Deep Research
Falcon
(USO1-deep-research-falcon.md)
Falcon
(USO1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Comprehensive Research Report: Human USO1 (p115) — Functional Annotation
Scope, identity verification, and nomenclature
The target is USO1 in Homo sapiens, encoding general vesicular transport factor p115 (also called p115/USO1; sometimes referenced as vesicle-docking protein), corresponding to UniProt O60763. Across retrieved sources, “USO1” consistently refers to the p115-family vesicular trafficking factor involved in early secretory pathway organization and Golgi/ERGIC tethering, with no conflicting alternative gene identity detected in this evidence set. (yakunin2025thegolgivesicle pages 1-5, yakunin2023characterisingthefactors pages 36-39)
1) Key concepts and definitions (current understanding)
1.1 Vesicle tethering vs fusion (where USO1/p115 acts)
In the secretory pathway, COPII vesicles bud from ER exit sites and must be captured and fused with early Golgi/ERGIC membranes. “Tethering” refers to the initial physical capture/bridging that promotes productive docking and sets the stage for membrane fusion. p115/USO1 is described as a Golgi vesicle tethering protein that promotes vesicle docking and fusion, in part through binding to SNARE machinery and Rab GTPase–dependent recruitment. (yakunin2025thegolgivesicle pages 1-5, yakunin2025thegolgivesicle pages 11-14)
1.2 Rab GTPase recruitment and SNARE assembly
A widely used framework is that Rab1 (RAB1A/B) on COPII vesicles recruits p115/USO1 at the ER–Golgi interface, and p115 then helps organize or stabilize cis-Golgi SNARE complex assembly, promoting membrane fusion. A 2023 thesis synthesis explicitly states that p115 has been shown to interact with Rab1A via its N-terminus and that p115 interacts with cis-Golgi SNAREs (e.g., Sec22b, Bet1, GOSR2, syntaxin-5) in proposed fusion models. (yakunin2023characterisingthefactors pages 36-39)
1.3 Domain architecture (annotation-level, supported mainly by homolog structural work)
Although the UniProt record specifies armadillo/ARM-like repeats for human USO1/p115, the strongest domain-resolved structural evidence in the retrieved set is from homologs: yeast/fungal Uso1/p115 head domains adopt an armadillo-repeat/α-solenoid organization, and the protein includes long coiled-coils implicated in tethering-distance/geometry. Human-centric sources in this set largely describe p115 as a dimer with an N-terminal head and coiled-coil tail. (heo2020crystalstructuresof pages 1-2, yakunin2023characterisingthefactors pages 36-39)
2) Molecular function and pathways (evidence-led functional annotation)
2.1 Primary function: early secretory pathway tether/fusion factor
Human p115/USO1 is positioned at the ER–Golgi interface (ERGIC/cis-Golgi) as a vesicle tether/fusion factor, linking COPII vesicle delivery with productive fusion. In a human-cell experimental study (preprint), p115/USO1 is described as a key player in delivery of COPII vesicles to early Golgi compartments and as binding SNARE proteins to promote fusion; this study adds that p115 can bind an ER exit-site organizer (Sec16A), implying a role in large-scale spatial organization of ER exit sites relative to early Golgi. (yakunin2025thegolgivesicle pages 1-5, yakunin2025thegolgivesicle pages 11-14)
2.2 Interaction network (validated and proposed)
Rab1/RAB1A: p115/USO1 is described as associating with Golgi membranes in a Rab1-dependent manner and is frequently placed downstream of Rab1 activation on COPII vesicles in mechanistic models. (yakunin2025thegolgivesicle pages 1-5, yakunin2023characterisingthefactors pages 36-39)
GM130/GRASP65 axis: A 2023 thesis synthesis states that GM130 binds p115 at its N-terminus and that this interaction can be blocked by Cdk1 phosphorylation of GM130 S37 during mitosis, consistent with mitotic reorganization of the Golgi. (yakunin2023characterisingthefactors pages 36-39)
