Deep Research
Falcon
(INS-deep-research-falcon.md)
Falcon
(INS-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Human INS (UniProt P01308) — Functional Annotation, Biology, and Applications (focus on 2023–2024)
0) Identity verification (critical disambiguation)
The target is human insulin encoded by INS (UniProt P01308) and synthesized as preproinsulin, a secreted peptide hormone precursor that is proteolytically processed to the mature insulin A and B chains plus C-peptide. Multiple sources explicitly describe the canonical preproinsulin organization (signal peptide + B chain + C-peptide + A chain), ER targeting/processing, granule storage, and regulated secretion in pancreatic β-cells, matching the UniProt description and insulin-family context. (urbaniak2025molecularpuzzleof pages 3-6, urbaniak2025molecularpuzzleof pages 2-3, ayan2023abriefatlas pages 2-3)
1) Key concepts and definitions (current understanding)
1.1 INS gene product and maturation pathway
INS encodes preproinsulin, a single-chain precursor that enters the regulated secretory pathway. After co-translational targeting to the rough ER, the signal peptide is removed to generate proinsulin, which folds in the ER and forms three disulfide bonds. Proinsulin then traffics through the Golgi and is packaged into secretory granules, where endoproteolytic processing yields mature insulin and C-peptide. (urbaniak2025molecularpuzzleof pages 2-3, ayan2023abriefatlas pages 2-3, szablewski2024changesincells pages 2-4)
A key definitional distinction:
- Preproinsulin: signal peptide-containing translation product targeted to the ER. (urbaniak2025molecularpuzzleof pages 2-3, ayan2023abriefatlas pages 2-3)
- Proinsulin: ER-luminal folding intermediate; precursor that contains B chain, C-peptide, and A chain. (ayan2023abriefatlas pages 2-3, szablewski2024changesincells pages 2-4)
- Mature insulin: disulfide-linked A and B chains; stored in granules (often as zinc-stabilized hexamers) and secreted upon stimulation. (ayan2023abriefatlas pages 2-3, urbaniak2025molecularpuzzleof pages 7-8)
Processing enzymes (canonical): prohormone convertases PC1/3 and PC2 cleave junctions flanking the C-peptide, and carboxypeptidase E (CPE) trims basic residues to yield mature insulin and C-peptide. (prefontaine2024investigatingtherelationship pages 8-13, szablewski2024changesincells pages 2-4, urbaniak2025molecularpuzzleof pages 7-8)
1.2 Subcellular localization of INS function
The INS product is primarily synthesized and matured across multiple compartments:
- Rough ER/ER lumen: signal peptide cleavage, oxidative folding, disulfide bond formation, quality control. (urbaniak2025molecularpuzzleof pages 2-3, rohli2024ametabolicredox pages 1-2)
- Golgi/TGN: trafficking and packaging into immature secretory granules. (ayan2023abriefatlas pages 2-3, urbaniak2025molecularpuzzleof pages 7-8)
- Immature/maturing secretory granules: proteolytic conversion of proinsulin → insulin + C-peptide; granule acidification/condensation and storage. (szablewski2024changesincells pages 2-4, urbaniak2025molecularpuzzleof pages 7-8)
- Plasma membrane/extracellular: Ca2+-triggered exocytosis releases insulin to circulation; insulin acts as a ligand for the insulin receptor in peripheral tissues. (szablewski2024changesincells pages 2-4, sakaguchi2024theroleof pages 1-3)
1.3 Glucose-stimulated insulin secretion (GSIS) and biphasic release
A core concept is biphasic insulin secretion:
- Triggering (first) phase: glucose metabolism increases ATP, closes KATP channels, depolarizes the β-cell, opens voltage-gated Ca2+ channels, and triggers exocytosis of a readily releasable granule pool. (dalle2024receptorsandsignaling pages 2-4, aamodt2023peelingtheonion pages 1-2)
- Amplifying (second) phase: sustained secretion driven by metabolic coupling factors and cytoskeletal/granule mobilization processes; involves actin remodeling and small GTPases. (szablewski2024changesincells pages 2-4, dalle2024receptorsandsignaling pages 2-4)
1.4 Downstream signaling: insulin receptor pathway
Insulin acts mainly by binding INSR, which autophosphorylates and recruits adaptor proteins (IRS, Shc), bifurcating into:
- PI3K→PIP3→AKT (metabolic branch): GLUT4 translocation, glycogen synthesis regulation via GSK3, mTOR-mediated protein synthesis, and transcriptional programs (e.g., FOXO regulation). (sakaguchi2024theroleof pages 1-3, szablewski2024changesincells pages 4-6, caturano2024insulin–heartaxisbridging pages 2-3)
- RAS→RAF→MEK→ERK (MAPK) (mitogenic branch): growth/proliferation and gene-expression programs. (sakaguchi2024theroleof pages 1-3, szablewski2024changesincells pages 4-6)
2) Recent developments and latest research (prioritizing 2023–2024)
2.1 Metabolism–redox coupling in ER proinsulin export (2024)
A major mechanistic advance is the explicit coupling of cellular metabolism to ER redox homeostasis, which in turn governs proinsulin export and insulin granule biogenesis. Rohli et al. (JCI Insight, 2024-06, https://doi.org/10.1172/jci.insight.178725) report that ER hyperoxidation delays proinsulin export; mitochondrial metabolism supplies reductive donors (e.g., NADPH/thioredoxin flux) that buffer ER redox to support proinsulin trafficking and granule formation. The study notes the folding challenge: a substantial fraction of proinsulin is misfolded and degraded even physiologically, making ER redox control central to insulin biogenesis. (rohli2024ametabolicredox pages 1-2)
2.2 ER calcium homeostasis and proinsulin processing (2024)
Evans-Molina (Diabetes, 2024-03, https://doi.org/10.2337/dbi23-0030) summarizes mechanistic work linking SERCA2/ER calcium to prohormone convertase maturation, ER→Golgi trafficking, and proinsulin processing; SERCA2 deficiency elevates proinsulin-to-insulin ratios and disrupts convertase activity/localization, implicating ER calcium as an upstream regulator of insulin maturation and secretory competence. (evansmolina2024theailingβcell pages 4-5, evansmolina2024theailingβcell pages 1-2)
2.3 Translational reprogramming in chronic hyperglycemia (“glucose toxicity”) (2024)
Cheruiyot et al. (JCI, 2024-11, https://doi.org/10.1172/jci173280) report that sustained hyperglycemia suppresses translation of not only insulin but also mRNAs required for secretory granule formation, exocytosis, and metabolism-coupled secretion, before global translation suppression and prior to ER stress. This identifies a targeted translational circuit contributing to β-cell secretory failure under chronic high glucose. (cheruiyot2024sustainedhyperglycemiaspecifically pages 1-2)
2.4 Phase separation/condensation and granule biogenesis (2023)
Toledo et al. (Protein Science, 2023-05, https://doi.org/10.1002/pro.4649) provide biophysical evidence that proinsulin/insulin can form condensates with the ICA512 RESP18 homology domain, supporting the idea that condensation/phase separation in early secretory compartments may contribute to secretory granule biogenesis, while requiring ER chaperoning to prevent premature condensation that would interfere with correct folding. (toledo2023condensationofthe pages 1-2)
3) Current applications and real-world implementations
3.1 Therapeutic insulin and insulin analogs
Clinical insulin therapy uses engineered formulations spanning rapid/ultra-rapid prandial analogs and long-acting basal analogs. ADA Standards of Care (Diabetes Care, 2024-12, https://doi.org/10.2337/dc24-s009) describe that rapid-acting analogs and ultra-rapid formulations improve postprandial control versus regular human insulin, and basal analogs such as U-300 glargine and degludec may reduce hypoglycemia risk versus U-100 glargine. (elsayed20249.pharmacologicapproaches pages 1-2)
3.2 Diabetes technology: CGM, pumps (CSII), and automated insulin delivery (AID)
The 2024 ADA Standards define diabetes technology to include CGM, insulin pumps, AID (closed-loop) systems, and connected pens, emphasizing that benefit requires education and support; consistent CGM access is important, as interruptions are associated with worsening outcomes, and backup blood glucose monitoring should be maintained. (Diabetes Care, 2024-12, https://doi.org/10.2337/dc24-s007) (elsayed20247.diabetestechnology pages 1-2, elsayed20247.diabetestechnology pages 1-1)
Automated insulin delivery (AID) as real-world implementation:
- AID/closed-loop systems are increasingly used in routine care for type 1 diabetes, and are expanding to other indications. (boughton2024theroleof pages 1-2, kadiyala2024closedloopsystemsrecent pages 1-2)
- AID systems consist of CGM + algorithm + pump, modulating insulin delivery every 5–10 minutes; current commercial systems are hybrid (need meal input). (kadiyala2024closedloopsystemsrecent pages 1-2)
