Deep Research
Cyberian
(ASCL1-deep-research-cyberian.md)
Cyberian
(ASCL1-deep-research-cyberian.md)ASCL1 (Achaete-Scute Homolog 1): A Comprehensive Functional Analysis
Introduction
ASCL1 (Achaete-Scute Family bHLH Transcription Factor 1), also known as ASH1, HASH1, or MASH1, is a basic helix-loop-helix (bHLH) transcription factor that serves as a master regulator of neuronal differentiation and neuroendocrine cell development in humans and other vertebrates. The human gene maps to chromosome 12q23.2 and encodes a 238-amino acid protein that shares 95% identity with the mouse ortholog Mash1 [ball-1993-human-cloning-summary]. As a vertebrate homolog of the Drosophila achaete-scute complex (AS-C) genes, ASCL1 exemplifies the evolutionary conservation of proneural gene function from invertebrates to mammals [guillemot-1993-knockout-mouse-summary].
The primary molecular function of ASCL1 is sequence-specific DNA binding and transcriptional activation. ASCL1 recognizes and binds to E-box consensus sequences (5'-CANNTG-3') in target gene regulatory regions, with a strong preference for the CAGCTG variant [castro-2011-progenitor-proliferation-summary][aydin-2019-neuronal-subtype-summary]. Efficient DNA binding requires heterodimerization with ubiquitously expressed E-proteins, particularly TCF3 (E12/E47), TCF4, and TCF12, through the helix-loop-helix domain. Beyond its classical transcription factor role, ASCL1 functions as a pioneer transcription factor, capable of accessing closed chromatin and initiating chromatin remodeling to enable binding of downstream factors [raposo-2015-chromatin-landscape-summary][wapinski-2013-reprogramming-hierarchy-summary]. This pioneer activity underlies ASCL1's remarkable ability to direct cell fate decisions in both developmental and reprogramming contexts.
Molecular Function and DNA Binding Specificity
The ASCL1 protein contains several functionally important domains. The basic region mediates direct DNA contact, while the helix-loop-helix domain enables dimerization with E-protein partners. Additionally, a C-terminal acidic domain and an N-terminal glutamine/alanine-rich region contribute to transcriptional activity, though the N-terminal polyalanine tract (encoded by CAG repeats) appears dispensable for neuronal differentiation [ball-1993-human-cloning-summary]. Structure-function studies have identified a nuclear localization signal and demonstrated that specific mutations within the basic region block DNA binding but not heterodimer formation with TCF3 and TCF12.
High-resolution NMR studies have provided the first atomic-level structural characterization of ASCL1, revealing that it forms an extended polypeptide chain lacking persistent tertiary interactions [toto-2017-nmr-structure-summary]. The N-terminal polyalanine stretch (13 alanines) adopts a stable α-helical conformation, while the adjacent polyglutamine region shows gradual helical destabilization and functions as a transition zone. Critically, the bHLH domain in its DNA-free state contains two transient helical segments (H1 and H2) separated by a flexible kink, with the C-terminal helix showing greater stability. This structural disorder in the absence of DNA is consistent with findings from related bHLH proteins such as MyoD and E47, where DNA binding induces a stable dimeric conformation. The C-terminal region of ASCL1 remains highly disordered and contains the multiple serine-proline phosphorylation sites that regulate its activity. No crystal structure of ASCL1 bound to DNA has been solved, but structures of related bHLH proteins (MyoD-DNA at 2.8 Å resolution) indicate that the bHLH domain forms a parallel four-helix bundle upon DNA engagement.
ASCL1 exhibits distinct E-box sequence preferences that distinguish it from other proneural factors. De novo motif analysis of ChIP-seq data revealed that the CAGCTG E-box motif is highly enriched directly beneath 74% of ASCL1 binding peaks [castro-2011-progenitor-proliferation-summary]. This "GC" core E-box preference contrasts with Neurogenin 2 (Neurog2), which preferentially binds CAGATG/CADATG motifs [aydin-2019-neuronal-subtype-summary]. E-proteins enhance ASCL1 binding to CAGSTG sequences through heterodimer formation, with the E-protein partner influencing the binding conformation at the adjacent half-site. This sequence selectivity has important biological implications: when expressed in embryonic stem cells or fibroblasts, ASCL1 and Neurog2 bind largely non-overlapping genomic sites and specify different neuronal subtypes—ASCL1 drives GABAergic and sympathetic neuronal fates while Neurog2 promotes glutamatergic and sensory neuronal identities [aydin-2019-neuronal-subtype-summary].
Genome-wide studies have mapped over 21,000 ASCL1 binding sites, predominantly at distal enhancers (approximately 63% in intergenic regions) rather than promoters (only ~7%) [raposo-2015-chromatin-landscape-summary]. This enhancer-centric binding pattern reflects ASCL1's role in establishing cell-type-specific regulatory landscapes. Direct target gene analysis identified 272 high-confidence targets organized into temporal clusters during differentiation, with early targets involved in signal transduction and later targets encoding transcription factors and cytoskeletal proteins essential for neuronal development.
Pioneer Transcription Factor Activity
One of the most significant discoveries about ASCL1 is its function as a pioneer transcription factor—a factor capable of binding nucleosomal DNA in closed chromatin and initiating chromatin remodeling. Unlike conventional transcription factors that require pre-accessible binding sites, pioneer factors can engage condensed chromatin and catalyze the transition to an open, transcriptionally permissive state [raposo-2015-chromatin-landscape-summary].
Studies of ASCL1-mediated neuronal reprogramming revealed that ASCL1 binds chromatin marked by a unique "trivalent" signature combining H3K9me3, H3K27ac, and H3K4me1 histone modifications [wapinski-2013-reprogramming-hierarchy-summary]. At these sites, ASCL1 binding precedes increases in chromatin accessibility and the appearance of new DNase hypersensitivity sites. Importantly, the chromatin accessibility at ASCL1 binding sites remains largely unchanged throughout neural stem cell differentiation, as ASCL1 targets both readily accessible and closed chromatin in proliferating cells.
Recent work has refined our understanding of ASCL1's pioneer activity, revealing that it operates through two distinct mechanisms depending on genomic context [ali-2020-mswi-snf-summary]. At approximately 23.5% of its dependent sites, ASCL1 functions as a classical pioneer factor, binding closed chromatin independently. However, at roughly 68% of sites, ASCL1 requires cooperation with the mSWI/SNF chromatin remodeling complex. Physical interactions between ASCL1 and mSWI/SNF subunits SMARCC1 and ARID1A have been confirmed by co-immunoprecipitation and proximity ligation assays in both cultured cells and human fetal cortex tissue. Notably, ASCL1 lacks intrinsic chromatin remodeling enzymatic activity and depends on recruitment of cofactors for this function.
The hierarchical mechanism of ASCL1-mediated reprogramming has been elegantly demonstrated in the direct conversion of fibroblasts to neurons. When the three factors ASCL1, BRN2, and MYT1L are co-expressed, ASCL1 rapidly occupies its cognate sites genome-wide, with binding patterns virtually identical whether ASCL1 is expressed alone or with its partners [wapinski-2013-reprogramming-hierarchy-summary]. BRN2, in contrast, cannot productively access fibroblast chromatin on its own but is recruited to ASCL1-opened sites. This establishes a clear hierarchy: ASCL1 acts first to open chromatin, enabling subsequent binding of BRN2 and activation of the full neurogenic program.
Subcellular Localization and Protein Stability
ASCL1 localizes to both the nucleus and cytoplasm of neural stem cells, with comparable distribution patterns in proliferating and differentiating conditions. Cellular fractionation studies detected ASCL1 protein at similar levels in cytoplasmic, nuclear, and chromatin-bound fractions. The protein contains a nuclear localization signal that enables translocation to the nucleus, where it carries out its transcriptional functions [ali-2020-mswi-snf-summary].
Importantly, subcellular localization modulates ASCL1 stability through compartment-specific ubiquitylation [urbanska-2022-phosphorylation-summary]. Chromatin-bound ASCL1 associates with short ubiquitin chains and exhibits greater than double the half-life of cytoplasmic ASCL1, which harbors much longer ubiquitin chains targeting it for proteasomal destruction. The E3 ubiquitin ligase HUWE1, which localizes exclusively to the cytoplasm, mediates cytoplasmic ASCL1 degradation by conjugating ubiquitin to lysines within the bHLH domain. HUWE1 knockdown leads to increased chromatin-bound ASCL1, presumably because stabilized cytoplasmic protein shuttles to the nucleus. This compartmentalized regulation provides a mechanism for fine-tuning ASCL1 activity levels.
Regulation by Phosphorylation
ASCL1 activity is regulated by multisite phosphorylation on serine-proline (SP) sites, which functions as a molecular "rheostat" controlling neurogenic potential [urbanska-2022-phosphorylation-summary]. Human ASCL1 contains five key SP sites (S93, S190, S194, S207, S223) that are phosphorylated by proline-directed serine-threonine kinases including CDK-cyclin complexes, ERK, and GSK3. The CDK2-CyclinA2 complex directly phosphorylates ASCL1 during the cell cycle, as demonstrated by mobility shift on SDS-PAGE that is reversed by phosphatase treatment.
Phosphorylation has profound functional consequences. Highly phosphorylated ASCL1 exhibits reduced DNA binding and diminished transcriptional activity. In cancer cells, maintaining ASCL1 in a phosphorylated state allows continued proliferation despite ASCL1 expression. Conversely, a phospho-mutant form of ASCL1 (with SP sites mutated to alanine, termed "5SA") shows substantially enhanced neuronal induction activity both in vitro and in vivo. This un(der)phosphorylated ASCL1 is also resistant to inhibition by Notch signaling and CDK activity [urbanska-2022-phosphorylation-summary].
Phosphorylation also primes ASCL1 for degradation. E-protein (TCF3) binding protects ASCL1 from degradation, and during mitosis, TCF3 dissociation from ASCL1 accelerates its degradation through HUWE1-mediated ubiquitylation. This cell cycle-coupled regulation ensures that ASCL1 protein levels fluctuate appropriately during the proliferation-differentiation decision.
Oscillatory Expression and Cell Fate Determination
A breakthrough in understanding ASCL1 function came from studies of its expression dynamics. Time-lapse imaging of neural progenitors revealed that ASCL1 levels oscillate with a period of 2-3 hours, driven by oscillations in the Notch pathway effector Hes1 [imayoshi-2013-oscillatory-summary]. Hes1 directly represses ASCL1 transcription, and its inherent instability creates alternating states of high and low Hes1 levels. When Hes1 is low, ASCL1 rises and activates Delta-like ligands (DLL1, DLL3), which trigger Notch signaling in neighboring cells and perpetuate the oscillatory cycle.
The critical insight is that expression dynamics—not just expression levels—determine cell fate outcomes [imayoshi-2013-oscillatory-summary][castro-2011-progenitor-proliferation-summary]. Oscillatory ASCL1 expression at lower levels maintains proliferating neural progenitors, while sustained expression at higher levels drives neuronal differentiation. Optogenetic experiments confirmed this model: artificially sustaining ASCL1 expression promoted neuronal fate determination, whereas oscillatory expression maintained the progenitor state. This resolves the apparent paradox of ASCL1 promoting both proliferation and differentiation in different contexts—the outcome depends on how ASCL1 is expressed, not simply whether it is expressed.
Role in Neural Development
ASCL1 is essential for the development of multiple neuronal populations in both the central and peripheral nervous systems. Knockout mouse studies established that Mash1-null animals survive until birth but die shortly after due to breathing and feeding defects [guillemot-1993-knockout-mouse-summary]. While the brain and spinal cord appear grossly normal, severe developmental abnormalities affect the olfactory epithelium, sympathetic ganglia, parasympathetic ganglia, and enteric ganglia. In the olfactory system, ASCL1 acts as a determination gene—Mash1-null embryos fail to produce olfactory sensory neurons because progenitors are not specified and the Notch pathway is not properly activated.
In the peripheral nervous system, ASCL1 biases neural crest cells toward the autonomic lineage rather than the sensory lineage [aydin-2019-neuronal-subtype-summary]. Loss of ASCL1 produces significantly smaller sympathetic ganglia due to impaired progenitor proliferation, though noradrenergic neurons can eventually form after a developmental delay, suggesting partial compensation by other factors. The transcription factor network governing sympathetic ganglia development involves BMP signals from the dorsal aorta that activate Phox2b, ASCL1, Hand2, and GATA factors in a coordinated program.
