Deep Research
Falcon
(HTT-deep-research-falcon.md)
Falcon
(HTT-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Human HTT (Huntingtin; UniProt P42858) — Functional Annotation, Mechanisms, Localization, and Translational Context
0) Target verification (required disambiguation)
The target gene symbol HTT in this report refers specifically to the human huntingtin gene/protein (Homo sapiens), located on chromosome 4p16.3, encoding a 3,144–amino-acid (~348 kDa) protein whose defining molecular feature is an N‑terminal CAG-repeat–encoded polyglutamine (polyQ) tract in exon 1 (pathogenic expansion in Huntington’s disease). These identifiers (genomic locus, size, exon‑1 polyQ, N17 region, HEAT/ARM-like repeat architecture) distinguish HTT/huntingtin from other similarly named proteins or orthologs. (bates2015huntingtondisease pages 4-7, kaushal2026genetherapyfor pages 1-2)
OpenTargets also links HTT (ENSG00000197386) to Huntington disease (MONDO_0007739) with curated evidence, consistent with the canonical human target identity. (OpenTargets Search: Huntington disease-HTT)
1) Key concepts and definitions (current understanding)
1.1 What kind of protein is huntingtin?
Huntingtin (HTT) is best characterized as a large, multi-domain, non-enzymatic scaffold/adaptor protein. Structurally it contains multiple HEAT/ARM-like repeat domains and a prominent N-terminal region containing an N17 amphipathic helix, a polyQ tract, and a proline-rich region; functionally, it interacts with many partners and coordinates intracellular trafficking and homeostatic pathways rather than catalyzing a specific chemical reaction. (tong2024huntington’sdiseasecomplex pages 1-2, krzystek2025navigatingtheneuronal pages 6-9, bates2015huntingtondisease pages 4-7)
1.2 PolyQ expansion and disease vs normal alleles
HTT’s exon-1 CAG repeat is polymorphic in humans. Normal alleles are typically 6–35 CAG repeats; ≥36 is disease-associated (with 36–39 reduced penetrance and ≥40 highly penetrant in many clinical contexts). (bates2015huntingtondisease pages 4-7, vauleon2023quantifyingmutanthuntingtin pages 1-2)
2) Functional roles, mechanisms, and subcellular localization (annotation-oriented)
2.1 Selective autophagy/lysosomal targeting via a defined ubiquitin-binding domain (2024 primary evidence)
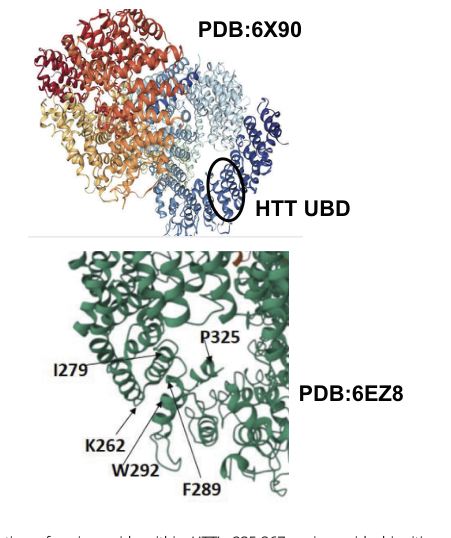
A major recent advance is direct mechanistic evidence that human HTT includes a ubiquitin-binding domain (UBD) localized to residues ~235–367 and that HTT regulates lysosomal targeting of specific cargo, including mitochondrial proteins and RNA-binding proteins (RBPs). Using CRISPR HTT knockout and lysosome immunoprecipitation (LysoIP) proteomics, HTT loss altered lysosomal cargo profiles, consistent with a role in selective autophagy/lysosomal delivery; experimentally, HTT UBD binding to ubiquitin provides a plausible physical mechanism for engaging ubiquitinated/ubiquitin-associated cargo. (fote2024huntingtincontainsan pages 2-3, fote2024huntingtincontainsan pages 1-2, fote2024huntingtincontainsan pages 3-4)
Visual support for these claims (UBD placement on HTT structure and KO/LysoIP results) is provided by figure panels retrieved from the 2024 PNAS study. (fote2024huntingtincontainsan media 6f73f1b2, fote2024huntingtincontainsan media f08bfae2, fote2024huntingtincontainsan media 42126a8d, fote2024huntingtincontainsan media 464dbd70, fote2024huntingtincontainsan media 5cfec8a3)
2.2 Vesicular transport and motor coordination (dynein/kinesin balance)
A core concept in HTT biology is that it acts as a scaffold that recruits or coordinates motor/adaptor proteins on cargoes to support bidirectional axonal transport. A key regulatory axis is HTT serine 421 phosphorylation, where dephosphorylation (e.g., by calcineurin) is linked to enhanced dynein-mediated retrograde transport, whereas phosphorylation (e.g., by Akt/S6K) supports kinesin-1 recruitment and anterograde movement. (fote2024huntingtincontainsan pages 2-3)
Neuronal autophagy literature further connects HTT with HAP1, dynein/dynactin, and kinesin-1, and with endolysosomal/autophagosomal RAB compartments involved in long-range transport. (krzystek2025navigatingtheneuronal pages 17-20)
2.3 Autophagosome maturation and axonal retrograde trafficking (neuronal specialization)
In neurons, HTT is repeatedly implicated in coordinating autophagy-related trafficking, including complexes involving STX17 and RAB7 on axonal endolysosomes and processes required for competent retrograde transport and fusion behavior. Mutant HTT (mHTT) is described as perturbing these processes by disrupting fusion/transport interactions (e.g., involving STX17, RILP–dynactin, OPTN–RAB8 complexes) in review syntheses of experimental work. (krzystek2025navigatingtheneuronal pages 17-20, krzystek2025navigatingtheneuronal pages 9-12)
2.4 Membrane targeting, lipid binding, and nuclear–cytoplasmic distribution
HTT’s N17 amphipathic helix is highlighted as a membrane-targeting element; perturbation of this N-terminal helix reduces membrane localization and increases nuclear accumulation, linking a structural feature to subcellular distribution. (krzystek2025navigatingtheneuronal pages 6-9)
A disease-focused review synthesis reports that HTT contains nuclear localization signals (NLS) and C-terminal nuclear export signals (NES), and that loss of these elements in N-terminal mutant fragments can favor abnormal nuclear localization via nuclear pore interactions. (tong2024huntington’sdiseasecomplex pages 1-2)
3) Recent developments and latest research (prioritizing 2023–2024)
3.1 2024: HTT ubiquitin-binding domain and lysosomal cargo routing
The 2024 PNAS study is a notable mechanistic development because it anchors HTT’s long-suspected scaffold role in selective autophagy/lysosomal function to a defined UBD (235–367) and to measurable cargo-routing phenotypes in HTT KO cells. (fote2024huntingtincontainsan pages 2-3, fote2024huntingtincontainsan pages 1-2)
3.2 2023: Clinical trial re-design and biomarker instrumentation
As HTT-lowering trials increasingly use CSF mutant huntingtin (mHTT) as a pharmacodynamic biomarker, assay development and validation have become central. A 2023 report describes validation of an assay intended to quantify CSF mHTT to support registrational trials, emphasizing polyQ-length–dependent signal behavior and cross-lab validation. (vauleon2023quantifyingmutanthuntingtin pages 1-2)
4) Current applications and real-world implementations
4.1 Genetics-based diagnosis and disease stratification
Huntington’s disease is genetically defined by expanded CAG repeats in HTT, and allele length categories (intermediate, reduced penetrance, highly penetrant) are used in counseling and risk stratification. (bates2015huntingtondisease pages 4-7, vauleon2023quantifyingmutanthuntingtin pages 1-2)
4.2 HTT-lowering therapeutics: modalities in real-world use/testing
Current disease-modifying strategies prominently include HTT lowering (non-allele-selective and allele-selective), implemented via:
- Intrathecal antisense oligonucleotides (ASOs) (e.g., tominersen; WVE-003). (NCT05686551 chunk 1, farag2024huntington’sdiseaseclinical pages 1-5)
- Oral small-molecule splice modulators that lower HTT in periphery and CSF (e.g., PTC518). (farag2024huntington’sdiseaseclinical pages 1-5, NCT05358717 chunk 1)
- Neurosurgically delivered AAV-based microRNA gene therapy targeting HTT (e.g., AMT-130), with imaging-based anatomical eligibility and surgical delivery constraints. (NCT04120493 chunk 2)
Example registry-level implementation details (ClinicalTrials.gov)
GENERATION HD2 (tominersen; NCT05686551; ClinicalTrials.gov)
- Phase: 2; design: randomized, double-blind, placebo-controlled (QUADRUPLE masking). (NCT05686551 chunk 1)
- Route: intrathecal; dosing: 60 mg Q16W or 100 mg Q16W vs placebo. (NCT05686551 chunk 1)
- Enrollment: 301 actual (registry); status: Active, not recruiting; start 2023-02-03; primary completion 2026-05-14. (NCT05686551 chunk 1)
PIVOT-HD (PTC518; NCT05358717; ClinicalTrials.gov)
- Phase: 2; route: oral tablets once daily; dosing arms included 5, 10, 20 mg vs placebo. (NCT05358717 chunk 1)
- Enrollment: 159; status: Completed; primary outcomes include TEAEs and % change in blood total HTT at Month 3. (NCT05358717 chunk 1)
5) Expert synthesis and authoritative opinions (consensus themes)
5.1 HTT as a multifunctional scaffold; loss-of-function vs toxic gain-of-function
Authoritative reviews converge on huntingtin as a multifunctional scaffold that participates in multiple cellular processes (transport, transcriptional regulation, proteostasis/autophagy, etc.), and emphasize that Huntington’s disease mechanisms likely include both toxic gain-of-function by mutant HTT and loss or perturbation of normal HTT functions. (tong2024huntington’sdiseasecomplex pages 1-2, fote2024huntingtincontainsan pages 2-3)
5.2 Safety and interpretation challenges in HTT-lowering
Clinical-trial experts emphasize that HTT lowering must balance target engagement with safety monitoring (notably CSF neurofilament light, NfL, as a marker of neuroaxonal injury) and interpretability (e.g., confounding inflammation or procedure-related effects). Trial updates highlight earlier safety signals (ventricular volume changes; neuropathy in some programs) and the need for careful biomarker packages alongside clinical endpoints. (estevezfraga2024huntington’sdiseaseclinical pages 2-4, farag2024huntington’sdiseaseclinical pages 1-5, estevezfraga2023huntington’sdiseaseclinical pages 1-3)
6) Relevant statistics and quantitative data (recent and authoritative)
6.1 Epidemiology (prevalence)
An authoritative primer reports HD prevalence of ~17.2 per 100,000 among those of European descent versus ~2.1 per 100,000 in the ethnically diverse remainder, noting founder populations with higher prevalence. (bates2015huntingtondisease pages 4-7)
6.2 Genetics and protein size
- Normal CAG range: 6–35; disease association begins at ≥36 with reduced penetrance for 36–39 and high penetrance for ≥40. (bates2015huntingtondisease pages 4-7, vauleon2023quantifyingmutanthuntingtin pages 1-2)
- Protein size: 3,144 aa (~348 kDa). (bates2015huntingtondisease pages 4-7)
6.3 Biomarkers decades before onset (2025 Nature Medicine; mechanistic relevance for HTT)
In a young-adult HD gene-expanded cohort studied ~23 years before predicted motor diagnosis, clinical measures did not significantly decline over 4.5 years, but CSF biomarkers already showed early neurodegeneration (elevated NfL, reduced PENK) with significant caudate/putamen atrophy signals; blood somatic CAG expansion metrics predicted subsequent striatal atrophy. (scahill2025somaticcagrepeat pages 1-2)
6.4 Quantitative outcomes from recent HTT-lowering trial updates (2024)
A 2024 clinical trials update reports:
- PTC518: mHTT decreased 22% and 43% in blood and 21% and 43% in CSF for 5 mg and 10 mg doses; motor score trends favored treatment vs placebo and no CSF NfL increase was reported in that update. (farag2024huntington’sdiseaseclinical pages 1-5)
- WVE-003 (SELECT-HD): mean CSF mHTT reduction 46% vs placebo at 24 weeks and 44% at 28 weeks, with broadly preserved wild-type HTT and no serious adverse events; however, NfL dynamics and interpretation remained an important concern. (farag2024huntington’sdiseaseclinical pages 1-5, farag2024huntington’sdiseaseclinical pages 5-9)
- Branaplam (VIBRANT-HD): CSF mHTT reduction up to 26.6% at 17 weeks, but accompanied by safety signals including serum NfL increases and ventricular volume increases up to 9.5% vs 1.6% on placebo, plus frequent peripheral neuropathy signs/symptoms. (estevezfraga2024huntington’sdiseaseclinical pages 2-4)
7) Practical functional-annotation summary table
The following table consolidates function, mechanisms, localization, and evidence types for annotation purposes.
