Deep Research
Falcon
(EDF1-deep-research-falcon.md)
Falcon
(EDF1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Comprehensive research report: Human EDF1 (UniProt O60869) — functional annotation
0) Target verification (critical disambiguation)
The literature summarized here is specifically for human EDF1 (gene symbol EDF1), whose protein product is also known as multiprotein-bridging factor 1 (MBF1) and endothelial differentiation-related factor 1. A primary cloning/functional study explicitly states human MBF1α is identical to EDF-1 and characterizes its coactivator activity (https://doi.org/10.1074/jbc.274.48.34196; published Nov 1999) (kabe1999theroleof pages 2-3, kabe1999theroleof pages 1-1). The major recent mechanistic study of ribosome collisions uses human EDF1 (UniProt O60869) and reports it as a ~16.4 kDa two-domain protein (N-terminal MBF1 domain; C-terminal Cro/C1-type HTH) (https://doi.org/10.7554/eLife.58828; published Aug 2020) (sinha2020edf1coordinatescellular pages 17-19, sinha2020edf1coordinatescellular pages 10-12). These features align with the UniProt identity provided in the prompt.
1) Key concepts and current understanding (definitions and primary function)
1.1 EDF1 as a ribosome-collision response factor (current dominant mechanistic model)
Ribosome collisions occur when translating ribosomes stall and are rear-ended by trailing ribosomes, forming “disomes” that trigger ribosome-associated quality control (RQC) and related stress signaling pathways. A central current concept is that EDF1 functions as an early collision-associated factor (“sensor/adaptor”) that binds a conserved site on the 40S subunit near the mRNA entry channel at or near the collision interface and then helps coordinate downstream responses (sinha2020edf1coordinatescellular pages 1-2, sinha2020edf1coordinatescellular pages 2-4).
Experimentally, EDF1 is robustly recruited to polysomes under collision-inducing conditions and is required for efficient recruitment of translational repressors that prevent new initiation on defective mRNAs (sinha2020edf1coordinatescellular pages 19-20, sinha2020edf1coordinatescellular pages 2-4). In this model EDF1 is not an enzyme or transporter; it is a protein–protein / protein–ribosome adaptor that couples a physical state (ribosome collision) to signaling and translation repression.
Key partners and pathways supported by primary evidence include:
- GIGYF2 and EIF4E2 (4EHP): EDF1 promotes their recruitment to collided ribosomes, initiating a negative-feedback loop that represses new rounds of initiation on problematic mRNAs (sinha2020edf1coordinatescellular pages 19-20).
- ZNF598: EDF1 is not required for collision-dependent ribosomal ubiquitylation, but it facilitates ZNF598 action and/or recruitment, since EDF1 loss reduces collision-stimulated ubiquitylation of ribosomal proteins eS10 and uS10 and causes a modest (~10–20%) decrease in ZNF598 recruitment under collision conditions (sinha2020edf1coordinatescellular pages 9-10, sinha2020edf1coordinatescellular pages 17-19).
- ZAKα→p38 signaling: EDF1 depletion reduces collision-triggered p38 phosphorylation, linking EDF1 to the ribotoxic stress response arm downstream of collisions (sinha2020edf1coordinatescellular pages 9-10, sinha2020edf1coordinatescellular pages 17-19).
1.2 EDF1 as a transcriptional coactivator (classical model; still relevant in specific contexts)
Historically, EDF1/MBF1 was characterized as a transcriptional coactivator—a “bridging factor” that can connect sequence-specific transcription factors to components of the basal transcription machinery. In vitro binding and cell-based reporter assays showed that human MBF1/EDF1 interacts with:
- TBP (TATA-binding protein)
- The nuclear receptor Ad4BP/SF-1
- bZIP/AP-1-family factors including ATF1, and binding was also detected with CREB/CREBP1, c-Jun, c-Fos (https://doi.org/10.1074/jbc.274.48.34196; Nov 1999) (kabe1999theroleof pages 1-2, kabe1999theroleof pages 5-6).
Functionally, EDF1/MBF1 increased Ad4BP/SF-1-dependent transcription and ATF1-dependent transcription in transient transfection assays; a central region (aa ~69–108) was implicated in interactions, and activation was on the order of ~3.5–4-fold for Ad4BP/SF-1-driven transcription in the reported conditions (kabe1999theroleof pages 5-6). Importantly, subcellular localization was regulated: EDF1/MBF1 was primarily cytoplasmic when expressed alone, but coexpression with nuclear Ad4BP/SF-1 induced nuclear accumulation, consistent with partner-driven nuclear localization (kabe1999theroleof pages 1-1, kabe1999theroleof pages 6-7).
A modern synthesis is that EDF1 is best supported as a cytoplasmic ribosome-collision factor under translational stress, while also having context-dependent nuclear/coactivator roles supported by earlier cell-based transcription assays and by signaling-regulated translocation studies (sinha2020edf1coordinatescellular pages 17-19, mariotti2000interactionbetweenendothelial pages 2-4).
2) Molecular function, interactions, and subcellular localization (evidence-based)
2.1 Subcellular localization: cytoplasm ↔ nucleus, and why the literature differs
Different experimental paradigms report different EDF1 localization behaviors:
- Collision/RQC-focused work: EDF1 is described as largely absent from polysomes in rapidly growing cells but robustly recruited to polysomes during collision induction; it binds ribosomes on the 40S mRNA entry channel (sinha2020edf1coordinatescellular pages 17-19, sinha2020edf1coordinatescellular pages 2-4).
- Recent ISR-focused work (2024): subcellular fractionation in the context of ISR induction reports that human EDF1 “exclusively resides in the cytoplasm” and that this does not change with stress in their tested conditions (https://doi.org/10.1016/j.molcel.2024.10.029; Dec 2024) (kim2024multiproteinbridgingfactor pages 10-11).
- Classical transcription-coactivator literature: EDF1/MBF1 is cytoplasmic in baseline conditions in some cell systems but can undergo nuclear translocation when signaling is activated (e.g., phorbol ester) or when nuclear partners are co-expressed (mariotti2000interactionbetweenendothelial pages 2-4, kabe1999theroleof pages 1-1).
These results can be reconciled by a model in which EDF1 is a high-copy cytoplasmic factor with a principal role at ribosomes (especially during collisions), while regulated nuclear enrichment can occur in certain signaling contexts and/or via binding to specific nuclear proteins (kim2024multiproteinbridgingfactor pages 10-11, mariotti2000interactionbetweenendothelial pages 2-4).
2.2 Signaling-linked binding partners: calmodulin and phosphorylation
EDF1 has an experimentally supported calmodulin (CaM) interaction that is regulated by phosphorylation and calcium:
- EDF1 binds CaM in vitro and in vivo; treatment with a phorbol ester (TPA) stimulates EDF1 phosphorylation in endothelial cells, and PKC phosphorylation prevents CaM binding. A phosphomimetic substitution at Thr-91 (T91→D) disrupted CaM binding, demonstrating a mechanistic phosphorylation switch (https://doi.org/10.1074/jbc.M001928200; Aug 2000) (mariotti2000interactionbetweenendothelial pages 2-4).
- A subsequent study reports EDF1 is phosphorylated by PKA in vitro and in vivo, that PKA modulates EDF1/CaM interaction, and that signaling (TPA; forskolin) increases nuclear-associated EDF1, reinforcing the concept of a signaling-regulated cytosolic↔nuclear functional duality (https://doi.org/10.1007/s00018-004-4016-0; Apr 2004) (mariotti2004thedualrole pages 1-2).
3) Structural biology and mechanistic details (high-confidence functional inference)
A key mechanistic anchor is cryo-EM structural evidence showing EDF1 bound to the ribosome:
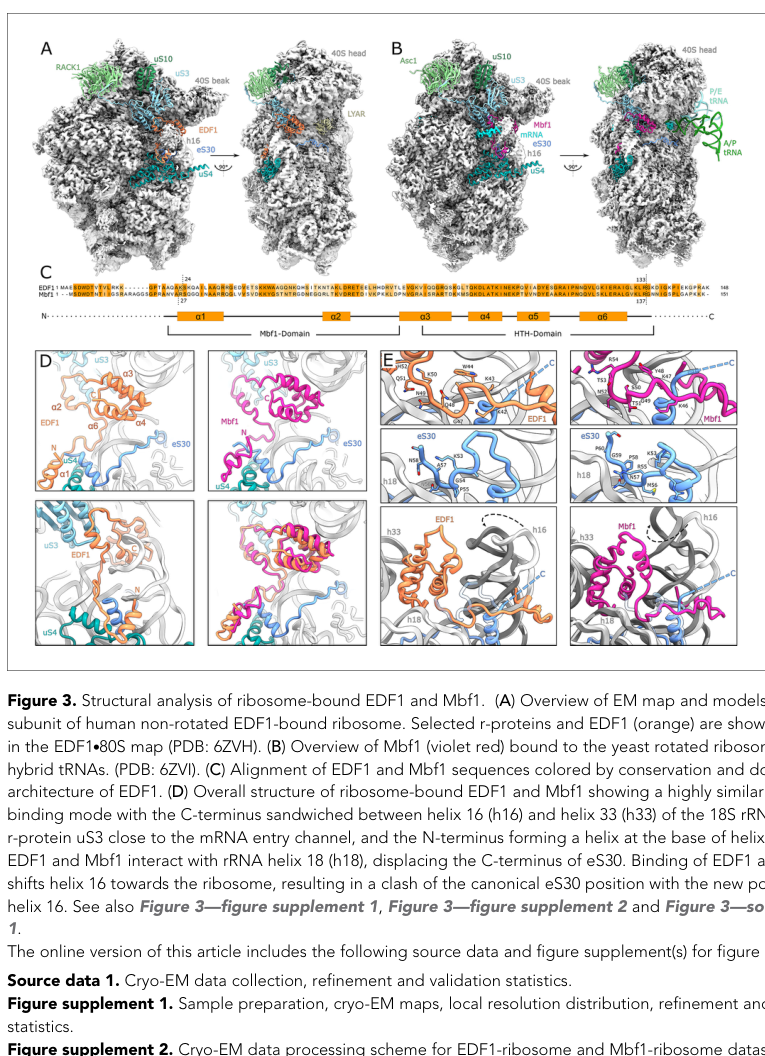
- Cryo-EM resolved EDF1 bound to a non-rotated human 80S complex at 2.9 Å, with a near-complete model spanning Ser-24 to Arg-133 (sinha2020edf1coordinatescellular pages 10-12).
- EDF1’s C-terminal HTH domain is positioned between 18S rRNA helices h16 and h33, while an N-terminal α-helix contacts the base of h16 and ribosomal proteins uS4 and eS30. EDF1 also engages h18 and contacts uS3 near the mRNA entry channel (sinha2020edf1coordinatescellular pages 10-12).
