Deep Research
Falcon
(EPYC-deep-research-falcon.md)
Falcon
(EPYC-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Comprehensive Research Report: Human EPYC (Epiphycan) — Functional Annotation (UniProt: Q99645)
0) Scope and identity verification (mandatory)
The literature and database context retrieved here consistently refers to EPYC as epiphycan, also called dermatan sulfate proteoglycan 3 (DSPG3) / proteoglycan-Lb (PG-Lb), in the small leucine-rich proteoglycan (SLRP) family, matching the user-specified UniProt identity (Q99645) and domain family expectations (LRR-containing extracellular proteoglycan). EPYC is described as a dermatan sulfate (DS) SLRP found in epiphyseal cartilage, and it is characterized by a leucine-rich repeat architecture (reported as 7 LRRs in one review). (hayes2018biodiversityofcsproteoglycan pages 7-8, halari2021rolesoftwo pages 4-6)
1) Key concepts and definitions (current understanding)
1.1 Small leucine-rich proteoglycans (SLRPs) and where EPYC fits
SLRPs are extracellular matrix (ECM) proteoglycans characterized by a protein core containing leucine-rich repeats (LRRs) and, in many family members, covalently attached glycosaminoglycan (GAG) chains such as dermatan sulfate (DS) or chondroitin sulfate (CS). In a review focused on SLRPs, EPYC is explicitly listed as epiphycan, a DS/CS# SLRP, with chromosomal location 12q21.33*, and SLRPs are described as ECM-resident components involved in tissue structure and collagen organization/fibrillogenesis. (halari2021rolesoftwo pages 4-6)
A second authoritative review focused on CS/DS proteoglycans places epiphycan (EPYC) among DS SLRPs found in epiphyseal cartilage, and it specifies that epiphycan contains 7 LRRs (fewer than the 10–11 LRRs typical of many other SLRPs). (hayes2018biodiversityofcsproteoglycan pages 7-8)
1.2 EPYC/epiphycan as an ECM structural regulator rather than an enzyme
Across the retrieved sources, EPYC is treated as a structural ECM proteoglycan rather than a catalytic enzyme/transport protein. Its repeatedly cited “primary” molecular role is in collagen fibril formation (fibrillogenesis) and cartilage matrix organization, which is consistent with SLRP family biology. (hayes2018biodiversityofcsproteoglycan pages 7-8, halari2021rolesoftwo pages 4-6, yang2024epycfunctionsas pages 1-2)
2) Core biological function, processes, and localization
2.1 Subcellular/extracellular localization
EPYC is described as an extracellular matrix (ECM) proteoglycan in SLRP reviews. (halari2021rolesoftwo pages 4-6)
More recent mechanistic work in cancer contexts treats EPYC as a secreted/soluble ECM-associated factor that can be supplied as recombinant protein (rhEPYC) and can act on other cells (e.g., endothelial cells), consistent with an extracellular role. Additionally, GO/KEGG enrichment in that laryngeal-cancer study associated EPYC-containing signatures with “extracellular matrix organization,” “extracellular region,” and “ECM-receptor interaction.” (zhou2024epycpromoteslaryngeal pages 1-5, zhou2024epycpromoteslaryngeal pages 5-8)
2.2 Biological process: cartilage matrix maturation and collagen fibrillogenesis
Evidence spanning cartilage-focused and broader ECM literature supports EPYC as part of cartilage ECM organization:
- Cartilage localization: epiphycan (EPYC) is explicitly reported as a DS SLRP found in epiphyseal cartilage. (hayes2018biodiversityofcsproteoglycan pages 7-8)
- Chondrogenic matrix maturation: in a human mesenchymal stem cell (MSC) chondrogenesis model, EPYC is described as a “novel” proteoglycan that appears in a later maturation phase of matrix development, with epiphycan described as characteristic of articular cartilage in that experimental context. (sorrell2018humanmesenchymalstem pages 10-14)
- Collagen fibril formation: a 2024 pancreatic-cancer paper reiterates a cartilage/ECM-centric concept of EPYC biology: EPYC “can regulate the fibril formation by interacting with collagen fibrils and other extracellular matrix proteins.” (yang2024epycfunctionsas pages 1-2)
Together, these sources support a functional annotation in which EPYC/epiphycan is a secreted ECM SLRP that contributes to collagen fibrillogenesis and cartilage ECM assembly, influencing chondrogenesis/matrix maturation. (hayes2018biodiversityofcsproteoglycan pages 7-8, sorrell2018humanmesenchymalstem pages 10-14, yang2024epycfunctionsas pages 1-2)
2.3 Tissue and cell-type expression highlights (including 2024 developments)
A 2024 preprint studying growth plate resting-zone biology used Epyc as a marker in scRNAseq-based tissue annotation and reports Epyc specifically expressed in growth plate chondrocytes and absent from articular chondrocytes and perichondrial cells (in the murine tissue analyzed). This supports a strong association with growth plate/epiphyseal chondrocytes, aligning with the epiphyseal cartilage localization reported in the proteoglycan review. (otsuru2024apolipoproteineis pages 1-4, hayes2018biodiversityofcsproteoglycan pages 7-8)
Note: another experimental system (human MSC chondrogenesis) described epiphycan as “characteristic of articular cartilage,” illustrating that EPYC expression can depend on developmental stage/model and that “cartilage” can encompass distinct zones with different gene programs. (sorrell2018humanmesenchymalstem pages 10-14, otsuru2024apolipoproteineis pages 1-4)
3) Recent developments and latest research (prioritizing 2023–2024)
Direct 2023–2024 mechanistic studies focused on EPYC in human cartilage biology were limited in the retrieved corpus; however, multiple 2024 studies substantially expand EPYC’s disease-context mechanistic literature, particularly in oncology, while still citing/leveraging EPYC’s ECM/collagen biology.
3.1 2024: EPYC as a mechanistic driver of angiogenesis and Wnt/VEGF signaling in laryngeal cancer
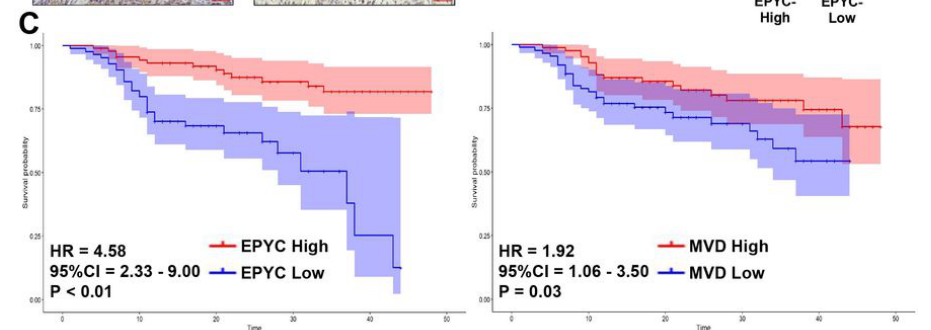
A 2024 Research Square preprint reports EPYC as an angiogenesis-related hub gene in laryngeal cancer and provides mechanistic evidence that EPYC promotes carcinogenesis and angiogenesis via Wnt/Axin/β-catenin and VEGFR1/VEGF axes:
- EPYC knockdown increased Axin and decreased nuclear β-catenin, consistent with EPYC promoting Wnt/β-catenin signaling by suppressing Axin. (zhou2024epycpromoteslaryngeal pages 10-13, zhou2024epycpromoteslaryngeal media e7b39943)
- EPYC was proposed to bind VEGFR1, with reported binding and downstream promotion of VEGFR1 degradation (proteasome dependence supported by MG132 rescue) and effects on VEGF signaling output; the study reports a binding affinity KD ~44.1 µM. (zhou2024epycpromoteslaryngeal pages 10-13, zhou2024epycpromoteslaryngeal pages 5-8)
- Single-cell analysis in this study highlighted EPYC expression in vascular endothelial cells, consistent with an extracellular factor active in the tumor microenvironment. (zhou2024epycpromoteslaryngeal pages 10-13, zhou2024epycpromoteslaryngeal media 717e387a)
These findings represent a notable 2024 expansion of EPYC biology into extracellular regulation of growth factor receptor availability and canonical Wnt signaling, albeit in a cancer microenvironment rather than cartilage. (zhou2024epycpromoteslaryngeal pages 10-13, zhou2024epycpromoteslaryngeal pages 1-5, zhou2024epycpromoteslaryngeal media e7b39943)
3.2 2024: EPYC in pancreatic cancer via PI3K–AKT signaling
A peer-reviewed 2024 Scientific Reports paper proposes EPYC as a prognostic biomarker for pancreatic cancer and reports functional assays in which EPYC promotes proliferation “via PI3K-AKT signaling in vivo and in vitro.” The paper also re-states EPYC’s ECM role in regulating fibril formation through collagen/ECM interactions. (yang2024epycfunctionsas pages 1-2)
3.3 2024: EPYC as a candidate breast cancer biomarker (with clinical qRT-PCR)