| Functional role | Key molecular mechanisms | Subcellular localization | Evidence type / key recent citations |
|---|---|---|---|
| Selective autophagy / lysosomal targeting | HTT acts as a scaffold for selective autophagy and lysosomal cargo delivery; a ubiquitin-binding domain (UBD) at residues 235–367 binds ubiquitin/ubiquitinated cargo; HTT interacts with ULK1, p62/SQSTM1, and LC3; HTT knockout alters lysosomal cargo composition, reducing lysosomal targeting of mitochondrial proteins and altering RNA-binding protein delivery | Cytoplasm; autophagosomes; lysosomes/endolysosomes; mitochondria-associated degradative pathway | Primary experimental evidence (biochemistry, CRISPR KO, LysoIP proteomics, imaging) and recent review synthesis (fote2024huntingtincontainsan pages 2-3, fote2024huntingtincontainsan pages 1-2, fote2024huntingtincontainsan pages 3-4, krzystek2025navigatingtheneuronal pages 9-12) |
| Vesicle / motor transport | HTT is a multifunctional scaffold for bidirectional vesicle transport; S421 phosphorylation biases motor engagement, with dephosphorylation favoring dynein-mediated retrograde transport and phosphorylation favoring kinesin-1 recruitment/anterograde transport; key partners include HAP1, kinesin-1, dynein/dynactin; HTT also associates with RAB-positive vesicles and endosomal machinery | Axons and neurites; cytoplasmic vesicles; endosomes; BDNF-containing transport vesicles | Primary experimental evidence and mechanistic review support (fote2024huntingtincontainsan pages 2-3, fote2024huntingtincontainsan pages 1-2, krzystek2025navigatingtheneuronal pages 17-20, turkalj2023mutanthuntingtinimpairs pages 16-21) |
| Autophagosome maturation / retrograde axonal trafficking | HTT cooperates with HAP1 and transport machinery to support retrograde autophagosome movement; HTT and STX17 co-localize on RAB7+ endolysosomes; STX17–RAB7 fusion state is linked to retrograde transport competence; mutant HTT perturbs STX17-mediated fusion and RILP–dynactin interactions | Distal axon; autophagosomes; RAB7+ endolysosomes; lysosome fusion compartments | Review integrating neuronal experimental literature (krzystek2025navigatingtheneuronal pages 17-20, krzystek2025navigatingtheneuronal pages 9-12) |
| Nuclear–cytoplasmic shuttling / transcription-related scaffolding | HTT contains NLS and C-terminal NES elements; the N17 N-terminal amphipathic helix promotes membrane association and limits nuclear accumulation; deletion/disruption of N17 increases nuclear localization; HTT interacts with transcription factors/co-regulators, supporting a scaffold role in transcriptional regulation | Nucleus; cytoplasm; membrane-associated compartments | Structural/functional review evidence and disease-focused review synthesis (krzystek2025navigatingtheneuronal pages 6-9, tong2024huntington’sdiseasecomplex pages 1-2, bates2015huntingtondisease pages 4-7) |
| Membrane targeting / lipid sensing | N17 forms an amphipathic α-helix important for membrane targeting; HTT shows preferential binding to phosphoinositides including PI3,5P2 and PI4,5P2; palmitoylation at C214 (via HIP14/ZDHHC17 and HIP14L/ZDHHC13) modulates membrane association and trafficking behavior | Plasma membrane; endolysosomal/autophagic membranes; ER/Golgi-associated membranes | Review synthesis grounded in prior structural/cell-biological studies (krzystek2025navigatingtheneuronal pages 6-9, krzystek2025navigatingtheneuronal pages 9-12) |
| Cytoskeleton / actin organization | Beyond microtubule-based transport, HTT directly organizes actin: the N-HEAT and Bridge domains wrap around F-actin, and HTT dimerization can bridge parallel actin filaments; this supports growth-cone morphology and cytoskeletal organization | F-actin networks; axonal growth cones; neuronal cytoskeleton | Recent structural primary study (2025) extending HTT functional annotation beyond trafficking (OpenTargets Search: Huntington disease-HTT) |
| Ciliogenesis / mitotic spindle / cell polarity (brief) | HTT has established roles in ciliogenesis, mitotic spindle positioning, and cell polarity/developmental organization; these functions are frequently interpreted as consistent with its broader scaffold/adaptor role, though mechanistic resolution is less complete than for transport/autophagy | Centrosome / ciliary base; mitotic apparatus; developing neural cells | Disease/developmental review and organoid-model evidence summaries (piao2025advancesingene pages 2-3, tong2024huntington’sdiseasecomplex pages 1-2) |
| Defining structural features for annotation | Large ~3144 aa (~348 kDa) HEAT/ARM-like scaffold protein with N-terminal polyQ tract, N17 amphipathic helix, polyproline-rich region, and resolved HTT–HAP40 architecture; these features support classification as a non-enzymatic scaffold/adaptor rather than enzyme/transporter | Broadly cytoplasmic with dynamic membrane and nuclear association | Authoritative review and structural evidence (bates2015huntingtondisease pages 4-7, tong2024huntington’sdiseasecomplex pages 1-2) |
Table: This table summarizes the main experimentally supported functions, mechanisms, and localizations of human HTT/huntingtin for annotation purposes. It prioritizes recent 2023–2024 evidence where available and highlights the scaffold/adaptor nature of HTT across autophagy, trafficking, and compartmental regulation.
8) URLs and publication dates (selected high-priority sources)
- Fote GM et al. “Huntingtin contains an ubiquitin-binding domain and regulates lysosomal targeting of mitochondrial and RNA-binding proteins.” PNAS. 2024-07. https://doi.org/10.1073/pnas.2319091121 (fote2024huntingtincontainsan pages 1-2)
- Farag M, Tabrizi SJ, Wild EJ. “Huntington’s Disease Clinical Trials Update: September 2024.” Journal of Huntington’s Disease. 2024-10. https://doi.org/10.1177/18796397241293955 (farag2024huntington’sdiseaseclinical pages 1-5)
- Tong H et al. “Huntington’s Disease: Complex Pathogenesis and Therapeutic Strategies.” IJMS. 2024-03. https://doi.org/10.3390/ijms25073845 (tong2024huntington’sdiseasecomplex pages 1-2)
- Vauleon S et al. “Quantifying mutant huntingtin protein in human cerebrospinal fluid…” Scientific Reports. 2023-04. https://doi.org/10.1038/s41598-023-32630-4 (vauleon2023quantifyingmutanthuntingtin pages 1-2)
- ClinicalTrials.gov: NCT05686551 (GENERATION HD2; tominersen). Registry record (start 2023-02-03). https://clinicaltrials.gov/study/NCT05686551 (NCT05686551 chunk 1)
- ClinicalTrials.gov: NCT05358717 (PTC518). Registry record (first posted 2022). https://clinicaltrials.gov/study/NCT05358717 (NCT05358717 chunk 1)
9) Notes on scope and evidence strength
HTT is not an enzyme or transporter; its “primary function” is best captured as a scaffold/adaptor coordinating cargo recognition (including ubiquitin-associated cargo), vesicular trafficking, and autophagy–lysosome pathway dynamics, with compartmental regulation via membrane targeting and nuclear–cytoplasmic shuttling. The strongest recent mechanistic evidence within the retrieved corpus is for UBD-mediated ubiquitin engagement and lysosomal targeting phenotypes (2024 PNAS) and for quantitative target engagement/safety monitoring in HTT-lowering trials (2024 trial updates + ClinicalTrials.gov). (fote2024huntingtincontainsan pages 2-3, farag2024huntington’sdiseaseclinical pages 1-5, NCT05686551 chunk 1)
References
-
(bates2015huntingtondisease pages 4-7): Gillian P. Bates, Ray Dorsey, James F. Gusella, Michael R. Hayden, Chris Kay, Blair R. Leavitt, Martha Nance, Christopher A. Ross, Rachael I. Scahill, Ronald Wetzel, Edward J. Wild, and Sarah J. Tabrizi. Huntington disease. Nature Reviews Disease Primers, Apr 2026. URL: https://doi.org/10.1038/nrdp.2015.5, doi:10.1038/nrdp.2015.5. This article has 1456 citations.
-
(kaushal2026genetherapyfor pages 1-2): Riya Kaushal, Mohit Yadav, Sourabh Kosey, and Madhaw Dwivedi. Gene therapy for huntington’s disease: advances, challenges, and future perspectives. Neurogenetics, Feb 2026. URL: https://doi.org/10.1007/s10048-026-00887-2, doi:10.1007/s10048-026-00887-2. This article has 0 citations and is from a peer-reviewed journal.
-
(OpenTargets Search: Huntington disease-HTT): Open Targets Query (Huntington disease-HTT, 8 results). Buniello, A. et al. (2025). Open Targets Platform: facilitating therapeutic hypotheses building in drug discovery. Nucleic Acids Research.
