---
provider: falcon
model: Edison Scientific Literature
cached: false
start_time: '2026-06-12T04:55:02.446279'
end_time: '2026-06-12T05:07:06.548239'
duration_seconds: 724.1
template_file: templates/gene_research_go_focused.md
template_variables:
organism: human
gene_id: RNF170
gene_symbol: RNF170
uniprot_accession: Q96K19
protein_description: 'RecName: Full=E3 ubiquitin-protein ligase RNF170; EC=2.3.2.27
{ECO:0000269|PubMed:31076723}; AltName: Full=Putative LAG1-interacting protein;
AltName: Full=RING finger protein 170; AltName: Full=RING-type E3 ubiquitin transferase
RNF170 {ECO:0000305};'
gene_info: Name=RNF170;
organism_full: Homo sapiens (Human).
protein_family: Not specified in UniProt
protein_domains: DUF1232. (IPR010652); RNF170. (IPR038896); Znf-RING_euk. (IPR027370);
Znf_RING. (IPR001841); Znf_RING/FYVE/PHD. (IPR013083)
provider_config:
timeout: null
max_retries: 3
parameters:
allowed_domains: []
temperature: 0.1
max_embedded_images: 8
citation_count: 21
artifact_count: 2
artifact_sources:
edison_answer_artifacts: 1
edison_message_content: 1
artifacts:
- filename: artifact-00.md
path: RNF170-deep-research-falcon_artifacts/artifact-00.md
media_type: text/markdown
source: edison_answer_artifacts
data_storage_id: null
description: Edison artifact artifact-00
- filename: image-1.png
path: RNF170-deep-research-falcon_artifacts/image-1.png
media_type: image/png
source: edison_message_content
data_storage_id: null
description: '## Context ID: pqac-00000027 Figure 5, panels A and B, quantifies
the effect of RNF170 knockdown (depletion) on IP3R1 processing. As described in
the text (page'
---
## Question
# Gene Research for Functional Annotation
## ⚠️ CRITICAL: Gene/Protein Identification Context
**BEFORE YOU BEGIN RESEARCH:** You MUST verify you are researching the CORRECT gene/protein. Gene symbols can be ambiguous, especially for less well-characterized genes from non-model organisms.
### Target Gene/Protein Identity (from UniProt):
- **UniProt Accession:** Q96K19
- **Protein Description:** RecName: Full=E3 ubiquitin-protein ligase RNF170; EC=2.3.2.27 {ECO:0000269|PubMed:31076723}; AltName: Full=Putative LAG1-interacting protein; AltName: Full=RING finger protein 170; AltName: Full=RING-type E3 ubiquitin transferase RNF170 {ECO:0000305};
- **Gene Information:** Name=RNF170;
- **Organism (full):** Homo sapiens (Human).
- **Protein Family:** Not specified in UniProt
- **Key Domains:** DUF1232. (IPR010652); RNF170. (IPR038896); Znf-RING_euk. (IPR027370); Znf_RING. (IPR001841); Znf_RING/FYVE/PHD. (IPR013083)
### MANDATORY VERIFICATION STEPS:
1. **Check if the gene symbol "RNF170" matches the protein description above**
2. **Verify the organism is correct:** Homo sapiens (Human).
3. **Check if protein family/domains align with what you find in literature**
4. **If you find literature for a DIFFERENT gene with the same or similar symbol, STOP**
### If Gene Symbol is Ambiguous or You Cannot Find Relevant Literature:
**DO NOT PROCEED WITH RESEARCH ON A DIFFERENT GENE.** Instead:
- State clearly: "The gene symbol 'RNF170' is ambiguous or literature is limited for this specific protein"
- Explain what you found (e.g., "Found extensive literature on a different gene with the same symbol in a different organism")
- Describe the protein based ONLY on the UniProt information provided above
- Suggest that the protein function can be inferred from domain/family information
### Research Target:
Please provide a comprehensive research report on the gene **RNF170** (gene ID: RNF170, UniProt: Q96K19) in human.
The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
## Output
Question: You are an expert researcher providing comprehensive, well-cited information.
Provide detailed information focusing on:
1. Key concepts and definitions with current understanding
2. Recent developments and latest research (prioritize 2023-2024 sources)
3. Current applications and real-world implementations
4. Expert opinions and analysis from authoritative sources
5. Relevant statistics and data from recent studies
Format as a comprehensive research report with proper citations. Include URLs and publication dates where available.
Always prioritize recent, authoritative sources and provide specific citations for all major claims.
# Gene Research for Functional Annotation
## ⚠️ CRITICAL: Gene/Protein Identification Context
**BEFORE YOU BEGIN RESEARCH:** You MUST verify you are researching the CORRECT gene/protein. Gene symbols can be ambiguous, especially for less well-characterized genes from non-model organisms.
### Target Gene/Protein Identity (from UniProt):
- **UniProt Accession:** Q96K19
- **Protein Description:** RecName: Full=E3 ubiquitin-protein ligase RNF170; EC=2.3.2.27 {ECO:0000269|PubMed:31076723}; AltName: Full=Putative LAG1-interacting protein; AltName: Full=RING finger protein 170; AltName: Full=RING-type E3 ubiquitin transferase RNF170 {ECO:0000305};
- **Gene Information:** Name=RNF170;
- **Organism (full):** Homo sapiens (Human).
- **Protein Family:** Not specified in UniProt
- **Key Domains:** DUF1232. (IPR010652); RNF170. (IPR038896); Znf-RING_euk. (IPR027370); Znf_RING. (IPR001841); Znf_RING/FYVE/PHD. (IPR013083)
### MANDATORY VERIFICATION STEPS:
1. **Check if the gene symbol "RNF170" matches the protein description above**
2. **Verify the organism is correct:** Homo sapiens (Human).
3. **Check if protein family/domains align with what you find in literature**
4. **If you find literature for a DIFFERENT gene with the same or similar symbol, STOP**
### If Gene Symbol is Ambiguous or You Cannot Find Relevant Literature:
**DO NOT PROCEED WITH RESEARCH ON A DIFFERENT GENE.** Instead:
- State clearly: "The gene symbol 'RNF170' is ambiguous or literature is limited for this specific protein"
- Explain what you found (e.g., "Found extensive literature on a different gene with the same symbol in a different organism")
- Describe the protein based ONLY on the UniProt information provided above
- Suggest that the protein function can be inferred from domain/family information
### Research Target:
Please provide a comprehensive research report on the gene **RNF170** (gene ID: RNF170, UniProt: Q96K19) in human.