- Conserved sequence motifs (including a KKW motif and a GQNKQ motif) contribute to a “clamp/headlock” over the mRNA path, consistent with a role in stabilizing collided ribosome states and limiting frameshifting under stall/collision conditions (sinha2020edf1coordinatescellular pages 10-12).
Image evidence supporting the binding site and contacts is available from the cryo-EM figure (Figure 3) highlighting EDF1 at the 40S entry channel and showing labeled interactions with uS3/uS4/eS30 and rRNA helices h16/h18/h33 (sinha2020edf1coordinatescellular media d3d9604d).
4) Recent developments (prioritizing 2023–2024)
4.1 2024: EDF1/Mbf1 links collided ribosomes to the integrated stress response (ISR)
A 2024 Molecular Cell paper on Mbf1 (yeast) and its human homologue EDF1 argues that EDF1 acts at collided ribosomes to promote robust ISR signaling, and reports EDF1 is exclusively cytoplasmic by fractionation in their system (https://doi.org/10.1016/j.molcel.2024.10.029; Dec 2024) (kim2024multiproteinbridgingfactor pages 10-11). The same work concludes that Mbf1/EDF1 plays little to no direct role in transcriptional coactivation of Gcn4 during ISR, reinforcing a ribosome-centric model for stress adaptation (kim2024multiproteinbridgingfactor pages 10-11).
4.2 2024: EDF1 in metabolic disease-related transcriptional ribonucleoprotein complexes
A 2024 review of lncRNAs in diet-induced metabolic diseases describes a liver transcriptional complex comprising lncRNA Blnc1 + EDF1 + LXRα, which induces lipogenic genes including Srebp1c, a regulator of de novo lipogenesis and triglyceride synthesis (https://doi.org/10.3390/ijms25115678; May 2024) (brandt2024longnoncodingrnas pages 4-5, brandt2024longnoncodingrnas pages 9-10). The review further summarizes that liver-specific Blnc1 knockout reduces high-fat-diet-associated weight gain, steatosis, and insulin resistance in mice, contextualizing EDF1 as a participant in a clinically relevant metabolic gene-regulatory module (brandt2024longnoncodingrnas pages 9-10).
4.3 2023–2024: EDF1 depletion observed in a ribosome-rescue genetic disease context (retinal dystrophy)
A human genetic disease study of HBS1L deficiency (a ribosomal rescue factor) reports that in a mouse model and patient, retinal dystrophy is associated with broad proteomic disruption and includes changes in the collision-response protein EDF1:
- In the 2024 peer-reviewed article, Edf1 protein was downregulated in 4-week-old Hbs1l hypomorph mouse retinas by quantitative proteomics and validated by western blot, while at 2 weeks Edf1 appeared similar, suggesting Edf1 decrease may be secondary to photoreceptor loss or progressive proteostasis impairment (https://doi.org/10.1242/dmm.050557; Jul 2024) (luo2024hbs1ldeficiencycauses pages 8-9, luo2024hbs1ldeficiencycauses pages 4-5).
- A 2023 preprint provides detailed quantitative statistics: TMT proteomics quantified 8114 retinal proteins (FDR <1%), identifying 169 increased and 480 decreased proteins (with fold-change and p-value cutoffs); EDF1 was among decreased proteins. The same work reports retinal thinning by OCT/histology (e.g., outer retina 55.56 ± 19.77 μm vs 93.92 ± 30.72 μm) and increased photoreceptor apoptosis at 2 weeks (105 ± 87 vs 17 ± 3, P=0.0012) (https://doi.org/10.1101/2023.10.18.562924; Oct 2023) (luo2023geneticdeficiencyof pages 6-8).
5) Current applications and real-world implementations
5.1 EDF1 as a mechanistic node and potential target in translational stress/RQC
EDF1 sits at an actionable interface: it is early-recruited to collided ribosomes and promotes recruitment of translational repressors (GIGYF2/EIF4E2), suggesting that perturbing EDF1 function could modulate how cells prioritize repression vs rescue during translational stress (sinha2020edf1coordinatescellular pages 19-20). While direct EDF1-targeted therapies are not established in the cited sources, EDF1’s positioning upstream of translational repression and stress signaling provides a rationale for considering it in diseases with proteostasis or translational stress components.
5.2 EDF1 in metabolic gene regulation via LXRα/lncRNA complexes
In liver metabolism, EDF1 is described as part of a Blnc1–EDF1–LXRα ribonucleoprotein transcriptional complex that drives lipogenic transcriptional programs (Srebp1c) (brandt2024longnoncodingrnas pages 4-5). This provides a concrete implementation of EDF1 as a co-regulator in a nuclear receptor pathway relevant to MASLD/NAFLD-like phenotypes, albeit primarily in mouse in vivo contexts summarized by review (brandt2024longnoncodingrnas pages 9-10).
5.3 Clinical association and biomarker contexts
OpenTargets lists low-to-moderate evidence associations for EDF1 with broad disease categories including neoplasm and neurodegenerative disease, but these are not yet strong or specific enough in the retrieved evidence to claim EDF1 as a validated clinical biomarker or causal gene (OpenTargets Search: -EDF1). In clinical-trial-scale proteomics, the MAGMA trial demonstrates the feasibility of large plasma-proteome endpoints (7596 proteins) in a cardiovascular/metabolic population with significant imaging outcome differences; EDF1 is mentioned in the retrieved snippet set as part of the broader proteomic context but EDF1-specific quantitative changes were not available from accessible text chunks (https://doi.org/10.1161/CIRCULATIONAHA.123.067620; Aug 2024) (rajagopalan2024mineralocorticoidreceptorantagonism pages 1-3).
6) Expert analysis and synthesis (authoritative interpretations)
A coherent, evidence-weighted interpretation is:
1. Highest-confidence primary function: EDF1 is a cytoplasmic ribosome-associated collision response factor that binds the 40S mRNA entry channel on stalled/collided ribosomes and coordinates downstream translation repression and signaling (sinha2020edf1coordinatescellular pages 17-19, sinha2020edf1coordinatescellular pages 10-12).
2. Mechanistic breadth: EDF1’s binding position and recruitment behavior support a role in collision-state stabilization, frameshift suppression, and adaptor-mediated recruitment of repression factors (GIGYF2/EIF4E2) (sinha2020edf1coordinatescellular pages 10-12, sinha2020edf1coordinatescellular pages 19-20).
3. Transcriptional coactivator evidence remains real but context-dependent: older but direct experiments support TBP and transcription factor binding and reporter activation, and show partner-driven nuclear enrichment (kabe1999theroleof pages 1-2, kabe1999theroleof pages 5-6). However, newer ISR-focused work finds little direct promoter coactivation role in their tested system and reports EDF1 as exclusively cytoplasmic, suggesting the nuclear coactivator role may not be universal across stresses/cell types (kim2024multiproteinbridgingfactor pages 10-11).
4. Signaling and shuttling: CaM binding and phosphorylation-dependent modulation provide a plausible biochemical basis for conditional redistribution and function switching, especially in endothelial contexts (mariotti2000interactionbetweenendothelial pages 2-4, mariotti2004thedualrole pages 1-2).
7) Key statistics and data points (from the most relevant studies)
- Structural resolution: EDF1-bound human 80S cryo-EM structure at 2.9 Å, model spanning Ser-24 to Arg-133 (2020) (sinha2020edf1coordinatescellular pages 10-12).
- ZNF598 recruitment effect size: EDF1 loss caused ~10–20% decrease in ZNF598 recruitment under collision conditions (2020) (sinha2020edf1coordinatescellular pages 9-10).
- Transcriptional coactivation magnitude: EDF1/MBF1 increased Ad4BP/SF-1-dependent transcription ~3.5–4-fold in the reported assay context (1999) (kabe1999theroleof pages 5-6).
- Retinal dystrophy proteomics (HBS1L deficiency): 8114 proteins quantified; 169 up and 480 down (FDR <1%); EDF1 among downregulated proteins (2023) (luo2023geneticdeficiencyof pages 6-8).
- Retinal thinning (mouse model): outer retina 55.56 ± 19.77 μm vs 93.92 ± 30.72 μm (mutant vs control), with extensive apoptosis at 2 weeks (105 ± 87 vs 17 ± 3, P=0.0012) (2023) (luo2023geneticdeficiencyof pages 6-8).
- MAGMA trial imaging endpoints: PWV change 7.1 ± 10.7% vs 0.87 ± 10.0% (p=0.028); ΔTWV 1.2 ± 1.7 cm³ vs 0.037 ± 1.9 cm³ (p=0.022); LV mass +3.1 ± 8.4 g vs −5.8 ± 8.4 g (p=0.001); native T1 +26.0 ± 41.9 ms vs −10.1 ± 36.3 ms (p=6.33×10⁻4) (2024) (rajagopalan2024mineralocorticoidreceptorantagonism pages 1-3).
