Deep Research
Falcon
(tam10-deep-research-falcon.md)
Falcon
(tam10-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: Schizosaccharomyces pombe tam10 (SPBC14C8.19; UniProt G2TRQ9) — functional annotation status
Executive summary
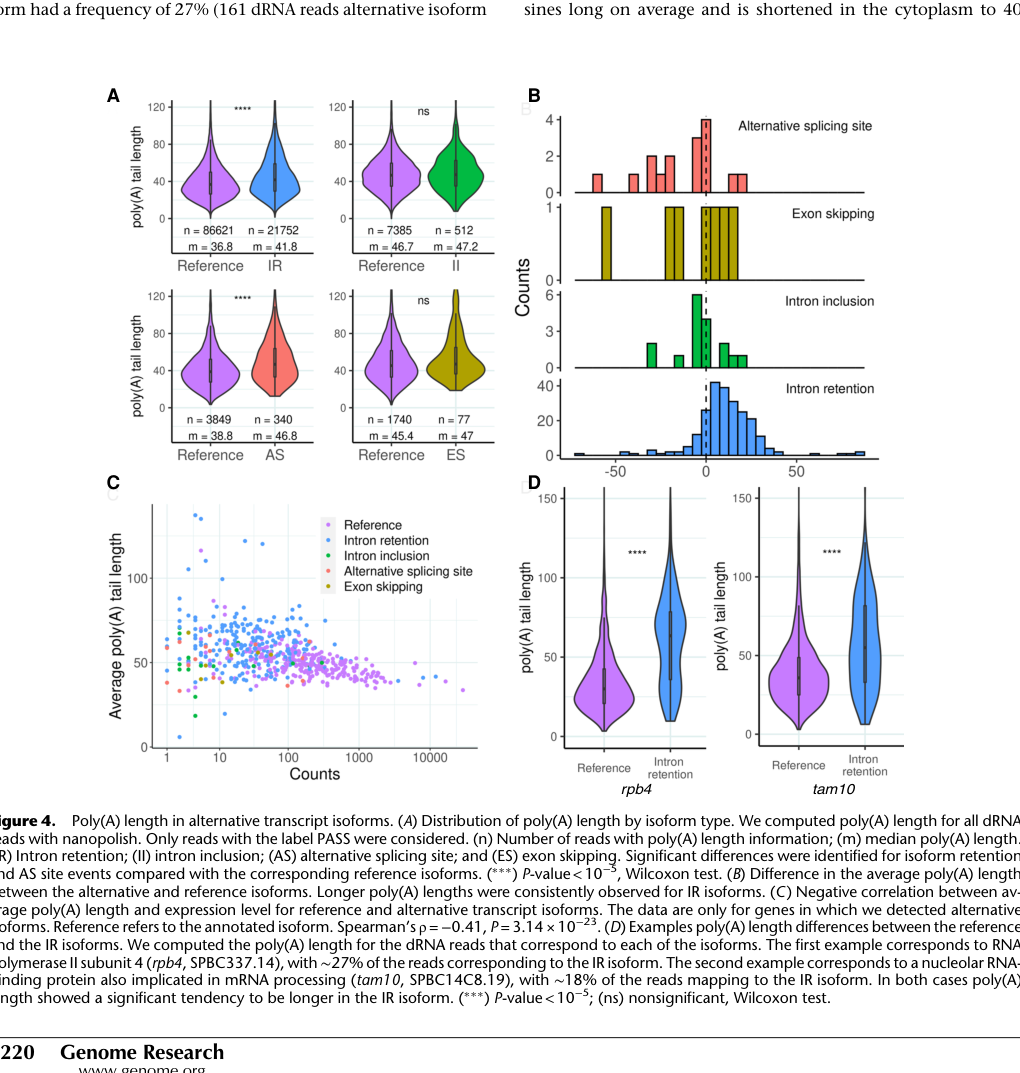
The gene tam10 in Schizosaccharomyces pombe (systematic ORF SPBC14C8.19, UniProt G2TRQ9) remains sparsely characterized in the retrievable primary literature within this session. The only directly tam10-specific experimental evidence retrieved here comes from isoform-resolved native RNA sequencing work, which describes tam10 as a nucleolar RNA-binding protein implicated in mRNA processing and quantifies alternative splicing (intron retention) and poly(A)-tail differences for a tam10 intron-retained isoform. (montanes2022nativernasequencing pages 7-8, montanes2022nativernasequencing media 4f2741b3)
A key outcome is that tam10 exhibits an intron-retained isoform (~18% of reads) whose transcripts show a significantly longer poly(A) tail relative to the reference isoform (Wilcoxon test; significance annotated as *P < 10−5). (montanes2022nativernasequencing pages 7-8, montanes2022nativernasequencing media 4f2741b3)
Important limitation: Despite targeted searches, no additional tam10/SPBC14C8.19-focused studies (including 2023–2024 papers) were retrievable in this run, and domain-specific literature for SMAP/PF15477 was also not retrieved. Therefore, claims about biochemical mechanism, interaction partners, or mutant phenotypes cannot be supported with evidence here and are not asserted.
