Deep Research
Falcon
(FAS-deep-research-falcon.md)
Falcon
(FAS-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: Human FAS (TNFRSF6/CD95/APO-1; UniProt P25445) functional annotation and current translational landscape
1) Key concepts, definitions, and current understanding (verified gene/protein identity)
Target verification (mandatory): The gene symbol FAS here refers to the human cell-surface death receptor CD95/APO-1, a type I transmembrane member of the TNF receptor superfamily (TNFRSF6) with extracellular cysteine-rich domains (CRDs) and a cytosolic death domain (DD) that recruits the adaptor FADD to assemble a death-inducing signaling complex (DISC), consistent with UniProt accession P25445. (hu2025fasmediatesapoptosis pages 1-2, seyrek2024thecrosstalkof pages 1-2)
Core definition: FAS/CD95 is a multifunctional receptor best known for initiating the extrinsic apoptosis pathway following engagement by its cognate ligand FASLG/CD95L, but it can also signal through non-apoptotic programs (e.g., NF-κB/MAPK) depending on stimulus strength, ligand form, and DISC composition. (seyrek2024thecrosstalkof pages 14-15, seyrek2024thecrosstalkof pages 2-4)
Structural concepts (functional regions):
- Extracellular region: 3 CRDs; ligand-binding is described primarily through CRD2 and part of CRD3. A pre-ligand assembly domain (PLAD) (reported residues 59–82) promotes pre-association at the plasma membrane, facilitating signaling competence. (seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof pages 1-2)
- Transmembrane segment: a single-pass (~17 aa) TM region; intramembrane features contribute to receptor trimerization/clustering. (seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof pages 1-2)
- Cytosolic death domain: ~80 aa arranged as six antiparallel α-helices; it mediates homotypic DD interactions and binding to FADD, enabling DISC assembly. (seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof pages 1-2)
Ligand forms matter (major checkpoint): Reviews emphasize that membrane-bound FASLG (mFASL) is a strong apoptosis trigger, while soluble FASLG (sFASL) more often biases toward non-apoptotic outputs such as NF-κB signaling. (seyrek2024thecrosstalkof pages 14-15, hu2025fasmediatesapoptosis pages 1-2)
2) Mechanism and pathway placement
2.1 Canonical apoptotic signaling (extrinsic apoptosis)
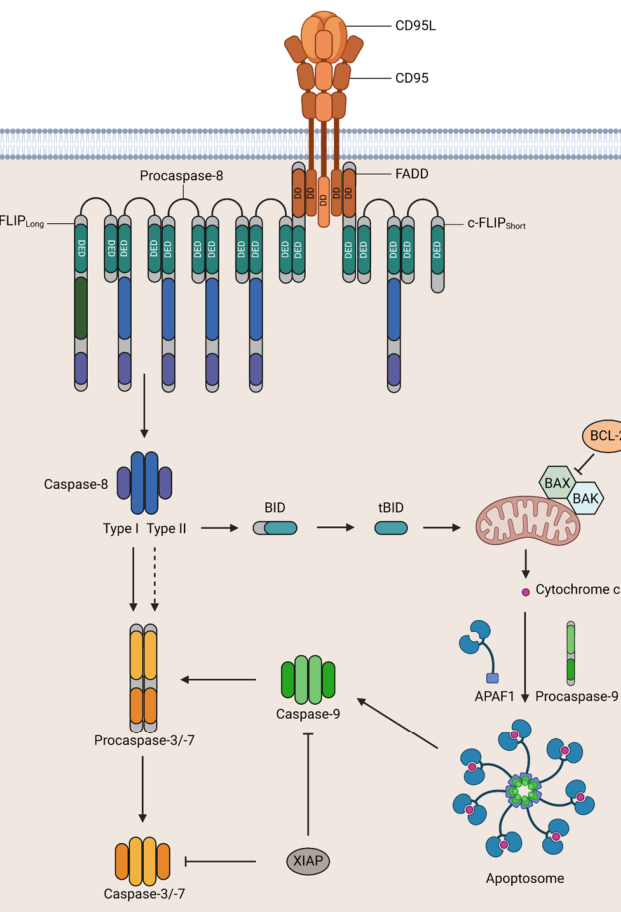
Upon ligand binding, conformational changes allow FAS DD engagement with FADD, which then recruits DED-containing proteins procaspase-8/10 and c-FLIP isoforms to form the membrane-associated DISC; this supports procaspase-8 activation, which then activates effector caspases (e.g., caspase-3/-7) to execute apoptosis. DISC assembly is described as rapid (on the order of seconds). (hu2025fasmediatesapoptosis pages 1-2, seyrek2024thecrosstalkof pages 2-4)
A common conceptual framework is the presence of type I vs type II apoptotic responses downstream of death receptors (direct effector caspase activation vs mitochondrial amplification), and this is explicitly illustrated in the CD95 pathway schematics. (seyrek2024thecrosstalkof media 9097b778)
Visual evidence (pathway schematic): Seyrek et al. provide a figure depicting CD95L–CD95 engagement, DISC formation (FADD, procaspase-8, c-FLIP), and downstream type I/type II apoptotic routes. (seyrek2024thecrosstalkof media 9097b778, seyrek2024thecrosstalkof media cf4ad51c)
2.2 Non-apoptotic (context-dependent) signaling
A key contemporary view is that CD95 can also act as a multifunctional signaling receptor, activating programs such as NF-κB and MAPK (ERK/JNK/p38), supporting outcomes including chemokine release and cellular migration/invasion in certain contexts. (seyrek2024thecrosstalkof pages 14-15, seyrek2024thecrosstalkof pages 2-4)
Mechanistic checkpoint—DISC composition and c-FLIP: The level and isoform composition of c-FLIP at the DISC is highlighted as a central decision point: high c-FLIP can disrupt procaspase-8 DED filament formation, favoring non-apoptotic signaling; additionally, signaling intermediates associated with c-FLIP processing (e.g., p43-FLIP) are described to recruit components (e.g., TRAF2/RIPK1/RAF1) that connect to non-apoptotic pathways. (seyrek2024thecrosstalkof pages 14-15)
Visual evidence (apoptotic vs non-apoptotic schematic): A second figure in Seyrek et al. schematizes apoptotic and non-apoptotic signaling and depicts NF-κB activation downstream of CD95-associated complexes (including FADD/RIPK1/c-FLIP/caspases-8/10). (seyrek2024thecrosstalkof media 39ce50a4, seyrek2024thecrosstalkof media 9276d39c)
3) Cellular localization and where FAS acts
Primary localization: FAS is primarily a plasma membrane/cell surface receptor, broadly expressed across human cells and prominently in immune contexts; signaling is initiated at the membrane where receptor pre-association/clustering supports rapid assembly of signaling complexes. (hu2025fasmediatesapoptosis pages 1-2, seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof pages 1-2)
Functional compartmentalization: Current models separate membrane-proximal DISC (apoptotic) versus alternative membrane-associated complexes (often described as MISC/FADDosome-like) associated with non-apoptotic NF-κB/MAPK signaling. (seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof media 9097b778)
4) Regulation and expert synthesis (authoritative review perspectives)
Recent expert synthesis highlights that cell fate downstream of CD95 is gated by multiple checkpoints, including (i) ligand form (mFASL vs sFASL), (ii) stimulus strength/threshold effects, (iii) availability and PTMs of DISC components, and (iv) particularly the c-FLIP:procaspase-8 balance at the DISC. (seyrek2024thecrosstalkof pages 14-15, seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof pages 1-2)
Post-translational regulation is discussed as a life/death determinant, including CD95 glycosylation and PTMs that affect stability/turnover of DISC regulators (e.g., nitrosylation/ubiquitylation control of c-FLIP stability is summarized in the review-level evidence). (seyrek2024thecrosstalkof pages 14-15, seyrek2024thecrosstalkof pages 1-2)
5) Recent developments and latest research (prioritizing 2023–2024)
5.1 Human genetics: FAS variants and autoimmune lymphoproliferative syndrome (ALPS)
A large 2024 cohort study of 802 individuals referred for ALPS NGS testing (May 2014–Jan 2023) provides quantitative, clinically grounded evidence linking FAS to immune homeostasis via apoptosis regulation:
- Definite diagnostic genetic findings in 7.7% (62/802).
- Among diagnostic cases, 84% (52/62) carried pathogenic/likely pathogenic FAS variants (6.5% of the total cohort).
- 46% of unique diagnostic FAS variants were novel (17/37).
- Pathogenic variants were enriched in the intracellular region: 58% (30/52) in the intracellular domain; among these, 57% (17/30) were missense variants in the death domain.
These data support the DD as a critical functional module for apoptotic signaling competence in vivo. (xu2024genetictestingin pages 2-4, xu2024genetictestingin pages 1-2)
5.2 Translational targeting: CD95 blockade by CD95-Fc (asunercept/APG101/CAN008)
A recurring translational theme is therapeutic blockade of CD95–CD95L using a soluble decoy/fusion protein (CD95 extracellular domain fused to Fc; commonly referred to as asunercept/APG101/CAN008), motivated by non-apoptotic tumor-promoting/invasive signaling in some contexts and by pathologic death signaling in others. (streuer2024treatmentwiththe pages 1-2)
Glioblastoma (GBM): A 2024 analysis of a phase 1/2 open-label trial cohort (NCT02853565) reports 10 enrolled and 9 evaluable newly diagnosed GBM patients receiving CAN008/asunercept plus standard chemoradiotherapy; high-dose weekly asunercept (400 mg/week; n=6) was associated with 2- and 5-year OS of 83% and 67%, compared with 40.1% and 8.8% in a historical Taiwanese GBM cohort (n=164), with historical median OS 20 months and a reported p=0.0103 for improved OS in the high-dose group. The study also reports biomarker associations including CD95L promoter methylation and higher tumor mutational burden in responders. (chang2024can008prolongsoverall pages 1-2)
Myelodysplastic neoplasms (MDS): The ClinicalTrials.gov record for APG101 in MDS (NCT01736436; registry 2013) describes a phase 1, single-arm, open-label study with 20 enrolled, dosing 100 mg IV weekly for 12 weeks, primary endpoint safety/tolerability, and secondary endpoints including transfusion frequency and bone marrow parameters. (NCT01736436 chunk 1)
A 2024 molecular substudy analyzing serial whole-exome sequencing in 12 low-risk MDS patients reports a molecular response in 75% (9/12), defined as ≥10% reduction in dominant clone VAF (mean decrease 20%, range 10.5–39.2%), with most decline after 12 weeks of treatment. (streuer2024treatmentwiththe pages 1-2)
Severe COVID-19 (trial design): The ClinicalTrials.gov record NCT04535674 (registry 2020) describes a multicenter, open-label, randomized phase 2 trial in hospitalized severe COVID-19 with 438 enrolled, comparing SoC vs SoC + weekly IV asunercept at 25/100/400 mg. The primary endpoint is time to sustained clinical improvement by ≥1 category on the WHO 9-point ordinal scale through day 29, with multiple secondary endpoints (oxygenation, ventilation, discharge, mortality). (NCT04535674 chunk 1)
6) Current applications and real-world implementations
Clinical implementation: The most concrete real-world implementation in the provided recent evidence is CD95L/CD95 blockade using asunercept/APG101/CAN008, evaluated across oncology (GBM), hematology (MDS), and infectious disease/inflammation (severe COVID-19) via interventional clinical trials with defined endpoints and substantial enrollment (e.g., NCT04535674). (chang2024can008prolongsoverall pages 1-2, NCT04535674 chunk 1, NCT01736436 chunk 1)
Biomarker-driven considerations: In GBM, CD95L promoter methylation and tumor mutational burden were linked to response, illustrating how CD95 pathway modulation is moving toward biomarker-informed use (hypothesis-generating at present). (chang2024can008prolongsoverall pages 1-2)
7) Summary tables (evidence-backed)
The following tables consolidate core functional annotation and quantitative translational evidence.