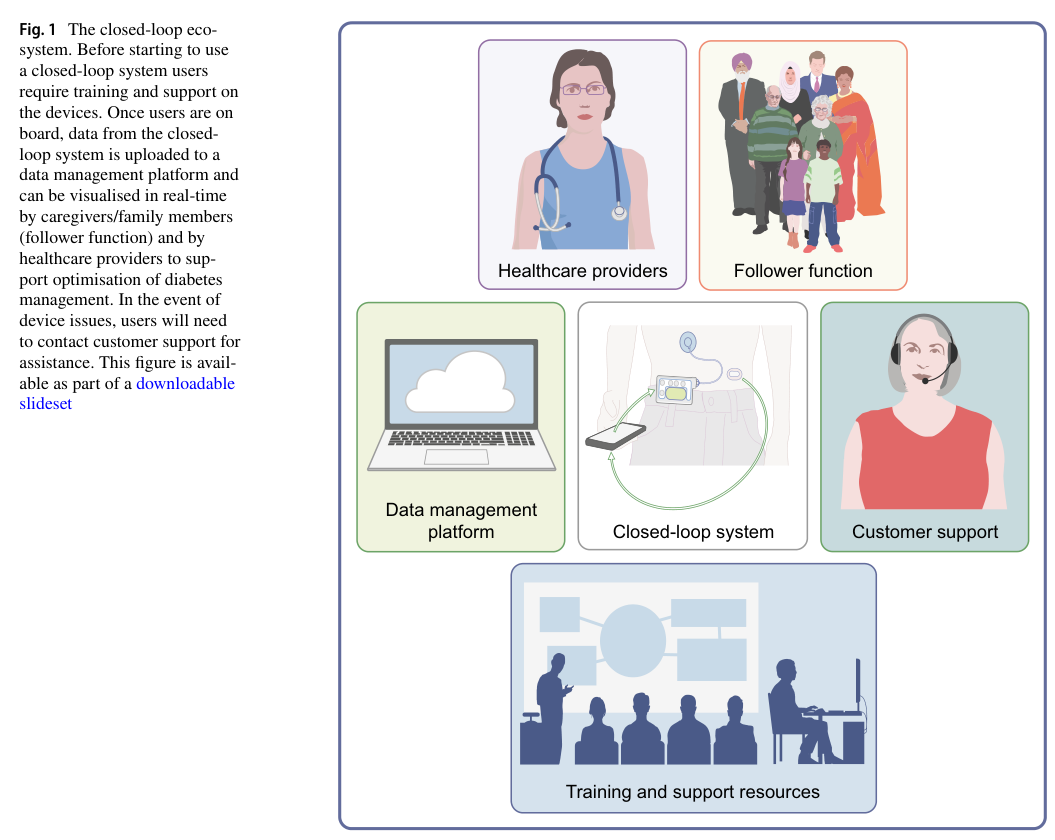
A key implementation theme from expert reviews is that outcomes depend on both device performance and the broader “ecosystem”: training, support, data-sharing, clinic resources, and connectivity reliability. (boughton2024theroleof pages 1-2)
4) Expert opinions and authoritative analyses (2023–2024)
4.1 Technology guidance and clinical standards (ADA & ISPAD, 2024)
- ADA Standards (2024) position early CGM initiation (even at diagnosis) as standard-of-care for most people with type 1 diabetes, and highlight clinician competency/training needs due to device complexity. (elsayed20249.pharmacologicapproaches pages 1-2, elsayed20247.diabetestechnology pages 1-1)
- ISPAD 2024 guidelines (Hormone Research in Paediatrics, 2024-12, https://doi.org/10.1159/000543034) recommend pump therapy for youth regardless of age or baseline HbA1c, emphasize infusion-set failure/DKA risk, and explicitly discuss disparities in access and reimbursement as implementation bottlenecks. (biester2024ispadclinicalpractice pages 2-3)
4.2 Closed-loop expert review perspective (2024)
Boughton & Hovorka (Diabetologia, 2024-05, https://doi.org/10.1007/s00125-024-06165-w) emphasize that real-world outcomes generally mirror trial results, but scaling AID requires addressing the full device-support ecosystem (training, customer support, data systems) and access inequities. (boughton2024theroleof pages 1-2)
5) Recent quantitative statistics and data points (from 2023–2024 sources)
5.1 Glycemic target attainment remains low (implementation gap)
- Registry data in a 2024 closed-loop review report that only 26% of people with type 1 diabetes met HbA1c <7% in 2021–2022, underscoring persistent unmet need. (kadiyala2024closedloopsystemsrecent pages 1-2)
- A 2024 pilot-system dissertation-style source cites Type 1 Diabetes Exchange registry (n=22,697; 2016–2018) target attainment of 17% (youth) and 21% (adults) meeting HbA1c targets. (odabassian2024pilotoutpatientassessment pages 22-26)
5.2 Quantitative benefits of AID/closed-loop systems
- A 2024 Expert Review of Medical Devices paper reports RCT/real-world improvements in time-in-range (70–180 mg/dL) of approximately +6 to +28 percentage points, with HbA1c reductions of approximately 0.1 to 1.4 percentage points (range depends on population/comparator). (kadiyala2024closedloopsystemsrecent pages 1-2)
- A 2024 Diabetologia review reports meta-analyses showing hybrid closed-loop improves time-in-range by approximately +8 to +12 percentage points, reduces hyperglycemia and mean glucose, and does not increase (and may reduce) hypoglycemia. (boughton2024theroleof pages 1-2)
5.3 CGM target metrics used in practice
A 2024 source summarizes widely used CGM targets: TIR >70% (3.9–10.0 mmol/L), time <3.9 mmol/L <4%, and time <3.0 mmol/L <1%. (odabassian2024pilotoutpatientassessment pages 22-26)
5.4 Quantitative biomarker changes reflecting β-cell function/stress (2023 trial)
In SURPASS-1 (Journal of the Endocrine Society, 2023-03, https://doi.org/10.1210/jendso/bvad056), tirzepatide monotherapy at 40 weeks reduced fasting proinsulin by ~49% and reduced proinsulin/C-peptide ratio by ~46–49% vs ~0% change with placebo, consistent with reduced β-cell stress and improved processing. (lee2023tirzepatideasmonotherapy pages 5-6, lee2023tirzepatideasmonotherapy pages 3-4)
Visual evidence: AID/closed-loop ecosystem
The figure below provides a schematic of the closed-loop ecosystem (sensor → algorithm → pump, plus supporting data/training/healthcare infrastructure), reflecting how real-world performance depends on the combined system.
(boughton2024theroleof media 294504cd)
Summary tables (artifacts)
The following tables consolidate the functional annotation and pathway context.
| Process/Function | Key molecular steps | Key molecules/enzymes | Subcellular localization | Evidence/source (short cite with year) |
|---|---|---|---|---|
| Preproinsulin synthesis and ER targeting | INS is translated as a 110-aa preproinsulin; N-terminal signal peptide directs cotranslational translocation into ER, where signal peptide is cleaved to form proinsulin | Signal peptide, SRP/translocon, signal peptidase | Rough ER / ER lumen of pancreatic β-cells | Ayan 2023; Urbaniak 2025 (ayan2023abriefatlas pages 2-3, urbaniak2025molecularpuzzleof pages 2-3) |
| Proinsulin folding and disulfide bond formation | Proinsulin folds in ER and forms three essential disulfide bonds; misfolded molecules are retained/degraded, linking folding efficiency to insulin output | PDI, ER oxidoreductases, chaperones (e.g., BiP/GRP78, GRP94) | ER lumen | Urbaniak 2025; Rohli 2024 (urbaniak2025molecularpuzzleof pages 3-6, rohli2024ametabolicredox pages 1-2) |
| Metabolic support for proinsulin export | Mitochondrial metabolism supplies reductive power needed to maintain ER redox; ER hyperoxidation delays proinsulin export and reduces granule biogenesis | NADPH, thioredoxin system, mitochondrial metabolism, ERO1/PDI network | ER linked to mitochondrial metabolic pathways | Rohli 2024 (rohli2024ametabolicredox pages 1-2) |
| ER stress and trafficking control | Chronic ER Ca2+ defects impair convertase maturation, proinsulin trafficking, and increase proinsulin/insulin ratio; altered ER→Golgi trafficking contributes to β-cell dysfunction | SERCA2, PC1/3, PC2, ER Ca2+ handling machinery | ER, cis-Golgi, intermediate secretory compartments | Evans-Molina 2024 (evansmolina2024theailingβcell pages 1-2, evansmolina2024theailingβcell pages 4-5) |
| Golgi trafficking and granule maturation | Folded proinsulin exits ER, traffics through Golgi/TGN, and is packaged into immature secretory granules for maturation | COPII machinery, Golgi sorting factors, Zn2+, Ca2+ | Golgi, trans-Golgi network, immature granules | Urbaniak 2025; Ayan 2023 (urbaniak2025molecularpuzzleof pages 7-8, ayan2023abriefatlas pages 2-3) |
| Proteolytic processing to mature insulin | Proinsulin is cleaved at B-chain/C-peptide and C-peptide/A-chain junctions; dibasic residues are removed to yield mature disulfide-linked insulin A and B chains plus C-peptide | PC1/3, PC2, carboxypeptidase E/CPE | Immature/maturing secretory granules | Szablewski 2024; Ayan 2023; Urbaniak 2025 (szablewski2024changesincells pages 2-4, ayan2023abriefatlas pages 2-3, urbaniak2025molecularpuzzleof pages 7-8) |
| Secretory granule condensation/biogenesis | Proinsulin has intrinsic self-condensation tendency; co-condensation with ICA512 RESP18HD may help segregate cargo and drive early secretory granule formation | Proinsulin/insulin, ICA512 RESP18HD | Early secretory pathway / nascent secretory granules | Toledo 2023 (toledo2023condensationofthe pages 1-2) |
| Glucose-stimulated insulin secretion (triggering phase) | Glucose metabolism raises ATP, closes KATP channels, depolarizes membrane, opens voltage-gated Ca2+ channels, and triggers exocytosis of readily releasable granules | GLUT1/2, glucokinase, KATP channels, VDCCs, Ca2+ | Plasma membrane, cytosol, docked insulin granules | Dalle 2024; Szablewski 2024 (dalle2024receptorsandsignaling pages 2-4, szablewski2024changesincells pages 2-4) |
| Biphasic/amplified secretion | First phase releases pre-docked granules; second phase recruits reserve granules via metabolic coupling factors and cytoskeletal remodeling | Cdc42, Rac1, NAD(P)H/NADPH, PLC/PKC, cAMP | Cortical actin network, cytoplasm, plasma membrane | Szablewski 2024; Dalle 2024 (szablewski2024changesincells pages 2-4, dalle2024receptorsandsignaling pages 2-4) |
| Translational adaptation and glucose toxicity | Acute glucose enhances translation of proinsulin and secretion-related proteins; sustained hyperglycemia selectively suppresses translation of insulin, granule biogenesis, exocytosis, and coupling-factor mRNAs before global translational collapse | Insulin mRNA 5′UTR regulatory elements, secretion-pathway mRNAs | Cytosol/ribosomes with downstream effects on ER-granule pathway | Cheruiyot 2024 (cheruiyot2024sustainedhyperglycemiaspecifically pages 1-2) |
| Endocrine signaling function after secretion | Mature insulin is released to circulation, dissociates to active monomers, binds insulin receptor, and activates metabolic and mitogenic signaling pathways | Insulin, INSR, IRS proteins, PI3K-AKT, MAPK | Extracellular space; target-cell plasma membrane and cytosol | Ayan 2023; Dalle 2024 (ayan2023abriefatlas pages 2-3, sakaguchi2024theroleof pages 1-3) |
Table: This table summarizes the core functional annotation of human INS/insulin from biosynthesis through secretion and downstream signaling. It integrates recent mechanistic and review evidence across subcellular compartments, highlighting the molecules and enzymes most relevant to precise functional interpretation.