-
(tong2024huntington’sdiseasecomplex pages 1-2): Huichun Tong, Tianqi Yang, Shuying Xu, Xinhui Li, Li Liu, Gongke Zhou, Sitong Yang, Shurui Yin, Xiao-Jiang Li, and Shihua Li. Huntington’s disease: complex pathogenesis and therapeutic strategies. International Journal of Molecular Sciences, 25:3845, Mar 2024. URL: https://doi.org/10.3390/ijms25073845, doi:10.3390/ijms25073845. This article has 126 citations.
-
(krzystek2025navigatingtheneuronal pages 6-9): Thomas J. Krzystek and Shermali Gunawardena. Navigating the neuronal recycling bin: another look at huntingtin in coordinating autophagy. Autophagy Reports, Jun 2025. URL: https://doi.org/10.1080/27694127.2025.2472450, doi:10.1080/27694127.2025.2472450. This article has 0 citations.
-
(vauleon2023quantifyingmutanthuntingtin pages 1-2): Stephanie Vauleon, Katharina Schutz, Benoit Massonnet, Nanda Gruben, Marianne Manchester, Alessandra Buehler, Eginhard Schick, Lauren Boak, and David J. Hawellek. Quantifying mutant huntingtin protein in human cerebrospinal fluid to support the development of huntingtin-lowering therapies. Scientific Reports, Apr 2023. URL: https://doi.org/10.1038/s41598-023-32630-4, doi:10.1038/s41598-023-32630-4. This article has 12 citations and is from a peer-reviewed journal.
-
(fote2024huntingtincontainsan pages 2-3): Gianna M. Fote, Vinay V. Eapen, Ryan G. Lim, Clinton Yu, Lisa Salazar, Nicolette R. McClure, Jharrayne McKnight, Thai B. Nguyen, Marie C. Heath, Alice L. Lau, Mark A. Villamil, Ricardo Miramontes, Ian H. Kratter, Steven Finkbeiner, Jack C. Reidling, Joao A. Paulo, Peter Kaiser, Lan Huang, David E. Housman, Leslie M. Thompson, and Joan S. Steffan. Huntingtin contains an ubiquitin-binding domain and regulates lysosomal targeting of mitochondrial and rna-binding proteins. Proceedings of the National Academy of Sciences of the United States of America, Jul 2024. URL: https://doi.org/10.1073/pnas.2319091121, doi:10.1073/pnas.2319091121. This article has 8 citations and is from a highest quality peer-reviewed journal.
-
(fote2024huntingtincontainsan pages 1-2): Gianna M. Fote, Vinay V. Eapen, Ryan G. Lim, Clinton Yu, Lisa Salazar, Nicolette R. McClure, Jharrayne McKnight, Thai B. Nguyen, Marie C. Heath, Alice L. Lau, Mark A. Villamil, Ricardo Miramontes, Ian H. Kratter, Steven Finkbeiner, Jack C. Reidling, Joao A. Paulo, Peter Kaiser, Lan Huang, David E. Housman, Leslie M. Thompson, and Joan S. Steffan. Huntingtin contains an ubiquitin-binding domain and regulates lysosomal targeting of mitochondrial and rna-binding proteins. Proceedings of the National Academy of Sciences of the United States of America, Jul 2024. URL: https://doi.org/10.1073/pnas.2319091121, doi:10.1073/pnas.2319091121. This article has 8 citations and is from a highest quality peer-reviewed journal.
-
(fote2024huntingtincontainsan pages 3-4): Gianna M. Fote, Vinay V. Eapen, Ryan G. Lim, Clinton Yu, Lisa Salazar, Nicolette R. McClure, Jharrayne McKnight, Thai B. Nguyen, Marie C. Heath, Alice L. Lau, Mark A. Villamil, Ricardo Miramontes, Ian H. Kratter, Steven Finkbeiner, Jack C. Reidling, Joao A. Paulo, Peter Kaiser, Lan Huang, David E. Housman, Leslie M. Thompson, and Joan S. Steffan. Huntingtin contains an ubiquitin-binding domain and regulates lysosomal targeting of mitochondrial and rna-binding proteins. Proceedings of the National Academy of Sciences of the United States of America, Jul 2024. URL: https://doi.org/10.1073/pnas.2319091121, doi:10.1073/pnas.2319091121. This article has 8 citations and is from a highest quality peer-reviewed journal.
-
(fote2024huntingtincontainsan media 6f73f1b2): Gianna M. Fote, Vinay V. Eapen, Ryan G. Lim, Clinton Yu, Lisa Salazar, Nicolette R. McClure, Jharrayne McKnight, Thai B. Nguyen, Marie C. Heath, Alice L. Lau, Mark A. Villamil, Ricardo Miramontes, Ian H. Kratter, Steven Finkbeiner, Jack C. Reidling, Joao A. Paulo, Peter Kaiser, Lan Huang, David E. Housman, Leslie M. Thompson, and Joan S. Steffan. Huntingtin contains an ubiquitin-binding domain and regulates lysosomal targeting of mitochondrial and rna-binding proteins. Proceedings of the National Academy of Sciences of the United States of America, Jul 2024. URL: https://doi.org/10.1073/pnas.2319091121, doi:10.1073/pnas.2319091121. This article has 8 citations and is from a highest quality peer-reviewed journal.
-
(fote2024huntingtincontainsan media f08bfae2): Gianna M. Fote, Vinay V. Eapen, Ryan G. Lim, Clinton Yu, Lisa Salazar, Nicolette R. McClure, Jharrayne McKnight, Thai B. Nguyen, Marie C. Heath, Alice L. Lau, Mark A. Villamil, Ricardo Miramontes, Ian H. Kratter, Steven Finkbeiner, Jack C. Reidling, Joao A. Paulo, Peter Kaiser, Lan Huang, David E. Housman, Leslie M. Thompson, and Joan S. Steffan. Huntingtin contains an ubiquitin-binding domain and regulates lysosomal targeting of mitochondrial and rna-binding proteins. Proceedings of the National Academy of Sciences of the United States of America, Jul 2024. URL: https://doi.org/10.1073/pnas.2319091121, doi:10.1073/pnas.2319091121. This article has 8 citations and is from a highest quality peer-reviewed journal.
-
(fote2024huntingtincontainsan media 42126a8d): Gianna M. Fote, Vinay V. Eapen, Ryan G. Lim, Clinton Yu, Lisa Salazar, Nicolette R. McClure, Jharrayne McKnight, Thai B. Nguyen, Marie C. Heath, Alice L. Lau, Mark A. Villamil, Ricardo Miramontes, Ian H. Kratter, Steven Finkbeiner, Jack C. Reidling, Joao A. Paulo, Peter Kaiser, Lan Huang, David E. Housman, Leslie M. Thompson, and Joan S. Steffan. Huntingtin contains an ubiquitin-binding domain and regulates lysosomal targeting of mitochondrial and rna-binding proteins. Proceedings of the National Academy of Sciences of the United States of America, Jul 2024. URL: https://doi.org/10.1073/pnas.2319091121, doi:10.1073/pnas.2319091121. This article has 8 citations and is from a highest quality peer-reviewed journal.
-
(fote2024huntingtincontainsan media 464dbd70): Gianna M. Fote, Vinay V. Eapen, Ryan G. Lim, Clinton Yu, Lisa Salazar, Nicolette R. McClure, Jharrayne McKnight, Thai B. Nguyen, Marie C. Heath, Alice L. Lau, Mark A. Villamil, Ricardo Miramontes, Ian H. Kratter, Steven Finkbeiner, Jack C. Reidling, Joao A. Paulo, Peter Kaiser, Lan Huang, David E. Housman, Leslie M. Thompson, and Joan S. Steffan. Huntingtin contains an ubiquitin-binding domain and regulates lysosomal targeting of mitochondrial and rna-binding proteins. Proceedings of the National Academy of Sciences of the United States of America, Jul 2024. URL: https://doi.org/10.1073/pnas.2319091121, doi:10.1073/pnas.2319091121. This article has 8 citations and is from a highest quality peer-reviewed journal.
-
(fote2024huntingtincontainsan media 5cfec8a3): Gianna M. Fote, Vinay V. Eapen, Ryan G. Lim, Clinton Yu, Lisa Salazar, Nicolette R. McClure, Jharrayne McKnight, Thai B. Nguyen, Marie C. Heath, Alice L. Lau, Mark A. Villamil, Ricardo Miramontes, Ian H. Kratter, Steven Finkbeiner, Jack C. Reidling, Joao A. Paulo, Peter Kaiser, Lan Huang, David E. Housman, Leslie M. Thompson, and Joan S. Steffan. Huntingtin contains an ubiquitin-binding domain and regulates lysosomal targeting of mitochondrial and rna-binding proteins. Proceedings of the National Academy of Sciences of the United States of America, Jul 2024. URL: https://doi.org/10.1073/pnas.2319091121, doi:10.1073/pnas.2319091121. This article has 8 citations and is from a highest quality peer-reviewed journal.
-
(krzystek2025navigatingtheneuronal pages 17-20): Thomas J. Krzystek and Shermali Gunawardena. Navigating the neuronal recycling bin: another look at huntingtin in coordinating autophagy. Autophagy Reports, Jun 2025. URL: https://doi.org/10.1080/27694127.2025.2472450, doi:10.1080/27694127.2025.2472450. This article has 0 citations.
-
(krzystek2025navigatingtheneuronal pages 9-12): Thomas J. Krzystek and Shermali Gunawardena. Navigating the neuronal recycling bin: another look at huntingtin in coordinating autophagy. Autophagy Reports, Jun 2025. URL: https://doi.org/10.1080/27694127.2025.2472450, doi:10.1080/27694127.2025.2472450. This article has 0 citations.
-
(NCT05686551 chunk 1): GENERATION HD2. A Study to Evaluate the Safety, Biomarkers, and Efficacy of Tominersen Compared With Placebo in Participants With Prodromal and Early Manifest Huntington's Disease. Hoffmann-La Roche. 2023. ClinicalTrials.gov Identifier: NCT05686551
-
(farag2024huntington’sdiseaseclinical pages 1-5): Mena Farag, Sarah J Tabrizi, and Edward J Wild. Huntington’s disease clinical trials update: september 2024. Journal of Huntington's Disease, 13:409-418, Oct 2024. URL: https://doi.org/10.1177/18796397241293955, doi:10.1177/18796397241293955. This article has 15 citations and is from a peer-reviewed journal.
-
(NCT05358717 chunk 1): A Study to Evaluate the Safety and Efficacy of PTC518 in Participants With Huntington's Disease (HD). PTC Therapeutics. 2022. ClinicalTrials.gov Identifier: NCT05358717
-
(NCT04120493 chunk 2): Safety and Proof-of-Concept (POC) Study With AMT-130 in Adults With Early Manifest Huntington's Disease. UniQure Biopharma B.V.. 2019. ClinicalTrials.gov Identifier: NCT04120493
-
(estevezfraga2024huntington’sdiseaseclinical pages 2-4): Carlos Estevez-Fraga, Sarah J. Tabrizi, and Edward J. Wild. Huntington’s disease clinical trials corner: march 2024. Journal of Huntington's Disease, 13:1-14, Mar 2024. URL: https://doi.org/10.3233/jhd-240017, doi:10.3233/jhd-240017. This article has 37 citations and is from a peer-reviewed journal.
