion of Vps34 (the only PI3K in yeast that generates PI3P for autophagy) results in loss of pexophagy, with a phenotype similar to that of snx41 deletion mutants, suggesting that PI3P generation is a prerequisite for SNX41 recruitment to pexophagosomes[51]. The importance of PI3K signaling for SNX41 function extends to selective autophagy pathways, where the PI3K complex localizes to the PAS through interactions with selective autophagy adaptor proteins such as Atg11, creating localized regions of high PI3P concentration that concentrate SNX41 and other PI3P-binding proteins[6][49][51].
The lipid kinase Vps34 has been shown to be associated with peroxisomes during their biogenesis, suggesting that SNX41 may interact with nascent or newly synthesized peroxisomes through PI3P-mediated recognition on peroxisomal membranes[51]. This observation supports a model in which pexophagy is initiated when peroxisomes are recognized through PI3P-dependent interactions with SNX41 and other autophagy proteins, leading to the wrapping of peroxisomes by phagophores and their ultimate degradation in the vacuole[51]. The dynamic regulation of PI3P levels on endosomal and autophagy-related membranes through the balanced activities of Class III PI3K (which generates PI3P) and PI3P-specific phosphatases (which degrade PI3P) provides a mechanism for temporally and spatially controlling SNX41 localization and activity[13][51][54]. The requirement for specific spatial organization of PI3P on membranes suggests that SNX41 recognizes not just PI3P lipids in isolation, but rather specific PI3P-containing membrane domains or microenvironments created through cooperative binding of multiple PI3P-binding proteins[13][31][36].
Evolutionary Conservation and Comparative Functional Analysis
SNX41 Homologues Across Eukaryotic Organisms
The sorting nexin protein family is highly conserved across eukaryotic evolution, with SNX41 homologues identified in organisms ranging from simple yeasts to complex multicellular organisms including plants and animals[11][26][37][42]. In budding yeast Saccharomyces cerevisiae, multiple SNX proteins related to S. pombe SNX41 have been identified, including Snx41 and Snx4, which perform both overlapping and distinct functions in endosomal sorting and autophagy[12][31][36][52][53]. Mammalian cells possess a larger complement of SNX-BAR proteins (at least twelve distinct members including SNX1, SNX2, SNX4, SNX5, SNX6, SNX7, SNX8, SNX9, SNX18, SNX30, SNX32, and SNX33), reflecting whole-genome duplications during vertebrate evolution and functional diversification of SNX genes[11][37][42][58][60]. The mammalian SNX4 homologue functions similarly to yeast SNX4, forming heterodimeric complexes with other SNX-BAR proteins such as SNX7 and SNX30 to regulate ATG9 trafficking and autophagy[37][58][60]. The mammalian SNX-BAR proteins engaged in autophagy regulation appear to perform analogous functions to the yeast Snx4 family proteins, mediating both retrograde trafficking and selective autophagy processes[37][58][60].
The expansion of the SNX gene family in mammals compared to simpler eukaryotes reflects both the increased complexity of mammalian endosomal systems and the evolution of new regulatory mechanisms for controlling membrane trafficking and autophagy in multicellular organisms[37][42][58][60]. Despite the expansion in gene number, the core biochemical principles underlying SNX function—including PI3P-dependent membrane targeting, BAR domain-mediated membrane remodeling, and cargo-specific sorting—appear to have remained largely conserved[37][42][58][60]. The functional consolidation evident in S. pombe SNX41, where a single protein performs roles analogous to multiple mammalian SNX-BARs, suggests that the ancestral SNX proteins may have had more promiscuous functions that became specialized through gene duplication and functional divergence in more complex organisms[2][19][26][37][42]. The conservation of SNX41 across diverse fungal species, including not only model organisms like S. pombe but also human pathogens such as M. oryzae and Fusarium graminearum, indicates that SNX41 is likely a suitable target for antifungal drug development, as inhibition of SNX41 function could potentially disrupt both basal cellular functions and specialized pathogenic processes in pathogenic fungi[2][19][27][30][35].
Functional Specialization and Redundancy in SNX Families
The SNX4 subfamily of sorting nexins, which includes SNX41, represents a specialized subset of the sorting nexin family with distinct functional properties and biological roles compared to other SNX subfamilies such as SNX-PX proteins (SNX3) and SNX-FERM proteins (SNX17, SNX27, SNX31)[11][26][37][42][52]. The SNX4 subfamily proteins are specifically defined by their possession of both PX and BAR domains, enabling both PI3P-dependent membrane targeting and BAR domain-mediated membrane curvature generation[11][26][37][42]. The SNX4 subfamily functions are largely distinct from those of retromer-associated SNX proteins (such as SNX-BARs with retromer or SNX3), which mediate different retrograde trafficking pathways with different cellular destinations and cargo specificities[11][26][37][42][52]. The distinct localizations and functions of different SNX subfamilies suggest that the SNX family has evolved through gene duplication and functional divergence to generate multiple, partially redundant systems for protein sorting and membrane trafficking, providing robustness to cellular trafficking networks[11][26][37][42][52]. The functional specialization is evident in the specific cargo recognition capacities of different SNX proteins, where SNX41 specifically recognizes Atg27 for retrograde trafficking, while SNX42/Atg20 recognizes Snc1 and other distinct cargo molecules[12][31][34][36].