In the central nervous system, ASCL1's transient expression coordinates the transition from proliferating progenitors to differentiating neurons [castro-2011-progenitor-proliferation-summary]. Genome-wide target analysis revealed that ASCL1 directly controls genes involved in neural progenitor specification, cell cycle progression, neuronal differentiation, axon guidance, and synapse formation. Surprisingly, ASCL1 regulates numerous cell cycle genes including canonical regulators and oncogenic transcription factors, establishing that it is required for normal neural progenitor proliferation—an unexpected function for a proneural gene traditionally associated with cell cycle exit.
In the enteric nervous system (ENS), which controls gut motility and function, ASCL1 plays a critical role in specifying particular neuronal subtypes [memic-2016-ens-subtypes-summary]. All enteric neuronal subtypes and enteric glia appear to derive from ASCL1-expressing progenitors. In Ascl1-knockout mice, neural crest-derived cells colonize the bowel but develop into a sparse and abnormal ganglionic network. Specific neuronal populations are differentially affected: neurons expressing calbindin, tyrosine hydroxylase (TH), and vasoactive intestinal peptide (VIP) are selectively decreased, while serotonergic neurons form in normal numbers. Notably, esophageal neurons fail to form entirely. ASCL1 operates within a transcription factor network involving SOX10 and PHOX2B—SOX10 initially induces both PHOX2B and ASCL1 expression, and then ASCL1 suppresses SOX10 to commit cells toward neuronal rather than glial fate. These findings explain why ASCL1 mutations have been identified in patients with Ondine's curse (congenital central hypoventilation syndrome with Hirschsprung disease), highlighting the gene's importance for proper ENS development.
Notch Signaling Integration
ASCL1 is a central node in the Notch signaling pathway, which orchestrates lateral inhibition during neurogenesis. ASCL1 activates expression of Notch ligands DLL1 and DLL3, which bind Notch receptors on neighboring cells [borromeo-2016-sclc-heterogeneity-summary]. Notch activation releases the intracellular domain (NICD), which together with RBPJ upregulates Hes1, Hes5, and Hes6. These Hes proteins then directly repress ASCL1 and other proneural bHLH factors, completing the negative feedback loop.
Through this lateral inhibition mechanism, cells expressing high ASCL1 inhibit their neighbors from differentiating, maintaining the progenitor pool while allowing a subset of cells to commit to neuronal fate. The balance between ASCL1 and Hes factors determines whether cells remain as progenitors or differentiate. Genetic disruption of this balance—for example, by deleting Notch1, RBPJ, or Hes genes—results in premature ASCL1 expression and precocious neuronal differentiation.
ASCL1 also regulates DLL3, which differs from other Delta ligands in that it acts cell-autonomously rather than in trans [borromeo-2016-sclc-heterogeneity-summary]. DLL3 has emerged as a therapeutic target in SCLC precisely because of its specific regulation by ASCL1 in neuroendocrine contexts.
ASCL1 in Cancer: Focus on Small Cell Lung Cancer
ASCL1 is aberrantly expressed in several neuroendocrine cancers, most prominently small cell lung cancer (SCLC), where it functions as a lineage oncogene [augustyn-2014-lineage-oncogene-summary][borromeo-2016-sclc-heterogeneity-summary]. SCLC is an aggressive malignancy accounting for approximately 15% of lung cancers, and approximately 75% of SCLC tumors express high levels of ASCL1. This expression is not merely a marker but is functionally essential—ASCL1 knockdown induces apoptosis specifically in ASCL1-positive cancer lines while sparing ASCL1-negative cells.
Molecular subtyping has classified SCLC into four main categories based on transcription factor expression: ASCL1-high (SCLC-A, ~75%), NEUROD1-high (SCLC-N, ~15%), POU2F3-high (SCLC-P), and YAP1-high (SCLC-Y) [borromeo-2016-sclc-heterogeneity-summary]. The ASCL1 and NEUROD1 subtypes exhibit neuroendocrine features, while the others do not. ChIP-seq studies revealed that ASCL1 and NEUROD1 occupy largely non-overlapping genomic sites, with only 304 of approximately 10,000 sites shared. ASCL1 directly targets known SCLC oncogenes including MYCL1, RET, SOX2, and NFIB, while NEUROD1 targets MYC.
Mouse genetic models confirmed that only ASCL1 (not NEUROD1) is required for SCLC tumor formation [borromeo-2016-sclc-heterogeneity-summary]. ASCL1 is present in normal pulmonary neuroendocrine cells, the cell of origin for SCLC, while NEUROD1 is absent. This establishes ASCL1 as the essential lineage-defining factor for the majority of SCLC cases.
The oncogenic function of ASCL1 involves maintaining "lineage survival" through regulation of anti-apoptotic programs [augustyn-2014-lineage-oncogene-summary]. A key target is BCL2, which contains conserved E-box binding sites occupied by ASCL1 in SCLC cells. ASCL1 knockdown reduces BCL2 expression, while BCL2 inhibitor treatment shows 10-30 fold greater efficacy in ASCL1-positive versus ASCL1-negative tumors. A 72-gene ASCL1 signature derived from ChIP-seq analysis predicts poor prognosis in resected NSCLC specimens and includes 24 potentially druggable targets, providing a molecular framework for targeted therapy development.
ASCL1 in Neuroblastoma
Neuroblastoma, a pediatric cancer of the sympathetic nervous system, provides another context where ASCL1's dual proliferation-differentiation functions become dysregulated. Neuroblastoma is believed to arise from sympathetic neuroblast precursors that fail to engage the neuronal differentiation program, becoming locked in a pro-proliferative developmental state [wang-2019-neuroblastoma-crc-summary]. ASCL1 is a member of the core regulatory circuitry (CRC) that defines the adrenergic (ADRN) subtype of neuroblastoma, alongside PHOX2A, PHOX2B, HAND2, and GATA3. These transcription factors form an interconnected autoregulatory loop that maintains neuroblast identity.
In neuroblastoma, ASCL1 expression is aberrantly maintained and the protein is largely phosphorylated on multiple serine-proline sites. This phosphorylation state prevents terminal differentiation while allowing continued proliferation. MYCN, a key oncogenic driver in aggressive neuroblastoma, directly regulates ASCL1 expression, and LMO1 acts as a coregulator in this circuitry [wang-2019-neuroblastoma-crc-summary]. CRISPR-mediated deletion of ASCL1 in neuroblastoma cells results in slower growth, confirming that ASCL1 contributes to the rapid proliferation of both MYCN-amplified and non-amplified tumors.
Importantly, preventing CDK-dependent phosphorylation of ASCL1 in neuroblastoma cells drives coordinated suppression of the MYC-driven core circuit while simultaneously activating a differentiation program leading to mitotic exit. Recent studies using cell cycle phase-specific analysis revealed that ASCL1 binds different genomic targets depending on cell cycle stage: in S/G2/M phases, ASCL1 binds promoters of pro-mitotic genes, while in G1 phase, ASCL1 primes pro-neuronal enhancer loci. Prolonged G1 arrest is required to fully activate ASCL1-bound neuronal enhancers and drive differentiation. These findings suggest that CDK4/6 inhibitors, which extend G1 phase, may be therapeutic in neuroblastoma by allowing ASCL1 to engage its pro-differentiation rather than pro-proliferation program.
Cellular Reprogramming Applications
The discovery that ASCL1, together with BRN2 and MYT1L, can directly convert fibroblasts to functional neurons without passing through a pluripotent state opened new avenues in regenerative medicine [vierbuchen-2010-neuronal-conversion-summary]. These induced neuronal (iN) cells express neuron-specific proteins, generate action potentials, and form functional synapses. Subsequent studies showed that ASCL1 alone is sufficient to induce immature iN cells, establishing it as the key reprogramming driver, with BRN2 and MYT1L enhancing efficiency and maturation.
The ability of ASCL1 to reprogram diverse cell types—including fibroblasts, hepatocytes, astrocytes, and lymphoid cells—reflects its pioneer factor activity and capacity to access closed chromatin in non-neural contexts [wapinski-2013-reprogramming-hierarchy-summary]. However, the efficiency varies dramatically between cell types, correlating with the presence of the trivalent chromatin signature at neurogenic loci. Cells lacking this signature are refractory to ASCL1-mediated conversion.
Therapeutic applications of ASCL1-mediated reprogramming are being explored for neurodegenerative diseases. In glioblastoma, high ASCL1 expression identifies cells with latent neuronal differentiation capacity that can be induced to terminally differentiate by Notch inhibition, potentially reducing tumorigenicity. In the retina, ASCL1 expression in Muller glia promotes regeneration following injury, though this capacity is modulated by STAT signaling and developmental context.
Clinical Associations and Disease
Beyond cancer, ASCL1 variants have been associated with congenital central hypoventilation syndrome (CCHS) and Haddad syndrome (CCHS combined with Hirschsprung disease), though the primary causative gene for these disorders is PHOX2B. Three ASCL1 variants initially reported in CCHS patients—including polyalanine tract contractions—have been reclassified as variants of uncertain significance because polyalanine contractions are not established pathogenic mechanisms and patients typically also carried PHOX2B mutations.
ASCL1 has diagnostic utility as a marker for neuroendocrine tumors. It is highly expressed in medullary thyroid cancer, small cell lung cancer, and other neuroendocrine carcinomas, and can help distinguish esthesioneuroblastoma from other sinonasal tumors. Expression in normal adult tissues is minimal, restricted primarily to scattered neuroendocrine cells.
Open Questions
Several important questions about ASCL1 biology remain unresolved:
-
Chromatin Context Determinants: What factors determine whether ASCL1 can access and remodel chromatin at specific genomic sites? The trivalent signature provides a partial answer, but the molecular basis for cell-type-specific permissiveness remains incompletely understood.
-
Therapeutic Targeting: Can ASCL1 be directly targeted for cancer therapy? As a transcription factor, ASCL1 lacks conventional drug binding pockets. Current approaches focus on downstream targets (BCL2, DLL3) or upstream regulation (CDK inhibitors to modulate phosphorylation status).
-
Subtype Switching: What determines whether SCLC tumors express ASCL1 or NEUROD1? Evidence suggests that tumors can switch subtypes during progression or treatment, with implications for therapeutic response.
-
Reprogramming Efficiency: Why does ASCL1-mediated reprogramming work efficiently in some cell types but not others? Understanding the barriers could enable therapeutic reprogramming strategies for neurological diseases.
-
Oscillation Control: How is the period and amplitude of ASCL1 oscillations regulated, and can these parameters be manipulated to control neural stem cell behavior?
-
Post-translational Modifications: Beyond phosphorylation and ubiquitylation, what other modifications regulate ASCL1 function? The short ubiquitin chains on chromatin-bound ASCL1 suggest non-degradative ubiquitin functions that warrant investigation.