The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
## Research report: Human RNF170 (UniProt Q96K19) — functional annotation and current understanding
### 0) Target verification (critical identity check)
The literature reviewed here matches the UniProt entry **Q96K19** for **Homo sapiens RNF170**, described experimentally as an **ER membrane, RING-type E3 ubiquitin ligase** that regulates **ER-associated degradation (ERAD)** of activated **inositol 1,4,5-trisphosphate receptors (IP3Rs)** and is genetically linked to **hereditary spastic paraplegia (HSP)**. (lu2011rnf170proteinan pages 4-5, wagner2019biallelicvariantsin pages 6-7)
### 1) Key concepts and definitions (current understanding)
#### 1.1 RNF170 as a RING-type E3 ubiquitin ligase
RNF170 is a **RING-HC E3 ubiquitin ligase**: it promotes transfer of ubiquitin from an E2 enzyme to a substrate protein, typically creating polyubiquitin chains that control substrate fate (often proteasomal degradation). In vitro, immunopurified RNF170 catalyzes ubiquitin conjugation in the presence of **E1 (UBE1), E2 (UbcH5b), and ubiquitin**, producing a high-molecular-weight ubiquitin “smear,” consistent with intrinsic ligase activity. (lu2011rnf170proteinan pages 4-5, lu2011rnf170proteinan pages 3-4)
A key mechanistic validation is that mutating zinc-coordinating residues in the RING domain (**Cys101/His103**) abolishes ligase activity, establishing the catalytic dependence on the RING motif. (lu2011rnf170proteinan pages 3-4)
#### 1.2 ERAD and stimulus-coupled degradation of activated IP3 receptors
Activated IP3Rs undergo rapid down-regulation via the **ubiquitin–proteasome pathway**, a form of ERAD-like quality control applied to an activated signaling channel. RNF170 has been described as (at the time of foundational studies) the only E3 ligase directly demonstrated to mediate IP3R ubiquitination in this context, with recruitment to activated IP3Rs preceding robust polyubiquitination and downstream processing. (lu2011rnf170proteinan pages 5-6, lu2011rnf170proteinan pages 3-4)
#### 1.3 ERLIN scaffolds as organizing platforms for RNF170
ERLIN1/2 are ER membrane SPFH-family proteins that assemble into large oligomeric scaffolds in cholesterol-enriched ER nanodomains. These scaffolds act as platforms to recruit factors including RNF170, and (per 2024 work) can bridge RNF170 to other clients such as TMUB1-L, connecting ubiquitin machinery to ER lipid organization and secretory function. (veronese2024erlin12scaffoldsbridge pages 1-2, veronese2024erlin12scaffoldsbridge pages 12-12)
### 2) Molecular function: enzymatic reaction, substrate specificity, and mechanism
#### 2.1 Subcellular localization and topology
RNF170 is an **integral ER membrane protein**, with topology predictions and biochemical fractionation placing the **N-terminus in the ER lumen** and the **RING domain/C-terminus in the cytosol**, consistent with ubiquitination of cytosolic lysines on substrates such as IP3Rs. (lu2011rnf170proteinan pages 3-4, lu2011rnf170proteinan pages 4-5)
#### 2.2 Core substrate: IP3 receptors (human cellular evidence)
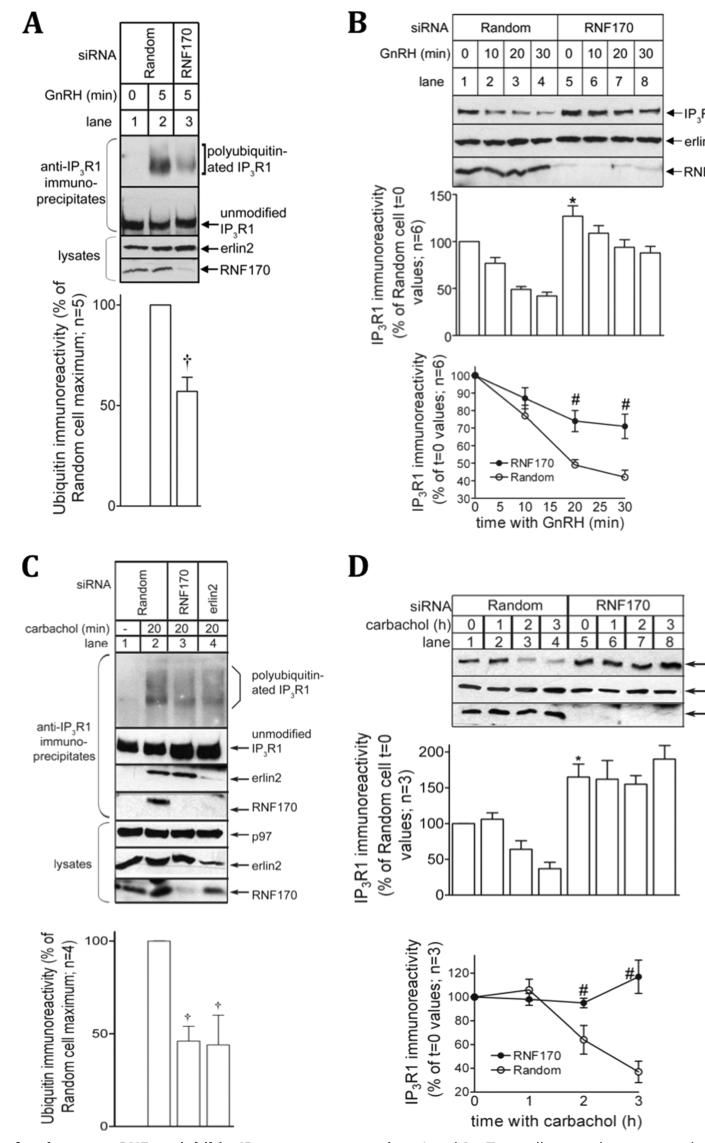
Foundational mechanistic work demonstrated that RNF170 rapidly associates with **activated IP3R1**, and depletion of RNF170 reduces stimulus-induced IP3R1 polyubiquitination and slows receptor down-regulation. (lu2011rnf170proteinan pages 4-5, lu2011rnf170proteinan pages 3-4)
Quantitatively, RNF170 knockdown reduced agonist-induced **IP3R1 polyubiquitination to 57 ± 7%** of control and inhibited **IP3R1 down-regulation by ~50%**, while also increasing basal IP3R1 levels. (lu2011rnf170proteinan pages 4-5, lu2011rnf170proteinan media b065ce9e)
Mechanistically, RNF170 recruitment to activated IP3Rs occurs via constitutive association with ERLIN1/2, which are required for efficient RNF170–IP3R coupling. (lu2011rnf170proteinan pages 8-9, lu2011rnf170proteinan pages 5-6)
#### 2.3 Additional substrate evidence: TLR3 (murine innate immunity; mechanistic details)
A separate, well-developed mechanistic line of evidence (in murine systems) identifies RNF170 as a negative regulator of innate immunity through direct targeting of **TLR3**. RNF170 binds the TLR3 TIR domain and catalyzes **K48-linked polyubiquitination** at **TLR3 Lys766**, promoting **proteasomal degradation**; this requires intact RNF170 RING residues (C101/H103) and is blocked by proteasome inhibition (MG132), supported by CHX chase assays and in vivo phenotyping in Rnf170−/− mice. (song2020e3ubiquitinligase pages 6-7, song2020e3ubiquitinligase pages 7-9)
Because this primary study is performed in **murine cells and mice**, extension to human physiology should be made cautiously, although the TLR3 K766 site is noted as conserved between mouse and human in the primary study. (song2020e3ubiquitinligase pages 7-9)
### 3) Protein–protein interactions and complexes
#### 3.1 ERLIN1/2–RNF170–IP3R module
RNF170 is reported to be constitutively associated with the **ERLIN1/2 complex**, which recruits RNF170 to activated IP3Rs to drive ubiquitination and proteasomal processing. (lu2011rnf170proteinan pages 8-9, lu2011rnf170proteinan pages 5-6)