Evidence table
The following table consolidates EDF1 functions, partners, localization, and key evidence:
| Functional role/Process | Mechanism & key partners | Subcellular location | Evidence type (assay/structure) | Key quantitative/structural details | Primary sources (with year, journal, URL) |
|---|---|---|---|---|---|
| Ribosome collision sensor/adaptor in ribosome-associated quality control | EDF1 is recruited to collided ribosomes independently of ZNF598 but promotes downstream recruitment of GIGYF2/EIF4E2 (4EHP) to repress new initiation on defective mRNAs; facilitates efficient ZNF598-dependent eS10/uS10 ubiquitylation and contributes to collision-triggered p38/ZAKα signaling; recruitment depends on RACK1. (sinha2020edf1coordinatescellular pages 9-10, sinha2020edf1coordinatescellular pages 19-20, sinha2020edf1coordinatescellular pages 1-2, sinha2020edf1coordinatescellular pages 2-4) | Predominantly cytoplasmic; accumulates on polysomes/collided ribosomes during translational distress. (sinha2020edf1coordinatescellular pages 9-10, sinha2020edf1coordinatescellular pages 17-19) | Sucrose-gradient polysome fractionation, TMT proteomics, KO/depletion, immunoblotting, cryo-EM. (sinha2020edf1coordinatescellular pages 9-10, sinha2020edf1coordinatescellular pages 10-12) | Human EDF1 is ~16.4 kDa; EDF1 loss caused a modest ~10–20% decrease in ZNF598 recruitment; cryo-EM resolved EDF1 bound to non-rotated human 80S at 2.9 Å near the 40S mRNA entry channel. (sinha2020edf1coordinatescellular pages 9-10, sinha2020edf1coordinatescellular pages 10-12) | Sinha et al. 2020, eLife, https://doi.org/10.7554/eLife.58828 (sinha2020edf1coordinatescellular pages 9-10, sinha2020edf1coordinatescellular pages 10-12) |
| Transcriptional coactivator bridging basal and sequence-specific transcription factors | hMBF1α is identical to EDF1 and binds TBP plus gene-specific activators including Ad4BP/SF-1 and ATF1/bZIP-family factors (ATF1, CREB/CREBP, c-Jun, c-Fos); enhances DNA binding of some partners and mediates Ad4BP/SF-1- and ATF1-dependent transcription. Nuclear accumulation is induced by coexpression with nuclear Ad4BP/SF-1. (kabe1999theroleof pages 7-8, kabe1999theroleof pages 1-2, kabe1999theroleof pages 1-1, kabe1999theroleof pages 6-7, kabe1999theroleof pages 5-6, kabe1999theroleof pages 2-3) | Cytoplasmic when expressed alone; nuclear enrichment upon interaction with nuclear partners such as Ad4BP/SF-1. (kabe1999theroleof pages 2-3, kabe1999theroleof pages 1-1, kabe1999theroleof pages 1-2) | GST pull-downs, co-immunoprecipitation, EMSA/DNA-binding assays, transient reporter assays, immunofluorescence. (kabe1999theroleof pages 7-8, kabe1999theroleof pages 2-3, kabe1999theroleof pages 1-2, kabe1999theroleof pages 5-6) | hMBF1a/b increased Ad4BP/SF-1-dependent transcription by ~3.5–4-fold; central region aa 69–108 required for interaction; basic-region contacts mapped on Ad4BP/SF-1 and bZIP factors. (kabe1999theroleof pages 5-6) | Kabe et al. 1999, J Biol Chem, https://doi.org/10.1074/jbc.274.48.34196 (kabe1999theroleof pages 7-8, kabe1999theroleof pages 5-6) |
| CaM-binding signaling-responsive shuttling factor | EDF1 binds calmodulin (CaM) in vitro and in vivo; PKC phosphorylation disrupts CaM binding, and PKA phosphorylation modulates EDF1/CaM interaction and correlates with signaling-dependent nuclear accumulation; native and PKC-phosphorylated EDF1 can still interact with TBP, supporting a cytosol-to-nucleus switch between CaM-associated and coactivator states. (mariotti2004thedualrole pages 1-2, mariotti2000interactionbetweenendothelial pages 2-4) | Basal distribution in cytosol and nucleus; TPA and forskolin increase nuclear-associated EDF1. (mariotti2004thedualrole pages 1-2, mariotti2000interactionbetweenendothelial pages 2-4) | In vitro/in vivo phosphorylation assays, CaM-binding assays, co-immunoprecipitation, localization studies in HUVEC/COS cells. (mariotti2004thedualrole pages 1-2, mariotti2000interactionbetweenendothelial pages 2-4) | Thr91→Asp phosphomimetic abolished CaM binding; PKC and PKA both regulate EDF1 localization/function; TPA stimulated EDF1 phosphorylation and nuclear translocation. (mariotti2004thedualrole pages 1-2, mariotti2000interactionbetweenendothelial pages 2-4) | Mariotti et al. 2000, J Biol Chem, https://doi.org/10.1074/jbc.M001928200; Mariotti et al. 2004, Cell Mol Life Sci, https://doi.org/10.1007/s00018-004-4016-0 (mariotti2004thedualrole pages 1-2, mariotti2000interactionbetweenendothelial pages 2-4) |
| Collision-coupled integrated stress response (ISR) activator / frameshift suppressor | Recent work argues EDF1/Mbf1 acts primarily at collided ribosomes, not as a direct nuclear coactivator in ISR; recruitment to collided ribosomes promotes upstream GCN2 pathway activation and eIF2α phosphorylation (shown mechanistically in yeast, inferred conserved for human EDF1), while Mbf1/EDF1 contacts rRNA, ribosomal proteins, and mRNA to prevent frameshifting. (kim2024multiproteinbridgingfactor pages 10-11) | Cytoplasmic; 2024 study reports human EDF1 “exclusively resides in the cytoplasm,” with no stress-induced relocalization detected. (kim2024multiproteinbridgingfactor pages 10-11) | Subcellular fractionation, promoter occupancy/ChIP analyses, collision/ISR assays, comparative mechanistic inference with structural data. (kim2024multiproteinbridgingfactor pages 10-11) | Yeast/human Mbf1-EDF1 present at concentrations orders of magnitude above typical transcription factors; supports stoichiometric ribosome-associated role rather than classical limiting nuclear coactivator role. (kim2024multiproteinbridgingfactor pages 10-11) | Kim et al. 2024, Molecular Cell, https://doi.org/10.1016/j.molcel.2024.10.029 (kim2024multiproteinbridgingfactor pages 10-11) |
| Structural basis of EDF1 domain function | EDF1 contains an N-terminal MBF1 domain and C-terminal Cro/C1-type HTH domain; on collided ribosomes, the HTH domain sits between h16 and h33, while the N-terminal helix contacts h16/uS4/eS30; EDF1 also engages h18 and uS3, consistent with UniProt domain annotation and a direct ribosome-binding mechanism. (sinha2020edf1coordinatescellular pages 17-19, sinha2020edf1coordinatescellular pages 10-12, sinha2020edf1coordinatescellular media d3d9604d) | 40S subunit mRNA entry channel on stalled/collided 80S ribosomes. (sinha2020edf1coordinatescellular pages 10-12, sinha2020edf1coordinatescellular media d3d9604d) | Cryo-EM structural modeling. (sinha2020edf1coordinatescellular pages 10-12, sinha2020edf1coordinatescellular media d3d9604d) | Near-complete human model built for Ser24–Arg133; conserved GQNKQ and KKW motifs contribute to rRNA/mRNA-path contacts; interface includes uS3, uS4, eS30 and rRNA helices h16, h18, h33. (sinha2020edf1coordinatescellular pages 10-12, sinha2020edf1coordinatescellular media d3d9604d) | Sinha et al. 2020, eLife, https://doi.org/10.7554/eLife.58828 (sinha2020edf1coordinatescellular pages 17-19, sinha2020edf1coordinatescellular pages 10-12, sinha2020edf1coordinatescellular media d3d9604d) |
Table: This table consolidates the main experimentally supported functions of human EDF1/MBF1 (UniProt O60869), emphasizing both the classical transcriptional coactivator literature and the newer ribosome-collision quality-control model. It is useful for reconciling EDF1’s localization, partners, mechanisms, and strongest primary evidence.
Key figure (structural localization)
A cryo-EM figure showing EDF1 bound at the 40S mRNA entry channel with labeled interacting ribosomal proteins and rRNA helices is available (sinha2020edf1coordinatescellular media d3d9604d).
References
-
(kabe1999theroleof pages 2-3): Yasuaki Kabe, Masahide Goto, Daisuke Shima, Takeshi Imai, Tadashi Wada, Ken-ichirou Morohashi, Masahiro Shirakawa, Susumu Hirose, and Hiroshi Handa. The role of human mbf1 as a transcriptional coactivator*. The Journal of Biological Chemistry, 274:34196-34202, Nov 1999. URL: https://doi.org/10.1074/jbc.274.48.34196, doi:10.1074/jbc.274.48.34196. This article has 117 citations.
-
(kabe1999theroleof pages 1-1): Yasuaki Kabe, Masahide Goto, Daisuke Shima, Takeshi Imai, Tadashi Wada, Ken-ichirou Morohashi, Masahiro Shirakawa, Susumu Hirose, and Hiroshi Handa. The role of human mbf1 as a transcriptional coactivator*. The Journal of Biological Chemistry, 274:34196-34202, Nov 1999. URL: https://doi.org/10.1074/jbc.274.48.34196, doi:10.1074/jbc.274.48.34196. This article has 117 citations.
-
(sinha2020edf1coordinatescellular pages 17-19): Niladri K Sinha, Alban Ordureau, Katharina Best, James A Saba, Boris Zinshteyn, Elayanambi Sundaramoorthy, Amit Fulzele, Danielle M Garshott, Timo Denk, Matthias Thoms, Joao A Paulo, J Wade Harper, Eric J Bennett, Roland Beckmann, and Rachel Green. Edf1 coordinates cellular responses to ribosome collisions. eLife, Aug 2020. URL: https://doi.org/10.7554/elife.58828, doi:10.7554/elife.58828. This article has 200 citations and is from a domain leading peer-reviewed journal.
-
(sinha2020edf1coordinatescellular pages 10-12): Niladri K Sinha, Alban Ordureau, Katharina Best, James A Saba, Boris Zinshteyn, Elayanambi Sundaramoorthy, Amit Fulzele, Danielle M Garshott, Timo Denk, Matthias Thoms, Joao A Paulo, J Wade Harper, Eric J Bennett, Roland Beckmann, and Rachel Green. Edf1 coordinates cellular responses to ribosome collisions. eLife, Aug 2020. URL: https://doi.org/10.7554/elife.58828, doi:10.7554/elife.58828. This article has 200 citations and is from a domain leading peer-reviewed journal.
-
(sinha2020edf1coordinatescellular pages 1-2): Niladri K Sinha, Alban Ordureau, Katharina Best, James A Saba, Boris Zinshteyn, Elayanambi Sundaramoorthy, Amit Fulzele, Danielle M Garshott, Timo Denk, Matthias Thoms, Joao A Paulo, J Wade Harper, Eric J Bennett, Roland Beckmann, and Rachel Green. Edf1 coordinates cellular responses to ribosome collisions. eLife, Aug 2020. URL: https://doi.org/10.7554/elife.58828, doi:10.7554/elife.58828. This article has 200 citations and is from a domain leading peer-reviewed journal.
-
(sinha2020edf1coordinatescellular pages 2-4): Niladri K Sinha, Alban Ordureau, Katharina Best, James A Saba, Boris Zinshteyn, Elayanambi Sundaramoorthy, Amit Fulzele, Danielle M Garshott, Timo Denk, Matthias Thoms, Joao A Paulo, J Wade Harper, Eric J Bennett, Roland Beckmann, and Rachel Green. Edf1 coordinates cellular responses to ribosome collisions. eLife, Aug 2020. URL: https://doi.org/10.7554/elife.58828, doi:10.7554/elife.58828. This article has 200 citations and is from a domain leading peer-reviewed journal.
-
(sinha2020edf1coordinatescellular pages 19-20): Niladri K Sinha, Alban Ordureau, Katharina Best, James A Saba, Boris Zinshteyn, Elayanambi Sundaramoorthy, Amit Fulzele, Danielle M Garshott, Timo Denk, Matthias Thoms, Joao A Paulo, J Wade Harper, Eric J Bennett, Roland Beckmann, and Rachel Green. Edf1 coordinates cellular responses to ribosome collisions. eLife, Aug 2020. URL: https://doi.org/10.7554/elife.58828, doi:10.7554/elife.58828. This article has 200 citations and is from a domain leading peer-reviewed journal.
-
(sinha2020edf1coordinatescellular pages 9-10): Niladri K Sinha, Alban Ordureau, Katharina Best, James A Saba, Boris Zinshteyn, Elayanambi Sundaramoorthy, Amit Fulzele, Danielle M Garshott, Timo Denk, Matthias Thoms, Joao A Paulo, J Wade Harper, Eric J Bennett, Roland Beckmann, and Rachel Green. Edf1 coordinates cellular responses to ribosome collisions. eLife, Aug 2020. URL: https://doi.org/10.7554/elife.58828, doi:10.7554/elife.58828. This article has 200 citations and is from a domain leading peer-reviewed journal.