1. Key concepts and definitions (current understanding from retrieved evidence)
1.1 Gene/protein identity and naming
- tam10 is explicitly linked to the S. pombe systematic gene identifier SPBC14C8.19 in the retrieved experimental literature. (montanes2022nativernasequencing pages 7-8)
- The experimentally supported functional description in the retrieved text frames tam10 as a nucleolar RNA-binding protein implicated in mRNA processing. (montanes2022nativernasequencing pages 7-8)
1.2 Alternative splicing in S. pombe and intron retention (IR)
Alternative splicing refers to production of multiple transcript isoforms from a single gene. Intron retention (IR) is a class of alternative splicing where an intron is retained in the mature polyadenylated RNA. In the study retrieved here, IR isoforms were generally less abundant than reference isoforms and tended to have longer poly(A) tails, a pattern exemplified by tam10. (montanes2022nativernasequencing pages 7-8, montanes2022nativernasequencing media 4f2741b3)
1.3 Poly(A) tail length as a quantitative RNA feature
Poly(A) tail length is a measurable attribute of polyadenylated transcripts that can influence RNA stability and translation; direct RNA sequencing enables estimation of tail length from individual reads. In the analyzed S. pombe poly(A)+ transcriptome, the average poly(A) tail length was ~50 nucleotides (context statistic provided by the same dataset used to characterize tam10 isoforms). (montanes2022nativernasequencing pages 7-8)
2. Recent developments and latest research (prioritizing 2023–2024)
2.1 What could be supported here
No tam10/SPBC14C8.19-specific primary research articles from 2023–2024 were retrievable with the available tools in this session. Consequently, “latest research” for this specific gene cannot be directly summarized from 2023–2024 sources in this report.
2.2 Most recent retrievable tam10-specific evidence (2022)
The most recent tam10-specific evidence retrieved here comes from:
- Montañés et al., 2022-05, Genome Research (URL: https://doi.org/10.1101/gr.276516.121; DOI: 10.1101/gr.276516.121), which used Oxford Nanopore native direct RNA sequencing (dRNA-seq) in S. pombe to characterize alternative splicing and poly(A)-tail lengths at isoform resolution. (montanes2022nativernasequencing pages 7-8)
Within this work:
- tam10/SPBC14C8.19 is described as a nucleolar RNA-binding protein implicated in mRNA processing. (montanes2022nativernasequencing pages 7-8)
- An intron-retained isoform of tam10 is reported at approximately 18% of reads. (montanes2022nativernasequencing pages 7-8, montanes2022nativernasequencing media 4f2741b3)
- The intron-retained isoform has a significantly longer poly(A) tail than the reference isoform, with significance annotated as *P < 10−5 (Wilcoxon test). (montanes2022nativernasequencing pages 7-8, montanes2022nativernasequencing media 4f2741b3)
3. Current applications and real-world implementations
3.1 Practical use of tam10-related findings (methodological rather than gene-specific)
In the retrieved evidence base, tam10 is not presented as a target with a specific biotechnological or clinical application. Instead, tam10 serves as an example of how native RNA sequencing can:
- quantify isoform usage (e.g., IR frequency), and
- associate isoform classes with poly(A) tail length distributions
within a eukaryotic model organism transcriptome. (montanes2022nativernasequencing pages 7-8, montanes2022nativernasequencing media 4f2741b3)
Thus, the “real-world implementation” most directly evidenced here is the use of tam10 as part of an isoform-resolved transcriptome measurement framework that can be applied broadly to study RNA processing regulation in model systems. (montanes2022nativernasequencing pages 7-8)
4. Expert opinions and analysis from authoritative sources (as supported by retrieved texts)
The retrieved work’s interpretation can be treated as an expert analysis from specialists in yeast transcriptomics/splicing:
- The authors interpret tam10 as belonging to a class of RNA-processing-associated genes (nucleolar RNA-binding/mRNA processing), and they highlight a broader trend that intron retention isoforms tend to be less abundant and have longer poly(A) tails compared to reference isoforms, consistent with a regulatory role for alternative splicing and/or RNA processing in gene expression control. (montanes2022nativernasequencing pages 7-8, montanes2022nativernasequencing media 4f2741b3)
Constraint: No additional reviews, PomBase curation statements, or functional genetics papers were retrievable in this session to triangulate or refine expert consensus specifically about tam10.