SNAREs: The same synthesis describes p115 interactions with cis-Golgi SNAREs (Sec22b, Bet1, GOSR2, syntaxin-5) and advances a model where p115 mediates SNARE complex assembly and vesicle fusion. (yakunin2023characterisingthefactors pages 36-39)
Sec16A (ER exit-site organizer): A human-cell study reports that the p115 head domain can bind directly to Sec16A, with mutations that disrupt motif binding reducing secretion efficiency, providing a mechanistic link between a Golgi tether and ER exit-site scaffolding. (yakunin2025thegolgivesicle pages 1-5)
3) Subcellular localization (where the protein carries out its role)
Human p115/USO1 localizes to the ERGIC and cis-Golgi, cycling on/off membranes and acting at early secretory trafficking steps. A human-cell study describes p115 as anchored to Golgi membranes, acting at the ER–Golgi interface, and adds functional coupling to ER exit sites via binding to Sec16A. (yakunin2025thegolgivesicle pages 1-5, yakunin2025thegolgivesicle pages 11-14)
Homolog systems reinforce this early-Golgi localization: fungal Uso1 resides on puncta overlapping early Golgi/ER–Golgi markers with dynamic residence times. (bravoplaza2023theuso1globular pages 1-2)
4) Recent developments and latest research (prioritizing 2023–2024)
4.1 2023: mechanistic shift toward “SNARE-regulatory essentiality” (homolog evidence)
A 2023 eLife study (fungal Uso1; homolog of human p115) reports that the globular head domain directly binds SNAREs (including Bet1 and Bos1/membrin family members) and that a monomeric head domain can complement loss of full-length Uso1 in vivo when overexpressed—arguing that long-range coiled-coil tethering can be dispensable and that the essential role can involve direct regulation of SNARE machinery. Quantitatively, Uso1 puncta had a reported half-life of ~60 s, and SNARE binding was modulated by specific point mutations. While not a human experiment, this work is influential because the machinery and architecture are conserved across eukaryotes and the study explicitly frames Uso1/p115 as a Rab1-recruited ER–Golgi tether with direct SNARE contacts. (bravoplaza2023theuso1globular pages 1-2, bravoplaza2023theuso1globular pages 27-30)
4.2 2024: phosphorylation/omics evidence in human disease tissue
A 2024 phosphoproteomics/proteomics study of treatment-naïve RA synovial biopsies (n=8) reports that USO1 Ser48 was the phosphosite with the highest inverse correlation with ESR in their analysis, illustrating that USO1 is phosphorylated in human inflammatory tissue and that specific phosphosites can correlate with clinical measures. This is associative and does not establish a trafficking mechanism for Ser48, but it provides human, recent evidence of regulated USO1 phosphorylation in vivo in disease. (cubuk2024phosphoproteomicprofilingof pages 1-2)
4.3 2024: systems-level contextualization in innate immunity
A 2024 review highlights the Golgi’s role in innate immune signaling and emphasizes that Golgi trafficking machinery (including tethers and SNARE-related processes) supports cytokine maturation and secretion pathways that pass through ER→ERGIC→Golgi→TGN routes. This supports the plausibility that perturbations in Golgi tethering proteins can influence immune signaling via secretory logistics, though it is not USO1-specific mechanistic proof. (maruntelu2024thegolgiapparatus pages 1-2)
Evidence gap note (important): within the retrieved corpus, there were relatively few 2023–2024 primary studies directly dissecting human USO1/p115 molecular mechanism (Rab1/GM130/SNARE binding biochemistry/structures) beyond the omics (RA) and broader Golgi/immune review context. Therefore, conserved homolog mechanistic evidence (2023 eLife) is explicitly labeled as such and used cautiously for functional inference. (bravoplaza2023theuso1globular pages 1-2, yakunin2023characterisingthefactors pages 36-39)
5) Current applications and real-world implementations
5.1 Host–pathogen biology: Legionella hijacking of p115
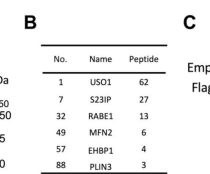