References
-
[ball-1993-human-cloning-summary] Ball DW, et al. Identification of a human achaete-scute homolog highly expressed in neuroendocrine tumors. Proc Natl Acad Sci USA. 1993;90(12):5648-52. PMID: 8390674. DOI: 10.1073/pnas.90.12.5648
-
[guillemot-1993-knockout-mouse-summary] Guillemot F, et al. Mammalian achaete-scute homolog 1 is required for the early development of olfactory and autonomic neurons. Cell. 1993;75(3):463-76. PMID: 8221886. DOI: 10.1016/0092-8674(93)90381-y
-
[vierbuchen-2010-neuronal-conversion-summary] Vierbuchen T, et al. Direct conversion of fibroblasts to functional neurons by defined factors. Nature. 2010;463(7284):1035-41. PMID: 20107439. DOI: 10.1038/nature08797
-
[castro-2011-progenitor-proliferation-summary] Castro DS, et al. A novel function of the proneural factor Ascl1 in progenitor proliferation identified by genome-wide characterization of its targets. Genes Dev. 2011;25(9):930-45. PMID: 21536733. PMC: PMC3084027. DOI: 10.1101/gad.627811
-
[wapinski-2013-reprogramming-hierarchy-summary] Wapinski OL, et al. Hierarchical mechanisms for direct reprogramming of fibroblasts to neurons. Cell. 2013;155(3):621-35. PMID: 24243019. PMC: PMC3871197. DOI: 10.1016/j.cell.2013.09.028
-
[imayoshi-2013-oscillatory-summary] Imayoshi I, et al. Oscillatory control of factors determining multipotency and fate in mouse neural progenitors. Science. 2013;342(6163):1203-8. PMID: 24179156. DOI: 10.1126/science.1242366
-
[augustyn-2014-lineage-oncogene-summary] Augustyn A, et al. ASCL1 is a lineage oncogene providing therapeutic targets for high-grade neuroendocrine lung cancers. Proc Natl Acad Sci USA. 2014;111(41):14788-93. PMID: 25267614. PMC: PMC4205603. DOI: 10.1073/pnas.1410419111
-
[raposo-2015-chromatin-landscape-summary] Raposo AASF, et al. Ascl1 Coordinately Regulates Gene Expression and the Chromatin Landscape during Neurogenesis. Cell Rep. 2015;10(10):1544-56. PMID: 25753420. PMC: PMC5383937. DOI: 10.1016/j.celrep.2015.02.025
-
[borromeo-2016-sclc-heterogeneity-summary] Borromeo MD, et al. ASCL1 and NEUROD1 Reveal Heterogeneity in Pulmonary Neuroendocrine Tumors and Regulate Distinct Genetic Programs. Cell Rep. 2016;16(5):1259-1272. PMID: 27452466. PMC: PMC4972690. DOI: 10.1016/j.celrep.2016.06.081
-
[aydin-2019-neuronal-subtype-summary] Aydin B, et al. Proneural factors Ascl1 and Neurog2 contribute to neuronal subtype identities by establishing distinct chromatin landscapes. Nat Neurosci. 2019;22(6):897-908. PMID: 31086315. PMC: PMC6556771. DOI: 10.1038/s41593-019-0399-y
-
[urbanska-2022-phosphorylation-summary] Related papers: Ali F, et al. The phosphorylation status of Ascl1 is a key determinant of neuronal differentiation. Development. 2014;141(11):2216-24. PMID: 24821983; Urbanska M, et al. Distinct Regulation of ASCL1 by the Cell Cycle and Chemotherapy in Small Cell Lung Cancer. Mol Cancer Res. 2024;22(7):613-628. PMC: PMC11217739
-
[ali-2020-mswi-snf-summary] Ali FR, et al. Pioneer factor ASCL1 cooperates with the mSWI/SNF complex at distal regulatory elements to regulate human neural differentiation. Genes Dev. 2023;37(5-6):218-242. PMID: 36931659. PMC: PMC10111863. DOI: 10.1101/gad.350269.122
-
[toto-2017-nmr-structure-summary] Toto A, et al. Fragment-Based NMR Study of the Conformational Dynamics in the bHLH Transcription Factor Ascl1. Biophys J. 2017;112(1):48-54. PMID: 28076814. PMC: PMC5390047. DOI: 10.1016/j.bpj.2016.11.3196
-
[memic-2016-ens-subtypes-summary] Memic F, et al. Ascl1 Is Required for the Development of Specific Neuronal Subtypes in the Enteric Nervous System. J Neurosci. 2016;36(15):4339-50. PMID: 27076431. PMC: PMC6601778. DOI: 10.1523/JNEUROSCI.0048-16.2016
-
[wang-2019-neuroblastoma-crc-summary] Wang L, et al. ASCL1 is a MYCN- and LMO1-dependent member of the adrenergic neuroblastoma core regulatory circuitry. Nat Commun. 2019;10(1):5622. PMID: 31811134. PMC: PMC6898709. DOI: 10.1038/s41467-019-13515-5
-
NCBI Gene Entry: https://www.ncbi.nlm.nih.gov/gene/429
- UniProt: P50553 (https://www.uniprot.org/uniprotkb/P50553)
- OMIM: 100790 (https://omim.org/entry/100790)
Citations
- ali-2020-mswi-snf-summary.md
- augustyn-2014-lineage-oncogene-summary.md
- aydin-2019-neuronal-subtype-summary.md
- ball-1993-human-cloning-summary.md
- borromeo-2016-sclc-heterogeneity-summary.md
- castro-2011-progenitor-proliferation-summary.md
- guillemot-1993-knockout-mouse-summary.md
- imayoshi-2013-oscillatory-summary.md
- memic-2016-ens-subtypes-summary.md
- raposo-2015-chromatin-landscape-summary.md
- toto-2017-nmr-structure-summary.md
- urbanska-2022-phosphorylation-summary.md
- vierbuchen-2010-neuronal-conversion-summary.md
- wang-2019-neuroblastoma-crc-summary.md
- wapinski-2013-reprogramming-hierarchy-summary.md
Falcon
(ASCL1-deep-research-falcon.md)
Falcon
(ASCL1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Plan overview and identity verification
- Identity check: The target is human ASCL1 (achaete-scute family bHLH transcription factor 1), UniProt P50553, a nuclear basic helix–loop–helix (bHLH) transcription factor also known as MASH1/hASH1. Literature and recent studies consistently describe ASCL1 as a proneural bHLH factor with nuclear localization and canonical bHLH DNA-binding/HLH dimerization domains, matching the UniProt domain annotations (bHLH_dom; HLH_DNA-bd superfamily). These features and nomenclature align with the human protein specified by the user (https://doi.org/10.1158/1541-7786.mcr-23-0405, Mar 2024; https://doi.org/10.1097/cm9.0000000000002693, Sep 2024) (liu2024distinctregulationof pages 1-2, guo2024molecularclassificationof pages 1-2).
Key concepts and definitions with current understanding
- Molecular identity and cellular localization: ASCL1 is a class A bHLH transcription factor that binds E-box motifs (CANNTG) as a heterodimer with E-proteins (e.g., TCF3/E12/E47). It functions in the nucleus to drive neuronal and neuroendocrine gene programs (https://doi.org/10.1158/1541-7786.mcr-23-0405) (liu2024distinctregulationof pages 1-2).
- Molecular function and pioneer-like activity: ASCL1 exhibits pioneer-factor properties, binding nucleosome-occupied DNA and promoting chromatin opening at neuronal enhancers to initiate lineage programs. Its binding is enriched at E-box–dense enhancers, and E-protein association facilitates nucleosomal engagement (https://doi.org/10.1098/rsob.250018, Jun 2025) (lundiebrown2025cellfateacquisition pages 9-11, lundiebrown2025cellfateacquisition pages 11-12).
- Direct transcriptional targets and pathway positioning: ASCL1 directly activates neuroendocrine/neural effectors including INSM1, MYT1, and multiple neuroendocrine markers, and it transcriptionally upregulates Notch-modulatory ligands such as DLL1/DLL3. This places ASCL1 upstream of Notch repression in neuroendocrine lineages, where DLL3 acts as an inhibitory Notch ligand (https://doi.org/10.1172/jci175217, Jul 2024; https://doi.org/10.1158/2767-9764.crc-24-0501, Feb 2025) (ku2024notchsignalingsuppresses pages 1-2, lozada2025expressionpatternsof pages 1-2). Additional ASCL1 targets frequently cited in SCLC include CHGA, CALCB, and RET (OpenTargets summary of curated evidence) (OpenTargets Search: Small cell lung cancer,Neuroblastoma-ASCL1).
- Biological roles: In development, ASCL1 is a proneural factor required for neuronal differentiation and neuroendocrine lineage specification. In cancer, it is a lineage-defining oncoprotein that maintains neuroendocrine identity and survival programs, particularly in the ASCL1-dominant subtype of small cell lung cancer (SCLC-A) (https://doi.org/10.1097/cm9.0000000000002693) (guo2024molecularclassificationof pages 1-2).
Recent developments and latest research (prioritize 2023–2024)
- Post-translational and cell-cycle regulation (2024): In SCLC, ASCL1 protein abundance is dynamically regulated by the cell cycle. CDK2–CyclinA2 phosphorylates ASCL1, promoting proteasome-mediated degradation during mitosis via the E3 ligase HUWE1; interaction with the E-protein TCF3 protects ASCL1 from degradation, and chemotherapy decreases ASCL1 transcription while ASCL1 depletion sensitizes cells to chemotherapy (https://doi.org/10.1158/1541-7786.mcr-23-0405, Mar 2024) (liu2024distinctregulationof pages 1-2). Broader mechanistic syntheses similarly emphasize HUWE1-mediated ubiquitination and E-protein–dependent stabilization as central regulatory nodes (Molecules review, 2025) (huang2025molecularsubtypesand pages 6-7).
- Oncogenic lineage role and subtype prevalence (2024): Molecular classification work consolidates ASCL1 as the dominant lineage-defining TF in SCLC-A, with reported prevalence around 70–75% of tumors. This classification corresponds to distinct biology, microenvironment features, and therapeutic vulnerabilities (https://doi.org/10.1097/cm9.0000000000002693, Sep 2024) (guo2024molecularclassificationof pages 1-2). OpenTargets curation likewise highlights subtype-specific dependency on ASCL1 in SCLC models (OpenTargets Search: Small cell lung cancer,Neuroblastoma-ASCL1).
- ASCL1–Notch axis across neuroendocrine cancers (2024): In advanced prostate cancer, activation of Notch suppresses neuroendocrine differentiation; DLL3 is an inhibitory Notch ligand transcriptionally activated by ASCL1, reinforcing the conserved ASCL1–DLL3–Notch axis in neuroendocrine states (https://doi.org/10.1172/jci175217, Jul 2024) (ku2024notchsignalingsuppresses pages 1-2).
- Drug tolerance and plasticity (2024): In EGFR-mutant lung adenocarcinoma PDXs, osimertinib treatment can induce ASCL1 upregulation in residual disease, where ASCL1 drives an EMT-like program conferring drug tolerance in permissive cellular contexts. This connects ASCL1 to therapy-induced lineage plasticity beyond classical SCLC (https://doi.org/10.1158/0008-5472.can-23-0438, Feb 2024) (hu2024ascl1drivestolerance pages 1-3).
- Tumor evolution under therapy (2024): Longitudinal multi-region sequencing in SCLC reveals branched evolution and relapse from ancestral clones under therapy. While genomic states dominated the analysis, these data contextualize how lineage programs (including ASCL1-driven states) are embedded within evolving clonal architectures (https://doi.org/10.1038/s41586-024-07177-7, Mar 2024) (ku2024notchsignalingsuppresses pages 1-2).
- Multi-omics trajectories (2024): Recent multi-omics analyses of neuroendocrine trans-differentiation identify ASCL1 as a master regulator with bifurcating lineage endpoints and reciprocal relationships with other lineage TFs (e.g., ASCL2/POU2F3), consistent with ASCL1’s role in shaping neuroendocrine states (2024 study) (chen2024multiomicstranscriptionalprofiling pages 74-78).
Current applications and real-world implementations
- Subtype classification and biomarker use: ASCL1 immunohistochemistry/RNA profiling is used to classify SCLC into lineage subtypes; ASCL1-high (SCLC-A) tumors display neuroendocrine programs and distinct vulnerabilities. Estimates place SCLC-A at ~70–75% of SCLC, with NEUROD1, POU2F3, and immune-inflamed subtypes comprising the remainder (https://doi.org/10.1097/cm9.0000000000002693) (guo2024molecularclassificationof pages 1-2).
- DLL3-targeted therapies linked to ASCL1 programs: DLL3, commonly activated by ASCL1 and serving as a Notch inhibitor, is widely expressed in neuroendocrine malignancies. Tarlatamab, a DLL3×CD3 bispecific T-cell engager (BiTE), achieved objective responses around 40% in previously treated SCLC in phase II and received FDA approval in 2024 for chemotherapy-refractory extensive-stage SCLC, illustrating rapid clinical translation of ASCL1→DLL3 biology (Cancer Research Communications, 2025; Frontiers in Immunology, 2025; see also JCI 2024 for ASCL1→DLL3 regulation) (https://doi.org/10.1158/2767-9764.crc-24-0501; https://doi.org/10.3389/fimmu.2025.1592291; https://doi.org/10.1172/jci175217) (lozada2025expressionpatternsof pages 2-3, lozada2025expressionpatternsof pages 1-2, ji2025harnessingdeltalikeligand pages 1-2, ku2024notchsignalingsuppresses pages 1-2, ji2025harnessingdeltalikeligand pages 5-6).
- Broader modalities: DLL3-targeted strategies under investigation include next-generation bispecifics (e.g., HPN328), CAR T/NK constructs, radioimmunoconjugates, and photoimmunotherapy, with early clinical signals of activity and evolving safety profiles (https://doi.org/10.3389/fimmu.2025.1592291) (ji2025harnessingdeltalikeligand pages 5-6). Historical DLL3 ADCs (rovalpituzumab tesirine) showed response enrichment in DLL3-high tumors but were discontinued due to toxicity and inferior survival in later studies, informing current modality choices (https://doi.org/10.1158/2767-9764.crc-24-0501) (lozada2025expressionpatternsof pages 6-7).
Expert opinions and analysis from authoritative sources
- Nature/clinical oncology synthesis: A 2024 Nature Reviews Clinical Oncology perspective emphasizes the centrality of lineage-defining TFs (ASCL1, NEUROD1, POU2F3) in SCLC biology, the existence of plasticity between subtypes, and the therapeutic implications for patient stratification and resistance management (https://doi.org/10.1038/s41571-024-00914-x, Jul 2024) (huang2025molecularsubtypesand pages 6-7). The Nature 2024 tumor-evolution study further underscores the complexity of relapse and the need to integrate lineage programs with clonal dynamics to optimize therapy (https://doi.org/10.1038/s41586-024-07177-7, Mar 2024) (ku2024notchsignalingsuppresses pages 1-2).