-
(estevezfraga2023huntington’sdiseaseclinical pages 1-3): Carlos Estevez-Fraga, Sarah J. Tabrizi, and Edward J. Wild. Huntington’s disease clinical trials corner: august 2023. Journal of Huntington's Disease, 12:169-185, Jul 2023. URL: https://doi.org/10.3233/jhd-239001, doi:10.3233/jhd-239001. This article has 43 citations and is from a peer-reviewed journal.
-
(scahill2025somaticcagrepeat pages 1-2): Rachael I. Scahill, Mena Farag, Michael J. Murphy, Nicola Z. Hobbs, Michela Leocadi, Christelle Langley, Harry Knights, Marc Ciosi, Kate Fayer, Mitsuko Nakajima, Olivia Thackeray, Johan Gobom, John Rönnholm, Sophia Weiner, Yara R. Hassan, Nehaa K. P. Ponraj, Carlos Estevez-Fraga, Christopher S. Parker, Ian B. Malone, Harpreet Hyare, Jeffrey D. Long, Amanda Heslegrave, Cristina Sampaio, Hui Zhang, Trevor W. Robbins, Henrik Zetterberg, Edward J. Wild, Geraint Rees, James B. Rowe, Barbara J. Sahakian, Darren G. Monckton, Douglas R. Langbehn, and Sarah J. Tabrizi. Somatic cag repeat expansion in blood associates with biomarkers of neurodegeneration in huntington’s disease decades before clinical motor diagnosis. Nature Medicine, 31:807-818, Jan 2025. URL: https://doi.org/10.1038/s41591-024-03424-6, doi:10.1038/s41591-024-03424-6. This article has 59 citations and is from a highest quality peer-reviewed journal.
-
(farag2024huntington’sdiseaseclinical pages 5-9): Mena Farag, Sarah J Tabrizi, and Edward J Wild. Huntington’s disease clinical trials update: september 2024. Journal of Huntington's Disease, 13:409-418, Oct 2024. URL: https://doi.org/10.1177/18796397241293955, doi:10.1177/18796397241293955. This article has 15 citations and is from a peer-reviewed journal.
-
(turkalj2023mutanthuntingtinimpairs pages 16-21): BA Turkalj. Mutant huntingtin impairs the recruitment and activation of motor proteins to bdnf-endosomes. Unknown journal, 2023.
-
(piao2025advancesingene pages 2-3): Xuejiao Piao, Dan Li, Hui Liu, Qing Guo, and Yang Yu. Advances in gene and cellular therapeutic approaches for huntington’s disease. Protein & Cell, 16:307-337, Aug 2025. URL: https://doi.org/10.1093/procel/pwae042, doi:10.1093/procel/pwae042. This article has 20 citations and is from a peer-reviewed journal.
Artifacts
Citations
- fote2024huntingtincontainsan pages 2-3
- krzystek2025navigatingtheneuronal pages 17-20
- krzystek2025navigatingtheneuronal pages 6-9
- vauleon2023quantifyingmutanthuntingtin pages 1-2
- bates2015huntingtondisease pages 4-7
- scahill2025somaticcagrepeat pages 1-2
- fote2024huntingtincontainsan pages 1-2
- kaushal2026genetherapyfor pages 1-2
- fote2024huntingtincontainsan pages 3-4
- krzystek2025navigatingtheneuronal pages 9-12
- turkalj2023mutanthuntingtinimpairs pages 16-21
- piao2025advancesingene pages 2-3
- https://doi.org/10.1073/pnas.2319091121
- https://doi.org/10.1177/18796397241293955
- https://doi.org/10.3390/ijms25073845
- https://doi.org/10.1038/s41598-023-32630-4
- https://clinicaltrials.gov/study/NCT05686551
- https://clinicaltrials.gov/study/NCT05358717
- https://doi.org/10.1038/nrdp.2015.5,
- https://doi.org/10.1007/s10048-026-00887-2,
- https://doi.org/10.3390/ijms25073845,
- https://doi.org/10.1080/27694127.2025.2472450,
- https://doi.org/10.1038/s41598-023-32630-4,
- https://doi.org/10.1073/pnas.2319091121,
- https://doi.org/10.1177/18796397241293955,
- https://doi.org/10.3233/jhd-240017,
- https://doi.org/10.3233/jhd-239001,
- https://doi.org/10.1038/s41591-024-03424-6,
- https://doi.org/10.1093/procel/pwae042,
OpenAI
(HTT-deep-research-openai.md)
OpenAI
(HTT-deep-research-openai.md)Introduction to the HTT Gene and Huntingtin Protein
Huntingtin (HTT) is a human gene whose product, the huntingtin protein, is a very large (≈348 kDa, 3,144-amino acid) protein with critical roles in cellular function (pubmed.ncbi.nlm.nih.gov). It is best known because mutations (polyglutamine expansions) in HTT cause Huntington’s disease, but the normal huntingtin protein is essential for life and widely involved in cellular processes (pubmed.ncbi.nlm.nih.gov). HTT is ubiquitously expressed in the body, with especially high expression in the brain (notably in cerebral cortex, striatum, cerebellar cortex, and hippocampus) (www.reactome.org). At the subcellular level, huntingtin is predominantly a cytosolic protein (pubmed.ncbi.nlm.nih.gov), often associated with membranes and cytoskeletal elements. It can shuttle between the cytoplasm and nucleus (www.reactome.org), though under normal conditions most huntingtin resides in the cytoplasm and neuronal processes, whereas mutant N-terminal fragments tend to accumulate pathologically in nuclei (pubmed.ncbi.nlm.nih.gov). The HTT protein is essential for development – mice completely lacking huntingtin die in mid-gestation with disorganized embryos and increased cell death (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov), underscoring that huntingtin has fundamental pro-survival functions. Modern research has revealed that huntingtin acts as a scaffolding protein that participates in numerous cellular pathways, including vesicle trafficking, cytoskeletal dynamics, autophagy, endocytosis, and gene regulation (pubmed.ncbi.nlm.nih.gov). Below, we discuss the structure of huntingtin and detail its primary functions, the biological processes it governs, where it carries out these functions in the cell, and the molecular mechanisms involved – with an emphasis on current understanding and recent findings (2023–2024).
Structure and Domain Organization
Huntingtin has no enzymatic activity; instead, it is a scaffold protein composed largely of repeated helical domains that mediate protein–protein interactions. A landmark cryo-electron microscopy study resolved the near full-length huntingtin structure (in complex with one of its partners, HAP40), revealing that the protein’s amino-terminal and carboxy-terminal regions each form extensive HEAT repeat domains (HEAT repeats are helical motifs named for proteins Huntingtin, Elongation factor 3, PP2A, TOR) (pubmed.ncbi.nlm.nih.gov). These HEAT repeats are arranged into superhelical solenoids (often termed the N-HEAT and C-HEAT domains) (pubmed.ncbi.nlm.nih.gov). Connecting the two major halves of huntingtin is a smaller “Bridge” domain containing other tandem repeats (pubmed.ncbi.nlm.nih.gov). This architecture produces an elongated, flexible protein well-suited for binding multiple partners simultaneously. Notably, the extreme N-terminus of huntingtin contains a polyglutamine tract (encoded by a CAG repeat in HTT exon 1) and a neighboring polyproline region. In the normal population this polyQ tract ranges from ~10–35 glutamines, whereas expansions beyond ~36 glutamines cause a pathogenic protein conformation associated with Huntington’s disease (www.reactome.org). The N-terminal 17 amino acids (adjacent to the polyQ stretch) are important for huntingtin’s interactions and also undergo posttranslational modifications (like phosphorylation and myristoylation) that can influence the protein’s localization and function (www.reactome.org). Overall, huntingtin’s solenoid-like HEAT repeat structure provides a large surface area for assembling molecular complexes, consistent with its role as an interaction hub coordinating diverse cellular activities (pubmed.ncbi.nlm.nih.gov). This structural framework gives insight into how huntingtin can simultaneously engage with motors, adaptors, and membranes.
Expression and Subcellular Localization
HTT is widely expressed in human tissues, indicating its general cellular importance, but its expression is highest in neurons of the central nervous system (www.reactome.org). Within the brain, huntingtin protein is abundant in neuron-rich regions and localized in neuronal cell bodies, dendrites, axons, and nerve terminals (www.reactome.org). Subcellularly, huntingtin is found largely in the cytosol and on cytoplasmic membranes. It associates with intracellular organelles and vesicles, consistent with its role in trafficking (discussed below). Under basal conditions, wild-type huntingtin is predominantly cytoplasmic (pubmed.ncbi.nlm.nih.gov). It has been observed on endosomal and autophagic vesicle membranes, the Golgi apparatus, and in the synaptic terminals of neurons (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov). Huntingtin can also transiently enter the nucleus (it contains no classical nuclear localization signal, but can translocate by alternative means (www.reactome.org)). However, in healthy cells the full-length protein does not accumulate to high levels in nuclei (pubmed.ncbi.nlm.nih.gov); instead, it likely shuttles in and out performing regulatory roles (for example, influencing gene transcription as described later). Notably, when huntingtin is mutated and cleaved, the resulting N-terminal fragments aberrantly accumulate in nuclei and form aggregates in Huntington’s disease, but normal huntingtin primarily functions in the cytoplasm at sites where it can interface with the cytoskeleton and vesicular organelles (pubmed.ncbi.nlm.nih.gov). This distribution is in line with huntingtin’s key involvement in cellular transport processes and organelle dynamics in the cytoplasmic compartment.
Scaffold for Intracellular Transport along Microtubules
One of huntingtin’s principal functions is to facilitate intracellular transport of vesicles and organelles, especially in neurons which have long-distance trafficking needs. Huntingtin serves as a scaffolding platform connecting cargo vesicles with motor proteins that move along the cytoskeletal tracks. In fact, huntingtin has been described as a “ubiquitously expressed scaffolding protein” that plays a central role in regulating the transport of various organelles and vesicles (pubmed.ncbi.nlm.nih.gov). It interacts with numerous adaptor proteins and motor proteins to form what researchers call the “huntingtin transport complex.” Through this network, huntingtin can engage microtubule-based motors — including kinesin (for anterograde transport) and dynein/dynactin (for retrograde transport) — as well as link to actin-based motors like myosin VI for short-range transport (pubmed.ncbi.nlm.nih.gov). By forming multiprotein complexes, huntingtin helps attach motor proteins to specific cargo and orchestrates their movement along microtubules or actin filaments (pubmed.ncbi.nlm.nih.gov).
