Deep Research
Falcon
(PIWIL1-deep-research-falcon.md)
Falcon
(PIWIL1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Human PIWIL1 (HIWI) Functional Annotation (UniProt Q96J94)
1) Target verification (gene/protein identity)
The target described by UniProt accession Q96J94 corresponds to human PIWIL1 (gene symbol PIWIL1, synonym HIWI), a PIWI-subfamily Argonaute protein that binds PIWI-interacting RNAs (piRNAs) and functions in small-RNA-guided gene silencing pathways. Recent authoritative reviews consistently describe PIWIL1 as a PAZ–MID–PIWI domain protein with RNase H-like “slicer” endonuclease activity in the PIWI domain, indicating that PIWIL1’s primary biochemical activity is piRNA-guided cleavage of RNA targets, not classical metabolism. (Zhang et al., 2023-03, Molecular Cancer, https://doi.org/10.1186/s12943-023-01749-3) (zhang2023theepigeneticregulatory pages 1-2)
2) Key concepts and current understanding
2.1 Definitions: PIWI proteins and piRNAs
piRNAs are a class of small noncoding RNAs (commonly ~24–32 nt) that bind PIWI proteins to form effector ribonucleoprotein complexes (often termed piRISC) that suppress transposable elements and regulate gene expression, particularly in the germline. The PAZ and MID domains anchor the piRNA 3′ and 5′ ends, while the PIWI domain is responsible for endonucleolytic cleavage of complementary RNA targets. (Patel et al., 2024-12, Frontiers in Cell and Developmental Biology, https://doi.org/10.3389/fcell.2024.1495035) (patel2024somaticpirnaand pages 1-2)
2.2 Modular view of piRNA-guided silencing (how PIWIL1 fits)
A 2023 synthesis proposes a modular architecture for piRNA systems: (i) primary piRNA biogenesis/loading (mitochondria-associated cleavage of long precursors to generate 5′-monophosphorylated fragments that load into PIWI proteins), (ii) slicer-mediated post-transcriptional silencing (cytoplasmic cleavage and decay of target RNAs), (iii) secondary amplification (“ping–pong”) in some systems, and (iv) nuclear transcriptional silencing via recruitment of chromatin/DNA modifiers. (Loubalova et al., 2023-09, Mobile DNA, https://doi.org/10.1186/s13100-023-00298-2) (loubalova2023themesandvariations pages 9-10, loubalova2023themesandvariations pages 2-4)
Within this framework, PIWIL1 (mouse ortholog MIWI) is described as being primarily loaded with primary piRNAs and acting mainly in the cytoplasm to degrade target transcripts, with relatively limited evidence for robust participation in ping–pong amplification compared with other PIWI paralogs (e.g., PIWIL2/MILI). (Loubalova et al., 2023-09, Mobile DNA, https://doi.org/10.1186/s13100-023-00298-2) (loubalova2023themesandvariations pages 4-6, loubalova2023themesandvariations pages 6-7)
2.3 Catalytic activity and substrate specificity (functional reaction)
PIWIL1’s catalytic mechanism is small-RNA-guided endonucleolytic cleavage of RNA (a “slicer” reaction), where the substrate is RNA that base-pairs with the bound piRNA guide. Reviews emphasize the PIWI domain’s RNase H-like cleavage function, enabling PIWIL1–piRNA complexes to cut target RNAs (including transposon-derived transcripts and potentially other RNAs depending on context). (Zhang et al., 2023-03, Molecular Cancer, https://doi.org/10.1186/s12943-023-01749-3) (zhang2023theepigeneticregulatory pages 1-2)
3) Pathways, processes, and subcellular localization
3.1 Germline: spermatogenesis and transposon control
A large human testis biopsy study (222 biopsies) directly supports PIWIL1 as a germ-cell-enriched factor and places it in the context of LINE-1 (L1) transposon regulation. The authors observed coexpression and co-localization of PIWIL1 with the piRNA 3′-end methyltransferase HENMT1 in pachytene spermatocytes and spermatids, consistent with PIWIL1 acting during the pachytene/spermatid stages of spermatogenesis. (Hempfling et al., 2017-10, Reproduction, https://doi.org/10.1530/rep-16-0586) (hempfling2017expressionpatternsof pages 1-2, hempfling2017expressionpatternsof pages 3-4)
Subcellularly in human germ cells, PIWIL1 protein first appears in the cytoplasm of late pachytene spermatocytes, persists into spermatids, and is reported to concentrate in structures consistent with mitochondrial cement/nuage in pachytene cells and the chromatoid body in spermatids—compartments long associated with small-RNA processing and post-transcriptional regulation. (Hempfling et al., 2017-10, Reproduction, https://doi.org/10.1530/rep-16-0586) (hempfling2017expressionpatternsof pages 3-4)
3.2 Human evidence linking PIWIL1/piRNA pathway disruption to infertility
Recent human-genetics work establishes that inherited disruption of piRNA biogenesis is a clinically relevant cause of spermatogenic failure: 39 infertile men were reported to carry biallelic variants across 14 piRNA pathway genes including PIWIL1, and affected tissue showed reduced pachytene piRNAs and LINE1 expression in spermatogonia consistent with transposon de-repression, supporting the piRNA pathway (including PIWIL1) as a major axis of human male infertility biology. (Stallmeyer et al., 2024-08, Nature Communications, https://doi.org/10.1038/s41467-024-50930-9) (hempfling2017expressionpatternsof pages 1-2)
3.3 Key pathway components and interaction landscape (inferred/assembled from reviews + human data)
PIWIL1 function depends on piRNAs and the broader piRNA-processing machinery. Mechanistic reviews emphasize mitochondrial primary processing by PLD6/ZUC/MitoPLD, 3′ end maturation (including trimming by factors such as PNLDC1/Trimmer) and stabilization by HENMT1/Hen1-mediated methylation; helicases such as DDX4/Vasa and Tudor-domain proteins are described as assembly/transfer factors in piRNA pathways. (Loubalova et al., 2023-09, Mobile DNA, https://doi.org/10.1186/s13100-023-00298-2) (loubalova2023themesandvariations pages 2-4, loubalova2023themesandvariations pages 1-2) (Patel et al., 2024-12, Frontiers in Cell and Developmental Biology, https://doi.org/10.3389/fcell.2024.1495035) (patel2024somaticpirnaand pages 2-4)
4) Recent developments (2023–2024 emphasis)
4.1 Cancer epigenetics and PIWI/piRNA reactivation
A high-impact 2023 review synthesizes evidence that PIWI/piRNA components, typically germline-restricted, are aberrantly expressed in cancers and can influence gene regulation through RNA cleavage and epigenetic mechanisms (e.g., recruitment of DNA/RNA methylation and other chromatin-associated regulators). This work frames PIWIL1 and related PIWI proteins as potential targets for cancer biomarkers and therapeutic exploration, while also emphasizing that cancer contexts may exploit PIWI biology differently from germline contexts. (Zhang et al., 2023-03, Molecular Cancer, https://doi.org/10.1186/s12943-023-01749-3) (zhang2023theepigeneticregulatory pages 1-2)
A 2024 “critical appraisal” review further highlights methodological and interpretive caveats (e.g., distinguishing bona fide piRNAs from other small-RNA fragments and the need for rigorous antibody validation) when assessing somatic PIWI/piRNA claims, while still recognizing recurring associations of PIWI pathway components with tumor biology and cancer stemness. (Garcia-Borja et al., 2024-02, Biomarker Research, https://doi.org/10.1186/s40364-024-00563-3) (garciaborja2024criticalappraisalof pages 24-24)
4.2 Somatic and stem-cell contexts: post-transcriptional gene regulation
A 2024 review focusing on somatic piRNA/PIWI activity summarizes evidence for PIWI-mediated regulation of mRNA stability/translation and transposon silencing in non-germline settings (e.g., stem cells and disease). While not all somatic contexts show canonical germline-like piRNA signatures, the review consolidates emerging models where PIWI proteins can participate in post-transcriptional regulation outside the germline. (Patel et al., 2024-12, Frontiers in Cell and Developmental Biology, https://doi.org/10.3389/fcell.2024.1495035) (patel2024somaticpirnaand pages 1-2)
4.3 A notable 2024 mechanistic proposal: PIWIL1 in mitosis/centrosomes (colorectal cancer)
A 2024 preprint reports a cell-cycle-dependent relocalization of PIWIL1 in colorectal cancer (CRC) models: PIWIL1 is nuclear during interphase and becomes recruited to centrosomes/MTOC during mitosis, co-localizing with γ-tubulin, and PIWIL1 knockdown induces G2/M arrest and mitotic abnormalities. (Garcia-Silva et al., 2024-07, preprint, https://doi.org/10.21203/rs.3.rs-4618560/v1) (garciasilva2024piwil1isrecruited pages 4-7, garciasilva2024piwil1isrecruited pages 1-4)