-
(kabe1999theroleof pages 1-2): Yasuaki Kabe, Masahide Goto, Daisuke Shima, Takeshi Imai, Tadashi Wada, Ken-ichirou Morohashi, Masahiro Shirakawa, Susumu Hirose, and Hiroshi Handa. The role of human mbf1 as a transcriptional coactivator*. The Journal of Biological Chemistry, 274:34196-34202, Nov 1999. URL: https://doi.org/10.1074/jbc.274.48.34196, doi:10.1074/jbc.274.48.34196. This article has 117 citations.
-
(kabe1999theroleof pages 5-6): Yasuaki Kabe, Masahide Goto, Daisuke Shima, Takeshi Imai, Tadashi Wada, Ken-ichirou Morohashi, Masahiro Shirakawa, Susumu Hirose, and Hiroshi Handa. The role of human mbf1 as a transcriptional coactivator*. The Journal of Biological Chemistry, 274:34196-34202, Nov 1999. URL: https://doi.org/10.1074/jbc.274.48.34196, doi:10.1074/jbc.274.48.34196. This article has 117 citations.
-
(kabe1999theroleof pages 6-7): Yasuaki Kabe, Masahide Goto, Daisuke Shima, Takeshi Imai, Tadashi Wada, Ken-ichirou Morohashi, Masahiro Shirakawa, Susumu Hirose, and Hiroshi Handa. The role of human mbf1 as a transcriptional coactivator*. The Journal of Biological Chemistry, 274:34196-34202, Nov 1999. URL: https://doi.org/10.1074/jbc.274.48.34196, doi:10.1074/jbc.274.48.34196. This article has 117 citations.
-
(mariotti2000interactionbetweenendothelial pages 2-4): Massino Mariotti, Lorenza De Benedictis, Eugenia Avon, and Jeanette A.M. Maier. Interaction between endothelial differentiation-related factor-1 and calmodulin in vitro and in vivo *. The Journal of Biological Chemistry, 275:24047-24051, Aug 2000. URL: https://doi.org/10.1074/jbc.m001928200, doi:10.1074/jbc.m001928200. This article has 57 citations.
-
(kim2024multiproteinbridgingfactor pages 10-11): Kyusik Q. Kim, Jeffrey J. Li, Ankanahalli N. Nanjaraj Urs, Miguel E. Pacheco, Victor Lasehinde, Timo Denk, Petr Tesina, Shota Tomomatsu, Yoshitaka Matsuo, Elesa McDonald, Roland Beckmann, Toshifumi Inada, Rachel Green, and Hani S. Zaher. Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes. Dec 2024. URL: https://doi.org/10.1016/j.molcel.2024.10.029, doi:10.1016/j.molcel.2024.10.029. This article has 13 citations and is from a highest quality peer-reviewed journal.
-
(mariotti2004thedualrole pages 1-2): M. Mariotti, L. De Benedictis, J. A. M. Maier, and E. Ballabio. The dual role of endothelial differentiation-related factor-1 in the cytosol and nucleus: modulation by protein kinase a. Cellular and Molecular Life Sciences CMLS, 61:1069-1074, Apr 2004. URL: https://doi.org/10.1007/s00018-004-4016-0, doi:10.1007/s00018-004-4016-0. This article has 38 citations.
-
(sinha2020edf1coordinatescellular media d3d9604d): Niladri K Sinha, Alban Ordureau, Katharina Best, James A Saba, Boris Zinshteyn, Elayanambi Sundaramoorthy, Amit Fulzele, Danielle M Garshott, Timo Denk, Matthias Thoms, Joao A Paulo, J Wade Harper, Eric J Bennett, Roland Beckmann, and Rachel Green. Edf1 coordinates cellular responses to ribosome collisions. eLife, Aug 2020. URL: https://doi.org/10.7554/elife.58828, doi:10.7554/elife.58828. This article has 200 citations and is from a domain leading peer-reviewed journal.
-
(brandt2024longnoncodingrnas pages 4-5): Annette Brandt and Florian Kopp. Long noncoding rnas in diet-induced metabolic diseases. International Journal of Molecular Sciences, 25:5678, May 2024. URL: https://doi.org/10.3390/ijms25115678, doi:10.3390/ijms25115678. This article has 7 citations.
-
(brandt2024longnoncodingrnas pages 9-10): Annette Brandt and Florian Kopp. Long noncoding rnas in diet-induced metabolic diseases. International Journal of Molecular Sciences, 25:5678, May 2024. URL: https://doi.org/10.3390/ijms25115678, doi:10.3390/ijms25115678. This article has 7 citations.
-
(luo2024hbs1ldeficiencycauses pages 8-9): Shiyu Luo, Bilal Alwattar, Qifei Li, Kiran Bora, Alexandra K. Blomfield, Jasmine Lin, Anne B. Fulton, Jing Chen, and Pankaj B. Agrawal. Hbs1l deficiency causes retinal dystrophy in a child and in a mouse model associated with defective development of photoreceptor cells. Disease Models & Mechanisms, Jul 2024. URL: https://doi.org/10.1242/dmm.050557, doi:10.1242/dmm.050557. This article has 2 citations and is from a domain leading peer-reviewed journal.
-
(luo2024hbs1ldeficiencycauses pages 4-5): Shiyu Luo, Bilal Alwattar, Qifei Li, Kiran Bora, Alexandra K. Blomfield, Jasmine Lin, Anne B. Fulton, Jing Chen, and Pankaj B. Agrawal. Hbs1l deficiency causes retinal dystrophy in a child and in a mouse model associated with defective development of photoreceptor cells. Disease Models & Mechanisms, Jul 2024. URL: https://doi.org/10.1242/dmm.050557, doi:10.1242/dmm.050557. This article has 2 citations and is from a domain leading peer-reviewed journal.
-
(luo2023geneticdeficiencyof pages 6-8): Shiyu Luo, Bilal Alwattar, Qifei Li, Kiran Bora, Alexandra K. Blomfield, Jasmine Lin, Anne B. Fulton, Jing Chen, and Pankaj B. Agrawal. Genetic deficiency of ribosomal rescue factor hbs1l causes retinal dystrophy associated with pelota and edf1 depletion. bioRxiv, Oct 2023. URL: https://doi.org/10.1101/2023.10.18.562924, doi:10.1101/2023.10.18.562924. This article has 1 citations.
-
(OpenTargets Search: -EDF1): Open Targets Query (-EDF1, 5 results). Buniello, A. et al. (2025). Open Targets Platform: facilitating therapeutic hypotheses building in drug discovery. Nucleic Acids Research.
-
(rajagopalan2024mineralocorticoidreceptorantagonism pages 1-3): Sanjay Rajagopalan, Mirela Dobre, Jean-Eudes Dazard, Armando Vergara-Martel, Kim Connelly, Michael E. Farkouh, Juan Gaztanaga, Heather Conger, Ann Dever, Laleh Razavi-Nematollahi, Anas Fares, Gabriel Pereira, Jonnelle Edwards-Glenn, Mark Cameron, Cheryl Cameron, Sadeer Al-Kindi, Robert D. Brook, Bertram Pitt, and Matthew Weir. Mineralocorticoid receptor antagonism prevents aortic plaque progression and reduces left ventricular mass and fibrosis in patients with type 2 diabetes and chronic kidney disease: the magma trial. Circulation, 150:663-676, Aug 2024. URL: https://doi.org/10.1161/circulationaha.123.067620, doi:10.1161/circulationaha.123.067620. This article has 21 citations and is from a highest quality peer-reviewed journal.
-
(kabe1999theroleof pages 7-8): Yasuaki Kabe, Masahide Goto, Daisuke Shima, Takeshi Imai, Tadashi Wada, Ken-ichirou Morohashi, Masahiro Shirakawa, Susumu Hirose, and Hiroshi Handa. The role of human mbf1 as a transcriptional coactivator*. The Journal of Biological Chemistry, 274:34196-34202, Nov 1999. URL: https://doi.org/10.1074/jbc.274.48.34196, doi:10.1074/jbc.274.48.34196. This article has 117 citations.
Artifacts
Citations
- kabe1999theroleof pages 5-6
- kim2024multiproteinbridgingfactor pages 10-11
- mariotti2000interactionbetweenendothelial pages 2-4
- mariotti2004thedualrole pages 1-2
- brandt2024longnoncodingrnas pages 9-10
- luo2023geneticdeficiencyof pages 6-8
- brandt2024longnoncodingrnas pages 4-5
- rajagopalan2024mineralocorticoidreceptorantagonism pages 1-3
- kabe1999theroleof pages 2-3
- kabe1999theroleof pages 1-1
- kabe1999theroleof pages 1-2
- kabe1999theroleof pages 6-7
- kabe1999theroleof pages 7-8
- https://doi.org/10.1074/jbc.274.48.34196;
- https://doi.org/10.7554/eLife.58828;
- https://doi.org/10.1016/j.molcel.2024.10.029;
- https://doi.org/10.1074/jbc.M001928200;
- https://doi.org/10.1007/s00018-004-4016-0;
- https://doi.org/10.3390/ijms25115678;
- https://doi.org/10.1242/dmm.050557;
- https://doi.org/10.1101/2023.10.18.562924;
- https://doi.org/10.1161/CIRCULATIONAHA.123.067620;
- https://doi.org/10.7554/eLife.58828
- https://doi.org/10.1074/jbc.274.48.34196
- https://doi.org/10.1007/s00018-004-4016-0
- https://doi.org/10.1016/j.molcel.2024.10.029
- https://doi.org/10.1074/jbc.274.48.34196,
- https://doi.org/10.7554/elife.58828,
- https://doi.org/10.1074/jbc.m001928200,
- https://doi.org/10.1016/j.molcel.2024.10.029,
- https://doi.org/10.1007/s00018-004-4016-0,
- https://doi.org/10.3390/ijms25115678,
- https://doi.org/10.1242/dmm.050557,
- https://doi.org/10.1101/2023.10.18.562924,
- https://doi.org/10.1161/circulationaha.123.067620,
OpenAI
(EDF1-deep-research-openai.md)
OpenAI
(EDF1-deep-research-openai.md)Functional Annotation of Human Endothelial Differentiation-Related Factor 1 (EDF1)
Gene Identity and Conservation
Endothelial differentiation-related factor 1 (EDF1), also known as Multiprotein Bridging Factor-1 (MBF1), is a small (~16 kDa) protein encoded by the human EDF1 gene (UniProt O60869). It was initially identified in the context of endothelial cell differentiation and is highly conserved across species (pmc.ncbi.nlm.nih.gov). The amino acid sequence of EDF1/MBF1 shows substantial conservation from yeast to humans (e.g. ~62% identity between human and silkworm MBF1) (pmc.ncbi.nlm.nih.gov), reflecting a fundamental role in cell physiology. EDF1 is classified as a transcriptional co-factor: its defining feature is the ability to form a bridge between gene-specific transcription factors and the basal transcription machinery (pmc.ncbi.nlm.nih.gov). This bridging activity is the basis for its alternate name “multiprotein bridging factor”, emphasizing its role in connecting multiple protein partners in gene regulation. EDF1 is also sometimes designated CFAP280 (cilia/flagella-associated protein 280) in databases, though its primary known functions are not cilia-specific. Its conservation and ubiquitous presence suggest an essential regulatory function maintained throughout eukaryotic evolution (pmc.ncbi.nlm.nih.gov).