Within the SNX4 subfamily itself, functional redundancy is evident, as evidenced by the fact that deletion of single SNX genes often does not result in complete loss of function, but rather reduced efficiency of the affected pathway[12][31][34]. In yeast, Snx4, Snx41, and Atg20 are not functionally interchangeable but exhibit partial redundancy in some contexts, suggesting that the three proteins have related but distinct functions and that backup mechanisms can partially compensate for loss of individual proteins[12][31][34][36][52][53]. The functional specificity of different SNX4 subfamily members is achieved through distinct protein-protein interaction domains (beyond the conserved PX and BAR domains) that enable preferential binding to particular partner proteins and cargo molecules[12][31][34]. This modular organization of SNX function, where conserved structural domains (PX and BAR) provide core trafficking functions while variable protein-interaction regions confer specificity, represents an elegant evolutionary solution to achieving both conservation of core function and diversification of regulatory mechanisms.
Significance for Understanding Cellular Homeostasis and Disease
Connections to Protein Quality Control and Cellular Stress Responses
SNX41 and related sorting nexins play important roles in cellular quality control and stress responses, where they contribute to the selective removal of damaged or aggregated proteins and organelles through autophagy-mediated pathways[37][42][52]. The participation of SNX41 in pexophagy represents one example of how SNX41 can serve quality control functions by mediating the selective degradation of damaged or senescent peroxisomes that may accumulate oxidative damage[2][19][30][51]. The role of SNX41 in maintaining proper localization of metabolic enzymes such as Oxp1 (involved in the gamma-glutamyl cycle) suggests that SNX41 also contributes to metabolic homeostasis by preventing the sequestration of essential metabolic enzymes in the vacuole where they would be degraded[50]. The connection between SNX41 function and glutathione biosynthesis through Oxp1 sorting highlights an indirect but important role for SNX41 in cellular redox homeostasis, as glutathione is a critical antioxidant that protects cells from oxidative stress[50]. The importance of SNX41 for maintaining antioxidant capacity appears to be particularly critical during developmental transitions such as sporulation or conidiation, where metabolic demands are high and oxidative stress may be elevated[50]. The selective autophagy pathways mediated by SNX41 can thus be understood not merely as catabolic processes for generating energy and amino acids during nutrient starvation, but as integral components of cellular quality control systems that maintain a clean, functional proteome and lipidome[37][42][52].
The integration of SNX41 functions into stress response pathways suggests that this protein may be subject to regulation by cellular signaling cascades that respond to nutrient availability, oxidative stress, and other environmental challenges. The upregulation of SNX41 during nutrient starvation (as evidenced by its meiotically-restricted expression pattern during sporulation triggered by nutrient depletion) indicates that SNX41 expression is responsive to stress signals[43][46]. The coordination of SNX41 activity with other autophagy proteins through phosphorylation and other post-translational modifications (as has been documented for related Atg20 protein) suggests that SNX41 is subject to multiple layers of regulation that allow cells to precisely tune autophagy responses to current metabolic conditions and cellular stress levels[6][49]. The dysregulation of SNX41 or related SNX proteins in human disease contexts has not been extensively studied, but the critical roles of these proteins in autophagy suggest that their dysfunction could contribute to disease phenotypes characterized by defective autophagy, such as neurodegenerative diseases and certain cancers[29][37][42][52].
Potential Targets for Therapeutic Intervention
The essential roles of SNX41 in fungal pathogenicity and the conservation of SNX41 homologues across diverse fungal species raise the possibility that SNX41 could serve as a target for antifungal drug development[2][19][27][30][35]. The requirement for SNX41 in conidiation of M. oryzae suggests that inhibitors of SNX41 function could prevent spore formation, potentially blocking the dispersal and transmission of fungal pathogens[2][19][30][50]. Similarly, the role of SNX41 in pathogenic development following infection suggests that SNX41 inhibitors could reduce the virulence of fungi that have already initiated infection[50]. The relative conservation of SNX41 between distantly related fungi versus differences in SNX protein composition between fungi and mammals suggests that it may be possible to develop SNX41-specific inhibitors with selectivity for fungal rather than mammalian sorting nexins, potentially reducing off-target effects in human cells[26][37][42]. The specific functions of SNX41 in maintaining the gamma-glutamyl cycle and glutathione biosynthesis in fungi might be exploited for selective toxicity, as fungi appear to depend more heavily on these pathways than mammalian cells for maintaining redox balance[50]. Potential inhibitory strategies could include disruption of SNX41 protein-protein interactions (preventing heterodimer formation with Snx4), inhibition of PI3P binding through the PX domain, or destabilization of the SNX41 protein through targeting of post-translational modification sites[2][19][26][37][42].