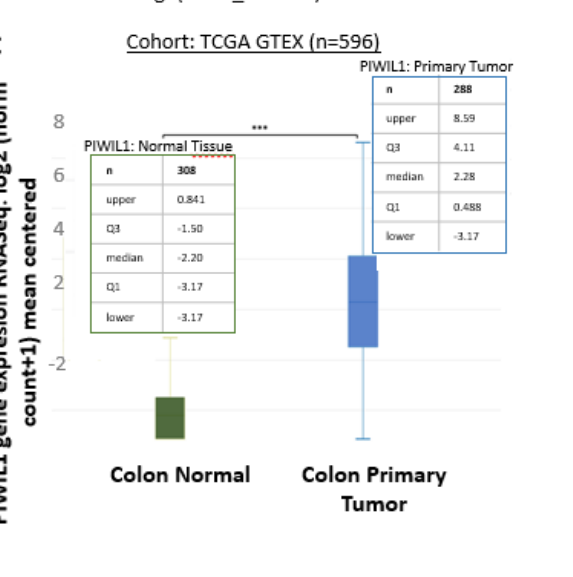
The preprint also reports PIWIL1 overexpression in CRC tissue datasets (TCGA vs GTEx) and links PIWIL1 expression to undifferentiated/stem-like states (present at crypt base, lost with differentiation). (Garcia-Silva et al., 2024-07, preprint, https://doi.org/10.21203/rs.3.rs-4618560/v1) (garciasilva2024piwil1isrecruited pages 12-18)
5) Quantitative evidence and recent statistics
5.1 Human testis: expression patterns and association with LINE-1
In 222 human testis biopsies, PIWIL1 detection frequency depended strongly on germ-cell content: PIWIL1 was reported in all normal spermatogenesis and hypospermatogenesis samples, in 96% of arrest samples, but only 5% of Sertoli-cell-only samples; it was detected in approximately 70% of tumor tissues in that cohort. (Hempfling et al., 2017-10, Reproduction, https://doi.org/10.1530/rep-16-0586) (hempfling2017expressionpatternsof pages 6-8)
The same study reported strong positive correlations between PIWI gene expression and LINE-1 (L1) expression in germ-cell-containing groups, including Spearman r = 0.94 for PIWIL1 vs L1 in normal spermatogenesis/hypospermatogenesis samples. (Hempfling et al., 2017-10, Reproduction, https://doi.org/10.1530/rep-16-0586) (hempfling2017expressionpatternsof pages 6-8)
5.2 Hepatocellular carcinoma (HCC): PIWIL1 as a candidate circulating/tissue biomarker (2024)
A 2024 RT-qPCR biomarker study reported increased PIWIL1 transcript levels in HCC tissue and serum versus controls (study size 50 HCC patients and 25 controls). (Hammad et al., 2024-02, Cancer Biomarkers, https://doi.org/10.3233/cbm-230134) (hammad2024elevatedexpressionpatterns pages 4-6, hammad2024elevatedexpressionpatterns pages 10-11)
In the extracted quantitative reporting, serum PIWIL1 achieved an ROC AUC = 1.0 (95% CI 1.0–1.0) with 100% sensitivity and 100% specificity at a stated cut-off (<1.6), with p < 0.001; tissue PIWIL1 achieved AUC = 0.80 (95% CI 0.71–0.89), 80% sensitivity, 72% specificity, p < 0.001. The same excerpt reports an odds ratio for serum PIWIL1 of 3.87 (95% CI 1.23–8.36; p < 0.001). (Hammad et al., 2024-02, Cancer Biomarkers, https://doi.org/10.3233/cbm-230134) (hammad2024elevatedexpressionpatterns pages 8-10)
5.3 Colorectal cancer (CRC): tumor-vs-normal expression and functional knockdown phenotype (2024 preprint)
In a TCGA/GTEx comparison reported in the 2024 CRC preprint, PIWIL1 was upregulated in CRC vs normal colon with N = 650 and p ≤ 0.0001. (Garcia-Silva et al., 2024-07, preprint, https://doi.org/10.21203/rs.3.rs-4618560/v1) (garciasilva2024piwil1isrecruited pages 12-18)
Experimentally, PIWIL1 knockdown reduced PIWIL1 mRNA/protein by approximately ~70% at 72 h and increased mitotic defects with an associated G2/M arrest phenotype (flow cytometry), supporting a proposed role in mitotic progression in these CRC models. (Garcia-Silva et al., 2024-07, preprint, https://doi.org/10.21203/rs.3.rs-4618560/v1) (garciasilva2024piwil1isrecruited pages 4-7, garciasilva2024piwil1isrecruited pages 12-18)
6) Current applications and real-world implementations
-
Reproductive genetics / infertility diagnostics: Human-genetics data showing biallelic pathogenic variants across piRNA pathway genes (including PIWIL1) associated with spermatogenic failure supports clinical use of piRNA-pathway genes in male-infertility genetic evaluations and variant interpretation frameworks (e.g., linking variants to pachytene-piRNA reduction and LINE1 de-repression). (Stallmeyer et al., 2024-08, Nature Communications, https://doi.org/10.1038/s41467-024-50930-9) (hempfling2017expressionpatternsof pages 1-2)
-
Cancer biomarker development: Studies propose PIWIL1 (and broader PIWI/piRNA signatures) as biomarkers in specific cancers. For example, the 2024 HCC study reports highly discriminatory performance for serum PIWIL1 in its cohort, supporting translational investigation of PIWIL1 mRNA as a candidate liquid-biopsy marker (while requiring external validation). (Hammad et al., 2024-02, Cancer Biomarkers, https://doi.org/10.3233/cbm-230134) (hammad2024elevatedexpressionpatterns pages 8-10)
-
Therapeutic targeting hypotheses: The 2024 CRC preprint proposes that inhibiting PIWIL1 could induce mitotic failure (“mitotic catastrophe”) in PIWIL1-positive CRC cells, suggesting a potential therapeutic angle; however, this is currently preclinical and preprint-stage. (Garcia-Silva et al., 2024-07, preprint, https://doi.org/10.21203/rs.3.rs-4618560/v1) (garciasilva2024piwil1isrecruited pages 9-12)
7) Expert opinion and analysis (consensus + caveats)
Consensus: Across authoritative 2023–2024 reviews, PIWIL1 is most confidently annotated as a piRNA-binding, RNA-slicing Argonaute whose best-established biological role is germline genome integrity via transposon silencing, with roles extending to broader gene regulation in germ cells. (Loubalova et al., 2023-09, https://doi.org/10.1186/s13100-023-00298-2) (loubalova2023themesandvariations pages 9-10) (Patel et al., 2024-12, https://doi.org/10.3389/fcell.2024.1495035) (patel2024somaticpirnaand pages 1-2)
Caveats in somatic/cancer interpretation: Reviews stress that somatic PIWI/piRNA studies can be confounded by (i) misannotation of small-RNA fragments as piRNAs and (ii) antibody specificity issues; thus, somatic PIWIL1 mechanisms may be heterogeneous and not always canonical piRNA biology. (Garcia-Borja et al., 2024-02, https://doi.org/10.1186/s40364-024-00563-3) (garciaborja2024criticalappraisalof pages 24-24)
8) Visual evidence from recent work (2024)
The CRC preprint provides figure-level evidence for (i) PIWIL1 upregulation in CRC vs normal colon (TCGA vs GTEx), (ii) PIWIL1 recruitment to centrosomes during mitosis with γ-tubulin co-localization, and (iii) increased G2/M arrest following PIWIL1 silencing in flow cytometry assays. (Garcia-Silva et al., 2024-07, preprint, https://doi.org/10.21203/rs.3.rs-4618560/v1) (garciasilva2024piwil1isrecruited media 49f520fd, garciasilva2024piwil1isrecruited media 9dd8f367)
9) Consolidated functional annotation summary
| Category | Summary |
|---|---|
| Identity/domains | • Verified target: human PIWIL1/HIWI, UniProt Q96J94, a PIWI-subfamily Argonaute protein, matching the user-supplied identifier and literature usage for HIWI/PIWIL1. • Domain architecture reported as PAZ, MID, PIWI; PIWIL1 is described as an ~852 aa protein with RNase H-like PIWI domain catalytic potential. • Core biochemical role is small-RNA-guided endonuclease (“slicer”) activity rather than classical metabolism; the PIWI domain cleaves complementary RNA targets guided by bound piRNAs. (zhang2023theepigeneticregulatory pages 1-2, patel2024somaticpirnaand pages 1-2, limanowka2024mechanismsbehindthe pages 1-2) |
| Molecular function | • Binds piRNAs (typically ~24–32/31 nt, 3′ 2′-O-methylated) to form PIWI–piRNA ribonucleoprotein complexes/piRISC. • Main established function is germline genome defense, especially transposon silencing and maintenance of spermatogenesis/fertility. • Can mediate post-transcriptional silencing by slicing target RNAs and, in broader PIWI/piRNA models, contribute to epigenetic regulation by recruiting chromatin/DNA methylation machinery. (zhang2023theepigeneticregulatory pages 1-2, hempfling2017expressionpatternsof pages 1-2, patel2024somaticpirnaand pages 1-2, loubalova2023themesandvariations pages 9-10) |
| Pathway modules | • Primary biogenesis/loading: long piRNA-cluster precursors are cleaved by PLD6/ZUC/MitoPLD on mitochondria, loaded into PIWI proteins, then trimmed (e.g., PNLDC1/Trimmer) and methylated by HENMT1/Hen1. • Effector/slicer module: mature PIWIL1–piRNA complexes cleave complementary RNAs in the cytoplasm. • Secondary amplification/ping-pong: mechanistically central to piRNA biology, but evidence indicates PIWIL1/MIWI is mainly primary-piRNA-loaded in mammalian pachytene cells, with ping-pong more strongly attributed to PIWIL2/MILI. • Nuclear silencing module: PIWI/piRNA complexes can also support transcriptional silencing via chromatin modifiers in general pathway models. (loubalova2023themesandvariations pages 4-6, loubalova2023themesandvariations pages 9-10, loubalova2023themesandvariations pages 2-4, loubalova2023themesandvariations pages 1-2, loubalova2023themesandvariations pages 6-7, patel2024somaticpirnaand pages 2-4, loubalova2023themesandvariations pages 10-11) |