Protein Structure and Domains
EDF1 is a relatively small protein composed largely of alpha-helical domains. It contains an N-terminal MBF1 domain and a C-terminal helix-turn-helix (HTH) motif characteristic of Cro/C1-type DNA-binding domains (IPR001387). Notably, EDF1 harbors an IQ motif (a short calmodulin-binding sequence), which overlaps with part of the MBF1 domain (www.abcam.com). This IQ motif allows EDF1 to bind the calcium sensor protein calmodulin (CaM). The C-terminal HTH domain (IPR013729, PF01381) is conserved in MBF1 proteins and enables DNA or nucleic acid binding in some contexts (elifesciences.org) (elifesciences.org). Indeed, structural analyses indicate that EDF1’s HTH and adjacent helices form a bundle that can interact with RNA and ribosomal components (as discussed below) (elifesciences.org) (elifesciences.org). In summary, EDF1’s domain architecture equips it with a bifunctional capacity: an N-terminal region for protein–protein interactions (with transcription factors and CaM) and a C-terminal HTH for nucleic acid or protein binding in larger complexes.
Expression and Subcellular Localization
The EDF1 gene is widely expressed in human tissues. RNA profiling data show ubiquitous expression, with especially high levels in the digestive tract (e.g. duodenum and small intestine) and consistent expression in many other tissues (www.ncbi.nlm.nih.gov). At the cellular level, EDF1 is found in both the cytoplasm and the nucleus. Under basal conditions, a significant fraction of EDF1 resides in the cytosol, often bound to calmodulin. However, upon certain stimuli EDF1 relocalizes to the nucleus (www.genecards.org). Notably, protein kinase A (PKA) activation (e.g. via forskolin treatment, which raises cAMP) causes EDF1 to be phosphorylated and accumulate in the nucleus (pubmed.ncbi.nlm.nih.gov). Phorbol ester (TPA) treatment has a similar effect, as does the binding of EDF1 to some of its partner transcription factors like NR5A1 (steroidogenic factor-1) that localize to the nucleus (www.genecards.org). In contrast, in quiescent cells EDF1 can be largely cytosolic. This dynamic localization indicates regulated shuttling: EDF1 acts as a cytosolic sensor and sequestering protein under some conditions, and as a nuclear coactivator under others. Phosphorylation by PKA modulates this balance – unphosphorylated EDF1 tends to stay in the cytoplasm bound to CaM, whereas phosphorylated EDF1 releases CaM and enters the nucleus to engage in transcriptional regulation (pubmed.ncbi.nlm.nih.gov).
Role as a Transcriptional Coactivator
In the nucleus, EDF1/MBF1 serves as a transcriptional coactivator, bridging between sequence-specific transcription activators and the general transcription machinery. It was first described in Drosophila as a cofactor for the FTZ-F1 transcription factor (an orphan nuclear receptor), facilitating FTZ-F1’s activation of target genes (academic.oup.com). This bridging function is conserved in humans: EDF1 has been shown to interact with the TATA-box binding protein (TBP), a core component of the pre-initiation complex, while simultaneously binding gene-specific activators (www.ncbi.nlm.nih.gov). By physically linking activator and TBP, EDF1 helps recruit or stabilize the transcriptional machinery at target promoters. Several studies have identified specific factors enhanced by EDF1. For example, EDF1 augments the DNA-binding and transactivation activity of certain bZIP family transcription factors like ATF1, ATF2, and CREB1 (www.genecards.org). It also acts as a coactivator for nuclear receptors: it was shown to stimulate the transcriptional activity of steroidogenic factor-1 (NR5A1) and the ligand-dependent receptors LXRα (NR1H3) and PPARγ (NR1C3) (pharos.nih.gov). These nuclear receptors regulate genes in steroid hormone biosynthesis and lipid metabolism; accordingly, an early study demonstrated that MBF1 enhances the activity of multiple lipid-metabolism regulators in this class (pubmed.ncbi.nlm.nih.gov) (pharos.nih.gov). Unlike classical coactivators such as p300/CBP, EDF1 does not have enzymatic histone acetyltransferase activity, but instead acts as an architectural tether – an adapter that brings together activator, TBP, and possibly other components. This function is essential for certain genes: for instance, in vitro experiments showed that without MBF1, an activator’s ability to stimulate a reporter gene via TBP was severely impaired (www.microbiologyresearch.org) (pmc.ncbi.nlm.nih.gov). In summary, EDF1’s primary function in the nucleus is to facilitate transcription initiation by bridging specific transcription factors to the general machinery, thereby boosting target gene expression.
Calmodulin Binding and Endothelial Cell Function
One distinctive feature of EDF1 is its interaction with calmodulin (CaM), a calcium-binding messenger protein. EDF1’s IQ motif allows it to bind CaM in a calcium-dependent manner, effectively sequestering CaM when EDF1 is in the cytoplasm (pubmed.ncbi.nlm.nih.gov). This has direct implications for endothelial cell function. In vascular endothelial cells, CaM is a crucial cofactor for endothelial nitric oxide synthase (eNOS), the enzyme that produces nitric oxide (NO). EDF1 can negatively regulate eNOS activity by competing for CaM. Under resting conditions, EDF1-bound calmodulin is not available to fully activate eNOS, thereby keeping NO release in check (pubmed.ncbi.nlm.nih.gov). Experimental studies support this model: silencing EDF1 in human endothelial cells leads to increased free CaM and enhanced NO production (pubmed.ncbi.nlm.nih.gov). Bolognese et al. (2010) reported that endothelial cells with shRNA-mediated EDF1 knockdown showed significantly higher NO output, which could be reversed by a CaM inhibitor, indicating that the effect was indeed through freed calmodulin activating eNOS (pubmed.ncbi.nlm.nih.gov). Interestingly, the loss of EDF1 in these cells also accelerated their organization into capillary-like networks (a sign of differentiation) and slowed their proliferation (pubmed.ncbi.nlm.nih.gov). Consistent with this, EDF1 levels are lower in quiescent or senescent endothelial cells and highest in actively proliferating endothelial cells (pubmed.ncbi.nlm.nih.gov). Thus, EDF1 appears to restrain full endothelial differentiation while promoting proliferation, in part by limiting NO signaling. Upon pro-angiogenic stimulation, this restraint is relieved: for example, vascular endothelial growth factor (VEGF) triggers a rise in endothelial Ca²⁺ that causes CaM to dissociate from EDF1 (pubmed.ncbi.nlm.nih.gov). VEGF treatment does not change total EDF1 levels, but it causes EDF1 to release CaM, which then binds to eNOS, coinciding with a burst of NO production (pubmed.ncbi.nlm.nih.gov). In parallel, as CaM is released and calcium levels rise, EDF1 can translocate to the nucleus (especially if PKA or other pathways phosphorylate it) (pubmed.ncbi.nlm.nih.gov). In the nucleus, it may then coactivate transcription of genes involved in angiogenesis or cell growth. In summary, EDF1 serves a dual role in endothelial cells: in the cytosol it is a CaM-binding protein that tonically represses NO synthesis and differentiation, and in the nucleus it can act as a coactivator for genes that promote endothelial cell proliferation and angiogenic responses (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov).
Involvement in Cardiomyocyte Hypertrophy
Beyond the endothelium, EDF1/MBF1 plays a role in the heart, particularly in the context of cardiac hypertrophy. Cardiac (ventricular) hypertrophy is an adaptive response to stress (e.g. hypertension or hormonal stimulation) characterized by enlarged cardiomyocyte size and reactivation of fetal cardiac genes. A study by Franco et al. found that MBF1 expression is upregulated during cardiomyocyte hypertrophy in vitro and in animal models (pubmed.ncbi.nlm.nih.gov). Cultured heart cells stimulated with phenylephrine (a hypertrophic agonist) showed increased MBF1 levels, and similarly, mice subjected to hypertrophic stimuli (angiotensin II infusion or pressure overload by aortic banding) had elevated cardiac MBF1 protein (pubmed.ncbi.nlm.nih.gov). Functionally, MBF1 is required for the hypertrophic gene program: using antisense oligonucleotides to knock down MBF1 markedly blocked the hypertrophic growth of cardiomyocytes in response to phenylephrine (pubmed.ncbi.nlm.nih.gov). Conversely, overexpression of MBF1 enhanced the activation of hypertrophy-associated genes such as atrial natriuretic peptide (ANP) under hormonal stimulation (pubmed.ncbi.nlm.nih.gov). Mechanistically, MBF1 was found to cooperate with the AP-1 transcription factor c-Jun in this process (pubmed.ncbi.nlm.nih.gov). c-Jun (part of the AP-1 complex) is known to drive expression of genes during hypertrophy; MBF1 likely bridges c-Jun to the basal machinery, boosting transcription of genes like ANP. These findings indicate that EDF1/MBF1 is a key co-factor in hormone-induced cardiomyocyte hypertrophy, linking neurohumoral signals to the genomic response in heart muscle. This aligns with its general role as a coactivator: in cardiomyocytes, it amplifies the effect of pro-hypertrophic transcription factors. Its inducibility and necessity in hypertrophy suggest that EDF1 could be a potential mediator of pathological cardiac remodeling. Indeed, one could speculate that targeting EDF1-MBF1 interactions might modulate the hypertrophic response, although no direct therapies exist yet.
Interaction with Metabolic Regulators
EDF1’s coactivator function also extends to metabolic regulation. As noted above, it can bind and stimulate nuclear receptors such as PPARγ and LXRα (pharos.nih.gov), which are master regulators of lipid metabolism and storage. A 2002 study in Molecular Endocrinology demonstrated that human MBF1 enhances the transcriptional activity of several non-steroid nuclear receptors involved in cholesterol and fatty acid metabolism (pubmed.ncbi.nlm.nih.gov). For instance, LXRα controls genes in cholesterol efflux and transport, while PPARγ activates adipogenic genes; MBF1’s presence boosts the expression of their target genes. In practical terms, EDF1 might influence processes like adipocyte differentiation or liver lipid homeostasis via these pathways. Additionally, EDF1 was shown to enhance the activity of SF-1 (NR5A1) (pharos.nih.gov), a nuclear receptor that regulates steroid hormone biosynthesis and certain aspects of lipid metabolism in endocrine tissues. Thus, EDF1 serves as a common co-factor linking diverse metabolic transcription factors to effective transcription. However, the physiological impact of EDF1 on metabolism in vivo remains to be fully elucidated. Given its ubiquitous expression, any metabolic phenotype of EDF1 dysfunction might be subtle or context-dependent. Some large-scale studies have annotated EDF1 with Gene Ontology terms like “lipid metabolism,” but direct experimental evidence (such as metabolic profiling of an EDF1 knockout) has not been widely reported. Nonetheless, the molecular interactions suggest that EDF1 could modulate metabolic gene networks in tissues like adipose, liver, and steroidogenic organs, by ensuring robust transcriptional activation by key metabolic regulators (pharos.nih.gov).