A 2024 BMC Cancer study identified EPYC among four candidate diagnostic biomarkers derived from integrative bioinformatics and validated via qRT-PCR in 55 breast cancer patients (tumor vs adjacent non-tumor). EPYC mRNA was significantly upregulated (P < 0.0001), but the reported overall survival association for EPYC was weak/non-significant (OS HR = 1.04; log-rank P = 0.72). (golestan2024unveilingpromisingbreast pages 6-8)
4) Current applications and real-world implementations
4.1 Biomarker development in oncology (2024)
Multiple 2024 studies frame EPYC as a candidate diagnostic/prognostic biomarker in distinct cancers (breast, pancreatic, laryngeal). These implementations are currently best characterized as transcript-level biomarkers and hypothesis-generating functional targets, rather than established clinical biomarkers:
- Breast cancer: EPYC among proposed diagnostic markers, validated by qRT-PCR in paired tissues. (golestan2024unveilingpromisingbreast pages 6-8)
- Pancreatic cancer: EPYC incorporated into prognostic modeling and functional validation, suggesting potential target/biomarker status. (yang2024epycfunctionsas pages 1-2, yang2024epycfunctionsas pages 8-11)
- Laryngeal cancer: EPYC used in ML-derived risk modeling and mechanistic angiogenesis assays, suggesting potential therapeutic target. (zhou2024epycpromoteslaryngeal pages 1-5, zhou2024epycpromoteslaryngeal pages 5-8)
4.2 Musculoskeletal biology: EPYC as a marker of cartilage zones/cell states
In 2024, EPYC/Epyc is used as a marker gene to distinguish growth plate chondrocytes from other cartilage-associated cell types in scRNAseq-based analyses. This is a practical implementation in developmental skeletal biology and spatial annotation workflows. (otsuru2024apolipoproteineis pages 1-4)
5) Expert opinions and analysis (authoritative synthesis)
5.1 What authoritative reviews imply about EPYC’s primary function
Review literature describing EPYC as a DS SLRP localized to epiphyseal cartilage and emphasizing SLRP roles in ECM organization/collagen fibrillogenesis supports a conservative “primary function” annotation: EPYC is likely a structural organizer/modulator of collagen fibrils and cartilage ECM assembly rather than a signaling ligand in the classical sense. (hayes2018biodiversityofcsproteoglycan pages 7-8, halari2021rolesoftwo pages 4-6)
5.2 Interpreting cancer-mechanism papers in the context of ECM SLRP biology
The 2024 laryngeal cancer study suggests EPYC can directly modulate receptor stability (VEGFR1) and Wnt signaling. A plausible integrative interpretation—consistent with known proteoglycan biology—is that an ECM proteoglycan can act as a context-dependent extracellular regulator by binding proteins at the cell surface or in the pericellular milieu, thereby influencing signaling gradients and receptor availability. However, these cancer-context mechanisms should be treated as disease-context expansions of function, not necessarily EPYC’s evolutionarily primary role (which the cartilage-focused and SLRP-focused sources point toward). (hayes2018biodiversityofcsproteoglycan pages 7-8, zhou2024epycpromoteslaryngeal pages 10-13, zhou2024epycpromoteslaryngeal pages 1-5)
6) Relevant statistics and data (recent studies)
Key quantitative values explicitly extractable from 2024 literature in the retrieved corpus include:
- Breast cancer (BMC Cancer; Jan 2024; https://doi.org/10.1186/s12885-024-11913-7): EPYC significantly upregulated in tumor vs adjacent tissue by qRT-PCR (n=55 pairs; P < 0.0001), but overall survival association reported as OS HR = 1.04 with log-rank P = 0.72. (golestan2024unveilingpromisingbreast pages 6-8)
- Pancreatic cancer (Scientific Reports; Jan 2024; https://doi.org/10.1038/s41598-024-51478-w): prognostic modeling reported c-index 0.72 (training) and 0.70 (test) for a model including EPYC. (yang2024epycfunctionsas pages 8-11)
- Laryngeal cancer (Research Square; Apr 2024; https://doi.org/10.21203/rs.3.rs-4182627/v1): EPYC included in an angiogenesis risk score with coefficient 0.25; reported EPYC–VEGFR1 binding affinity KD ~44.1 µM. (zhou2024epycpromoteslaryngeal pages 5-8, zhou2024epycpromoteslaryngeal pages 10-13)
- Cartilage disease model (Disease Models & Mechanisms; Nov 2020; https://doi.org/10.1242/dmm.046425): Epyc among the most strongly downregulated transcripts in hip articular cartilage from Gnptgko mice (log2 ratio ≈ −4). (westermann2020imbalancedcellularmetabolism pages 9-11)
7) Disease associations and genetics resources (database-level)
Open Targets disease–target association results linked EPYC to several phenotypes/diseases including osteoarthritis, abnormality of the skeletal system, and hip dysplasia (Beukes type) (among others), indicating that human-genetics and curated evidence streams consider EPYC relevant to skeletal/cartilage phenotypes (though the evidence details were not expanded in the retrieved context). (OpenTargets Search: -EPYC)
8) Visual evidence from 2024 mechanistic work
Cropped figure panels retrieved from the 2024 laryngeal-cancer study show (i) EPYC-associated survival and (ii) mechanistic experiments implicating Axin/β-catenin regulation and VEGFR1 binding/degradation as the basis of pro-angiogenic effects, supporting the textual mechanism summary. (zhou2024epycpromoteslaryngeal media e7b39943, zhou2024epycpromoteslaryngeal media 717e387a, zhou2024epycpromoteslaryngeal media f7ef9b71, zhou2024epycpromoteslaryngeal media c240e33d)
9) Summary of functional annotation (concise)
EPYC (epiphycan; UniProt Q99645) is best annotated as a secreted extracellular matrix small leucine-rich proteoglycan (DS/CS SLRP) with LRR architecture (reported 7 LRRs in one review), localized to cartilage (including epiphyseal/growth plate cartilage) and implicated in collagen fibrillogenesis and cartilage matrix maturation. Recent 2024 literature extends EPYC’s known biology into cancer microenvironment signaling, including regulation of PI3K–AKT (pancreatic cancer) and Wnt/Axin/β-catenin plus VEGFR1/VEGF-linked angiogenesis (laryngeal cancer), supporting a model in which an ECM proteoglycan can modulate cell signaling in a context-dependent fashion. (hayes2018biodiversityofcsproteoglycan pages 7-8, yang2024epycfunctionsas pages 1-2, zhou2024epycpromoteslaryngeal pages 10-13, otsuru2024apolipoproteineis pages 1-4)
Evidence map table
| Aspect | Summary | Key sources with publication date and URL |
|---|---|---|
| Identity/Family | EPYC encodes epiphycan (human UniProt Q99645), a small leucine-rich proteoglycan (SLRP) carrying dermatan sulfate and/or chondroitin sulfate (DS/CS) chains; reviewed as an ECM proteoglycan at 12q21.33. Open Targets also maps the approved symbol EPYC to disease associations including osteoarthritis and skeletal-system abnormalities, supporting correct human gene identity. (hayes2018biodiversityofcsproteoglycan pages 7-8, halari2021rolesoftwo pages 4-6, OpenTargets Search: -EPYC) | Hayes et al., 2018 — https://doi.org/10.1042/bcj20170820; Halari et al., 2021 — https://doi.org/10.3390/ijms221910584; Open Targets context (OpenTargets Search: -EPYC) |
| Domains/structure | EPYC is an SLRP with leucine-rich repeats (LRRs); the cited review notes epiphycan contains 7 LRRs, fewer than the 10–11 LRRs typical of many other SLRPs. SLRPs share a core LRR-rich protein architecture with extracellular matrix structural roles. (hayes2018biodiversityofcsproteoglycan pages 7-8, halari2021rolesoftwo pages 4-6) | Hayes et al., 2018 — https://doi.org/10.1042/bcj20170820; Halari et al., 2021 — https://doi.org/10.3390/ijms221910584 |
| Localization | Evidence supports EPYC as a secreted/extracellular matrix proteoglycan. Reviews place SLRPs in the ECM, and 2024 laryngeal-cancer work explicitly treats EPYC as a soluble/secreted ECM-associated SLRP acting on endothelial/tumor microenvironment signaling; GO/KEGG enrichment linked EPYC-containing signatures to extracellular matrix organization, extracellular region, and ECM-receptor interaction. (halari2021rolesoftwo pages 4-6, zhou2024epycpromoteslaryngeal pages 1-5, zhou2024epycpromoteslaryngeal pages 5-8) | Halari et al., 2021 — https://doi.org/10.3390/ijms221910584; Zhou et al., 2024 — https://doi.org/10.21203/rs.3.rs-4182627/v1 |