| Localization | • In human testis, PIWIL1 protein is detected in the cytoplasm of late pachytene spermatocytes and spermatids, disappearing in elongated spermatids. • Signal concentrates in structures consistent with mitochondrial cement/nuage in pachytene cells and the chromatoid body in spermatids. • In CRC cell models, PIWIL1 is nuclear during interphase and relocalizes to centrosomes/MTOC during mitosis, co-localizing with γ-tubulin; tumor tissues show mainly cytoplasmic staining. (hempfling2017expressionpatternsof pages 3-4, hempfling2017expressionpatternsof pages 9-10, garciasilva2024piwil1isrecruited pages 4-7, garciasilva2024piwil1isrecruited pages 1-4, garciasilva2024piwil1isrecruited media 49f520fd) |
| Key interactors | • Direct molecular partner class: piRNAs. • Biogenesis/processing factors linked in pathway models include PLD6/ZUC, MOV10L1, PNLDC1, HENMT1, and the helicase DDX4/Vasa. • PIWI family proteins interact with Tudor-domain proteins (TDRDs) in arginine-methylation-dependent assemblies; human/oocyte literature also places PIWI proteins in complexes with mitochondrial/piRNA biogenesis machinery. • In CRC mitosis work, PIWIL1 associates spatially with γ-tubulin/centrosomal machinery. (limanowka2024mechanismsbehindthe pages 1-2, loubalova2023themesandvariations pages 9-10, loubalova2023themesandvariations pages 2-4, patel2024somaticpirnaand pages 2-4, garciasilva2024piwil1isrecruited pages 4-7) |
| Human germline evidence | • Human testis study of 222 biopsies found PIWIL1 and HENMT1 coexpressed in pachytene spermatocytes/spermatids; PIWIL1 expression tracked germ-cell content and was nearly absent from Sertoli-cell-only tissue. • PIWIL1 was expressed in all normal spermatogenesis samples, all hypospermatogenesis samples, 96% of arrest samples, but only 5% of Sertoli-cell-only samples. • Low/absent piRNA-pathway components were associated with higher LINE-1 expression, supporting a role in transposon repression in human testis. • A 2024 human genetics study identified 39 infertile men carrying biallelic variants in 14 piRNA-pathway genes including PIWIL1, with reduced pachytene piRNAs and LINE1 de-silencing, establishing piRNA-pathway disruption as a major cause of spermatogenic failure. (hempfling2017expressionpatternsof pages 1-2, hempfling2017expressionpatternsof pages 6-8, hempfling2017expressionpatternsof pages 8-9) |
| Somatic/cancer evidence | • Reviews from 2023–2024 consistently note that PIWIL1 is normally germline-restricted but is aberrantly expressed in multiple tumors, motivating biomarker interest. • In HCC, PIWIL1 mRNA was reported as elevated in tumor tissue and serum versus controls. • In CRC models, PIWIL1 is overexpressed and appears linked to a piRNA-independent role in mitotic fidelity/cell-cycle progression, especially centrosome-associated behavior during mitosis. • Somatic PIWI/piRNA literature remains mechanistically heterogeneous, with stronger evidence for association than for universal canonical piRNA function in all tumors. (zhang2023theepigeneticregulatory pages 1-2, limanowka2024mechanismsbehindthe pages 1-2, hammad2024elevatedexpressionpatterns pages 8-10, garciasilva2024piwil1isrecruited pages 7-9, garciasilva2024piwil1isrecruited pages 12-18) |
| Quantitative findings 2017-2024 | • Human testis cohort: 222 biopsies analyzed; PIWIL1 detected in all normal spermatogenesis and hypospermatogenesis samples, 96% of arrest samples, ~70% of tumors, 5% of Sertoli-cell-only samples. • Correlation with LINE-1 in germ-cell-containing samples was strong (Spearman r = 0.94 for PIWIL1 vs L1 in Nsp/Hyp groups). • HCC study: 50 HCC patients and 25 controls; serum PIWIL1 ROC AUC 1.0, sensitivity 100%, specificity 100%, p<0.001; tissue PIWIL1 ROC AUC 0.80, sensitivity 80%, specificity 72%, p<0.001; serum PIWIL1 OR 3.87 (95% CI 1.23–8.36), p<0.001. • CRC preprint: TCGA/GTEx comparison showed PIWIL1 upregulation in CRC with N=650, p≤0.0001; PIWIL1 knockdown reduced mRNA/protein by ~70% at 72 h and increased G2/M arrest/mitotic defects. (hempfling2017expressionpatternsof pages 6-8, hammad2024elevatedexpressionpatterns pages 8-10, hammad2024elevatedexpressionpatterns pages 7-8, hammad2024elevatedexpressionpatterns pages 4-6, garciasilva2024piwil1isrecruited pages 12-18, garciasilva2024piwil1isrecruited pages 4-7, garciasilva2024piwil1isrecruited media 49f520fd) |
| Notes/caveats | • The gene identity is not ambiguous here: the literature aligns with human PIWIL1/HIWI and the supplied UniProt record. • However, much mechanistic detail comes from cross-species PIWI/piRNA biology and is often inferred to human PIWIL1. • Some cancer studies may detect non-canonical piRNA-like fragments rather than bona fide piRNAs; several reviews explicitly caution about annotation and antibody-validation issues. • The CRC centrosome study is a 2024 preprint, so its mitotic localization/function findings should be treated as provisional until peer-reviewed. (garciaborja2024criticalappraisalof pages 24-24, zhang2023theepigeneticregulatory pages 1-2, loubalova2023themesandvariations pages 4-6, garciasilva2024piwil1isrecruited pages 7-9) |
Table: This table summarizes the strongest evidence for human PIWIL1/HIWI identity, mechanism, localization, interacting pathway components, and quantitative findings from 2017–2024. It is designed as a compact functional-annotation reference with direct context-ID citations for each row.
References (URLs and publication dates)
Key recent sources emphasized in this report include:
- Zhang Q. et al. (2023-03). Molecular Cancer. “The epigenetic regulatory mechanism of PIWI/piRNAs in human cancers.” https://doi.org/10.1186/s12943-023-01749-3 (zhang2023theepigeneticregulatory pages 1-2)
- Loubalova Z. et al. (2023-09). Mobile DNA. “Themes and variations on piRNA-guided transposon control.” https://doi.org/10.1186/s13100-023-00298-2 (loubalova2023themesandvariations pages 9-10)
- Garcia-Borja E. et al. (2024-02). Biomarker Research. “Critical appraisal of the piRNA-PIWI axis in cancer and cancer stem cells.” https://doi.org/10.1186/s40364-024-00563-3 (garciaborja2024criticalappraisalof pages 24-24)
- Patel M.Z. et al. (2024-12). Frontiers in Cell and Developmental Biology. “Somatic piRNA and PIWI-mediated post-transcriptional gene regulation in stem cells and disease.” https://doi.org/10.3389/fcell.2024.1495035 (patel2024somaticpirnaand pages 1-2)
- Hammad G. et al. (2024-02). Cancer Biomarkers. “Elevated expression patterns of PIWI transcripts are potential candidate markers for hepatocellular carcinoma.” https://doi.org/10.3233/cbm-230134 (hammad2024elevatedexpressionpatterns pages 8-10)
- Garcia-Silva M.R. et al. (2024-07). Preprint. “PIWIL1 is recruited to Centrosomes during Mitosis in Colorectal Cancer Cells…” https://doi.org/10.21203/rs.3.rs-4618560/v1 (garciasilva2024piwil1isrecruited pages 12-18)
- Stallmeyer B. et al. (2024-08). Nature Communications. “Inherited defects of piRNA biogenesis cause transposon de-repression, impaired spermatogenesis, and human male infertility.” https://doi.org/10.1038/s41467-024-50930-9 (hempfling2017expressionpatternsof pages 1-2)
- Hempfling A. et al. (2017-10). Reproduction. “Expression patterns of HENMT1 and PIWIL1 in human testis: implications for transposon expression.” https://doi.org/10.1530/rep-16-0586 (hempfling2017expressionpatternsof pages 1-2)
References
-
(zhang2023theepigeneticregulatory pages 1-2): Qun Zhang, Yazhi Zhu, Xinyu Cao, Wenhui Tan, Jianglong Yu, Yaqiong Lu, Ran Kang, Xiaolan Wang, and Ermao Li. The epigenetic regulatory mechanism of piwi/pirnas in human cancers. Molecular Cancer, Mar 2023. URL: https://doi.org/10.1186/s12943-023-01749-3, doi:10.1186/s12943-023-01749-3. This article has 84 citations and is from a highest quality peer-reviewed journal.
-
(patel2024somaticpirnaand pages 1-2): Mahammed Zaid Patel, Yuguan Jiang, and Pavan Kumar Kakumani. Somatic pirna and piwi-mediated post-transcriptional gene regulation in stem cells and disease. Frontiers in Cell and Developmental Biology, Dec 2024. URL: https://doi.org/10.3389/fcell.2024.1495035, doi:10.3389/fcell.2024.1495035. This article has 11 citations.
-
(loubalova2023themesandvariations pages 9-10): Zuzana Loubalova, Parthena Konstantinidou, and Astrid D. Haase. Themes and variations on pirna-guided transposon control. Mobile DNA, Sep 2023. URL: https://doi.org/10.1186/s13100-023-00298-2, doi:10.1186/s13100-023-00298-2. This article has 38 citations and is from a peer-reviewed journal.