| Feature | Human FAS/CD95 annotation | Evidence note | Citation |
|---|---|---|---|
| Protein type / identity | Type I transmembrane death receptor of the TNF receptor superfamily; also called CD95, APO-1, TNFRSF6; human FAS corresponds to UniProt P25445 | Recent reviews explicitly match human FAS with CD95/APO-1 and describe it as a cell-surface TNFR-family death receptor mediating apoptotic and non-apoptotic signaling | (hu2025fasmediatesapoptosis pages 1-2, seyrek2024thecrosstalkof pages 1-2) |
| Extracellular cysteine-rich domains (CRDs) | N-terminal extracellular region contains three cysteine-rich domains; CRD2 and part of CRD3 contribute to ligand recognition | Structural summary identifies 3 CRDs and maps ligand-binding activity mainly to CRD2 plus part of CRD3, consistent with TNFR-family architecture | (seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof pages 1-2) |
| PLAD (pre-ligand assembly domain) | PLAD spans residues 59–82 and supports receptor pre-association/oligomerization before ligand binding | CD95 pre-oligomerization at the plasma membrane is highlighted as a signaling-facilitating feature, with PLAD specifically assigned to residues 59–82 | (seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof pages 1-2) |
| Transmembrane region | Single-pass transmembrane segment (~17 aa); intramembrane interactions contribute to receptor trimerization/clustering | Reviews describe a 17-aa TM region and note a proline-containing TM motif/intramembrane trimerization that helps organize signaling-competent receptor assemblies | (seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof pages 1-2) |
| Cytoplasmic death domain | Intracellular death domain (~80 aa) composed of six antiparallel α-helices; binds FADD through homotypic DD interactions | This domain is the core signaling module that converts ligand-induced receptor conformational changes into DISC assembly | (seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof pages 1-2) |
| Main ligand(s) | FAS ligand/CD95L is the cognate ligand; exists as membrane-bound (mFASL) and soluble (sFASL) forms | mFASL is emphasized as the potent apoptosis-inducing form, whereas sFASL more often favors non-apoptotic outputs such as NF-κB-linked signaling | (hu2025fasmediatesapoptosis pages 1-2, seyrek2024thecrosstalkof pages 14-15) |
| DISC adaptor proteins | FADD is the central adaptor; DISC recruits procaspase-8, procaspase-10, and c-FLIP isoforms | After FAS activation, FADD bridges the receptor death domain to DED-containing effectors/regulators, forming the membrane-bound DISC | (hu2025fasmediatesapoptosis pages 1-2, seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof media 9097b778) |
| Apoptotic signaling branch | Canonical extrinsic apoptosis via caspase-8 activation; type I cells rely mainly on direct effector caspase activation, type II cells amplify through mitochondria/BID/apoptosome | Figure-based and text evidence show DISC-driven caspase-8 activation followed by direct caspase-3/7 activation or mitochondrial amplification depending on cellular context | (seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof media 9097b778) |
| Non-apoptotic signaling branch | Can activate NF-κB and MAPK pathways (ERK, JNK, p38), supporting inflammation, migration, survival, regeneration, and tumor-promoting programs in some contexts | Current understanding is that FAS is multifunctional: low/altered stimulation or modified DISC composition can redirect signaling away from apoptosis toward survival/inflammatory programs | (seyrek2024thecrosstalkof pages 14-15, seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof media 9097b778) |
| Typical cellular localization | Primarily plasma membrane/cell surface receptor; signaling platforms include pre-associated oligomers and membrane microdomains; DISC assembles at the membrane | Reviews describe ubiquitous surface expression, especially on immune cells, with signaling initiated at membrane-associated receptor clusters | (hu2025fasmediatesapoptosis pages 1-2, seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof pages 1-2) |
| Key intracellular signaling compartments | Membrane-associated DISC for apoptotic signaling; non-apoptotic complexes including FADDosome/MISC have been proposed for NF-κB/MAPK signaling | Figure summary distinguishes DISC from non-apoptotic FADDosome-like assemblies containing FADD, RIPK1, c-FLIP, and caspases-8/10 | (seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof media 9097b778) |
| Ligand-form checkpoint | mFASL generally drives robust apoptotic signaling; sFASL preferentially supports non-apoptotic signaling | Ligand biochemical form is repeatedly cited as a major determinant of life/death outcome downstream of FAS | (hu2025fasmediatesapoptosis pages 1-2, seyrek2024thecrosstalkof pages 14-15) |
| c-FLIP checkpoint | c-FLIPL, c-FLIPS, and c-FLIPR modulate procaspase-8 activation; short isoforms inhibit caspase-8 activation, while c-FLIPL can be pro- or anti-apoptotic depending on abundance/ratio | DISC composition and especially c-FLIP level are presented as central switches controlling whether CD95 signaling remains apoptotic or becomes non-apoptotic | (seyrek2024thecrosstalkof pages 1-2, seyrek2024thecrosstalkof pages 14-15) |
| PTM / receptor regulation checkpoint | Glycosylation of CD95 and PTMs of DISC proteins influence signaling outcome; nitrosylation and ubiquitination control c-FLIP stability and apoptosis resistance | Reviews highlight PTMs as major checkpoints, though effects are context dependent and often act by reshaping DISC assembly or turnover of regulators | (seyrek2024thecrosstalkof pages 1-2, seyrek2024thecrosstalkof pages 14-15) |
Table: This table summarizes the verified identity, domain architecture, signaling mechanisms, localization, and major regulatory checkpoints of human FAS/CD95 (UniProt P25445). It is useful as a compact functional annotation linking receptor structure to apoptotic versus non-apoptotic outcomes.
| Evidence area | Study / source | Key quantitative or translational findings | URL | Publication / registry date | Citation |
|---|---|---|---|---|---|
| ALPS genetic testing cohort | Xu et al., Journal of Clinical Immunology; retrospective ALPS NGS testing cohort | 802 patients tested (May 2014–Jan 2023); median age 12 years; 63% male (504/802). Definite diagnostic yield 7.7% (62/802). Of diagnostic cases, 84% (52/62) had pathogenic/likely pathogenic FAS variants, equal to 6.5% of the full cohort (52/802). Diagnostic yield increased to 30% among patients also meeting abnormal ALPS immunology criteria. Among 37 unique diagnostic FAS variants, 46% (17/37) were novel. Domain distribution: 58% (30/52) of FAS-positive cases had heterozygous variants in the intracellular domain; of these, 57% (17/30) were missense variants in the death domain. | https://doi.org/10.1007/s10875-024-01772-z | Jul 2024 | (xu2024genetictestingin pages 2-4, xu2024genetictestingin pages 1-2) |
| GBM CAN008 / asunercept outcomes | Chang et al., Biomedical Journal; phase 1/2 open-label trial analysis in newly diagnosed glioblastoma | Trial NCT02853565 enrolled 10 patients; 9 evaluable. Dosing groups: 200 mg/week (n=3) and 400 mg/week (n=6) CAN008/asunercept plus standard CCRT. Compared with historical Taiwanese GBM cohort (n=164), high-dose CAN008 group showed OS at 2 and 5 years of 83% and 67% vs 40.1% and 8.8% in the historical cohort; historical cohort median OS 20 months; improved OS with high-dose CAN008, p=0.0103. Biomarker findings: low CD95L promoter CpG2 methylation associated with better response; better responders had higher variant count and tumor mutational burden. | https://doi.org/10.1016/j.bj.2023.100660 | Aug 2024 | (chang2024can008prolongsoverall pages 1-2) |
| MDS APG101 registry parameters | ClinicalTrials.gov NCT01736436; APG101/asunercept in transfusion-dependent low/intermediate-risk MDS | Phase 1, single-arm, open-label interventional study; enrollment 20. Intervention: APG101 100 mg IV weekly for 12 weeks with 6-month follow-up (37 weeks total). Primary objective/outcome: safety and tolerability (AEs/SAEs, ECGs, abdominal ultrasound, anti-drug antibodies, lymphocyte subsets/activation markers, ECOG). Secondary outcomes: overall survival, changes in transfusion frequency, bone marrow parameters (histologic/cytologic/cytogenetic), and hemoglobin levels. Key eligibility included adult WHO-classified de novo low/intermediate-risk MDS, blast count <5%, transfusion dependence, refractory/unlikely to respond to ESA. | https://clinicaltrials.gov/study/NCT01736436 | Registry 2013 | (NCT01736436 chunk 1) |
| MDS APG101 molecular substudy | Streuer et al., Annals of Hematology; molecular follow-up from NCT01736436 | Bone marrow from 12 low-risk MDS patients analyzed at 58 time points by serial whole-exome sequencing. Mean 3.5 molecularly defined subclones per patient (range 2–6). Molecular response defined as dominant clone VAF decrease ≥10% occurred in 9/12 patients (75%); mean VAF decrease 20%, range 10.5–39.2%. Most clonal decline occurred after completion of 12 weeks treatment. Early pronounced reductions were noted in subclones with DNMT3A, IDH2, or TET2 mutations. | https://doi.org/10.1007/s00277-024-05664-5 | Feb 2024 | (streuer2024treatmentwiththe pages 1-2) |
| Severe COVID-19 trial design | ClinicalTrials.gov NCT04535674; asunercept in hospitalized severe COVID-19 | Multicenter, open-label, randomized, phase 2 trial; enrollment 438 (actual). Parallel arms: Standard of Care alone vs SoC + asunercept at 25 mg, 100 mg, or 400 mg weekly IV. Primary endpoint: time to sustained clinical improvement of at least 1 category on the 9-category WHO ordinal scale, sustained for 2 consecutive days, measured Days 1–29. Secondary endpoints included NEWS-based recovery/discharge, oxygenation-free days, ventilator-free days, hospital/ICU outcomes, and mortality at days 15, 29, 60, and 90. | https://clinicaltrials.gov/study/NCT04535674 | Registry 2020 | (NCT04535674 chunk 1) |
Table: This table compiles recent quantitative and clinical evidence for human FAS/CD95, spanning inherited disease genetics, oncology trials, hematology studies, and COVID-19 trial design. It is useful as a compact evidence map linking FAS biology to real-world human datasets and therapeutic development.