| Core biological role | Current evidence supports a primary structural ECM role in cartilage, especially collagen fibrillogenesis / fibril formation and cartilage matrix maturation. Reviews identify EPYC as a cartilage SLRP; chondrogenic human MSC cultures show EPYC appearing during matrix maturation and being characteristic of a more mature articular-cartilage-like matrix. A 2024 pancreatic-cancer study reiterates that EPYC regulates fibril formation by interacting with collagen fibrils and other ECM proteins. (hayes2018biodiversityofcsproteoglycan pages 7-8, sorrell2018humanmesenchymalstem pages 10-14, yang2024epycfunctionsas pages 1-2) | Hayes et al., 2018 — https://doi.org/10.1042/bcj20170820; Sorrell et al., 2018 — https://doi.org/10.1002/jor.23820; Yang et al., 2024 — https://doi.org/10.1038/s41598-024-51478-w |
| Key tissues/cell types | EPYC is reported in epiphyseal/growth plate cartilage and cartilage-forming cells. Review evidence places epiphycan in epiphyseal cartilage; human MSC chondrogenesis links EPYC to articular-cartilage-type matrix maturation; mouse work found Epyc specifically expressed in growth plate chondrocytes and absent from articular chondrocytes/perichondrial cells in that model; separate cartilage transcriptomics showed strong downregulation in articular cartilage under disease-model conditions. (hayes2018biodiversityofcsproteoglycan pages 7-8, sorrell2018humanmesenchymalstem pages 10-14, otsuru2024apolipoproteineis pages 1-4, westermann2020imbalancedcellularmetabolism pages 9-11) | Hayes et al., 2018 — https://doi.org/10.1042/bcj20170820; Sorrell et al., 2018 — https://doi.org/10.1002/jor.23820; Otsuru et al., 2024 — https://doi.org/10.21203/rs.3.rs-4656728/v1; Westermann et al., 2020 — https://doi.org/10.1242/dmm.046425 |
| Pathways/signaling links | Native/cartilage biology evidence mainly supports ECM assembly roles rather than enzymatic catalysis. In 2024 cancer studies, EPYC was linked to PI3K-AKT signaling in pancreatic cancer and to Wnt/Axin/β-catenin plus VEGFR1/VEGF signaling in laryngeal cancer. The laryngeal study further reported direct EPYC–VEGFR1 binding and VEGFR1 degradation, consistent with extracellular signaling modulation by a secreted matrix proteoglycan. (yang2024epycfunctionsas pages 1-2, zhou2024epycpromoteslaryngeal pages 10-13, zhou2024epycpromoteslaryngeal pages 1-5, zhou2024epycpromoteslaryngeal media e7b39943) | Yang et al., 2024 — https://doi.org/10.1038/s41598-024-51478-w; Zhou et al., 2024 — https://doi.org/10.21203/rs.3.rs-4182627/v1 |
| Disease/biomarker evidence (2023-2024) | Recent studies propose EPYC as a biomarker/functional contributor in several diseases: pancreatic cancer prognostic biomarker with functional proliferation effects; laryngeal cancer angiogenesis/carcinogenesis promoter; breast cancer diagnostic biomarker candidate; Open Targets lists associations with osteoarthritis, abnormality of the skeletal system, and hip dysplasia, Beukes type. These disease links are promising but context-dependent and do not supersede EPYC’s likely core ECM/cartilage function. (yang2024epycfunctionsas pages 1-2, zhou2024epycpromoteslaryngeal pages 1-5, golestan2024unveilingpromisingbreast pages 6-8, OpenTargets Search: -EPYC) | Yang et al., 2024 — https://doi.org/10.1038/s41598-024-51478-w; Zhou et al., 2024 — https://doi.org/10.21203/rs.3.rs-4182627/v1; Golestan et al., 2024 — https://doi.org/10.1186/s12885-024-11913-7; Open Targets context (OpenTargets Search: -EPYC) |
| Quantitative stats (2024) | Reported 2024 quantitative findings include: breast cancer qRT-PCR in 55 paired samples showed EPYC upregulation (P < 0.0001), but overall-survival association was weak/non-significant (HR 1.04, log-rank P = 0.72). Pancreatic-cancer prognostic modeling reported c-index 0.72 (training) and 0.70 (test) in a model retaining EPYC. Laryngeal-cancer work reported EPYC coefficient = 0.25 in its risk score and EPYC–VEGFR1 binding KD ~44.1 µM. In cartilage disease-model tissue, Epyc log2 ratio ~ -4 in hip articular cartilage was reported. (golestan2024unveilingpromisingbreast pages 6-8, yang2024epycfunctionsas pages 8-11, zhou2024epycpromoteslaryngeal pages 5-8, zhou2024epycpromoteslaryngeal pages 10-13, westermann2020imbalancedcellularmetabolism pages 9-11) | Golestan et al., 2024 — https://doi.org/10.1186/s12885-024-11913-7; Yang et al., 2024 — https://doi.org/10.1038/s41598-024-51478-w; Zhou et al., 2024 — https://doi.org/10.21203/rs.3.rs-4182627/v1; Westermann et al., 2020 — https://doi.org/10.1242/dmm.046425 |
Table: This table summarizes the verified identity, structure, localization, biological role, tissue expression, signaling links, and recent disease evidence for human EPYC (epiphycan). It is useful as a compact evidence map for functional annotation grounded in the cited contexts only.
References
-
(hayes2018biodiversityofcsproteoglycan pages 7-8): Anthony Hayes, Kazuyuki Sugahara, Brooke Farrugia, John M. Whitelock, Bruce Caterson, and James Melrose. Biodiversity of cs-proteoglycan sulphation motifs: chemical messenger recognition modules with roles in information transfer, control of cellular behaviour and tissue morphogenesis. The Biochemical journal, 475 3:587-620, Feb 2018. URL: https://doi.org/10.1042/bcj20170820, doi:10.1042/bcj20170820. This article has 63 citations.
-
(halari2021rolesoftwo pages 4-6): Chidambra D. Halari, Michael Zheng, and Peeyush K. Lala. Roles of two small leucine-rich proteoglycans decorin and biglycan in pregnancy and pregnancy-associated diseases. International Journal of Molecular Sciences, 22:10584, Sep 2021. URL: https://doi.org/10.3390/ijms221910584, doi:10.3390/ijms221910584. This article has 35 citations.
-
(yang2024epycfunctionsas pages 1-2): Zhen Yang, Honglin Li, Jie Hao, Hanwei Mei, Minghan Qiu, Huaqing Wang, and Ming Gao. Epyc functions as a novel prognostic biomarker for pancreatic cancer. Scientific Reports, Jan 2024. URL: https://doi.org/10.1038/s41598-024-51478-w, doi:10.1038/s41598-024-51478-w. This article has 15 citations and is from a peer-reviewed journal.
-
(zhou2024epycpromoteslaryngeal pages 1-5): Dongmei Zhou, Fan Liu, Xiaoqi Li, and Fei Lv. Epyc promotes laryngeal cancer carcinogenesis and angiogenesis via thewnt-axin-β-catenin/vegfr1 pathway. Unknown journal, Apr 2024. URL: https://doi.org/10.21203/rs.3.rs-4182627/v1, doi:10.21203/rs.3.rs-4182627/v1.
-
(zhou2024epycpromoteslaryngeal pages 5-8): Dongmei Zhou, Fan Liu, Xiaoqi Li, and Fei Lv. Epyc promotes laryngeal cancer carcinogenesis and angiogenesis via thewnt-axin-β-catenin/vegfr1 pathway. Unknown journal, Apr 2024. URL: https://doi.org/10.21203/rs.3.rs-4182627/v1, doi:10.21203/rs.3.rs-4182627/v1.
-
(sorrell2018humanmesenchymalstem pages 10-14): J. Michael Sorrell, Rodrigo A. Somoza, and Arnold I. Caplan. Human mesenchymal stem cells induced to differentiate as chondrocytes follow a biphasic pattern of extracellular matrix production. Journal of Orthopaedic Research®, 36:1757-1766, Jun 2018. URL: https://doi.org/10.1002/jor.23820, doi:10.1002/jor.23820. This article has 25 citations.
-
(otsuru2024apolipoproteineis pages 1-4): Satoru Otsuru, Joe Kodama, Takeshi Oichi, Kevin Wilkinson, Joshua Abzug, Takashi Kaito, Motomi Iwamoto-Enomoto, and Masahiro Iwamoto. Apolipoprotein e is a novel marker for chondrocytes in the growth plate resting zone. Research Square, Aug 2024. URL: https://doi.org/10.21203/rs.3.rs-4656728/v1, doi:10.21203/rs.3.rs-4656728/v1. This article has 1 citations.
-
(zhou2024epycpromoteslaryngeal pages 10-13): Dongmei Zhou, Fan Liu, Xiaoqi Li, and Fei Lv. Epyc promotes laryngeal cancer carcinogenesis and angiogenesis via thewnt-axin-β-catenin/vegfr1 pathway. Unknown journal, Apr 2024. URL: https://doi.org/10.21203/rs.3.rs-4182627/v1, doi:10.21203/rs.3.rs-4182627/v1.