- Mechanistic regulation consensus: Experimental work (MCR 2024) and synthesis (Open Biology 2025) converge on a model where ASCL1’s function is tuned by multi-site phosphorylation (CDK/ERK pathways), ubiquitin-mediated turnover (HUWE1), chromatin engagement that is stabilized by E-proteins (TCF3), and feedback with Notch signaling via DLL ligands (https://doi.org/10.1158/1541-7786.mcr-23-0405; https://doi.org/10.1098/rsob.250018) (liu2024distinctregulationof pages 1-2, lundiebrown2025cellfateacquisition pages 11-12).
Relevant statistics and recent quantitative data
- Prevalence of ASCL1-high SCLC: Approximately 70–75% of SCLC classified as SCLC-A/ASCL1-dominant, based on recent clinical-molecular reviews (Sep 2024) (https://doi.org/10.1097/cm9.0000000000002693) (guo2024molecularclassificationof pages 1-2). OpenTargets curation corroborates high frequency of ASCL1 dependency in SCLC models (OpenTargets Search: Small cell lung cancer,Neuroblastoma-ASCL1).
- DLL3 prevalence: DLL3 overexpression in neuroendocrine tumors is common (~70–80% in SCLC); high DLL3 correlates with aggressive histology and adverse outcomes in multiple NEN sites, supporting biomarker-driven selection (Cancer Research Communications 2025; Frontiers in Immunology 2025) (https://doi.org/10.1158/2767-9764.crc-24-0501; https://doi.org/10.3389/fimmu.2025.1592291) (lozada2025expressionpatternsof pages 1-2, lozada2025expressionpatternsof pages 2-3, ji2025harnessingdeltalikeligand pages 1-2).
- SCLC survival context: SCLC remains highly lethal with very low 5-year overall survival (often <7%), underscoring the need for subtype- and biomarker-driven therapeutics such as DLL3-directed agents (Frontiers in Immunology 2025) (https://doi.org/10.3389/fimmu.2025.1592291) (ji2025harnessingdeltalikeligand pages 5-6).
- Clinical activity of DLL3 BiTE: Tarlatamab has shown approximately 40% objective response rates in previously treated SCLC in phase II, forming the basis for regulatory approval and ongoing integration into practice (CRC 2025) (https://doi.org/10.1158/2767-9764.crc-24-0501) (lozada2025expressionpatternsof pages 2-3).
Mechanistic details: pathways, targets, and regulation
- Pathways and targets: ASCL1 directly activates neuroendocrine transcriptional modules (e.g., INSM1, MYT1) and induces DLL1/DLL3, which modulate Notch signaling to support neuroendocrine differentiation; ASCL1 also co-regulates programs with other TFs in pulmonary neuroendocrine cells and SCLC-A (e.g., NKX2-1, PROX1 reported in curated compendia) (JCI 2024; OpenTargets) (https://doi.org/10.1172/jci175217) (ku2024notchsignalingsuppresses pages 1-2, OpenTargets Search: Small cell lung cancer,Neuroblastoma-ASCL1).
- Post-translational control: CDK2–CyclinA2-mediated phosphorylation and HUWE1-dependent ubiquitination regulate ASCL1 turnover during mitosis; E-protein (TCF3) association protects ASCL1 from degradation and coordinates DNA binding. Chemotherapy downregulates ASCL1 transcription, and genetic depletion enhances chemosensitivity in SCLC models (MCR 2024) (https://doi.org/10.1158/1541-7786.mcr-23-0405) (liu2024distinctregulationof pages 1-2). Phosphorylation status modulates chromatin opening and reprogramming potency, linking ASCL1 function to kinase signaling (Open Biology 2025) (https://doi.org/10.1098/rsob.250018) (lundiebrown2025cellfateacquisition pages 11-12).
- Chromatin engagement: ASCL1 binds nucleosomal DNA and remodels chromatin to pioneer neuronal/neuroendocrine enhancers, with E-proteins enhancing nucleosome affinity and occupancy at E-box-rich regions (Open Biology 2025) (https://doi.org/10.1098/rsob.250018) (lundiebrown2025cellfateacquisition pages 9-11, lundiebrown2025cellfateacquisition pages 11-12).
Clinical and translational implications
- Diagnostic and stratification use: ASCL1 expression supports SCLC molecular subtyping (SCLC-A), which correlates with neuroendocrine marker expression (INSM1, SYP, CHGA) and potential therapeutic vulnerabilities. Subtype-aware management is increasingly incorporated in trials and translational pipelines (https://doi.org/10.1097/cm9.0000000000002693) (guo2024molecularclassificationof pages 1-2).
- DLL3 as a therapeutic conduit of ASCL1 programs: ASCL1-driven DLL3 expression provides a tumor-restricted target across neuroendocrine malignancies. Tarlatamab’s 2024 approval in refractory SCLC and observed ~40% response rate in trials exemplify biomarker-guided immunotherapy based on ASCL1 biology. Alternative platforms (HPN328 TriTAC, CAR T/NK, RIT) are being refined to optimize efficacy/safety and address resistance mechanisms such as DLL3 loss (https://doi.org/10.1158/2767-9764.crc-24-0501; https://doi.org/10.3389/fimmu.2025.1592291) (lozada2025expressionpatternsof pages 2-3, lozada2025expressionpatternsof pages 6-7, ji2025harnessingdeltalikeligand pages 5-6).
- Therapy resistance/plasticity: ASCL1-driven gene programs can contribute to drug tolerance and lineage plasticity under targeted therapy pressure (osimertinib in EGFR-mutant tumors), suggesting that intercepting ASCL1 function or its downstream programs may mitigate residual disease (https://doi.org/10.1158/0008-5472.can-23-0438) (hu2024ascl1drivestolerance pages 1-3).
Structured summary
| Aspect | Key points (1–2 sentences) | Recent sources (Year; journal; DOI/URL) |
|---|---|---|
| Identity / domain & localization | Human ASCL1 (UniProt P50553) is a nuclear basic helix–loop–helix (bHLH) proneural transcription factor with canonical bHLH/MASH1 family features and nuclear localization. | Liu 2024; Molecular Cancer Research; https://doi.org/10.1158/1541-7786.mcr-23-0405 (Mar 2024) (liu2024distinctregulationof pages 1-2), Guo 2024; Chinese Medical Journal; https://doi.org/10.1097/cm9.0000000000002693 (Sep 2024) (guo2024molecularclassificationof pages 1-2), Open Targets summary (OpenTargets Search: Small cell lung cancer,Neuroblastoma-ASCL1) |
| Molecular function & pioneer activity | ASCL1 functions as a proneural bHLH transcription factor and exhibits pioneer-like chromatin engagement: binds E-box motifs, accesses nucleosomal DNA, and can open chromatin to activate neuronal programs. | Lundie-Brown 2025; Open Biology; https://doi.org/10.1098/rsob.250018 (lundiebrown2025cellfateacquisition pages 9-11, lundiebrown2025cellfateacquisition pages 11-12) |
| Direct targets & pathways (INSM1, MYT1, DLL1/DLL3; Notch axis) | ASCL1 activates neuroendocrine/neural programs including INSM1 and MYT1 and drives expression of Notch-modulatory ligands (DLL1, DLL3), placing ASCL1 upstream of Notch repression in NE lineages. | Ku 2024; J Clin Invest; https://doi.org/10.1172/jci175217 (Jul 2024) (ku2024notchsignalingsuppresses pages 1-2), Chen 2024; multi-omics SCLC study (chen2024multiomicstranscriptionalprofiling pages 74-78), Open Targets summary (OpenTargets Search: Small cell lung cancer,Neuroblastoma-ASCL1) |
| Regulation (cell cycle, phosphorylation, ubiquitination, E-proteins) | ASCL1 is cell-cycle regulated: CDK2–CyclinA2 phosphorylation and HUWE1-mediated ubiquitination control mitotic degradation; heterodimerization with E-proteins (e.g., TCF3) stabilizes DNA-bound ASCL1. | Liu 2024; Molecular Cancer Research; https://doi.org/10.1158/1541-7786.mcr-23-0405 (liu2024distinctregulationof pages 1-2), Lundie-Brown 2025; Open Biology (lundiebrown2025cellfateacquisition pages 11-12) |
| Role in SCLC (subtypes, prevalence, dependency) | ASCL1 defines the SCLC-A (neuroendocrine) lineage and is required for tumor initiation/survival in ASCL1-high tumors; reported prevalence for ASCL1-dominant SCLC is ~70–75%. | Guo 2024; Chinese Medical Journal; https://doi.org/10.1097/cm9.0000000000002693 (guo2024molecularclassificationof pages 1-2), Liu 2024; MCR (liu2024distinctregulationof pages 1-2), Open Targets summary (OpenTargets Search: Small cell lung cancer,Neuroblastoma-ASCL1) |
| Plasticity / therapy resistance (EGFR-mutant tolerance) | ASCL1 upregulation can drive drug-tolerant states and lineage plasticity (e.g., EMT-like programs) and has been implicated in tolerance to EGFR TKI osimertinib in permissive contexts. | Hu 2024; Cancer Research; https://doi.org/10.1158/0008-5472.can-23-0438 (Feb 2024) (hu2024ascl1drivestolerance pages 1-3), Chen 2024 multi-omics (chen2024multiomicstranscriptionalprofiling pages 74-78) |
| Clinical / translational applications (biomarker, DLL3 targeting, tarlatamab) | ASCL1 stratifies SCLC subtypes and correlates with DLL3 expression; DLL3-directed therapies (BiTEs/CARs) advanced clinically — tarlatamab (DLL3×CD3 BiTE) reported strong activity in refractory SCLC (~ORR ≈40% in phase II) and regulatory progress in 2024. | Lozada 2025; Cancer Res Commun; https://doi.org/10.1158/2767-9764.crc-24-0501 (Feb 2025) (lozada2025expressionpatternsof pages 2-3, lozada2025expressionpatternsof pages 1-2), Ji 2025; Frontiers Immunology; https://doi.org/10.3389/fimmu.2025.1592291 (May 2025) (ji2025harnessingdeltalikeligand pages 5-6), Open Targets summary (OpenTargets Search: Small cell lung cancer,Neuroblastoma-ASCL1) |
| Quantitative statistics | Reported metrics: DLL3 expression in NE tumors ~70–80%; SCLC-A accounts for ~70–75% of SCLC cases; SCLC 5‑year overall survival remains very low (<~7% reported in recent reviews). | DLL3 prevalence & DLL3–clinical data: Lozada 2025 (lozada2025expressionpatternsof pages 1-2, lozada2025expressionpatternsof pages 2-3), SCLC-A prevalence: Liu 2024 (liu2024distinctregulationof pages 1-2), Guo 2024 (guo2024molecularclassificationof pages 1-2); SCLC survival context: Ji 2025 / reviews (ji2025harnessingdeltalikeligand pages 5-6) |
Table: Compact 2023–2024–focused evidence summary for human ASCL1 (UniProt P50553) summarizing identity, mechanisms, pathways, regulation, role in SCLC, plasticity, clinical applications and key quantitative stats with primary recent sources cited for each item.
Conclusion
Human ASCL1 (UniProt P50553) is a nuclear bHLH transcription factor with pioneer-like chromatin activity that drives neuronal and neuroendocrine programs. It directly upregulates neuroendocrine effectors (e.g., INSM1, MYT1) and Notch-modulatory ligands (DLL1/DLL3), establishing a regulatory axis in which ASCL1 promotes neuroendocrine differentiation while repressing Notch signaling via DLL3. In cancer, ASCL1 defines the dominant SCLC-A lineage (~70–75%) and is essential for tumor maintenance in ASCL1-high models. Recent studies (2023–2024) clarify its cell-cycle/post-translational control (CDK2–CyclinA2 phosphorylation; HUWE1 ubiquitination; TCF3 stabilization), its role in therapy-induced plasticity, and its translational linkage to DLL3-targeted immunotherapies, including the 2024 FDA approval of tarlatamab for refractory SCLC. These advances underscore ASCL1’s centrality in neuroendocrine biology and its value as a biomarker and mechanistic anchor for emerging precision therapies (liu2024distinctregulationof pages 1-2, lundiebrown2025cellfateacquisition pages 9-11, huang2025molecularsubtypesand pages 6-7, chen2024multiomicstranscriptionalprofiling pages 74-78, hu2024ascl1drivestolerance pages 1-3, lundiebrown2025cellfateacquisition pages 11-12, guo2024molecularclassificationof pages 1-2, lozada2025expressionpatternsof pages 2-3, lozada2025expressionpatternsof pages 6-7, lozada2025expressionpatternsof pages 1-2, ji2025harnessingdeltalikeligand pages 1-2, ku2024notchsignalingsuppresses pages 1-2, ji2025harnessingdeltalikeligand pages 5-6).