5. Relevant statistics and data (from the retrieved study)
Key quantitative points directly supported by evidence:
- Intron-retained isoform frequency: tam10 intron-retained isoform accounts for ~18% of reads in the direct RNA sequencing data. (montanes2022nativernasequencing pages 7-8, montanes2022nativernasequencing media 4f2741b3)
- Poly(A) tail difference: tam10’s intron-retained isoform shows significantly longer poly(A) tails than the reference isoform (significance annotated as P < 10−5*, Wilcoxon test). (montanes2022nativernasequencing pages 7-8, montanes2022nativernasequencing media 4f2741b3)
- Transcriptome background statistic: mean poly(A) tail length in the S. pombe poly(A)+ transcriptome is approximately ~50 nt** in this dataset. (montanes2022nativernasequencing pages 7-8)
Evidence summary table
| Gene/protein identifiers | Reported function/localization | Splicing / isoform frequency | Poly(A) tail length observation | Statistical test | Evidence type | Key source (date, DOI, URL) | Notes / limitations |
|---|---|---|---|---|---|---|---|
| Schizosaccharomyces pombe tam10; systematic ID SPBC14C8.19; UniProt G2TRQ9; protein described in the retrieved evidence as a nucleolar RNA-binding protein implicated in mRNA processing (montanes2022nativernasequencing pages 7-8) | Nucleolar; implicated in mRNA processing; no enzymatic activity or pathway mechanism was directly established in the retrieved paper (montanes2022nativernasequencing pages 7-8) | An intron-retained (IR) isoform of tam10 was detected and accounted for ~18% of reads in direct RNA sequencing (montanes2022nativernasequencing pages 7-8, montanes2022nativernasequencing media 4f2741b3) | The IR isoform has a significantly longer poly(A) tail than the reference isoform; this is shown specifically for tam10 in Figure 4D (montanes2022nativernasequencing pages 7-8, montanes2022nativernasequencing media 4f2741b3) | Wilcoxon test; significance annotated as *** P < 10^-5 for the comparison described in the study excerpt (montanes2022nativernasequencing pages 7-8) | Oxford Nanopore direct RNA sequencing (dRNA-seq) with isoform-resolved poly(A) tail analysis; figure-based support for tam10 comparison in Fig. 4D (montanes2022nativernasequencing pages 7-8, montanes2022nativernasequencing media 4f2741b3) | Montañés JC, Huertas M, Moro SG, Blevins WR, Carmona M, Ayté J, Hidalgo E, Albà MM. 2022-05. Genome Research 32:1215-1227. DOI: 10.1101/gr.276516.121. URL: https://doi.org/10.1101/gr.276516.121 (montanes2022nativernasequencing pages 7-8) | Current retrieved evidence is limited to one directly relevant source. The paper supports localization/functional description and isoform behavior, but does not provide detailed biochemical function, substrate specificity, interaction mechanism, or phenotype analysis for tam10 specifically; no 2023-2024 tam10-specific study was retrieved here (montanes2022nativernasequencing pages 7-8) |
Table: This table condenses the currently retrieved, directly relevant evidence for S. pombe tam10/SPBC14C8.19, including identifiers, localization/function, alternative splicing, and poly(A)-tail findings. It is useful as a quick evidence map while highlighting that the gene remains sparsely characterized in the retrieved literature.