8) Limitations of this synthesis
- While recent review evidence provides detailed mechanistic models (DISC vs non-apoptotic complexes), precise numeric kinetic/biophysical parameters (e.g., binding constants, absolute molecule counts per DISC) were not present in the extracted text snippets; the review points to such primary studies but they were not directly retrievable in the current evidence set. (seyrek2024thecrosstalkof pages 2-4, seyrek2024thecrosstalkof pages 20-21)
- Some highly relevant primary immunology work on non-apoptotic FAS signaling in human B cells (Science Immunology, 2024; doi:10.1126/sciimmunol.adj5948) was flagged as unobtainable in tool retrieval and therefore is not cited directly here.
9) Key recent sources (with dates and URLs)
- Seyrek K, Espe J, Reiss E, Lavrik IN. The Crosstalk of Apoptotic and Non-Apoptotic Signaling in CD95 System. Cells. Nov 2024. https://doi.org/10.3390/cells13211814 (seyrek2024thecrosstalkof pages 1-2)
- Xu X et al. Genetic Testing in Patients with Autoimmune Lymphoproliferative Syndrome: Experience of 802 Patients… Journal of Clinical Immunology. Jul 2024. https://doi.org/10.1007/s10875-024-01772-z (xu2024genetictestingin pages 1-2)
- Chang IY-F et al. CAN008 prolongs overall survival… newly diagnosed GBM… Biomedical Journal. Aug 2024. https://doi.org/10.1016/j.bj.2023.100660 (chang2024can008prolongsoverall pages 1-2)
- Streuer A et al. Treatment with the apoptosis inhibitor Asunercept reduces clone sizes… lower risk MDS. Annals of Hematology. Feb 2024. https://doi.org/10.1007/s00277-024-05664-5 (streuer2024treatmentwiththe pages 1-2)
- ClinicalTrials.gov. NCT01736436: APG101 in Myelodysplastic Syndrome. Registry 2013. https://clinicaltrials.gov/study/NCT01736436 (NCT01736436 chunk 1)
- ClinicalTrials.gov. NCT04535674: Asunercept in Patients With Severe COVID-19. Registry 2020. https://clinicaltrials.gov/study/NCT04535674 (NCT04535674 chunk 1)
References
-
(hu2025fasmediatesapoptosis pages 1-2): Liying Hu, Juane Lu, Hongfei Fan, Changcheng Niu, Yanping Han, Qinggele Caiyin, Hao Wu, and Jianjun Qiao. Fas mediates apoptosis, inflammation, and treatment of pathogen infection. Frontiers in Cellular and Infection Microbiology, Apr 2025. URL: https://doi.org/10.3389/fcimb.2025.1561102, doi:10.3389/fcimb.2025.1561102. This article has 25 citations.
-
(seyrek2024thecrosstalkof pages 1-2): Kamil Seyrek, Johannes Espe, Elisabeth Reiss, and Inna N. Lavrik. The crosstalk of apoptotic and non-apoptotic signaling in cd95 system. Cells, 13:1814, Nov 2024. URL: https://doi.org/10.3390/cells13211814, doi:10.3390/cells13211814. This article has 12 citations.
-
(seyrek2024thecrosstalkof pages 14-15): Kamil Seyrek, Johannes Espe, Elisabeth Reiss, and Inna N. Lavrik. The crosstalk of apoptotic and non-apoptotic signaling in cd95 system. Cells, 13:1814, Nov 2024. URL: https://doi.org/10.3390/cells13211814, doi:10.3390/cells13211814. This article has 12 citations.
-
(seyrek2024thecrosstalkof pages 2-4): Kamil Seyrek, Johannes Espe, Elisabeth Reiss, and Inna N. Lavrik. The crosstalk of apoptotic and non-apoptotic signaling in cd95 system. Cells, 13:1814, Nov 2024. URL: https://doi.org/10.3390/cells13211814, doi:10.3390/cells13211814. This article has 12 citations.
-
(seyrek2024thecrosstalkof media 9097b778): Kamil Seyrek, Johannes Espe, Elisabeth Reiss, and Inna N. Lavrik. The crosstalk of apoptotic and non-apoptotic signaling in cd95 system. Cells, 13:1814, Nov 2024. URL: https://doi.org/10.3390/cells13211814, doi:10.3390/cells13211814. This article has 12 citations.
-
(seyrek2024thecrosstalkof media cf4ad51c): Kamil Seyrek, Johannes Espe, Elisabeth Reiss, and Inna N. Lavrik. The crosstalk of apoptotic and non-apoptotic signaling in cd95 system. Cells, 13:1814, Nov 2024. URL: https://doi.org/10.3390/cells13211814, doi:10.3390/cells13211814. This article has 12 citations.
-
(seyrek2024thecrosstalkof media 39ce50a4): Kamil Seyrek, Johannes Espe, Elisabeth Reiss, and Inna N. Lavrik. The crosstalk of apoptotic and non-apoptotic signaling in cd95 system. Cells, 13:1814, Nov 2024. URL: https://doi.org/10.3390/cells13211814, doi:10.3390/cells13211814. This article has 12 citations.
-
(seyrek2024thecrosstalkof media 9276d39c): Kamil Seyrek, Johannes Espe, Elisabeth Reiss, and Inna N. Lavrik. The crosstalk of apoptotic and non-apoptotic signaling in cd95 system. Cells, 13:1814, Nov 2024. URL: https://doi.org/10.3390/cells13211814, doi:10.3390/cells13211814. This article has 12 citations.
-
(xu2024genetictestingin pages 2-4): Xinxiu Xu, James Denton, Yaning Wu, Jie Liu, Qiaoning Guan, D. Brian Dawson, Jack Bleesing, and Wenying Zhang. Genetic testing in patients with autoimmune lymphoproliferative syndrome: experience of 802 patients at cincinnati children’s hospital medical center. Journal of Clinical Immunology, Jul 2024. URL: https://doi.org/10.1007/s10875-024-01772-z, doi:10.1007/s10875-024-01772-z. This article has 5 citations and is from a domain leading peer-reviewed journal.
-
(xu2024genetictestingin pages 1-2): Xinxiu Xu, James Denton, Yaning Wu, Jie Liu, Qiaoning Guan, D. Brian Dawson, Jack Bleesing, and Wenying Zhang. Genetic testing in patients with autoimmune lymphoproliferative syndrome: experience of 802 patients at cincinnati children’s hospital medical center. Journal of Clinical Immunology, Jul 2024. URL: https://doi.org/10.1007/s10875-024-01772-z, doi:10.1007/s10875-024-01772-z. This article has 5 citations and is from a domain leading peer-reviewed journal.
-
(streuer2024treatmentwiththe pages 1-2): Alexander Streuer, Johann-Christoph Jann, Tobias Boch, Maximilian Mossner, Vladimir Riabov, Nanni Schmitt, Eva Altrock, Qingyu Xu, Marie Demmerle, Verena Nowak, Julia Oblaender, Iris Palme, Nadine Weimer, Felicitas Rapp, Georgia Metzgeroth, Anna Hecht, Thomas Höger, Christian Merz, Wolf-Karsten Hofmann, Florian Nolte, and Daniel Nowak. Treatment with the apoptosis inhibitor asunercept reduces clone sizes in patients with lower risk myelodysplastic neoplasms. Annals of Hematology, 103:1221-1233, Feb 2024. URL: https://doi.org/10.1007/s00277-024-05664-5, doi:10.1007/s00277-024-05664-5. This article has 3 citations and is from a peer-reviewed journal.
-
(chang2024can008prolongsoverall pages 1-2): Ian Yi-Feng Chang, Hong-Chieh Tsai, Chia-Hua Chen, Hsiu-Chi Chen, Chia-Wen Huang, Gerald F. Cox, Fang-Min Huang, You-Yu Lin, Ko-Ting Chen, Ya-Jui Lin, and Kuo-Chen Wei. Can008 prolongs overall survival in patients with newly diagnosed gbm characterized by high tumor mutational burden. Aug 2024. URL: https://doi.org/10.1016/j.bj.2023.100660, doi:10.1016/j.bj.2023.100660. This article has 1 citations.
-
(NCT01736436 chunk 1): APG101 in Myelodysplastic Syndrome. Apogenix GmbH. 2013. ClinicalTrials.gov Identifier: NCT01736436
-
(NCT04535674 chunk 1): Asunercept in Patients With Severe COVID-19. Apogenix AG. 2020. ClinicalTrials.gov Identifier: NCT04535674
-
(seyrek2024thecrosstalkof pages 20-21): Kamil Seyrek, Johannes Espe, Elisabeth Reiss, and Inna N. Lavrik. The crosstalk of apoptotic and non-apoptotic signaling in cd95 system. Cells, 13:1814, Nov 2024. URL: https://doi.org/10.3390/cells13211814, doi:10.3390/cells13211814. This article has 12 citations.