A major real-world implementation of USO1/p115 biology is in infection cell biology: a 2022 Science Advances study shows that the Legionella effector LegA15 hijacks host p115, inducing host Golgi fragmentation and altering lipid-droplet homeostasis. Proteomics identified p115/USO1 as a top putative interactor with high peptide support (see quantitative section and figure-based evidence below). This provides a concrete example where p115 is a host dependency exploited by pathogens. (chen2022atypicallegionellagtpase pages 1-2, chen2022atypicallegionellagtpase media 41416518, chen2022atypicallegionellagtpase media b798ae8b)
5.2 Cancer cell biology and biomarker/isoform studies
In hepatocellular carcinoma, an isoform-level USO1 study reports tumor-associated isoforms with distinct functional consequences on Golgi structure and ER→Golgi→plasma membrane trafficking, linking ER–Golgi network dysregulation to cancer aggressiveness and prognosis association. (yoon2021uso1isoformsdifferentially pages 1-2, yoon2021uso1isoformsdifferentially pages 7-8)
5.3 Clinical proteomics/phosphoproteomics for stratification
The 2024 RA synovial phosphoproteomics study illustrates a practical pipeline for label-free MS-based phosphoproteomics in patient tissue and supports the concept that pathway activity/phosphosite signatures can stratify disease pathotypes; USO1 Ser48 emerges as one of the phosphosites associated with a clinical metric (ESR). (cubuk2024phosphoproteomicprofilingof pages 1-2)
6) Expert opinions and analysis (authoritative synthesis)
A 2023 thesis synthesis (University of Cambridge repository) summarizes multiple models for p115/USO1 action, including: (i) tethering via C-terminal binding to golgins such as GM130/giantin, (ii) Rab1A binding via the N-terminus, and (iii) promotion of SNARE assembly/fusion. Importantly, it emphasizes that the field has debated whether p115’s golgin-binding is essential (conflicting results between models), consistent with more recent homolog mechanistic work suggesting essentiality may lie in SNARE regulation rather than long-distance tethering alone. (yakunin2023characterisingthefactors pages 36-39, bravoplaza2023theuso1globular pages 1-2)
7) Relevant statistics and recent data (quantitative highlights)
7.1 Proteomics: p115/USO1 peptide support in host–pathogen interaction mapping
In the LegA15 immunoprecipitation/MS analysis, p115/USO1 was identified with 62 unique peptides among top candidate binding proteins (figure-based evidence). (chen2022atypicallegionellagtpase media 41416518)
7.2 Live-cell dynamics and colocalization (homolog system)
In fungal Uso1 (homolog), Uso1 puncta recurrence had an estimated half-life of ~60 seconds, and quantitative colocalization with early Golgi markers was reported (e.g., Pearson correlations with early Golgi factors and Rab1). While not human, these data quantify early Golgi association dynamics for a conserved tether/fusion factor. (bravoplaza2023theuso1globular pages 1-2, bravoplaza2022tetheringbyuso1 pages 11-13)
7.3 Human phosphoproteomics cohort size and USO1 phosphosite association
In the 2024 RA synovium study, 8 pre-treatment synovial biopsies were analyzed by label-free MS; within correlated phosphosite analyses, USO1 Ser48 was highlighted as having the strongest inverse correlation with ESR (as reported in the study text describing Fig. 4C). (cubuk2024phosphoproteomicprofilingof pages 1-2)
7.4 Human trafficking assay quantitation framework (HCC context)
In the HCC isoform study, ER→Golgi trafficking was quantified using a VSVG-ts045 trafficking assay with readouts reported as mean % of cells ± SEM and sampling of n=5–8 fields with 10–50 cells per field (exact percentages are not included in the available excerpt). (yoon2021uso1isoformsdifferentially pages 9-10)
Summary table of supported annotation
The following table consolidates function, localization, partners, regulation, disease links, and quantitative points, explicitly distinguishing direct human evidence from homolog inference:
| Category | Evidence summary | Key references with DOI/URL and publication date | Citation IDs |