Proteomics of ER-resident E3 ligase complexes has independently recovered RNF170 with high-confidence interactors including **ERLIN1, ERLIN2, and ITPR3**, supporting the recurring architecture of this module in ERAD/proteostasis networks. (lari2016resolutionofproteotoxic pages 111-115)
#### 3.2 2024 expansion: ERLIN scaffolds bridging RNF170 and TMUB1-L (cholesterol/secretory pathway link)
A major 2024 development is the proposal that ERLIN1/2 scaffolds bind a conserved luminal N-terminal motif present in RNF170 and the long isoform of **TMUB1 (TMUB1-L)**, bridging these proteins in cholesterol-rich ER nanodomains. AlphaFold-Multimer modeling supports interaction interfaces between the conserved motifs and adjacent ERLIN subunits. (veronese2024erlin12scaffoldsbridge pages 4-6, veronese2024erlin12scaffoldsbridge pages 12-12)
Functionally, ERLIN loss (DKO) leads to increased cholesterol esterification and lipid droplet accumulation, ER tubule collapse, Golgi fragmentation, and impaired secretory trafficking, with rescue by ERLIN re-expression or pharmacologic inhibition of cholesterol esterification (SOAT1 inhibitor **avasimibe**). (veronese2024erlin12scaffoldsbridge pages 10-12, veronese2024erlin12scaffoldsbridge pages 12-12)
### 4) Pathways and biological roles
#### 4.1 ER calcium signaling homeostasis via IP3R turnover
By controlling stimulus-dependent degradation of activated IP3Rs, RNF170 is positioned as a regulator of **ER Ca2+ release signaling dynamics**, linking receptor activation state to ERAD engagement. (wagner2019biallelicvariantsin pages 6-7, lu2011rnf170proteinan pages 5-6)
In patient-derived or engineered RNF170-deficient models, increased basal IP3R levels (cell-type-specific isoforms) and failure of stimulus-dependent IP3R reduction support the concept that RNF170 constrains IP3R abundance and signaling. (wagner2019biallelicvariantsin pages 6-7)
#### 4.2 Innate immune modulation through TLR3 control (murine evidence)
RNF170-mediated proteasomal removal of TLR3 reduces downstream IRF3/NF-κB/STAT1-linked transcriptional activity and cytokine production in TLR3 pathways, positioning RNF170 as a negative regulator of TLR3-dependent antiviral inflammation in murine models. (song2020e3ubiquitinligase pages 6-7, song2020e3ubiquitinligase pages 7-9)
2024 reviews summarize RNF170 in this role primarily by citing the 2020 primary study, without adding additional mechanistic detail in the excerpted sections. (wang2024hostfactorsmodulate pages 13-14, li2024thernabindingproteins pages 12-13)
### 5) Human disease associations and genetics (with recent emphasis)
#### 5.1 Autosomal recessive hereditary spastic paraplegia from biallelic RNF170 variants
A key primary genetics study provides evidence that **biallelic RNF170 variants** are a cause of **autosomal recessive HSP**, supported by functional validation across patient fibroblasts, neuronal cells, and zebrafish assays. (wagner2019biallelicvariantsin pages 6-7)
Mechanistically, patient-derived fibroblasts fail to show physiological stimulus-dependent degradation of IP3R-3, and RNF170 knockout neuronal SH-SY5Y cells show elevated basal IP3R-1 with rescue by re-expression of wild-type RNF170. (wagner2019biallelicvariantsin pages 6-7)
Quantitatively, this study reports zebrafish developmental phenotypes and statistical testing (e.g., embryo length and eye size differences with adjusted P < 0.0001) supporting functional impairment of tested patient variants relative to RNF170wt. (wagner2019biallelicvariantsin pages 6-7)
#### 5.2 Database-level disease-target support (Open Targets)
Open Targets disease–target evidence links **RNF170** to **hereditary spastic paraplegia** and **complex hereditary spastic paraplegia**, with literature evidence pointing to PubMed ID 31636353 / PMC6803694. (OpenTargets Search: hereditary spastic paraplegia,spastic paraplegia,sensory ataxia,spinocerebellar ataxia,neuroaxonal dystrophy-RNF170)
#### 5.3 Expert synthesis (2024): ER homeostasis and ubiquitin-mediated quality control as convergent neurogenetic mechanisms
A 2024 Trends in Neurosciences review emphasizes ER structure and protein quality-control pathways as central mechanisms in inherited neuropathies, HSP, and ataxias, and explicitly notes RNF170 as an E3 ligase associated with recessive HSP and inherited peripheral neuropathy. (vondel2024overarchingpathomechanismsin pages 5-8)
Consistent pathway-context evidence from 2024 human genetics (ERLIN1 series) supports the broader ERLIN1/2–RNF170–IP3R ERAD module as disease-relevant: in a cohort of **13 individuals** with biallelic ERLIN1 variants (SPG62), the authors describe ERLIN1/2 as recruiting RNF170 to degrade activated IP3R1 and provide cohort-level statistics (e.g., mean onset 1.8 years; corpus callosum anomalies 5/13; founder splice variant in 6 individuals). Although ERLIN1 is not RNF170, these data reinforce the clinical relevance of the RNF170-centered module. (cogan2024biallelicvariantsin pages 1-6, cogan2024biallelicvariantsin pages 12-16)