Artifacts
Citations
- seyrek2024thecrosstalkof pages 14-15
- streuer2024treatmentwiththe pages 1-2
- seyrek2024thecrosstalkof pages 1-2
- xu2024genetictestingin pages 1-2
- hu2025fasmediatesapoptosis pages 1-2
- seyrek2024thecrosstalkof pages 2-4
- xu2024genetictestingin pages 2-4
- seyrek2024thecrosstalkof pages 20-21
- https://doi.org/10.1007/s10875-024-01772-z
- https://doi.org/10.1016/j.bj.2023.100660
- https://clinicaltrials.gov/study/NCT01736436
- https://doi.org/10.1007/s00277-024-05664-5
- https://clinicaltrials.gov/study/NCT04535674
- https://doi.org/10.3390/cells13211814
- https://doi.org/10.3389/fcimb.2025.1561102,
- https://doi.org/10.3390/cells13211814,
- https://doi.org/10.1007/s10875-024-01772-z,
- https://doi.org/10.1007/s00277-024-05664-5,
- https://doi.org/10.1016/j.bj.2023.100660,
Perplexity
(FAS-deep-research-perplexity.md)
Perplexity
(FAS-deep-research-perplexity.md)The Fas Receptor (CD95/TNFRSF6): Molecular Architecture, Death-Inducing Signaling, and the Emerging Complexity of Non-Apoptotic Functions in Human Cellular Regulation
Executive Summary
The Fas receptor, encoded by the FAS gene located at chromosome 10q24.1 in humans and officially designated as tumor necrosis factor receptor superfamily member 6 (TNFRSF6), represents one of the most extensively characterized death receptors in cellular biology[1][3]. This 319-amino acid transmembrane glycoprotein, also known by its alternative designations CD95, APO-1, and APT1, functions as a primary mediator of programmed cell death through its interaction with Fas ligand (FasL/CD95L), initiating apoptosis through assembly of the death-inducing signaling complex (DISC)[1][2]. Beyond its canonical role in eliminating unwanted or autoreactive lymphocytes, emerging evidence from recent structural studies and clinical observations reveals that Fas possesses significant non-apoptotic functions mediated through alternative signaling pathways that regulate immune cell survival, migration, inflammatory responses, and metabolic processes[20][37][40]. The Fas/FasL system represents a critical regulatory node in immune homeostasis, with mutations in the FAS gene causing autoimmune lymphoproliferative syndrome (ALPS), while evolutionary changes in human FasL have paradoxically weakened immune surveillance against certain solid tumors through increased susceptibility to plasmin-mediated proteolytic inactivation[44]. This comprehensive analysis examines the molecular mechanisms of Fas-mediated apoptosis, the structural basis of DISC assembly, the tissue-specific differences in signaling pathway utilization, the regulatory mechanisms controlling Fas activity, and the expanding recognition of non-apoptotic Fas functions that fundamentally reshape our understanding of this death receptor's biological role.
Molecular Structure and Genomic Organization of the Fas Receptor
Gene Structure and Chromosomal Localization
The human FAS gene spans approximately 25,255 base pairs on the long arm of chromosome 10 at position 10q24.1, organized in a characteristic arrangement of nine protein-encoding exons distributed along the Watson (plus) strand[1][13]. This genomic organization reflects the evolutionary conservation of the TNF receptor superfamily structure, with exons arranged to encode distinct functional domains of the mature receptor protein[1]. The gene demonstrates remarkable evolutionary conservation across mammalian species, with orthologs identified in mice (chromosome 19), primates, and other vertebrates, indicating that the fundamental architecture and biological roles of Fas have been maintained through hundreds of millions of years of evolution[1][14]. Alternative splicing of the FAS transcript represents an important regulatory mechanism, with the most notable variant involving the skipping of exon 6, which encodes the transmembrane domain, resulting in production of a soluble form of Fas (sFas) that functions as a dominant-negative inhibitor by competing with membrane-bound Fas for FasL binding in the extracellular space[33]. This alternative splicing mechanism provides cells with an intrinsic mechanism to modulate their susceptibility to Fas-mediated apoptosis, particularly important in immune cell populations that must balance activation with controlled elimination[33].
Protein Domain Organization and Structural Features
The mature Fas receptor protein comprises 319 amino acids organized into three functionally and structurally distinct domains: an extracellular domain (157 amino acids), a transmembrane domain (17 amino acids), and an intracellular domain (145 amino acids) with predicted molecular weight of 48 kilodaltons[1][13]. The extracellular region, encoded by exons 1 through 5, is characterized by extraordinary richness in cysteine residues (18 cysteines within 157 amino acids), which form disulfide bonds stabilizing a series of cysteine-rich domains (CRDs) that display a characteristic "ladder-like" structure typical of TNF receptor superfamily members[1][3][48]. Specifically, the extracellular domain encompasses three well-structured cysteine-rich domains (CRD1, CRD2, and the upper portion of CRD3), with CRD1 spanning amino acids 56 to 82, stabilized by numerous disulfide bonds that provide structural rigidity and enable specific ligand recognition[48]. Critically, within the first 55 amino acids preceding CRD1 exists a flexible, intrinsically disordered region that contains the pre-ligand assembly domain (PLAD), a region essential for the constitutive oligomerization of Fas molecules even in the absence of ligand binding[14][48]. The PLAD spans residues 49 to 82 and contains a minimal domain (residues 59-82) sufficient to mediate homotypic Fas-Fas receptor interactions, enabling receptors to pre-associate on the cell surface in a ligand-independent manner that conditions them for rapid activation upon FasL engagement[3][14].
The transmembrane domain, encoded by exon 6 and consisting of 17 amino acids, exhibits remarkable functional importance beyond simple membrane anchoring. Recent nuclear magnetic resonance spectroscopic studies of the transmembrane region in lipid bilayer-mimicking bicelles revealed that the Fas transmembrane domain forms a specific homotrimeric structure utilizing proline-containing motifs to create optimal packing geometry distinct from classical coiled-coil arrangements observed in solution[8]. This transmembrane trimerization appears directly coupled to apoptotic signaling competence, as cancer-associated and structure-based mutations disrupting the precise three-dimensional packing of the transmembrane helices substantially reduce both trimerization efficiency and apoptosis induction in intact cells[8]. The intracellular region, encoded by exons 7 through 9, contains the crucial death domain (DD) spanning amino acids 210 to 303, an 85-amino acid module homologous to similar domains in other death receptors and serving as the primary platform for recruitment of downstream signaling proteins[3]. This death domain is characterized by a high number of charged amino acids (24 basic and 19 acidic residues), properties that mediate protein-protein interactions essential for death signal transduction[2]. The extreme carboxyl terminus (amino acids 303-319), comprising the final 15 amino acids, represents a functionally mysterious region that can interact with regulatory proteins such as protein tyrosine phosphatase FAP-1 and the scaffolding protein Dlg1, which antagonize Fas-mediated apoptosis through mechanisms remaining incompletely characterized[3].
Ligand Binding and Pre-Ligand Assembly in Fas Activation
The Nature of Fas Ligand and Its Structural Requirements
Fas ligand exists in two primary forms with distinctly different biological consequences: membrane-bound Fas ligand (mFasL) and soluble Fas ligand (sFasL) generated through metalloprotease-mediated ectodomain shedding[34][40][55]. Membrane-bound FasL presents as a multimeric complex of membrane-anchored homotrimeric units, with increasing evidence suggesting that FasL exists as aggregated trimers creating higher-order oligomeric complexes on the cell surface[1][10]. This multivalency of membrane-bound FasL proves absolutely essential for apoptotic signaling, as synthetic monomeric or even simple homotrimeric soluble FasL fails to trigger apoptosis, whereas engineered hexameric forms of FasL robustly induce cell death by clustering six Fas receptors into a signaling-competent complex[10][35]. The soluble form of FasL (sFasL) generated by proteolytic cleavage predominantly exists as a homotrimer, and critically, this trimeric form shows substantially diminished apoptotic capability compared to its membrane-anchored counterpart[40][55]. Instead, sFasL preferentially triggers non-apoptotic signaling pathways by activating a distinct molecular complex designated the motility-inducing signaling complex (MISC) that recruits different adaptor proteins and kinases than the apoptosis-mediating DISC[40][55]. The structural basis for this functional divergence relates to the presence of a "stalk region" in the sFasL molecule that connects the extracellular trimeric ligand head to the membrane anchoring point in mFasL; removal of this stalk region by certain metalloproteases produces a sFasL molecule incapable of triggering apoptosis regardless of its oligomeric status[34][40][55].
Pre-Ligand Receptor Assembly and the PLAD
One of the most profound paradigm shifts in understanding Fas biology involved the recognition that Fas receptor exists as pre-assembled oligomers on the cell surface prior to ligand binding, a property mediated by the pre-ligand assembly domain (PLAD) residing within the extracellular cysteine-rich domain[14][17]. This pre-assembly occurs independently of FasL, with approximately 46% of wild-type Fas molecules existing in oligomeric states (clusters of 2-3 molecules representing 75% of such clusters) in unstimulated cells[8]. Single-molecule microscopy studies employing photoactivated localization microscopy (PALM) combined with photoactivatable green fluorescent protein (PA-GFP) directly visualized these pre-formed Fas receptor assemblies at the plasma membrane, establishing that receptor oligomerization is an intrinsic property of the receptor rather than a ligand-induced phenomenon[8]. The biological significance of PLAD-mediated pre-assembly became apparent when disruption of PLAD-PLAD interactions completely eliminated Fas signaling capacity, despite preservation of ligand binding capability, indicating that pre-assembled trimers create a conformational platform essential for subsequent ligand-induced signaling[14][17][48]. Molecular models reveal that pre-assembled Fas likely exists in anti-parallel dimer or trimer configurations where critical ligand-binding regions are masked by inter-receptor interfaces, creating an inactive state that requires ligand-induced conformational rearrangement to expose the FasL-binding surfaces[48]. Upon binding of multivalent membrane-bound FasL, the ligand trimerizes and links multiple pre-assembled Fas trimers into increasingly large receptor aggregates, with evidence suggesting that a minimum of six Fas receptors (2 Fas trimers) becomes necessary for achieving full signaling competence[10][48].
The Death-Inducing Signaling Complex: Structure and Assembly Mechanism
Molecular Architecture of DISC Assembly
The death-inducing signaling complex represents the proximal signaling platform where Fas oligomerization couples to proteolytic activation of the caspase cascade initiating programmed cell death. Recent cryogenic electron microscopy (cryo-EM) structures have provided unprecedented mechanistic insight into DISC assembly, revealing that Fas and FADD death domains form an asymmetric dodecameric complex composed of seven Fas death domains and five FADD death domains arranged in a distinctive three-layered architecture[10][45]. The upper and middle layers comprise the seven Fas death domains, while the five FADD death domains form the bottom layer, creating a geometry reminiscent of the caspase-2-activating PIDDosome complex[10][45]. This 7:5 stoichiometry appears to represent the physiologically relevant complex under native conditions, though mass spectrometry studies have documented that Fas-FADD complexes can assemble with varying stoichiometries ranging from 5:5 to 7:5 depending on crystallization conditions and protein concentrations, suggesting that the oligomeric state represents an equilibrium process responsive to local protein concentrations and cellular conditions[10][45]. The Fas death domain interaction interfaces display exquisite specificity, with multiple distinct interaction surfaces operating at different stages of complex assembly. Residues within the first 49 amino acids of the Fas death domain (encompassing the pre-ligand assembly domain) mediate crucial homo-oligomerization contacts, while other death domain residues form the primary interface for FADD death domain binding[3].