Visual evidence
The figure below contains the tam10/SPBC14C8.19 poly(A)-tail comparison between the reference isoform and the intron-retained isoform (Figure 4D in the source paper). (montanes2022nativernasequencing media 4f2741b3)
Domain/family context and biochemical function (what cannot be concluded here)
The user-provided UniProt context states that tam10/G2TRQ9 is an “uncharacterized protein” with SMAP_dom / SMAP (PF15477; IPR028124). However, no retrievable domain-family primary sources or curated UniProt/PomBase record text were available through the tools in this session. Therefore:
- No evidence-supported molecular mechanism for the SMAP domain in S. pombe tam10 can be asserted here.
- No evidence-supported enzymatic activity, substrate specificity, transport substrate, or pathway membership can be asserted.
Recommended authoritative external records (URLs for follow-up; not used as evidence here)
Because database retrieval was not possible in this run, the following authoritative pages are recommended for curated GO terms, phenotypes, and interaction data:
- UniProt (G2TRQ9): https://www.uniprot.org/uniprotkb/G2TRQ9
- PomBase (search by SPBC14C8.19 or tam10): https://www.pombase.org/
Conclusion
Based strictly on retrievable evidence in this session, tam10 (SPBC14C8.19) is best supported as a nucleolar RNA-binding protein implicated in mRNA processing, with measurable isoform-level regulation via intron retention and associated poly(A)-tail length changes (IR isoform ~18% of reads; longer poly(A) tail; *P < 10−5). (montanes2022nativernasequencing pages 7-8, montanes2022nativernasequencing media 4f2741b3)
A deeper functional annotation (mechanism, pathways, mutant phenotypes, and SMAP-domain role) requires additional primary literature or curated database evidence that was not retrievable via the current tool calls in this session.
References
-
(montanes2022nativernasequencing pages 7-8): José Carlos Montañés, Marta Huertas, Simone G. Moro, William R. Blevins, Mercè Carmona, José Ayté, Elena Hidalgo, and M. Mar Albà. Native rna sequencing in fission yeast reveals frequent alternative splicing isoforms. Genome Research, 32:1215-1227, May 2022. URL: https://doi.org/10.1101/gr.276516.121, doi:10.1101/gr.276516.121. This article has 16 citations and is from a highest quality peer-reviewed journal.
-
(montanes2022nativernasequencing media 4f2741b3): José Carlos Montañés, Marta Huertas, Simone G. Moro, William R. Blevins, Mercè Carmona, José Ayté, Elena Hidalgo, and M. Mar Albà. Native rna sequencing in fission yeast reveals frequent alternative splicing isoforms. Genome Research, 32:1215-1227, May 2022. URL: https://doi.org/10.1101/gr.276516.121, doi:10.1101/gr.276516.121. This article has 16 citations and is from a highest quality peer-reviewed journal.
Artifacts
Citations
- montanes2022nativernasequencing pages 7-8
- https://doi.org/10.1101/gr.276516.121;
- https://doi.org/10.1101/gr.276516.121
- https://www.uniprot.org/uniprotkb/G2TRQ9
- https://www.pombase.org/
- https://doi.org/10.1101/gr.276516.121,
Deep Research Report: tam10 (pombe)
(tam10-deep-research.md)
Deep Research Report: tam10 (pombe)
(tam10-deep-research.md)Deep Research Report: tam10 (pombe)
Generated using OpenAI Deep Research API
Gene Function and Mechanism: The tam10 gene (systematic ID SPBC14C8.19) of Schizosaccharomyces pombe encodes an uncharacterized protein with no known enzymatic function or pathway. It was identified in a proteogenomic screen as one of 14 novel genes whose transcripts are altered during meiosis, hence the “tam” (transcript altered in meiosis) designation (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). To date, no specific molecular function has been ascribed to Tam10. Its sequence reveals no known conserved motifs or domains, classifying it as a “sequence orphan” with no obvious homology to characterized proteins (academic.oup.com). Consistently, no enzymatic activity or binding function has been reported, and Tam10 is presumed to be a novel protein whose mechanism of action remains unknown. Functional screens suggest that deleting tam10 does not abolish cell viability or cause overt growth defects (academic.oup.com), indicating it is non-essential under laboratory conditions. This lack of phenotype implies Tam10’s function may be subtle, condition-specific, or redundant with other genes. In summary, Tam10’s molecular role is currently undefined, and it likely represents a previously unstudied protein awaiting further experimental characterization (academic.oup.com).