Importantly, huntingtin regulates a wide range of cargoes. For example, it is required for the axonal transport of brain-derived neurotrophic factor (BDNF) containing secretory vesicles (pubmed.ncbi.nlm.nih.gov). In healthy neurons, huntingtin binding to motors (via adaptors like HAP1 and dynactin) enhances BDNF vesicle movement along microtubules, ensuring trophic support is delivered to synapses (pubmed.ncbi.nlm.nih.gov). Experimental studies have shown that introducing normal huntingtin significantly increases the speed of BDNF vesicle transport, whereas mutant huntingtin or loss of huntingtin function impairs this movement (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Huntingtin similarly affects other vesicular cargo, such as vesicles carrying amyloid precursor protein (APP) (pubmed.ncbi.nlm.nih.gov), and it helps transport organelles including early endosomes, lysosomes, autophagosomes, and even mitochondria (pubmed.ncbi.nlm.nih.gov). Through these interactions, huntingtin effectively “guides” cargo through different stages of their life cycle – from biosynthetic pathways (Golgi to outposts), to signaling events (e.g. delivering trophic factors), to degradation pathways (autophagosome and lysosome delivery) (pubmed.ncbi.nlm.nih.gov). Disruption of huntingtin’s transport function (for instance by polyglutamine expansion) leads to trafficking defects; neurons with mutant huntingtin show accumulated or mislocalized cargoes and impaired axonal transport, which is thought to contribute to neurodegeneration (pubmed.ncbi.nlm.nih.gov). In summary, huntingtin’s primary role is as a scaffold that links motor proteins to their cargo, enabling efficient microtubule-based transport of vital cargos like growth factor vesicles and organelles throughout the cell (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov). This transport function is particularly crucial in neurons for maintaining synaptic function and survival.
Molecular mechanisms: Huntingtin’s transport facilitation involves several specific molecular interactions. It binds to HAP1 (Huntingtin-associated protein 1) and p150^Glued (a component of the dynein/dynactin motor complex), which together attach vesicles to the dynein motor (pmc.ncbi.nlm.nih.gov). It can also interact with kinesin-1 through adaptors like TRAK1/2 and others to drive anterograde movement. Additionally, huntingtin forms a complex with Rab5A and HAP40 on early endosomes (www.reactome.org), functioning as a Rab5 effector that recruits huntingtin to endosomal membranes. This allows huntingtin to help coordinate endocytic vesicle trafficking and maturation (www.reactome.org). Huntingtin’s scaffolding seems to require its large size and HEAT domain structure to simultaneously bind motors, adaptors, and cargo. Notably, specific regions of huntingtin are dedicated to transport functions: experiments mapping functional domains indicate the N-terminal region of huntingtin (within the first ~400–600 amino acids) is crucial for binding HAP1 and stimulating BDNF-vesicle transport (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Meanwhile, other regions bind different partners (e.g. a C-terminal region may engage kinesin or myosin adaptors). By recruiting multiple motor types (dynein, kinesin, myosin) to cargo, huntingtin even helps transfer cargo between microtubule and actin transport systems – for instance, secretory vesicles might use microtubule motors for long-range travel then switch to myosin for short-range movement in the actin-rich periphery, with huntingtin coordinating this handoff (pubmed.ncbi.nlm.nih.gov). This multi-motor coordination by huntingtin is a sophisticated mechanism ensuring cargo reach their correct destination in the cell’s complex architecture.
Role in Cytoskeletal Organization (Microtubules and Actin)
Beyond simply serving as a static link between motors and cargo, huntingtin also directly interacts with cytoskeletal filaments, influencing their organization. Historically, huntingtin was linked mostly to microtubule-based transport, but new research shows it has a direct role in actin cytoskeletal dynamics as well. A 2023 structural and cell-biological study demonstrated that huntingtin binds to F-actin (filamentous actin) and can crosslink actin filaments (pubmed.ncbi.nlm.nih.gov). Specifically, huntingtin’s N-terminal HEAT domain and Bridge domain can wrap around actin filaments, while the C-terminal HEAT domain is displaced upon binding actin (pubmed.ncbi.nlm.nih.gov). Huntingtin appears to form dimers that bridge two parallel actin filaments approximately 20 nm apart (pubmed.ncbi.nlm.nih.gov). Functionally, loss of huntingtin was shown to disturb the morphology and function of axonal growth cones in neurons (pubmed.ncbi.nlm.nih.gov). Neurons lacking huntingtin have disorganized actin networks in their growth cones, leading to stunted axon outgrowth and altered growth cone structure (pubmed.ncbi.nlm.nih.gov). Thus, huntingtin helps organize F-actin into bundles within growth cones and perhaps other subcellular regions, thereby supporting proper neuronal connectivity development. This actin-binding capability was a newly elucidated aspect of huntingtin’s function (pubmed.ncbi.nlm.nih.gov), expanding our understanding of huntingtin as a cytoskeletal organizer. It complements earlier evidence that huntingtin influences microtubule-based processes (indirectly via motors); now we know huntingtin can also directly stabilize or arrange the actin cytoskeleton.
In terms of microtubules, huntingtin’s role is more indirect but still critical. It doesn’t bind microtubules strongly itself (huntingtin is not a motor or MAP in the classic sense), but by recruiting motors and cargo, huntingtin effectively links organelles to the microtubule network. In doing so, it can impact microtubule organization and dynamics indirectly. Moreover, huntingtin interacts with proteins like profilin 1 (PFN1) (www.reactome.org) and others that modulate the cytoskeleton. Some data suggest huntingtin may also stabilize microtubules via complex formation. For example, huntingtin’s interaction with the dynactin complex (which itself can stabilize microtubules and mediate cargo binding) means loss of huntingtin could weaken microtubule-based transport stability (pubmed.ncbi.nlm.nih.gov). Additionally, huntingtin is implicated in mitotic spindle orientation and cell division in some cells (research in stem cells and development indicates that huntingtin might help position microtubule organizing centers, thereby affecting spindle orientation during mitosis (pmc.ncbi.nlm.nih.gov), although this is an emerging area).
Overall, huntingtin serves as a multifaceted cytoskeletal regulator: it links molecular motors to microtubules for long-range transport and bundles actin filaments for local structural organization. These activities ensure proper cellular architecture and material delivery, particularly in neurons where long axons and dynamic growth cones demand tight coordination between transport and the cytoskeletal scaffold (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov). Huntingtin’s ability to interface with both major cytoskeletal systems (microtubules and actin) underscores its central position in maintaining cell structure and intracellular trafficking routes.
Regulation of Autophagy and Vesicle Degradation
Huntingtin also plays a significant role in autophagy, the cellular pathway for degrading and recycling proteins and organelles via lysosomal machinery. Notably, huntingtin functions as a scaffold for selective macroautophagy, helping to link cargo destined for degradation with the autophagy machinery (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Research in mammalian cells has shown that huntingtin physically interacts with p62/sequestosome-1, which is an autophagy cargo receptor that binds ubiquitinated proteins slated for degradation (pmc.ncbi.nlm.nih.gov). At the same time, huntingtin can bind ULK1, a kinase that initiates autophagosome formation (pmc.ncbi.nlm.nih.gov). By binding both the cargo receptor (p62) and an autophagy initiator complex (ULK1 and likely other Atg proteins), huntingtin brings the cargo and the autophagosome precursor together (pmc.ncbi.nlm.nih.gov). In other words, huntingtin serves as a scaffold that bridges “cargo recognition” and “autophagosome initiation” during selective autophagy (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). This greatly increases the efficiency by which specific cargo (like misfolded proteins or damaged organelles) are recognized and enclosed in autophagosomes for degradation.
Experimental evidence supports this dual role: cells lacking huntingtin show reduced selective autophagy – they form autophagosomes, but the incorporation of specific cargo (e.g. ubiquitin-tagged protein aggregates) is less efficient (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In Drosophila models, huntingtin genetically interacts with autophagy pathway components, and in mammalian cells loss of huntingtin impairs the clearance of selective substrates while leaving bulk (non-selective) autophagy largely intact (pmc.ncbi.nlm.nih.gov). These findings led to the model that huntingtin is analogous to the yeast scaffold Atg11, which organizes selective autophagy cargo and machinery (pmc.ncbi.nlm.nih.gov). Huntingtin’s HEAT-repeat architecture may provide a flexible platform to tether p62 (and bound cargo) in proximity to the ULK1 complex that nucleates the autophagosome membrane. A study in Nature Cell Biology (2015) showed that eliminating huntingtin or disrupting its interaction with p62 abrogated the efficient clearance of mutant protein aggregates, whereas wild-type huntingtin strongly promoted their autophagic removal (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Therefore, huntingtin is considered dispensable for bulk autophagy but crucial for selective autophagy (pmc.ncbi.nlm.nih.gov) – it ensures that specific targeted cargos are recognized and handed off to the forming autophagosome. This role is another facet of huntingtin’s broader function in vesicle trafficking, here applied to the degradative pathway (autophagosomes are essentially vesicles that capture cargo for degradation). Furthermore, huntingtin’s interaction with the autophagosomal and lysosomal system ties into observations that huntingtin helps transport autophagosomes and lysosomes along microtubules (pubmed.ncbi.nlm.nih.gov); it likely facilitates their movement to fusion sites. In summary, huntingtin acts as a scaffold and coordinator in the autophagy pathway, promoting the clearance of cellular debris by linking cargo to the autophagy initiation machinery and aiding the trafficking of autophagic vesicles (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). This function explains, in part, why loss of normal huntingtin function can lead to accumulation of toxic proteins and organelles, contributing to cellular stress.
Influence on Gene Expression and Cell Signaling
Although huntingtin predominantly operates in the cytoplasm, it also influences gene expression and signaling pathways, underscoring its multifaceted regulatory role. One well-characterized function of huntingtin is its ability to modulate transcription of specific neuronal genes through interaction with transcriptional regulators. In particular, wild-type huntingtin enhances the expression of Brain-Derived Neurotrophic Factor (BDNF) and other neuronal genes by interfering with the repressor REST/NRSF (RE1 Silencing Transcription Factor). REST is a transcriptional repressor that silences neuron-specific genes by binding NRSE/RE1 elements. Huntingtin binds to REST and prevents REST from entering the nucleus and repressing target genes (pubmed.ncbi.nlm.nih.gov). In the case of the BDNF gene, huntingtin was shown to inhibit REST’s silencer function at the BDNF promoter, thereby increasing BDNF gene transcription (pubmed.ncbi.nlm.nih.gov). Wild-type (normal) huntingtin accomplishes this by sequestering REST in the cytoplasm or otherwise blocking its activity on chromatin (pubmed.ncbi.nlm.nih.gov). Mutant huntingtin (with expanded polyQ) cannot bind REST effectively, leading to REST accumulation in the nucleus and repression of BDNF and other genes (pubmed.ncbi.nlm.nih.gov). This mechanism was first elucidated in the early 2000s, when researchers found that cells expressing normal HTT had higher BDNF expression, whereas those with mutant HTT lost this transcriptional stimulation (pubmed.ncbi.nlm.nih.gov). Thus, one role of huntingtin in the nucleus is as a positive regulator of transcription for neurotrophic factors and possibly other neuron-specific genes, achieved by antagonizing transcriptional repressors like REST (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov). This function connects to neuronal survival: by boosting BDNF production (a key survival factor for certain neurons), huntingtin supports neuronal health at the gene expression level in addition to its vesicular transport of BDNF protein.