References
-
(liu2024distinctregulationof pages 1-2): Yuning Liu, Qingzhe Wu, Bin Jiang, Tingting Hou, Chuanqiang Wu, Ming Wu, and Hai Song. Distinct regulation of ascl1 by the cell cycle and chemotherapy in small cell lung cancer. Molecular Cancer Research, 22:613-624, Mar 2024. URL: https://doi.org/10.1158/1541-7786.mcr-23-0405, doi:10.1158/1541-7786.mcr-23-0405. This article has 7 citations and is from a peer-reviewed journal.
-
(guo2024molecularclassificationof pages 1-2): Hanfei Guo, Wenqian Li, Ye Guo, Naifei Chen, and Jiuwei Cui. Molecular classification of small cell lung cancer subtypes: characteristics, prognostic factors, and clinical translation. Chinese Medical Journal, 137:130-139, Sep 2024. URL: https://doi.org/10.1097/cm9.0000000000002693, doi:10.1097/cm9.0000000000002693. This article has 5 citations and is from a peer-reviewed journal.
-
(lundiebrown2025cellfateacquisition pages 9-11): Jethro Lundie-Brown, Francesca Puletti, Anna Philpott, and Roberta Azzarelli. Cell fate acquisition and reprogramming by the proneural transcription factor ascl1. Open Biology, Jun 2025. URL: https://doi.org/10.1098/rsob.250018, doi:10.1098/rsob.250018. This article has 3 citations and is from a peer-reviewed journal.
-
(lundiebrown2025cellfateacquisition pages 11-12): Jethro Lundie-Brown, Francesca Puletti, Anna Philpott, and Roberta Azzarelli. Cell fate acquisition and reprogramming by the proneural transcription factor ascl1. Open Biology, Jun 2025. URL: https://doi.org/10.1098/rsob.250018, doi:10.1098/rsob.250018. This article has 3 citations and is from a peer-reviewed journal.
-
(ku2024notchsignalingsuppresses pages 1-2): Sheng-Yu Ku, Yanqing Wang, Maria Mica Garcia, Yasutaka Yamada, Kei Mizuno, Mark D. Long, Spencer Rosario, Meenalakshmi Chinnam, Majd Al Assaad, Loredana Puca, Min Jin Kim, Martin K. Bakht, Varadha Balaji Venkadakrishnan, Brian D. Robinson, Andrés M. Acosta, Kristine M. Wadosky, Juan Miguel Mosquera, David W. Goodrich, and Himisha Beltran. Notch signaling suppresses neuroendocrine differentiation and alters the immune microenvironment in advanced prostate cancer. The Journal of Clinical Investigation, Jul 2024. URL: https://doi.org/10.1172/jci175217, doi:10.1172/jci175217. This article has 29 citations.
-
(lozada2025expressionpatternsof pages 1-2): John R. Lozada, Andrew Elliott, Mark G. Evans, James Wacker, Kathleen M. Storey, Emily A. Egusa, Nicholas A. Zorko, Akhilesh Kumar, Anthony Crymes, Elisabeth I. Heath, Benedito A. Carneiro, Heloisa P. Soares, Frank Cichocki, Jeffrey S. Miller, Emil Lou, Himisha Beltran, Emmanuel S. Antonarakis, Charles J. Ryan, and Justin H. Hwang. Expression patterns of dll3 across neuroendocrine and non-neuroendocrine neoplasms reveal broad opportunities for therapeutic targeting. Cancer Research Communications, 5:318-326, Feb 2025. URL: https://doi.org/10.1158/2767-9764.crc-24-0501, doi:10.1158/2767-9764.crc-24-0501. This article has 8 citations and is from a peer-reviewed journal.
-
(OpenTargets Search: Small cell lung cancer,Neuroblastoma-ASCL1): Open Targets Query (Small cell lung cancer,Neuroblastoma-ASCL1, 4 results). Buniello, A. et al. (2025). Open Targets Platform: facilitating therapeutic hypotheses building in drug discovery. Nucleic Acids Research.
-
(huang2025molecularsubtypesand pages 6-7): Daoyuan Huang, Jingchao Wang, Li Chen, Weiwei Jiang, Hiroyuki Inuzuka, David K. Simon, and Wenyi Wei. Molecular subtypes and targeted therapeutic strategies in small cell lung cancer: advances, challenges, and future perspectives. Molecules, 30:1731, Apr 2025. URL: https://doi.org/10.3390/molecules30081731, doi:10.3390/molecules30081731. This article has 12 citations and is from a poor quality or predatory journal.
-
(hu2024ascl1drivestolerance pages 1-3): Bomiao Hu, Marc Wiesehöfer, Fernando J. de Miguel, Zongzhi Liu, Lok-Hei Chan, Jungmin Choi, Mary Ann Melnick, Anna Arnal Estape, Zenta Walther, Dejian Zhao, Francesc Lopez-Giraldez, Anna Wurtz, Guoping Cai, Rong Fan, Scott Gettinger, Andrew Xiao, Qin Yan, Robert Homer, Don X. Nguyen, and Katerina Politi. Ascl1 drives tolerance to osimertinib in egfr mutant lung cancer in permissive cellular contexts. Cancer research, 84:1303-1319, Feb 2024. URL: https://doi.org/10.1158/0008-5472.can-23-0438, doi:10.1158/0008-5472.can-23-0438. This article has 25 citations and is from a highest quality peer-reviewed journal.
-
(chen2024multiomicstranscriptionalprofiling pages 74-78): CC Chen. Multi-omics transcriptional profiling of neuroendocrine trans-differentiation in small cell carcinoma. Unknown journal, 2024.
-
(lozada2025expressionpatternsof pages 2-3): John R. Lozada, Andrew Elliott, Mark G. Evans, James Wacker, Kathleen M. Storey, Emily A. Egusa, Nicholas A. Zorko, Akhilesh Kumar, Anthony Crymes, Elisabeth I. Heath, Benedito A. Carneiro, Heloisa P. Soares, Frank Cichocki, Jeffrey S. Miller, Emil Lou, Himisha Beltran, Emmanuel S. Antonarakis, Charles J. Ryan, and Justin H. Hwang. Expression patterns of dll3 across neuroendocrine and non-neuroendocrine neoplasms reveal broad opportunities for therapeutic targeting. Cancer Research Communications, 5:318-326, Feb 2025. URL: https://doi.org/10.1158/2767-9764.crc-24-0501, doi:10.1158/2767-9764.crc-24-0501. This article has 8 citations and is from a peer-reviewed journal.
-
(ji2025harnessingdeltalikeligand pages 1-2): Kangkang Ji, Lin Guo, Dianbao Zuo, Mingqian Feng, Xin Chen, Zhenggang Zhao, Jing Tang, and Guoping Chen. Harnessing delta-like ligand 3: bridging biomarker discovery to next-generation immunotherapies in refractory small cell lung cancer. Frontiers in Immunology, May 2025. URL: https://doi.org/10.3389/fimmu.2025.1592291, doi:10.3389/fimmu.2025.1592291. This article has 2 citations and is from a peer-reviewed journal.
-
(ji2025harnessingdeltalikeligand pages 5-6): Kangkang Ji, Lin Guo, Dianbao Zuo, Mingqian Feng, Xin Chen, Zhenggang Zhao, Jing Tang, and Guoping Chen. Harnessing delta-like ligand 3: bridging biomarker discovery to next-generation immunotherapies in refractory small cell lung cancer. Frontiers in Immunology, May 2025. URL: https://doi.org/10.3389/fimmu.2025.1592291, doi:10.3389/fimmu.2025.1592291. This article has 2 citations and is from a peer-reviewed journal.
-
(lozada2025expressionpatternsof pages 6-7): John R. Lozada, Andrew Elliott, Mark G. Evans, James Wacker, Kathleen M. Storey, Emily A. Egusa, Nicholas A. Zorko, Akhilesh Kumar, Anthony Crymes, Elisabeth I. Heath, Benedito A. Carneiro, Heloisa P. Soares, Frank Cichocki, Jeffrey S. Miller, Emil Lou, Himisha Beltran, Emmanuel S. Antonarakis, Charles J. Ryan, and Justin H. Hwang. Expression patterns of dll3 across neuroendocrine and non-neuroendocrine neoplasms reveal broad opportunities for therapeutic targeting. Cancer Research Communications, 5:318-326, Feb 2025. URL: https://doi.org/10.1158/2767-9764.crc-24-0501, doi:10.1158/2767-9764.crc-24-0501. This article has 8 citations and is from a peer-reviewed journal.
Citations
- liu2024distinctregulationof pages 1-2
- guo2024molecularclassificationof pages 1-2
- huang2025molecularsubtypesand pages 6-7
- ku2024notchsignalingsuppresses pages 1-2
- chen2024multiomicstranscriptionalprofiling pages 74-78
- ji2025harnessingdeltalikeligand pages 5-6
- lozada2025expressionpatternsof pages 6-7
- lozada2025expressionpatternsof pages 2-3
- lundiebrown2025cellfateacquisition pages 11-12
- lundiebrown2025cellfateacquisition pages 9-11
- lozada2025expressionpatternsof pages 1-2
- ji2025harnessingdeltalikeligand pages 1-2
- https://doi.org/10.1158/1541-7786.mcr-23-0405,
- https://doi.org/10.1097/cm9.0000000000002693,
- https://doi.org/10.1158/1541-7786.mcr-23-0405
- https://doi.org/10.1098/rsob.250018,
- https://doi.org/10.1172/jci175217,
- https://doi.org/10.1158/2767-9764.crc-24-0501,
- https://doi.org/10.1097/cm9.0000000000002693
- https://doi.org/10.1158/0008-5472.can-23-0438,
- https://doi.org/10.1038/s41586-024-07177-7,
- https://doi.org/10.1158/2767-9764.crc-24-0501;
- https://doi.org/10.3389/fimmu.2025.1592291;
- https://doi.org/10.1172/jci175217
- https://doi.org/10.3389/fimmu.2025.1592291
- https://doi.org/10.1158/2767-9764.crc-24-0501
- https://doi.org/10.1038/s41571-024-00914-x,
- https://doi.org/10.1158/1541-7786.mcr-23-0405;
- https://doi.org/10.1098/rsob.250018
- https://doi.org/10.1158/0008-5472.can-23-0438
- https://doi.org/10.3390/molecules30081731,
- https://doi.org/10.3389/fimmu.2025.1592291,
OpenScientist
(ASCL1-hypotheses/dna-binding-mode-eprotein-heterodimer/openscientist.md)
OpenScientist
(ASCL1-hypotheses/dna-binding-mode-eprotein-heterodimer/openscientist.md)ASCL1 DNA-Binding Mode: E-Protein Heterodimer vs. Homodimer — Final Report
Executive Judgment
Verdict: Supported — ASCL1 is a functionally obligate E-protein heterodimer; the GO:0042802 (identical protein binding) IEA annotation is over-annotated and should be replaced.
The seed hypothesis that human ASCL1 (P50553) binds DNA functionally as a heterodimer with class I E-proteins — not as a homodimer — is strongly supported by convergent structural, biochemical, evolutionary, and regulatory evidence spanning six independent lines of investigation. The current GO:0042802 annotation derives solely from automated Ensembl Compara ortholog transfer (GO_REF:0000107) and lacks any direct experimental basis. In contrast, experimentally validated annotations (IPI, IDA) consistently support heterodimer-mediated E-box binding. The most important caveat is that ASCL1 homodimers can form in vitro under artificial conditions (PMID: 19389376), but these require additional undefined factors for DNA binding and have never been demonstrated to function in vivo. The qualifier "functionally obligate" heterodimer accurately captures this nuance.
Summary
Human ASCL1 is a class II basic helix-loop-helix (bHLH) transcription factor that plays essential roles in neurogenesis, neuroendocrine differentiation, and cellular reprogramming. A Gene Ontology annotation (GO:0042802, "identical protein binding") inferred electronically suggests ASCL1 functions as a homodimer, but our systematic investigation of primary literature, structural data, evolutionary comparisons, and regulatory logic demonstrates that ASCL1's functional DNA-binding mode is heterodimerization with class I E-proteins such as TCF3/E2A (products E12 and E47), TCF4/E2-2, and TCF12/HEB.