|---|---|---|---|
| Identity/domains | Verified target: human USO1 encodes general vesicular transport factor p115 (also called p115/USO1), a Golgi/ER-Golgi tethering factor. Structural evidence from homolog work shows an N-terminal globular head with armadillo-repeat/α-solenoid architecture and a long coiled-coil tail; human-focused studies and reviews describe p115 as a dimer with N-terminal head and coiled-coil tail. Note: direct domain-resolution evidence here is mainly from yeast/fungal homologs and review synthesis, not a new 2023–2024 human structure. (heo2020crystalstructuresof pages 1-2, yakunin2023characterisingthefactors pages 36-39) | Heo Y et al. Sci Rep (2020), doi:10.1038/s41598-020-66480-1, https://doi.org/10.1038/s41598-020-66480-1, published Jun 2020; Yakunin I. PhD thesis (2023), doi:10.17863/cam.93885, https://doi.org/10.17863/cam.93885, published Feb 2023 | (heo2020crystalstructuresof pages 1-2, yakunin2023characterisingthefactors pages 36-39) |
| Core molecular function | Human evidence: p115/USO1 is a Golgi vesicle tether required for ER-to-Golgi/early secretory trafficking, vesicle docking, and secretion efficiency; recent human-cell work also supports direct binding of p115 to Sec16A, an ER exit site organizer, linking p115 to early secretory pathway organization. Homolog evidence: 2023 fungal work argues the globular head directly binds SNAREs and that long-range coiled-coil tethering may be partly dispensable, emphasizing a key role in SNARE regulation/fusion competence. (yakunin2025thegolgivesicle pages 1-5, yakunin2025thegolgivesicle pages 11-14, bravoplaza2023theuso1globular pages 1-2) | Yakunin IG et al. bioRxiv preprint, doi:10.1101/2025.10.16.682774, https://doi.org/10.1101/2025.10.16.682774, posted Oct 2025; Bravo-Plaza I et al. eLife (2023), doi:10.7554/eLife.85079, https://doi.org/10.7554/elife.85079, published May 30 2023 | (yakunin2025thegolgivesicle pages 1-5, yakunin2025thegolgivesicle pages 11-14, bravoplaza2023theuso1globular pages 1-2) |
| Key interaction partners | Human-supported/reviewed partners: RAB1A binds the N-terminus; GM130 binds p115 at its N-terminus in a mitotically regulated manner; p115 also interacts with giantin at the C-terminus and with cis-Golgi SNAREs including Sec22b, Bet1, GOSR2, syntaxin-5 in proposed SNARE-assembly/fusion models. Human disease/pathogen context: Legionella effector LegA15 physically hijacks host p115. Homolog support: fungal/yeast studies support direct binding to Bet1/Bos1/Sec22-family SNAREs and recruitment dependence on Rab1/golgin systems. (chen2022atypicallegionellagtpase pages 1-2, yakunin2023characterisingthefactors pages 36-39, bravoplaza2022tetheringbyuso1 pages 20-23) | Chen T-T et al. Sci Adv (2022), doi:10.1126/sciadv.add7945, https://doi.org/10.1126/sciadv.add7945, published Dec 16 2022; Yakunin I. PhD thesis (2023), doi:10.17863/cam.93885, https://doi.org/10.17863/cam.93885, published Feb 2023; Bravo-Plaza I et al. bioRxiv (2022), doi:10.1101/2022.11.30.518472, https://doi.org/10.1101/2022.11.30.518472, posted Dec 2022 | (chen2022atypicallegionellagtpase pages 1-2, yakunin2023characterisingthefactors pages 36-39, bravoplaza2022tetheringbyuso1 pages 20-23) |
| Subcellular localization | Human evidence/review: p115 localizes to the ERGIC and cis-Golgi, associates with Golgi membranes in a Rab1-dependent manner, and functions at the ER–Golgi interface; recent human-cell work also links it to ER exit sites via Sec16A. Homolog evidence: fungal Uso1 cycles on early Golgi/ER-Golgi puncta. (yakunin2025thegolgivesicle pages 1-5, yakunin2025thegolgivesicle pages 11-14, bravoplaza2023theuso1globular pages 1-2) | Yakunin IG et al. bioRxiv preprint, doi:10.1101/2025.10.16.682774, https://doi.org/10.1101/2025.10.16.682774, posted Oct 2025; Bravo-Plaza I et al. eLife (2023), doi:10.7554/eLife.85079, https://doi.org/10.7554/elife.85079, published May 30 2023 | (yakunin2025thegolgivesicle pages 1-5, yakunin2025thegolgivesicle pages 11-14, bravoplaza2023theuso1globular pages 1-2) |