-
(zhou2024epycpromoteslaryngeal media e7b39943): Dongmei Zhou, Fan Liu, Xiaoqi Li, and Fei Lv. Epyc promotes laryngeal cancer carcinogenesis and angiogenesis via thewnt-axin-β-catenin/vegfr1 pathway. Unknown journal, Apr 2024. URL: https://doi.org/10.21203/rs.3.rs-4182627/v1, doi:10.21203/rs.3.rs-4182627/v1.
-
(zhou2024epycpromoteslaryngeal media 717e387a): Dongmei Zhou, Fan Liu, Xiaoqi Li, and Fei Lv. Epyc promotes laryngeal cancer carcinogenesis and angiogenesis via thewnt-axin-β-catenin/vegfr1 pathway. Unknown journal, Apr 2024. URL: https://doi.org/10.21203/rs.3.rs-4182627/v1, doi:10.21203/rs.3.rs-4182627/v1.
-
(golestan2024unveilingpromisingbreast pages 6-8): Ali Golestan, Ahmad Tahmasebi, Nafiseh Maghsoodi, Seyed Nooreddin Faraji, Cambyz Irajie, and Amin Ramezani. Unveiling promising breast cancer biomarkers: an integrative approach combining bioinformatics analysis and experimental verification. BMC Cancer, Jan 2024. URL: https://doi.org/10.1186/s12885-024-11913-7, doi:10.1186/s12885-024-11913-7. This article has 37 citations and is from a peer-reviewed journal.
-
(yang2024epycfunctionsas pages 8-11): Zhen Yang, Honglin Li, Jie Hao, Hanwei Mei, Minghan Qiu, Huaqing Wang, and Ming Gao. Epyc functions as a novel prognostic biomarker for pancreatic cancer. Scientific Reports, Jan 2024. URL: https://doi.org/10.1038/s41598-024-51478-w, doi:10.1038/s41598-024-51478-w. This article has 15 citations and is from a peer-reviewed journal.
-
(westermann2020imbalancedcellularmetabolism pages 9-11): Lena Marie Westermann, Lutz Fleischhauer, Jonas Vogel, Zsuzsa Jenei-Lanzl, Nataniel Floriano Ludwig, Lynn Schau, Fabio Morellini, Anke Baranowsky, Timur A. Yorgan, Giorgia Di Lorenzo, Michaela Schweizer, Bruna de Souza Pinheiro, Nicole Ruas Guarany, Fernanda Sperb-Ludwig, Fernanda Visioli, Thiago Oliveira Silva, Jamie Soul, Gretl Hendrickx, J. Simon Wiegert, Ida V. D. Schwartz, Hauke Clausen-Schaumann, Frank Zaucke, Thorsten Schinke, Sandra Pohl, and Tatyana Danyukova. Imbalanced cellular metabolism compromises cartilage homeostasis and joint function in a mouse model of mucolipidosis type iii gamma. Disease Models & Mechanisms, Nov 2020. URL: https://doi.org/10.1242/dmm.046425, doi:10.1242/dmm.046425. This article has 10 citations and is from a domain leading peer-reviewed journal.
-
(OpenTargets Search: -EPYC): Open Targets Query (-EPYC, 5 results). Buniello, A. et al. (2025). Open Targets Platform: facilitating therapeutic hypotheses building in drug discovery. Nucleic Acids Research.
-
(zhou2024epycpromoteslaryngeal media f7ef9b71): Dongmei Zhou, Fan Liu, Xiaoqi Li, and Fei Lv. Epyc promotes laryngeal cancer carcinogenesis and angiogenesis via thewnt-axin-β-catenin/vegfr1 pathway. Unknown journal, Apr 2024. URL: https://doi.org/10.21203/rs.3.rs-4182627/v1, doi:10.21203/rs.3.rs-4182627/v1.
-
(zhou2024epycpromoteslaryngeal media c240e33d): Dongmei Zhou, Fan Liu, Xiaoqi Li, and Fei Lv. Epyc promotes laryngeal cancer carcinogenesis and angiogenesis via thewnt-axin-β-catenin/vegfr1 pathway. Unknown journal, Apr 2024. URL: https://doi.org/10.21203/rs.3.rs-4182627/v1, doi:10.21203/rs.3.rs-4182627/v1.
Artifacts
Citations
- halari2021rolesoftwo pages 4-6
- hayes2018biodiversityofcsproteoglycan pages 7-8
- sorrell2018humanmesenchymalstem pages 10-14
- yang2024epycfunctionsas pages 1-2
- golestan2024unveilingpromisingbreast pages 6-8
- otsuru2024apolipoproteineis pages 1-4
- yang2024epycfunctionsas pages 8-11
- westermann2020imbalancedcellularmetabolism pages 9-11
- zhou2024epycpromoteslaryngeal pages 1-5
- zhou2024epycpromoteslaryngeal pages 5-8
- zhou2024epycpromoteslaryngeal pages 10-13
- https://doi.org/10.1186/s12885-024-11913-7
- https://doi.org/10.1038/s41598-024-51478-w
- https://doi.org/10.21203/rs.3.rs-4182627/v1
- https://doi.org/10.1242/dmm.046425
- https://doi.org/10.1042/bcj20170820;
- https://doi.org/10.3390/ijms221910584;
- https://doi.org/10.3390/ijms221910584
- https://doi.org/10.1002/jor.23820;
- https://doi.org/10.21203/rs.3.rs-4656728/v1;
- https://doi.org/10.1038/s41598-024-51478-w;
- https://doi.org/10.21203/rs.3.rs-4182627/v1;
- https://doi.org/10.1186/s12885-024-11913-7;
- https://doi.org/10.1042/bcj20170820,
- https://doi.org/10.3390/ijms221910584,
- https://doi.org/10.1038/s41598-024-51478-w,
- https://doi.org/10.21203/rs.3.rs-4182627/v1,
- https://doi.org/10.1002/jor.23820,
- https://doi.org/10.21203/rs.3.rs-4656728/v1,
- https://doi.org/10.1186/s12885-024-11913-7,
- https://doi.org/10.1242/dmm.046425,
Perplexity
(EPYC-deep-research-perplexity.md)
Perplexity
(EPYC-deep-research-perplexity.md)Epiphycan (EPYC): A Comprehensive Functional and Molecular Analysis of a Small Leucine-Rich Proteoglycan
Opening Summary
Epiphycan (EPYC), also known as dermatan sulfate proteoglycan 3 (DSPG3), represents a significant member of the small leucine-rich proteoglycan (SLRP) family with essential roles in extracellular matrix organization, tissue development, and homeostasis. Located on chromosome 12 and encoding a 309-amino acid protein with a molecular mass of approximately 35.5 kilodaltons, EPYC functions primarily as a modulator of collagen fibrillogenesis and a regulator of cell signaling through multiple pathways including transforming growth factor-beta (TGF-β), phosphatidylinositol 3-kinase/protein kinase B (PI3K/AKT), and focal adhesion signaling[6][13][16]. The protein is secreted as a decorated extracellular matrix component bearing chondroitin/dermatan sulfate glycosaminoglycan chains, through which it orchestrates critical interactions between cells and their matrix environment[2][3]. Recent research has revealed diverse roles for EPYC extending from cartilage development and osteoarthritis pathogenesis to tumor progression in pancreatic and ovarian cancers, establishing it as both a fundamental regulator of tissue architecture and an emerging biomarker for disease prognosis[3][47][55]. This comprehensive report synthesizes current knowledge of EPYC's structural features, localization patterns, biochemical functions, signaling mechanisms, and clinical relevance.
Classification and Definition of Small Leucine-Rich Proteoglycans
The small leucine-rich proteoglycan family represents the largest proteoglycan family in humans, comprising eighteen distinct members organized into five distinct classes based on their evolutionary conservation, genomic organization, and structural features[7][52]. These molecules are defined fundamentally by the presence of tandem leucine-rich repeat (LRR) domains flanked by N- and C-terminal cysteine-rich clusters, which create the characteristic curved solenoid structure characteristic of all SLRP family members[7]. The core protein domains of SLRPs typically range from 36 to 77 kilodaltons and may be substituted with various types of glycosaminoglycan (GAG) side chains, including chondroitin sulfate (CS), dermatan sulfate (DS), and keratan sulfate (KS), although some SLRP members exist as non-glycosylated proteins depending on tissue context and developmental stage[7][29]. Because SLRPs are present in virtually all extracellular matrices of bilaterian animals and exhibit wide tissue distribution, they represent ancient and highly conserved molecules with deep evolutionary significance[2]. The functional diversity of this protein family emerges not merely from amino acid sequence variation but fundamentally from the differential organization of their leucine-rich repeat domains, their capacity for post-translational modification, and their tissue-specific regulation of glycosaminoglycan substitution[2][7].