| Step | Key molecules | Cellular location | Main functional outputs | Key evidence (source, year) |
|---|---|---|---|---|
| Ligand binding and receptor activation | Insulin, insulin receptor (INSR) α2β2 heterotetramer, receptor Tyr1158/Tyr1162/Tyr1163 autophosphorylation sites | Plasma membrane | Initiates insulin signaling by activating the receptor’s intrinsic tyrosine kinase and creating docking sites for adaptor proteins | Sakaguchi 2024; Szablewski 2024; Caturano 2024 (sakaguchi2024theroleof pages 1-3, szablewski2024changesincells pages 2-4, caturano2024insulin–heartaxisbridging pages 2-3) |
| Adaptor recruitment | IRS1/IRS2, Shc, PTB/PH domains, NPEpY972 receptor motif | Inner leaflet of plasma membrane / receptor-proximal cytosol | Couples activated INSR to downstream metabolic and mitogenic pathways | Sakaguchi 2024; Szablewski 2024 (sakaguchi2024theroleof pages 1-3, szablewski2024changesincells pages 2-4) |
| PI3K activation and PIP3 generation | IRS proteins, PI3K p85/p110, PIP2, PIP3 | Plasma membrane / cytosolic membrane interface | Establishes the main metabolic signaling branch and recruits PDK1/AKT machinery | Sakaguchi 2024; Szablewski 2024; Caturano 2024 (sakaguchi2024theroleof pages 1-3, szablewski2024changesincells pages 4-6, caturano2024insulin–heartaxisbridging pages 2-3) |
| AKT activation | PIP3, PDK1, AKT/PKB, mTORC2 | Plasma membrane then cytosol | Central integration node for glucose uptake, glycogen synthesis, lipogenesis, protein synthesis, and survival signaling | Sakaguchi 2024; Caturano 2024 (sakaguchi2024theroleof pages 1-3, caturano2024insulin–heartaxisbridging pages 2-3) |
| Glucose transporter trafficking | AKT, GLUT4 vesicles, atypical PKC | Cytosol to plasma membrane in muscle/adipose cells | Increases cellular glucose uptake by translocating GLUT4 to the cell surface | Szablewski 2024; Caturano 2024; Sakaguchi 2024 (szablewski2024changesincells pages 4-6, caturano2024insulin–heartaxisbridging pages 5-7, sakaguchi2024theroleof pages 1-3) |
| Glycogen synthesis control | AKT, GSK3, glycogen synthase | Cytosol | Promotes glycogen synthesis by inhibiting GSK3 and relieving inhibition of glycogen synthase | Szablewski 2024; Caturano 2024; Sakaguchi 2024 (szablewski2024changesincells pages 4-6, caturano2024insulin–heartaxisbridging pages 2-3, sakaguchi2024theroleof pages 1-3) |
| Protein synthesis and anabolic signaling | AKT, TSC2, PRAS40, Rheb, mTORC1, 4E-BP1, p70S6K | Cytosol / lysosomal-mTOR signaling compartments | Stimulates protein synthesis, cell growth, and anabolic metabolism | Caturano 2024; Sakaguchi 2024; Szablewski 2024 (caturano2024insulin–heartaxisbridging pages 2-3, sakaguchi2024theroleof pages 1-3, szablewski2024changesincells pages 4-6) |
| Transcriptional regulation of fasting/feeding programs | AKT, FOXO transcription factors, SREBPs | Cytosol and nucleus | Suppresses FOXO-driven gluconeogenic gene expression and promotes lipogenic/anabolic gene programs during feeding | Sakaguchi 2024; Szablewski 2024 (sakaguchi2024theroleof pages 1-3, szablewski2024changesincells pages 4-6) |
| MAPK mitogenic branch | Shc, Grb2, SOS, Ras, Raf, MEK, ERK/MAPK | Plasma membrane to cytosol to nucleus | Drives growth, proliferation, differentiation, and gene-expression responses distinct from the core metabolic branch | Sakaguchi 2024; Szablewski 2024; Caturano 2024 (sakaguchi2024theroleof pages 1-3, szablewski2024changesincells pages 4-6, caturano2024insulin–heartaxisbridging pages 5-7) |
| Tissue-level physiological outputs | INSR-B (metabolic isoform), PI3K-AKT branch, MAPK branch, eNOS/NO in heart | Whole-body target tissues including liver, muscle, adipose tissue, and heart | Coordinates glucose disposal, reduced hepatic gluconeogenesis, glycogen and lipid storage, protein anabolism, and in cardiomyocytes supports substrate utilization and survival signaling | Szablewski 2024; Caturano 2024 (szablewski2024changesincells pages 4-6, caturano2024insulin–heartaxisbridging pages 2-3) |
| Negative feedback / insulin resistance nodes | Serine-phosphorylated IRS, reduced INSR content/activity, mTORC1-GRB10 feedback | Plasma membrane and cytosol | Weakens IRS–PI3K coupling, blunts GLUT4 translocation and glycogen synthesis, and contributes to insulin resistance | Caturano 2024; Szablewski 2024 (caturano2024insulin–heartaxisbridging pages 5-7, szablewski2024changesincells pages 6-7) |
Table: This table summarizes the major steps of insulin receptor signaling downstream of INS-encoded insulin, linking key molecules and cellular locations to metabolic and mitogenic outputs. It is useful for functional annotation because it connects secreted insulin to its principal target-cell mechanisms and physiological effects.
Conclusions (functional annotation synthesis)
Human INS (P01308) encodes a secreted peptide hormone precursor whose primary function is to produce mature insulin (A/B chains) that is stored in dense-core secretory granules and released in a glucose- and Ca2+-regulated manner from β-cells to maintain systemic metabolic homeostasis. Its biosynthesis is tightly coupled to ER folding capacity, redox state, ER Ca2+ homeostasis, and secretory trafficking; recent 2023–2024 work clarifies how metabolism-driven redox buffering, chronic hyperglycemia-induced translational programs, and cargo condensation mechanisms can regulate or impair insulin production and secretion. (ayan2023abriefatlas pages 2-3, szablewski2024changesincells pages 2-4, rohli2024ametabolicredox pages 1-2, evansmolina2024theailingβcell pages 4-5, cheruiyot2024sustainedhyperglycemiaspecifically pages 1-2, toledo2023condensationofthe pages 1-2)
URLs and publication dates for key 2023–2024 sources cited
- Ayan & Demirci. Current Diabetes Reviews (2023-06). https://doi.org/10.2174/1573399819666220610150342 (ayan2023abriefatlas pages 2-3)
- Aamodt & Powers. J Clin Invest (2023-04). https://doi.org/10.1172/jci169718 (aamodt2023peelingtheonion pages 1-2)
- Toledo et al. Protein Science (2023-05). https://doi.org/10.1002/pro.4649 (toledo2023condensationofthe pages 1-2)
- Lee et al. J Endocrine Society (2023-03). https://doi.org/10.1210/jendso/bvad056 (lee2023tirzepatideasmonotherapy pages 5-6)
- Szablewski. Int J Mol Sci (2024-02). https://doi.org/10.3390/ijms25042397 (szablewski2024changesincells pages 2-4)
- Dalle & Abderrahmani. Cells (2024-07). https://doi.org/10.3390/cells13151244 (dalle2024receptorsandsignaling pages 2-4)
- Sakaguchi. Endocrine Journal (2024-07). https://doi.org/10.1507/endocrj.ej24-0205 (sakaguchi2024theroleof pages 1-3)
- Caturano et al. Int J Mol Sci (2024-07). https://doi.org/10.3390/ijms25158369 (caturano2024insulin–heartaxisbridging pages 2-3)
- Evans-Molina. Diabetes (2024-03). https://doi.org/10.2337/dbi23-0030 (evansmolina2024theailingβcell pages 4-5)
- Rohli et al. JCI Insight (2024-06). https://doi.org/10.1172/jci.insight.178725 (rohli2024ametabolicredox pages 1-2)
- Cheruiyot et al. J Clin Invest (2024-11). https://doi.org/10.1172/jci173280 (cheruiyot2024sustainedhyperglycemiaspecifically pages 1-2)
- Boughton & Hovorka. Diabetologia (2024-05). https://doi.org/10.1007/s00125-024-06165-w (boughton2024theroleof pages 1-2)
- Kadiyala et al. Expert Rev Med Devices (2024-10). https://doi.org/10.1080/17434440.2024.2406901 (kadiyala2024closedloopsystemsrecent pages 1-2)
- ADA Standards: Diabetes Technology. Diabetes Care (2024-12). https://doi.org/10.2337/dc24-s007 (elsayed20247.diabetestechnology pages 1-1)
- ADA Standards: Pharmacologic Approaches. Diabetes Care (2024-12). https://doi.org/10.2337/dc24-s009 (elsayed20249.pharmacologicapproaches pages 1-2)
- ISPAD Guidelines: Insulin Delivery. Hormone Research in Paediatrics (2024-12). https://doi.org/10.1159/000543034 (biester2024ispadclinicalpractice pages 2-3)
References
-
(urbaniak2025molecularpuzzleof pages 3-6): Edyta Urbaniak, Sara Henry, Maciej Lalowski, and Malgorzata Borowiak. Molecular puzzle of insulin: structural assembly pathways and their role in diabetes. Frontiers in Cell and Developmental Biology, Feb 2025. URL: https://doi.org/10.3389/fcell.2025.1502469, doi:10.3389/fcell.2025.1502469. This article has 4 citations.
-
(urbaniak2025molecularpuzzleof pages 2-3): Edyta Urbaniak, Sara Henry, Maciej Lalowski, and Malgorzata Borowiak. Molecular puzzle of insulin: structural assembly pathways and their role in diabetes. Frontiers in Cell and Developmental Biology, Feb 2025. URL: https://doi.org/10.3389/fcell.2025.1502469, doi:10.3389/fcell.2025.1502469. This article has 4 citations.
-
(ayan2023abriefatlas pages 2-3): Esra Ayan and H. Demirci. A brief atlas of insulin. Current diabetes reviews, Jun 2023. URL: https://doi.org/10.2174/1573399819666220610150342, doi:10.2174/1573399819666220610150342. This article has 18 citations.
-
(szablewski2024changesincells pages 2-4): Leszek Szablewski. Changes in cells associated with insulin resistance. International Journal of Molecular Sciences, 25:2397, Feb 2024. URL: https://doi.org/10.3390/ijms25042397, doi:10.3390/ijms25042397. This article has 76 citations.
-
(urbaniak2025molecularpuzzleof pages 7-8): Edyta Urbaniak, Sara Henry, Maciej Lalowski, and Malgorzata Borowiak. Molecular puzzle of insulin: structural assembly pathways and their role in diabetes. Frontiers in Cell and Developmental Biology, Feb 2025. URL: https://doi.org/10.3389/fcell.2025.1502469, doi:10.3389/fcell.2025.1502469. This article has 4 citations.
-
(prefontaine2024investigatingtherelationship pages 8-13): C Préfontaine. Investigating the relationship between insulin processing and senescence in β-cells. Unknown journal, 2024.
-
(rohli2024ametabolicredox pages 1-2): Kristen E. Rohli, Nicole J. Stubbe, Emily M. Walker, Gemma L. Pearson, Scott A. Soleimanpour, and Samuel B. Stephens. A metabolic redox relay supports er proinsulin export in pancreatic islet β cells. JCI Insight, Jun 2024. URL: https://doi.org/10.1172/jci.insight.178725, doi:10.1172/jci.insight.178725. This article has 14 citations and is from a domain leading peer-reviewed journal.
-
(sakaguchi2024theroleof pages 1-3): Masaji Sakaguchi. The role of insulin signaling with foxo and foxk transcription factors. Endocrine Journal, 71:939-944, Jul 2024. URL: https://doi.org/10.1507/endocrj.ej24-0205, doi:10.1507/endocrj.ej24-0205. This article has 12 citations and is from a peer-reviewed journal.
