Deep Research
Bioreason Pro
(SPDL1-deep-research-bioreason-sft.md)
Bioreason Pro
(SPDL1-deep-research-bioreason-sft.md)Thinking Trace
The architecture begins with IPR028593 (Protein Spindly, chordates family) spanning residues 1–605, covering the full polypeptide and defining a chordate-specific kinetochore adaptor scaffold. Nested within this, residues 3–468 carry IPR051149 (Spindly/BICDR Dynein Motor Adapter family), a hallmark of coiled-coil adaptors that couple cytoplasmic dynein to diverse cargo. The placement of the dynein-adapter core inside the broader Spindly family envelope indicates a bipartite design: an N-terminal-to-mid-region motor-adapter module that binds dynein and its regulatory subunits, and a C-terminal region within the full-length family signature that engages kinetochore receptors. This arrangement causes selective recruitment of dynein to kinetochores and spindle poles, and it provides a platform for regulatory enzymes at the kinetochore.
From this domain logic, the molecular function centers on assembling and positioning a motor-enzymatic module. The dynein-adapter core enforces dynein targeting to microtubule minus ends and kinetochore receptors, while the scaffolded surfaces accommodate regulatory enzymes. This supports GO:0019899 enzyme binding, as the adaptor is poised to bind kinases and phosphatases that tune kinetochore-microtubule attachments and checkpoint signaling.
The same architecture drives specific biological processes. By docking dynein at kinetochores, the adaptor generates poleward forces that align chromosomes at the metaphase plate, matching GO:0007080 mitotic metaphase plate congression. Dynein recruitment also biases spindle geometry and microtubule flux, enabling GO:0000132 establishment of mitotic spindle orientation. The scaffold’s ability to capture outer-kinetochore receptors and concentrate regulatory enzymes directly implements GO:0034501 protein localization to kinetochore. Outside mitosis, dynein-based transport along microtubules contributes to directed cell movement; by positioning dynein at specific cortical or Golgi sites, the adaptor can influence cytoskeletal polarity and vesicle trafficking, aligning with GO:0016477 cell migration.
The cellular context follows from the same chain. The dynein-adapter module and kinetochore-binding surfaces place the protein at the GO:0000940 outer kinetochore during mitosis, where it forms the GO:0098806 Spindly-BICDR complex that couples dynein to kinetochore receptors. Minus-end-directed transport and motor loading explain accumulation at the GO:0000922 spindle pole. Between divisions and during interphase transport, a soluble pool resides in the GO:0005829 cytosol, while regulatory cycles and chromatin-proximal functions rationalize presence in the GO:0005634 nucleus.
Mechanistically, the adaptor binds outer-kinetochore receptors such as BUB1B/BUB3 and the NDC80 complex to anchor dynein and its cofactors, creating a dynein–dynactin–adaptor assembly that exerts poleward forces on attached kinetochores. Phosphorylation by PLK1 and checkpoint factors modulates its affinity and timing, while association with MKLN1 (BICDR) tunes dynein loading in interphase. Through these regulated interactions, it promotes bi-orientation, error correction, and timely anaphase onset. The same scaffold can engage additional kinetochore modules (SKA1/2/3, CENP-F) to stabilize end-on attachments and coordinate with APC/C activation via CDC20. In non-mitotic contexts, adaptor-mediated dynein targeting to Golgi membranes and cortical sites can drive vesicle trafficking and polarity, contributing to cell migration. The observed partners—BUB1 beta, CENP-F, SKA1/2/3, BIO1, PLK1, CDC20, NDC80, and even developmental regulators like HOX-D13—fit a model in which the adaptor integrates checkpoint, structural, and regulatory inputs to choreograph dynein-dependent transport and kinetochore mechanics.
## Functional Summary
A dynein-recruiting kinetochore adaptor that assembles a motor–regulatory module to capture and load dynein–dynactin at kinetochores and spindle poles. By anchoring to outer-kinetochore receptors and engaging checkpoint and structural factors, it generates poleward forces for chromosome alignment and proper spindle orientation, coordinates the timing of anaphase onset, and modulates kinetochore–microtubule attachments. In interphase, association with a dynein cofactor tunes dynein loading and extends its role to Golgi-directed transport and polarity programs that support cell migration. The protein cycles between cytosol and nucleus, concentrates at kinetochores during mitosis, and forms a dedicated adaptor complex with its dynein-binding partner.
## UniProt Summary
Kinetochore adapter protein which recruits the dynein motor complex to kinetochores. Required for chromosome alignment and spindle orientation during mitosis. May also regulate the onset of anaphase and kinetochore microtubule attachment. Association with MKLN1/BICDR regulates dynein loading by SPINDLY onto the Golgi apparatus.