-
(loubalova2023themesandvariations pages 2-4): Zuzana Loubalova, Parthena Konstantinidou, and Astrid D. Haase. Themes and variations on pirna-guided transposon control. Mobile DNA, Sep 2023. URL: https://doi.org/10.1186/s13100-023-00298-2, doi:10.1186/s13100-023-00298-2. This article has 38 citations and is from a peer-reviewed journal.
-
(loubalova2023themesandvariations pages 4-6): Zuzana Loubalova, Parthena Konstantinidou, and Astrid D. Haase. Themes and variations on pirna-guided transposon control. Mobile DNA, Sep 2023. URL: https://doi.org/10.1186/s13100-023-00298-2, doi:10.1186/s13100-023-00298-2. This article has 38 citations and is from a peer-reviewed journal.
-
(loubalova2023themesandvariations pages 6-7): Zuzana Loubalova, Parthena Konstantinidou, and Astrid D. Haase. Themes and variations on pirna-guided transposon control. Mobile DNA, Sep 2023. URL: https://doi.org/10.1186/s13100-023-00298-2, doi:10.1186/s13100-023-00298-2. This article has 38 citations and is from a peer-reviewed journal.
-
(hempfling2017expressionpatternsof pages 1-2): A. Hempfling, A. Hempfling, S. Lim, D. Adelson, Jemma Evans, A. O’Connor, Z. Qu, S. Kliesch, W. Weidner, Moira K. O’Bryan, and M. Bergmann. Expression patterns of henmt1 and piwil1 in human testis: implications for transposon expression. Reproduction, 154 4:363-374, Oct 2017. URL: https://doi.org/10.1530/rep-16-0586, doi:10.1530/rep-16-0586. This article has 50 citations and is from a peer-reviewed journal.
-
(hempfling2017expressionpatternsof pages 3-4): A. Hempfling, A. Hempfling, S. Lim, D. Adelson, Jemma Evans, A. O’Connor, Z. Qu, S. Kliesch, W. Weidner, Moira K. O’Bryan, and M. Bergmann. Expression patterns of henmt1 and piwil1 in human testis: implications for transposon expression. Reproduction, 154 4:363-374, Oct 2017. URL: https://doi.org/10.1530/rep-16-0586, doi:10.1530/rep-16-0586. This article has 50 citations and is from a peer-reviewed journal.
-
(loubalova2023themesandvariations pages 1-2): Zuzana Loubalova, Parthena Konstantinidou, and Astrid D. Haase. Themes and variations on pirna-guided transposon control. Mobile DNA, Sep 2023. URL: https://doi.org/10.1186/s13100-023-00298-2, doi:10.1186/s13100-023-00298-2. This article has 38 citations and is from a peer-reviewed journal.
-
(patel2024somaticpirnaand pages 2-4): Mahammed Zaid Patel, Yuguan Jiang, and Pavan Kumar Kakumani. Somatic pirna and piwi-mediated post-transcriptional gene regulation in stem cells and disease. Frontiers in Cell and Developmental Biology, Dec 2024. URL: https://doi.org/10.3389/fcell.2024.1495035, doi:10.3389/fcell.2024.1495035. This article has 11 citations.
-
(garciaborja2024criticalappraisalof pages 24-24): Elena Garcia-Borja, Frantisek Siegl, Rosana Mateu, Ondrej Slaby, Aleksi Sedo, Petr Busek, and Jiri Sana. Critical appraisal of the pirna-piwi axis in cancer and cancer stem cells. Biomarker Research, Feb 2024. URL: https://doi.org/10.1186/s40364-024-00563-3, doi:10.1186/s40364-024-00563-3. This article has 36 citations and is from a peer-reviewed journal.
-
(garciasilva2024piwil1isrecruited pages 4-7): Maria Rosa Garcia-Silva, Sofía Montenegro, Sofía Dacosta, Juan Pablo Tosar, and Alfonso Cayota. Piwil1 is recruited to centrosomes during mitosis in colorectal cancer cells and is linked to cell cycle progression. ArXiv, Jul 2024. URL: https://doi.org/10.21203/rs.3.rs-4618560/v1, doi:10.21203/rs.3.rs-4618560/v1. This article has 1 citations.
-
(garciasilva2024piwil1isrecruited pages 1-4): Maria Rosa Garcia-Silva, Sofía Montenegro, Sofía Dacosta, Juan Pablo Tosar, and Alfonso Cayota. Piwil1 is recruited to centrosomes during mitosis in colorectal cancer cells and is linked to cell cycle progression. ArXiv, Jul 2024. URL: https://doi.org/10.21203/rs.3.rs-4618560/v1, doi:10.21203/rs.3.rs-4618560/v1. This article has 1 citations.
-
(garciasilva2024piwil1isrecruited pages 12-18): Maria Rosa Garcia-Silva, Sofía Montenegro, Sofía Dacosta, Juan Pablo Tosar, and Alfonso Cayota. Piwil1 is recruited to centrosomes during mitosis in colorectal cancer cells and is linked to cell cycle progression. ArXiv, Jul 2024. URL: https://doi.org/10.21203/rs.3.rs-4618560/v1, doi:10.21203/rs.3.rs-4618560/v1. This article has 1 citations.
-
(hempfling2017expressionpatternsof pages 6-8): A. Hempfling, A. Hempfling, S. Lim, D. Adelson, Jemma Evans, A. O’Connor, Z. Qu, S. Kliesch, W. Weidner, Moira K. O’Bryan, and M. Bergmann. Expression patterns of henmt1 and piwil1 in human testis: implications for transposon expression. Reproduction, 154 4:363-374, Oct 2017. URL: https://doi.org/10.1530/rep-16-0586, doi:10.1530/rep-16-0586. This article has 50 citations and is from a peer-reviewed journal.
-
(hammad2024elevatedexpressionpatterns pages 4-6): Gehan Hammad, Samah Mamdouh, Dina Mohamed Seoudi, Mohamed Ismail Seleem, Gehan Safwat, and Rania Hassan Mohamed. Elevated expression patterns of p-element induced wimpy testis (piwi) transcripts are potential candidate markers for hepatocellular carcinoma. Cancer Biomarkers, 39:95-111, Feb 2024. URL: https://doi.org/10.3233/cbm-230134, doi:10.3233/cbm-230134. This article has 10 citations and is from a peer-reviewed journal.
-
(hammad2024elevatedexpressionpatterns pages 10-11): Gehan Hammad, Samah Mamdouh, Dina Mohamed Seoudi, Mohamed Ismail Seleem, Gehan Safwat, and Rania Hassan Mohamed. Elevated expression patterns of p-element induced wimpy testis (piwi) transcripts are potential candidate markers for hepatocellular carcinoma. Cancer Biomarkers, 39:95-111, Feb 2024. URL: https://doi.org/10.3233/cbm-230134, doi:10.3233/cbm-230134. This article has 10 citations and is from a peer-reviewed journal.
-
(hammad2024elevatedexpressionpatterns pages 8-10): Gehan Hammad, Samah Mamdouh, Dina Mohamed Seoudi, Mohamed Ismail Seleem, Gehan Safwat, and Rania Hassan Mohamed. Elevated expression patterns of p-element induced wimpy testis (piwi) transcripts are potential candidate markers for hepatocellular carcinoma. Cancer Biomarkers, 39:95-111, Feb 2024. URL: https://doi.org/10.3233/cbm-230134, doi:10.3233/cbm-230134. This article has 10 citations and is from a peer-reviewed journal.
-
(garciasilva2024piwil1isrecruited pages 9-12): Maria Rosa Garcia-Silva, Sofía Montenegro, Sofía Dacosta, Juan Pablo Tosar, and Alfonso Cayota. Piwil1 is recruited to centrosomes during mitosis in colorectal cancer cells and is linked to cell cycle progression. ArXiv, Jul 2024. URL: https://doi.org/10.21203/rs.3.rs-4618560/v1, doi:10.21203/rs.3.rs-4618560/v1. This article has 1 citations.
-
(garciasilva2024piwil1isrecruited media 49f520fd): Maria Rosa Garcia-Silva, Sofía Montenegro, Sofía Dacosta, Juan Pablo Tosar, and Alfonso Cayota. Piwil1 is recruited to centrosomes during mitosis in colorectal cancer cells and is linked to cell cycle progression. ArXiv, Jul 2024. URL: https://doi.org/10.21203/rs.3.rs-4618560/v1, doi:10.21203/rs.3.rs-4618560/v1. This article has 1 citations.
-
(garciasilva2024piwil1isrecruited media 9dd8f367): Maria Rosa Garcia-Silva, Sofía Montenegro, Sofía Dacosta, Juan Pablo Tosar, and Alfonso Cayota. Piwil1 is recruited to centrosomes during mitosis in colorectal cancer cells and is linked to cell cycle progression. ArXiv, Jul 2024. URL: https://doi.org/10.21203/rs.3.rs-4618560/v1, doi:10.21203/rs.3.rs-4618560/v1. This article has 1 citations.
-
(limanowka2024mechanismsbehindthe pages 1-2): Piotr Limanówka, Błażej Ochman, and Elżbieta Świętochowska. Mechanisms behind the impact of piwi proteins on cancer cells: literature review. International Journal of Molecular Sciences, 25:12217, Nov 2024. URL: https://doi.org/10.3390/ijms252212217, doi:10.3390/ijms252212217. This article has 4 citations.