### 6) Recent developments (prioritizing 2023–2024)
#### 6.1 2024: Linking the ERLIN–RNF170 axis to cholesterol esterification and secretory pathway control
Veronese et al. (Life Science Alliance; **May 2024**) propose that ERLIN scaffolds directly bind cholesterol and restrain cholesterol esterification, thereby maintaining ER cholesterol accessibility for ER→Golgi transport; the ERLIN scaffolds concurrently organize RNF170 and TMUB1-L via conserved luminal motifs. Loss of ERLINs increases cholesterol esterification (including CE 18:1), enlarges lipid droplets, fragments Golgi, and disrupts secretory trafficking and migration phenotypes; SOAT1 inhibition by **avasimibe** rescues multiple phenotypes. (veronese2024erlin12scaffoldsbridge pages 10-12, veronese2024erlin12scaffoldsbridge pages 12-12)
Quantitative details reported include proteomics showing a trend toward increased SOAT1 abundance (log2FC 0.40; q=0.07) and phenotyping with N=3 biological replicates and large cell counts (≥130 cells for lipid droplet size; ≥340 for Golgi fragmentation) analyzed by ANOVA with Tukey post hoc tests. (veronese2024erlin12scaffoldsbridge pages 12-12, veronese2024erlin12scaffoldsbridge pages 10-12)
#### 6.2 2024: Updated clinical genetics context for the ERLIN–RNF170–IP3R module
Cogan et al. (Human Genetics; **Oct 2024**) frame ERLIN1/2 as ERAD organizers that associate with RNF170 to target activated IP3Rs, and expand genotype–phenotype characterization in ERLIN-related HSP. This strengthens the view (also consistent with RNF170 HSP genetics) that motor neurons are unusually sensitive to disruption of this ERAD-linked calcium signaling module. (cogan2024biallelicvariantsin pages 1-6, cogan2024biallelicvariantsin pages 12-16)
### 7) Current applications and real-world implementations
#### 7.1 Genetic diagnostics and variant interpretation
RNF170 is now a disease gene supported by primary genetics and functional validation for HSP, supporting its inclusion in **neurogenetic diagnostic panels** for spastic paraplegia/ataxia phenotypes and for variant interpretation workflows. (wagner2019biallelicvariantsin pages 6-7, vondel2024overarchingpathomechanismsin pages 5-8)
#### 7.2 Therapeutic implications: pathway prioritization and mechanism-informed interventions
The RNF170-centered mechanism suggests two translationally relevant intervention layers:
1) **IP3R signaling / ER Ca2+ homeostasis**: human genetics and functional data prioritize the IP3R degradation/signaling axis as a candidate therapeutic pathway in HSP. (wagner2019biallelicvariantsin pages 6-7)
2) **ER cholesterol esterification / secretory pathway**: 2024 mechanistic work identifies cholesterol esterification control as a modifiable node in ERLIN-module dysfunction, with **SOAT1 inhibition (avasimibe)** rescuing cell phenotypes (lipid droplet size, Golgi morphology, and gene-expression readouts). This is not a direct RNF170-targeting therapy, but it provides a mechanism-based proof-of-concept for pharmacologic modulation of ERLIN–RNF170-associated ER nanodomain functions. (veronese2024erlin12scaffoldsbridge pages 10-12, veronese2024erlin12scaffoldsbridge pages 12-12)
### 8) Expert opinions and analysis (authoritative sources)
A 2024 expert review in Trends in Neurosciences highlights ER structure and protein quality-control pathways (UPS/autophagy) as overarching mechanisms in inherited neuropathies, HSP, and ataxias, and includes RNF170 among implicated E3 ligases, supporting a convergent “ER homeostasis / QC” framing for RNF170-related disease mechanisms. (vondel2024overarchingpathomechanismsin pages 5-8)
Genetics-focused 2024 work on ERLIN1 (SPG62) similarly emphasizes that the ERLIN1/2–RNF170 module is expected to impair IP3R1 turnover and Ca2+ signaling, providing additional disease-mechanism coherence at the pathway level. (cogan2024biallelicvariantsin pages 12-16)
### 9) Recent statistics and quantitative data highlights
- **RNF170 knockdown in human cell models**: stimulus-induced IP3R1 polyubiquitination reduced to **57 ± 7%** of control and IP3R1 down-regulation inhibited by **~50%**. (lu2011rnf170proteinan pages 4-5, lu2011rnf170proteinan media b065ce9e)
- **RNF170 loss-of-function cellular phenotypes in HSP genetics study**: increased basal IP3R-1 in RNF170 KO SH-SY5Y cells with rescue; zebrafish phenotypes with adjusted P values including **P < 0.0001** in quantitative measures. (wagner2019biallelicvariantsin pages 6-7)
- **2024 ERLIN scaffold phenotyping/omics**: SOAT1 proteomics trend (log2FC **0.40**, q=**0.07**) and high-powered imaging quantification with **≥130** and **≥340** cells across readouts (N=3 biological replicates). (veronese2024erlin12scaffoldsbridge pages 12-12, veronese2024erlin12scaffoldsbridge pages 10-12)
### 10) Quick reference summary table
The following table consolidates the most evidence-supported statements for functional annotation.
| Category | Summary |
|---|---|
| Identity/domains | - Human **RNF170** corresponds to UniProt **Q96K19**, a **RING-type E3 ubiquitin ligase** studied as an ER-membrane regulator of protein turnover and signaling.