Structural Organization and Molecular Composition of Epiphycan
Epiphycan occupies a specific position within SLRP classification as a Class III member alongside opticin and osteoglycin (mimecan)[7][8][50]. The EPYC gene comprises seven exons organized on chromosome 12, position 12q21.33, with a genomic span of approximately 41.3 kilobases[13][48]. The resulting protein contains between six and eight leucine-rich repeat domains, which represents the defining characteristic of Class III SLRPs and distinguishes them from the more elaborate Class I and II members carrying ten to twelve LRRs[7][8]. Each individual leucine-rich repeat in EPYC spans approximately twenty to twenty-nine amino acids and consists of highly conserved segments displaying characteristic patterns such as LxxLxLxxNxL and variable segments that form diverse secondary structures including alpha-helices and beta-turns[19]. The curved solenoid architecture arising from these repeating structures creates a characteristic molecular shape wherein the concave inner surface serves as the ligand-binding interface, while the convex exterior provides structural scaffolding[19]. The N- and C-terminal regions flanking the LRR domains are rich in cysteine residues that form disulfide bonds, creating terminal caps essential for protein stability and proper three-dimensional folding[2][7].
The epiphycan protein is synthesized as a precursor containing an N-terminal signal peptide approximately twenty amino acids in length, which directs the nascent polypeptide chain to the endoplasmic reticulum during translation and is subsequently cleaved to yield the mature secreted protein[41]. Following synthesis, EPYC undergoes extensive post-translational modification within the secretory pathway, with glycosaminoglycan chain attachment occurring primarily through the Golgi apparatus via coordinated enzymatic reactions[7][37]. The protein accepts chondroitin and dermatan sulfate GAG chains as post-translational modifications, with these structures being constructed from repeating disaccharide units comprising N-acetylgalactosamine and uronic acid residues[27][30]. The specific composition and sulfation pattern of these GAG chains undergo further enzymatic processing including epimerization of glucuronic acid to iduronic acid and sulfation at specific positions on both the hexosamine and uronic acid components, reactions catalyzed by dedicated sulfotransferase and epimerase enzymes that confer functional diversity to the modified proteoglycan[27][37].
Post-translational modifications beyond GAG attachment include N-linked and O-linked glycosylation through mannose and glucose residues, modifications that prove critical for protein folding, structural stability, and proper presentation of ligand-binding epitopes[37]. Tyrosine residues within specific regions of EPYC undergo sulfation, a relatively common but underappreciated post-translational modification catalyzed by tyrosylprotein sulfotransferase in the trans-Golgi network[37][40]. These tyrosine sulfations appear particularly important in Class II SLRPs such as fibromodulin but require further investigation in Class III members like EPYC[37]. The overall modification state of epiphycan varies significantly depending on the tissue source, developmental stage, aging, and pathological context, reflecting dynamic regulation of the enzymatic machinery responsible for GAG biosynthesis[7][37]. This dynamic composition explains in part how a single protein core can function in remarkably diverse biological contexts and disease states.
Subcellular Localization and Tissue Distribution of Epiphycan
EPYC functions exclusively in extracellular contexts, with the protein predicted and annotated as secreted based on its N-terminal signal peptide sequence and absence of transmembrane domains[41]. The secretion process delivers epiphycan to the extracellular space where it assembles into complex three-dimensional matrices alongside other proteoglycans, structural proteins including collagens and elastin, and various glycoproteins that together constitute the functional extracellular matrix[2][6][9]. The mature protein contains no intracellular localization signals and demonstrates no evidence of intracellular accumulation in normal physiological conditions, consistent with its role as a purely secreted matrix molecule[41][6]. Following secretion, EPYC becomes integrated into the organized three-dimensional scaffold of the extracellular matrix through multiple molecular interactions with collagen fibrils, hyaluronan, and other matrix components, positions that prove critical for its regulatory functions[2][9].
EPYC shows preferential expression in skeletal tissues, with particularly high levels in the growth plate cartilage, articular cartilage, and intervertebral discs where it participates in establishing and maintaining the ordered matrix architecture essential for skeletal function[6][24]. During embryonic development, epiphycan mRNA transcripts appear initially around embryonic day 12.5 in developing cartilage elements and exhibit a tightly spatially and temporally regulated expression pattern during chondrogenesis[25]. The expression timing of EPYC relative to other cartilage markers reveals important developmental relationships, with EPYC mRNA expression occurring later than type II collagen, which marks early chondrogenic differentiation, but appearing in the growth plate before and being excluded from the zone of hypertrophic chondrocytes expressing collagen type X[25]. Protein localization studies using immunofluorescence histochemistry demonstrate that EPYC protein persists throughout all zones of growth plate cartilage, including the hypertrophic zone, even in regions where the gene is no longer actively transcribed[25][28]. This discordance between mRNA and protein localization patterns suggests that epiphycan protein accumulates in the extracellular matrix and persists there during tissue remodeling and cartilage maturation.
Beyond skeletal tissues, EPYC distribution extends to virtually all tissues containing specialized extracellular matrices, including fibrous tissues such as tendons, ligaments, and the cornea, and to the encapsulating matrices of major organs including the meninges and pericardium[7][29]. The widespread distribution of EPYC across diverse connective tissues, combined with its particularly high expression in fibrous tissues, reflects the fundamental importance of this molecule in organizing and regulating extracellular matrix properties across multiple tissue types[7]. However, the expression level and GAG modification state of EPYC varies substantially between different tissues, with such variation likely reflecting tissue-specific functional requirements and developmental history[6][7]. Single-cell expression analysis indicates that EPYC production occurs in chondrocytes of cartilage tissues and in fibroblasts of connective tissues, with these specialized matrix-producing cells serving as the primary sources of EPYC protein in their respective tissues[41].
Primary Biological Functions: Collagen Fibrillogenesis and Matrix Organization
The primary and best-characterized function of EPYC involves the regulation of collagen fibrillogenesis and the maintenance of ordered collagen matrix architecture, functions shared with other SLRP family members but with specific characteristics reflecting EPYC's unique structural features and biochemical properties[8][24][29]. Collagen fibrils form through a multistep process beginning with self-assembly of collagen triple helix monomers followed by lateral association and cross-linking of individual fibrils into larger organized fiber bundles, processes that generate the mechanical properties essential for tissue function[9][26]. In this assembly process, EPYC and other SLRPs function as regulators, controlling the kinetics of fibril formation, limiting lateral growth of individual fibrils, and maintaining appropriate intermolecular spacing between adjacent fibrils[7][29][31]. Through direct binding interactions with collagen fibrils, particularly with type II collagen present in cartilage and other collagen types in different tissues, EPYC exerts steric hindrance effects that prevent excessive lateral fusion of fibril structures[24][31]. This regulatory function proves essential for maintaining the optimal biomechanical properties of tissues, as aberrant collagen fibrillogenesis results in structural defects affecting tissue stiffness, elasticity, and load-bearing capacity[31].
The role of EPYC in collagen regulation becomes evident through analysis of knockout mouse models wherein EPYC-deficient mice display minimal developmental abnormalities at birth but develop progressive osteoarthritis with age, with the severity of degeneration increasing dramatically in mice deficient in both EPYC and biglycan, another SLRP family member[8][24][33]. Histological examination of cartilage from these doubly deficient animals reveals increased collagen loss, particularly in superficial zones of articular cartilage, associated with a deficiency of superficial chondrocytes and profound disruption of normal joint architecture[24]. These findings demonstrate that EPYC, while not absolutely essential for embryonic development, proves critical for the long-term maintenance of joint integrity and the prevention of cartilage degeneration under the mechanical stresses imposed during normal skeletal loading[8][24][33].
Beyond its role in regulating the physical assembly of collagen matrices, EPYC participates in organizing the three-dimensional architecture of the extracellular matrix through interactions with multiple other matrix components[3][6]. The proteoglycan functions as an organization node, facilitating interactions between collagen fibrils, hyaluronic acid, link proteins, and other proteoglycans that together generate the complex viscoelastic properties characteristic of functional cartilage and other connective tissues[2][9][34]. Through these multi-molecular interactions, EPYC contributes to what is often termed "matrix organization," a process that involves the spatial arrangement of matrix molecules into their proper three-dimensional configuration essential for tissue function[3]. This organizational role extends to regulating the accessibility of matrix components to proteolytic degradation by matrix metalloproteinases and other enzymes, a function with clear implications for understanding tissue remodeling and pathological matrix degradation observed in diseases such as osteoarthritis[26][36].
Bone Formation and Cartilage Development
EPYC demonstrates important functions beyond simple structural organization, with documented roles in bone formation and in establishing the conditions necessary for normal chondrogenesis[4][6]. The protein influences bone development through mechanisms that may involve both direct effects on osteoblast function and indirect effects mediated through matrix organization and sequestration of bone-active growth factors[6]. During endochondral ossification, the developmental process through which cartilage serves as a temporary template for bone formation, EPYC participates in the remodeling of epiphyseal cartilage preceding the formation of secondary ossification centers[15]. The protein may contribute to this process through its interactions with transforming growth factor-beta (TGF-β) family members, including bone morphogenetic proteins (BMPs), which regulate both chondrocyte differentiation and osteoblast maturation[6][14].