-
(dalle2024receptorsandsignaling pages 2-4): Stéphane Dalle and Amar Abderrahmani. Receptors and signaling pathways controlling beta-cell function and survival as targets for anti-diabetic therapeutic strategies. Cells, 13:1244, Jul 2024. URL: https://doi.org/10.3390/cells13151244, doi:10.3390/cells13151244. This article has 18 citations.
-
(aamodt2023peelingtheonion pages 1-2): Kristie I. Aamodt and Alvin C. Powers. Peeling the onion: another layer in the regulation of insulin secretion. The Journal of clinical investigation, Apr 2023. URL: https://doi.org/10.1172/jci169718, doi:10.1172/jci169718. This article has 3 citations.
-
(szablewski2024changesincells pages 4-6): Leszek Szablewski. Changes in cells associated with insulin resistance. International Journal of Molecular Sciences, 25:2397, Feb 2024. URL: https://doi.org/10.3390/ijms25042397, doi:10.3390/ijms25042397. This article has 76 citations.
-
(caturano2024insulin–heartaxisbridging pages 2-3): Alfredo Caturano, Raffaele Galiero, Erica Vetrano, Celestino Sardu, Luca Rinaldi, Vincenzo Russo, Marcellino Monda, Raffaele Marfella, and Ferdinando Carlo Sasso. Insulin–heart axis: bridging physiology to insulin resistance. International Journal of Molecular Sciences, 25:8369, Jul 2024. URL: https://doi.org/10.3390/ijms25158369, doi:10.3390/ijms25158369. This article has 46 citations.
-
(evansmolina2024theailingβcell pages 4-5): Carmella Evans-Molina. The ailing β-cell in diabetes: insights from a trip to the er: the 2023 outstanding scientific achievement award lecture. Diabetes, 73 4:545-553, Mar 2024. URL: https://doi.org/10.2337/dbi23-0030, doi:10.2337/dbi23-0030. This article has 8 citations and is from a highest quality peer-reviewed journal.
-
(evansmolina2024theailingβcell pages 1-2): Carmella Evans-Molina. The ailing β-cell in diabetes: insights from a trip to the er: the 2023 outstanding scientific achievement award lecture. Diabetes, 73 4:545-553, Mar 2024. URL: https://doi.org/10.2337/dbi23-0030, doi:10.2337/dbi23-0030. This article has 8 citations and is from a highest quality peer-reviewed journal.
-
(cheruiyot2024sustainedhyperglycemiaspecifically pages 1-2): Abigael Cheruiyot, Jennifer Hollister-Lock, Brooke Sullivan, Hui Pan, Jonathan M. Dreyfuss, Susan Bonner-Weir, and Jean E. Schaffer. Sustained hyperglycemia specifically targets translation of mrnas for insulin secretion. The Journal of Clinical Investigation, Nov 2024. URL: https://doi.org/10.1172/jci173280, doi:10.1172/jci173280. This article has 18 citations.
-
(toledo2023condensationofthe pages 1-2): Pamela L. Toledo, Diego S. Vazquez, Alejo R. Gianotti, Milagros B. Abate, Carolin Wegbrod, Juha M. Torkko, Michele Solimena, and Mario R. Ermácora. Condensation of the β‐cell secretory granule luminal cargoes pro/insulin and
ica512 resp18 homology domain. Protein Science, May 2023. URL: https://doi.org/10.1002/pro.4649, doi:10.1002/pro.4649. This article has 9 citations and is from a peer-reviewed journal. -
(elsayed20249.pharmacologicapproaches pages 1-2): Nuha A. ElSayed, G. Aleppo, Raveendhara R. Bannuru, Dennis Bruemmer, B. Collins, Laya Ekhlaspour, Jason L. Gaglia, Marisa E. Hilliard, Eric L. Johnson, K. Khunti, Ildiko Lingvay, Glenn Matfin, Rozalina G. McCoy, M. L. Perry, Scott J. Pilla, Sarit Polsky, P. Prahalad, R. Pratley, Alissa R. Segal, J. Seley, Robert C. Stanton, and Robert A. Gabbay. 9. pharmacologic approaches to glycemic treatment: standards of care in diabetes-2024. Diabetes care, 47 Suppl 1:S158-S178, Dec 2024. URL: https://doi.org/10.2337/dc24-s009, doi:10.2337/dc24-s009. This article has 541 citations and is from a highest quality peer-reviewed journal.
-
(elsayed20247.diabetestechnology pages 1-2): Nuha A. ElSayed, G. Aleppo, Raveendhara R. Bannuru, Dennis Bruemmer, B. Collins, Laya Ekhlaspour, Marisa E. Hilliard, Eric L. Johnson, K. Khunti, Ildiko Lingvay, Glenn Matfin, Rozalina G. McCoy, M. L. Perry, Scott J. Pilla, Sarit Polsky, P. Prahalad, R. Pratley, Alissa R. Segal, J. Seley, Robert C. Stanton, and Robert A. Gabbay. 7. diabetes technology: standards of care in diabetes-2024. Diabetes care, 47 Suppl 1:S126-S144, Dec 2024. URL: https://doi.org/10.2337/dc24-s007, doi:10.2337/dc24-s007. This article has 203 citations and is from a highest quality peer-reviewed journal.
-
(elsayed20247.diabetestechnology pages 1-1): Nuha A. ElSayed, G. Aleppo, Raveendhara R. Bannuru, Dennis Bruemmer, B. Collins, Laya Ekhlaspour, Marisa E. Hilliard, Eric L. Johnson, K. Khunti, Ildiko Lingvay, Glenn Matfin, Rozalina G. McCoy, M. L. Perry, Scott J. Pilla, Sarit Polsky, P. Prahalad, R. Pratley, Alissa R. Segal, J. Seley, Robert C. Stanton, and Robert A. Gabbay. 7. diabetes technology: standards of care in diabetes-2024. Diabetes care, 47 Suppl 1:S126-S144, Dec 2024. URL: https://doi.org/10.2337/dc24-s007, doi:10.2337/dc24-s007. This article has 203 citations and is from a highest quality peer-reviewed journal.
-
(boughton2024theroleof pages 1-2): Charlotte K. Boughton and Roman Hovorka. The role of automated insulin delivery technology in diabetes. Diabetologia, 67:2034-2044, May 2024. URL: https://doi.org/10.1007/s00125-024-06165-w, doi:10.1007/s00125-024-06165-w. This article has 54 citations and is from a highest quality peer-reviewed journal.
-
(kadiyala2024closedloopsystemsrecent pages 1-2): Nithya Kadiyala, Roman Hovorka, and Charlotte K. Boughton. Closed-loop systems: recent advancements and lived experiences. Expert Review of Medical Devices, 21:927-941, Oct 2024. URL: https://doi.org/10.1080/17434440.2024.2406901, doi:10.1080/17434440.2024.2406901. This article has 22 citations and is from a peer-reviewed journal.
-
(biester2024ispadclinicalpractice pages 2-3): Torben Biester, Cari Berget, Charlotte Boughton, Laura Cudizio, Laya Ekhlaspour, Marisa E. Hilliard, Leenatha Reddy, Suzanne Sap Ngo Um, Melissa Schoelwer, Jennifer L. Sherr, and Klemen Dovc. Ispad clinical practice consensus guidelines 2024: diabetes technologies: insulin delivery. Hormone research in paediatrics, 97:1-34, Dec 2024. URL: https://doi.org/10.1159/000543034, doi:10.1159/000543034. This article has 24 citations and is from a peer-reviewed journal.
-
(odabassian2024pilotoutpatientassessment pages 22-26): M Odabassian. Pilot outpatient assessment of a fully closed-loop dual-hormone (insulin and pramlintide) artificial pancreas system. Unknown journal, 2024.
-
(lee2023tirzepatideasmonotherapy pages 5-6): Clare J Lee, Huzhang Mao, Vivian T Thieu, Laura Fernández Landó, and Melissa K Thomas. Tirzepatide as monotherapy improved markers of beta-cell function and insulin sensitivity in type 2 diabetes (surpass-1). Journal of the Endocrine Society, Mar 2023. URL: https://doi.org/10.1210/jendso/bvad056, doi:10.1210/jendso/bvad056. This article has 43 citations and is from a peer-reviewed journal.
-
(lee2023tirzepatideasmonotherapy pages 3-4): Clare J Lee, Huzhang Mao, Vivian T Thieu, Laura Fernández Landó, and Melissa K Thomas. Tirzepatide as monotherapy improved markers of beta-cell function and insulin sensitivity in type 2 diabetes (surpass-1). Journal of the Endocrine Society, Mar 2023. URL: https://doi.org/10.1210/jendso/bvad056, doi:10.1210/jendso/bvad056. This article has 43 citations and is from a peer-reviewed journal.
-
(boughton2024theroleof media 294504cd): Charlotte K. Boughton and Roman Hovorka. The role of automated insulin delivery technology in diabetes. Diabetologia, 67:2034-2044, May 2024. URL: https://doi.org/10.1007/s00125-024-06165-w, doi:10.1007/s00125-024-06165-w. This article has 54 citations and is from a highest quality peer-reviewed journal.
-
(caturano2024insulin–heartaxisbridging pages 5-7): Alfredo Caturano, Raffaele Galiero, Erica Vetrano, Celestino Sardu, Luca Rinaldi, Vincenzo Russo, Marcellino Monda, Raffaele Marfella, and Ferdinando Carlo Sasso. Insulin–heart axis: bridging physiology to insulin resistance. International Journal of Molecular Sciences, 25:8369, Jul 2024. URL: https://doi.org/10.3390/ijms25158369, doi:10.3390/ijms25158369. This article has 46 citations.
-
(szablewski2024changesincells pages 6-7): Leszek Szablewski. Changes in cells associated with insulin resistance. International Journal of Molecular Sciences, 25:2397, Feb 2024. URL: https://doi.org/10.3390/ijms25042397, doi:10.3390/ijms25042397. This article has 76 citations.