Huntingtin also intersects with intracellular signaling pathways. A notable example is its involvement in the NF-κB pathway in neurons. NF-κB is a transcription factor that can be activated at synapses and then needs to translocate to the nucleus to turn on stress-response genes. Wild-type huntingtin facilitates the retrograde transport of activated NF-κB from synapses to the nucleus (pmc.ncbi.nlm.nih.gov). Experiments using live-cell imaging demonstrated that when neurons receive excitatory stimuli at dendritic spines (activating NF-κB there), huntingtin helps move the NF-κB (specifically the p65 subunit) out of the dendritic spine and along the axon towards the nucleus (pmc.ncbi.nlm.nih.gov). In cells with normal HTT, a high level of active NF-κB reaches the nucleus in response to stimulation, whereas in cells with mutant HTT (polyQ-expanded), this NF-κB transport is impaired (pmc.ncbi.nlm.nih.gov). The result is a blunted transcriptional response to synaptic stimuli when huntingtin’s function is compromised (pmc.ncbi.nlm.nih.gov). This suggests that huntingtin acts as a facilitator of certain signaling cascades, linking synaptic events to nuclear gene regulation by literally transporting signaling molecules. Another signaling interface is huntingtin’s modulation of protein phosphorylation networks; for instance, some evidence indicates huntingtin may sequester protein phosphatase complexes or interact with kinases (CDK5 can phosphorylate huntingtin under DNA damage conditions, which protects neurons (www.reactome.org)). Moreover, huntingtin appears to have a role in mitochondrial homeostasis and calcium signaling indirectly: mutant huntingtin disrupts mitochondrial transport and network dynamics, whereas normal huntingtin might support mitochondrial trafficking to areas of high calcium or energy demand (pubmed.ncbi.nlm.nih.gov). All these points illustrate that huntingtin’s scaffolding functionality extends into the realm of signaling – it can bind components of signaling pathways or help ferry them to the right location.
In summary, huntingtin contributes to the regulation of gene expression and signaling in neurons by two main mechanisms: (1) Transcriptional modulation – by sequestering or interacting with transcription factors/co-factors (e.g. REST), it ensures proper expression of vital neuronal genes (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov). (2) Signal transduction facilitation – by transporting signaling molecules (e.g. NF-κB, perhaps others like calcium regulators or mRNA) to their site of action (pmc.ncbi.nlm.nih.gov). These nuclear and signaling roles, while less understood than its cytoplasmic transport functions, highlight huntingtin as a coordinator of cellular responses to stimuli. Notably, many of the downstream effects seen in Huntington’s disease (such as transcriptional dysregulation and impaired stress responses) reflect the loss of huntingtin’s normal function in these pathways (pmc.ncbi.nlm.nih.gov).
Current Research and Clinical Implications
Research into HTT and huntingtin is very active, with recent developments further clarifying its function. For example, a 2023 cryo-electron tomography study provided the first direct visualization of huntingtin bound to actin filaments, solidifying the concept that huntingtin structurally organizes the actin cytoskeleton (pubmed.ncbi.nlm.nih.gov). This adds to our understanding that huntingtin is not only a microtubule transport scaffold but also an actin-bundling factor, especially relevant in neuronal growth cones and possibly other dynamic actin-rich structures. Additionally, comprehensive proteomics and interaction studies (e.g. mapping the huntingtin interactome) continue to identify new partners and pathways involving huntingtin. Hundreds of huntingtin-interacting proteins have been catalogued, ranging from motor/adaptor proteins to regulators of transcription, metabolism, and signal transduction (www.preprints.org) (www.preprints.org). Modern high-throughput studies (2022–2024) are connecting these interactions with functional outcomes, helping to piece together an integrative model of how huntingtin coordinates cellular physiology. For instance, recent data implicate huntingtin in pathways like DNA damage response (phosphorylated huntingtin can protect against DNA-damage-induced toxicity (www.reactome.org)) and synaptic vesicle recycling, showing the breadth of its influence. Despite this, researchers note that “the functions of HTT are still not fully understood” (www.preprints.org), reflecting the complexity of this large protein.
From a clinical perspective, understanding huntingtin’s normal functions is vital. Therapeutic approaches for Huntington’s disease are being developed to reduce the levels of mutant huntingtin (for example, via antisense oligonucleotides or gene-silencing vectors). While lowering mutant HTT can alleviate toxicity, these strategies must contend with the fact that wild-type huntingtin is necessary for neurons. Indeed, adult mice in which HTT is globally knocked out eventually develop severe neurodegeneration, indicating that ongoing huntingtin function is required even beyond development (pmc.ncbi.nlm.nih.gov). This has prompted efforts to design allele-specific therapies (reducing mutant HTT while sparing normal HTT) or temporal control of HTT lowering. A first-in-human gene therapy trial (using an HTT-targeting vector) reported in 2023 that partial lowering of HTT in patients is achievable (www.livescience.com), but long-term outcomes will depend on maintaining enough huntingtin for its normal roles. The current consensus among experts is that huntingtin’s normal activities – supporting axonal transport, neurotrophic factor release, proteostasis via autophagy, etc. – are crucial to preserve. As one recent review put it, huntingtin is involved “here, there, everywhere” in the cell (pmc.ncbi.nlm.nih.gov), making it a hub of cellular homeostasis. Thus, a comprehensive understanding of HTT’s functional biology guides not only basic science but also the development of safe interventions for Huntington’s disease.
Conclusion
Huntingtin (HTT) emerges from decades of research as a versatile scaffolding protein essential for neuronal function and survival. Its primary role is to serve as a platform that brings together cargo and the machinery needed to move or process that cargo – whether it be transporting growth factor vesicles along microtubules, bundling actin filaments in a growth cone, or bridging autophagy cargo to the autophagosome initiation complex. Through these activities, huntingtin touches a remarkably wide range of biological processes: axonal transport, vesicle trafficking, cytoskeletal assembly, endocytosis, autophagy, and gene regulation (pubmed.ncbi.nlm.nih.gov). It predominantly carries out these functions in the cytoplasm, at organelle membranes and cytoskeletal tracks, but also has important influences in the nucleus (indirectly affecting transcription). In molecular pathways, huntingtin is a key node in maintaining neuronal health – promoting the delivery of BDNF to neurons, removal of misfolded proteins via autophagy, proper synapse-to-nucleus signaling, and expression of survival genes. These precise roles have been illuminated by a combination of genetic, biochemical, and structural studies, with strong experimental evidence underlining each function: e.g. the embryonic lethality of HTT knockout shows its necessity (pubmed.ncbi.nlm.nih.gov), live-cell imaging confirms its transport of vesicles (pubmed.ncbi.nlm.nih.gov), and high-resolution structural studies depict its binding to actin and autophagy proteins (pubmed.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). As research continues (especially with advanced techniques in 2023–2024), our picture of huntingtin is increasingly that of a master coordinator of intracellular logistics and homeostasis. Its broad, pleiotropic effects in the cell stem from a unifying principle: huntingtin ensures that the right components (proteins, organelles, signals) are in the right place at the right time. This central functionality explains why disrupting HTT leads to widespread cellular dysfunction. Therefore, when functionally annotating the HTT gene, one must emphasize that huntingtin’s biological significance lies in its scaffolding capacity, enabling critical processes in neuronal cells and beyond – a role that is as complex as it is vital (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov).
References: The information above is supported by recent high-impact studies and reviews, including structural biology findings (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov), cellular and molecular biology experiments (pubmed.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov), and authoritative reviews on huntingtin’s function (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov) (see inline citations). Each citation corresponds to a source detailing the claim: for example, Saudou et al. (2004) showed huntingtin’s role in BDNF vesicle transport (pubmed.ncbi.nlm.nih.gov), Rui et al. (2015) demonstrated the autophagy scaffolding function (pmc.ncbi.nlm.nih.gov), and Shirasaki et al. (2023) revealed the actin-binding mechanism (pubmed.ncbi.nlm.nih.gov). This integrated, evidence-based view provides a current and comprehensive functional annotation of the human HTT gene product, huntingtin.