## InterPro Domains
- IPR028593: Protein Spindly, chordates (family) [1-605]
- IPR051149: Spindly/BICDR Dynein Motor Adapter (family) [3-468]
## GO Term Predictions
### Molecular Function
### Biological Process
### Cellular Component
Falcon
(SPDL1-deep-research-falcon.md)
Falcon
(SPDL1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Human SPDL1 (Spindly; UniProt Q96EA4) Functional Annotation
0) Target verification (mandatory)
SPDL1 in this report refers specifically to human Spindly (hSpindly) encoded by SPDL1 (also referenced as CCDC99) with UniProt accession Q96EA4 and a length of ~605 aa. This matches the mitotic dynein–dynactin activating adaptor described in kinetochore/corona literature, including explicit UniProt mapping and domain schematics in a 2023 review. (gassmann2023dyneinatthe pages 5-6, gassmann2023dyneinatthe pages 4-5, d’amico2022conformationaltransitionsof pages 1-2, feng2024emergingroleand pages 1-2)
1) Key concepts, definitions, and current understanding
1.1 What SPDL1/Spindly is
Spindly (SPDL1) is a coiled-coil activating adaptor that links the microtubule minus-end motor cytoplasmic dynein-1 and its cofactor dynactin to the kinetochore fibrous corona during mitosis, enabling dynein recruitment/activation at kinetochores. (gassmann2023dyneinatthe pages 4-5, gassmann2023dyneinatthe pages 5-6)
A core organizing concept is that “activating adaptors” are not merely tethers: they stabilize the dynein–dynactin interaction and promote formation of a motile dynein–dynactin–adaptor complex (here, DDS = dynein–dynactin–Spindly). (gassmann2023dyneinatthe pages 4-5)
1.2 Kinetochore/corona and spindle assembly checkpoint (SAC) silencing
The fibrous corona is a dynamic outer kinetochore layer that concentrates microtubule-binding and checkpoint proteins early in mitosis. Spindly operates within the RZZ–Spindly pathway to recruit dynein–dynactin to this region and to promote dynein-dependent “stripping” (poleward transport/removal) of corona material, including checkpoint effectors, as kinetochores achieve productive microtubule attachments—an important mechanism contributing to SAC silencing and error avoidance. (gassmann2023dyneinatthe pages 4-5, gassmann2023dyneinatthe pages 3-4)
2) Molecular mechanism: domains, interactions, and regulation
2.1 Domain architecture and dynein/dynactin binding logic
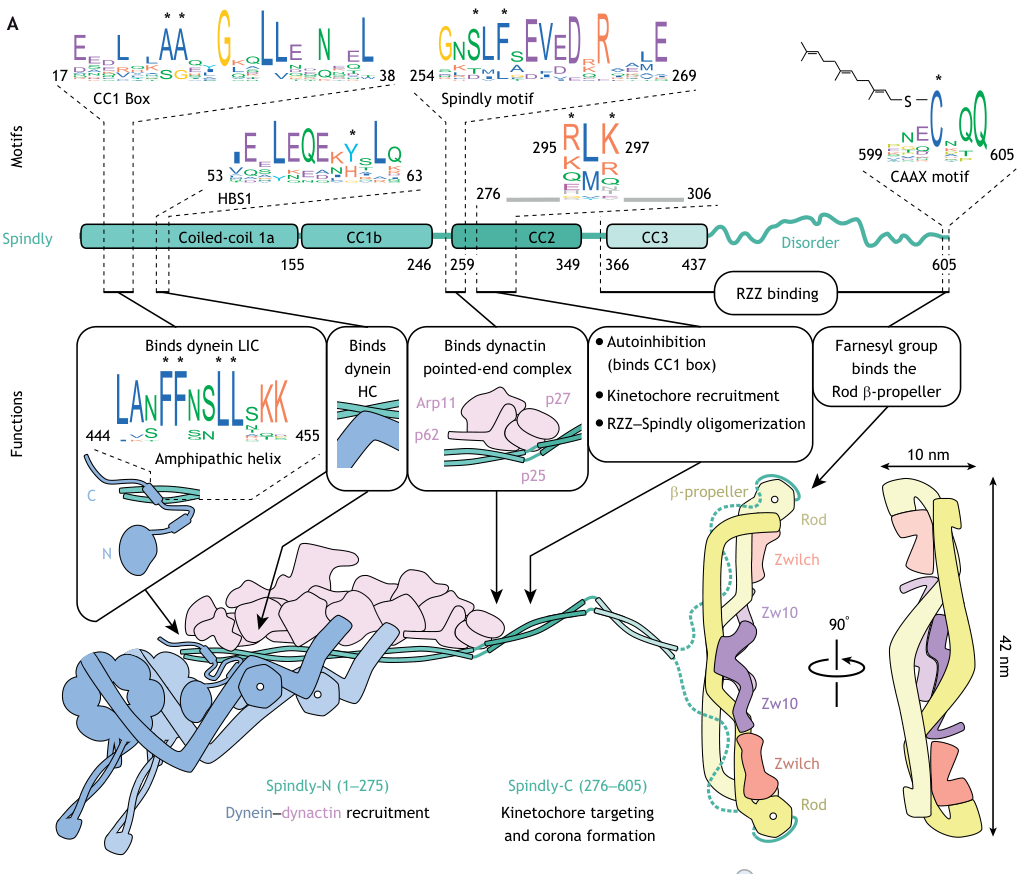
A domain schematic from a 2023 review summarizes Spindly’s key motifs and post-translational targeting signal, including:
- CC1 box: creates a binding pocket for a conserved amphipathic helix in dynein LIC (light intermediate chain).