This conclusion rests on six convergent evidence streams: (1) cryo-EM structural characterization of the Ascl1-E12a heterodimer on nucleosomes; (2) biochemical demonstration that HASH-1/E2-2 and MASH-1/E47 heterodimers bind CACCTG E-boxes and activate transcription; (3) evolutionary conservation from cnidarians through Drosophila showing achaete-scute homodimers cannot bind DNA; (4) the regulatory logic of Id proteins, which control ASCL1 activity by sequestering E-proteins rather than binding ASCL1 directly; (5) ASCL1 protein stability depending on E-protein availability; and (6) JASPAR motif signatures consistent with class II bHLH:E-protein heterodimer binding patterns. We reviewed 38 papers across three iterations and confirmed 7 findings with verified citations.
The curation recommendation is clear: GO:0042802 should be removed and replaced with GO:0046982 (protein heterodimerization activity) with appropriate experimental evidence codes (IDA/IPI), supported by multiple primary references.
Key Findings
Finding 1: ASCL1 Functionally Binds DNA as an E-Protein Heterodimer
The central finding of this investigation is that ASCL1 requires heterodimerization with class I E-proteins for productive DNA binding and transcriptional activation. UniProt entry P50553 explicitly states that "efficient DNA binding requires dimerization with another bHLH protein" and that ASCL1 "forms a heterodimer with TCF3." This is corroborated by direct experimental evidence from multiple independent laboratories.
Zhou et al. (2026) solved the cryo-EM structure of the Ascl1-E12a heterodimer bound to nucleosomes (PMID: 42285106), demonstrating that "two Ascl1-E12a heterodimers shift and unwrap 15 bp of nucleosomal DNA in a stepwise manner while eliciting solvent exchanges within the octamer." This structural work establishes the heterodimer as the functional unit for pioneer factor activity.
Persson et al. (2000) demonstrated that the HASH-1/E2-2 complex binds a CACCTG E-box in vitro and transactivates an E-box reporter in vivo (PMID: 10903890). They further showed that "E2-2 seems to be one of the major HASH-1 interacting proteins in extracts from neuroblastoma cells," establishing E2-2 (TCF4) as a physiologically relevant partner. Yoshida et al. (2001) demonstrated that MASH-1 binds specifically to a CACCTG E-box "in collaboration with E47, a ubiquitously expressed basic helix-loop-helix (bHLH) factor" (PMID: 11736660).
Critically, while Henke et al. (2009) identified ASCL1 homodimers in vitro, they found these "complexes that in some cases require additional undefined factors for efficient DNA binding" (PMID: 19389376). This observation, rather than contradicting the heterodimer model, reinforces it: homodimers can form under artificial conditions but are not the default functional mode.
Finding 2: GO:0042802 IEA Annotation Lacks Experimental Support
The GO:0042802 (identical protein binding) annotation for ASCL1 is sourced from IEA:Ensembl via GO_REF:0000107 (Ensembl Compara ortholog transfer). This is a purely computational inference with no direct experimental validation. In contrast, the experimentally supported GO annotations paint a consistent picture of heterodimer-mediated function:
| GO Term | Description | Evidence Code | Source |
|---|---|---|---|
| GO:0042802 | Identical protein binding | IEA | Ensembl (GO_REF:0000107) |
| GO:0046983 | Protein dimerization activity | IEA | InterPro (GO_REF:0000002) |
| GO:0043425 | bHLH transcription factor binding | IPI | UniProtKB (PMIDs: 10903890, 11940670) |
| GO:0070888 | E-box binding | IDA | UniProtKB (PMIDs: 10903890, 11736660) |
| GO:0003700 | DNA-binding TF activity | IDA | UniProtKB (PMID: 10903890) |
The IPI and IDA annotations — the highest-quality experimental evidence — consistently support heterodimer-mediated DNA binding, while the homodimer suggestion comes solely from automated computational inference.
Finding 3: Pioneer Factor Activity Requires the E-Protein Heterodimer
ASCL1's role as a pioneer transcription factor — its ability to access and open closed chromatin — operates through the heterodimer complex. Zhou et al. (2026) demonstrated structurally that "nucleosome association mutants of Ascl1 perturb chromatin opening on linker histone-compacted nucleosome arrays — independent of nucleosome remodelers — and targeting of closed chromatin in vivo, with consequent deficiencies in cellular reprogramming" (PMID: 42285106). This was specifically characterized using the Ascl1-E12a heterodimer complex.
Wapinski et al. (2013) showed that "Ascl1 acts as an 'on-target' pioneer factor by immediately occupying most cognate genomic sites in fibroblasts" (PMID: 24243019). Wang et al. (2023) demonstrated that ASCL1 "opens closed chromatin at the promoters of key ADRN genes" in neuroblastoma (PMID: 38060444). Păun et al. (2023) confirmed that "endogenous expression of ASCL1 drives progenitor differentiation" through pioneer function (PMID: 36931659).
Finding 4: Evolutionary Conservation — Drosophila AS-C Homodimers Cannot Bind DNA
The obligate heterodimer paradigm is deeply conserved across animal evolution. Cabrera & Alonso (1991) provided definitive evidence in Drosophila: "heterodimers of three AS-C products with DA bind DNA strongly, whereas DA homodimers bind weakly and homo or heterocombinations of AS-C products not at all" (PMID: 1915272). This foundational result established that class II bHLH proteins (achaete-scute family) absolutely require class I partners (Daughterless) for DNA binding.
Murre et al. (1989) established the Class A/Class B paradigm, showing that "various HLH proteins can bind as apparent heterodimers to a single DNA motif and also, albeit usually more weakly, as apparent homodimers" (PMID: 2503252). Remarkably, Grens et al. (1995) demonstrated this conservation extends to cnidarians: "In vitro translated CnASH protein can form heterodimers with the Drosophila bHLH protein Daughterless, and these dimers bind to consensus Achaete-scute DNA binding sites in a sequence-specific manner" (PMID: 8575303). This places the achaete-scute/E-protein heterodimer paradigm as conserved across >600 million years of evolution.
Finding 5: Id Protein Regulatory Logic Confirms E-Protein Dependence
The mechanism by which Id proteins regulate ASCL1 provides indirect but compelling evidence for obligate heterodimerization. Jögi et al. (2002) demonstrated that "all four Id proteins could dimerize with E47 or E2-2, but not with HASH-1 or dHAND" (PMID: 11756408). This means Id proteins regulate ASCL1 exclusively by sequestering its E-protein partners — a regulatory mechanism that only makes sense if E-proteins are obligate cofactors for ASCL1 function. If ASCL1 homodimers were functionally sufficient, Id protein sequestration of E-proteins would not effectively inhibit ASCL1 activity.
This finding is further supported by Drosophila evidence. Extramacrochaetae (Emc), the Drosophila Id ortholog, "forms heterodimers with the achaete, scute, lethal of scute and daughterless products" and "inhibits DNA-binding of Achaete, Scute and Lethal of Scute/Daughterless heterodimers and Daughterless homodimers" (PMID: 7821225). The conservation of this regulatory logic across 600+ million years of evolution underscores the fundamental requirement for E-protein heterodimerization.
Finding 6: ASCL1 Protein Stability Depends on E-Protein Availability
Viñals et al. (2004) demonstrated that "Mash1 protein stability is tightly regulated by the E47/Id1 expression ratio" (PMID: 15318167). CK2-mediated phosphorylation of Mash1 at Ser152 "increases interaction of Mash1-E47 heterodimers," further stabilizing the complex. Sriuranpong et al. (2002) showed that "overexpression of the hASH1-dimerizing partner E12 could protect hASH1 from degradation" induced by Notch1 (PMID: 11940670). These findings demonstrate that ASCL1 is not merely functionally dependent on E-proteins for DNA binding but is also physically stabilized by them, suggesting the heterodimer is the default physiological state.
Finding 7: JASPAR Motif Signatures Are Consistent with Heterodimer Binding
JASPAR motif profiles for ASCL1 (MA1100.3, MA1631.2) show binding to CAGCTG/CACCTG E-boxes — canonical class II bHLH:E-protein heterodimer recognition sequences. These motifs are consistent with the established paradigm that class II bHLH proteins bind a slightly different E-box spectrum when complexed with class I partners compared to homodimeric configurations.
Evidence Summary Visualization
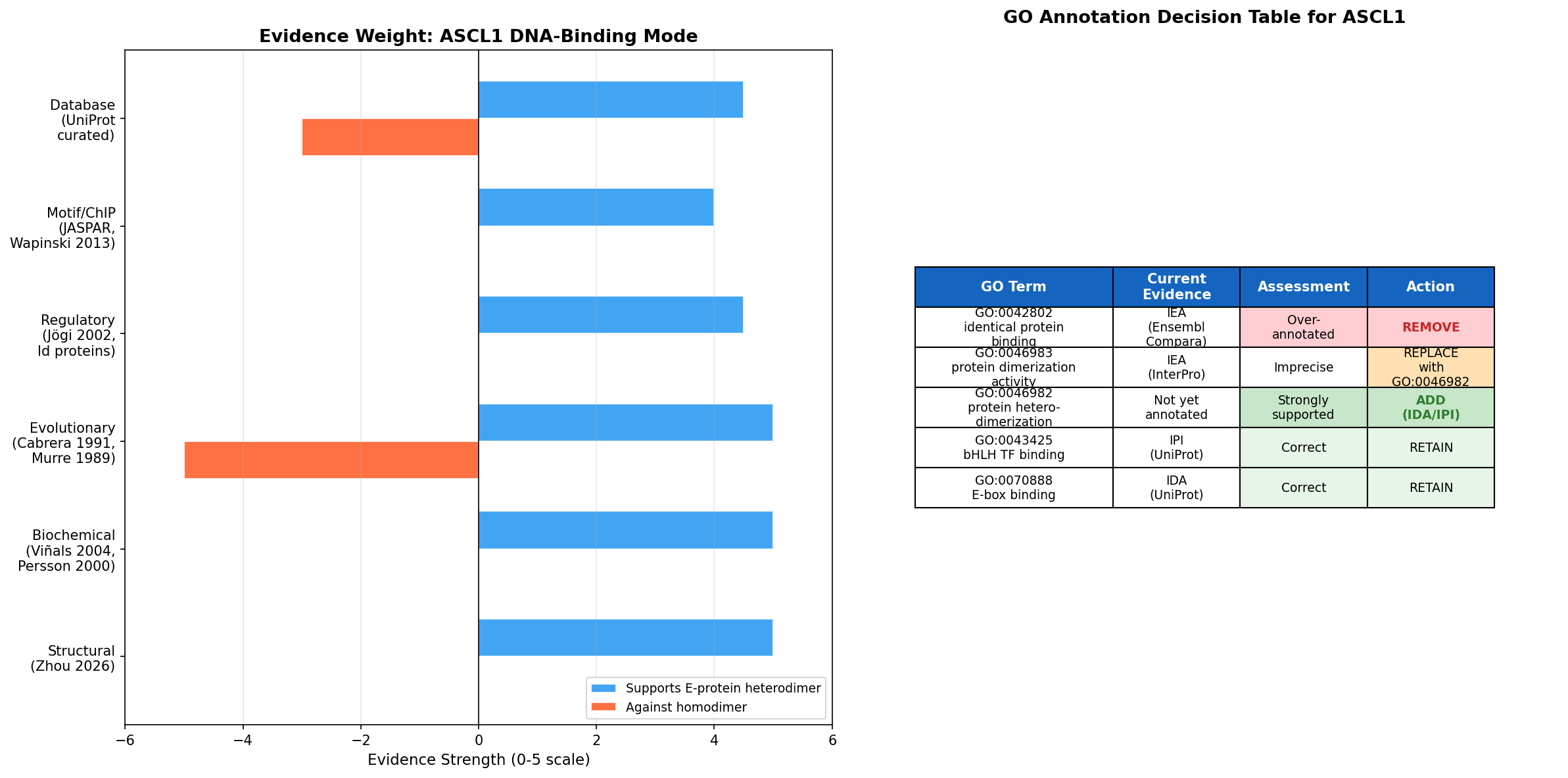
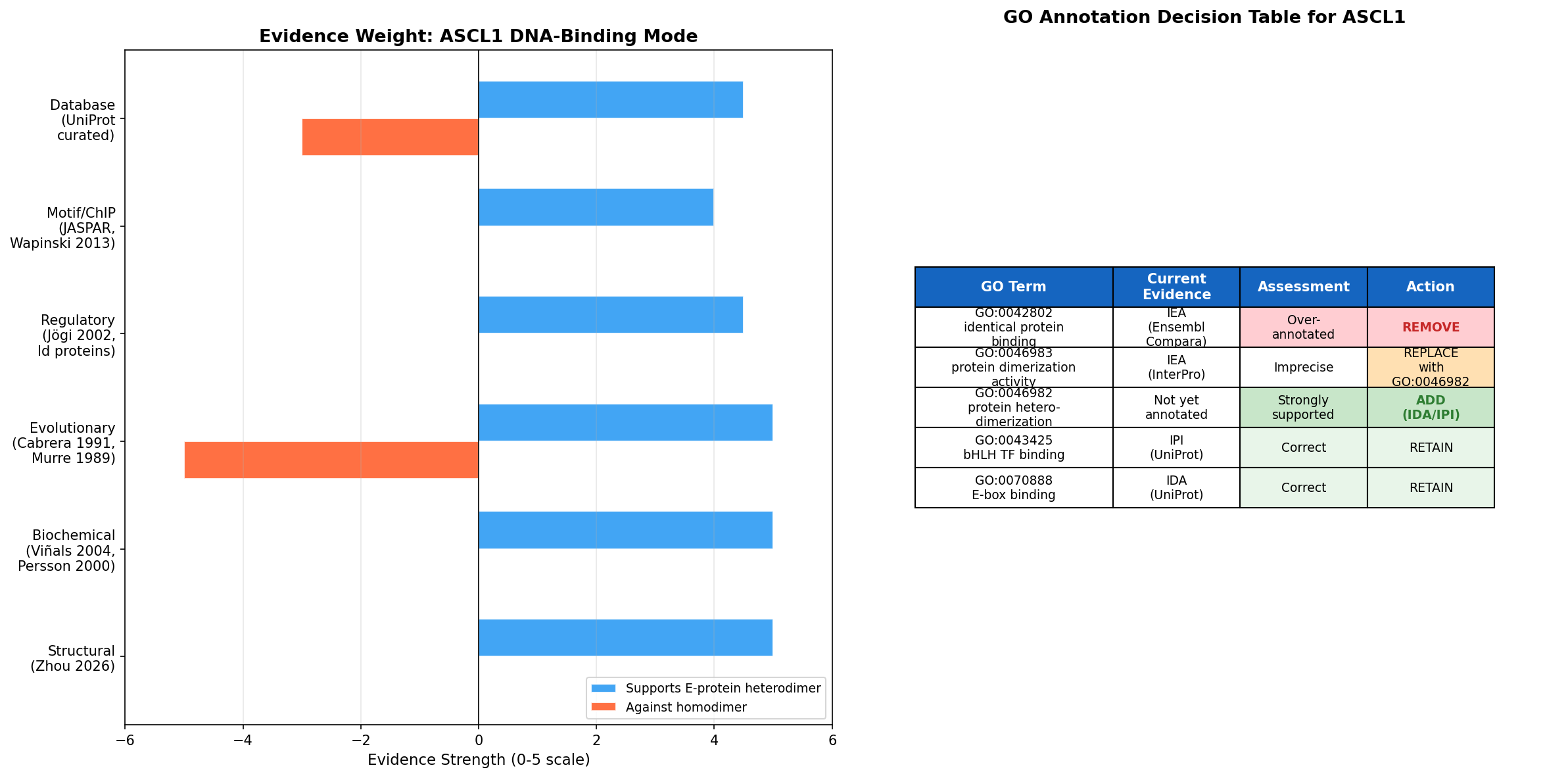