- Foundational work defines RNF170 as a **257 aa RING-HC protein** with catalytic dependence on **Cys101/His103**; this matches the UniProt RING-domain annotation and ER-associated function. (lu2011rnf170proteinan pages 4-5, lu2011rnf170proteinan pages 3-4) |
| Localization/topology | - RNF170 is an **integral endoplasmic reticulum (ER) membrane** protein.
- Topology predictions and biochemical fractionation place its **N-terminus in the ER lumen** and its **RING domain/C-terminus in the cytosol**, positioning the catalytic machinery to ubiquitinate cytosolic receptor lysines.
- RNF170 localizes in **ERLIN-positive cholesterol-rich ER nanodomains**. (lu2011rnf170proteinan pages 4-5, lu2011rnf170proteinan pages 3-4, veronese2024erlin12scaffoldsbridge pages 1-2) |
| Core enzymatic activity | - RNF170 catalyzes **E3 ubiquitin transfer** in vitro using **UBE1 + UbcH5b** and ubiquitin, generating a high-molecular-weight ubiquitin smear typical of ligase activity.
- Catalysis is lost with **RING mutant C101S/H103A**, confirming dependence on the RING domain.
- Functionally, RNF170 promotes **proteasome-directed ER-associated degradation (ERAD)** of selected membrane/signaling proteins. (lu2011rnf170proteinan pages 4-5, lu2011rnf170proteinan pages 3-4, song2020e3ubiquitinligase pages 6-7) |
| Key substrates | - Best-supported human substrate class: **activated IP3 receptors (IP3Rs)**, especially **IP3R1/IP3R3**, which undergo RNF170-dependent ubiquitination and degradation after stimulation.
- In murine innate immunity studies, RNF170 also targets **TLR3**, catalyzing **K48-linked polyubiquitination** at **K766** to drive proteasomal degradation.
- TLR3 regulation is strongly supported experimentally, but species context should be noted because the primary paper is in **murine cells/mice**. (wagner2019biallelicvariantsin pages 6-7, lu2011rnf170proteinan pages 4-5, song2020e3ubiquitinligase pages 6-7, song2020e3ubiquitinligase pages 1-2, song2020e3ubiquitinligase pages 7-9) |
| Key interactors/complex | - RNF170 is constitutively associated with the **ERLIN1/2 complex**, which recruits it to activated IP3Rs.
- It is also found in complexes containing **p97/VCP-associated ERAD machinery** and, in 2024 work, **TMUB1-L** bridged by ERLIN scaffolds.
- Proteomics and co-IP studies repeatedly enrich **ERLIN1, ERLIN2, ITPR3, TMUB1, TMEM259** with RNF170-centered complexes. (lu2011rnf170proteinan pages 8-9, lari2016resolutionofproteotoxic pages 111-115, veronese2024erlin12scaffoldsbridge pages 1-2, veronese2024erlin12scaffoldsbridge pages 12-12) |
| Pathways | - RNF170 acts in **ERAD/proteostasis**, especially stimulus-coupled degradation of activated **IP3Rs**.
- Through IP3R turnover, RNF170 regulates **ER Ca2+ release signaling** and is therefore connected to neurodegeneration-relevant calcium homeostasis pathways.
- Separate immune work places RNF170 in **TLR3 innate immune signaling** as a negative regulator limiting IRF3/NF-kB/STAT1 outputs by degrading TLR3. (wagner2019biallelicvariantsin pages 6-7, lu2011rnf170proteinan pages 5-6, song2020e3ubiquitinligase pages 6-7, vondel2024overarchingpathomechanismsin pages 5-8) |
| Disease associations | - **Biallelic loss-of-function RNF170 variants** cause **autosomal recessive hereditary spastic paraplegia (HSP)**, with Open Targets evidence mapped to hereditary/complex HSP.
- Earlier literature also linked a **dominant sensory ataxia** phenotype to RNF170 mutation, and mouse knockout models show age-dependent gait abnormalities.
- Expert 2024 synthesis places RNF170 within an **ER homeostasis/quality-control disease module** shared across HSP, ataxia, and related neurodegenerative disorders. (wagner2019biallelicvariantsin pages 6-7, lu2011rnf170proteinan pages 8-9, OpenTargets Search: hereditary spastic paraplegia,spastic paraplegia,sensory ataxia,spinocerebellar ataxia,neuroaxonal dystrophy-RNF170, vondel2024overarchingpathomechanismsin pages 5-8) |
| Recent 2024 developments | - **Veronese et al. 2024** propose that **ERLIN1/2 ring-like scaffolds** bind a conserved luminal motif in **RNF170 and TMUB1-L**, organizing them in cholesterol-rich ER nanodomains.
- This work expands RNF170 biology beyond IP3R degradation by linking the ERLIN–RNF170 module to **cholesterol esterification control, Golgi morphology, and secretory pathway regulation**.