The assembly process follows an ordered mechanism: upon massive clustering of Fas receptors by membrane-bound FasL, a striking conformational transition occurs where Fas death domains transition from a "closed" conformation (as observed in isolated death domain structures) to an "open" configuration that exposes critical binding surfaces for FADD interaction[11]. This opening mechanism appears coupled to the lateral diffusion and clustering of pre-assembled Fas trimers, whereby the stabilization of these trimers in large aggregates on the membrane provides the mechanical force necessary to induce and stabilize the conformational transitions required for FADD recruitment[10][11]. The opening transition specifically involves a repositioning of helix 6 of the Fas death domain, a structural element that, when mutated to favor the open conformation, actually enhances apoptotic signaling despite potentially destabilizing the closed state, underscoring the biological importance of achieving and maintaining the open conformation for efficient DISC function[3]. Once FADD death domains bind to the open Fas death domain surfaces, they position the death effector domains (DEDs) at the appropriate spatial orientation and density to nucleate the recruitment and subsequent oligomerization of procaspase-8 molecules.
FADD-Mediated Caspase-8 Recruitment and Activation
Fas-associated death domain (FADD) protein, also designated MORT1 (mediator of receptor-induced toxicity 1), serves as the critical adaptor molecule linking Fas death domain signaling to caspase-8 activation[2][5][7]. FADD consists of two functionally distinct domains connected by a flexible linker: an amino-terminal death effector domain (DED) and a carboxyl-terminal death domain (DD) that binds to Fas[3][7]. The death domain of FADD exhibits remarkable flexibility, with amino acid residues at the interface between FADD and Fas showing variability in their exact spatial positioning, a property that may contribute to the dynamic nature of DISC assembly and disassembly[3]. The DED of FADD contains tandemly repeated DED modules (DED1 and DED2) separated by a linker region, creating a structure that closely resembles the similar DED architecture of caspase-8[10][45]. Upon recruitment to the Fas-FADD death domain complex, FADD death effector domains undergo a remarkable conformational transition, assembling into elongated helical filament structures that nucleate the subsequent polymerization of caspase-8 tandem DED molecules[10][45]. These FADD DED filaments, which exhibit near-complete structural overlap with caspase-8 DED filaments when DED1 and DED2 are considered as equivalent structures, appear to serve as a molecular scaffold that promotes both the positioning and the proximity-induced dimerization of caspase-8 zymogens[10][45].
Procaspase-8, the inactive zymogen form of caspase-8 (also designated FLICE for FADD-like IL-1β-converting enzyme), exists as a monomer in the cytosol until recruited into the DISC through its death effector domains[5][10]. The recruitment of multiple procaspase-8 molecules through their DEDs into the FADD DED filament brings multiple procaspase-8 molecules into sufficient proximity that enables autocatalytic activation through intermolecular cleavage[10][45]. Specifically, the enzymatic domain of one procaspase-8 molecule cleaves the activation loop of a proximate procaspase-8 molecule, resulting in removal of the inhibitory prodomain and generation of the active heterodimeric form of caspase-8 composed of a p18 (large) and p10 (small) subunit[1][2]. This proximity-induced or induced-dimerization mechanism of caspase-8 activation represents a fundamental strategy utilized throughout death receptor signaling, wherein the spacing and positioning afforded by adaptor proteins creates the necessary structural framework for catalytic activation of proteases that are otherwise kept inactive through spatial segregation[5][7][10].
Once activated within the DISC, caspase-8 exists in a state of limited activity, constrained by its association with other DISC components and by the limited availability of certain substrates at the DISC locale[2][5]. Active caspase-8 subsequently dissociates from the DISC, translocating into the cytosolic compartment where it gains unrestricted access to its full complement of proteolytic substrates[1][2]. This physical separation between DISC assembly (at the plasma membrane and in endocytic compartments) and the executioner phase of apoptosis (in the cytosol) provides an additional layer of regulatory control, permitting signal amplification and checkpoint control before commitment to complete cellular demolition[1][2][5].
Type I Versus Type II Cell Death Pathways
Differential Utilization of Apoptotic Cascades in Distinct Cell Types
Fas-expressing cells exhibit profound differences in their mechanisms of death in response to FasL stimulation, leading to their classification as either Type I or Type II cells based on whether they require amplification of the caspase cascade through the intrinsic mitochondrial apoptosis pathway[32][35]. Type I cells, exemplified by thymocytes, mature T lymphocytes, and certain transformed cell lines, respond to Fas engagement with robust and rapid caspase-3 and -7 activation that suffices for efficient apoptosis without requiring mitochondrial outer membrane permeabilization (MOMP) or release of cytochrome c[32][35]. In these cells, the amount of caspase-8 activated within the DISC proves sufficient to directly cleave and activate effector caspases (caspase-3 and -7) to levels exceeding the inhibitory capacity of X-chromosome-linked inhibitor of apoptosis protein (XIAP), permitting rapid progression to the executioner phase of apoptosis[32][35]. The rapid kinetics of Type I cell death, often occurring within 2-5 hours of Fas stimulation, reflects both the efficient apoptotic machinery present in these cells and their historical evolutionary role as cells requiring rapid elimination when autoreactive or aberrantly activated[32].
In sharp contrast, Type II cells, including hepatocytes, pancreatic β-cells, and certain fibroblast populations, demonstrate substantially slower kinetics of Fas-induced death, requiring amplification through the intrinsic apoptotic pathway for efficient killing[32][35]. In these cells, the amount of caspase-8 directly activated by the DISC generates insufficient effector caspase activity to overcome XIAP-mediated inhibition, necessitating engagement of the mitochondrial apoptotic program[32][35]. This amplification pathway utilizes the BH3-interacting domain death agonist (Bid), a BH3-only member of the Bcl-2 protein family that exists in an inactive state in resting cells[32][35]. Activated caspase-8 within the DISC cleaves Bid at a specific aspartate residue, generating a truncated fragment (tBid) that inserts into the outer mitochondrial membrane and promotes conformational changes in the pro-apoptotic Bcl-2 family members Bax and Bak, leading to mitochondrial outer membrane permeabilization[32][35]. This permeabilization releases multiple pro-apoptotic factors including cytochrome c, which combines with cytoplasmic Apaf-1 and procaspase-9 to assemble the apoptosome, a macromolecular platform that catalyzes the activation of caspase-9, which in turn cleaves and activates caspase-3 and -7 in a feed-forward manner[5][32][35]. Additionally, mitochondrial permeabilization releases Smac/DIABLO, an antagonist of IAPs, which reduces XIAP-mediated caspase inhibition and further amplifies the caspase cascade[32][35].
The mechanistic basis for Type I versus Type II cell classification appears linked to differential regulation of XIAP protein levels in response to Fas activation. In Type I cells (thymocytes), Fas stimulation triggers rapid proteolytic degradation of XIAP through a caspase-3-dependent mechanism, reducing inhibitory capacity and permitting efficient effector caspase activation[32]. Remarkably, in Type II cells (hepatocytes), the identical caspase-3 activation triggered by Fas stimulation leads to an increase in XIAP protein levels, a seemingly paradoxical effect that reflects differential regulation through distinct post-translational modification pathways dependent on the cell type[32]. This enhanced XIAP levels in Type II cells effectively increases the apoptotic threshold, necessitating the amplification provided by Bid-mediated mitochondrial engagement to achieve sufficient caspase-3/7 activation for cell death[32]. The differential XIAP regulation appears to be a fundamental property distinguishing these cell types, as deletion of Xiap in Bid-deficient Type II cells reverses their resistance to Fas-mediated lethality, restoring death kinetics approaching those of wild-type cells despite the absence of functional Bid protein[32][35].
Tissue-Specific Differences and Physiological Significance
The differential involvement of Type I versus Type II pathways in various tissues reflects their distinct biological roles and the need for tissue-specific regulation of inflammatory responses. The liver demonstrates exceptional sensitivity to Fas-mediated apoptosis, with systemic administration of Fas agonist antibodies triggering fulminant hepatitis and death within hours through massive hepatocyte apoptosis[49][52]. This hepatic sensitivity relates to the tissue's high expression of Fas receptor and the Type II status of hepatocytes, requiring mitochondrial pathway amplification[49][52]. Importantly, the hepatotoxic response to Fas activation appears synergistically enhanced by TNF signaling, wherein low-level TNF exposure sensitizes hepatocytes to Fas-mediated apoptosis through mechanisms involving the activation of stress signaling kinases including JNK, p38, and the transcription factor NF-κB[49]. This TNF-Fas interplay is physiologically relevant to drug-induced liver failure, wherein drugs such as acetaminophen trigger both TNF expression and Fas receptor upregulation, explaining the often catastrophic nature of acute liver failure in some drug overdose situations[49].
The lymphoid organs, particularly the thymus and germinal centers of secondary lymphoid tissues, represent sites where Fas-mediated apoptosis exerts profound regulatory effects on immune development and the resolution of immune responses. In the thymus, Fas/FasL interactions participate in the deletion of autoreactive T cells, although the relative contribution of Fas-mediated death versus other mechanisms remains incompletely understood[5][15]. In the peripheral immune system, Fas-mediated apoptosis plays an essential role in activation-induced cell death (AICD), a mechanism whereby repeated TCR stimulation induces expression of both Fas and FasL on T cells, creating an autocrine/paracrine death signal that terminates immune responses[5][22]. The germinal center reaction, wherein B cells with low-affinity or autoreactive B cell receptors are eliminated through Fas-FasL interactions with CD4+ helper T cells, represents another critical physiological application of Fas-mediated death in maintaining immune tolerance[5][27].