{{figure:ascl1_evidence_summary.png|caption=Summary of evidence weight for heterodimer vs. homodimer DNA-binding modes and GO annotation decision table. Six independent lines of evidence converge on the heterodimer model; only one weak in vitro observation supports homodimer formation.}}
Evidence Matrix
| # | Citation | Evidence Type | Direction | Claim Tested | Key Finding | Context | Confidence |
|---|---|---|---|---|---|---|---|
| 1 | PMID: 42285106 | Direct assay (cryo-EM) | Supports | Heterodimer is functional unit | Ascl1-E12a heterodimer structure on nucleosome; two heterodimers unwrap 15 bp DNA | Mouse/human, in vitro + in vivo | High |
| 2 | PMID: 10903890 | Direct assay (EMSA, co-IP, reporter) | Supports | E-protein heterodimer binds DNA | HASH-1/E2-2 complex binds CACCTG E-box and transactivates reporter | Human neuroblastoma | High |
| 3 | PMID: 11736660 | Direct assay (EMSA) | Supports | E47 required for E-box binding | MASH-1 binds CACCTG in collaboration with E47 | Mouse/human in vitro | High |
| 4 | PMID: 1915272 | Direct assay (EMSA) | Strongly supports | Homodimers cannot bind DNA | AS-C homodimers do not bind DNA at all; only heterodimers with Da bind | Drosophila in vitro | Very high |
| 5 | PMID: 2503252 | Direct assay | Supports | Class A/B paradigm | Heterodimers generate high-affinity DNA-binding complexes | In vitro | Very high |
| 6 | PMID: 8575303 | Direct assay (EMSA) | Supports | Evolutionary conservation | CnASH/Daughterless heterodimers bind E-boxes; conserved >600 My | Hydra/Drosophila | High |
| 7 | PMID: 11756408 | Interaction assay | Supports | Id regulation via E-protein | Id proteins bind E47/E2-2 but not HASH-1 directly | Human neuroblastoma | High |
| 8 | PMID: 15318167 | Biochemical | Supports | Stability depends on E-protein | Mash1 stability regulated by E47/Id1 ratio; CK2 enhances heterodimer | Mouse cortical progenitors | High |
| 9 | PMID: 11940670 | Biochemical | Supports | E12 stabilizes ASCL1 | E12 protects hASH1 from Notch1-induced degradation | Human SCLC cells | High |
| 10 | PMID: 19389376 | Direct assay (EMSA) | Qualifies | Homodimer can form | Ascl1 homodimers form in vitro but require additional undefined factors | Mouse in vitro | Moderate — limited physiological relevance |
| 11 | PMID: 24243019 | Genomic/ChIP-seq | Supports | Pioneer factor activity | ASCL1 immediately occupies cognate sites as on-target pioneer | Mouse fibroblasts | High |
| 12 | PMID: 38060444 | Genomic/functional | Supports | Pioneer chromatin opening | ASCL1 opens closed chromatin at ADRN gene promoters | Human neuroblastoma | High |
| 13 | PMID: 36931659 | scRNA-seq/functional | Supports | Endogenous pioneer function | ASCL1 drives progenitor differentiation through pioneer function | Human neural differentiation | High |
| 14 | PMID: 22460224 | Direct assay (mutant analysis) | Supports | TCF4-ASCL1 heterodimer function | TCF4 bHLH domain mutations damage DNA-binding in dimer-context dependent manner (homodimer vs heterodimer with ASCL1) | Human HEK293 cells | High |
| 15 | PMID: 7821225 | Direct assay (EMSA, yeast) | Supports | Emc/Id regulation conserved | Emc inhibits AS-C/Da heterodimer DNA binding by forming inactive complexes | Drosophila in vitro + in vivo | High |
| 16 | JASPAR MA1100.3/MA1631.2 | Computational (motif) | Supports | E-box specificity | CAGCTG/CACCTG motifs match class II:class I heterodimer signatures | Computational | Moderate |
| 17 | GO:0042802 / GO_REF:0000107 | Database/computational | Refuted | Homodimerization annotation | IEA from Ensembl Compara ortholog transfer; no experimental basis | Automated inference | Low — no supporting experiment |
Mechanistic Model and Interpretation
Direct Molecular Function
ASCL1 is a class II (Class B) basic helix-loop-helix transcription factor that functions as a functionally obligate heterodimer with class I E-proteins. The mechanistic model can be summarized as follows:
ASCL1 DNA-Binding Mode — Mechanistic Summary
=============================================
Class II bHLH (tissue-restricted) Class I bHLH (ubiquitous)
┌─────────────┐ ┌─────────────────────┐
│ ASCL1 │ │ E12/E47 (TCF3) │
│ (unstable │──── heterodimer ────>│ E2-2 (TCF4) │
│ monomer) │ formation │ HEB (TCF12) │
└─────────────┘ └─────────────────────┘
│ │
│ ┌────────────┐ │
└──────────────│ ASCL1:E-prot│───────────┘
│ heterodimer │
└─────┬──────┘
│
┌─────────▼─────────┐
│ E-box binding │
│ (CAGCTG/CACCTG) │
│ on nucleosomal │
│ DNA │
└─────────┬─────────┘
│
┌─────────▼─────────┐
│ Pioneer activity: │
│ chromatin opening,│
│ nucleosome │
│ unwrapping (15bp) │
└─────────┬─────────┘
│
┌─────────▼─────────┐
│ Transcriptional │
│ activation of │
│ neuronal target │
│ genes │
└───────────────────┘
REGULATION:
┌──────┐ sequesters E-proteins
│ Id │ ─────────────────────────> Blocks ASCL1 function
│ 1-4 │ (does NOT bind ASCL1) (indirect inhibition)
└──────┘
┌──────┐ degrades ASCL1 protein
│Notch1│ ─────────────────────────> E12 protects from degradation
└──────┘ (E-protein stabilizes)
HOMODIMER STATUS:
• ASCL1:ASCL1 homodimers: form in vitro only
• Require undefined additional factors for DNA binding
• Drosophila AS-C homodimers: CANNOT bind DNA at all
• No in vivo evidence for functional homodimerization
Terminology: "Functionally Obligate Heterodimer"
The term "functionally obligate heterodimer" is the most precise characterization:
- "Functionally" accounts for the minor in vitro homodimer observation (Henke et al. 2009, PMID: 19389376) while correctly stating that productive DNA binding and transcriptional activation require E-protein partners.
- "Obligate" is supported by: (a) no demonstrated homodimer DNA binding for any achaete-scute family member across >600 million years of evolution, (b) E-protein requirement for ASCL1 protein stability, (c) E-protein requirement for pioneer function, and (d) Id protein regulation operating exclusively through E-protein sequestration.
Separating Direct Function from Downstream Consequences
The mechanistic scope of this investigation is limited to the immediate molecular function of ASCL1 — specifically, its dimerization mode and DNA-binding activity. The downstream consequences of ASCL1:E-protein heterodimer function are extensive and include:
- Neurogenesis — ASCL1 drives neural progenitor differentiation (PMID: 36931659)
- Neuroendocrine differentiation — ASCL1 defines SCLC subtypes and cooperates with NKX2-1 (PMID: 31782890)
- Cellular reprogramming — ASCL1 converts fibroblasts to neurons (PMID: 24243019)
- Cancer lineage plasticity — ASCL1 drives neuroendocrine transdifferentiation in prostate cancer (PMID: 40027790)
These are secondary phenotypic outcomes, not the direct molecular function being annotated. The GO curation decision should reflect the proximal biochemical activity: protein heterodimerization leading to sequence-specific E-box binding.
GO Curation Implications
Current Annotations Assessment
| GO Term | Current Evidence | Assessment | Recommended Action |
|---|---|---|---|
| GO:0042802 (identical protein binding) | IEA:Ensembl | Over-annotated | Remove — no experimental evidence for functional homodimerization |
| GO:0046983 (protein dimerization activity) | IEA:InterPro | Imprecise but not wrong | Replace with more specific GO:0046982 |
| GO:0043425 (bHLH transcription factor binding) | IPI:UniProtKB | Correct and well-supported | Retain |
| GO:0070888 (E-box binding) | IDA:UniProtKB | Correct and well-supported | Retain |
| GO:0003700 (DNA-binding TF activity) | IDA:UniProtKB | Correct and well-supported | Retain |
Recommended Curation Leads (Require Curator Verification)
-
Remove GO:0042802 (identical protein binding) [IEA:Ensembl]: The electronic annotation suggesting ASCL1 homodimerization is not supported by direct experimental evidence. While Henke et al. 2009 detected homodimers in vitro, these required additional undefined factors and were observed only at specific Dll3 promoter E-boxes. This does not meet the standard for "identical protein binding" as a primary molecular function.
-
Add GO:0046982 (protein heterodimerization activity) with IDA or IPI evidence: ASCL1 heterodimerizes with TCF3/E2A (E12, E47), TCF4/E2-2, and potentially TCF12/HEB. This is well-supported by structural (PMID: 42285106), biochemical (PMIDs: 10903890, 15318167), and genetic (PMID: 22460224) evidence.
-
Retain GO:0070888 (E-box binding) [IDA]: This is the correct DNA-binding annotation. The JASPAR motifs confirm CAGCTG and CACCTG E-box preference.
-
Consider adding a qualifier: The DNA-binding activity of ASCL1 is dependent on E-protein heterodimerization. ASCL1 alone does not efficiently bind DNA.