Artifacts
Citations
- rohli2024ametabolicredox pages 1-2
- cheruiyot2024sustainedhyperglycemiaspecifically pages 1-2
- toledo2023condensationofthe pages 1-2
- kadiyala2024closedloopsystemsrecent pages 1-2
- boughton2024theroleof pages 1-2
- biester2024ispadclinicalpractice pages 2-3
- odabassian2024pilotoutpatientassessment pages 22-26
- ayan2023abriefatlas pages 2-3
- aamodt2023peelingtheonion pages 1-2
- lee2023tirzepatideasmonotherapy pages 5-6
- szablewski2024changesincells pages 2-4
- dalle2024receptorsandsignaling pages 2-4
- sakaguchi2024theroleof pages 1-3
- urbaniak2025molecularpuzzleof pages 3-6
- urbaniak2025molecularpuzzleof pages 2-3
- urbaniak2025molecularpuzzleof pages 7-8
- prefontaine2024investigatingtherelationship pages 8-13
- szablewski2024changesincells pages 4-6
- lee2023tirzepatideasmonotherapy pages 3-4
- szablewski2024changesincells pages 6-7
- https://doi.org/10.1172/jci.insight.178725
- https://doi.org/10.2337/dbi23-0030
- https://doi.org/10.1172/jci173280
- https://doi.org/10.1002/pro.4649
- https://doi.org/10.2337/dc24-s009
- https://doi.org/10.2337/dc24-s007
- https://doi.org/10.1159/000543034
- https://doi.org/10.1007/s00125-024-06165-w
- https://doi.org/10.1210/jendso/bvad056
- https://doi.org/10.2174/1573399819666220610150342
- https://doi.org/10.1172/jci169718
- https://doi.org/10.3390/ijms25042397
- https://doi.org/10.3390/cells13151244
- https://doi.org/10.1507/endocrj.ej24-0205
- https://doi.org/10.3390/ijms25158369
- https://doi.org/10.1080/17434440.2024.2406901
- https://doi.org/10.3389/fcell.2025.1502469,
- https://doi.org/10.2174/1573399819666220610150342,
- https://doi.org/10.3390/ijms25042397,
- https://doi.org/10.1172/jci.insight.178725,
- https://doi.org/10.1507/endocrj.ej24-0205,
- https://doi.org/10.3390/cells13151244,
- https://doi.org/10.1172/jci169718,
- https://doi.org/10.3390/ijms25158369,
- https://doi.org/10.2337/dbi23-0030,
- https://doi.org/10.1172/jci173280,
- https://doi.org/10.1002/pro.4649,
- https://doi.org/10.2337/dc24-s009,
- https://doi.org/10.2337/dc24-s007,
- https://doi.org/10.1007/s00125-024-06165-w,
- https://doi.org/10.1080/17434440.2024.2406901,
- https://doi.org/10.1159/000543034,
- https://doi.org/10.1210/jendso/bvad056,
Perplexity
(INS-deep-research-perplexity.md)
Perplexity
(INS-deep-research-perplexity.md)Comprehensive Analysis of the Human Insulin Gene (INS) and Its Protein Product: Structure, Synthesis, Signaling, and Metabolic Functions
This comprehensive report synthesizes current knowledge regarding the human insulin gene (INS, UniProt: P01308) and its encoded protein product, one of the most thoroughly characterized and clinically significant hormones in human physiology. Insulin functions as a 51-amino acid peptide hormone composed of two polypeptide chains linked by disulfide bonds, with the primary physiological role of regulating blood glucose homeostasis through complex signaling cascades that influence glucose uptake, glycogen synthesis, lipogenesis, and protein synthesis across multiple tissues[1][2][3]. The hormone is synthesized as a single-chain precursor molecule in pancreatic beta cells and undergoes sophisticated post-translational processing involving multiple proteolytic cleavage events to generate the mature, active form. Beyond its canonical role in glucose metabolism, insulin activates pleiotropic signaling pathways that regulate cell growth, proliferation, and differentiation through distinct intracellular cascades. Recent structural and mechanistic studies have revealed that insulin receptor activation proceeds through a stepwise conformational transition mechanism involving initial ligand binding followed by complex autophosphorylation events, providing molecular insights into both normal physiology and pathological states such as diabetes mellitus. This report provides an integrated analysis of insulin gene organization, protein biosynthesis, three-dimensional structure, receptor interactions, downstream signaling mechanisms, tissue-specific metabolic effects, and the molecular basis of insulin resistance and monogenic forms of diabetes resulting from INS gene mutations.
Protein Structure and Molecular Architecture of Insulin
Primary and Secondary Structure Organization
The mature human insulin protein is composed of 51 amino acid residues organized into two distinct polypeptide chains designated the A-chain and B-chain, which are covalently linked through disulfide bonds rather than a peptide backbone[9][12]. The A-chain consists of 21 amino acid residues, while the B-chain contains 30 residues, with molecular mass of approximately 5808 Daltons[9]. Three disulfide bonds maintain the structural integrity of the mature protein: two interchain disulfide bonds connect the A and B chains at positions A7-B7 and A20-B19, while a third intrachain disulfide bond exists within the A-chain between cysteine residues at positions A6 and A11[9][12]. These disulfide bonds are evolutionarily conserved across the insulin/insulin-like growth factor superfamily and represent a defining structural feature essential for biological activity and stability[2][12].
The three-dimensional structure of insulin reveals a compact, globular configuration with distinct secondary structural elements distributed between the two chains[9]. The A-chain exhibits two α-helical regions spanning residues A1-A8 and A12-A19, which are oriented in an antiparallel arrangement[9]. In contrast, the B-chain contains a central α-helix covering residues B9-B19, flanked on both sides by the interchain disulfide bonds, while two β-sheet structures exist at positions B7-B10 and B20-B23[9]. This architecture creates a highly ordered protein structure stabilized by multiple hydrogen bonding networks and the disulfide bond framework, enabling insulin to form higher-order oligomeric assemblies through protein-protein interactions. The specific amino acid composition and spatial organization of the A and B chains create two major binding epitopes that interact with the insulin receptor in a cooperative manner[16][26].
Zinc Coordination and Oligomeric Assembly
An important but frequently overlooked aspect of insulin structure involves the coordination of divalent zinc ions, which play critical roles in both storage and bioactivity of the hormone. Insulin possesses a histidine residue at position B10 that coordinates zinc ions, enabling the formation of zinc-stabilized hexamers through a toroidal geometry in which two zinc ions coordinate the B10 histidine residues of three insulin dimers[32][35][45]. These zinc-insulin hexamers represent the predominant storage form of insulin within pancreatic beta cell secretory granules, where they exist as water-insoluble crystals that can contain up to 100 millimolar concentrations of insulin[31]. The hexameric assembly is further stabilized by additional interactions involving other histidine residues and glutamic acid residues distributed across multiple insulin molecules within the complex[32][45].
Recent single-molecule studies have revealed that insulin oligomerization occurs through multiple competing assembly pathways depending on the solution concentration and presence of chemical additives[45]. At nanomolar concentrations mimicking physiological secretion, insulin assembly primarily proceeds through monomeric addition to growing oligomeric species, although dimeric and tetrameric addition pathways also exist[45]. At higher concentrations resembling pharmaceutical insulin formulations, the assembly pathway shifts toward dimer-mediated hexamer formation due to increased dimer-to-monomer ratio[45]. The composition of the oligomeric species at any given concentration can be predicted from kinetic rate constants and has important implications for insulin absorption kinetics: monomeric insulin is absorbed rapidly within 5-10 minutes of subcutaneous injection, dimers are absorbed over 20-30 minutes, while hexamers require 1-2 hours for absorption and must first dissociate into monomers to become physiologically active[48].
Evolutionary Conservation and Species Variation
The amino acid sequence of insulin has been highly conserved throughout vertebrate evolution, reflecting strong selective pressure to maintain biological function[2]. Human insulin differs from porcine insulin only at position B30, where human insulin possesses threonine while porcine insulin contains alanine[44]. This minimal difference explains why porcine insulin was historically used to treat insulin-dependent diabetic patients before recombinant human insulin could be produced in large quantities through recombinant DNA technologies[9]. The conservation of the disulfide bond pattern across all vertebrate insulin homologs demonstrates the evolutionary importance of the cysteine residues and their specific geometric arrangement[2]. Some species, such as guinea pig and hagfish, possess insulin molecules lacking histidine at position B10 and therefore do not form zinc-stabilized hexameric crystals; nevertheless, these species maintain normal glucose homeostasis, indicating that while zinc coordination and crystallization enhance insulin storage efficiency, they are not absolutely required for basal insulin function[35].
Biosynthesis and Post-Translational Processing of Insulin
Initial Synthesis and Translocation into the Endoplasmic Reticulum
Insulin synthesis within pancreatic beta cells is initiated from a precursor molecule designated preproinsulin, a single polypeptide chain of 110 amino acids that is the immediate product of translation from insulin mRNA[7][9]. The synthesis begins in the cytosol but is rapidly redirected to the rough endoplasmic reticulum (RER) through recognition of an N-terminal signal peptide comprising 24 hydrophobic residues that constitute the signal sequence[7][44]. This signal peptide directs the nascent preproinsulin chain to the ER membrane through interaction with the signal recognition particle, enabling co-translational translocation into the lumen of the RER[7]. The translocation process is surprisingly inefficient, with recent studies demonstrating that approximately 5-15% of newly synthesized preproinsulin molecules fail to translocate across the ER membrane during the initial 10-minute synthesis period, relying instead on post-translational translocation mechanisms[7]. The signal peptide is cleaved by a signal peptidase located on the lumenal surface of the RER membrane, yielding proinsulin as the immediate product of signal sequence removal[7][12][44].
The efficiency of preproinsulin translocation is influenced by expression levels of the translocon-associated protein (TRAP) complex, whose components are upregulated in response to glucose stimulation in pancreatic beta cell lines[7]. A genetic polymorphism in the TRAPα gene has been associated with both type 2 diabetes and gestational diabetes, suggesting that variations in translocation efficiency may contribute to altered insulin secretion capacity in these disease states[7]. The translocation process must be completed efficiently to prevent accumulation of misfolded preproinsulin in the cytosol, where it would trigger cellular stress responses and potentially undergo degradation through the proteasome pathway[7].
Proinsulin Folding and Disulfide Bond Formation
Following translocation into the ER lumen, preproinsulin undergoes signal peptide cleavage to generate proinsulin, the 86-amino acid single-chain precursor of insulin[12][44]. Proinsulin then embarks upon a complex folding pathway that involves formation of the three disulfide bonds that define the mature insulin structure[2][7]. The ER environment is specifically optimized to facilitate this disulfide bond formation through multiple specialized mechanisms, including high local concentrations of oxidoreductase enzymes such as protein disulfide isomerases and the overall oxidative atmosphere of the ER compartment[7]. The process of proinsulin folding is tightly coupled to disulfide bond pairing, and studies using nuclear magnetic resonance spectroscopy have revealed that the folding pathway proceeds through specific disulfide bonding intermediates rather than forming all three bonds simultaneously[2][7].
Significant evolutionary pressures have shaped the primary sequence of proinsulin to optimize its "foldability," the intrinsic propensity of the protein to fold correctly to the native conformation[7]. These evolutionary pressures are distinct from those governing the ultimate biological activity of the mature insulin molecule, reflecting the different functional requirements of the precursor versus the mature hormone[7]. A pancreatic beta cell is capable of synthesizing approximately 6000 preproinsulin molecules per second, representing an enormous translational load[7]. Even in the basal state without external stimulation, pancreatic beta cells experience significant endoplasmic reticulum stress as measured by markers such as spliced XBP1 protein, suggesting that the high biosynthetic rate of proinsulin pushes the ER protein folding machinery to near maximal capacity[7]. Following feeding, insulin biosynthesis increases further, and in metabolic states characterized by insulin resistance such as pregnancy or obesity, beta cells upregulate proinsulin synthesis even more dramatically to compensate for the reduced peripheral insulin sensitivity[7].