Citations
- AnnotationURLCitation(end_index=347, start_index=222, title='The cryo-electron microscopy structure of huntingtin - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/29466333/#:~:text=Huntingtin%20,of%20HTT%20is%20lacking%2C%20the')
- AnnotationURLCitation(end_index=669, start_index=544, title='The cryo-electron microscopy structure of huntingtin - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/29466333/#:~:text=Huntingtin%20,of%20HTT%20is%20lacking%2C%20the')
- AnnotationURLCitation(end_index=968, start_index=834, title='Reactome | UniProt:P42858 HTT', type='url_citation', url='https://www.reactome.org/content/schema/instance/browser/56424#:~:text=%28PubMed%3A16476778%29,PubMed%3A10770929')
- AnnotationURLCitation(end_index=1189, start_index=1044, title='Nuclear localization of N-terminal mutant huntingtin is cell cycle dependent - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12169117/#:~:text=Nuclear%20localization%20of%20N,in%20the%20nucleus%20of%20affected')
- AnnotationURLCitation(end_index=1429, start_index=1299, title='Reactome | UniProt:P42858 HTT', type='url_citation', url='https://www.reactome.org/content/schema/instance/browser/56424#:~:text=%28PubMed%3A16476778%29,rich%20domain')
- AnnotationURLCitation(end_index=1752, start_index=1607, title='Nuclear localization of N-terminal mutant huntingtin is cell cycle dependent - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12169117/#:~:text=Nuclear%20localization%20of%20N,in%20the%20nucleus%20of%20affected')
- AnnotationURLCitation(end_index=2034, start_index=1912, title="Increased apoptosis and early embryonic lethality in mice nullizygous for the Huntington's disease gene homologue - PubMed", type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/7550343/#:~:text=The%20expansion%20of%20CAG%20triplet,5%20and')
- AnnotationURLCitation(end_index=2210, start_index=2035, title="Increased apoptosis and early embryonic lethality in mice nullizygous for the Huntington's disease gene homologue - PubMed", type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/7550343/#:~:text=become%20developmentally%20retarded%20and%20disorganized%2C,we%20propose%20that%20huntingtin%20is')
- AnnotationURLCitation(end_index=2636, start_index=2511, title='The cryo-electron microscopy structure of huntingtin - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/29466333/#:~:text=Huntingtin%20,of%20HTT%20is%20lacking%2C%20the')
- AnnotationURLCitation(end_index=3603, start_index=3484, title='The cryo-electron microscopy structure of huntingtin - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/29466333/#:~:text=domains.%20The%20amino,HAP40%20is%20also')
- AnnotationURLCitation(end_index=3829, start_index=3710, title='The cryo-electron microscopy structure of huntingtin - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/29466333/#:~:text=domains.%20The%20amino,HAP40%20is%20also')
- AnnotationURLCitation(end_index=4061, start_index=3942, title='The cryo-electron microscopy structure of huntingtin - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/29466333/#:~:text=domains.%20The%20amino,HAP40%20is%20also')
- AnnotationURLCitation(end_index=4684, start_index=4529, title='Reactome | UniProt:P42858 HTT', type='url_citation', url='https://www.reactome.org/content/schema/instance/browser/56424#:~:text=Myristoylated%20at%20Gly,disease%20is%20caused%20by%20variants')
- AnnotationURLCitation(end_index=5080, start_index=4946, title='Reactome | UniProt:P42858 HTT', type='url_citation', url='https://www.reactome.org/content/schema/instance/browser/56424#:~:text=equilibrium%20of%20alpha,550.POLYMORPHISM')
- AnnotationURLCitation(end_index=5423, start_index=5298, title='The cryo-electron microscopy structure of huntingtin - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/29466333/#:~:text=Huntingtin%20,of%20HTT%20is%20lacking%2C%20the')
- AnnotationURLCitation(end_index=5892, start_index=5758, title='Reactome | UniProt:P42858 HTT', type='url_citation', url='https://www.reactome.org/content/schema/instance/browser/56424#:~:text=%28PubMed%3A16476778%29,PubMed%3A10770929')
- AnnotationURLCitation(end_index=6177, start_index=6043, title='Reactome | UniProt:P42858 HTT', type='url_citation', url='https://www.reactome.org/content/schema/instance/browser/56424#:~:text=%28PubMed%3A16476778%29,PubMed%3A10770929')
- AnnotationURLCitation(end_index=6611, start_index=6466, title='Nuclear localization of N-terminal mutant huntingtin is cell cycle dependent - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12169117/#:~:text=Nuclear%20localization%20of%20N,in%20the%20nucleus%20of%20affected')
- AnnotationURLCitation(end_index=6912, start_index=6747, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=multitude%20of%20cellular%20roles%2C%20including,kinesin%20and%20dynein%2C%20as%20well')
- AnnotationURLCitation(end_index=7088, start_index=6913, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=autophagosomes%2C%20lysosomes%2C%20and%20mitochondria,cargoes%20and%20guides%20cargoes%20through')
- AnnotationURLCitation(end_index=7366, start_index=7236, title='Reactome | UniProt:P42858 HTT', type='url_citation', url='https://www.reactome.org/content/schema/instance/browser/56424#:~:text=%28PubMed%3A16476778%29,rich%20domain')
- AnnotationURLCitation(end_index=7609, start_index=7464, title='Nuclear localization of N-terminal mutant huntingtin is cell cycle dependent - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12169117/#:~:text=Nuclear%20localization%20of%20N,in%20the%20nucleus%20of%20affected')
- AnnotationURLCitation(end_index=8192, start_index=8047, title='Nuclear localization of N-terminal mutant huntingtin is cell cycle dependent - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12169117/#:~:text=Nuclear%20localization%20of%20N,in%20the%20nucleus%20of%20affected')
- AnnotationURLCitation(end_index=9030, start_index=8900, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=Huntingtin%20,kinesin%20and%20dynein%2C%20as%20well')
- AnnotationURLCitation(end_index=9597, start_index=9422, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=autophagosomes%2C%20lysosomes%2C%20and%20mitochondria,cargoes%20and%20guides%20cargoes%20through')
- AnnotationURLCitation(end_index=9905, start_index=9761, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=autophagosomes%2C%20lysosomes%2C%20and%20mitochondria,Accordingly')
- AnnotationURLCitation(end_index=10268, start_index=10103, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=multitude%20of%20cellular%20roles%2C%20including,kinesin%20and%20dynein%2C%20as%20well')
- AnnotationURLCitation(end_index=10623, start_index=10458, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=multitude%20of%20cellular%20roles%2C%20including,kinesin%20and%20dynein%2C%20as%20well')
- AnnotationURLCitation(end_index=10982, start_index=10832, title='pARIS-htt: an optimised expression platform to study huntingtin reveals functional domains required for vesicular trafficking - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC2887845/#:~:text=encoded%20by%20exon%201%2C%20stimulates,length%20protein%20context')
- AnnotationURLCitation(end_index=11073, start_index=10983, title='pARIS-htt: an optimised expression platform to study huntingtin reveals functional domains required for vesicular trafficking - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC2887845/#:~:text=BDNF,p')
- AnnotationURLCitation(end_index=11354, start_index=11189, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=multitude%20of%20cellular%20roles%2C%20including,kinesin%20and%20dynein%2C%20as%20well')
- AnnotationURLCitation(end_index=11639, start_index=11474, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=multitude%20of%20cellular%20roles%2C%20including,kinesin%20and%20dynein%2C%20as%20well')
- AnnotationURLCitation(end_index=12053, start_index=11923, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=huntingtin%20to%20control%20microtubule,Accordingly')
- AnnotationURLCitation(end_index=12497, start_index=12328, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=huntingtin%20to%20control%20microtubule,defects%20in%20transport%20and%20neurodegeneration')
- AnnotationURLCitation(end_index=12884, start_index=12730, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/40971423/#:~:text=The%20Huntingtin%20protein%20,parallel%20actin%20filaments%20separated%20by')
- AnnotationURLCitation(end_index=13020, start_index=12885, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=Huntingtin%20,cargoes%20and%20guides%20cargoes%20through')
- AnnotationURLCitation(end_index=13573, start_index=13420, title='pARIS-htt: an optimised expression platform to study huntingtin reveals functional domains required for vesicular trafficking - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC2887845/#:~:text=Htt%20function%20in%20MT,trafficking%20in%20cells%2C%20whereas%20each')
- AnnotationURLCitation(end_index=13971, start_index=13772, title='Reactome | UniProt:P42858 HTT', type='url_citation', url='https://www.reactome.org/content/schema/instance/browser/56424#:~:text=F8A1%2FF8A2%2FF8A3%20%28PubMed%3A16476778%2C%20PubMed%3A29466333%29,the%20brain%2C%20the%20regions%20where')
- AnnotationURLCitation(end_index=14339, start_index=14140, title='Reactome | UniProt:P42858 HTT', type='url_citation', url='https://www.reactome.org/content/schema/instance/browser/56424#:~:text=F8A1%2FF8A2%2FF8A3%20%28PubMed%3A16476778%2C%20PubMed%3A29466333%29,the%20brain%2C%20the%20regions%20where')
- AnnotationURLCitation(end_index=14884, start_index=14749, title='pARIS-htt: an optimised expression platform to study huntingtin reveals functional domains required for vesicular trafficking - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC2887845/#:~:text=pARIS,httQ23%2FQ100%20or%20dynein%2FHAP1%20deletion')
- AnnotationURLCitation(end_index=15038, start_index=14885, title='pARIS-htt: an optimised expression platform to study huntingtin reveals functional domains required for vesicular trafficking - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC2887845/#:~:text=Htt%20function%20in%20MT,trafficking%20in%20cells%2C%20whereas%20each')
- AnnotationURLCitation(end_index=15694, start_index=15519, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=autophagosomes%2C%20lysosomes%2C%20and%20mitochondria,cargoes%20and%20guides%20cargoes%20through')
- AnnotationURLCitation(end_index=16563, start_index=16398, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/40971423/#:~:text=molecular%20motors%20along%20microtubules,and%20organizes%20the%20actin%20cytoskeleton')
- AnnotationURLCitation(end_index=16897, start_index=16735, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/40971423/#:~:text=function%20of%20the%20axonal%20growth,parallel%20actin%20filaments%20separated%20by')
- AnnotationURLCitation(end_index=17171, start_index=17000, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/40971423/#:~:text=structural%20analyses%20reveal%20that%20HTT%27s,and%20organizes%20the%20actin%20cytoskeleton')
- AnnotationURLCitation(end_index=17447, start_index=17293, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/40971423/#:~:text=The%20Huntingtin%20protein%20,parallel%20actin%20filaments%20separated%20by')
- AnnotationURLCitation(end_index=17765, start_index=17600, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/40971423/#:~:text=molecular%20motors%20along%20microtubules,and%20organizes%20the%20actin%20cytoskeleton')
- AnnotationURLCitation(end_index=18183, start_index=18029, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/40971423/#:~:text=The%20Huntingtin%20protein%20,parallel%20actin%20filaments%20separated%20by')
- AnnotationURLCitation(end_index=19049, start_index=18894, title='Reactome | UniProt:P42858 HTT', type='url_citation', url='https://www.reactome.org/content/schema/instance/browser/56424#:~:text=%28PubMed%3A16476778%29,the%20brain%2C%20the%20regions%20where')
- AnnotationURLCitation(end_index=19560, start_index=19391, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=huntingtin%20to%20control%20microtubule,defects%20in%20transport%20and%20neurodegeneration')
- AnnotationURLCitation(end_index=20006, start_index=19839, title='Effect of early embryonic deletion of huntingtin from pyramidal neurons on the development and long-term survival of neurons in cerebral cortex and striatum - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC5821111/#:~:text=Skip%20to%20main%20content%20Neurobiol,huntingtin%20from%20pyramidal%20neurons%20on')
- AnnotationURLCitation(end_index=20627, start_index=20473, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/40971423/#:~:text=The%20Huntingtin%20protein%20,parallel%20actin%20filaments%20separated%20by')
- AnnotationURLCitation(end_index=20793, start_index=20628, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/40971423/#:~:text=molecular%20motors%20along%20microtubules,and%20organizes%20the%20actin%20cytoskeleton')
- AnnotationURLCitation(end_index=21525, start_index=21359, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=a%20scaffold%20protein%20for%20selective,p62%20to%20facilitate%20its%20association')
- AnnotationURLCitation(end_index=21625, start_index=21526, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=processes,First')
- AnnotationURLCitation(end_index=21991, start_index=21825, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=a%20scaffold%20protein%20for%20selective,p62%20to%20facilitate%20its%20association')
- AnnotationURLCitation(end_index=22188, start_index=22089, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=processes,First')