- 2024 disease reviews and genetics papers further emphasize the **ERLIN1/2–RNF170–IP3R** axis as a recurrent neurogenetic mechanism in spastic paraplegia/ataxia. (veronese2024erlin12scaffoldsbridge pages 12-12, veronese2024erlin12scaffoldsbridge pages 4-6, veronese2024erlin12scaffoldsbridge pages 10-12, cogan2024biallelicvariantsin pages 1-6, vondel2024overarchingpathomechanismsin pages 5-8) |
| Quantitative data points | - **RNF170 knockdown** reduced agonist-induced **IP3R1 polyubiquitination to 57 ± 7% of control**, inhibited IP3R1 down-regulation by roughly **~50%**, and increased basal IP3R1 by **~27 ± 11%**. (lu2011rnf170proteinan pages 4-5)
- In RNF170-deficient models, **IP3R3** rose by about **~4-fold** in patient fibroblasts and **IP3R1** by **~1.8-fold** in RNF170-knockout SH-SY5Y cells. (gehweiler2024rnf170anoveldisease pages 68-72)
- 2024 ERLIN-loss proteomics/lipidomics reported **SOAT1 log2FC = 0.40, q = 0.07** and phenotyping with **N = 3** biological replicates, including **≥130 cells** for lipid-droplet size and **≥340 cells** for Golgi fragmentation. (veronese2024erlin12scaffoldsbridge pages 12-12, veronese2024erlin12scaffoldsbridge pages 10-12) |
*Table: This table condenses the key verified facts about human RNF170 (UniProt Q96K19), including its identity, ER localization, E3 ligase function, substrates, complexes, disease relevance, and notable 2024 advances. It is useful as a quick-reference annotation scaffold anchored to specific evidence contexts.*
### 11) Key figure evidence
A figure supporting RNF170’s quantitative effect on IP3R1 ubiquitination and down-regulation is available from Lu et al. 2011 (Figure 5A-B). (lu2011rnf170proteinan media b065ce9e)
### 12) Limitations and open questions
- **Human vs murine innate-immunity substrate**: the strongest mechanistic TLR3 evidence is murine; human relevance is plausible (conserved K766) but not established here as a human-cell mechanism within the retrieved excerpts. (song2020e3ubiquitinligase pages 7-9)
- **Direct RNF170 catalytic specificity determinants** (e.g., how RNF170 selects IP3Rs vs other ER membrane clients, and how ERLIN nanodomains tune substrate choice) remain incompletely resolved; 2024 work suggests a broader organizational role for ERLIN scaffolds that may impact RNF170 client selection. (veronese2024erlin12scaffoldsbridge pages 12-12)
### 13) Core references (URLs and publication dates)
- Lu JP et al. **“RNF170 Protein, an Endoplasmic Reticulum Membrane Ubiquitin Ligase, Mediates Inositol 1,4,5-Trisphosphate Receptor Ubiquitination and Degradation”**. *Journal of Biological Chemistry*. **Jul 2011**. https://doi.org/10.1074/jbc.m111.251983 (lu2011rnf170proteinan pages 4-5, lu2011rnf170proteinan pages 3-4)
- Wagner M et al. **“Bi-allelic variants in RNF170 are associated with hereditary spastic paraplegia”**. *Nature Communications*. **Oct 2019**. https://doi.org/10.1038/s41467-019-12620-9 (wagner2019biallelicvariantsin pages 6-7)
- Song X et al. **“E3 ubiquitin ligase RNF170 inhibits innate immune responses by targeting and degrading TLR3 in murine cells”**. *Cellular & Molecular Immunology*. **May 2020**. https://doi.org/10.1038/s41423-019-0236-y (song2020e3ubiquitinligase pages 6-7)
- Veronese M et al. **“ERLIN1/2 scaffolds bridge TMUB1 and RNF170 and restrict cholesterol esterification to regulate the secretory pathway”**. *Life Science Alliance*. **May 2024**. https://doi.org/10.26508/lsa.202402620 (veronese2024erlin12scaffoldsbridge pages 10-12)
- Van de Vondel L et al. **“Overarching pathomechanisms in inherited peripheral neuropathies, spastic paraplegias, and cerebellar ataxias”**. *Trends in Neurosciences*. **Mar 2024**. https://doi.org/10.1016/j.tins.2024.01.004 (vondel2024overarchingpathomechanismsin pages 5-8)
- Cogan G et al. **“Biallelic variants in ERLIN1: a series of 13 individuals with spastic paraparesis.”** *Human Genetics*. **Oct 2024**. https://doi.org/10.1007/s00439-024-02702-0 (cogan2024biallelicvariantsin pages 1-6, cogan2024biallelicvariantsin pages 12-16)
- Open Targets Platform (RNF170–HSP association evidence; literature includes PMID 31636353). Accessed via tool output. (OpenTargets Search: hereditary spastic paraplegia,spastic paraplegia,sensory ataxia,spinocerebellar ataxia,neuroaxonal dystrophy-RNF170)
References
1. (lu2011rnf170proteinan pages 4-5): Justine P. Lu, Yuan Wang, Danielle A. Sliter, Margaret M.P. Pearce, and Richard J.H. Wojcikiewicz. Rnf170 protein, an endoplasmic reticulum membrane ubiquitin ligase, mediates inositol 1,4,5-trisphosphate receptor ubiquitination and degradation. Journal of Biological Chemistry, 286:24426-24433, Jul 2011. URL: https://doi.org/10.1074/jbc.m111.251983, doi:10.1074/jbc.m111.251983. This article has 134 citations and is from a domain leading peer-reviewed journal.
2. (wagner2019biallelicvariantsin pages 6-7): Matias Wagner, Daniel P. S. Osborn, Ina Gehweiler, Maike Nagel, Ulrike Ulmer, Somayeh Bakhtiari, Rim Amouri, Reza Boostani, Faycal Hentati, Maryam M. Hockley, Benedikt Hölbling, Thomas Schwarzmayr, Ehsan Ghayoor Karimiani, Christoph Kernstock, Reza Maroofian, Wolfgang Müller-Felber, Ege Ozkan, Sergio Padilla-Lopez, Selina Reich, Jennifer Reichbauer, Hossein Darvish, Neda Shahmohammadibeni, Abbas Tafakhori, Katharina Vill, Stephan Zuchner, Michael C. Kruer, Juliane Winkelmann, Yalda Jamshidi, and Rebecca Schüle. Bi-allelic variants in rnf170 are associated with hereditary spastic paraplegia. Nature Communications, Oct 2019. URL: https://doi.org/10.1038/s41467-019-12620-9, doi:10.1038/s41467-019-12620-9. This article has 62 citations and is from a highest quality peer-reviewed journal.