The vast storage pool of older insulin and C-peptide in mature secretory granules is considerably larger than the daily production of newly synthesized insulin, yet evidence indicates a "relative preference" for the release of newly synthesized insulin, particularly during the extended second-phase insulin secretion response[7]. This finding underscores the physiological importance of maintaining adequate proinsulin biosynthesis capacity and the precise regulation of biosynthetic pathway dynamics, as beta cell homeostasis critically depends upon both the biosynthetic pathway that generates new insulin and the granule turnover processes that maintain the releasable insulin pool[7].
Proteolytic Processing and Generation of Mature Insulin
The conversion of proinsulin to mature insulin occurs through the sequential action of specialized endoproteases and an exopeptidase that cleave the connecting C-domain peptide from between the A and B chains[10][12][44]. The C-domain, which links the B-chain to the A-chain in the proinsulin molecule, consists of approximately 30-35 amino acid residues flanked at each end by dibasic residue pairs (Arg-Arg and Lys-Arg)[12][44]. Two prohormone convertases designated PC1/3 (prohormone convertase 1/3) and PC2 (prohormone convertase 2) mediate the primary endoproteolytic cleavage events, with the relative contribution of each enzyme to proinsulin processing having been the subject of considerable investigation[10][12].
Historically, the prevailing model proposed that PC1/3 first cleaves proinsulin at the B-chain-C-peptide junction, generating an intermediate des-64,65 proinsulin product, which is then preferentially cleaved by PC2 at the C-peptide-A-chain junction to yield mature insulin[10][12]. However, recent evidence from human pancreatic islets has challenged this model. Immunofluorescence studies using validated antibodies revealed that PC2 protein is essentially undetectable in human beta cells, with PC2 mRNA signal localized almost exclusively to alpha cells and glucagon-producing cells rather than insulin-producing beta cells[10]. In contrast, PC1/3 is robustly expressed within human beta cells. Biochemical pulse-chase experiments in human islets demonstrated that despite pharmacological and molecular inhibition of PC2 function through multiple approaches including temperature blockade, brefeldin A, chloroquine, and specific protease inhibitors, human beta cells retained full capacity to process proinsulin to mature insulin[10]. Conversely, selective suppression of PC1/3 activity completely blocked proinsulin processing in human beta cells[10].
These findings establish that human proinsulin processing is mediated primarily or exclusively by PC1/3 without requiring PC2, representing a significant revision to the long-standing model based on rodent studies[10]. This species difference has important implications for understanding both normal insulin biosynthesis and disease mechanisms, as mutations affecting PC1/3 but not PC2 would be expected to impair insulin processing in humans, yet potential PC2 mutations might have little impact on insulin production. The PC1/3-mediated cleavage occurs within specialized secretory granules where pH is maintained at 5.0-5.5 by an intrinsic proton pump, creating conditions optimal for both prohormone processing and zinc-insulin crystal formation[12][57].
Following the endoproteolytic cleavage by PC1/3, the dibasic residues remaining at the junctions are removed by a carboxypeptidase B-like exopeptidase activity, generating the final mature insulin A and B chains with the correct terminal amino acid sequences[12][44]. The excised C-peptide, which lacks the dibasic residues, remains in the secretory granule along with insulin but is released into the circulation as an equimolar product of insulin secretion, serving as a useful clinical marker of endogenous insulin secretion[12]. The efficiency of proinsulin conversion to insulin within granules varies with physiological state and metabolic demand, and dysregulation of proinsulin processing has been implicated in the pathogenesis of both type 1 and type 2 diabetes mellitus[10].
Insulin Receptor Structure and Ligand Binding Mechanisms
Insulin Receptor Molecular Architecture
The insulin receptor (IR) is a transmembrane receptor tyrosine kinase that exists as a covalent homodimer composed of two α-subunits and two β-subunits linked by disulfide bonds, forming an (αβ)₂ quaternary structure[3][26]. The α-subunits are entirely extracellular and contain the ligand-binding domains, while the β-subunits are transmembrane proteins composed of an extracellular domain, a single transmembrane helix, and extensive intracellular domains harboring the catalytic tyrosine kinase activity[3][13][16]. The extracellular region of the α-subunit contains multiple functional domains including two leucine-rich globular domains designated L1 (residues 1-120) and L2 (residues 211-428) separated by a cysteine-rich region[13][26]. Additionally, the α-subunit contains two fibronectin type III (FnIII) domains and an α-helical C-terminal domain (αCT) that undergo critical conformational changes during receptor activation[16][26].
Alternative splicing of exon 11 in the insulin receptor gene creates two receptor isoforms, designated IR-A and IR-B, which differ by 12 amino acids in the αCT domain length[16][43]. These isoforms display different biological properties, with IR-B being the predominant form in most metabolic tissues such as liver, muscle, and adipose tissue, while IR-A is more abundant in certain tissues including the brain and is expressed at higher levels during fetal development[26]. The insulin receptor kinase domain shares approximately 84% sequence homology with the insulin-like growth factor-1 receptor (IGF-1R) kinase domain, while the overall sequence homology between IR and IGF-1R ranges from 41-84% depending on the specific domain analyzed[13][26].
Insulin-Binding Sites and Receptor Activation Mechanism
Biochemical and structural studies have established that insulin binds to the insulin receptor through two distinct binding sites designated site-1 and site-2, which have markedly different binding affinities[16][26][46]. Site-1, the primary insulin-binding site, is composed of the L1 domain of one α-subunit and the αCT domain of the adjacent α-subunit; insulin binds to this site with high affinity (Kd approximately 10-30 nanomolar)[16]. Site-2 is a secondary binding site that is composed primarily of the FnIII-1 domain and has much weaker affinity (Kd approximately 400 nanomolar)[16]. Insulin binding to site-2 occurs only at excessive insulin concentrations above 100 nanomolar, which exceeds physiological conditions where circulating insulin levels typically range from low picomolar to nanomolar concentrations[16].
Structural studies have revealed that the insulin receptor exists in distinct conformational states that change dramatically upon insulin binding[16][46]. In the unliganded state, the insulin receptor adopts a "Λ-shaped" auto-inhibited conformation in which the two receptor α-subunits are held in close proximity through extensive inter-protomer interactions that keep the receptor in a signaling-incompetent state[46]. Upon binding of a single insulin molecule to site-1, the receptor undergoes a dramatic conformational transition to a "Γ-shaped" intermediate state, where the FnIII-2 domains move away from the L1 domain, initiating separation of the two receptor protomers[16][46]. This conformational change brings the two intracellular kinase domains into closer spatial proximity, enabling trans-autophosphorylation whereby one kinase domain phosphorylates tyrosine residues on the other kinase domain[43][46].
When multiple insulin molecules bind, the receptor transitions to a fully activated "T-shaped" conformation with both kinase domains held in optimal alignment for maximal kinase activity[16][46]. The structural transitions between the Λ, Γ, and T conformations are mediated by mechanical rearrangement of the α-helical C-terminal (αCT) domains, which are linked by a disulfide bond at position C683[16]. The upshift of one αCT domain from a four-turn α-helix in the apo state to a seven-turn helix in the active state forces the opposite αCT domain downward due to their disulfide linkage, resulting in a coordinated conformational change that propagates from the extracellular binding domains to the transmembrane regions and ultimately to the intracellular kinase domains[16][46].
Insulin Signaling Cascades and Intracellular Signal Transduction
Initial Receptor Autophosphorylation Events
The binding of insulin to its extracellular receptor leads to rapid and sequential autophosphorylation of multiple tyrosine residues distributed across different domains of the intracellular region[43]. The first step in insulin receptor activation involves trans-autophosphorylation of tyrosine residues in three distinct intracellular regions: the juxtamembrane (JM) domain, the kinase domain activation loop, and the C-terminal (CT) domain[43]. Specifically, three tyrosine residues in the kinase domain activation loop (Tyr1146, Tyr1150, and Tyr1151 using IR isoform A numbering) are phosphorylated in a sequential manner, with phosphorylation of Tyr1150 occurring first, followed by Tyr1148 and Tyr1151[43]. These phosphorylation events are critical because in the inactive receptor, the unphosphorylated activation loop is inserted into the catalytic site of the kinase domain, physically blocking substrate and ATP from accessing the active site[43]. Following insulin-binding-induced phosphorylation, the activated loop is released from the catalytic site, dramatically increasing kinase activity by permitting substrate and ATP to interact with the kinase[43][46].
Additional autophosphorylation events occur at Tyr953 and Tyr960 in the JM domain, and at Tyr1316 and Tyr1322 in the CT domain[43][46]. Among these sites, the phosphorylated Tyr960 (designated pY960) is of particular functional importance, as it functions as an NPXY motif that serves as a docking site for the phosphotyrosine-binding (PTB) domains of insulin receptor substrates (IRS) proteins and Src homology 2 domain-containing transforming protein (SHC)[43][46]. This phosphorylation creates a recruitment platform for downstream signaling effectors, allowing direct physical interaction between the phosphorylated insulin receptor and key adaptor proteins that initiate the downstream signaling cascades[43].
The Phosphatidylinositol 3-Kinase and AKT Signaling Pathway
The phosphatidylinositol 3-kinase (PI3K) pathway represents the primary route through which insulin exerts its metabolic effects, particularly regarding glucose homeostasis, glycogen synthesis, and protein synthesis[3][6][40]. The binding of phosphorylated IRS proteins (IRS-1 and IRS-2 being the predominant forms in most tissues) to the activated insulin receptor leads to phosphorylation of multiple tyrosine residues within the IRS proteins by the receptor kinase[3][6]. These phosphorylated IRS tyrosine residues then serve as docking sites for the regulatory subunit (p85) of PI3K, which contains SH2 domains that specifically recognize phosphotyrosine residues in the correct sequence context[3][6].
Recruitment of PI3K to the membrane leads to activation of the catalytic subunit (p110), which phosphorylates the lipid phosphatidylinositol 4,5-bisphosphate (PIP2) on the inner leaflet of the plasma membrane to generate phosphatidylinositol 3,4,5-trisphosphate (PIP3)[3][6]. The accumulation of PIP3 at the plasma membrane creates a docking site for proteins containing pleckstrin homology (PH) domains, including phosphoinositide-dependent protein kinase-1 (PDK1) and the serine/threonine kinase AKT (also known as protein kinase B or PKB)[3][6][40]. PDK1 phosphorylates and activates AKT at threonine 308 within the activation loop, while additional phosphorylation of AKT at serine 473 in the C-terminal hydrophobic motif by mTORC2 complex leads to maximal AKT activation[40].