- Spindly motif: contributes to interaction with the dynactin pointed-end.
- C-terminal CAAX motif enabling farnesylation.
- N-terminal functional partitioning (roughly 1–275 for dynein/dynactin recruitment; 276–605 for kinetochore targeting/corona formation). (gassmann2023dyneinatthe pages 4-5, gassmann2023dyneinatthe pages 5-6)
Image evidence: The Spindly domain architecture and the kinetochore recruitment model are shown schematically in the cited review figures. (gassmann2023dyneinatthe media 046141ea, gassmann2023dyneinatthe media fb7cb4cf)
2.2 Recruitment to kinetochores via RZZ and farnesylation
Spindly is recruited to kinetochores primarily through the RZZ complex (ROD–ZW10–ZWILCH). A key targeting mechanism is C-terminal farnesylation, with the Rod β-propeller acting as a farnesyl receptor in humans. (gassmann2023dyneinatthe pages 6-7, barbosa2020rzzspindlydyneinyougot pages 5-7)
2.3 Autoinhibition and kinetochore-dependent activation (structural/biochemical understanding)
A major mechanistic insight from structure–function work is that full-length Spindly is autoinhibited: it adopts a folded/closed conformation that occludes the CC1 box and Spindly motif, preventing productive binding to dynein–dynactin in solution. (d’amico2022conformationaltransitionsof pages 1-2, d’amico2022conformationaltransitionsof pages 11-13)
Importantly, RZZ binding alone is insufficient to fully “open” Spindly into a dynein–dynactin-binding competent state; the data support a multi-trigger activation model at kinetochores (RZZ plus at least one additional kinetochore cue/receptor). (d’amico2022conformationaltransitionsof pages 11-13, d’amico2022conformationaltransitionsof pages 13-14, gassmann2023dyneinatthe pages 6-7)
A 2023 synthesis describes a regulatory segment in CC2 (residues ~276–306) folding back onto the CC1 box to enforce autoinhibition, thereby preventing inappropriate dynein activation away from kinetochores and coupling activation to corona assembly. (gassmann2023dyneinatthe pages 6-7)
2.4 Kinase regulation and corona dynamics
Multiple mitotic kinases regulate the RZZ–Spindly–dynein module:
- MPS1 activity is required for robust RZZ–Spindly recruitment/corona expansion; MPS1-dependent ROD phosphorylation promotes expansion. (barbosa2020rzzspindlydyneinyougot pages 5-7, barbosa2022theroleof pages 17-18, gassmann2023dyneinatthe pages 7-8)
- PLK1/Polo phosphorylates Spindly to tune coupling/uncoupling between RZZ and dynein, helping prevent premature stabilization of erroneous attachments early in mitosis and allowing later transition to stable end-on attachments. (barbosa2020rzzspindlydyneinyougot pages 1-3, barbosa2020rzzspindlydyneinyougot pages 3-5, barbosa2020rzzspindlydyneinyougot pages 5-7)
3) Cellular functions (experimentally supported)
3.1 Spindle assembly checkpoint silencing via corona disassembly (updated model)
A key 2023 mechanistic advance is that kinetochore dynein’s checkpoint-silencing role is restricted primarily to disassembly of the fibrous corona, rather than wholesale removal of checkpoint proteins from the entire outer kinetochore. Spindly is central because it recruits dynein to the corona. (ide2023theroleof pages 1-2, ide2023theroleof pages 9-10)
Quantitative evidence from the 2023 study: a Spindly mutant (SpindlyΔCCS) that fails to recruit dynein but allows corona expansion yields strong mitotic arrest phenotypes (mitotic index 63.9% with wild-type Mad1). Disrupting corona-localized Mad1/Mad2 (Mad1-3EK) in the same SpindlyΔCCS background reduces the mitotic index (30.8%) and increases mitotic exit over 15 h (49% vs 8% for SpindlyΔCCS + WT Mad1), supporting that dynein recruited through Spindly is essential to remove a potent corona-derived inhibitory signal when few kinetochores remain unattached. (ide2023theroleof pages 9-10)
3.2 Promoting correct kinetochore–microtubule attachments and limiting segregation errors
A 2023 review describes that Spindly-dependent dynein stripping is triggered after end-coupled microtubule attachment, enabling removal of corona components (including Mad1–Mad2) and promoting kinetochore remodeling/compaction. Perturbations that prevent Spindly-dependent dynein recruitment/stripping can lead to merotelic attachments and chromosome mis-segregation. (gassmann2023dyneinatthe pages 4-5, gassmann2023dyneinatthe pages 3-4)
4) Subcellular localization
A 2024 SPDL1-focused review summarizes Spindly localization dynamics:
- Interphase: nuclear localization.
- Prometaphase: kinetochore localization.
- Pre-metaphase/metaphase: relocalization toward spindle poles.