- AnnotationURLCitation(end_index=22478, start_index=22379, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=processes,First')
- AnnotationURLCitation(end_index=22790, start_index=22623, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=scaffolding%20for%20selective%20autophagy%20by,it%20does%20not%20affect%20autophagy')
- AnnotationURLCitation(end_index=22890, start_index=22791, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=processes,First')
- AnnotationURLCitation(end_index=23471, start_index=23305, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=a%20scaffold%20protein%20for%20selective,p62%20to%20facilitate%20its%20association')
- AnnotationURLCitation(end_index=23638, start_index=23472, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=match%20at%20L79%20These%20data,cargo%20recognition%20and%20autophagy%20initiation')
- AnnotationURLCitation(end_index=24044, start_index=23878, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=a%20scaffold%20protein%20for%20selective,p62%20to%20facilitate%20its%20association')
- AnnotationURLCitation(end_index=24342, start_index=24196, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=match%20at%20L58%20cargo%20for,6%7D.%20A%20second%20difference')
- AnnotationURLCitation(end_index=24940, start_index=24774, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=a%20scaffold%20protein%20for%20selective,p62%20to%20facilitate%20its%20association')
- AnnotationURLCitation(end_index=25040, start_index=24941, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=processes,First')
- AnnotationURLCitation(end_index=25315, start_index=25149, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=a%20scaffold%20protein%20for%20selective,p62%20to%20facilitate%20its%20association')
- AnnotationURLCitation(end_index=25978, start_index=25813, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=multitude%20of%20cellular%20roles%2C%20including,kinesin%20and%20dynein%2C%20as%20well')
- AnnotationURLCitation(end_index=26437, start_index=26270, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=scaffolding%20for%20selective%20autophagy%20by,it%20does%20not%20affect%20autophagy')
- AnnotationURLCitation(end_index=26537, start_index=26438, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=processes,First')
- AnnotationURLCitation(end_index=27677, start_index=27518, title='Huntingtin interacts with REST/NRSF to modulate the transcription of NRSE-controlled neuronal genes - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12881722/#:~:text=REST%2FNRSF%20in%20the%20nucleus%20is,acts%20as%20a%20positive%20transcriptional')
- AnnotationURLCitation(end_index=27982, start_index=27839, title='Huntingtin interacts with REST/NRSF to modulate the transcription of NRSE-controlled neuronal genes - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12881722/#:~:text=Huntingtin%20protein%20is%20mutated%20in,We%20show%20that%20this')
- AnnotationURLCitation(end_index=28277, start_index=28118, title='Huntingtin interacts with REST/NRSF to modulate the transcription of NRSE-controlled neuronal genes - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12881722/#:~:text=REST%2FNRSF%20in%20the%20nucleus%20is,acts%20as%20a%20positive%20transcriptional')
- AnnotationURLCitation(end_index=28591, start_index=28432, title='Huntingtin interacts with REST/NRSF to modulate the transcription of NRSE-controlled neuronal genes - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12881722/#:~:text=REST%2FNRSF%20in%20the%20nucleus%20is,acts%20as%20a%20positive%20transcriptional')
- AnnotationURLCitation(end_index=28944, start_index=28801, title='Huntingtin interacts with REST/NRSF to modulate the transcription of NRSE-controlled neuronal genes - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12881722/#:~:text=Huntingtin%20protein%20is%20mutated%20in,We%20show%20that%20this')
- AnnotationURLCitation(end_index=29314, start_index=29171, title='Huntingtin interacts with REST/NRSF to modulate the transcription of NRSE-controlled neuronal genes - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12881722/#:~:text=Huntingtin%20protein%20is%20mutated%20in,We%20show%20that%20this')
- AnnotationURLCitation(end_index=29474, start_index=29315, title='Huntingtin interacts with REST/NRSF to modulate the transcription of NRSE-controlled neuronal genes - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12881722/#:~:text=REST%2FNRSF%20in%20the%20nucleus%20is,acts%20as%20a%20positive%20transcriptional')
- AnnotationURLCitation(end_index=30261, start_index=30116, title="The Huntington's disease mutation impairs Huntingtin's role in the transport of NF-κB from the synapse to the nucleus - PMC", type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC2957321/#:~:text=and%20neuronal%20transcription,a%20high%20level%20of%20active')
- AnnotationURLCitation(end_index=30677, start_index=30532, title="The Huntington's disease mutation impairs Huntingtin's role in the transport of NF-κB from the synapse to the nucleus - PMC", type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC2957321/#:~:text=and%20neuronal%20transcription,a%20high%20level%20of%20active')
- AnnotationURLCitation(end_index=31015, start_index=30870, title="The Huntington's disease mutation impairs Huntingtin's role in the transport of NF-κB from the synapse to the nucleus - PMC", type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC2957321/#:~:text=and%20neuronal%20transcription,a%20high%20level%20of%20active')
- AnnotationURLCitation(end_index=31250, start_index=31128, title="The Huntington's disease mutation impairs Huntingtin's role in the transport of NF-κB from the synapse to the nucleus - PMC", type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC2957321/#:~:text=genes%29,to%20the%20etiology%20of%20HD')
- AnnotationURLCitation(end_index=31927, start_index=31738, title='Reactome | UniProt:P42858 HTT', type='url_citation', url='https://www.reactome.org/content/schema/instance/browser/56424#:~:text=expanded%20polyglutamine%20expansion%20are%20specifically,region%20is%20also%20polymorphic%20and')
- AnnotationURLCitation(end_index=32387, start_index=32222, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=multitude%20of%20cellular%20roles%2C%20including,kinesin%20and%20dynein%2C%20as%20well')
- AnnotationURLCitation(end_index=33028, start_index=32885, title='Huntingtin interacts with REST/NRSF to modulate the transcription of NRSE-controlled neuronal genes - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12881722/#:~:text=Huntingtin%20protein%20is%20mutated%20in,We%20show%20that%20this')
- AnnotationURLCitation(end_index=33188, start_index=33029, title='Huntingtin interacts with REST/NRSF to modulate the transcription of NRSE-controlled neuronal genes - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12881722/#:~:text=REST%2FNRSF%20in%20the%20nucleus%20is,acts%20as%20a%20positive%20transcriptional')
- AnnotationURLCitation(end_index=33499, start_index=33354, title="The Huntington's disease mutation impairs Huntingtin's role in the transport of NF-κB from the synapse to the nucleus - PMC", type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC2957321/#:~:text=and%20neuronal%20transcription,a%20high%20level%20of%20active')
- AnnotationURLCitation(end_index=34004, start_index=33882, title="The Huntington's disease mutation impairs Huntingtin's role in the transport of NF-κB from the synapse to the nucleus - PMC", type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC2957321/#:~:text=genes%29,to%20the%20etiology%20of%20HD')
- AnnotationURLCitation(end_index=34549, start_index=34384, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/40971423/#:~:text=molecular%20motors%20along%20microtubules,and%20organizes%20the%20actin%20cytoskeleton')
- AnnotationURLCitation(end_index=35243, start_index=35121, title='Recent Advances in the Study of Huntingtin Interacting Proteins[v1] | Preprints.org', type='url_citation', url='https://www.preprints.org/manuscript/202307.0615/v1#:~:text=HTT,HIP1%29%20have%20been%20extensively')
- AnnotationURLCitation(end_index=35392, start_index=35244, title='Recent Advances in the Study of Huntingtin Interacting Proteins[v1] | Preprints.org', type='url_citation', url='https://www.preprints.org/manuscript/202307.0615/v1#:~:text=The%20functions%20of%20HTT%20are,other%20aspects%20of%20HTT%20and')
- AnnotationURLCitation(end_index=35947, start_index=35758, title='Reactome | UniProt:P42858 HTT', type='url_citation', url='https://www.reactome.org/content/schema/instance/browser/56424#:~:text=expanded%20polyglutamine%20expansion%20are%20specifically,region%20is%20also%20polymorphic%20and')
- AnnotationURLCitation(end_index=36261, start_index=36113, title='Recent Advances in the Study of Huntingtin Interacting Proteins[v1] | Preprints.org', type='url_citation', url='https://www.preprints.org/manuscript/202307.0615/v1#:~:text=The%20functions%20of%20HTT%20are,other%20aspects%20of%20HTT%20and')
- AnnotationURLCitation(end_index=37079, start_index=36916, title='Global huntingtin knockout in adult mice leads to fatal neurodegeneration that spares the pancreas - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11272958/#:~:text=Global%20huntingtin%20knockout%20in%20adult,old%20animals.%20The%20levels%20of')
- AnnotationURLCitation(end_index=37623, start_index=37369, title="2025-09-24 | 'Groundbreaking' gene therapy is first treatment for Huntington's disease to slow the condition", type='url_citation', url='https://www.livescience.com/health/medicine-drugs/groundbreaking-gene-therapy-is-first-treatment-for-huntingtons-disease-to-slow-the-condition#:~:text=Huntington%27s%20disease%20to%20slow%20the,130%20targets%20the%20condition%27s')
- AnnotationURLCitation(end_index=38142, start_index=38004, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC12448071/#:~:text=cytoskeleton%20organization%20,2%2C%20395%E2%80%93403')
- AnnotationURLCitation(end_index=39175, start_index=39050, title='The cryo-electron microscopy structure of huntingtin - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/29466333/#:~:text=Huntingtin%20,of%20HTT%20is%20lacking%2C%20the')
- AnnotationURLCitation(end_index=39987, start_index=39865, title="Increased apoptosis and early embryonic lethality in mice nullizygous for the Huntington's disease gene homologue - PubMed", type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/7550343/#:~:text=The%20expansion%20of%20CAG%20triplet,5%20and')
- AnnotationURLCitation(end_index=40207, start_index=40042, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=multitude%20of%20cellular%20roles%2C%20including,kinesin%20and%20dynein%2C%20as%20well')
- AnnotationURLCitation(end_index=40460, start_index=40299, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/40971423/#:~:text=function%20of%20the%20axonal%20growth,and%20organizes%20the%20actin%20cytoskeleton')
- AnnotationURLCitation(end_index=40560, start_index=40461, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=processes,First')
- AnnotationURLCitation(end_index=41434, start_index=41299, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=Huntingtin%20,cargoes%20and%20guides%20cargoes%20through')
- AnnotationURLCitation(end_index=41560, start_index=41435, title='The cryo-electron microscopy structure of huntingtin - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/29466333/#:~:text=Huntingtin%20,of%20HTT%20is%20lacking%2C%20the')
- AnnotationURLCitation(end_index=41850, start_index=41696, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/40971423/#:~:text=The%20Huntingtin%20protein%20,parallel%20actin%20filaments%20separated%20by')
- AnnotationURLCitation(end_index=41970, start_index=41851, title='The cryo-electron microscopy structure of huntingtin - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/29466333/#:~:text=domains.%20The%20amino,HAP40%20is%20also')
- AnnotationURLCitation(end_index=42154, start_index=42015, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=multitude%20of%20cellular%20roles%2C%20including,Accordingly')
- AnnotationURLCitation(end_index=42254, start_index=42155, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=processes,First')
- AnnotationURLCitation(end_index=42432, start_index=42307, title='The cryo-electron microscopy structure of huntingtin - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/29466333/#:~:text=Huntingtin%20,of%20HTT%20is%20lacking%2C%20the')
- AnnotationURLCitation(end_index=42576, start_index=42433, title='Huntingtin interacts with REST/NRSF to modulate the transcription of NRSE-controlled neuronal genes - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/12881722/#:~:text=Huntingtin%20protein%20is%20mutated%20in,We%20show%20that%20this')
- AnnotationURLCitation(end_index=42910, start_index=42745, title='The Huntingtin Transport Complex - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/39909923/#:~:text=multitude%20of%20cellular%20roles%2C%20including,kinesin%20and%20dynein%2C%20as%20well')
- AnnotationURLCitation(end_index=43077, start_index=42978, title='Huntingtin Functions as a Scaffold for Selective Macroautophagy - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4344873/#:~:text=processes,First')
- AnnotationURLCitation(end_index=43309, start_index=43144, title='Structure of the Huntingtin F-actin complex reveals its role in cytoskeleton organization - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/40971423/#:~:text=molecular%20motors%20along%20microtubules,and%20organizes%20the%20actin%20cytoskeleton')