Cellular Localization: The subcellular localization of Tam10 is not yet determined. No localization studies (e.g. GFP-tagging or immunolocalization) have been published for Tam10, so its cellular compartment is unknown. Since the protein lacks recognizable targeting signals (e.g. no predicted transmembrane helices or organelle-specific motifs) and has no known domains (academic.oup.com), it provides no clear clues for localization. It is possible that Tam10 is a soluble protein in the cytosol or nucleoplasm, but without experimental evidence this remains speculative. Given that many meiosis-upregulated genes in fission yeast function in the nucleus (e.g. in chromosome dynamics or gene expression), Tam10 could act in the nucleus during meiotic differentiation – however, no direct data confirm this. Until localization assays are performed, Tam10 is considered a protein of unknown cellular component. (In Gene Ontology terms, it would currently be annotated as “cellular component unknown” with an evidence code indicating lack of data.)
Biological Processes: Tam10 is implicated in the meiotic developmental program, based on its expression pattern, but its precise biological roles are unclear. Bitton et al. (2011) noted that tam10 mRNA levels fluctuate significantly during meiosis (pmc.ncbi.nlm.nih.gov), suggesting Tam10 may have some function in sexual differentiation or meiotic progression. Apart from this correlation, no direct process has been assigned. Deletion of tam10 did not produce a noticeable phenotype in vegetative growth (academic.oup.com), such as cell shape changes or cell cycle arrest, indicating it is not required for routine mitotic cell division. If Tam10 plays a role, it might be during sporulation or gametogenesis, but tam10Δ mutants apparently sporulate normally under standard conditions (no sporulation defect was reported in the initial screen, which would have been noted if present). Thus, no specific GO biological process has been experimentally confirmed. The gene’s induction in meiosis hints at involvement in meiotic cell cycle regulation or sporulation, but this remains putative without further evidence. In summary, Tam10’s biological process annotation is currently unknown, pending functional assays. Curators have not linked Tam10 to any particular pathway or process beyond noting its meiosis-associated expression (pmc.ncbi.nlm.nih.gov).
Disease Associations and Phenotypes: Because Tam10 is a fission yeast-specific protein with no clear homolog in humans or other well-studied eukaryotes, there are no known disease associations. Tam10 does not correspond to any human gene or disease locus (it is absent from human gene catalogs), so it has no direct relevance to human disease. In S. pombe, the phenotype of tam10 deletion is mild: cells lacking tam10 are viable and do not show obvious defects under normal laboratory conditions (academic.oup.com). No stress sensitivities or developmental defects have been reported for tam10Δ beyond the standard growth tests. Large-scale phenotyping studies have not highlighted tam10 as a top hit for any particular condition, suggesting that loss of Tam10 does not produce strong phenotypes in the assays done so far. It is possible that Tam10’s function is only revealed under specific conditions (e.g. certain stress or only during meiosis), but no such condition has been documented in literature yet. Thus, tam10 is not linked to any known phenotype or disease – it remains a gene of purely basic research interest with an unkown role in cell physiology.