The potential drawbacks of targeting SNX41 for therapeutic purposes must also be considered, particularly regarding the redundancy and multiple functions of sorting nexin proteins in cellular physiology. The fact that SNX41-defective cells remain viable under laboratory conditions (albeit with defects in specific developmental or pathogenic processes) suggests that complete loss of SNX41 function may be tolerated by cells, but partial or conditional inhibition might be necessary to achieve therapeutic effects while minimizing toxicity[2][30]. The off-target effects of SNX41 inhibitors on other sorting nexin functions and on general autophagy pathways would need to be carefully evaluated, as disruption of autophagy generally can have complex effects on cell survival and disease progression[37][42][52].
Conclusion
Sorting nexin-41 (SNX41) represents a multifunctional protein that integrates several critical cellular processes including retrograde trafficking, selective autophagy, and cellular stress responses in the model organism Schizosaccharomyces pombe and in fungal pathogens such as Magnaporthe oryzae. Through its dual PX and BAR domain architecture, SNX41 localizes to PI3P-rich endosomal and autophagy-related membranes, where it functions as a core component of heterodimeric complexes with other SNX4 subfamily members to mediate cargo-specific sorting and membrane remodeling. The primary functions of SNX41 involve the retrograde trafficking of specific integral membrane proteins such as Atg27 and the participation in selective autophagy pathways including pexophagy, processes that are coordinated through dynamic regulation by Class III PI3K-generated PI3P. The meiotically-restricted expression of SNX41 in S. pombe and its essential roles in conidiation and pathogenicity in M. oryzae indicate that SNX41 has specialized roles in fungal developmental programs and pathogenic processes beyond its contributions to basal cellular functions. The conservation of SNX41 homologues across eukaryotic evolution, coupled with functional specialization reflecting gene family expansion in complex organisms, highlights the evolutionary importance of this protein family in eukaryotic cell biology. Future research directions should focus on elucidating the precise molecular mechanisms of SNX41-mediated cargo recognition, identifying post-translational modifications that regulate SNX41 activity, characterizing the full spectrum of SNX41 cargo molecules, and exploring the therapeutic potential of SNX41 inhibitors for controlling fungal pathogenicity while minimizing effects on host cell autophagy.
Citations
- https://www.pombase.org/gene/SPBC14F5.11c
- https://journals.plos.org/plosone/article?id=10.1371%2Fjournal.pone.0079128
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5766950/
- https://www.uniprot.org/uniprotkb/O60107/entry
- https://www.yeastgenome.org/locus/S000002833
- https://www.pnas.org/doi/10.1073/pnas.1708367114
- https://www.uniprot.org/uniprotkb/O14201/entry
- https://pmc.ncbi.nlm.nih.gov/articles/PMC117945/
- https://www.molbiolcell.org/doi/10.1091/mbc.E17-12-0743
- https://en.wikipedia.org/wiki/Sorting_nexin
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5262529/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC140746/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1073682/
- https://www.yeastgenome.org/complex/CPX-1378
- https://thebiogrid.org/276275/table/schizosaccharomyces-pombe/mug186.html
- https://journals.biologists.com/jcs/article/115/6/1099/2813/The-PX-domain-a-new-phosphoinositide-binding
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3841179/
- https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1356-9597.2004.00744.x
- https://ecommons.cornell.edu/bitstreams/38d453c4-1263-490f-ae1b-f91d902d4d06/download
- https://ediss.uni-goettingen.de/bitstream/handle/11858/14120/PhDThesis_MatthewTaylor_2022.pdf?sequence=1&isAllowed=y
- https://pubmed.ncbi.nlm.nih.gov/15189449/
- https://pubmed.ncbi.nlm.nih.gov/25256216/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6249802/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7823608/
- https://pubmed.ncbi.nlm.nih.gov/32281734/
- https://www.molbiolcell.org/doi/abs/10.1091/mbc.E17-12-0743
- https://pubmed.ncbi.nlm.nih.gov/38874037/
- https://www.molbiolcell.org/doi/prev/20190508-aop/pdf/10.1091/mbc.E19-02-0117
- https://pubmed.ncbi.nlm.nih.gov/28026081/
- https://onlinelibrary.wiley.com/doi/abs/10.1111/cmi.13208
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4832598/
- https://elifesciences.org/articles/77035
- https://elifesciences.org/articles/22311
- https://repository.upenn.edu/entities/publication/3911ff00-a4c5-4250-8392-c818f95d5510
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9006456/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9214231/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7086038/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4355233/
- https://pubmed.ncbi.nlm.nih.gov/19778961/
- https://www.molbiolcell.org/doi/10.1091/mbc.e11-03-0255
- https://journals.biologists.com/jcs/article/117/3/389/27963/Forespore-membrane-assembly-in-yeast-coordinating
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3429543/
- https://pubmed.ncbi.nlm.nih.gov/21121900/
- https://febs.onlinelibrary.wiley.com/doi/10.1016/j.febslet.2010.01.011
- https://www.molbiolcell.org/doi/10.1091/mbc.E19-02-0117
- https://onlinelibrary.wiley.com/doi/abs/10.1111/tra.12462
- https://www.uniprot.org/uniprotkb/Q5AZC9/entry
- https://journals.biologists.com/jcs/article/133/14/jcs246306/224907/A-heterodimeric-SNX4-SNX7-SNX-BAR-autophagy