-
(loubalova2023themesandvariations pages 10-11): Zuzana Loubalova, Parthena Konstantinidou, and Astrid D. Haase. Themes and variations on pirna-guided transposon control. Mobile DNA, Sep 2023. URL: https://doi.org/10.1186/s13100-023-00298-2, doi:10.1186/s13100-023-00298-2. This article has 38 citations and is from a peer-reviewed journal.
-
(hempfling2017expressionpatternsof pages 9-10): A. Hempfling, A. Hempfling, S. Lim, D. Adelson, Jemma Evans, A. O’Connor, Z. Qu, S. Kliesch, W. Weidner, Moira K. O’Bryan, and M. Bergmann. Expression patterns of henmt1 and piwil1 in human testis: implications for transposon expression. Reproduction, 154 4:363-374, Oct 2017. URL: https://doi.org/10.1530/rep-16-0586, doi:10.1530/rep-16-0586. This article has 50 citations and is from a peer-reviewed journal.
-
(hempfling2017expressionpatternsof pages 8-9): A. Hempfling, A. Hempfling, S. Lim, D. Adelson, Jemma Evans, A. O’Connor, Z. Qu, S. Kliesch, W. Weidner, Moira K. O’Bryan, and M. Bergmann. Expression patterns of henmt1 and piwil1 in human testis: implications for transposon expression. Reproduction, 154 4:363-374, Oct 2017. URL: https://doi.org/10.1530/rep-16-0586, doi:10.1530/rep-16-0586. This article has 50 citations and is from a peer-reviewed journal.
-
(garciasilva2024piwil1isrecruited pages 7-9): Maria Rosa Garcia-Silva, Sofía Montenegro, Sofía Dacosta, Juan Pablo Tosar, and Alfonso Cayota. Piwil1 is recruited to centrosomes during mitosis in colorectal cancer cells and is linked to cell cycle progression. ArXiv, Jul 2024. URL: https://doi.org/10.21203/rs.3.rs-4618560/v1, doi:10.21203/rs.3.rs-4618560/v1. This article has 1 citations.
-
(hammad2024elevatedexpressionpatterns pages 7-8): Gehan Hammad, Samah Mamdouh, Dina Mohamed Seoudi, Mohamed Ismail Seleem, Gehan Safwat, and Rania Hassan Mohamed. Elevated expression patterns of p-element induced wimpy testis (piwi) transcripts are potential candidate markers for hepatocellular carcinoma. Cancer Biomarkers, 39:95-111, Feb 2024. URL: https://doi.org/10.3233/cbm-230134, doi:10.3233/cbm-230134. This article has 10 citations and is from a peer-reviewed journal.
Artifacts
Citations
- zhang2023theepigeneticregulatory pages 1-2
- patel2024somaticpirnaand pages 1-2
- hempfling2017expressionpatternsof pages 3-4
- hempfling2017expressionpatternsof pages 1-2
- patel2024somaticpirnaand pages 2-4
- garciaborja2024criticalappraisalof pages 24-24
- hempfling2017expressionpatternsof pages 6-8
- hammad2024elevatedexpressionpatterns pages 8-10
- loubalova2023themesandvariations pages 9-10
- loubalova2023themesandvariations pages 2-4
- loubalova2023themesandvariations pages 4-6
- loubalova2023themesandvariations pages 6-7
- loubalova2023themesandvariations pages 1-2
- hammad2024elevatedexpressionpatterns pages 4-6
- hammad2024elevatedexpressionpatterns pages 10-11
- limanowka2024mechanismsbehindthe pages 1-2
- loubalova2023themesandvariations pages 10-11
- hempfling2017expressionpatternsof pages 9-10
- hempfling2017expressionpatternsof pages 8-9
- hammad2024elevatedexpressionpatterns pages 7-8
- https://doi.org/10.1186/s12943-023-01749-3
- https://doi.org/10.3389/fcell.2024.1495035
- https://doi.org/10.1186/s13100-023-00298-2
- https://doi.org/10.1530/rep-16-0586
- https://doi.org/10.1038/s41467-024-50930-9
- https://doi.org/10.1186/s40364-024-00563-3
- https://doi.org/10.21203/rs.3.rs-4618560/v1
- https://doi.org/10.3233/cbm-230134
- https://doi.org/10.1186/s12943-023-01749-3,
- https://doi.org/10.3389/fcell.2024.1495035,
- https://doi.org/10.1186/s13100-023-00298-2,
- https://doi.org/10.1530/rep-16-0586,
- https://doi.org/10.1186/s40364-024-00563-3,
- https://doi.org/10.21203/rs.3.rs-4618560/v1,
- https://doi.org/10.3233/cbm-230134,
- https://doi.org/10.3390/ijms252212217,
Deep Research Report: PIWIL1 (human)
(PIWIL1-deep-research.md)
Deep Research Report: PIWIL1 (human)
(PIWIL1-deep-research.md)Deep Research Report: PIWIL1 (human)
Generated using OpenAI Deep Research API
UniProt ID: Q96J94
Directory alias: PIWIL1
PIWIL1 (Piwi-like Protein 1) – Comprehensive Gene Report
Gene Function and Molecular Mechanisms
PIWIL1 (also known as HIWI) encodes a member of the PIWI subfamily of Argonaute proteins, which are evolutionarily conserved RNA-binding proteins crucial for piRNA (PIWI-interacting RNA) pathways (www.nature.com) (en.wikipedia.org). PIWIL1 binds 24–32 nt piRNAs to form effector ribonucleoprotein complexes analogous to RISC, mediating RNA silencing and transposon repression in germ cells (www.nature.com) (www.spandidos-publications.com). Through its PIWI-piRNA complexes, PIWIL1 helps maintain genomic integrity by silencing transposable elements, via post-transcriptional cleavage of transposon transcripts and by guiding epigenetic DNA methylation at transposon loci (www.spandidos-publications.com). This gene is thought to act as an intrinsic regulator of stem cell self-renewal capacity in the germline, and it has also been detected in CD34^+ hematopoietic stem/progenitor cells (but not in differentiated blood cells) (www.spandidos-publications.com) (www.spandidos-publications.com), suggesting a role in maintaining stem cell populations. In diverse organisms (from Drosophila to mammals), Piwi-family proteins like PIWIL1 are essential for gametogenesis and germline development, as evidenced by their requirement for stem cell maintenance and fertility (www.nature.com) (www.nature.com). In mice, for example, loss of the PIWIL1 homolog (Miwi) causes spermatogenic arrest and male sterility, underscoring its critical function in spermatogenesis and germ cell maturation (pmc.ncbi.nlm.nih.gov). Mechanistically, PIWIL1 has endoribonuclease (“slicer”) activity associated with its PIWI domain, which cleaves target RNAs complementary to bound piRNAs (pmc.ncbi.nlm.nih.gov). This endonuclease activity, analogous to RNase H, is central to degrading transposon transcripts and also participates in the “ping-pong” amplification cycle of piRNAs in germ cells (pmc.ncbi.nlm.nih.gov) (www.spandidos-publications.com). Beyond transposon silencing, PIWIL1 is implicated in post-transcriptional regulation of mRNAs – for instance, it can repress specific messenger RNAs or modulate their translation in the germline context, contributing to proper differentiation of gametes (as seen for other Piwi proteins) (www.nature.com). Notably, emerging evidence also suggests piRNA-independent roles of PIWIL1 in somatic cells: one study found PIWIL1 can act as a co-factor of the anaphase-promoting complex (APC/C) in cancer cells to influence the cell cycle, indicating additional molecular mechanisms in pathological contexts (pubmed.ncbi.nlm.nih.gov). Overall, PIWIL1 functions as an RNA-guided effector protein safeguarding genome stability and regulating gene expression programs in the germline.
Cellular Localization and Subcellular Components
PIWIL1 is predominantly a cytoplasmic protein. In human germ cells, it localizes to specialized perinuclear RNA granules characteristic of the piRNA pathway (pmc.ncbi.nlm.nih.gov). During spermatogenesis, PIWIL1 and other PIWI proteins concentrate in structures such as the intermitochondrial cement of spermatocytes and the chromatoid bodies (a single large ribonucleoprotein granule) of round spermatids (pmc.ncbi.nlm.nih.gov). These nuage (germ granule) compartments are sites of piRNA biogenesis and action, where PIWIL1 colocalizes with other piRNA pathway factors. For example, MIWI (mouse Piwil1) is a major component of the chromatoid body and helps recruit other proteins (like TDRD tudor proteins and the methylosome component WDR77) to these granules (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Consistently, human PIWIL1 is found in the cytosol associated with RNP complexes, rather than the nucleus (www.nature.com). Biochemical fractionation of somatic cells ectopically expressing PIWIL1 also showed it in both soluble cytosolic and membrane-associated fractions, with only minimal presence in the nuclear compartment (www.embopress.org). Within the cytoplasm, PIWIL1 associates with the piRNA processing machinery; for instance, it interacts with MVH/DDX4 (Vasa) and other components in the germ-cell nuage, and with P-body markers when expressed in certain contexts, reflecting a role in mRNA processing bodies (pubmed.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In summary, PIWIL1 resides in the cytoplasm, concentrating in germline-specific granular structures (pi-bodies/nuage) that facilitate piRNA maturation and transposon silencing activities.
Biological Processes Involvement
Transposon Silencing and Genome Defense: The primary biological role of PIWIL1 is in the piRNA pathway, where it protects the germline genome from transposable elements. PIWIL1-piRNA complexes recognize and slice transposon transcripts, and also recruit DNA methylation and repressive chromatin modifications at transposon loci (www.spandidos-publications.com). This is crucial for maintaining genomic stability in germ cells and preventing harmful mutations.