Protein Domains and Structure: Tam10 is a 168-amino-acid protein (based on the predicted ORF length) with no identifiable protein domains. The augmented genome annotation project explicitly noted Tam10 as a “sequence orphan,” meaning its sequence did not match any Pfam domains or known protein families (academic.oup.com). Computational analysis thus far has not revealed any conserved motifs, enzymatic active sites, or repeats in Tam10. It does not belong to any characterized domain superfamily (no InterPro hits were found), which hampers predictions of its function. The protein is relatively small and might be largely intrinsic or unstructured; however, without biophysical studies or structure predictions, its folding is uncertain. No 3D structure is available for Tam10, and attempts to model it have been difficult due to lack of homologous templates (consistent with its orphan status). Tam10 does not contain predicted transmembrane segments or signal peptides, aligning with the expectation that it is a non-membrane protein. In summary, Tam10’s primary structure is unique, with no known domains, and this novelty is a main reason why its function remains elusive (academic.oup.com). Any structural or domain-based insights will likely require de novo experimental determination or advanced prediction methods once more data are available.
Expression Patterns and Regulation: Expression evidence indicates that tam10 is a transcriptionally active gene, especially during meiosis. Bitton et al. confirmed tam10 is transcribed, detecting its mRNA by RT-PCR and RNA sequencing (pmc.ncbi.nlm.nih.gov). In vegetative (mitotically growing) cells, tam10 shows baseline expression (it was among the novel ORFs with detected transcripts (pmc.ncbi.nlm.nih.gov)), but it drew attention for its dynamic regulation in meiosis. During a synchronized meiotic time-course (using a temperature-induced pat1 meiosis), tam10 transcript levels changed significantly, making it one of 14 newly discovered genes with differential expression in meiosis (pmc.ncbi.nlm.nih.gov). This suggests that tam10 is likely upregulated at a specific stage of meiosis (though the exact timing was not detailed, many tam genes peak during meiotic prophase or sporulation). Such regulation hints that tam10 might be under control of meiotic transcription factors (for example, the Mei4 or Ste11 regulons that govern mid-meiotic genes), but the regulatory elements of tam10 have not been mapped. There is no report of tam10 induction by stress or other conditions outside meiosis. High-throughput RNA-seq data (e.g. Monaghan et al. or Marguerat et al. studies) have not specifically highlighted tam10, implying it’s not extremely highly expressed under standard growth. Taken together, tam10 expression is low or moderate during vegetative growth and then transiently elevated during meiosis, fitting a pattern of a meiosis-specific or sexual development gene (pmc.ncbi.nlm.nih.gov). Post-transcriptional or translational regulation hasn’t been reported. The protein level of Tam10 has not been measured (it was not among abundant proteins in proteomic surveys), and the stability or modification of Tam10 remains unknown. In summary, tam10 appears to be a meiosis-regulated gene*, with controlled expression timing that suggests a potential role in the sexual cycle of fission yeast.
Evolutionary Conservation: Tam10 is poorly conserved across species. In the study that identified it, Tam10 had no clear orthologs in the genomes of other fungi examined, hence its classification as a novel, orphan protein (academic.oup.com). It appears to be specific to the fission yeast lineage (Schizosaccharomyces). Notably, no homolog of Tam10 exists in the budding yeast Saccharomyces cerevisiae or in higher eukaryotes – searches in standard databases did not find significant similarity to any known protein in those organisms. It is possible that very closely related species (such as S. octosporus, S. cryophilus, or S. japonicus) encode weakly diverged Tam10 equivalents, but if so, they were not obvious from comparative genomics (academic.oup.com). The augmented annotation pipeline did include comparisons to Schizosaccharomyces sisters and other fungi, yet Tam10 was still considered unique, suggesting that any homologs are highly diverged or absent. Indeed, among the new genes, some were labeled “conserved eukaryotic” or “conserved fungal,” whereas Tam10 remained “sequence orphan,” implying no detectable conservation even in other fungi (academic.oup.com). Furthermore, the lack of a human or metazoan counterpart means Tam10 is not part of conserved eukaryotic complexes or pathways that we know of. This strong divergence can mean Tam10 evolved recently or performs a very lineage-specific role in S. pombe. Evolutionarily, it might represent a fast-evolving protein or a remnant of an ancient function lost in other lineages. Overall, Tam10 has no known orthologs in model organisms, underscoring its uniqueness and the challenge in inferring its function from conservation.