| Pathways/processes | USO1/p115 functions in the early secretory pathway, especially COPII-derived ER-to-Golgi transport, vesicle tethering/docking, and likely cis-SNARE complex assembly/fusion. In broader cell-biology/immune reviews, Golgi tethers including p115 are discussed as supporting cytokine secretion and Golgi-dependent innate immune signaling logistics, but this is contextual rather than USO1-specific mechanistic proof. (heo2020crystalstructuresof pages 1-2, yakunin2023characterisingthefactors pages 36-39, maruntelu2024thegolgiapparatus pages 1-2) | Heo Y et al. Sci Rep (2020), doi:10.1038/s41598-020-66480-1, https://doi.org/10.1038/s41598-020-66480-1, published Jun 2020; Yakunin I. PhD thesis (2023), doi:10.17863/cam.93885, https://doi.org/10.17863/cam.93885, published Feb 2023; Mărunțelu I et al. Int J Mol Sci (2024), doi:10.3390/ijms25074120, https://doi.org/10.3390/ijms25074120, published Apr 8 2024 | (heo2020crystalstructuresof pages 1-2, yakunin2023characterisingthefactors pages 36-39, maruntelu2024thegolgiapparatus pages 1-2) |
| Regulation/PTMs | Human review evidence: the GM130–p115 interaction is blocked by Cdk1 phosphorylation of GM130 S37 during mitosis, indicating cell-cycle regulation of p115 tethering. Human phosphoproteomics: USO1 Ser48 was reported in RA synovial phosphoproteomic analysis as inversely correlated with ESR, suggesting regulated phosphorylation in disease tissue, but this is associative and not mechanistically assigned to trafficking. (yakunin2023characterisingthefactors pages 36-39, cubuk2024phosphoproteomicprofilingof pages 1-2) | Yakunin I. PhD thesis (2023), doi:10.17863/cam.93885, https://doi.org/10.17863/cam.93885, published Feb 2023; Çubuk C et al. Arthritis Res Ther (2024), doi:10.1186/s13075-024-03351-4, https://doi.org/10.1186/s13075-024-03351-4, published Jun 2024 | (yakunin2023characterisingthefactors pages 36-39, cubuk2024phosphoproteomicprofilingof pages 1-2) |
| Disease/clinical associations | Cancer: in human HCC, a tumor-associated isoform USO1-T correlates with poorer prognosis, promotes aggressive behavior, causes Golgi unstacking, and accelerates trafficking from ER to Golgi/plasma membrane; modeling suggests weakened dimerization/GM130 tethering and involvement of ERK and GRASP65. Additional literature summaries link USO1/p115 to colon cancer, gastric cancer, multiple myeloma, NSCLC, and leukemia vulnerability/fusions, though quantitative details are limited in the available snippets. Infection: Legionella LegA15 hijacks host p115, causing Golgi fragmentation and altered lipid-droplet homeostasis. (yoon2021uso1isoformsdifferentially pages 1-2, yoon2021uso1isoformsdifferentially pages 13-13, li2025uso1coordinatescentriolar pages 13-14, chen2022atypicallegionellagtpase pages 1-2) | Yoon S et al. Carcinogenesis (2021), doi:10.1093/carcin/bgab067, https://doi.org/10.1093/carcin/bgab067, published Jul 22 2021; Chen T-T et al. Sci Adv (2022), doi:10.1126/sciadv.add7945, https://doi.org/10.1126/sciadv.add7945, published Dec 16 2022 | (yoon2021uso1isoformsdifferentially pages 1-2, yoon2021uso1isoformsdifferentially pages 13-13, li2025uso1coordinatescentriolar pages 13-14, chen2022atypicallegionellagtpase pages 1-2) |
| Recent developments 2023-2024 | 2023: homolog-based mechanistic work showed the Uso1/p115 globular head directly binds SNAREs and can sustain essential function without the coiled-coil tether, sharpening the model from “long tether” to SNARE-regulatory tether/fusion factor. 2024: Golgi/innate-immunity review highlights p115 among Golgi tethers relevant to secretory and immune processes; RA phosphoproteomics identified USO1 Ser48 as a disease-associated phosphosite. Caveat: among retrieved items, truly human USO1-specific primary mechanistic advances in 2023–2024 were limited. (bravoplaza2023theuso1globular pages 1-2, maruntelu2024thegolgiapparatus pages 1-2, cubuk2024phosphoproteomicprofilingof pages 1-2) | Bravo-Plaza I et al. eLife (2023), doi:10.7554/eLife.85079, https://doi.org/10.7554/elife.85079, published May 30 2023; Mărunțelu I et al. Int J Mol Sci (2024), doi:10.3390/ijms25074120, https://doi.org/10.3390/ijms25074120, published Apr 8 2024; Çubuk C et al. Arthritis Res Ther (2024), doi:10.1186/s13075-024-03351-4, https://doi.org/10.1186/s13075-024-03351-4, published Jun 2024 | (bravoplaza2023theuso1globular pages 1-2, maruntelu2024thegolgiapparatus pages 1-2, cubuk2024phosphoproteomicprofilingof pages 1-2) |