3. (lu2011rnf170proteinan pages 3-4): Justine P. Lu, Yuan Wang, Danielle A. Sliter, Margaret M.P. Pearce, and Richard J.H. Wojcikiewicz. Rnf170 protein, an endoplasmic reticulum membrane ubiquitin ligase, mediates inositol 1,4,5-trisphosphate receptor ubiquitination and degradation. Journal of Biological Chemistry, 286:24426-24433, Jul 2011. URL: https://doi.org/10.1074/jbc.m111.251983, doi:10.1074/jbc.m111.251983. This article has 134 citations and is from a domain leading peer-reviewed journal.
4. (lu2011rnf170proteinan pages 5-6): Justine P. Lu, Yuan Wang, Danielle A. Sliter, Margaret M.P. Pearce, and Richard J.H. Wojcikiewicz. Rnf170 protein, an endoplasmic reticulum membrane ubiquitin ligase, mediates inositol 1,4,5-trisphosphate receptor ubiquitination and degradation. Journal of Biological Chemistry, 286:24426-24433, Jul 2011. URL: https://doi.org/10.1074/jbc.m111.251983, doi:10.1074/jbc.m111.251983. This article has 134 citations and is from a domain leading peer-reviewed journal.
5. (veronese2024erlin12scaffoldsbridge pages 1-2): Matteo Veronese, Sebastian Kallabis, Alexander Tobias Kaczmarek, Anushka Das, Lennart Robers, Simon Schumacher, Alessia Lofrano, Susanne Brodesser, Stefan Müller, Kay Hofmann, Marcus Krüger, and Elena I Rugarli. Erlin1/2 scaffolds bridge tmub1 and rnf170 and restrict cholesterol esterification to regulate the secretory pathway. Life Science Alliance, 7:e202402620, May 2024. URL: https://doi.org/10.26508/lsa.202402620, doi:10.26508/lsa.202402620. This article has 8 citations and is from a peer-reviewed journal.
6. (veronese2024erlin12scaffoldsbridge pages 12-12): Matteo Veronese, Sebastian Kallabis, Alexander Tobias Kaczmarek, Anushka Das, Lennart Robers, Simon Schumacher, Alessia Lofrano, Susanne Brodesser, Stefan Müller, Kay Hofmann, Marcus Krüger, and Elena I Rugarli. Erlin1/2 scaffolds bridge tmub1 and rnf170 and restrict cholesterol esterification to regulate the secretory pathway. Life Science Alliance, 7:e202402620, May 2024. URL: https://doi.org/10.26508/lsa.202402620, doi:10.26508/lsa.202402620. This article has 8 citations and is from a peer-reviewed journal.
7. (lu2011rnf170proteinan media b065ce9e): Justine P. Lu, Yuan Wang, Danielle A. Sliter, Margaret M.P. Pearce, and Richard J.H. Wojcikiewicz. Rnf170 protein, an endoplasmic reticulum membrane ubiquitin ligase, mediates inositol 1,4,5-trisphosphate receptor ubiquitination and degradation. Journal of Biological Chemistry, 286:24426-24433, Jul 2011. URL: https://doi.org/10.1074/jbc.m111.251983, doi:10.1074/jbc.m111.251983. This article has 134 citations and is from a domain leading peer-reviewed journal.
8. (lu2011rnf170proteinan pages 8-9): Justine P. Lu, Yuan Wang, Danielle A. Sliter, Margaret M.P. Pearce, and Richard J.H. Wojcikiewicz. Rnf170 protein, an endoplasmic reticulum membrane ubiquitin ligase, mediates inositol 1,4,5-trisphosphate receptor ubiquitination and degradation. Journal of Biological Chemistry, 286:24426-24433, Jul 2011. URL: https://doi.org/10.1074/jbc.m111.251983, doi:10.1074/jbc.m111.251983. This article has 134 citations and is from a domain leading peer-reviewed journal.
9. (song2020e3ubiquitinligase pages 6-7): Xiaoqi Song, Shuo Liu, Wen-die Wang, Zhong-fei Ma, Xuetao Cao, and Minghong Jiang. E3 ubiquitin ligase rnf170 inhibits innate immune responses by targeting and degrading tlr3 in murine cells. Cellular & Molecular Immunology, 17:865-874, May 2020. URL: https://doi.org/10.1038/s41423-019-0236-y, doi:10.1038/s41423-019-0236-y. This article has 28 citations and is from a peer-reviewed journal.
10. (song2020e3ubiquitinligase pages 7-9): Xiaoqi Song, Shuo Liu, Wen-die Wang, Zhong-fei Ma, Xuetao Cao, and Minghong Jiang. E3 ubiquitin ligase rnf170 inhibits innate immune responses by targeting and degrading tlr3 in murine cells. Cellular & Molecular Immunology, 17:865-874, May 2020. URL: https://doi.org/10.1038/s41423-019-0236-y, doi:10.1038/s41423-019-0236-y. This article has 28 citations and is from a peer-reviewed journal.
11. (lari2016resolutionofproteotoxic pages 111-115): F Lari. Resolution of proteotoxic stress in the endoplasmic reticulum by ubiquitin ligase complexes. Unknown journal, 2016.
12. (veronese2024erlin12scaffoldsbridge pages 4-6): Matteo Veronese, Sebastian Kallabis, Alexander Tobias Kaczmarek, Anushka Das, Lennart Robers, Simon Schumacher, Alessia Lofrano, Susanne Brodesser, Stefan Müller, Kay Hofmann, Marcus Krüger, and Elena I Rugarli. Erlin1/2 scaffolds bridge tmub1 and rnf170 and restrict cholesterol esterification to regulate the secretory pathway. Life Science Alliance, 7:e202402620, May 2024. URL: https://doi.org/10.26508/lsa.202402620, doi:10.26508/lsa.202402620. This article has 8 citations and is from a peer-reviewed journal.
13. (veronese2024erlin12scaffoldsbridge pages 10-12): Matteo Veronese, Sebastian Kallabis, Alexander Tobias Kaczmarek, Anushka Das, Lennart Robers, Simon Schumacher, Alessia Lofrano, Susanne Brodesser, Stefan Müller, Kay Hofmann, Marcus Krüger, and Elena I Rugarli. Erlin1/2 scaffolds bridge tmub1 and rnf170 and restrict cholesterol esterification to regulate the secretory pathway. Life Science Alliance, 7:e202402620, May 2024. URL: https://doi.org/10.26508/lsa.202402620, doi:10.26508/lsa.202402620. This article has 8 citations and is from a peer-reviewed journal.