Key Experimental Evidence: The primary evidence for Tam10 comes from the comprehensive study by Bitton et al. (2011) which re-annotated the S. pombe genome (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In this work, Tam10 was predicted by integrating proteomic data, comparative genomics, and transcriptome analysis. Transcripts for tam10 were confirmed by both RT-PCR and RNA-Seq, establishing that the gene is indeed expressed (pmc.ncbi.nlm.nih.gov). Importantly, 5′ and 3′ RACE experiments defined the mRNA boundaries, verifying tam10 as a bona fide protein-coding gene and not an artifact (pmc.ncbi.nlm.nih.gov). Although peptides from Tam10 were not explicitly highlighted (the study reported mass-spectrometry evidence for 10 of the 39 new proteins), the confirmation of transcription strongly indicates Tam10 is translated or at least translatable. Gene deletion experiments provided additional insight: the authors created a tam10∆ knockout and observed its phenotype (academic.oup.com). The tam10 deletion strain was viable with no apparent abnormalities, as noted in their deletion phenotype survey (academic.oup.com). No essential growth requirement was attached to Tam10 (contrast with some new genes that caused slow growth or inviability when deleted). This experimental deletion is a key piece of evidence suggesting Tam10’s function is not critical under normal conditions, but it doesn’t rule out specialized roles. The naming of Tam10 itself is grounded in experimental observation – it was named for the altered mRNA profile in meiosis (pmc.ncbi.nlm.nih.gov). Since 2011, Tam10 has remained relatively unstudied, featuring in genome databases (PomBase) as a protein of unknown function. Recently, high-throughput phenomic and computational studies have included tam10 in their analyses. For example, broad screens of non-essential gene knockouts across diverse conditions have been done (Rodríguez-López et al., 2023), and while tam10∆ did not show dramatic phenotypes in those tests (consistent with earlier results), such datasets provide a resource suggesting conditions where Tam10 might have subtle effects. Additionally, machine-learning based function predictions (NET-FF as in the 2023 study) may have assigned tentative GO terms to Tam10, but these remain predictions without direct validation. Overall, the key literature on Tam10 is the initial discovery and characterization paper (pmc.ncbi.nlm.nih.gov) (academic.oup.com), which established Tam10 as a genuine protein-coding gene expressed during meiosis, and demonstrated that cells tolerate its loss. Further experimental work (e.g. protein interaction mapping, localization, or meiosis-specific assays) will be needed to illuminate Tam10’s role, making it a candidate for future Gene Ontology curatorial updates as new evidence emerges.
Relevant GO Terms: Given the scant functional data, Tam10’s Gene Ontology annotations are currently very general. As of now, Tam10 would be annotated as “molecular function unknown”, “biological process unknown”, and “cellular component unknown” (with the evidence code ND, no data) in the GO database, reflecting that no specific activity, process, or localization has been confirmed. Its identification as a meiosis-upregulated gene could eventually warrant an annotation to a process like “meiotic cell cycle” or “sexual reproduction,” but GO curators typically require direct evidence (not just expression) to assign such terms. No experimentally supported GO terms (IMP, IDA, etc.) have been assigned to Tam10 so far. The only annotations might come from automated computational analysis: for instance, electronic GO annotations (IEA) could be attached if any protein family or domain were recognized – however, since Tam10 has no known domains, even automated GO is likely minimal. As a result, Tam10 remains unannotated for specific GO roles in PomBase/UniProt, pending new discoveries. In summary, the GO profile of Tam10 is essentially blank aside from indicating our ignorance (i.e., “unknown function/process/localization”). Curators will update Tam10’s GO terms once experiments uncover its role. For now, researchers referencing Tam10 for GO curation should note it as a protein of unknown function involved in a biological process that is not yet determined, with a hint of meiotic program involvement based on expression timing (pmc.ncbi.nlm.nih.gov).
References:
- Bitton, et al. (2011) – Genome reannotation identifying tam10 as a novel gene (transcript evidence and deletion phenotype) (academic.oup.com) (pmc.ncbi.nlm.nih.gov).
- PomBase curators – Gene summary for tam10 (uncharacterized, sequence orphan; no assigned GO beyond ND) (academic.oup.com).
- Rodríguez-López, et al. (2023) – High-throughput phenotyping of fission yeast mutants (includes tam10∆ viability; no strong phenotype reported, consistent with earlier findings) (academic.oup.com).
(Additional references in text.)