Regulation of Fas Signaling Through Adaptor Proteins and Post-Translational Modifications
The c-FLIP Protein and Bifurcation of Apoptotic Versus Non-Apoptotic Signaling
The cellular FLICE-inhibitory protein (c-FLIP), which exists in both long (c-FLIP_L) and short (c-FLIP_S) isoforms generated through alternative splicing, represents perhaps the most critical endogenous regulator of the balance between Fas-mediated apoptosis and non-apoptotic signaling[5][57]. c-FLIP contains two death effector domains nearly identical to those of caspase-8 and can therefore bind to FADD through DED-DED interactions, competing with caspase-8 for positions within the DISC[5]. The c-FLIP_S isoform, containing only the DEDs without the catalytic domain, functions as a dominant inhibitor of DISC-mediated caspase-8 activation, with overexpression of c-FLIP_S reducing Fas-induced apoptosis by 80-95% depending on the cell type[5][51]. The c-FLIP_L isoform, containing an intact but catalytically inactive caspase-like domain, exhibits more complex effects, retaining the ability to inhibit apoptosis while simultaneously providing a scaffolding platform for recruitment of kinases that mediate non-apoptotic signaling pathways[5][37].
Genetic studies utilizing c-FLIP-deficient mice revealed that this protein possesses critical non-apoptotic functions beyond simple inhibition of caspase-8, as its complete loss causes embryonic lethality phenocopying that observed in caspase-8-deficient mice[5]. This surprising observation indicated that c-FLIP, when not simply inhibiting caspase-8, must serve important roles in promoting non-apoptotic signaling necessary for immune cell development and homeostasis[5]. Selective deletion of c-FLIP in T lymphocytes or fibroblasts paradoxically accelerated Fas-induced apoptosis in these cells, confirming its apoptosis-inhibitory role in isolation[5]. However, deletion of c-FLIP in all tissues produced embryonic lethality associated with defective immune development and impaired NF-κB signaling, indicating its role in supporting non-apoptotic Fas signaling essential for proper immune function[5]. This led to the recognition that the relative abundance of c-FLIP versus caspase-8 recruited into the DISC effectively determines the signaling outcome: high caspase-8 relative to c-FLIP favors apoptosis, whereas high c-FLIP relative to caspase-8 promotes non-apoptotic kinase signaling through mechanisms involving recruitment of RIP, TRAF proteins, and MAP kinases[5][37][57].
Post-Translational Modifications Regulating Fas Signaling
The Fas receptor and its associated signaling machinery are subject to multiple post-translational modifications that fine-tune the balance between apoptotic and non-apoptotic signaling responses. Ubiquitination represents one such modification, with E3 ubiquitin ligases targeting Fas and FADD for polyubiquitin chain formation that can either promote signaling (through non-degradative K63-linked chains) or antagonize signaling (through K48-linked chains promoting proteasomal degradation)[29]. Phosphorylation of Fas and downstream signaling components by various kinases including Src family kinases, Akt, ERK1/2, and JNK modulates DISC formation efficiency and kinase pathway activation[40][57]. Notably, phosphorylation of specific tyrosine residues on Fas can promote the recruitment of SH2-domain-containing proteins including c-yes and Src kinases, directly coupling Fas engagement to non-apoptotic kinase signaling pathways[40].
S-glutathionylation of Fas at cysteine 294 (murine sequence, corresponding to human cysteine residue 295) represents another regulatory modification that markedly promotes Fas receptor aggregation and subsequent caspase-8 activation[3]. This post-translational modification can be reversed by glutaredoxin enzymes, creating a regulatory cycle whereby reactive oxygen species generated during cellular stress enhance Fas signaling capacity through promoting S-glutathionylation, while antioxidant enzymes antagonize this response[3]. This mechanism provides a molecular link between oxidative stress and Fas-mediated apoptosis, with implications for understanding how cells integrate stress signals to make life-death decisions. Interestingly, caspase-8 itself cleaves and inactivates glutaredoxin-1 (Grx1), creating a positive feedback loop wherein Fas activation enhances oxidative conditions that further promote Fas signaling through continued glutathionylation[3].
Internalization and Membrane Trafficking of Fas Receptor Complexes
Following Fas activation, the receptor undergoes rapid internalization through multiple endocytic pathways, with the internalized DISC compartments concentrating around the Golgi apparatus in a polarized distribution distinct from the general plasma membrane endocytic patterns[39]. Fas stimulation preferentially enhances clathrin-independent endocytic pathways, particularly those dependent on Rho GTPases including CDC42, more than classical clathrin-mediated endocytosis[28][39]. This selective enhancement of specific endocytic portals suggests that Fas-activated signaling drives internalization through routes that may spatially segregate the apoptotic DISC from potential inhibitory factors present in early endosomes or at the plasma membrane. Activated caspases associated with the internalized DISC complexes exhibit perinuclear concentration in the vicinity of recycling endosomes around the Golgi region, indicating that the compartmentalization of active caspases to distinct cellular microdomains may contribute to controlled and localized apoptotic signaling[28][39]. The trafficking of GPI-anchored proteins such as CD59 is markedly altered upon Fas stimulation, with rapid internalization and peri-Golgi accumulation that parallels the trafficking of the apoptotic machinery itself, suggesting that Fas-triggered membrane reorganization may facilitate the internalization of specific lipid raft-associated signaling platforms[28][39].
Lipid raft compartmentalization appears to significantly influence Fas signaling outcome, with Fas receptor and its ligand showing enriched localization to cholesterol- and sphingolipid-rich membrane microdomains[42]. The initial clustering of Fas molecules into lipid raft structures upon FasL binding may represent an early organizing event that facilitates DISC assembly by concentrating the necessary signaling components into discrete membrane domains. Soluble FasL, lacking the membrane anchoring and multivalency of membrane-bound FasL, fails to similarly concentrate Fas into rafts and instead promotes the assembly of alternative signaling complexes (MISC) that activate non-apoptotic pathways. This mechanistic distinction between membrane-bound and soluble ligand signaling highlights how the physical organization of signaling components within specialized membrane microdomains fundamentally determines the biological outcome of Fas engagement.
Non-Apoptotic Functions of Fas Receptor Signaling
Discovery and Characterization of Non-Apoptotic Fas Pathways
Contemporaneous with the accumulation of structural and biochemical evidence for the apoptotic function of Fas, numerous observations emerged indicating that Fas can engage additional signaling pathways distinct from the apoptotic cascade[23][37][40][55]. The most direct evidence came from cell populations that proved refractory or resistant to Fas-induced apoptosis despite expressing high levels of functional Fas receptor and intact downstream apoptotic machinery. For instance, Th2 cells exhibit marked resistance to Fas-mediated apoptosis while nonetheless responding to FasL stimulation with activation of NF-κB, enhanced gene expression, and cellular differentiation[37]. Similarly, glioblastoma and other tumor cell lines develop resistance to Fas-induced apoptosis through either mutation of the Fas gene itself or through overexpression of anti-apoptotic proteins, yet these same cells respond to Fas engagement with enhanced migration, invasion, and metastatic dissemination through activation of non-apoptotic kinase cascades[37][40][55].
The molecular basis for non-apoptotic Fas signaling involves an alternative configuration of the Fas signaling complex wherein soluble FasL (rather than membrane-bound FasL) engages the receptor, or wherein sufficient c-FLIP concentration within DISC-like complexes redirects the signaling outcome from caspase-8-mediated apoptosis toward kinase activation[40][55]. Recent studies characterizing this alternative complex designated the motility-inducing signaling complex (MISC) revealed that it completely lacks both FADD and caspase-8 proteins, instead recruiting the Src family kinase c-yes through a mechanism involving elevated intracellular reactive oxygen species (ROS) generated through NADPH oxidase 3[40]. The MISC appears to form primarily in response to soluble FasL stimulation and lacks the adaptor proteins critical for apoptotic signaling, instead assembling kinases and signaling molecules that activate the PI3K/Akt, ERK1/2 MAP kinase, and NF-κB pathways[40][55]. These kinase pathways promote cell survival, proliferation, migration, and inflammatory cytokine production rather than apoptosis, creating a scenario where Fas stimulation can either kill cells or promote their activation and survival depending on the cellular context and the form of FasL encountered[40][55].
Non-Apoptotic Signaling and Immune Function
Recent investigations utilizing mice bearing Fas mutations that specifically block apoptotic signaling while preserving non-apoptotic pathway capacity revealed the critical importance of non-apoptotic Fas functions in immune homeostasis[20]. Studies of patients with autoimmune lymphoproliferative syndrome (ALPS) bearing FAS mutations demonstrated that defective non-apoptotic Fas signaling, independent of apoptosis resistance, contributes to impaired B cell differentiation and germinal center formation[20]. Mechanistically, transient engagement of FAS upon T-dependent B cell activation decreases mTOR activation through a process dependent on caspase-8 and involving nuclear exclusion of PTEN, a critical negative regulator of PI3K signaling[20]. This non-apoptotic mTOR modulation by FAS signaling acts as a molecular switch determining whether activated B cells commit to germinal center differentiation (associated with lower mTOR activity and appropriate transcriptional programs) versus extrafollicular differentiation producing short-lived plasma cells with impaired antibody affinity maturation[20]. In B cells from ALPS-FAS patients, the defect in FASL-mediated PTEN nuclear exclusion and mTOR inhibition correlates with expansion of extrafollicular B cell responses and reduced germinal center formation, despite intact apoptotic resistance[20]. This finding reveals that non-apoptotic Fas signaling provides essential instructions for proper immune development independent of its death-inducing capacity.
The role of non-apoptotic Fas signaling in regulating Th2-mediated inflammation further exemplifies the importance of these alternative pathways[37]. In allergic airway inflammation models, Fas-deficiency on T cells delays resolution of type 2 inflammation and eosinophilia, but this defect cannot be overcome by increasing T cell survival through deletion of pro-apoptotic molecules (Bim) or overexpression of anti-apoptotic molecules (Bcl-xL)[37]. Instead, using mice bearing Fas mutations that specifically ablate apoptotic signaling while preserving non-apoptotic functions, investigators demonstrated that non-apoptotic Fas signaling in Th2 cells drives resolution of inflammation[37]. This non-apoptotic signaling, occurring specifically in Th2 cells which exhibit inherent resistance to Fas-induced apoptosis and preferentially activate NF-κB upon FasL exposure, appears critical for implementing the transcriptional and functional changes necessary for inflammation resolution[37]. These findings suggest that Fas may function in distinct cellular contexts as either a death receptor (in type I cells) or as an activating receptor promoting cellular processes essential for immune homeostasis (in apoptosis-resistant cell populations).