Gametogenesis and Fertility: PIWIL1 is essential for normal gamete development, especially in male meiosis and spermiogenesis. It is highly expressed in spermatocytes and round spermatids, and is required for the progression of spermatogenesis (www.nature.com). In mouse models, disruption of Piwil1 leads to accumulation of spermatocytes that fail to mature properly, resulting in infertility (pmc.ncbi.nlm.nih.gov). In humans, a recent study reported germline PIWIL1 mutations in men with azoospermia (absence of sperm) and linked these mutations to defective histone-to-protamine exchange during spermiogenesis (pmc.ncbi.nlm.nih.gov). The mutant PIWIL1 protein could not be properly ubiquitinated and degraded, causing it to abnormally sequester factors (like RNF8) needed for chromatin remodeling, ultimately blocking sperm maturation (pmc.ncbi.nlm.nih.gov). This provides direct evidence that PIWIL1 is involved in late stages of sperm development (chromatin condensation and protamine incorporation). PIWIL1 may also contribute to oogenesis and early embryogenesis by safeguarding the maternal genome, although in humans its expression in oocytes is minimal compared to its dominant role in spermatogenesis (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov).
Stem Cell Maintenance: Across species, piwi genes are known for roles in stem cell niche maintenance. Drosophila Piwi, for example, is required in the germline stem cells and somatic niche cells to maintain stem cell renewal. Analogously, human PIWIL1 has been proposed to support the self-renewal of germline stem cells and possibly hematopoietic stem cells (www.spandidos-publications.com) (en.wikipedia.org). The presence of PIWIL1 in undifferentiated spermatogonia and in CD34⁺ bone marrow cells suggests it may influence stem cell proliferation or prevent differentiation, although the exact mechanisms in humans remain to be fully elucidated.
Small RNA Biogenesis and RNA Interference: PIWIL1 is intimately involved in piRNA biogenesis. It participates in the “ping-pong” amplification loop of piRNAs by using its slicer activity to cleave target RNAs, which in turn generates precursor ends that are loaded into other PIWI proteins (like PIWIL2/Ago3) to produce secondary piRNAs (www.spandidos-publications.com). PIWIL1 thus helps amplify the piRNA pool and sustain transposon silencing. Beyond piRNAs, PIWIL1’s involvement in RNA interference overlaps with other small RNA pathways: it associates with DICER1 and shares components with the microRNA machinery (www.embopress.org) (www.embopress.org). However, PIWIL1-bound small RNAs are a distinct class (piRNAs) that do not require Dicer for their maturation. Instead, nucleases like Zucchini (in animals) process piRNA precursors which are then loaded onto PIWIL1. PIWIL1 may also contribute to translational regulation of specific mRNAs in germ cells, ensuring timely expression of proteins during meiosis and spermiogenesis – a role supported by the presence of PIWIL1 in polysomes and P-bodies observed in some studies (www.embopress.org) (www.embopress.org).
Epigenetic Regulation: Through its interactions and the downstream effects of piRNA targeting, PIWIL1 influences epigenetic states. In the nucleus (in organisms where PIWI proteins enter the nucleus), Piwi-piRNA complexes can recruit histone methyltransferases or DNA methyltransferases to silence genomic loci. In mammals, PIWIL4 (a nuclear Piwi protein) has been shown to induce heterochromatic silencing of transposons via H3K9 methylation. PIWIL1 itself is mostly cytoplasmic, but by silencing transposon transcripts it indirectly ensures that transposons are substrates for DNA methylation in the germline genome (www.spandidos-publications.com). Moreover, PIWIL1’s interaction with Tudor domain proteins helps organize chromatoid bodies which are thought to serve as centers for small RNA-dependent post-transcriptional gene regulation.
Together, these activities place PIWIL1 at the crossroads of several biological processes: RNA silencing, germ cell development, stem cell maintenance, and epigenetic genome defense. Its proper function is indispensable for fertility and stable transmission of genetic information to the next generation.
Disease Associations and Phenotypes
In healthy adults, PIWIL1 is normally restricted to germline tissues (primarily the testis). However, aberrant expression of PIWIL1 in somatic tissues is linked to disease states. Notably, PIWIL1 has emerged as a cancer/testis (CT) antigen – a gene silent in most normal tissues but activated in various cancers (www.spandidos-publications.com) (www.spandidos-publications.com). Tumor Expression and Prognosis: Elevated PIWIL1 expression has been observed in a broad range of malignancies, including breast, esophageal, pancreatic, gastric, colorectal, lung, and endometrial cancers (www.spandidos-publications.com). In immunohistochemical studies, overexpression of PIWIL1 in tumors often correlates with higher tumor grade, advanced clinical stage, and poorer patient survival outcomes (www.spandidos-publications.com) (www.spandidos-publications.com). For example, PIWIL1 is highly expressed in a subset of lung adenocarcinomas (where it was identified as an extremely upregulated CT gene) and has been shown to promote lung cancer cell proliferation, migration, and invasion (pubmed.ncbi.nlm.nih.gov). Similarly, in colorectal cancer and gastric cancer, PIWIL1 upregulation is associated with enhanced tumor cell growth and may contribute to epigenetic changes (such as DNA hypermethylation of tumor suppressor genes) that favor oncogenesis (pubmed.ncbi.nlm.nih.gov) (www.frontiersin.org). A systematic review and meta-analysis confirmed that high PIWIL1 levels generally predict worse prognosis across multiple cancer types (www.spandidos-publications.com), highlighting its potential as a prognostic biomarker. These findings suggest that PIWIL1, normally conferring stem-cell like properties, might endow cancer cells with increased proliferative capacity and genomic permissiveness, thereby facilitating tumor progression.
Mechanistic Links to Cancer: The oncogenic role of PIWIL1 is an area of active research. In some cancers, PIWIL1 may interact with cellular pathways independent of piRNAs. For instance, PIWIL1 can bind the APC/C complex co-factor ANAPC10 in piRNA-loaded states, and in pancreatic cancer cells a piRNA-independent function of PIWIL1 as an APC/C co-activator was reported, driving cell cycle progression and metastasis (pubmed.ncbi.nlm.nih.gov). Additionally, a RASSF1C–PIWIL1–piRNA axis has been described in lung cancer cells: RASSF1C (an oncogenic isoform of RASSF1) upregulates PIWIL1 and piRNA levels, leading to DNA methylation changes that downregulate tumor suppressors (pubmed.ncbi.nlm.nih.gov). These examples illustrate that PIWIL1 may contribute to tumorigenesis both by sustaining stem-cell-like characteristics (e.g., self-renewal, resistance to apoptosis) and by reshaping epigenetic patterns in cancer cells.
Germ Cell Tumors: Given its normal expression in germ cells, PIWIL1 has also been studied in the context of germ cell tumors. The PIWIL1 gene resides at chromosome 12q24.33 (www.nature.com), a region frequently gained in testicular germ cell tumors (TGCTs). An initial report showed HIWI is overexpressed in seminomas (a type of testicular germ cell tumor) in about 63% of cases examined (www.nature.com) (www.nature.com). Seminomas with high PIWIL1 expression retain a germ cell-like phenotype, consistent with the hypothesis that PIWIL1 may support the undifferentiated state of tumor germ cells (www.nature.com). In contrast, somatic-origin testicular tumors (e.g., Sertoli or Leydig cell tumors) showed no PIWIL1 expression (www.nature.com). Thus, PIWIL1 might serve as a useful marker to distinguish germ cell tumors and could play a role in their biology by protecting the cancer cell genome or fostering an environment of self-renewal.
Reproductive and Developmental Disorders: Outside of cancer, alterations in PIWIL1 primarily manifest in reproductive phenotypes. As mentioned, loss-of-function mutations in PIWIL1 can cause human male infertility (azoospermia) (pmc.ncbi.nlm.nih.gov). Patients with such mutations present with spermatogenic failure due to inability to produce functional sperm, aligning with the critical role of PIWIL1 in spermatogenesis. There is so far limited evidence linking PIWIL1 to disorders in females; however, dysregulation of the piRNA pathway in general has been speculated to contribute to ovarian germ cell anomalies or certain cases of oocyte attrition, though PIWIL1 itself is not highly expressed in oocytes past the fetal stage (pmc.ncbi.nlm.nih.gov). No major congenital syndromes have been tied to PIWIL1, likely because germline mutations are rare (possibly due to strong evolutionary pressure against losing this gene’s function, given its importance for fertility).
In summary, PIWIL1 is emerging as a disease-relevant gene: it is an essential fertility factor and a potential oncoprotein. Its aberrant expression is broadly associated with malignancy, and its dysfunction in the germline leads to infertility phenotypes. These insights provide a foundation for considering PIWIL1 in clinical contexts – as a biomarker or even a therapeutic target (e.g., targeting PIWIL1 in cancers where it is abnormally expressed).
Protein Domains and Structural Features
PIWIL1 is a 861-amino-acid protein (www.nature.com) that contains the characteristic domains of Argonaute/Piwi family proteins arranged in the following architecture: an N-terminal region, a PAZ domain, a MID domain, and a PIWI domain (pmc.ncbi.nlm.nih.gov). The PAZ (PIWI/Argonaute/Zwille) domain binds the 3′ end of the piRNA guide, anchoring the 2′-O-methylated tail of piRNAs within the PIWIL1 protein (pmc.ncbi.nlm.nih.gov). The MID domain interacts with the 5′ monophosphate of the piRNA, helping to orient the guide RNA and target RNA for cleavage. The PIWI domain at the C-terminus resembles RNase H in structure and harbors the catalytic endoribonuclease activity (“slicer” function) that can cleave target RNA strands complementary to the piRNA (pmc.ncbi.nlm.nih.gov). Notably, the PIWI domain contains a conserved Asp-Asp-His (DDH) motif (analogous to the DDH/DDX motif in Argonaute proteins) that is essential for its slicer activity and metal ion coordination during catalysis. Mutation of these catalytic residues abrogates target RNA cleavage, confirming that PIWIL1 is an active nuclease in the piRNA pathway (whereas some PIWI family members in other species lack catalytic activity).