- Loss after chromosome congression. (feng2024emergingroleand pages 1-2)
Consistent with the mechanistic literature, Spindly is specifically enriched at the kinetochore fibrous corona in early mitosis where it enables dynein–dynactin recruitment. (d’amico2022conformationaltransitionsof pages 1-2, ide2023theroleof pages 1-2)
5) Recent developments and latest research (prioritizing 2023–2024)
5.1 2023: dynein/Spindly in checkpoint silencing refined to corona disassembly
The 2023 Molecular Biology of the Cell study provides a refined checkpoint-silencing model emphasizing that dynein recruited via Spindly primarily removes corona-localized checkpoint effectors, effectively “priming” the checkpoint system for stable end-on attachment signals. (ide2023theroleof pages 1-2, ide2023theroleof pages 9-10)
5.2 2024: disease-focused synthesis of SPDL1 (review)
A 2024 Open Medicine review provides a consolidated picture of SPDL1/Spindly biology and disease relevance, summarizing SPDL1 structure (~605 aa; farnesylation; kinetochore-binding segment), core mitotic functions (SAC silencing, microtubule attachment), and diverse disease associations including pulmonary fibrosis and multiple cancers. (feng2024emergingroleand pages 1-2, feng2024emergingroleand pages 6-7)
5.3 2024: oncology example—ESCC functional targeting and prognostic modeling
A 2024 PLOS ONE study in esophageal squamous cell carcinoma (ESCC) reports SPDL1 is overexpressed in tumor tissue datasets (TCGA/GEO), correlates with survival through SPDL1-related lncRNA risk scores (P<0.05), and that SPDL1 inhibition in TE-1 cells suppresses proliferation, migration, and invasion (P<0.05), supporting biomarker/target hypotheses in a real tumor setting. (liu2024inhibitingtheexpression pages 1-2)
5.4 2024: large-scale human genetics (preprint) implicating SPDL1 in prostate cancer risk
A May 2024 medRxiv meta-analysis of rare coding germline variation in prostate cancer (37,184 cases; 331,329 male controls overall) reports rare non-synonymous variants in SPDL1 among genes associated with decreased prostate cancer risk (preprint; not peer-reviewed). (mitchell2024characterisingthecontribution pages 1-4)
6) Human disease associations and statistics (with emphasis on fibrosis genetics)
6.1 Idiopathic pulmonary fibrosis (IPF): replicated SPDL1 missense association
A 2021 Communications Biology sequencing study identifies a rare SPDL1 missense variant p.Arg20Gln (rs116483731) associated with IPF:
- Discovery: 752 sporadic IPF cases vs 119,055 controls; allele frequency 2.2% in cases vs 0.78% controls; OR 2.87 (95% CI 2.03–4.07), p=2.4×10−7.
- Replication: FinnGen 1028 cases vs 196,986 controls; case frequency 6.9% vs control 3.0%; OR 3.13 (95% CI 2.37–4.14), p=1.0×10−15.
- Combined evidence: p=2.2×10−20.
The authors highlight SPDL1/Spindly as a mitotic checkpoint signaling gene not previously tied to fibrosis, suggesting a new mechanistic axis for IPF biology and drug discovery. (dhindsa2021identificationofa pages 2-3, dhindsa2021identificationofa pages 1-2)
Clinical context reported in the same study: median diagnosis age 71 years, median survival 39.4 months, underscoring unmet need and the potential value of genetic stratification for etiology and therapy development. (dhindsa2021identificationofa pages 1-2)
6.2 Antagonistic pleiotropy (IPF risk vs cancer protection) and somatic chromosomal alterations (preprint)
A 2021 medRxiv analysis using FinnGen/UK Biobank data reports the same SPDL1 missense allele shows antagonistic pleiotropy: increased IPF risk but decreased cancer risk. In FinnGen, the SPDL1 missense allele is associated with IPF risk (HR 2.27, 95% CI 1.86–2.76; P=4.46×10−16) and reduced overall cancer risk (OR 0.81, 95% CI 0.77–0.85; P=2.05×10−15; longitudinal cancer HR 0.83). The allele is also associated with reduced mosaic chromosomal alterations (OR 0.71, 95% CI 0.63–0.79; P=5.36×10−9), supporting a hypothesis that reduced accumulation of chromosomal alterations may protect against cancer while predisposing to fibrosis/senescence pathways (preprint). (koskela2021geneticvariantin pages 7-10)
7) Current applications and real-world implementations
-
Clinical genetics / risk stratification in IPF: The replicated association of SPDL1 p.Arg20Gln with IPF (OR ~2.9–3.1; strong replication; large cohorts) supports its use as a genetic risk marker in research and potentially in clinical genetic panels for pulmonary fibrosis risk assessment/stratification, alongside known loci such as MUC5B and telomere-related genes (implementation depends on local practice and validation studies). (dhindsa2021identificationofa pages 2-3, dhindsa2021identificationofa pages 1-2)
-
Therapeutic discovery for IPF: By implicating a mitotic checkpoint/kinetochore–dynein pathway in IPF—distinct from telomere-length mechanisms—SPDL1 provides a mechanistically distinct hypothesis space for target discovery, though direct SPDL1-targeted therapies are not established. (dhindsa2021identificationofa pages 1-2, dhindsa2021identificationofa pages 3-5)
-