Non-Apoptotic Fas Signaling in Cell Migration and Metastasis
The soluble form of FasL detected in serum of patients with aggressive solid tumors, particularly triple-negative breast cancer, colon cancer, and ovarian cancer, activates non-apoptotic signaling pathways that enhance tumor cell migration, invasion, and metastatic dissemination[40][55]. The MISC complex assembled in response to sFasL stimulation recruits Src family kinases and promotes phospholipase C-γ1 (PLC-γ1) activation, generating inositol 1,4,5-trisphosphate (IP3) and diacylglycerol that trigger intracellular calcium mobilization and protein kinase C activation[40][56]. These signaling cascades, collectively termed calcium-dependent signaling, promote cell migration, focal adhesion dynamics, and cytoskeletal rearrangement necessary for metastatic dissemination[40][56]. The elevated plasmin levels in the tumor microenvironment of solid tumors creates a particularly hostile context for immune cell killing, as plasmin proteolytically cleaves human FasL at a unique cleavage site created by a single evolutionary amino acid change (serine at position 153 instead of proline, unique to humans compared to non-human primates) that renders human FasL uniquely susceptible to plasmin-mediated inactivation[44]. This evolutionary vulnerability in human FasL, apparently selected for in our species to enhance neural development, creates a substantial disadvantage in immune surveillance against plasmin-expressing solid tumors, explaining the frequent failure of CAR-T and other T cell immunotherapies in solid tumors despite their efficacy in blood cancers lacking plasmin expression[44].
Clinical Manifestations of Fas Dysfunction: Autoimmune Lymphoproliferative Syndrome
Molecular Basis of ALPS Pathogenesis
Autoimmune lymphoproliferative syndrome (ALPS) represents a rare but clinically significant genetic disorder caused by germline mutations affecting Fas-mediated apoptosis[21][24][47]. Most ALPS patients carry heterozygous missense mutations in the FAS gene that encode defective Fas proteins exhibiting dominant-negative effects on wild-type Fas signaling[47]. The disease manifests with characteristic clinical features including early-onset lymphadenopathy (often presenting in childhood), splenomegaly, immune cytopenias (low blood counts due to autoimmune destruction), and significantly elevated risk for B cell lymphomas, reflecting the loss of Fas-mediated elimination of autoreactive lymphocytes and impaired immune homeostasis[21][24][47]. The hallmark lymphoproliferative phenotype involves massive expansion of CD4-CD8- double-negative (DN) T cells expressing the B cell marker B220, creating a distinctive circulating population of unusual T cells not present in healthy individuals[21][38][47].
The molecular mechanisms by which FAS mutations cause ALPS demonstrate remarkable diversity despite the unified clinical phenotype[47]. Mutations affecting the extracellular domain of Fas frequently result in reduced surface expression of the receptor through nonsense-mediated decay of the mutant transcript or proteasomal degradation of the misfolded protein, a mechanism termed haploinsufficiency[47]. These extracellular-region mutations attenuate DISC formation and apoptosis signaling to a lesser extent than intracellular domain mutations, and notably, patients with haploinsufficient FAS mutations show a lower penetrance of clinical symptoms and, strikingly, have not developed lymphomas despite decades of observation, in contrast to patients bearing intracellular domain mutations who frequently develop lymphomas[47]. This observation suggests that residual Fas function, even when reduced to 50% of normal levels through haploinsufficiency, may suffice for tumor surveillance function. Mutations affecting the intracellular death domain generally produce dominant-negative effects wherein the defective Fas protein interferes with signaling even from wild-type Fas on the same cell[47]. These intracellular mutations typically prevent recruitment of FADD or cause instability of the DISC complex, demonstrating the critical importance of death domain integrity for Fas signaling[7][47].
Pathogenesis of Lymphoproliferation and Autoimmunity
The expansion of double-negative T cells in ALPS represents an enigmatic aspect of the disease, as these unusual cells rarely develop in healthy individuals yet proliferate massively when Fas is defective[38]. Recent investigations revealed that dysregulated expression of Eomesodermin (Eomes), a T-box transcription factor critical for effector and memory CD8+ T cell function, characterizes DN T cells in both ALPS mice (lpr mice) and ALPS patients[38]. T cell-specific deletion of Eomes substantially ameliorated the DN T cell expansion and lymphoproliferation of ALPS mice, establishing Eomes dysregulation as essential for DN T cell pathogenesis[38]. However, residual lymphoproliferation persisted in ALPS mice lacking T cell Eomes, indicating that additional Fas-dependent mechanisms beyond Eomes-regulated DN T cell expansion contribute to the overall lymphoproliferative phenotype[38]. The expansion of DN T cells, while dramatic, appears less important than B cell dysfunction for driving the autoimmune manifestations of ALPS, as selective deletion of Fas from B cells or antigen-presenting cells suffices to reproduce the autoimmune phenotype of systemic ALPS[19][27].
Investigations of Fas-mediated control of antigen-presenting cells (APCs) revealed that mature antigen-presenting cells, particularly dendritic cells, express elevated levels of Fas receptor following exposure to microbial stimuli, predisposing them to Fas-mediated death by activated T cells[19]. The elimination of Fas-positive antigen-presenting cells by T cells bearing FasL represents a negative feedback mechanism that terminates immune responses by destroying the cells presenting antigenic stimuli[19]. Tissue-specific deletion of Fas in dendritic cells proved sufficient to cause systemic autoimmunity and lymphoproliferation, establishing that Fas-mediated elimination of activated antigen-presenting cells represents a critical control mechanism preventing persistent activation of self-reactive T cells[19]. Similarly, deletion of Fas specifically from B cells resulted in their accumulation and systemic autoimmunity characterized by elevated autoantibodies and immune complex deposition[19][27]. The B cell-intrinsic defect in ALPS appears to involve both failure of Fas-mediated elimination of low-affinity and autoreactive B cells within germinal centers and defective generation of memory B cells, reflecting both apoptotic and non-apoptotic functions of Fas in B cell physiology[20][27].
Evolutionary Significance and Comparative Biology of Fas
Conservation and Evolutionary Divergence
The Fas receptor belongs to an evolutionarily ancient signaling system, with orthologs identified throughout vertebrate species and even invertebrate organisms, indicative of its fundamental importance in organismal biology[43][46]. Molecular phylogenetic analysis of death receptors including Fas, TNFR1, DR3, DR4, DR5, and DR6 revealed that Fas represents a somewhat divergent member of the TNF receptor family, having evolved from a common ancestor with TNFR family members more than 500 million years ago, before the radiation of modern animal phyla[43][46]. Despite this evolutionary distance, Fas maintains the characteristic structural organization of TNF receptors, including the cysteine-rich extracellular domains with their disulfide-bonded ladder-like architecture and the intracellular death domain critical for signaling[43][46]. The strong conservation of specific cysteine residues throughout the extracellular domain across diverse species suggests that these residues provide essential structural constraints maintaining the overall three-dimensional organization necessary for ligand binding and receptor activation[43]. Notably, more amino acid residues exhibit conservation in TNFR1, DR3, and TNFBR compared to Fas (16 highly conserved residues in TNFR1, DR3, TNFBR versus only 7 in Fas), suggesting that Fas may be more permissive of sequence variation while maintaining overall functional capacity, or alternatively, that Fas evolved more rapidly following its divergence from common ancestors.
Evolutionary Trade-offs: Human FasL Vulnerability to Plasmin
A striking and evolutionarily significant finding emerged from comparative analysis of Fas ligand sequences across primates and humans[44]. Humans possess a unique evolutionary substitution in the FasL protein at position 153, where a proline residue present in all non-human primates (including chimpanzees) is replaced with serine in humans[44]. This single-residue change creates a novel proteolytic cleavage site for plasmin, a serine protease frequently elevated in aggressive solid tumors that are heavily dependent on this enzyme for metastasis[44]. The consequence of this human-specific mutation appears paradoxical: while it may have contributed to the evolutionary expansion of the human cerebral cortex (perhaps through effects on neural development), it simultaneously renders human FasL extremely vulnerable to proteolytic inactivation by plasmin in the tumor microenvironment of solid malignancies[44]. In vitro experiments demonstrated that human FasL is cleaved and inactivated by plasmin with high efficiency, rendering immune cells unable to trigger apoptosis in tumor cells even when properly activated and armed with FasL molecules[44]. Remarkably, non-human primates such as chimpanzees exhibit much lower rates of solid tumors compared to humans, a difference that may partially reflect this evolutionary vulnerability in human FasL to plasmin-mediated inactivation in tumor environments[44].
This discovery has direct implications for cancer immunotherapy, providing a mechanistic explanation for the frequent failure of CAR-T and T cell-based therapies against solid tumors while these same approaches show substantial efficacy against blood cancers that typically do not express high levels of plasmin[44]. The differential dependence of blood cancers versus solid tumors on plasmin-mediated proteolysis and matrix degradation creates an environment wherein human immune cells' primary death-inducing weapon (FasL) becomes disabled specifically in solid tumor contexts[44]. Blocking plasmin or engineering protective antibodies that shield FasL from plasmin cleavage restored apoptosis-inducing capacity in in vitro experiments, suggesting that combination approaches combining anti-plasmin strategies with FasL-based or T cell immunotherapies may enhance efficacy against solid tumors[44]. This example illustrates how evolutionary changes in immune molecules, apparently selected for beneficial effects on neural development, can create substantial vulnerabilities to modern age-related diseases not encountered in our evolutionary history.
Integration of Signaling Pathways: Type I and Type II Mechanisms Revisited
Recent Structural Insights into DISC Assembly and Signaling Amplification
The latest cryo-EM structures of the Fas-FADD complex, published in 2024, provide unprecedented mechanistic clarity regarding how the asymmetric 7:5 oligomeric architecture translates receptor clustering into caspase-8 activation and apoptotic signal amplification[10][45]. The structural analysis reveals that the FADD death effector domains emanating from the bottom layer of the complex form extended filamentous structures that closely resemble the native architecture of caspase-8 tandem DED filaments[10][45]. This structural similarity is not coincidental but appears essential for the mechanism of caspase-8 assembly and activation, as FADD DED filaments effectively serve as nucleation scaffolds that promote the polymerization of caspase-8 tandem DED domains into similar helical structures[10][45]. The hypothesis emerging from this structural work proposes that FADD DED filaments lower the kinetic barrier for caspase-8 tandem DED assembly by providing a pre-organized template of charged and hydrophilic residues that precisely match those required for productive caspase-8 DED-DED interactions[10][45]. Three distinct DED interface types (Type I hydrophobic, Type II and III hydrophilic/charged) mediate both FADD-FADD interactions within the filament and FADD-caspase-8 interactions, with the electrostatic properties of FADD DED closely resembling those of individual caspase-8 DED domains, enabling seamless integration of caspase-8 molecules into the growing FADD filament[10][45].