14. (wang2024hostfactorsmodulate pages 13-14): Jingjing Wang, Yirui Dong, Xuewei Zheng, Haodi Ma, Mengjiao Huang, Dongliao Fu, Jiangbo Liu, and Qinan Yin. Host factors modulate virus-induced ifn production via pattern recognition receptors. Journal of Inflammation Research, 17:3737-3752, Jun 2024. URL: https://doi.org/10.2147/jir.s455035, doi:10.2147/jir.s455035. This article has 6 citations and is from a peer-reviewed journal.
15. (li2024thernabindingproteins pages 12-13): Jianguo Li, Jingge Yu, Ao Shen, Suwen Lai, Zhiping Liu, and Tian-Sheng He. The rna-binding proteins regulate innate antiviral immune signaling by modulating pattern recognition receptors. Virology Journal, Sep 2024. URL: https://doi.org/10.1186/s12985-024-02503-x, doi:10.1186/s12985-024-02503-x. This article has 13 citations and is from a peer-reviewed journal.
16. (OpenTargets Search: hereditary spastic paraplegia,spastic paraplegia,sensory ataxia,spinocerebellar ataxia,neuroaxonal dystrophy-RNF170): Open Targets Query (hereditary spastic paraplegia,spastic paraplegia,sensory ataxia,spinocerebellar ataxia,neuroaxonal dystrophy-RNF170, 2 results). Buniello, A. et al. (2025). Open Targets Platform: facilitating therapeutic hypotheses building in drug discovery. Nucleic Acids Research.
17. (vondel2024overarchingpathomechanismsin pages 5-8): Liedewei Van de Vondel, Jonathan De Winter, Vincent Timmerman, and Jonathan Baets. Overarching pathomechanisms in inherited peripheral neuropathies, spastic paraplegias, and cerebellar ataxias. Mar 2024. URL: https://doi.org/10.1016/j.tins.2024.01.004, doi:10.1016/j.tins.2024.01.004. This article has 11 citations and is from a highest quality peer-reviewed journal.
18. (cogan2024biallelicvariantsin pages 1-6): Guillaume Cogan, Maha S. Zaki, Mahmoud Issa, Boris Keren, Marine Guillaud-Bataille, Florence Renaldo, Arnaud Isapof, Pauline Lallemant, Giovanni Stevanin, Lena Guillot-Noel, Thomas Courtin, Julien Buratti, Cécile Freihuber, Joseph G. Gleeson, Robyn Howarth, Alexandra Durr, Jean-Madeleine de Sainte Agathe, and Cyril Mignot. Biallelic variants in erlin1: a series of 13 individuals with spastic paraparesis. Human genetics, 143:1353-1362, Oct 2024. URL: https://doi.org/10.1007/s00439-024-02702-0, doi:10.1007/s00439-024-02702-0. This article has 3 citations and is from a peer-reviewed journal.
19. (cogan2024biallelicvariantsin pages 12-16): Guillaume Cogan, Maha S. Zaki, Mahmoud Issa, Boris Keren, Marine Guillaud-Bataille, Florence Renaldo, Arnaud Isapof, Pauline Lallemant, Giovanni Stevanin, Lena Guillot-Noel, Thomas Courtin, Julien Buratti, Cécile Freihuber, Joseph G. Gleeson, Robyn Howarth, Alexandra Durr, Jean-Madeleine de Sainte Agathe, and Cyril Mignot. Biallelic variants in erlin1: a series of 13 individuals with spastic paraparesis. Human genetics, 143:1353-1362, Oct 2024. URL: https://doi.org/10.1007/s00439-024-02702-0, doi:10.1007/s00439-024-02702-0. This article has 3 citations and is from a peer-reviewed journal.
20. (song2020e3ubiquitinligase pages 1-2): Xiaoqi Song, Shuo Liu, Wen-die Wang, Zhong-fei Ma, Xuetao Cao, and Minghong Jiang. E3 ubiquitin ligase rnf170 inhibits innate immune responses by targeting and degrading tlr3 in murine cells. Cellular & Molecular Immunology, 17:865-874, May 2020. URL: https://doi.org/10.1038/s41423-019-0236-y, doi:10.1038/s41423-019-0236-y. This article has 28 citations and is from a peer-reviewed journal.
21. (gehweiler2024rnf170anoveldisease pages 68-72): I Gehweiler. Rnf170-a novel disease gene causing hereditary spastic paraplegia. Unknown journal, 2024.
## Artifacts
- [Edison artifact artifact-00](RNF170-deep-research-falcon_artifacts/artifact-00.md)
## Citations
1. lari2016resolutionofproteotoxic pages 111-115
2. wagner2019biallelicvariantsin pages 6-7
3. vondel2024overarchingpathomechanismsin pages 5-8
4. cogan2024biallelicvariantsin pages 12-16
5. wang2024hostfactorsmodulate pages 13-14
6. li2024thernabindingproteins pages 12-13
7. cogan2024biallelicvariantsin pages 1-6
8. https://doi.org/10.1074/jbc.m111.251983
9. https://doi.org/10.1038/s41467-019-12620-9
10. https://doi.org/10.1038/s41423-019-0236-y
11. https://doi.org/10.26508/lsa.202402620
12. https://doi.org/10.1016/j.tins.2024.01.004
13. https://doi.org/10.1007/s00439-024-02702-0
14. https://doi.org/10.1074/jbc.m111.251983,
15. https://doi.org/10.1038/s41467-019-12620-9,
16. https://doi.org/10.26508/lsa.202402620,
17. https://doi.org/10.1038/s41423-019-0236-y,
18. https://doi.org/10.2147/jir.s455035,
19. https://doi.org/10.1186/s12985-024-02503-x,
20. https://doi.org/10.1016/j.tins.2024.01.004,
21. https://doi.org/10.1007/s00439-024-02702-0,