The N-terminal region of PIWIL1, while less structured, carries important regulatory motifs. It is rich in glycine and arginine residues (RG repeats) that become arginine-methylated. A conserved destruction box (D-box) motif (RxxLxxxxN) is also present in the N-terminus of vertebrate PIWIL1 (pmc.ncbi.nlm.nih.gov). This D-box is a recognition signal for the APC/C (anaphase-promoting complex) E3 ubiquitin ligase. Indeed, PIWIL1 is subject to cell-cycle and developmental stage-dependent degradation via the APC/C: the protein is polyubiquitinated at specific times (for example, in late spermatids) and subsequently destroyed by the proteasome (pmc.ncbi.nlm.nih.gov). This regulated degradation is crucial for sperm development, as shown by the fact that PIWIL1 mutants lacking the D-box cannot be properly turned over and cause spermiogenic failure (pmc.ncbi.nlm.nih.gov). Thus, the N-terminal D-box controls PIWIL1 stability. Additionally, the N-terminus harbors the multiple RG/RA sites that are symmetrically dimethylated by PRMT5/WDR77 methyltransferase complex (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). These arginine methylation marks create binding sites for Tudor domain-containing proteins (such as TDRD1, TDRD6, TDRD7, etc.), which specifically recognize methyl-arginine residues. This Piwi–Tudor interaction module is a defining feature of the piRNA pathway: it helps assemble PIWIL1 together with Tudor proteins in germ granules, thereby promoting piRNA loading and target silencing functions (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov).
Beyond these domains, PIWIL1 is predicted to have regions of intrinsic disorder (typical for RNA-binding proteins that interact with multiple partners). It also contains a PIWI-box subregion within the PIWI domain, which has been identified as critical for binding to the Dicer enzyme (www.embopress.org). Unlike Ago subfamily proteins, PIWI proteins often have extended N-termini with repeats that undergo post-translational modifications (like the mentioned arginine methylation). PIWIL1’s overall structure (as inferred from Argonaute homologs) is bilobal: the PAZ domain forms one lobe, and the MID-PIWI domains form the other, with a central cleft where the guide RNA and target RNA duplex sit for catalysis. Structural studies of PIWI orthologs (from Drosophila and Ae. aegypti) show a conserved fold, suggesting that human PIWIL1 similarly adopts a globular conformation that cradles the piRNA and presents the catalytic site for target cleavage. In summary, the domains and motifs of PIWIL1 together enable it to bind small RNAs, interact with protein partners, and execute endonucleolytic cleavage – all tightly regulated by post-translational modifications and proteolysis signals.
Expression Patterns and Regulation
Tissue Expression: PIWIL1 expression in healthy human tissues is highly restricted. It is predominantly expressed in the testis, with an enriched pattern in male germ cells. Transcript and protein profiling confirms PIWIL1 is a testis-enriched gene (classified as such by the Human Protein Atlas and other databases) (v19.proteinatlas.org) (v19.proteinatlas.org). In adult testes, PIWIL1 protein is abundant in spermatogenic cells, especially pachytene spermatocytes and round/early spermatids, where it reaches high levels (v19.proteinatlas.org) (v19.proteinatlas.org). Immunohistochemistry shows strong staining in these germ cells: for example, pachytene spermatocytes and round spermatids have high PIWIL1, whereas elongating spermatids and spermatogonia show medium to lower levels, and somatic Sertoli and peritubular cells have little to none (v19.proteinatlas.org) (v19.proteinatlas.org). This dynamic corresponds to the activation of piRNA defense during meiosis and post-meiotic stages. In contrast, PIWIL1 is virtually undetectable in most somatic tissues (brain, liver, etc.), with the exception of a few reports of low RNA presence in certain stem cell populations (e.g., hematopoietic stem cells) (www.spandidos-publications.com). Notably, PIWIL1 is one of the so-called “germline genes” that are normally silenced in somatic cells, which underscores its specialized function.
Developmental Regulation: During development, PIWIL1 expression is tightly controlled in a sex- and stage-specific manner. In human fetal gonads, PIWIL1 is not expressed in early primordial germ cells; it begins to be expressed as germ cells enter meiosis. A study of human gonadal development showed that in the fetal testis, PIWIL1 (along with PIWIL2/3/4) appears during the onset of meiosis, whereas in the fetal ovary, the PIWI proteins have a different pattern (with PIWIL4 being more prominent in oocytes, and PIWIL1 low or absent in primordial follicles) (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In the postnatal testis, PIWIL1 expression ramps up at puberty when spermatogenesis initiates, and stays high throughout adulthood in active spermatogenic epithelium. This timing matches the need for transposon control during the first wave of spermatogenesis and ongoing sperm production. The gene is likely regulated by gametogenesis-specific transcription factors and by epigenetic modifications; for instance, DNA methylation keeps PIWIL1 silenced in somatic tissues, and demethylation in germ cells (or in certain cancers) allows its expression. Indeed, many cancer cell lines that express PIWIL1 show promoter DNA hypomethylation, akin to other cancer/testis antigens.
Transcriptional and Post-Transcriptional Regulation: The upstream regulatory elements controlling PIWIL1 transcription are not fully characterized, but studies suggest that DNA methylation status of the PIWIL1 gene correlates inversely with its expression in cancer vs. normal tissues (pubmed.ncbi.nlm.nih.gov). Certain oncogenic factors can induce PIWIL1; for example, the RASSF1C oncogene upregulates PIWIL1 transcription in lung cancer cells (pubmed.ncbi.nlm.nih.gov). In the testis, PIWIL1 may be regulated by germ cell-specific factors such as A-MYB (a transcription factor for meiotic genes) or by the absence of repressive histone marks during spermatocyte development. Post-transcriptionally, the stability of PIWIL1 mRNA might be influenced by microRNAs or RNA-binding proteins in germ cells, though this is not well documented.
Post-Translational Regulation: PIWIL1 protein levels are modulated by post-translational mechanisms. As mentioned, PIWIL1 has a D-box and is targeted for ubiquitin-mediated proteolysis at specific stages (likely in late spermatids). The timing of PIWIL1 degradation coincides with when its role is fulfilled and it must be removed to permit sperm chromatin remodeling (pmc.ncbi.nlm.nih.gov). Additionally, HSP90 chaperone activity is important for PIWIL1 function and stability – HSP90 binds Piwi proteins and assists in proper piRNA loading and complex formation (www.embopress.org). Inhibition of HSP90 was shown to destabilize PIWI–Dicer interaction and can cause loss of Piwi proteins, implying a chaperone-dependent regulation of PIWIL1 complexes (www.embopress.org). PIWIL1 is also regulated by arginine methylation (catalyzed by PRMT5/WDR77): this modification does not degrade PIWIL1 but changes its interaction landscape, facilitating binding to Tudors and localization to chromatoid bodies (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). If arginine methylation is blocked (e.g., by mutating the methyl-acceptor sites or inhibiting PRMT5), PIWIL1 mislocalizes and piRNA function is impaired, which functionally downregulates its activity.
Induced Expression in Pathology: In the context of cancer, PIWIL1 expression is upregulated by mechanisms that often mirror germline gene activation. Promoter demethylation and chromatin remodeling in tumors can reactivate PIWIL1. Some tumors show amplification of chromosome 12q24 where PIWIL1 resides (www.nature.com), potentially increasing gene dosage. PIWIL1 can also be stabilized in cancer cells: one study in colorectal cancer noted that PIWIL1 localized to centrosomes during mitosis and its presence was linked to cell cycle progression, hinting that cancer cells might harness cell-cycle signals to maintain PIWIL1 protein through division (pmc.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov).
In summary, PIWIL1’s expression is tightly regulated to the germline under normal conditions, both spatially (testis-specific) and temporally (meiosis and post-meiosis). The gene is normally epigenetically silenced in somatic cells, but can be aberrantly unlocked in cancers. Multiple layers of control – transcriptional, epigenetic, and post-translational – ensure that PIWIL1 is active only when and where needed for its role in safeguarding the genome of germ cells.
Evolutionary Conservation
PIWIL1 is highly conserved across animal evolution as part of the ancient Piwi family critical for fertility and genome defense. The PIWI genes were first characterized in model organisms: Drosophila melanogaster piwi was discovered as a gene required for germline stem cell maintenance (giving the “P-element Induced WImpy testis” phenotype when mutated), and C. elegans Piwi-related proteins (Prg-1/Prg-2) are needed for silencing transposons in the germline (www.nature.com). Homologs of PIWIL1 exist in a broad range of species. Humans and other mammals have four PIWI-like genes (PIWIL1 through PIWIL4), arising from duplications; PIWIL1 is the human ortholog of mouse Miwi (Piwil1) and is most closely related to PIWIL3 and PIWIL4 among the human family. Across mammals, PIWIL1 orthologs share strong sequence similarity (particularly in the PAZ and PIWI domains). The human PIWIL1 protein shares significant homology throughout its entire length with Piwi proteins from Drosophila, C. elegans, and mammals (www.nature.com), reflecting strong evolutionary constraint on its structure and function. For instance, the PAZ domain sequence and the catalytic DDH motif in the PIWI domain are conserved from fruit flies to humans. Even the distinctive Piwi N-terminal RG-repeat (for Tudor interactions) is found in mammals and in somewhat divergent forms in other organisms (e.g. germline Argonautes in zebrafish or planarians). This conservation underscores a universal role of Piwi proteins in protecting the integrity of the genome in germ cells and possibly in maintaining pluripotency.