Once activated, AKT phosphorylates numerous downstream substrates that collectively mediate the metabolic effects of insulin[40]. These substrates include glycogen synthase kinase-3 (GSK3), which is phosphorylated and inactivated, thereby allowing dephosphorylated and active glycogen synthase to catalyze glycogen synthesis[40]. AKT also phosphorylates and activates mTOR complex 1 (mTORC1), which stimulates protein synthesis through phosphorylation of ribosomal protein S6 kinase (S6K) and eIF4E-binding protein 1 (4E-BP1), relieving translational repression[40]. Additionally, AKT phosphorylates and inactivates forkhead box protein O1 (FoxO1), a transcription factor that normally activates genes involved in gluconeogenesis; phosphorylation of FoxO1 by AKT causes its cytoplasmic retention and proteasomal degradation, thereby suppressing hepatic glucose production[40][38].
The Mitogen-Activated Protein Kinase and ERK1/2 Signaling Pathway
In parallel with the PI3K/AKT pathway, insulin also activates the mitogen-activated protein kinase (MAPK) pathway primarily through interaction of the SHC adaptor protein with the phosphorylated insulin receptor[3][6]. SHC binding to the phosphorylated receptor leads to SHC phosphorylation by the receptor kinase, which then recruits the growth factor receptor-bound protein 2 (Grb2) adaptor protein in complex with the guanyl nucleotide exchange factor SOS[3][6]. This complex promotes GDP/GTP exchange on the small G protein Ras, leading to Ras activation and consequent activation of the Raf serine/threonine kinase[6]. Raf then phosphorylates and activates MEK1/2 (MAPK/ERK kinase), which phosphorylates and activates the extracellular signal-regulated kinases ERK1 and ERK2[6][40].
The MAPK pathway mediated by ERK1/2 has distinct biological effects compared to the PI3K/AKT pathway, with the MAPK pathway playing a more prominent role in the regulation of gene expression and cell proliferation rather than acute metabolic effects[3][6][40]. Phosphorylated ERK1/2 translocates to the nucleus and phosphorylates numerous transcription factors and chromatin regulators that alter the expression of genes involved in cell growth, differentiation, and proliferation[6]. The differential activation of the PI3K and MAPK pathways may depend on insulin concentration and the duration of receptor activation, with sustained insulin signaling producing more pronounced MAPK pathway activation[6].
Metabolic Functions of Insulin in Glucose Homeostasis
Stimulation of Glucose Uptake in Skeletal Muscle and Adipose Tissue
The most physiologically important function of insulin is the stimulation of glucose uptake from the bloodstream into insulin-responsive tissues, particularly skeletal muscle and adipose tissue, which together account for the majority of postprandial glucose disposal[5][8][22]. This metabolic action is mediated through the insulin-dependent translocation of glucose transporter type-4 (GLUT4) from intracellular membrane compartments to the plasma membrane, which is a critical control point for glucose homeostasis[8][19][22]. In the basal state without insulin stimulation, approximately 90% of GLUT4 resides in specialized intracellular vesicles and recycling endosomes, with only about 5% located at the plasma membrane[6][19][22]. Upon insulin stimulation, GLUT4-containing vesicles rapidly translocate to the plasma membrane, increasing cell surface GLUT4 by 2-3 fold and dramatically enhancing glucose transport capacity[6][19][22].
The GLUT4 translocation process is initiated by the PI3K/AKT signaling pathway downstream of insulin receptor activation[6][19][22]. Following AKT activation, the serine/threonine kinase AS160 (also known as TBC1D4) is phosphorylated by AKT, leading to its release from GLUT4-containing storage vesicles[22]. This phosphorylation event activates the RabGAP activity of AS160, converting Rab small GTPases to their inactive GDP-bound state and enabling GLUT4 vesicle mobilization[22]. Additionally, phosphorylation of TBC1D1 by AKT promotes GLUT4 trafficking through effects on the exocyst complex and actin cytoskeleton dynamics[22]. The coordinated action of these signaling molecules results in translocation of GLUT4 to the plasma membrane, t-tubules in muscle, and sarcolemmal membranes, where GLUT4 can mediate glucose transport[8][22].
The translocated GLUT4 facilitates glucose entry through ATP-independent facilitative diffusion with a Michaelis constant (Km) of approximately 5 millimolar, enabling steep gradients for rapid glucose clearance at physiological glucose concentrations[8][19][22]. Once inside the cell, glucose is phosphorylated by hexokinase to glucose-6-phosphate, which is then retained in the cell and utilized for glycolysis or stored as glycogen. In skeletal muscle, approximately 80% of postprandial glucose uptake is mediated through this GLUT4-dependent insulin-stimulated mechanism, underscoring the critical importance of this process for maintaining blood glucose homeostasis[8][22].
Hepatic Glucose Metabolism and Suppression of Hepatic Glucose Output
The liver plays a critical role in maintaining glucose homeostasis through its capacity to produce endogenous glucose via both glycogenolysis (breakdown of stored glycogen) and gluconeogenesis (synthesis of new glucose from precursors such as lactate, glycerol, and amino acids)[21][24]. Insulin potently suppresses hepatic glucose production through both direct actions on the liver and indirect actions mediated by effects on adipose tissue, muscle, and central nervous system signaling[21][24]. Direct hepatic insulin effects occur through insulin receptor activation in hepatocytes, leading to suppression of glycogenolysis and gluconeogenesis through multiple mechanisms[21][24].
Insulin rapidly inhibits hepatic glycogenolysis through mechanisms involving protein phosphatase-1 (PP1), which is activated through insulin-mediated suppression of glycogen phosphorylase kinase, thereby preventing phosphorylation of glycogen phosphorylase and allowing its inactivation[15][21]. Simultaneously, insulin activates glycogen synthase through PP1-mediated dephosphorylation, promoting the synthesis of new glycogen from glucose-6-phosphate[15][21]. These complementary effects result in a rapid net transition from glucose output to glucose storage in the liver in response to rising postprandial insulin levels[15][21].
The suppression of hepatic gluconeogenesis by insulin involves inhibition of the transcription factors CREB and FoxO1, which normally activate the expression of the rate-limiting gluconeogenic enzymes phosphoenolpyruvate carboxylase (PEPCK) and glucose-6-phosphatase[15][38][41]. The AKT-mediated phosphorylation and inactivation of FoxO1, combined with reduced CREB activity following insulin treatment, leads to decreased expression of gluconeogenic enzymes and reduced hepatic glucose production[15][38][41]. Additionally, an important indirect mechanism through which insulin suppresses hepatic glucose production involves inhibition of adipose tissue lipolysis, leading to decreased circulating free fatty acid and glycerol concentrations[14][21][24]. Since fatty acid oxidation in hepatocytes provides ATP and acetyl-CoA that drive gluconeogenesis, the reduction in hepatic fatty acid oxidation resulting from decreased NEFA availability represents a dominant mechanism by which systemic insulin indirectly suppresses hepatic glucose production[14][24].
Lipid and Protein Metabolism Regulated by Insulin
Stimulation of Fatty Acid and Triglyceride Synthesis
Insulin is a potent anabolic hormone that stimulates fatty acid synthesis and triglyceride storage in adipose tissue and liver through multiple transcriptional and post-translational mechanisms[15][18][50]. The key transcription factors mediating insulin-induced lipogenic gene expression are the sterol regulatory element binding proteins (SREBPs), particularly SREBP-1c in lipogenic tissues, along with upstream stimulatory factors (USF) and liver X receptors (LXRα)[18][50][53]. Insulin stimulates SREBP-1c expression through the mTOR pathway in a manner that does not require S6K1, indicating activation of mTOR substrates distinct from S6K1[18][50][53]. Upon insulin stimulation, SREBP-1c undergoes proteolytic activation in the Golgi apparatus, releasing the transcriptionally active N-terminal fragment that translocates to the nucleus and activates transcription of genes encoding enzymes required for fatty acid synthesis including fatty acid synthase (FAS), acetyl-CoA carboxylase (ACC), stearoyl-CoA desaturase (SCD), and glycerol-3-phosphate acyltransferase[18][50][53].
The induced fatty acid synthesized in response to insulin is then esterified into triglycerides through sequential esterification reactions with glycerol-3-phosphate, generating triglyceride molecules that accumulate in the adipose tissue as energy storage or are exported from the liver as circulating VLDL particles[18][50]. This process is facilitated by insulin-induced suppression of the AMP-activated protein kinase (AMPK), which normally inhibits mTORC1 and acetyl-CoA carboxylase; suppression of AMPK by elevated glucose and insulin allows maximal activation of acetyl-CoA carboxylase and fatty acid synthesis[18][50].
Inhibition of Lipolysis and Regulation of Lipid Mobilization
Reciprocally, insulin potently inhibits lipolysis in adipose tissue through inactivation of hormone-sensitive lipase (HSL) via a PKA-independent mechanism[15]. Insulin-activated AKT phosphorylates and inactivates phosphodiesterase 3B (PDE3B), reducing the degradation of cAMP and preventing PKA activation[15]. Additionally, insulin signaling leads to dephosphorylation of HSL, which suppresses its catalytic activity and prevents the hydrolysis of stored triglycerides into free fatty acids and glycerol[15]. The net result is decreased adipose tissue lipolysis and reduced circulating free fatty acid levels, diminishing the substrate for hepatic ketone body synthesis and gluconeogenesis, while also allowing increased glucose uptake and lipid storage in adipose tissue in the fed state[15].
Effects on Protein Metabolism
Insulin is also a powerful regulator of protein turnover, stimulating protein synthesis while simultaneously inhibiting protein degradation, thereby promoting net protein anabolism[20][23]. Insulin stimulates protein synthesis through multiple mechanisms including increased cellular uptake of amino acids through upregulation of amino acid transporters, particularly for branched-chain amino acids (BCAAs) such as leucine[20]. Leucine is a particularly important signal for mTORC1 activation and protein synthesis initiation through its capacity to activate the Rag family of GTPases that recruit mTORC1 to the lysosomal membrane[20][23]. Following insulin stimulation and increased leucine availability, mTORC1 phosphorylates S6K1 and 4E-BP1, leading to translation initiation factor phosphorylation and enhanced translation of mRNA molecules, particularly those encoding protein synthesis machinery and growth-promoting proteins[20][23].
In contrast, insulin suppresses hepatic protein degradation through inhibition of the ATP-ubiquitin-dependent proteasome pathway and ATP-independent lysosomal proteases[15][20]. The mechanism involves AKT-mediated phosphorylation and inactivation of FoxO transcription factors, which normally activate genes encoding ubiquitin ligases and other proteolytic machinery[15][20]. In skeletal muscle, insulin stimulates protein synthesis and simultaneously inhibits protein degradation through effects on the same signaling pathways, promoting net muscle protein accumulation[20].