| Quantitative data points | Homolog quantitative data: fungal Uso1 puncta half-life ~60 s; Pearson colocalization with GeaA/Gea1,2 0.52 ± 0.06 (n=16) and with RAB1 0.61 ± 0.07 (n=20). Human/pathogen quantitative data: LegA15 interactome identified host p115/USO1 with 62 unique peptides in MS. Human trafficking assay framework: HCC study quantified VSVG trafficking as mean % cells ± SEM with n=5–8 fields, 10–50 cells/field, but exact values were not in the available snippet. Human interaction assay parameters (preprint): RUSH assay used 1 mM biotin, 35 min progression, 3 biological repeats. (bravoplaza2023theuso1globular pages 1-2, bravoplaza2022tetheringbyuso1 pages 11-13, chen2022atypicallegionellagtpase media 41416518, yoon2021uso1isoformsdifferentially pages 9-10, yakunin2025thegolgivesicle pages 11-14) | Bravo-Plaza I et al. eLife (2023), doi:10.7554/eLife.85079, https://doi.org/10.7554/elife.85079, published May 30 2023; Bravo-Plaza I et al. bioRxiv (2022), doi:10.1101/2022.11.30.518472, https://doi.org/10.1101/2022.11.30.518472, posted Dec 2022; Chen T-T et al. Sci Adv (2022), doi:10.1126/sciadv.add7945, https://doi.org/10.1126/sciadv.add7945, published Dec 16 2022; Yoon S et al. Carcinogenesis (2021), doi:10.1093/carcin/bgab067, https://doi.org/10.1093/carcin/bgab067, published Jul 22 2021; Yakunin IG et al. bioRxiv preprint, doi:10.1101/2025.10.16.682774, https://doi.org/10.1101/2025.10.16.682774, posted Oct 2025 | (bravoplaza2023theuso1globular pages 1-2, bravoplaza2022tetheringbyuso1 pages 11-13, chen2022atypicallegionellagtpase media 41416518, yoon2021uso1isoformsdifferentially pages 9-10, yakunin2025thegolgivesicle pages 11-14) |
Table: This table summarizes supported functional annotation for human USO1/p115 (UniProt O60763), separating direct human evidence from inference based on yeast/fungal homologs. It highlights function, localization, partners, disease links, and the strongest quantitative findings available from the gathered evidence.
Figure-based evidence (visual)
The following figure crops provide visual support for key quantitative and mechanistic claims regarding host p115/USO1 in Legionella infection biology:
- Proteomics evidence identifying p115/USO1 with 62 unique peptides as a top LegA15 interactor. (chen2022atypicallegionellagtpase media 41416518)
- Model schematic of LegA15 hijacking host p115 to impact LD/LCV interactions and Golgi fragmentation. (chen2022atypicallegionellagtpase media b798ae8b)
Concluding functional annotation (most defensible claims)
- Core role: Human USO1/p115 is a conserved ERGIC/cis-Golgi vesicle tether/fusion factor in early secretory trafficking, connecting Rab1-regulated vesicle capture/docking with SNARE-dependent membrane fusion. (yakunin2025thegolgivesicle pages 1-5, yakunin2023characterisingthefactors pages 36-39)
- Key interactions: Robustly supported interaction axes include Rab1A, GM130 (mitotically regulated via GM130 S37 phosphorylation), and cis-Golgi SNAREs (Sec22b/Bet1/GOSR2/STX5 in proposed models); newer work also links p115 to Sec16A at ER exit sites. (yakunin2023characterisingthefactors pages 36-39, yakunin2025thegolgivesicle pages 1-5)
- Localization: p115/USO1 localizes to ERGIC and cis-Golgi and functionally couples to ER exit sites (Sec16A linkage). (yakunin2025thegolgivesicle pages 1-5)
- Human disease relevance: USO1 is implicated in cancer trafficking/Golgi organization phenotypes (HCC isoform effects) and appears in human disease phosphoproteomes (USO1 Ser48 association in RA tissue). (yoon2021uso1isoformsdifferentially pages 1-2, cubuk2024phosphoproteomicprofilingof pages 1-2)