The role of EPYC in bone formation becomes apparent through examination of knockout mouse phenotypes, which while showing relatively subtle skeletal abnormalities early in development, display progressive changes in long bone morphometry with age, including effects on femur length in males as assessed at nine months of age[8][24]. These age-dependent skeletal phenotypes suggest that EPYC functions in the maintenance of normal bone homeostasis and the prevention of age-related skeletal changes rather than serving as an absolutely required factor for bone formation itself[8]. The protein may function by regulating local growth factor availability within bone matrix compartments or by maintaining the mechanical properties of the bone matrix necessary for proper skeletal loading and remodeling[6].
Growth Factor Interactions and Signaling Modulation
A major class of EPYC functions involves the binding and sequestration of soluble growth factors and cytokines, particularly the transforming growth factor-beta superfamily and bone morphogenetic proteins[3][6][29]. Like other SLRP family members, EPYC binds TGF-β isoforms through a mechanism that reduces TGF-β bioavailability and modulates its signaling activity in a dose-dependent manner[26][29]. This function proves particularly significant in tissues where TGF-β signaling drives pathological fibrosis or inflammation, a context in which SLRP-mediated sequestration of TGF-β provides a biological brake on excess fibrogenic signaling[26][29]. The chondroitin and dermatan sulfate GAG chains attached to EPYC contribute substantially to these growth factor interactions, with the sulfated structures serving as high-affinity ligands for multiple bioactive molecules[27][30].
The interaction between EPYC and growth factors extends beyond simple sequestration to include functional modulation of signaling outcomes. Through binding to growth factors or to their cognate receptors, EPYC can either attenuate or enhance specific signaling pathways depending on the cellular context and the nature of the EPYC-growth factor-receptor complex[3][6]. Recent evidence suggests that EPYC may interact with specific signaling molecules including phospholipase Cγ2 (PLCG2) and the adaptor protein CRK, both implicated in signal transduction cascades activated by receptor tyrosine kinases and non-receptor tyrosine kinases[51]. These interactions position EPYC not merely as a passive sequestrant of diffusible signals but as an active participant in cellular decision-making regarding proliferation, survival, and differentiation. The interaction with CRK protein, a known effector in insulin-like growth factor (IGF) signaling pathways, suggests potential cross-talk between ECM-resident EPYC and growth factor receptor signaling cascades that regulate cell survival and metabolism[51].
Involvement in Osteoarthritis and Joint Pathology
Evidence from multiple sources demonstrates that EPYC dysfunction associates with osteoarthritis development and progression, a relationship emerging from both genetic studies and functional investigations of EPYC expression in disease tissues. Epiphycan-deficient mice develop osteoarthritis spontaneously with age, with the onset of pathological changes appearing earliest in mice carrying deletions in both EPYC and biglycan, suggesting synergistic effects between these two Class I and Class III SLRP family members[8][24][33]. The osteoarthritis phenotype in EPYC-deficient animals includes classic hallmarks of the disease including articular cartilage erosions, proteoglycan loss from the matrix, subchondral bone sclerosis, and osteophyte growth, morphological changes that closely resemble those observed in human osteoarthritis patients[33].
Mechanistic investigation of EPYC's role in osteoarthritis has identified alterations in matrix protein expression and signaling pathway activation in animals lacking EPYC[8]. Microarray analysis of knee joints from EPYC/biglycan double-deficient mice reveals increased matrix protein expression compared with wild-type mice, including upregulation of other SLRP family members such as asporin, fibromodulin, and lumican[8][24][33]. This compensatory upregulation of alternative SLRPs likely reflects attempts by the tissue to restore normal matrix organization in the face of EPYC deficiency, but these compensatory mechanisms prove insufficient to prevent cartilage degeneration[33]. The expression alterations suggest a model in which EPYC loss results in matrix disorganization that triggers secondary responses including altered transcription of matrix genes and potential activation of catabolic pathways[33].
Gene expression profiling studies employing machine learning approaches identify EPYC as a major feature gene distinguishing osteoarthritis tissue from normal cartilage, with EPYC expression elevated in osteoarthritic tissue samples[55]. Paradoxically, while EPYC knockout mice develop osteoarthritis, osteoarthritic tissue shows elevated EPYC expression, a pattern suggesting that increased EPYC production may represent a reparative response by chondrocytes attempting to restore damaged matrix architecture[55]. This interpretation aligns with the observation that in normal cartilage, EPYC expression remains relatively low and may become transiently upregulated during periods of matrix damage or stress[25][55]. The apparent contradiction between EPYC deficiency causing osteoarthritis and EPYC upregulation in osteoarthritic tissue may reflect distinct roles in prevention of degeneration versus response to matrix damage, a distinction that requires further investigation.
Role in Corneal Dystrophies and Ocular Pathology
EPYC associations extend beyond skeletal tissues to include roles in corneal development and maintenance, revealed through genetic studies identifying EPYC mutations in patients with corneal dystrophies. Posterior amorphous corneal dystrophy (PACD), a rare autosomal dominant corneal disorder characterized by partial or complete posterior lamellar corneal opacification and decreased corneal thickness, shows genetic linkage to the chromosomal region harboring EPYC[39]. Affected patients exhibit disorganization of posterior corneal stromal lamellae with an abnormal collagenous layer within or adjacent to Descemet's membrane, architectural changes consistent with disrupted collagen matrix organization similar to defects observed when SLRP function is compromised[39]. While no mutations in EPYC were identified in the initially reported PACD family, the candidate gene approach highlighting EPYC alongside other SLRPs including keratocan, lumican, and decorin emphasizes the importance of this proteoglycan family in maintaining corneal architecture[39][26].
EPYC additionally associates with cornea plana (keratoconus), another inherited corneal disorder characterized by corneal flattening and stromal thinning[39][42]. The shared candidate gene status of EPYC with other SLRP family members in corneal dystrophies reflects the universal dependence of corneal transparency and structure on proper collagen matrix organization, a process in which multiple SLRP family members participate redundantly[26][29]. The cornea, with its requirement for exceptional transparency to permit light transmission, demonstrates particular sensitivity to disruptions in matrix organization, such that even modest alterations in collagen fibril diameter or interfibrillar spacing result in clinically apparent stromal opacification[26]. This extreme sensitivity of the cornea to matrix disorganization makes it a sensitive biosensor for SLRP dysfunction and has facilitated the identification of multiple SLRP mutations in corneal disease through molecular genetic studies[26].
Cancer-Associated Functions and Emerging Biomarker Roles
Recent research reveals that EPYC expression patterns and functions extend into malignant disease contexts, where the proteoglycan promotes tumor progression and serves as a prognostic indicator of poor outcomes. In pancreatic cancer, integrated bioinformatics analysis of The Cancer Genome Atlas (TCGA) and Gene Expression Omnibus (GEO) datasets identifies EPYC as an independent risk factor predicting poor prognosis[47][58]. Functional studies demonstrate that EPYC promotes pancreatic cancer cell proliferation through activation of the PI3K-AKT signaling pathway both in cell culture and in vivo tumor models[47][58]. Knockdown of EPYC in pancreatic cancer cell lines reduces cancer cell proliferation, demonstrating direct pro-tumorigenic effects of this ECM proteoglycan[47]. The emergence of EPYC as a prognostic biomarker in pancreatic cancer exemplifies how ECM components traditionally viewed as passive structural molecules now emerge as active regulators of tumor cell behavior.
In ovarian cancer, EPYC expression levels correlate with metastatic potential and clinical outcomes, with EPYC showing higher expression in metastatic ovarian cancer tissue compared to primary tumors, and higher expression in primary cancers compared to normal ovaries[3][44]. Functional interference studies in which EPYC expression is suppressed using small interfering RNA demonstrate that EPYC knockdown reduces ovarian cancer cell migration, invasion, and proliferation, confirming active roles for EPYC in promoting hallmark cancer behaviors[3]. Mechanistically, EPYC appears to promote epithelial-to-mesenchymal transition (EMT), a cellular process associated with increased migratory and invasive capacity through multiple signaling pathways including TGF-β/PI3K-AKT and focal adhesion pathways[3][44]. EPYC-mediated activation of these pathways leads to suppression of the epithelial marker E-cadherin (CDH1) through upregulation of the transcriptional repressor SNAI2, a critical step in EMT[44].
Gene ontology enrichment analysis demonstrates that EPYC primarily functions in extracellular matrix organization, confirming its fundamental role as a matrix protein[3][44]. Pathway enrichment analysis reveals that EPYC-associated gene expression changes activate multiple signaling cascades including PI3K/AKT, focal adhesion, and TGF-β pathways[3][44]. This multiplicity of activated pathways reflects the complex mechanisms through which ECM components influence cancer cell behavior through bidirectional signaling between matrix-resident proteins and cell surface receptors. The findings establish EPYC as an emerging therapeutic target in cancers where ECM-promoted tumor progression contributes to poor outcomes.