Role in Ribosome Quality Control and Stress Response
One of the most exciting recent developments (2020 onwards) in understanding EDF1 is the discovery of its role in ribosome-associated quality control and the integrated stress response. While historically known as a transcription factor coactivator, EDF1 has now been implicated in managing stalled ribosomes and maintaining translational fidelity. Studies in yeast and human cells found that EDF1 (and its yeast homolog Mbf1) is recruited to collided ribosomes – situations where multiple ribosomes jam on an mRNA due to a translational stall (elifesciences.org). Cryo-electron microscopy mapping shows EDF1/Mbf1 binding at the interface of two collided ribosomes, near the mRNA entry channel of the 40S subunit (elifesciences.org) (elifesciences.org). In this position, EDF1 acts as a sensor and mediator of the collision response. Sinha et al. (2020) showed that EDF1 recruits the translational repressors GIGYF2 and eIF4E2 (also known as the 4EHP–GIGYF2 complex) to the stalled ribosome complex (elifesciences.org). By bringing in this complex, EDF1 helps initiate a negative-feedback mechanism that prevents new ribosomes from loading onto the defective mRNA (elifesciences.org). In other words, EDF1 helps shut down translation initiation on messages that are broken or stalled, which is part of a process called No-Go Decay/Ribosome Quality Control (RQC). Consistently, cells lacking EDF1/Mbf1 show aberrant translation re-initiation and frameshifting on problematic mRNAs (elifesciences.org) (elifesciences.org), indicating that EDF1 normally prevents such errors by stabilizing the stalled ribosome in a conformation that halts translation and signals for rescue. Indeed, EDF1/Mbf1 binding to collided ribosomes was found to physically block the mRNA path and displace certain ribosomal proteins, thereby acting as a “clamp” to stop ribosomes from proceeding on a damaged template (elifesciences.org) (elifesciences.org).
Beyond halting local translation, EDF1 activates cellular stress responses stemming from ribosome collisions. Recent work has shown that Mbf1 is required to fully activate the Integrated Stress Response (ISR) in yeast and mammals (www.sciencedirect.com). The ISR is a conserved pathway where the kinase GCN2 (in yeast) or analogous eIF2α kinases in mammals detect translation stress and phosphorylate eIF2α, attenuating global protein synthesis and inducing stress-responsive genes. In yeast, deletion of MBF1 leads to blunted activation of GCN2: cells lacking Mbf1 have significantly lower eIF2α phosphorylation under stress despite the presence of collided ribosomes (www.sciencedirect.com) (www.sciencedirect.com). Without Mbf1, the downstream induction of GCN4 (a transcription factor produced upon eIF2α phosphorylation) is impaired, and the entire GCN4-dependent gene regulon is under-expressed during stress (www.sciencedirect.com) (www.sciencedirect.com). These defects resemble the phenotype of a GCN2 knockout, suggesting Mbf1 is an upstream activator of GCN2. Mechanistically, Mbf1 appears to cooperate with the known ribosome collision sensor GCN1 to stimulate GCN2 when collisions occur (www.sciencedirect.com) (www.sciencedirect.com). In fact, Wang et al. (2018) and Tesina et al. (2020) earlier reported that Mbf1 and the ribosomal protein Asc1/RACK1 act together to prevent +1 frameshifting and to promote appropriate stalling signals for GCN2 (elifesciences.org) (elifesciences.org). Building on that, a 2024 study in Molecular Cell concluded that Mbf1/EDF1 is a “core” factor for collision-induced stress signaling: it links the mechanical event of ribosome stalling to the biochemical activation of the ISR kinase (www.sciencedirect.com). Notably, that study found Mbf1’s traditional transcription coactivator role is not required for the stress response – when GCN4 was expressed constitutively (bypassing the need for translation control), Mbf1 deletion no longer affected stress gene induction (www.sciencedirect.com). Instead, Mbf1’s critical function is at the ribosome: facilitating robust GCN2 activation and subsequent eIF2α phosphorylation during stress (www.sciencedirect.com) (www.sciencedirect.com). Structurally, the N-terminal region of EDF1/Mbf1 that binds the collided ribosome was shown to be essential for this signaling, as mutations that disrupt ribosome binding also compromise GCN2 activation (www.sciencedirect.com). Thus, EDF1 serves as a molecular linchpin in the ribosome surveillance pathway – it not only halts aberrant protein synthesis but also triggers cellular stress remediation programs (both translational arrest via eIF2α phosphorylation and an “immediate early” transcriptional response to stress) (elifesciences.org). This dual action ensures proteostasis is maintained when cells encounter translation errors or damage.
Biological and Clinical Significance
Through its multiple roles, EDF1 integrates into several critical biological pathways. In the nucleus it participates in gene expression programs for development, metabolism, and stress, while in the cytosol it modulates signaling pathways like Ca²⁺/calmodulin–NO signaling and ribosome-associated stress signaling. The pleiotropic effects of EDF1 are increasingly being understood in specific physiological contexts:
-
Vascular function: By regulating nitric oxide production in endothelial cells, EDF1 can influence blood vessel dilation, angiogenesis, and vascular remodeling. Knockdown experiments suggest that lowering EDF1 raises NO levels and promotes endothelial differentiation, which could be beneficial for repairing blood vessels (pubmed.ncbi.nlm.nih.gov). On the other hand, excessive NO can be deleterious; thus, EDF1 may act as a brake to prevent unwarranted NO release. Its expression is required for proper endothelial proliferation and organization (pubmed.ncbi.nlm.nih.gov), implicating EDF1 in maintaining vascular integrity. These findings hint that EDF1 could play a role in cardiovascular diseases: for instance, in atherosclerosis or thrombosis, where endothelial dysfunction is key, the balance of EDF1 and NO might be a factor (though direct clinical correlations remain to be investigated).
-
Cardiac hypertrophy: EDF1 (MBF1) is clearly induced in hypertrophic hearts (pubmed.ncbi.nlm.nih.gov), and it appears necessary for the full hypertrophic gene expression response to neurohormonal stimuli (pubmed.ncbi.nlm.nih.gov). This makes it a potential marker or mediator in cardiac stress. Some have proposed it as part of the network controlling fetal gene reactivation in heart failure. While not yet a clinical target, EDF1’s cooperation with c-Jun/AP-1 in cardiomyocytes links it to pathways (like MAPK and adrenergic signaling) known to drive heart disease (pubmed.ncbi.nlm.nih.gov).
-
Metabolism: Through nuclear receptors like PPARγ and LXRα, EDF1 could influence metabolic syndrome components. For example, PPARγ is a drug target in type 2 diabetes (thiazolidinediones activate PPARγ); if EDF1 amplifies PPARγ activity, variations in EDF1 levels might affect adipogenesis or insulin sensitivity. Similarly, LXRα helps clear cholesterol; EDF1 might enhance LXR-driven anti-atherogenic genes (ABCA1, etc.). More research is needed to connect EDF1 with metabolic phenotypes, but its coactivator role places it at key nodes of metabolic regulation (pharos.nih.gov).
-
Protein homeostasis and neurodegeneration: The newly discovered role of EDF1 in ribosomal quality control may have implications for diseases caused by protein misfolding or translational stress. For instance, neurodegenerative diseases often involve stress granule formation and ISR activation. Indeed, mutations in tRNA or ribosome recycling factors that elevate ribosome collisions can lead to neurological disorders (www.sciencedirect.com). EDF1’s action in preventing frameshifts and activating rescue pathways suggests it might be protective in such settings. There is emerging evidence that if this system fails (e.g., in EDF1/MBF1 loss-of-function conditions), cells are less able to handle proteotoxic stress (www.sciencedirect.com) (www.sciencedirect.com). While no human diseases have yet been directly linked to mutations in EDF1, its categorization as “core ISR factor” (www.sciencedirect.com) raises the possibility that it could be a vulnerability factor in conditions from viral infection (where ISR is triggered) to cancer (tumors experience translation stress) – or conversely, a target to modulate these responses.
-
Viral interactions: Interestingly, one report indicates that HIV-1 Tat protein can downregulate EDF1 expression in endothelial cells (www.ncbi.nlm.nih.gov). Tat is known to cause vascular dysfunction in HIV patients; by suppressing EDF1, Tat might lead to excess NO release or aberrant endothelial behavior, contributing to HIV-related vascular pathology. This is a specific example of how pathogens might exploit EDF1’s pathway.
In summary, EDF1 is a multifaceted regulatory protein that links several major cellular processes – transcription, signal transduction, and translation surveillance. Its ability to interface with different partners (TBP at gene promoters, CaM and eNOS in the cytosol, ribosomes and GCN2 in the cytoplasm) is unique and underscores a unifying theme: EDF1 is a bridge between molecular systems. As one group of experts described, EDF1 “plays a central role in facilitating multiple steps” of cellular stress responses, acting upstream of key stress sensors and downstream of collision signals (elifesciences.org) (www.sciencedirect.com). Likewise, decades earlier it was recognized as an evolutionarily conserved coactivator essential for bridging certain activators to TBP (pmc.ncbi.nlm.nih.gov). These expert analyses convey that the current understanding of EDF1 is as a versatile adapter protein – one that cells utilize in different contexts to ensure proper communication between signaling pathways and gene expression outcomes. Ongoing research (particularly recent work in 2023–2024) is expanding our appreciation of EDF1, from a TBP tether to a guardian of the genome’s translational output (www.sciencedirect.com). This breadth of function makes EDF1 an intriguing subject for further study, as well as a potential node for therapeutic intervention in diseases where these pathways go awry.
References: Publications and data supporting this overview include: Takemaru et al., 1997 (PNAS) (pmc.ncbi.nlm.nih.gov); Bolognese et al., 2004 (J. Biol. Chem.) (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov); Bolognese et al., 2010 (Cardiovasc. Res.) (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov); Franco et al., 2003 (J. Biol. Chem.) (pubmed.ncbi.nlm.nih.gov); Liu et al., 2002 (Mol. Endocrinol.) (pubmed.ncbi.nlm.nih.gov); Sinha et al., 2020 (eLife) (elifesciences.org); and Efremov et al., 2024 (Molecular Cell) (www.sciencedirect.com) (www.sciencedirect.com), among others. These sources provide detailed experimental evidence for EDF1’s functions, interactions, and regulatory importance as discussed above.