- Real-world exploitation: Host p115/USO1 is directly hijacked by Legionella effector LegA15, supported by proteomics (62 unique peptides) and mechanistic modeling. (chen2022atypicallegionellagtpase media 41416518, chen2022atypicallegionellagtpase media b798ae8b)
Reference URLs and publication dates (from retrieved sources)
- Bravo-Plaza I. et al. eLife (published 2023-05-30): https://doi.org/10.7554/elife.85079 (bravoplaza2023theuso1globular pages 1-2)
- Çubuk C. et al. Arthritis Research & Therapy (published 2024-06, DOI page): https://doi.org/10.1186/s13075-024-03351-4 (cubuk2024phosphoproteomicprofilingof pages 1-2)
- Mărunțelu I. et al. International Journal of Molecular Sciences (published 2024-04-08): https://doi.org/10.3390/ijms25074120 (maruntelu2024thegolgiapparatus pages 1-2)
- Chen T.-T. et al. Science Advances (published 2022-12-16): https://doi.org/10.1126/sciadv.add7945 (chen2022atypicallegionellagtpase pages 1-2)
- Yoon S. et al. Carcinogenesis (advance access 2021-07-22): https://doi.org/10.1093/carcin/bgab067 (yoon2021uso1isoformsdifferentially pages 1-2)
- Yakunin I. PhD thesis/dissertation (published 2023-02): https://doi.org/10.17863/cam.93885 (yakunin2023characterisingthefactors pages 36-39)
References
-
(yakunin2025thegolgivesicle pages 1-5): Igor Yakunin, Alison K Gillingham, Conceição Pereira, David C Gershlick, and Sean Munro. The golgi vesicle tether p115/uso1 can bind directly to the er exit site organiser sec16a. BioRxiv, Oct 2026. URL: https://doi.org/10.1101/2025.10.16.682774, doi:10.1101/2025.10.16.682774. This article has 1 citations.
-
(yakunin2023characterisingthefactors pages 36-39): Characterising the factors involved in ER-to-Golgi anterograde trafficking This article has 0 citations.
-
(yakunin2025thegolgivesicle pages 11-14): Igor Yakunin, Alison K Gillingham, Conceição Pereira, David C Gershlick, and Sean Munro. The golgi vesicle tether p115/uso1 can bind directly to the er exit site organiser sec16a. BioRxiv, Oct 2026. URL: https://doi.org/10.1101/2025.10.16.682774, doi:10.1101/2025.10.16.682774. This article has 1 citations.
-
(heo2020crystalstructuresof pages 1-2): Yoonyoung Heo, Hye-Jin Yoon, Hanseo Ko, Soonmin Jang, and Hyung Ho Lee. Crystal structures of uso1 membrane tether reveal an alternative conformation in the globular head domain. Scientific Reports, Jun 2020. URL: https://doi.org/10.1038/s41598-020-66480-1, doi:10.1038/s41598-020-66480-1. This article has 14 citations and is from a peer-reviewed journal.
-
(bravoplaza2023theuso1globular pages 1-2): Ignacio Bravo-Plaza, Victor G Tagua, Herbert N Arst, Ana Alonso, Mario Pinar, Begoña Monterroso, Antonio Galindo, and Miguel A Peñalva. The uso1 globular head interacts with snares to maintain viability even in the absence of the coiled-coil domain. eLife, May 2023. URL: https://doi.org/10.7554/elife.85079, doi:10.7554/elife.85079. This article has 10 citations and is from a domain leading peer-reviewed journal.
-
(bravoplaza2023theuso1globular pages 27-30): Ignacio Bravo-Plaza, Victor G Tagua, Herbert N Arst, Ana Alonso, Mario Pinar, Begoña Monterroso, Antonio Galindo, and Miguel A Peñalva. The uso1 globular head interacts with snares to maintain viability even in the absence of the coiled-coil domain. eLife, May 2023. URL: https://doi.org/10.7554/elife.85079, doi:10.7554/elife.85079. This article has 10 citations and is from a domain leading peer-reviewed journal.
-
(cubuk2024phosphoproteomicprofilingof pages 1-2): Cankut Çubuk, Rachel Lau, Pedro Cutillas, Vinothini Rajeeve, Christopher R. John, Anna E. A. Surace, Rebecca Hands, Liliane Fossati-Jimack, Myles J. Lewis, and Costantino Pitzalis. Phosphoproteomic profiling of early rheumatoid arthritis synovium reveals active signalling pathways and differentiates inflammatory pathotypes. Arthritis Research & Therapy, Jun 2024. URL: https://doi.org/10.1186/s13075-024-03351-4, doi:10.1186/s13075-024-03351-4. This article has 12 citations and is from a domain leading peer-reviewed journal.