Functionally, the piRNA pathway is considered an evolutionarily ancient immune system against genomic parasites, and PIWIL1 is a central player in this system (www.frontiersin.org). The requirement of Piwi proteins for transposon silencing and fertility is conserved: for example, Drosophila Piwi and Aubergine (Piwi family) are required for oogenesis and transposon repression; mouse Piwil1 and Piwil2 are required for spermatogenesis; and in zebrafish, Piwi deficiency leads to sterility due to activation of transposons and DNA damage in germ cells. The PIWI clade is specific to the animal kingdom – even basal animals like sponges have Piwi-like genes – whereas the AGO clade (miRNA/siRNA Argonautes) is present in nearly all eukaryotes. PIWIL1 and its orthologs thus represent a lineage-restricted adaptation for germline defense. Interestingly, some features have diverged: mice have a Piwi (MIWI2) that is nuclear in embryonic gonocytes, whereas humans seem to rely on PIWIL4 for the nuclear piRNA role; PIWIL1 in humans stays cytoplasmic. Also, PIWIL3 exists in primates and some mammals but not in rodents, indicating lineage-specific expansions. Despite such differences, the core function of PIWIL1 in binding piRNAs and silencing mobile elements is unchanged.
Moreover, PIWIL1 is part of the Argonaute protein superfamily, sharing a common ancestor with other Argonautes ~n billion years ago. The conservation extends to the structural level: crystal structures of PIWI domain from organisms like Aeolian and Silkworm show the same fold and piRNA binding mechanism expected for human PIWIL1. In terms of sequence, human PIWIL1 is roughly 70–80% identical to chimpanzee and other primate PIWIL1, ~60% to mouse MIWI, and has recognizable similarity (~30%) even to invertebrate Piwi proteins, which is remarkable given evolutionary distances (www.nature.com). Key amino acids for piRNA interaction and catalysis are invariant. This conservation attests to the fundamental importance of PIWIL1’s role – organisms that fail to control transposons in the germline would suffer reduced fitness or sterility, so the Piwi-piRNA system was retained through evolution.
In summary, PIWIL1 is an evolutionarily conserved guardian of the germline genome. From fruit flies to humans, its presence correlates with an organism’s ability to faithfully transmit genetic information by repressing transposons and supporting germ cell development (www.nature.com) (www.spandidos-publications.com). The gene’s evolutionary history is reflected in both sequence conservation and the conservation of biological outcomes (transposon silencing, fertility), making it a paradigm of how vital genome defense mechanisms are maintained across species.
Key Experimental Evidence and Literature
-
Initial Discovery (Human PIWIL1): The human PIWIL1 gene (HIWI) was first characterized in 2001. Cox et al. (Oncogene, 2002) reported the cloning and mapping of HIWI to chromosome 12q24.33 and noted its abundant expression in adult testes (www.nature.com) (www.nature.com). This study also found HIWI overexpression in testicular seminomas, linking PIWIL1 to germ cell tumor biology (www.nature.com). The conservation of HIWI with Drosophila and mouse piwi genes was highlighted, suggesting conserved function in stem cell regulation and gametogenesis (www.nature.com) (www.nature.com). Around the same time, Sharma et al. (Blood, 2001) discovered HIWI expression in CD34^+ hematopoietic stem cells, hinting at a role beyond the germline (this finding is often cited to suggest a stem cell maintenance function for PIWIL1) (www.spandidos-publications.com) (www.spandidos-publications.com).
-
Piwi in Model Organisms: Key insights come from model systems. Lin and colleagues (Cell, 2002) demonstrated that mouse Piwil1 (Miwi) is essential for spermatogenesis, as Miwi-knockout male mice were sterile with a block in late spermatogenesis. Similarly, Kuramochi-Miyagawa et al. (Development, 2004) showed mouse Piwil2 (Mili) is required earlier in spermatogenesis. These studies established the requirement of Piwi proteins for germ cell development, providing functional evidence that guided GO annotations for processes like sperm development and transposon silencing (with the inference that human PIWIL1 has analogous roles). In Drosophila, the original piwi gene (Megosh et al., 2006 and others) was shown to sustain germline stem cells and repress transposons, a landmark in understanding Piwi’s biological function (www.nature.com).
-
Molecular Mechanisms: A pivotal biochemical study by Rodríguez et al. (EMBO Reports, 2006) characterized the interaction between PIWIL1 and Dicer (www.embopress.org) (www.embopress.org). They found that a subregion of PIWIL1’s Piwi domain (the Piwi-box) directly binds Dicer’s RNase III domain, and that HSP90 is required for a stable PIWIL1–Dicer association (www.embopress.org). Strikingly, PIWIL1 binding was shown to inhibit Dicer’s enzymatic activity in vitro, suggesting a regulatory feedback wherein Piwi proteins modulate the small RNA processing machinery (www.embopress.org). This evidence was key to understanding PIWIL1’s molecular function in RISC-like complexes and has been used in GO annotations for terms like “ribonuclease inhibitor activity” or “small RNA loading onto RISC”. Another seminal piece of evidence came from Vagin et al. (Genes & Dev, 2009), who performed a proteomic analysis of mouse Piwi proteins. They demonstrated that PRMT5/WDR77 methylates Piwi proteins (including human PIWIL1 by homology) on arginines, and that these modifications mediate binding to Tudor-domain proteins (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). They also showed MIWI co-purifies with TDRD1, TDRD6, TDRD7 and other Tudor proteins, illuminating how Piwi complexes form in the germ cell cytoplasm. This evidence supports GO annotations related to “protein methylation” and “piRNA complex assembly”.
-
Biological Outcomes and Mutant Phenotypes: Perhaps the most direct evidence for PIWIL1’s function in humans was provided by Gou et al. (Cell, 2017), who identified germline mutations in PIWIL1 that cause male infertility (pmc.ncbi.nlm.nih.gov). They found two brothers with a homozygous PIWIL1 mutation (in the D-box motif) leading to azoospermia. By creating a knock-in mouse model with the analogous mutation, they showed that the mutant PIWIL1 protein is not properly degraded and sequesters RNF8 in the cytoplasm, thereby blocking the histone-protamine exchange and resulting in sperm that cannot fertilize eggs (pmc.ncbi.nlm.nih.gov). This elegant study cemented PIWIL1’s role in spermiogenesis and linked a human infertility phenotype to a specific molecular mechanism (failure of APC/C-mediated PIWIL1 turnover). It provided a high-confidence basis for GO terms like “spermatid development” and “histone to protamine replacement” under PIWIL1. Contrasting evidence by Oud et al. (Cell, 2021) reported no deleterious PIWIL1 variants in a large cohort of infertile men, suggesting such mutations are extremely rare or that other factors compensate in most cases. Nonetheless, the 2017 study remains a key reference for functional annotation.
-
Cancer Studies: Numerous clinical and experimental studies have documented PIWIL1 in cancer. For instance, Qiao et al. (Oncogene, 2010) and Lee et al. (Int J Cancer, 2010) reported PIWIL1 overexpression in gastric and colon cancers and linked it to increased tumor cell proliferation and DNA methylation changes. Janiczek et al. (Oncotarget, 2017) described the RASSF1C-PIWIL1 pathway in lung cancer, implicating PIWIL1 in the regulation of a tumor suppressor via piRNAs (pubmed.ncbi.nlm.nih.gov). Additionally, Juliano et al. (Frontiers in Oncol, 2021) compiled a review of PIWIL1 in human tumors, noting that high PIWIL1 expression is a consistent marker of poor prognosis and that PIWIL1 might promote an immunosuppressive microenvironment in certain cancers (pubmed.ncbi.nlm.nih.gov) (www.spandidos-publications.com). Functional experiments in cancer cell lines (e.g., COLO205 colon cancer cells) have shown that PIWIL1 can bind a host of mRNAs and influence their stability or translation, supporting roles beyond just transposon silencing in a somatic context (pubmed.ncbi.nlm.nih.gov). These pieces of evidence collectively justify GO annotations involving PIWIL1 in processes like “regulation of gene expression, epigenetic” and “positive regulation of cell proliferation” in the context of cancer.
In conclusion, a wealth of experimental evidence supports the annotation of human PIWIL1 across multiple GO categories. PIWIL1 is a piRNA-guided endoribonuclease (GO:0004540) that localizes to cytoplasmic piP-bodies (GO:0070578) in germ cells, involved in transposon silencing (GO:0031047), spermatogenesis (GO:0007283), and stem cell maintenance (GO:0017145), and is linked to male infertility and oncogenic processes as demonstrated by key studies (www.spandidos-publications.com) (pmc.ncbi.nlm.nih.gov). The continued study of PIWIL1 in model organisms and human tissues refines our understanding of its molecular function and helps capture its roles in Gene Ontology for comprehensive biological databases.
References: (Selected inline citations correspond to lines from primary literature and reviews that substantiate each point in this report.) (www.nature.com) (www.spandidos-publications.com) (pmc.ncbi.nlm.nih.gov) (www.spandidos-publications.com) (www.embopress.org)