Citations
- AnnotationURLCitation(end_index=558, start_index=434, title='Multiprotein bridging factor 1 (MBF1) is an evolutionarily conserved transcriptional coactivator that connects a regulatory factor and TATA element-binding\u2009protein - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC23807/#:~:text=showed%20that%20the%20MBF1%20sequence,mays')
- AnnotationURLCitation(end_index=826, start_index=702, title='Multiprotein bridging factor 1 (MBF1) is an evolutionarily conserved transcriptional coactivator that connects a regulatory factor and TATA element-binding\u2009protein - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC23807/#:~:text=showed%20that%20the%20MBF1%20sequence,mays')
- AnnotationURLCitation(end_index=1226, start_index=1068, title='The plant MBF1 protein family: a bridge between stress and transcription - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC7094072/#:~:text=MBF1%20proteins%20and%20stress%20responses,MBF1%20belong%20to%20the%20bZIP')
- AnnotationURLCitation(end_index=1812, start_index=1688, title='Multiprotein bridging factor 1 (MBF1) is an evolutionarily conserved transcriptional coactivator that connects a regulatory factor and TATA element-binding\u2009protein - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC23807/#:~:text=showed%20that%20the%20MBF1%20sequence,mays')
- AnnotationURLCitation(end_index=2364, start_index=2204, title='EDF1 | Abcam', type='url_citation', url='https://www.abcam.com/en-us/targets/edf1/19619#:~:text=JavaScript%20is%20disabled%20in%20your,calmodulin%20binding%2C%20overlaps%20with%20the')
- AnnotationURLCitation(end_index=2706, start_index=2579, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=In%20these%20structures%2C%20the%20C,domain%20of%20both')
- AnnotationURLCitation(end_index=2840, start_index=2707, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=From%20uS3%2C%20EDF1%20and%20Mbf1,Figure%203E%2C%20top%20four')
- AnnotationURLCitation(end_index=3142, start_index=3003, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=%23%23%23%20Structural%20analysis%20of%20ribosome,EDF1%20and%20Mbf1')
- AnnotationURLCitation(end_index=3275, start_index=3143, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=interacting%20with%20EDF1%20and%20Mbf1,Figure%204A%E2%80%93B')
- AnnotationURLCitation(end_index=3977, start_index=3815, title='EDF1 endothelial differentiation related factor 1 [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene/8721#:~:text=Expression%20Ubiquitous%20expression%20in%20duodenum,Try%20the%20new%20Transcript%20table')
- AnnotationURLCitation(end_index=4350, start_index=4227, title='EDF1 Gene - GeneCards | EDF1 Protein | EDF1 Antibody', type='url_citation', url='https://www.genecards.org/cgi-bin/carddisp.pl?gene=EDF1#:~:text=,cells%20with%20TPA%20or%20forskolin')
- AnnotationURLCitation(end_index=4659, start_index=4512, title='The dual role of endothelial differentiation-related factor-1 in the cytosol and nucleus: modulation by protein kinase A - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/15112053/#:~:text=vitro%20and%20in%20vivo%20phosphorylated,in%20the%20nucleus%20as%20a')
- AnnotationURLCitation(end_index=4974, start_index=4851, title='EDF1 Gene - GeneCards | EDF1 Protein | EDF1 Antibody', type='url_citation', url='https://www.genecards.org/cgi-bin/carddisp.pl?gene=EDF1#:~:text=,cells%20with%20TPA%20or%20forskolin')
- AnnotationURLCitation(end_index=5597, start_index=5450, title='The dual role of endothelial differentiation-related factor-1 in the cytosol and nucleus: modulation by protein kinase A - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/15112053/#:~:text=vitro%20and%20in%20vivo%20phosphorylated,in%20the%20nucleus%20as%20a')
- AnnotationURLCitation(end_index=6077, start_index=5983, title='Multiprotein Bridging Factor-1 (MBF-1) Is a Cofactor for Nuclear Receptors that Regulate Lipid Metabolism | Molecular Endocrinology | Oxford Academic', type='url_citation', url='https://academic.oup.com/mend/article/16/6/1367/2741839#:~:text=Abstract')
- AnnotationURLCitation(end_index=6420, start_index=6301, title='EDF1 endothelial differentiation related factor 1 [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene/8721#:~:text=This%20gene%20encodes%20a%20protein,Expression')
- AnnotationURLCitation(end_index=6931, start_index=6761, title='EDF1 Gene - GeneCards | EDF1 Protein | EDF1 Antibody', type='url_citation', url='https://www.genecards.org/cgi-bin/carddisp.pl?gene=EDF1#:~:text=Transcriptional%20coactivator%20stimulating%20NR5A1%20and,%28%20EDF1_HUMAN%2CO60869')
- AnnotationURLCitation(end_index=7328, start_index=7157, title='Pharos : Target Details - EDF1', type='url_citation', url='https://pharos.nih.gov/targets/EDF1#:~:text=Transcriptional%20coactivator%20stimulating%20NR5A1%20and,general%20transcription%20factor%20TATA%20element')
- AnnotationURLCitation(end_index=7704, start_index=7550, title='Multiprotein bridging factor-1 (MBF-1) is a cofactor for nuclear receptors that regulate lipid metabolism - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12040021/#:~:text=Multiprotein%20bridging%20factor,are%20implicated%20in%20lipid%20metabolism')
- AnnotationURLCitation(end_index=7876, start_index=7705, title='Pharos : Target Details - EDF1', type='url_citation', url='https://pharos.nih.gov/targets/EDF1#:~:text=Transcriptional%20coactivator%20stimulating%20NR5A1%20and,general%20transcription%20factor%20TATA%20element')
- AnnotationURLCitation(end_index=8495, start_index=8311, title='Differential expression and interaction of transcription co-activator MBF1 with TATA-binding protein (TBP) in the apicomplexan Cryptosporidium parvum | Microbiology Society', type='url_citation', url='https://www.microbiologyresearch.org/content/journal/micro/10.1099/mic.0.26891-0#:~:text=Differential%20expression%20and%20interaction%20of,1%7D%20and')
- AnnotationURLCitation(end_index=8654, start_index=8496, title='The plant MBF1 protein family: a bridge between stress and transcription - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC7094072/#:~:text=MBF1%20proteins%20and%20stress%20responses,MBF1%20belong%20to%20the%20bZIP')
- AnnotationURLCitation(end_index=9335, start_index=9161, title='The dual role of endothelial differentiation-related factor-1 in the cytosol and nucleus: modulation by protein kinase A - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/15112053/#:~:text=stimulates%20nuclear%20accumulation%20of%20EDF,nucleus%20as%20a%20transcriptional%20coactivator')
- AnnotationURLCitation(end_index=9912, start_index=9738, title='The dual role of endothelial differentiation-related factor-1 in the cytosol and nucleus: modulation by protein kinase A - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/15112053/#:~:text=stimulates%20nuclear%20accumulation%20of%20EDF,nucleus%20as%20a%20transcriptional%20coactivator')
- AnnotationURLCitation(end_index=10238, start_index=10056, title='The effects of silencing EDF-1 in human endothelial cells - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/20185128/#:~:text=phosphorylation%20of%20endothelial%20NO%20synthase,ultimately%20activates%20endothelial%20NO%20synthase')
- AnnotationURLCitation(end_index=10630, start_index=10495, title='The effects of silencing EDF-1 in human endothelial cells - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/20185128/#:~:text=EDF,ultimately%20activates%20endothelial%20NO%20synthase')
- AnnotationURLCitation(end_index=10932, start_index=10803, title='The effects of silencing EDF-1 in human endothelial cells - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/20185128/#:~:text=Results%3A%20EDF,amounts%20and%20the%20degree%20of')
- AnnotationURLCitation(end_index=11207, start_index=11078, title='The effects of silencing EDF-1 in human endothelial cells - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/20185128/#:~:text=Results%3A%20EDF,amounts%20and%20the%20degree%20of')
- AnnotationURLCitation(end_index=11702, start_index=11538, title='EDF-1 contributes to the regulation of nitric oxide release in VEGF-treated human endothelial cells - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/20605058/#:~:text=%28eNOS%29%20activity.%20Because%20Endothelial%20Differentiation,1%20in%20endothelial')
- AnnotationURLCitation(end_index=12020, start_index=11856, title='EDF-1 contributes to the regulation of nitric oxide release in VEGF-treated human endothelial cells - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/20605058/#:~:text=%28eNOS%29%20activity.%20Because%20Endothelial%20Differentiation,1%20in%20endothelial')
- AnnotationURLCitation(end_index=12317, start_index=12170, title='The dual role of endothelial differentiation-related factor-1 in the cytosol and nucleus: modulation by protein kinase A - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/15112053/#:~:text=vitro%20and%20in%20vivo%20phosphorylated,in%20the%20nucleus%20as%20a')
- AnnotationURLCitation(end_index=12859, start_index=12712, title='The dual role of endothelial differentiation-related factor-1 in the cytosol and nucleus: modulation by protein kinase A - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/15112053/#:~:text=vitro%20and%20in%20vivo%20phosphorylated,in%20the%20nucleus%20as%20a')
- AnnotationURLCitation(end_index=13017, start_index=12860, title='The effects of silencing EDF-1 in human endothelial cells - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/20185128/#:~:text=Conclusions%3A%20Since%20EDF,and%2C%20therefore%2C%20to%20vascular%20integrity')
- AnnotationURLCitation(end_index=13677, start_index=13517, title='Multiprotein bridging factor 1 cooperates with c-Jun and is necessary for cardiac hypertrophy in vitro - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12729799/#:~:text=Cardiac%20hypertrophy%20is%20induced%20by,MBF1%20antisense%20oligodeoxynuclotides')
- AnnotationURLCitation(end_index=14098, start_index=13938, title='Multiprotein bridging factor 1 cooperates with c-Jun and is necessary for cardiac hypertrophy in vitro - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12729799/#:~:text=Cardiac%20hypertrophy%20is%20induced%20by,MBF1%20antisense%20oligodeoxynuclotides')
- AnnotationURLCitation(end_index=14437, start_index=14310, title='Multiprotein bridging factor 1 cooperates with c-Jun and is necessary for cardiac hypertrophy in vitro - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12729799/#:~:text=angiotensin%20II%20treatment%20and%20aortic,MBF1')
- AnnotationURLCitation(end_index=14727, start_index=14600, title='Multiprotein bridging factor 1 cooperates with c-Jun and is necessary for cardiac hypertrophy in vitro - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12729799/#:~:text=angiotensin%20II%20treatment%20and%20aortic,MBF1')
- AnnotationURLCitation(end_index=14962, start_index=14835, title='Multiprotein bridging factor 1 cooperates with c-Jun and is necessary for cardiac hypertrophy in vitro - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12729799/#:~:text=angiotensin%20II%20treatment%20and%20aortic,MBF1')
- AnnotationURLCitation(end_index=16109, start_index=15938, title='Pharos : Target Details - EDF1', type='url_citation', url='https://pharos.nih.gov/targets/EDF1#:~:text=Transcriptional%20coactivator%20stimulating%20NR5A1%20and,general%20transcription%20factor%20TATA%20element')
- AnnotationURLCitation(end_index=16526, start_index=16372, title='Multiprotein bridging factor-1 (MBF-1) is a cofactor for nuclear receptors that regulate lipid metabolism - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12040021/#:~:text=Multiprotein%20bridging%20factor,are%20implicated%20in%20lipid%20metabolism')