Tissue-Specific Insulin Actions and the Metabolic Effects Across Organ Systems
Central Nervous System Regulation of Energy Metabolism
Beyond its peripheral metabolic actions, insulin also acts within the central nervous system to regulate appetite, feeding behavior, and overall energy expenditure[39][42]. Insulin receptors are widely distributed throughout the hypothalamus and other brain regions involved in energy homeostasis regulation[42]. In the healthy metabolic state, insulin acts in an anorexigenic fashion, reducing appetite and food intake through mechanisms involving PI3K/AKT signaling in the hypothalamus and interconnected brain regions[39][42]. Chronic overfeeding with high-caloric diets rich in sugar and saturated fat leads to development of brain insulin resistance characterized by blunted responsiveness of feeding-related brain regions to insulin signaling despite persistent peripheral insulin resistance[39].
Recent studies have demonstrated that even short-term consumption of high-caloric diet (8 weeks of supplemental high-caloric foods in addition to regular diet) produces prolonged alterations in brain insulin responsiveness in the absence of significant changes in body weight or peripheral insulin sensitivity[39]. These alterations are particularly evident in the reward-processing brain regions including the midbrain and insular cortex, where augmented insulin responsiveness correlates with increased liver fat content[39]. Notably, the diminished insulin responsiveness in the hippocampus and fusiform gyrus outlasts the duration of high-caloric diet consumption, suggesting that diet-induced changes in brain insulin responsiveness may represent a precursor to the development of systemic insulin resistance[39].
Hepatic Lipid Accumulation and Multiorgan Insulin Resistance
The accumulation of intrahepatic triglycerides (IHTG) represents a central feature linking insulin resistance across multiple tissues[14]. In nondiabetic obese subjects, IHTG content is the best independent predictor of insulin action across liver, skeletal muscle, and adipose tissue, accounting for 34-44% of the variance in tissue-specific insulin sensitivity independent of body mass index and total percent body fat[14]. The association between elevated IHTG content and impaired hepatic insulin sensitivity, as reflected in reduced suppression of endogenous glucose production, involves multiple mechanisms including competition of lipid-derived signals (such as diacylglycerol and ceramides) with glucose-derived signals for IRS-1 binding capacity at the insulin receptor[14].
This metabolic crosstalk between hepatic lipid content and systemic insulin sensitivity illustrates the integrated nature of metabolic regulation, whereby defects in one tissue's insulin signaling capacity propagate to impair insulin action in other tissues through effects on circulating nutrient and lipid signals[14]. In this context, nonalcoholic fatty liver disease should be conceptualized not as an isolated hepatic pathology but rather as a manifestation of multiorgan insulin resistance affecting liver, muscle, and adipose tissue simultaneously[14].
Genetic Variants and Monogenic Forms of Diabetes Associated with INS Gene Mutations
Structural Consequences of Proinsulin Mutations
Dominant mutations in the insulin gene (INS) cause monogenic syndromes of diabetes mellitus, most prominently including permanent neonatal-onset diabetes (PNDM) due to toxic misfolding of proinsulin variants[12][27][30]. Seven distinct heterozygous mutations in the INS gene have been identified as causes of permanent neonatal diabetes, with the mutations resulting in defects in insulin protein folding and secretion that lead to endoplasmic reticulum stress and apoptosis of pancreatic beta cells[30]. When these mutant proinsulins are expressed in cell culture systems, they demonstrate impaired folding and secretion compared to wild-type proinsulin, and expression of the mutant forms is associated with increased Grp78 protein expression and XBP1 mRNA splicing, both markers of endoplasmic reticulum stress response[30].
The molecular mechanism by which proinsulin mutations cause diabetes involves generation of misfolded protein intermediates that accumulate in the ER lumen, triggering the unfolded protein response (UPR) and ultimately leading to beta cell apoptosis[30]. The critical disulfide bonds that stabilize insulin structure are often disrupted by amino acid substitutions that affect cysteine residues, and studies in the Akita and Munich mouse models that harbor cysteine mutations in the insulin gene have demonstrated that alterations in proinsulin disulfide bonding patterns severely impair insulin production and lead to diabetes[7][30].
Genetic Imprinting Effects on Neonatal Diabetes Severity
An unusual feature of monogenic INS mutations is that the disease phenotype is affected by whether the mutated allele is inherited from the mother or father, reflecting epigenetic phenomena known as genomic imprinting[27]. The INS gene locus undergoes parent-of-origin-specific methylation, resulting in selective silencing of either the maternal or paternal allele depending on its origin[27]. In the case of maternally inherited INS mutations, the normal paternal INS allele is expressed, whereas in paternally inherited mutations, the maternal INS allele is silenced, resulting in functional haploinsufficiency where only the single mutated paternal allele is expressed[27]. This results in considerably more severe phenotypes for paternally inherited mutations: children inheriting mutated INS alleles from their fathers develop neonatal diabetes at a median age of 2.5 months, while affected parents typically did not manifest diabetes until 84 months of age, demonstrating markedly earlier disease onset in children[27].
In contrast, the difference in age at diagnosis between children and parents was not statistically significant for maternally transmitted mutations, as the imprinting pattern ensures that the normal maternal allele is silenced regardless of whether the child inherits the mutation[27]. These imprinting-related differences in disease severity highlight the importance of allele dosage effects in the pathogenesis of monogenic diabetes, with the complete absence of functional insulin production (as occurs with paternal transmission) producing more severe neonatal diabetes than the situation in which a normal allele is functional[27].
Conclusion: Integration of Insulin Biology into a Comprehensive Framework
The insulin gene (INS) and its protein product represent one of the most thoroughly characterized and physiologically critical components of human metabolism. Insulin functions as a multifaceted signaling hormone that coordinates the anabolic and catabolic processes of the body in response to nutrient availability, serving as the primary signal of the fed state and governing energy storage, utilization, and expenditure[2][5][15]. The sophisticated mechanisms governing insulin biosynthesis, from preproinsulin synthesis and translocation through complex proteolytic processing generating mature insulin, have been refined through evolution to ensure reliable production of the massive quantities of insulin required for metabolic homeostasis[7][2].
The insulin receptor and its associated signaling cascade represent a model system for understanding receptor tyrosine kinase activation and signal transduction, with recent structural studies revealing the dynamic conformational transitions that convert the inactive receptor to a fully activated signaling-competent state[16][43][46]. The divergent downstream signaling pathways—particularly the PI3K/AKT and MAPK/ERK cascades—enable insulin to coordinately regulate acute metabolic processes such as glucose uptake and glycogen synthesis while simultaneously controlling long-term gene expression and cellular proliferation[3][6][40]. The tissue-specific effects of insulin reflect the differential expression of signaling proteins, receptor isoforms, and downstream effectors in distinct tissue compartments, enabling coordinate regulation of glucose utilization in muscle, hepatic glucose production and lipogenesis in liver, and lipid storage and NEFA release in adipose tissue[15][22][24].
Dysregulation of insulin secretion, insulin receptor signaling, or the post-receptor signaling cascades leads to the spectrum of metabolic diseases collectively termed diabetes mellitus, encompassing both type 1 diabetes resulting from loss of insulin-producing beta cells and type 2 diabetes characterized by insulin resistance and eventual beta cell failure[2][5][6]. Monogenic forms of diabetes resulting from INS gene mutations demonstrate the critical importance of insulin structure and production for maintaining beta cell viability and insulin secretion capacity[27][30]. Understanding the precise molecular mechanisms governing insulin synthesis, secretion, receptor binding, and intracellular signaling has opened new therapeutic avenues for diabetes treatment, including development of insulin receptor sensitizers, GLP-1 receptor agonists that enhance endogenous insulin secretion, and novel formulations of recombinant insulin with improved pharmacokinetics[5][48].
The complexity and elegance of insulin's mechanisms of action—from the intricate biosynthetic pathway that generates the hormone in pancreatic beta cells, through the stepwise conformational transitions of its receptor, to the multiple interconnected signaling cascades that coordinate metabolic responses across tissues—exemplify the sophistication of human physiology and provide a framework for understanding both normal metabolic homeostasis and the molecular basis of metabolic disease. Continued investigation of insulin biology at the molecular, cellular, and systems levels remains essential for developing improved therapeutic strategies for diabetes and obesity, which represent major global health challenges affecting over 450 million individuals worldwide[31].
Citations
- https://www.uniprot.org/uniprotkb/P01308/entry
- https://pubmed.ncbi.nlm.nih.gov/21204007/
- https://www.ncbi.nlm.nih.gov/books/NBK378978/
- https://www.ncbi.nlm.nih.gov/gene/3630
- https://www.cdc.gov/diabetes/about/insulin-resistance-type-2-diabetes.html
- https://www.frontiersin.org/journals/endocrinology/articles/10.3389/fendo.2023.1226655/full
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6463291/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2838661/
- https://en.wikipedia.org/wiki/Insulin
- https://diabetesjournals.org/diabetes/article/69/7/1451/39562/Revisiting-Proinsulin-Processing-Evidence-That
- https://febs.onlinelibrary.wiley.com/doi/full/10.1046/j.1432-1327.1999.00043.x
- https://www.ncbi.nlm.nih.gov/books/NBK279029/
- https://www.ncbi.nlm.nih.gov/books/NBK6192/
- https://pubmed.ncbi.nlm.nih.gov/18355813/
- https://www.ncbi.nlm.nih.gov/books/NBK525983/
- https://www.nature.com/articles/s12276-023-01101-1
- https://physoc.onlinelibrary.wiley.com/doi/10.14814/phy2.14607
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3092640/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7672939/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10780923/
- https://diabetesjournals.org/diabetes/article/55/Supplement_2/S65/12042/The-Inhibitory-Effects-of-Insulin-on-Hepatic
- https://www.ncbi.nlm.nih.gov/books/NBK537322/
- https://diabetesjournals.org/diabetes/article/21/Supplement_2/447/8663/Effect-of-Insulin-on-Protein-Synthesis
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5358484/
- https://diabetesjournals.org/diabetes/article/65/4/981/35131/Glucose-Induces-Mouse-Cell-Proliferation-via-IRS2
- https://diabetesjournals.org/care/article/34/1/e9/27610/Paternally-Inherited-Proinsulin-Mutations-May
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1951771/
- https://www.nature.com/articles/ncomms14892
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2350430/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5866358/
- https://pubs.acs.org/doi/10.1021/acs.cgd.1c01463
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9450567/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5710702/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2736467/
- https://diabetesjournals.org/diabetes/article/73/3/412/153920/High-Doses-of-Exogenous-Glucagon-Stimulate-Insulin
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7981362/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4133859/
- https://www.nature.com/articles/s42255-025-01226-9
- https://www.cellsignal.com/pathways/insulin-receptor-signaling-pathway
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8233149/
- https://pubmed.ncbi.nlm.nih.gov/16933179/
- https://www.nature.com/articles/s42003-022-04386-6
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10398885/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7716884/
- https://www.nature.com/articles/s41467-022-29722-6
- https://www.nature.com/articles/s41419-023-06154-8
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9298385/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5923433/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5783513/
- https://www.nature.com/articles/s41467-023-41306-6
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4020243/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2276387/
- https://www.nature.com/articles/s42255-023-00964-y
- https://journals.plos.org/plosone/article?id=10.1371%2Fjournal.pone.0036989