Oncology biomarker/target exploration: A 2024 ESCC study supports SPDL1 as a prognostic/biology-linked marker and suggests suppressing SPDL1 can inhibit malignant cell phenotypes in vitro, motivating exploratory targeting strategies in specific tumor contexts; however, prognostic directionality appears cancer-type dependent per review synthesis. (liu2024inhibitingtheexpression pages 1-2, feng2024emergingroleand pages 6-7)
8) Expert synthesis and interpretation (authoritative perspectives)
-
A 2023 Journal of Cell Science review frames SPDL1/Spindly as a kinetochore-localized activating adaptor that couples dynein recruitment to motor activation, and emphasizes that dynein/Spindly-dependent corona stripping is integrated with establishment of correct end-on attachments and checkpoint silencing; several mechanistic parameters (stoichiometry of DDS; triggers of Spindly activation; full corona architecture) remain open. (gassmann2023dyneinatthe pages 4-5, gassmann2023dyneinatthe pages 3-4, gassmann2023dyneinatthe pages 6-7)
-
A 2024 SPDL1-focused review synthesizes broader disease connections, emphasizing mitotic checkpoint silencing, microtubule attachment and potential roles in migration/drug response as recurring biological themes; it highlights heterogeneity and contradictions in cancer associations as an area requiring more rigorous clinical validation. (feng2024emergingroleand pages 1-2, feng2024emergingroleand pages 6-7)
9) Summary tables
9.1 Core functional annotation (mechanism-level)
| Aspect | Current best-supported annotation for human SPDL1/Spindly | Key sources |
|---|---|---|
| Molecular role | Kinetochore-localized dynein–dynactin activating adaptor; assembles DDS at fibrous corona | (gassmann2023dyneinatthe pages 4-5) |
| Recruitment platform | RZZ complex (ROD/ZW10/ZWILCH); oligomerization contributes to corona organization | (gassmann2023dyneinatthe pages 6-7, gassmann2023dyneinatthe pages 7-8) |
| Targeting PTM | C-terminal farnesylation (CAAX); Rod β-propeller is farnesyl receptor | (gassmann2023dyneinatthe pages 6-7, barbosa2020rzzspindlydyneinyougot pages 5-7) |
| Dynein binding | CC1 box → dynein LIC amphipathic helix; additional motifs engage dynein/dynactin | (gassmann2023dyneinatthe pages 4-5, barbosa2020rzzspindlydyneinyougot pages 3-5) |
| Dynactin binding | Spindly motif engages dynactin pointed-end | (gassmann2023dyneinatthe pages 4-5, barbosa2020rzzspindlydyneinyougot pages 3-5) |
| Regulation | Autoinhibition (closed conformation occludes CC1 box/Spindly motif); activated at kinetochores | (d’amico2022conformationaltransitionsof pages 1-2, d’amico2022conformationaltransitionsof pages 11-13) |
| Checkpoint role | Dynein recruited by Spindly disassembles corona and removes corona-localized checkpoint effectors | (ide2023theroleof pages 1-2, ide2023theroleof pages 9-10) |
9.2 Selected recent/impactful studies with quantitative data
| Year | Study | Topic | Key quantitative findings | URL |
|---|---|---|---|---|
| 2023 | Ide et al., Mol Biol Cell | Dynein/Spindly in SAC silencing via corona disassembly | SpindlyΔCCS + WT Mad1 mitotic index 63.9% vs SpindlyΔCCS + Mad1-3EK 30.8%; mitotic exit over 15h 8% vs 49% | https://doi.org/10.1091/mbc.e23-04-0130 |
| 2024 | Feng et al., Open Medicine (review) | SPDL1 in health/disease synthesis | Reports 605-aa Q96EA4 protein; summarizes mitotic roles and disease links | https://doi.org/10.1515/med-2024-0922 |
| 2024 | Liu et al., PLOS ONE | ESCC biomarker/functional study | SPDL1 inhibition suppresses TE-1 proliferation/migration/invasion (P<0.05); risk models correlate with OS (P<0.05) | https://doi.org/10.1371/journal.pone.0302312 |
| 2024 | Mitchell et al., medRxiv (preprint) | Prostate cancer rare variant meta-analysis | Cohort sizes 37,184 cases; 331,329 controls; SPDL1 rare variants associated with decreased risk (no OR in excerpt) | https://doi.org/10.1101/2024.05.10.24307164 |
| 2021 | Dhindsa et al., Commun Biol | IPF genetics | p.Arg20Gln (rs116483731) OR 2.87 (p=2.4×10−7) discovery; OR 3.13 (p=1.0×10−15) replication; combined p=2.2×10−20 | https://doi.org/10.1038/s42003-021-01910-y |
| 2021 | Koskela et al., medRxiv (preprint) | IPF–cancer antagonistic pleiotropy | IPF HR 2.27; cancer OR 0.81; mosaic chromosomal alterations OR 0.71 | https://doi.org/10.1101/2021.05.07.21255988 |
10) Notes on evidence limitations
- Some 2024 oncology/genetics results are preprints (medRxiv) and should be interpreted as provisional until peer reviewed. (mitchell2024characterisingthecontribution pages 1-4, koskela2021geneticvariantin pages 7-10)
- Cancer associations for SPDL1 appear context-dependent (tumor type–specific directionality), per 2024 synthesis. (feng2024emergingroleand pages 6-7)
Key cited visuals
- Spindly domain architecture and kinetochore recruitment pathway cartoons (used as visual evidence in this report). (gassmann2023dyneinatthe media 046141ea, gassmann2023dyneinatthe media fb7cb4cf)
References
-
(gassmann2023dyneinatthe pages 5-6): Reto Gassmann. Dynein at the kinetochore. Journal of cell science, Mar 2023. URL: https://doi.org/10.1242/jcs.220269, doi:10.1242/jcs.220269. This article has 24 citations and is from a domain leading peer-reviewed journal.