PI3K/AKT Signaling and Focal Adhesion Pathway Activation
The phosphatidylinositol 3-kinase/protein kinase B (PI3K/AKT) signaling cascade represents a major effector pathway activated by EPYC in both cancer and normal physiological contexts[3][47][55]. This pathway initiates when ligands bind to receptor tyrosine kinases or when integrins engage ECM molecules including EPYC-containing matrix through focal adhesion complexes[20][32][35]. Upon receptor activation, PI3K catalyzes the phosphorylation of phosphatidylinositol 4,5-bisphosphate to generate the signaling lipid phosphatidylinositol 3,4,5-trisphosphate (PIP3), which serves as a membrane docking site for proteins containing pleckstrin homology domains, including AKT and its upstream activator PDK1[20][23]. Once recruited to the membrane, AKT becomes phosphorylated and activated, whereupon it phosphorylates multiple downstream effectors regulating cell survival, proliferation, metabolism, and cytoskeletal dynamics[20][23]. In the context of EPYC signaling, PI3K/AKT activation promotes cell survival through suppression of pro-apoptotic proteins and cell proliferation through modulation of cell cycle checkpoint proteins[47][58].
The focal adhesion pathway constitutes another major signaling cascade activated downstream of EPYC-ECM interactions, with focal adhesions forming complex multi-protein assemblies at sites where cell surface integrins engage ECM ligands[20][32][35]. Within focal adhesions, the non-receptor tyrosine kinase focal adhesion kinase (FAK) becomes activated through interactions with integrin cytoplasmic domains, leading to FAK autophosphorylation at tyrosine 397 and subsequent recruitment and activation of Src-family kinases[32][35]. FAK and Src then phosphorylate multiple adaptor proteins including paxillin, p130Cas, and Talin, creating a signaling scaffold that activates downstream pathways regulating cell migration, survival, and proliferation[20][32][35]. These FAK-Src signaling complexes activate additional kinases including the integrin-linked kinase (ILK), which further propagates signals through multiple effector pathways[20][32][35]. The activation of focal adhesion kinase downstream of EPYC-integrin interactions demonstrates how ECM-resident proteoglycans participate in bidirectional signaling between cells and their matrix environment.
Developmental Regulation and Tissue Remodeling Functions
EPYC plays crucial roles in developmental processes involving tissue remodeling and matrix reorganization, functions reflecting the general dependence of development on properly regulated ECM assembly and proteoglycan function[6][14][15]. During embryonic skeletal development, EPYC expression precisely correlates with stages of chondrogenesis and endochondral ossification, appearing in developing cartilage elements in the precise temporal and spatial pattern necessary for proper tissue organization[25]. The protein emerges as an important intermediate marker for chondrogenesis, appearing later than early markers of chondrocyte differentiation such as type II collagen but before terminal differentiation markers[25]. This intermediate expression timing positions EPYC in a critical developmental window when cartilage tissue requires specialized matrix organization distinct from both early cartilage anlagen and mature, mineralized cartilage.
The regulatory factors controlling EPYC expression during development remain incompletely characterized but likely include both transcriptional regulation responsive to developmental signals and post-transcriptional regulation of GAG modification state. The growth plate cartilage, a tissue continuously undergoing developmental remodeling through zones of resting, proliferating, and hypertrophic chondrocytes, shows EPYC protein localization throughout its organized structure even though the EPYC gene becomes progressively downregulated as chondrocytes differentiate toward terminal hypertrophy[25][28]. This persistence of EPYC protein in matrix compartments despite reduced transcription suggests accumulation of long-lived matrix-resident EPYC that persists through tissue remodeling cycles and may provide substrate for matrix repair mechanisms. The developmental remodeling of EPYC expression thus reflects complex interplay between synthetic, secretory, and degradative processes governing matrix homeostasis.
Comparative Analysis with Other SLRP Family Members and Functional Redundancy
EPYC functions within the context of a larger proteoglycan family displaying both functional redundancy and functional specialization, characteristics that emerge from structural similarities combined with evolutionarily divergent sequences and regulatory patterns[7][29][31][52]. The Class III SLRP family, comprising EPYC, opticin, and osteoglycin (mimecan), represents a distinct lineage within the broader SLRP family tree, with sequence and structural characteristics distinguishing these molecules from Class I and II members[7][50][52]. While all SLRPs share fundamental structural organization based on leucine-rich repeat domains and typically interact with collagen matrices to regulate fibrillogenesis, individual SLRP members display specialized functions reflecting their distinct binding specificities and tissue expression patterns[7][29].
Evidence for functional redundancy among SLRP family members emerges from studies of doubly deficient knockout mice, wherein deletion of both EPYC and biglycan produces a phenotype more severe than deletion of either molecule alone[8][24][33]. The enhanced osteoarthritis phenotype in EPYC/biglycan double-deficient mice compared to singly deficient animals suggests that while individual SLRPs are partly dispensable due to compensatory activity of other family members, combined loss of multiple SLRPs reveals their essential aggregate function in joint maintenance[33]. Moreover, molecular characterization of joint tissues from doubly deficient mice reveals compensatory upregulation of other SLRP family members including asporin, fibromodulin, and lumican, demonstrating that tissues attempt to restore normal SLRP levels when individual members are deleted[8][24][33]. This compensation appears partially effective in delaying osteoarthritis onset but ultimately insufficient to prevent age-related cartilage degeneration, suggesting that individual SLRPs possess at least partially specialized functions not entirely redundant with their family members[33].
EPYC demonstrates particular functional relationships with Class I SLRPs like biglycan and decorin through shared interactions with TGF-β and other growth factors, interactions that influence wound healing, tissue remodeling, and fibrosis[26][29][31]. However, Class III SLRPs like EPYC differ from Class I and II members in their GAG composition, with EPYC carrying chondroitin/dermatan sulfate chains while Class II SLRPs typically bear keratan sulfate modifications, differences that confer distinct ligand-binding specificities and functional properties[7][50]. The evolutionary divergence of SLRP family members, evident from phylogenetic analysis showing distinct branches separating different SLRP classes, suggests that specialization for distinct tissue roles occurred during vertebrate evolution, with different SLRP members becoming adapted for particular connective tissue types[52].
Post-Translational Modification and Functional Diversity
The biological activity of EPYC depends critically on post-translational modifications including glycosaminoglycan chain attachment and sulfation, modifications that substantially alter the proteoglycan's biochemical and biophysical properties[7][37]. The composition of attached GAG chains, including the degree of sulfation and the specific positioning of sulfate groups on hexosamine and uronic acid residues, directly influences EPYC's capacity to interact with growth factors, matrix molecules, and cell surface receptors[27][30][37]. This structural variability in GAG modification emerges from regulated expression of sulfotransferase and epimerase enzymes that catalyze the post-translational modifications of GAG chains within the Golgi apparatus and extracellular space[27][37]. Remarkably, a single EPYC core protein can exist in multiple biochemically and functionally distinct forms depending on the degree and pattern of GAG sulfation, a phenomenon that introduces substantial complexity into proteoglycan function[7][37].
The sulfation patterns of GAG chains attached to EPYC prove especially important for the protein's growth factor-binding activities, with specific sulfate positioning enhancing affinity for molecules like transforming growth factor-beta and determining whether EPYC acts as a high-affinity ligand sequestering growth factors or a lower-affinity scaffold permitting growth factor access to cell surface receptors[26][29][30]. Sulfated GAG chains also provide charged environments that influence local pH and ion concentrations in the perifibrillar matrix compartment surrounding collagen fibrils, biophysical properties that may influence both collagen stability and cell behavior[9][21]. The variable GAG sulfation state of EPYC in different tissues and developmental stages suggests that fine-tuning of proteoglycan post-translational modification represents a regulatory mechanism allowing the EPYC core protein to assume diverse functional roles depending on biosynthetic and enzymatic context[7][37].
Evolutionary Conservation and Phylogenetic Relationships
EPYC represents an evolutionarily conserved protein, with homologous molecules present across vertebrate species and even certain invertebrate representatives, suggesting ancient biological importance[49][52]. Phylogenetic analysis reveals that vertebrate SLRP family members, including EPYC alongside opticin and osteoglycin, cluster within a distinct evolutionary lineage, the Class III SLRP clade that likely arose through gene duplication and divergence from ancestral SLRP sequences[49][52]. The presence of characteristic structural features including the defining six to eight leucine-rich repeats and specific cysteine-rich N-terminal domains distinguishing Class III members indicates that EPYC evolved specialized functions adapted to distinct tissue requirements[49][52]. During teleost fish evolution, EPYC remained as a single gene copy per haploid genome, unlike certain other SLRP members that underwent whole-genome duplication events creating multiple gene copies with subfunctionalization[49]. This single-copy retention through vertebrate evolution suggests that EPYC has maintained a specialized biological role that does not benefit from gene duplication and functional specialization observed in other SLRP family members.