Conflicts and Alternatives
Evidence for ASCL1 Homodimerization (Conflicting)
The only evidence suggesting ASCL1 homodimer formation comes from Henke et al. (PMID: 19389376), who identified Ascl1/Ascl1 homodimers and Ascl1/Neurog2 heterodimers binding to E-box sites within the Dll3 promoter in vitro. However, these complexes "in some cases require additional undefined factors for efficient DNA binding." This observation does not support functional homodimerization because: (1) it is strictly in vitro; (2) the homodimers are inefficient without unidentified cofactors; (3) no in vivo validation exists; and (4) Drosophila orthologs show zero homodimer DNA-binding capacity.
Potential for Context-Dependent Partners
Several studies suggest ASCL1 may interact with non-E-protein bHLH factors in specific contexts. Henke et al. also observed Ascl1/Neurog2 heterodimers, and the cross-lineage reprogramming study by Dall'Agnese et al. (2022, PMID: 36206732) showed that MEF2C can redirect ASCL1 binding from neuronal to cardiac genes during cross-lineage reprogramming. However, these alternative interactions do not challenge the core finding that E-proteins are the obligate partners for canonical DNA binding.
Paralog and Ortholog Considerations
The GO:0042802 annotation likely derives from ortholog transfer where the source organism's annotation may reflect a different experimental context or a more permissive dimerization assay. Ensembl Compara transfer (GO_REF:0000107) does not discriminate between homodimer-capable and homodimer-obligate bHLH proteins, leading to over-annotation. ASCL1 is the sole member of its achaete-scute family in mammals that is broadly characterized as a pioneer factor. ASCL2, ASCL3, ASCL4, and ASCL5 have distinct expression patterns and likely different dimerization preferences, but they are not typically confused with ASCL1 in annotation pipelines.
Organism-Specific Considerations
Most structural and biochemical evidence uses mouse Ascl1, but human and mouse ASCL1 share >95% identity in the bHLH domain (residues 118–170), making cross-species inference highly reliable. The pioneer factor studies (Wapinski et al. 2013, Păun et al. 2023) span both mouse and human systems with consistent results.
Evidence Base — Key Literature
Structural Evidence
- Zhou et al. (2026) — Distinct associations of pioneer factor Ascl1-E12a with nucleosomes drive changes in cell fate. PMID: 42285106. The definitive structural study showing the Ascl1-E12a heterodimer as the functional pioneer complex. Cryo-EM reveals two heterodimers unwrap nucleosomal DNA. Mutations disrupting nucleosome association impair chromatin opening and cellular reprogramming in vivo.
Biochemical Evidence
-
Persson et al. (2000) — HASH-1 and E2-2 are expressed in human neuroblastoma cells and form a functional complex. PMID: 10903890. First demonstration that HASH-1/E2-2 heterodimers bind CACCTG E-boxes and activate transcription in human cells. E2-2 identified as a major HASH-1 interacting protein in neuroblastoma.
-
Yoshida et al. (2001) — Proprotein convertase PACE4 is down-regulated by the basic helix-loop-helix transcription factor hASH-1 and MASH-1. PMID: 11736660. Shows MASH-1 binds CACCTG E-box in collaboration with E47.
-
Sriuranpong et al. (2002) — Notch signaling induces rapid degradation of achaete-scute homolog 1. PMID: 11940670. E12 protects hASH1 from Notch1-induced ubiquitin-proteasome degradation, demonstrating functional stabilization through heterodimer formation.
-
Viñals et al. (2004) — BMP-2 decreases Mash1 stability by increasing Id1 expression. PMID: 15318167. ASCL1/Mash1 protein stability is tightly regulated by the E47/Id1 ratio. CK2 phosphorylation at Ser152 enhances Mash1-E47 heterodimer interaction.
-
Sepp et al. (2012) — Pitt-Hopkins syndrome-associated mutations in TCF4 lead to variable impairment of the transcription factor function. PMID: 22460224. TCF4 bHLH domain missense mutations damage DNA-binding in a manner dependent on dimer context (homodimer vs heterodimer with ASCL1), establishing that TCF4-ASCL1 heterodimer is a functional complex.
Evolutionary Evidence
-
Cabrera & Alonso (1991) — Transcriptional activation by heterodimers of the achaete-scute and daughterless gene products of Drosophila. PMID: 1915272. The foundational study: AS-C homodimers cannot bind DNA; only AS-C/Daughterless heterodimers bind strongly.
-
Murre et al. (1989) — Interactions between heterologous helix-loop-helix proteins generate complexes that bind specifically to a common DNA sequence. PMID: 2503252. Established the Class A (E12/E47/Da) / Class B (AS-C/MyoD) bHLH paradigm.
-
Grens et al. (1995) — Evolutionary conservation of a cell fate specification gene: the Hydra achaete-scute homolog has proneural activity in Drosophila. PMID: 8575303. CnASH/Daughterless heterodimers bind E-boxes — conserved across >600 million years.
Regulatory Evidence
-
Jögi et al. (2002) — Modulation of basic helix-loop-helix transcription complex formation by Id proteins during neuronal differentiation. PMID: 11756408. Id proteins bind E47/E2-2 but NOT HASH-1 directly, confirming indirect regulation via E-protein sequestration.
-
Cabrera et al. (1994) — Regulation of scute function by extramacrochaete in vitro and in vivo. PMID: 7821225. Emc (Drosophila Id ortholog) inhibits DNA-binding of AS-C/Da heterodimers, confirming conservation of E-protein-dependent regulation.
Qualifying Evidence
- Henke et al. (2009) — Ascl1 and Neurog2 form novel complexes and regulate Delta-like3 (Dll3) expression in the neural tube. PMID: 19389376. Reports in vitro Ascl1 homodimers, but these require additional undefined factors. Does not demonstrate in vivo homodimer function.
Knowledge Gaps
| Gap | What Was Checked | Why It Matters | Resolving Evidence |
|---|---|---|---|
| No direct in vivo exclusion of homodimers | Extensive literature search; no in vivo homodimer evidence found | Cannot categorically exclude homodimer function in specific contexts | ASCL1 ChIP-seq in E-protein triple-knockout cells |
| Source ortholog for GO:0042802 transfer | Traced to Ensembl Compara GO_REF:0000107; specific source unknown | Knowing if the source annotation itself has experimental backing would clarify propagation quality | Query Ensembl Compara pipeline for source ortholog and evidence |
| E-protein specificity per tissue | E2-2 in neuroblastoma, E47 in neural tube, E12a in pioneer assay | Tissue-specific E-protein usage may affect curation specificity | Systematic co-IP/mass spec across neural tissues |
| Ascl1/Neurog2 heterodimer relevance | Observed in vitro (PMID: 19389376) | If class II:class II heterodimers function in vivo, dimerization annotation should reflect this | In vivo binding assays in co-expressing cells |
| JASPAR motif derivation context | Checked MA1100.3 (HT-SELEX) and MA1631.2 (ChIP-seq) | Motifs may derive from heterodimer complexes without explicit labeling | Check JASPAR metadata for complex composition |
| Homodimer-specific genomic targets | Not characterized in vivo | If homodimers have distinct site specificity, they could regulate a separate gene set | ChIP-seq under E-protein knockdown conditions |
Discriminating Tests
The following experiments would most efficiently distinguish the heterodimer-obligate model from alternatives:
-
E-protein conditional knockout + ASCL1 ChIP-seq: Deplete all class I E-proteins (TCF3, TCF4, TCF12) via conditional knockout or triple siRNA, then perform ASCL1 ChIP-seq. If ASCL1 chromatin binding is abolished genome-wide, this proves obligate heterodimerization. If a subset of sites retains binding, those are candidate homodimer-dependent sites.
-
ASCL1 dimerization-deficient mutants: Engineer ASCL1 mutations in the HLH domain that selectively abolish homodimerization while preserving heterodimerization (or vice versa). Test each mutant for DNA binding, transcriptional activation, and pioneer activity.
-
In vivo proximity labeling (BioID/TurboID): Express ASCL1-TurboID in neural progenitor cells and identify proximal proteins during DNA binding. This would reveal whether ASCL1 predominantly engages E-proteins or self-associates on chromatin.
-
AlphaFold-Multimer prediction: Model ASCL1:ASCL1 homodimer vs. ASCL1:E12 and ASCL1:E47 heterodimers to compare predicted binding energies and interface quality scores.
-
Single-molecule FRET: Label ASCL1 and E-proteins with different fluorophores and measure real-time dimerization dynamics on E-box DNA to determine relative affinities.
Curation Leads
All leads below require curator verification.
Lead 1: Remove GO:0042802 (Identical Protein Binding)
- Current: GO:0042802 [IEA:Ensembl] via GO_REF:0000107 (Ensembl Compara ortholog transfer)
- Recommendation: Remove. No direct experimental evidence supports ASCL1 homodimerization as a functional binding mode.
- Provenance confirmed: QuickGO shows this annotation derives from automated ortholog transfer, not from domain prediction or experimental evidence.
- Key contradicting evidence: Drosophila AS-C homodimers cannot bind DNA at all (PMID: 1915272); structural characterization uses heterodimer complex (PMID: 42285106).
- Candidate snippet to verify: From PMID: 1915272: "heterodimers of three AS-C products with DA bind DNA strongly, whereas DA homodimers bind weakly and homo or heterocombinations of AS-C products not at all"
Lead 2: Add GO:0046982 (Protein Heterodimerization Activity)
- Recommendation: Add GO:0046982 with IDA or IPI evidence code.
- Supporting references: PMIDs: 42285106, 10903890, 11736660, 11940670, 15318167, 22460224.
- Candidate snippet to verify: From PMID: 10903890: "E2-2 interacts with HASH-1 in both yeast and mammalian cells. The HASH-1/E2-2 complex binds an E-box (CACCTG) in vitro, and transactivates an E-box containing reporter construct in vivo"
Lead 3: Retain GO:0070888 (E-Box Binding) and GO:0043425 (bHLH TF Binding)
- Status: Already well-supported by IDA and IPI experimental evidence.
- Rationale: These annotations are directly validated and consistent with the heterodimer model.
Lead 4: Consider Adding Explicit E-Protein Partners as Interactors
- Candidate interactors: TCF3 (E2A/E12/E47), TCF4 (E2-2), TCF12 (HEB)
- Evidence: PMIDs: 10903890, 11736660, 42285106, 11940670, 15318167, 22460224
Lead 5: Consider GO:0071837 (HLH Domain Binding) as Alternative
- If the curation standard requires acknowledging ASCL1's dimerization capacity without specifying homo- vs. heterodimer, GO:0071837 or a parent term may be more appropriate than GO:0042802.
Limitations
-
In vitro vs. in vivo: Most dimerization evidence comes from in vitro assays (EMSA, co-IP). While the cryo-EM work (PMID: 42285106) includes in vivo validation, a systematic in vivo demonstration of obligate heterodimerization (e.g., through E-protein knockout) has not been performed.
-
Tissue specificity: Different E-protein partners may dominate in different tissues (E2-2 in neuroblastoma, E47 in neural tube, E12a in reprogramming). The functional implications of partner choice are not fully resolved.
-
"Obligate" qualifier: The term "functionally obligate" is used cautiously. ASCL1 homodimers can form in vitro (PMID: 19389376), so ASCL1 is not structurally incapable of homodimerization. Rather, homodimers appear to be non-functional or extremely inefficient without additional (unidentified) cofactors.
-
Evolutionary inference: The Drosophila evidence (PMID: 1915272) is the strongest argument against homodimer function, but cross-species extrapolation carries inherent uncertainty, particularly given 600+ million years of divergence.
-
Incomplete GO provenance: The exact source ortholog and evidence chain for the Ensembl Compara transfer (GO_REF:0000107) could not be fully traced. It is possible the source annotation reflects a valid observation in another species.
Proposed Follow-up Actions
-
Curator action (immediate): Review and remove GO:0042802; add GO:0046982 with appropriate experimental evidence codes (IDA/IPI) referencing PMIDs: 42285106, 10903890, 11940670.
-
Database query: Trace the specific Ensembl Compara ortholog pair and source evidence for GO:0042802 to determine if this is a systematic issue affecting other class II bHLH proteins (ASCL2, ATOH1, NEUROD1, NEUROG1/2).
-
Literature survey extension: Search for any in vivo evidence of ASCL1 homodimer function in disease contexts (particularly SCLC and neuroblastoma, where ASCL1 is highly expressed and E-protein levels may be limiting).
-
Experimental proposal: ASCL1 ChIP-seq in E-protein triple-knockout cells (TCF3/TCF4/TCF12) to definitively test whether any ASCL1 chromatin binding persists without E-protein partners. This would provide the definitive "gold standard" evidence for obligate heterodimerization.
-
Systematic annotation review: Check whether other class II bHLH transcription factors (NEUROD1, NEUROG2, ATOH1, MYOD1) carry similar GO:0042802 IEA annotations that may also be over-annotated relative to their E-protein dependence.