-
(gassmann2023dyneinatthe pages 4-5): Reto Gassmann. Dynein at the kinetochore. Journal of cell science, Mar 2023. URL: https://doi.org/10.1242/jcs.220269, doi:10.1242/jcs.220269. This article has 24 citations and is from a domain leading peer-reviewed journal.
-
(d’amico2022conformationaltransitionsof pages 1-2): Ennio A. d’Amico, Misbha Ud Din Ahmad, Verena Cmentowski, Mathias Girbig, Franziska Müller, Sabine Wohlgemuth, Andreas Brockmeyer, Stefano Maffini, Petra Janning, Ingrid R. Vetter, Andrew P. Carter, Anastassis Perrakis, and Andrea Musacchio. Conformational transitions of the spindly adaptor underlie its interaction with dynein and dynactin. The Journal of Cell Biology, Sep 2022. URL: https://doi.org/10.1083/jcb.202206131, doi:10.1083/jcb.202206131. This article has 34 citations.
-
(feng2024emergingroleand pages 1-2): Yuejiao Feng, Donghao Tang, and Jie Wang. Emerging role and function of spdl1 in human health and diseases. Open Medicine, Jan 2024. URL: https://doi.org/10.1515/med-2024-0922, doi:10.1515/med-2024-0922. This article has 4 citations and is from a peer-reviewed journal.
-
(gassmann2023dyneinatthe pages 3-4): Reto Gassmann. Dynein at the kinetochore. Journal of cell science, Mar 2023. URL: https://doi.org/10.1242/jcs.220269, doi:10.1242/jcs.220269. This article has 24 citations and is from a domain leading peer-reviewed journal.
-
(gassmann2023dyneinatthe media 046141ea): Reto Gassmann. Dynein at the kinetochore. Journal of cell science, Mar 2023. URL: https://doi.org/10.1242/jcs.220269, doi:10.1242/jcs.220269. This article has 24 citations and is from a domain leading peer-reviewed journal.
-
(gassmann2023dyneinatthe media fb7cb4cf): Reto Gassmann. Dynein at the kinetochore. Journal of cell science, Mar 2023. URL: https://doi.org/10.1242/jcs.220269, doi:10.1242/jcs.220269. This article has 24 citations and is from a domain leading peer-reviewed journal.
-
(gassmann2023dyneinatthe pages 6-7): Reto Gassmann. Dynein at the kinetochore. Journal of cell science, Mar 2023. URL: https://doi.org/10.1242/jcs.220269, doi:10.1242/jcs.220269. This article has 24 citations and is from a domain leading peer-reviewed journal.
-
(barbosa2020rzzspindlydyneinyougot pages 5-7): João Barbosa, Carlos Conde, and Claudio Sunkel. Rzz-spindly-dynein: you got to keep ‘em separated. Cell Cycle, 19:1716-1726, Jun 2020. URL: https://doi.org/10.1080/15384101.2020.1780382, doi:10.1080/15384101.2020.1780382. This article has 10 citations and is from a peer-reviewed journal.
-
(d’amico2022conformationaltransitionsof pages 11-13): Ennio A. d’Amico, Misbha Ud Din Ahmad, Verena Cmentowski, Mathias Girbig, Franziska Müller, Sabine Wohlgemuth, Andreas Brockmeyer, Stefano Maffini, Petra Janning, Ingrid R. Vetter, Andrew P. Carter, Anastassis Perrakis, and Andrea Musacchio. Conformational transitions of the spindly adaptor underlie its interaction with dynein and dynactin. The Journal of Cell Biology, Sep 2022. URL: https://doi.org/10.1083/jcb.202206131, doi:10.1083/jcb.202206131. This article has 34 citations.
-
(d’amico2022conformationaltransitionsof pages 13-14): Ennio A. d’Amico, Misbha Ud Din Ahmad, Verena Cmentowski, Mathias Girbig, Franziska Müller, Sabine Wohlgemuth, Andreas Brockmeyer, Stefano Maffini, Petra Janning, Ingrid R. Vetter, Andrew P. Carter, Anastassis Perrakis, and Andrea Musacchio. Conformational transitions of the spindly adaptor underlie its interaction with dynein and dynactin. The Journal of Cell Biology, Sep 2022. URL: https://doi.org/10.1083/jcb.202206131, doi:10.1083/jcb.202206131. This article has 34 citations.