This structural mechanism of signal amplification through filamentous assembly explains several previously enigmatic observations. The requirement for hexameric (or higher-order oligomeric) FasL to trigger apoptosis, despite the trimeric nature of individual FasL molecules, now appears to reflect the need to cluster sufficient numbers of Fas receptors to achieve the critical density of Fas death domains required to form the minimal signaling-competent oligomeric platform[10][45]. The dodecameric 7:5 complex may represent one such minimal unit, though higher-order clustering likely generates additional complexes that further amplify signaling through positive feedback mechanisms. The proline-containing sequences in the Fas transmembrane domain create optimal packing geometry for trimerization, and the precise spatial organization of these transmembrane trimers appears essential for communicating the conformational changes induced by extracellular ligand binding down to the intracellular death domains, enabling the opening transitions necessary for FADD recruitment[8][10].
Differential Regulation of Apoptosis in Type I and Type II Cells: Molecular Mechanisms
The emerging understanding of how Type I and Type II cell classification relates to XIAP abundance and regulation provides a molecular explanation for the differential kinetics of Fas-induced apoptosis in various cell types[32][35]. Recent mechanistic studies indicate that the key decision point determining whether a cell exhibits Type I or Type II signaling relates to the ability of activated caspase-3 (itself a product of the initial caspase-8 activation) to trigger the degradation of XIAP protein[32]. In Type I cells such as thymocytes, the caspase-8 levels activated within the DISC prove sufficient to directly cleave and activate caspase-3 to concentrations that exceed XIAP inhibitory capacity; additionally, activated caspase-3 directly cleaves and destroys XIAP, creating a positive feedback loop that ensures complete apoptotic commitment[32]. In Type II cells including hepatocytes, the initial caspase-8 and caspase-3 activation is more modest, allowing XIAP to maintain inhibitory control; however, the same activated caspase-3 that is inhibited by XIAP in the initial phase somehow promotes XIAP upregulation through unknown post-translational mechanisms, perhaps involving altered ubiquitination or phosphorylation of XIAP or its regulatory proteins[32][35]. This elevated XIAP then effectively increases the apoptotic threshold for Type II cells, forcing them to engage the Bid-BaxBak-mitochondrial pathway amplification mechanism[32][35].
Remarkably, administration of SMAC mimetics, which antagonize IAP proteins and mimic Smac/DIABLO released from mitochondria, can reverse the resistance of Bid-deficient Type II cells to Fas-induced apoptosis, restoring death kinetics approaching those of wild-type cells[32]. This observation indicates that the Caspase-XIAP-Smac axis forms a central regulatory nexus in Fas-mediated apoptosis, with the relative abundances and activities of these molecules determining the cellular response[32]. The Type II pathway may represent an evolutionary strategy for tissues such as the liver that require heightened apoptotic thresholds to prevent inadvertent death from basal activation of death receptors and stress signals, while Type I cells such as lymphocytes represent cell types where rapid elimination upon activation or autoreactivity proves paramount to immune homeostasis.
Tissue-Specific Expression and Physiological Functions
Fas Expression Patterns in Embryonic Development and Adult Tissues
Comprehensive analysis of Fas and FasL expression during murine embryonic development revealed that Fas expression becomes detectable in distinct cell types of developing tissues from embryonic day 16.5 onwards, with particular prominence in developing sinusoid endothelium, thymic primordium, lung, and liver[15]. FasL expression during embryogenesis, in contrast, shows much more restricted distribution, primarily detected in submaxillary gland epithelial cells and the developing nervous system[15]. This spatial and temporal separation of Fas and FasL expression during development suggests that embryonic Fas/FasL interactions, while demonstrating significant expression in some tissues, may not be primarily responsible for the programmed cell death events characteristic of neural development, tissue remodeling, and organ formation during embryogenesis[15]. This finding contrasts with the substantial expression observed in adult tissues and suggests that the Fas system acquires greater importance in adult tissues than in embryonic development.
In adult tissues, remarkably wide expression of both Fas and FasL has been documented through sensitive RNase protection assays, with constitutive coexpression noted in thymus, lung, spleen, small intestine, large intestine, seminal vesicles, prostate, and uterus[15]. Tissues exhibiting the highest coexpression of Fas and FasL generally are characterized by rapid apoptotic cell turnover and active cell proliferation, consistent with a role for Fas-FasL interactions in regulating physiological cell turnover[15]. Tissues expressing FasL are notably enriched for immune-privileged sites including eye, brain, placenta, and testis, tissues that must actively protect their resident cells from immune attack through various mechanisms, with FasL expression on specialized cell types within these tissues triggering apoptosis of infiltrating lymphocytes bearing Fas receptor[15][18]. The cardiac expression of FasL, normally absent or minimal in wild-type hearts, becomes pathogenic when artificially enforced through transgenic expression, triggering a proinflammatory response characterized by mild leukocyte infiltration, interstitial fibrosis, and concentric cardiac hypertrophy, demonstrating that FasL expression in cardiac myocytes can actively drive pathological inflammation despite the coexpression of Fas in these cells[18].
Conclusion: Fas as a Central Regulatory Hub in Immune Homeostasis and Cellular Death
The Fas receptor emerges from decades of intensive investigation as far more than a simple death-inducing receptor, representing instead a sophisticated molecular platform integrating apoptotic and non-apoptotic signaling capable of flexibly responding to diverse cellular contexts and external signals. The structural advances achieved through cryo-EM analysis of the DISC complex have unveiled how massive clustering of pre-assembled Fas receptor trimers couples to formation of asymmetric oligomeric death domain platforms that nucleate caspase-8 activation through FADD-mediated DED filament assembly and polymerization. The recognition that Fas signaling outcome depends critically on ligand form (membrane-bound versus soluble), receptor oligomerization state, adaptor protein composition, and post-translational modifications has revealed a far more nuanced picture than the simple binary "live or die" model that dominated earlier understanding. The dramatic tissue-specific differences in apoptotic pathway utilization, exemplified by Type I and Type II cells, reflect elegant regulatory mechanisms wherein XIAP levels and regulation create different apoptotic thresholds appropriate for distinct tissue functions. The non-apoptotic functions of Fas—ranging from regulating mTOR-dependent B cell fate decisions to promoting immune cell migration and inflammation resolution—have emerged as equally critical to organismal biology, suggesting that the name "death receptor" substantially underestimates the biological complexity of this molecule.
The clinical importance of Fas dysfunction extends beyond the relatively rare ALPS syndrome to encompass the majority of human cancers, which frequently upregulate anti-apoptotic mechanisms countering Fas signaling, and to autoimmune diseases including systemic lupus erythematosus and various vasculitides, where soluble FasL and defective Fas signaling promote pathological inflammation[40][55]. The evolutionary history of human FasL, wherein a single amino acid change apparently selected for neural development created a vulnerability to plasmin-mediated inactivation in solid tumors, exemplifies how evolutionary trade-offs can create unexpected vulnerabilities to modern diseases. Future therapeutic strategies targeting Fas signaling must account for the tissue-specific roles of apoptotic versus non-apoptotic pathways, the impact of soluble versus membrane-bound ligand forms, and the complex regulation of adaptor proteins and post-translational modifications that determine signaling outcome. The convergence of structural biology, evolutionary analysis, and clinical investigation on the Fas system continues to yield insights into fundamental mechanisms of programmed cell death while revealing previously unsuspected roles in immune regulation, development, and disease pathogenesis.
Citations
- https://en.wikipedia.org/wiki/Fas_receptor
- https://www.ncbi.nlm.nih.gov/books/NBK6345/
- https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2020.00314/full
- https://www.ncbi.nlm.nih.gov/gene/355
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2956119/
- https://pubmed.ncbi.nlm.nih.gov/25153377/
- https://pubmed.ncbi.nlm.nih.gov/20935634/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4761300/
- https://pubmed.ncbi.nlm.nih.gov/11525634/
- https://www.pnas.org/doi/10.1073/pnas.2504819122
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2788920/
- https://www.frontiersin.org/journals/molecular-neuroscience/articles/10.3389/fnmol.2018.00101/full
- https://pubmed.ncbi.nlm.nih.gov/17449269/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2120795/
- https://geneglobe.qiagen.com/us/knowledge/gene/ENSG00000026103
- https://www.science.org/doi/10.1126/science.288.5475.2328
- https://www.jci.org/articles/view/8212
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2575811/
- https://www.science.org/doi/10.1126/sciimmunol.adj5948
- https://primaryimmune.org/understanding-primary-immunodeficiency/types-of-pi/autoimmune-lymphoproliferative-syndrome-alps
- https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2017.00237/full
- https://pubmed.ncbi.nlm.nih.gov/12485619/
- https://ashpublications.org/blood/article/123/13/1989/32605/Natural-history-of-autoimmune-lymphoproliferative
- https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2021.631274/full
- https://rupress.org/jcb/article/218/6/1776/61850/Post-translational-regulation-of-ubiquitin
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3470429/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2626565/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8946176/
- https://med.stanford.edu/content/dam/sm/shared/community/department-of-medicine/Immuno/JournalClub/19-20/JC-week-11_Hunter.pdf
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7112278/
- https://academic.oup.com/jmcb/article/2/2/63/894704
- https://elifesciences.org/articles/80953
- https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2022.1074099/full
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3252833/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7772363/
- https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2018.02521/full
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2997140/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5623854/
- https://ashpublications.org/bloodict/article-split/1/3/100010/547286/Lipid-nanoparticle-Fas-gene-therapy-suppresses
- https://ashpublications.org/blood/article-abstract/107/6/2384/22094/Fas-ligand-is-localized-to-membrane-rafts-where-it
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3964250/
- https://www.sciencedaily.com/releases/2025/07/250702214136.htm
- https://journals.biologists.com/jcs/article/129/1/108/138738/Evolutionarily-conserved-primary-TNF-sequences
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3725553/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC12432253/
- https://www.jci.org/articles/view/42686
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11049660/
- https://haematologica.org/article/view/4668
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3228663/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1523423/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC358392/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2933433/
- https://www.thermofisher.com/us/en/home/life-science/antibodies/antibodies-learning-center/antibodies-resource-library/cell-signaling-pathways/fas-signaling-pathway.html
- https://febs.onlinelibrary.wiley.com/doi/10.1111/febs.14292
- https://journals.plos.org/plosone/article?id=10.1371%2Fjournal.pone.0031258