The conservation of EPYC protein sequence across mammalian species reflects functional constraints on amino acid positions critical for ligand binding, structural stability, and interactions with other matrix molecules[49][52]. Comparative genomic analysis reveals that the EPYC gene structure, including the pattern of exons and introns, shows remarkable conservation across mammalian species, a finding consistent with purifying selection against changes that would disrupt proper gene expression or protein function[49]. The evolutionary conservation of EPYC extends to regulatory regions controlling the gene's expression in specific tissues and developmental stages, suggesting that temporal and spatial patterns of EPYC expression have been optimized through evolution for proper skeletal development and tissue homeostasis[49][52].
Mechanisms of Collagen Interaction and Fibrillogenesis Regulation
The molecular mechanisms through which EPYC regulates collagen fibrillogenesis involve direct binding interactions between the EPYC protein core and collagen molecules, interactions mediated by the leucine-rich repeat domains serving as the binding interface[7][29][26]. Biochemical studies employing collagen-proteoglycan complex formation assays and surface plasmon resonance measurements reveal specific binding sites for SLRP interaction with collagen molecules, with certain leucine-rich repeat domains showing particularly high-affinity interactions[7]. For EPYC and other Class III SLRPs, the binding site on collagen involves specific regions of the collagen triple helix that become accessible as the collagen molecules assemble into progressively larger fibrillar structures[29]. The binding of EPYC to collagen fibrils affects the kinetics of collagen fibrillogenesis, with proteomic and biophysical studies demonstrating that SLRP presence slows the rate of fibril formation and reduces the overall turbidity of collagen gels, evidence that SLRP binding inhibits lateral fusion of collagen fibrils[7][29][34].
The mechanism of SLRP-mediated inhibition of collagen fibril growth likely involves steric hindrance, wherein the curved solenoid structure of the SLRP core protein physically limits the lateral approach and fusion of adjacent collagen fibrils by occupying space on the collagen fibril surface[7][29]. This sterically mediated regulation differs from alternative mechanisms such as proteolytic cleavage or covalent modification of collagen, instead representing a reversible, regulator-dependent mechanism that permits dynamic adjustment of fibril growth in response to changing matrix requirements[29][31]. The strength of SLRP binding to collagen, and consequently the degree of fibrillogenesis inhibition, varies among different SLRP family members, reflecting their evolutionary specialization for different tissues and functional roles[7][29]. EPYC demonstrates intermediate-strength binding to collagen compared to some other SLRPs, a characteristic potentially reflecting its specialized role in cartilage and skeletal tissues where moderate regulation of fibrillogenesis optimizes tissue biomechanical properties[29][31].
The GAG chains attached to EPYC contribute substantially to the protein's collagen-binding properties through multiple mechanisms including providing additional binding surface area and creating charged microenvironments that influence protein-protein interactions[7][29][27]. Removal of GAG chains from SLRP molecules through enzymatic treatment with chondroitinase or other GAG-degrading enzymes reduces SLRP binding to collagen and alters the effects of SLRPs on collagen fibrillogenesis kinetics[29][34]. This dependence of function on GAG presence explains the importance of proper post-translational modification for SLRP biological activity and the variable functional states of EPYC depending on tissue-specific GAG modification patterns.
Emerging Therapeutic Implications and Future Research Directions
The emerging recognition of EPYC as both a fundamental regulator of tissue homeostasis and an important player in disease pathogenesis suggests potential therapeutic applications in multiple clinical contexts. In osteoarthritis, strategies aimed at modulating EPYC expression or function might target either the excessive matrix degradation associated with advanced disease or the compensatory matrix synthesis responses occurring in early degeneration[55]. The observation that EPYC functions partially redundantly with other SLRP family members suggests that therapeutic interventions targeting EPYC might require simultaneous modulation of other SLRP molecules to achieve maximal efficacy[8][33]. Conversely, the specific roles of EPYC distinct from other SLRPs might enable selective therapeutic targeting of EPYC to achieve desired effects on cartilage organization without disrupting other SLRP-dependent functions.
In cancer contexts, the emerging role of EPYC as a promoter of tumor progression and metastasis suggests that targeting EPYC expression or its signaling functions might impede cancer progression[47][58]. Approaches might include direct inhibition of EPYC using blocking antibodies or antisense oligonucleotides, modulation of EPYC expression through targeting upstream regulators, or indirect approaches targeting downstream signaling pathways activated by EPYC including the PI3K-AKT cascade[47]. The identification of PLCG2 and CRK as EPYC-interacting proteins opens possibilities for targeting these specific protein-protein interactions to modulate EPYC signaling functions[51]. Combination strategies targeting EPYC alongside other ECM components or growth factors might prove particularly effective, reflecting the multifactorial nature of both osteoarthritis and cancer involving complex interactions between cells and their matrix environment.
Future research directions include detailed characterization of the molecular mechanisms through which EPYC activates specific signaling pathways in different cellular contexts, investigation of tissue-specific regulation of EPYC expression and post-translational modification, and functional analysis of the predicted EPYC interactions with PLCG2, CRK, and other signaling molecules[51]. Structure-function studies mapping the specific amino acid residues and domains of EPYC required for interactions with distinct ligands including growth factors, other matrix proteins, and cell surface receptors would provide molecular-level insights enabling rational therapeutic targeting[7][19]. The development of improved animal models permitting inducible deletion or modification of EPYC in specific tissues or at specific developmental timepoints would clarify the tissue-specific roles of EPYC and distinguish between functions required during development versus maintenance of adult tissues[8]. Investigation of whether and how aberrant EPYC expression or modification contribute to disease pathogenesis in tissues beyond those currently characterized would identify new disease associations and potential therapeutic targets[47][55][58].
Conclusion
Epiphycan represents a prototypical small leucine-rich proteoglycan with fundamental roles in organizing extracellular matrices, regulating growth factor signaling, and maintaining tissue homeostasis across multiple biological systems. The protein's multifaceted functions reflect both its structural characteristics as a curved solenoid molecule with multiple binding surfaces and its sophisticated post-translational modification with chondroitin and dermatan sulfate glycosaminoglycans that enable dynamic regulation of biochemical properties in response to tissue requirements and developmental signals. EPYC demonstrates critical importance in maintaining skeletal integrity, preventing osteoarthritis progression, and regulating corneal structure, functions that become apparent from genetic studies employing knockout animal models and bioinformatic analysis of patient tissues. Beyond these classical roles in tissue organization, EPYC emerges as an important regulator of cancer cell behavior, promoting tumor progression through activation of PI3K-AKT and focal adhesion signaling pathways, functions establishing EPYC as an emerging biomarker for cancer prognosis[47][55][58]. Future therapeutic strategies targeting EPYC may prove valuable in both preventing cartilage degeneration in osteoarthritis and restricting cancer progression in malignancies where aberrant EPYC expression promotes metastasis and poor outcomes. The continued investigation of EPYC structure-function relationships, tissue-specific regulation, and signaling mechanisms will enhance understanding of proteoglycan biology while identifying novel approaches for modulating EPYC in diseased states.
Citations
- https://www.ncbi.nlm.nih.gov/gene/1833
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3527888/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8650094/
- https://www.uniprot.org/uniprotkb/Q99645/entry
- https://www.proteinatlas.org/ENSG00000083782-EPYC/structure+interaction
- https://www.prospecbio.com/epyc-protein
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11092928/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3013283/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4883592/
- https://anatomypubs.onlinelibrary.wiley.com/doi/10.1002/(SICI)1097-0177(199912)216:4/5%3C499::AID-DVDY18%3E3.0.CO;2-S
- https://www.genecards.org/cgi-bin/carddisp.pl?gene=EPYC
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6829314/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3820861/
- https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=1833
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3606489/
- https://pubmed.ncbi.nlm.nih.gov/36920035/
- https://www.wikipathways.org/pathways/WP3932.html
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10432810/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6852186/
- https://pubmed.ncbi.nlm.nih.gov/10633869/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5204247/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9957132/
- https://academic.oup.com/glycob/article/12/9/117R/548591
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9378794/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10537162/
- https://www.nature.com/articles/s41598-020-76107-0
- https://www.nature.com/articles/s41392-022-01259-6
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3671949/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11678238/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2910638/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6272617/
- https://www.proteinatlas.org/ENSG00000083782-EPYC/subcellular
- https://diseases.jensenlab.org/Entity?documents=10&type1=9606&id1=ENSP00000261172&type2=-26&id2=DOID%3A0060287
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11745771/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2763122/
- https://www.semanticscholar.org/paper/EPYC-functions-as-a-novel-prognostic-biomarker-for-Yang-Li/b28771fc4013c5a30fb201e814fe08d5c53918d6
- https://www.nature.com/articles/s41598-024-51478-w
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6293640/
- https://febs.onlinelibrary.wiley.com/doi/10.1111/j.1742-4658.2010.07797.x
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4859157/
- https://maayanlab.cloud/Harmonizome/gene_set/connective+tissue/TISSUES+Text-mining+Tissue+Protein+Expression+Evidence+Scores
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9330685/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3527881/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC12254123/
- https://onlinelibrary.wiley.com/doi/abs/10.1002/jor.1100090602
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2547145/