- AnnotationURLCitation(end_index=17073, start_index=16902, title='Pharos : Target Details - EDF1', type='url_citation', url='https://pharos.nih.gov/targets/EDF1#:~:text=Transcriptional%20coactivator%20stimulating%20NR5A1%20and,general%20transcription%20factor%20TATA%20element')
- AnnotationURLCitation(end_index=18139, start_index=17968, title='Pharos : Target Details - EDF1', type='url_citation', url='https://pharos.nih.gov/targets/EDF1#:~:text=Transcriptional%20coactivator%20stimulating%20NR5A1%20and,general%20transcription%20factor%20TATA%20element')
- AnnotationURLCitation(end_index=18891, start_index=18752, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=structures%20of%20EDF1%20and%20its,EDF1%20functions%20upstream%20of')
- AnnotationURLCitation(end_index=19151, start_index=19041, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=Cryo,feedback%20loop%20that%20prevents')
- AnnotationURLCitation(end_index=19250, start_index=19152, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=,are%20shown%20as%20models')
- AnnotationURLCitation(end_index=19610, start_index=19503, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=collision%20interface,Our%20results')
- AnnotationURLCitation(end_index=19867, start_index=19760, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=collision%20interface,Our%20results')
- AnnotationURLCitation(end_index=20327, start_index=20168, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=previous%20genetic%20studies%20had%20implicated,in%20our%20polysome%20proteomics%20data')
- AnnotationURLCitation(end_index=20460, start_index=20328, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=interacting%20with%20EDF1%20and%20Mbf1,Figure%204A%E2%80%93B')
- AnnotationURLCitation(end_index=20984, start_index=20831, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=proteomic%20analysis%20of%20the%20yeast,2018%3B%20Wolf%20and%20Grayhack%2C%202015')
- AnnotationURLCitation(end_index=21141, start_index=20985, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=we%20observed%20Mbf1%20occupying%20the,associated%20with%20the%20collided%20ribosome')
- AnnotationURLCitation(end_index=21556, start_index=21382, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=ribosomes%2C%20a%20known%20activator%20of,by%20acting%20as%20a%20direct')
- AnnotationURLCitation(end_index=22159, start_index=21976, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=However%2C%20contrary%20to%20initial%20studies,of%20eIF2%CE%B1%20by%20Gcn2%20and')
- AnnotationURLCitation(end_index=22327, start_index=22160, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=important%20for%20Gcn2%20activation,Collectively%2C%20our%20data')
- AnnotationURLCitation(end_index=22717, start_index=22526, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=required%20for%20robust%20Gcn2%20activation,recruitment%20to%20collided%20ribosomes%20is')
- AnnotationURLCitation(end_index=22899, start_index=22718, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=Gcn2%20activation%20is%20severely%20compromised,in%20the%20absence%20of%20Mbf1')
- AnnotationURLCitation(end_index=23318, start_index=23137, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=mechanism%20of%20Gcn2%20activation%20in,less%20so%20on%20another%20coactivator')
- AnnotationURLCitation(end_index=23499, start_index=23319, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=Gcn2%20is%20activated%20by%20collided,showed%20that%20Gcn1%20binds%20collided')
- AnnotationURLCitation(end_index=23869, start_index=23716, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=proteomic%20analysis%20of%20the%20yeast,2018%3B%20Wolf%20and%20Grayhack%2C%202015')
- AnnotationURLCitation(end_index=24017, start_index=23870, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=to%20cooperate%20with%20Mbf1%20to,2018%3B%20Wolf%20and%20Grayhack%2C%202015')
- AnnotationURLCitation(end_index=24431, start_index=24257, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=ribosomes%2C%20a%20known%20activator%20of,by%20acting%20as%20a%20direct')
- AnnotationURLCitation(end_index=24884, start_index=24693, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=required%20for%20robust%20Gcn2%20activation,recruitment%20to%20collided%20ribosomes%20is')
- AnnotationURLCitation(end_index=25210, start_index=25027, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=otherwise%20seemingly%20disparate%20functions%20of,by%20acting%20as%20a%20direct')
- AnnotationURLCitation(end_index=25378, start_index=25211, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=important%20for%20Gcn2%20activation,Collectively%2C%20our%20data')
- AnnotationURLCitation(end_index=25750, start_index=25583, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=important%20for%20Gcn2%20activation,Collectively%2C%20our%20data')
- AnnotationURLCitation(end_index=26158, start_index=26051, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=collision%20interface,Our%20results')
- AnnotationURLCitation(end_index=27222, start_index=27065, title='The effects of silencing EDF-1 in human endothelial cells - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/20185128/#:~:text=Conclusions%3A%20Since%20EDF,and%2C%20therefore%2C%20to%20vascular%20integrity')
- AnnotationURLCitation(end_index=27579, start_index=27422, title='The effects of silencing EDF-1 in human endothelial cells - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/20185128/#:~:text=Conclusions%3A%20Since%20EDF,and%2C%20therefore%2C%20to%20vascular%20integrity')
- AnnotationURLCitation(end_index=28147, start_index=27987, title='Multiprotein bridging factor 1 cooperates with c-Jun and is necessary for cardiac hypertrophy in vitro - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12729799/#:~:text=Cardiac%20hypertrophy%20is%20induced%20by,MBF1%20antisense%20oligodeoxynuclotides')
- AnnotationURLCitation(end_index=28377, start_index=28250, title='Multiprotein bridging factor 1 cooperates with c-Jun and is necessary for cardiac hypertrophy in vitro - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12729799/#:~:text=angiotensin%20II%20treatment%20and%20aortic,MBF1')
- AnnotationURLCitation(end_index=28842, start_index=28715, title='Multiprotein bridging factor 1 cooperates with c-Jun and is necessary for cardiac hypertrophy in vitro - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12729799/#:~:text=angiotensin%20II%20treatment%20and%20aortic,MBF1')
- AnnotationURLCitation(end_index=29583, start_index=29412, title='Pharos : Target Details - EDF1', type='url_citation', url='https://pharos.nih.gov/targets/EDF1#:~:text=Transcriptional%20coactivator%20stimulating%20NR5A1%20and,general%20transcription%20factor%20TATA%20element')
- AnnotationURLCitation(end_index=30196, start_index=30011, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=match%20at%20L129%20elevated%20ribosome,ribosomes%2C%20and%20in%20particular%20the')
- AnnotationURLCitation(end_index=30658, start_index=30475, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=However%2C%20contrary%20to%20initial%20studies,of%20eIF2%CE%B1%20by%20Gcn2%20and')
- AnnotationURLCitation(end_index=30850, start_index=30659, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=required%20for%20robust%20Gcn2%20activation,recruitment%20to%20collided%20ribosomes%20is')
- AnnotationURLCitation(end_index=31148, start_index=30974, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=ribosomes%2C%20a%20known%20activator%20of,by%20acting%20as%20a%20direct')
- AnnotationURLCitation(end_index=31613, start_index=31520, title='EDF1 endothelial differentiation related factor 1 [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene/8721#:~:text=Protein%20%20,PubMed')
- AnnotationURLCitation(end_index=32631, start_index=32492, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=structures%20of%20EDF1%20and%20its,EDF1%20functions%20upstream%20of')
- AnnotationURLCitation(end_index=32806, start_index=32632, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=ribosomes%2C%20a%20known%20activator%20of,by%20acting%20as%20a%20direct')
- AnnotationURLCitation(end_index=33104, start_index=32946, title='The plant MBF1 protein family: a bridge between stress and transcription - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC7094072/#:~:text=MBF1%20proteins%20and%20stress%20responses,MBF1%20belong%20to%20the%20bZIP')
- AnnotationURLCitation(end_index=33690, start_index=33516, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=ribosomes%2C%20a%20known%20activator%20of,by%20acting%20as%20a%20direct')
- AnnotationURLCitation(end_index=34098, start_index=33974, title='Multiprotein bridging factor 1 (MBF1) is an evolutionarily conserved transcriptional coactivator that connects a regulatory factor and TATA element-binding\u2009protein - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC23807/#:~:text=showed%20that%20the%20MBF1%20sequence,mays')
- AnnotationURLCitation(end_index=34289, start_index=34142, title='The dual role of endothelial differentiation-related factor-1 in the cytosol and nucleus: modulation by protein kinase A - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/15112053/#:~:text=vitro%20and%20in%20vivo%20phosphorylated,in%20the%20nucleus%20as%20a')
- AnnotationURLCitation(end_index=34464, start_index=34290, title='The dual role of endothelial differentiation-related factor-1 in the cytosol and nucleus: modulation by protein kinase A - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/15112053/#:~:text=stimulates%20nuclear%20accumulation%20of%20EDF,nucleus%20as%20a%20transcriptional%20coactivator')
- AnnotationURLCitation(end_index=34650, start_index=34510, title='The effects of silencing EDF-1 in human endothelial cells - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/20185128/#:~:text=synthase,ultimately%20activates%20endothelial%20NO%20synthase')
- AnnotationURLCitation(end_index=34808, start_index=34651, title='The effects of silencing EDF-1 in human endothelial cells - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/20185128/#:~:text=Conclusions%3A%20Since%20EDF,and%2C%20therefore%2C%20to%20vascular%20integrity')
- AnnotationURLCitation(end_index=34976, start_index=34849, title='Multiprotein bridging factor 1 cooperates with c-Jun and is necessary for cardiac hypertrophy in vitro - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12729799/#:~:text=angiotensin%20II%20treatment%20and%20aortic,MBF1')
- AnnotationURLCitation(end_index=35170, start_index=35016, title='Multiprotein bridging factor-1 (MBF-1) is a cofactor for nuclear receptors that regulate lipid metabolism - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12040021/#:~:text=Multiprotein%20bridging%20factor,are%20implicated%20in%20lipid%20metabolism')
- AnnotationURLCitation(end_index=35308, start_index=35201, title='EDF1 coordinates cellular responses to ribosome collisions | eLife', type='url_citation', url='https://elifesciences.org/articles/58828#:~:text=collision%20interface,Our%20results')
- AnnotationURLCitation(end_index=35528, start_index=35354, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=ribosomes%2C%20a%20known%20activator%20of,by%20acting%20as%20a%20direct')
- AnnotationURLCitation(end_index=35720, start_index=35529, title='Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes - ScienceDirect', type='url_citation', url='https://www.sciencedirect.com/science/article/pii/S1097276524008694#:~:text=required%20for%20robust%20Gcn2%20activation,recruitment%20to%20collided%20ribosomes%20is')