-
(barbosa2022theroleof pages 17-18): João Barbosa, Claudio E. Sunkel, and Carlos Conde. The role of mitotic kinases and the rzz complex in kinetochore-microtubule attachments: doing the right link. Frontiers in Cell and Developmental Biology, Jan 2022. URL: https://doi.org/10.3389/fcell.2022.787294, doi:10.3389/fcell.2022.787294. This article has 18 citations.
-
(gassmann2023dyneinatthe pages 7-8): Reto Gassmann. Dynein at the kinetochore. Journal of cell science, Mar 2023. URL: https://doi.org/10.1242/jcs.220269, doi:10.1242/jcs.220269. This article has 24 citations and is from a domain leading peer-reviewed journal.
-
(barbosa2020rzzspindlydyneinyougot pages 1-3): João Barbosa, Carlos Conde, and Claudio Sunkel. Rzz-spindly-dynein: you got to keep ‘em separated. Cell Cycle, 19:1716-1726, Jun 2020. URL: https://doi.org/10.1080/15384101.2020.1780382, doi:10.1080/15384101.2020.1780382. This article has 10 citations and is from a peer-reviewed journal.
-
(barbosa2020rzzspindlydyneinyougot pages 3-5): João Barbosa, Carlos Conde, and Claudio Sunkel. Rzz-spindly-dynein: you got to keep ‘em separated. Cell Cycle, 19:1716-1726, Jun 2020. URL: https://doi.org/10.1080/15384101.2020.1780382, doi:10.1080/15384101.2020.1780382. This article has 10 citations and is from a peer-reviewed journal.
-
(ide2023theroleof pages 1-2): Amy H. Ide, Keith F. DeLuca, O’Neil Wiggan, Steven M. Markus, and Jennifer G. DeLuca. The role of kinetochore dynein in checkpoint silencing is restricted to disassembly of the corona. Jun 2023. URL: https://doi.org/10.1091/mbc.e23-04-0130, doi:10.1091/mbc.e23-04-0130. This article has 10 citations and is from a domain leading peer-reviewed journal.
-
(ide2023theroleof pages 9-10): Amy H. Ide, Keith F. DeLuca, O’Neil Wiggan, Steven M. Markus, and Jennifer G. DeLuca. The role of kinetochore dynein in checkpoint silencing is restricted to disassembly of the corona. Jun 2023. URL: https://doi.org/10.1091/mbc.e23-04-0130, doi:10.1091/mbc.e23-04-0130. This article has 10 citations and is from a domain leading peer-reviewed journal.
-
(feng2024emergingroleand pages 6-7): Yuejiao Feng, Donghao Tang, and Jie Wang. Emerging role and function of spdl1 in human health and diseases. Open Medicine, Jan 2024. URL: https://doi.org/10.1515/med-2024-0922, doi:10.1515/med-2024-0922. This article has 4 citations and is from a peer-reviewed journal.
-
(liu2024inhibitingtheexpression pages 1-2): Tao Liu, Juan Xu, Qun-xian Zhang, Yan-Jiao Huang, Wei Wang, and Zhu Fu. Inhibiting the expression of spindle appendix cooled coil protein 1 can suppress tumor cell growth and metastasis and is associated with cancer immune cells in esophageal squamous cell carcinoma. PLOS ONE, 19(8):e0302312, Aug 2024. URL: https://doi.org/10.1371/journal.pone.0302312, doi:10.1371/journal.pone.0302312. This article has 1 citations and is from a peer-reviewed journal.
-
(mitchell2024characterisingthecontribution pages 1-4): Jonathan Mitchell, Niedzica Camacho, Patrick Shea, Konrad H. Stopsack, Vijai Joseph, Oliver Burren, Ryan Dhindsa, Abhishek Nag, Jacob E. Berchuck, Amanda O’Neill, Ali Abbasi, Anthony W. Zoghbi, Jesus Alegre-Díaz, Pablo Kuri-Morales, Jaime Berumen, Roberto Tapia-Conyer, Jonathan Emberson, Jason M. Torres, Rory Collins, Quanli Wang, David Goldstein, Athena Matakidou, Carolina Haefliger, Lauren Anderson-Dring, Ruth March, Vaidehi Jobanputra, Brian Dougherty, Keren Carss, Slavé Petrovski, Philip W. Kantoff, Kenneth Offit, Lorelei A. Mucci, Mark Pomerantz, and Margarete A. Fabre. Characterising the contribution of rare protein-coding germline variants to prostate cancer risk and severity in 37,184 cases. medRxiv, May 2024. URL: https://doi.org/10.1101/2024.05.10.24307164, doi:10.1101/2024.05.10.24307164. This article has 4 citations.
-
(dhindsa2021identificationofa pages 2-3): Ryan S. Dhindsa, Johan Mattsson, Abhishek Nag, Quanli Wang, Louise V. Wain, Richard Allen, Eleanor M. Wigmore, Kristina Ibanez, Dimitrios Vitsios, Sri V. V. Deevi, Sebastian Wasilewski, Maria Karlsson, Glenda Lassi, Henric Olsson, Daniel Muthas, Susan Monkley, Alex Mackay, Lynne Murray, Simon Young, Carolina Haefliger, Toby M. Maher, Maria G. Belvisi, Gisli Jenkins, Philip L. Molyneaux, Adam Platt, and Slavé Petrovski. Identification of a missense variant in spdl1 associated with idiopathic pulmonary fibrosis. Communications Biology, Mar 2021. URL: https://doi.org/10.1038/s42003-021-01910-y, doi:10.1038/s42003-021-01910-y. This article has 69 citations and is from a peer-reviewed journal.
-
(dhindsa2021identificationofa pages 1-2): Ryan S. Dhindsa, Johan Mattsson, Abhishek Nag, Quanli Wang, Louise V. Wain, Richard Allen, Eleanor M. Wigmore, Kristina Ibanez, Dimitrios Vitsios, Sri V. V. Deevi, Sebastian Wasilewski, Maria Karlsson, Glenda Lassi, Henric Olsson, Daniel Muthas, Susan Monkley, Alex Mackay, Lynne Murray, Simon Young, Carolina Haefliger, Toby M. Maher, Maria G. Belvisi, Gisli Jenkins, Philip L. Molyneaux, Adam Platt, and Slavé Petrovski. Identification of a missense variant in spdl1 associated with idiopathic pulmonary fibrosis. Communications Biology, Mar 2021. URL: https://doi.org/10.1038/s42003-021-01910-y, doi:10.1038/s42003-021-01910-y. This article has 69 citations and is from a peer-reviewed journal.
-
(koskela2021geneticvariantin pages 7-10): Jukka T. Koskela, Paavo Häppölä, Aoxing Liu, Juulia Partanen, Giulio Genovese, Mykyta Artomov, Mikko N.M. Myllymäki, Masahiro Kanai, Wei Zhou, Juha M. Karjalainen, Teemu Palviainen, Justiina Ronkainen, Sylvain Sebert, Taru Tukiainen, Priit Palta, Jaakko Kaprio, Mitja Kurki, Andrea Ganna, Aarno Palotie, Tarja Laitinen, Marjukka Myllärniemi, and Mark J. Daly. Genetic variant in spdl1 reveals novel mechanism linking pulmonary fibrosis risk and cancer protection. MedRxiv, May 2021. URL: https://doi.org/10.1101/2021.05.07.21255988, doi:10.1101/2021.05.07.21255988. This article has 11 citations.
-
(dhindsa2021identificationofa pages 3-5): Ryan S. Dhindsa, Johan Mattsson, Abhishek Nag, Quanli Wang, Louise V. Wain, Richard Allen, Eleanor M. Wigmore, Kristina Ibanez, Dimitrios Vitsios, Sri V. V. Deevi, Sebastian Wasilewski, Maria Karlsson, Glenda Lassi, Henric Olsson, Daniel Muthas, Susan Monkley, Alex Mackay, Lynne Murray, Simon Young, Carolina Haefliger, Toby M. Maher, Maria G. Belvisi, Gisli Jenkins, Philip L. Molyneaux, Adam Platt, and Slavé Petrovski. Identification of a missense variant in spdl1 associated with idiopathic pulmonary fibrosis. Communications Biology, Mar 2021. URL: https://doi.org/10.1038/s42003-021-01910-y, doi:10.1038/s42003-021-01910-y. This article has 69 citations and is from a peer-reviewed journal.
Artifacts
Citations
- gassmann2023dyneinatthe pages 4-5
- gassmann2023dyneinatthe pages 6-7
- ide2023theroleof pages 9-10
- feng2024emergingroleand pages 1-2
- liu2024inhibitingtheexpression pages 1-2
- mitchell2024characterisingthecontribution pages 1-4
- dhindsa2021identificationofa pages 1-2
- koskela2021geneticvariantin pages 7-10
- feng2024emergingroleand pages 6-7
- gassmann2023dyneinatthe pages 5-6
- gassmann2023dyneinatthe pages 3-4
- barbosa2020rzzspindlydyneinyougot pages 5-7
- barbosa2022theroleof pages 17-18
- gassmann2023dyneinatthe pages 7-8
- barbosa2020rzzspindlydyneinyougot pages 1-3
- barbosa2020rzzspindlydyneinyougot pages 3-5
- ide2023theroleof pages 1-2
- dhindsa2021identificationofa pages 2-3
- dhindsa2021identificationofa pages 3-5
- https://doi.org/10.1091/mbc.e23-04-0130
- https://doi.org/10.1515/med-2024-0922
- https://doi.org/10.1371/journal.pone.0302312
- https://doi.org/10.1101/2024.05.10.24307164
- https://doi.org/10.1038/s42003-021-01910-y
- https://doi.org/10.1101/2021.05.07.21255988
- https://doi.org/10.1242/jcs.220269,
- https://doi.org/10.1083/jcb.202206131,
- https://doi.org/10.1515/med-2024-0922,
- https://doi.org/10.1080/15384101.2020.1780382,
- https://doi.org/10.3389/fcell.2022.787294,
- https://doi.org/10.1091/mbc.e23-04-0130,
- https://doi.org/10.1371/journal.pone.0302312,
- https://doi.org/10.1101/2024.05.10.24307164,
- https://doi.org/10.1038/s42003-021-01910-y,
- https://doi.org/10.1101/2021.05.07.21255988,