-
(maruntelu2024thegolgiapparatus pages 1-2): Ion Mărunţelu, Alexandra-Elena Constantinescu, Razvan-Adrian Covache-Busuioc, and Ileana Constantinescu. The golgi apparatus: a key player in innate immunity. International Journal of Molecular Sciences, 25:4120, Apr 2024. URL: https://doi.org/10.3390/ijms25074120, doi:10.3390/ijms25074120. This article has 20 citations.
-
(chen2022atypicallegionellagtpase pages 1-2): Tao-Tao Chen, Yanling Lin, Shijun Zhang, Shuxin Liu, Lei Song, Wenhong Zhong, Zhao-Qing Luo, and Aidong Han. Atypical legionella gtpase effector hijacks host vesicular transport factor p115 to regulate host lipid droplet. Science Advances, Dec 2022. URL: https://doi.org/10.1126/sciadv.add7945, doi:10.1126/sciadv.add7945. This article has 15 citations and is from a highest quality peer-reviewed journal.
-
(chen2022atypicallegionellagtpase media 41416518): Tao-Tao Chen, Yanling Lin, Shijun Zhang, Shuxin Liu, Lei Song, Wenhong Zhong, Zhao-Qing Luo, and Aidong Han. Atypical legionella gtpase effector hijacks host vesicular transport factor p115 to regulate host lipid droplet. Science Advances, Dec 2022. URL: https://doi.org/10.1126/sciadv.add7945, doi:10.1126/sciadv.add7945. This article has 15 citations and is from a highest quality peer-reviewed journal.
-
(chen2022atypicallegionellagtpase media b798ae8b): Tao-Tao Chen, Yanling Lin, Shijun Zhang, Shuxin Liu, Lei Song, Wenhong Zhong, Zhao-Qing Luo, and Aidong Han. Atypical legionella gtpase effector hijacks host vesicular transport factor p115 to regulate host lipid droplet. Science Advances, Dec 2022. URL: https://doi.org/10.1126/sciadv.add7945, doi:10.1126/sciadv.add7945. This article has 15 citations and is from a highest quality peer-reviewed journal.
-
(yoon2021uso1isoformsdifferentially pages 1-2): Sarah Yoon, Ji-Hye Choi, Masaud Shah, So Mee Kwon, Jieun Yang, Young Nyun Park, Hee-Jung Wang, and Hyun Goo Woo. Uso1 isoforms differentially promote liver cancer progression by dysregulating the er-golgi network. Carcinogenesis, 42:1208-1220, Jul 2021. URL: https://doi.org/10.1093/carcin/bgab067, doi:10.1093/carcin/bgab067. This article has 13 citations and is from a peer-reviewed journal.
-
(yoon2021uso1isoformsdifferentially pages 7-8): Sarah Yoon, Ji-Hye Choi, Masaud Shah, So Mee Kwon, Jieun Yang, Young Nyun Park, Hee-Jung Wang, and Hyun Goo Woo. Uso1 isoforms differentially promote liver cancer progression by dysregulating the er-golgi network. Carcinogenesis, 42:1208-1220, Jul 2021. URL: https://doi.org/10.1093/carcin/bgab067, doi:10.1093/carcin/bgab067. This article has 13 citations and is from a peer-reviewed journal.
-
(bravoplaza2022tetheringbyuso1 pages 11-13): Ignacio Bravo-Plaza, Víctor G. Tagua, Herbert N. Arst, Ana Alonso, Mario Pinar, Begoña Monterroso, Antonio Galindo, and Miguel Á. Peñalva. Tethering by uso1 is dispensable: the uso1 monomeric globular head domain interacts with snares to maintain viability. bioRxiv, Dec 2022. URL: https://doi.org/10.1101/2022.11.30.518472, doi:10.1101/2022.11.30.518472. This article has 0 citations.
-
(yoon2021uso1isoformsdifferentially pages 9-10): Sarah Yoon, Ji-Hye Choi, Masaud Shah, So Mee Kwon, Jieun Yang, Young Nyun Park, Hee-Jung Wang, and Hyun Goo Woo. Uso1 isoforms differentially promote liver cancer progression by dysregulating the er-golgi network. Carcinogenesis, 42:1208-1220, Jul 2021. URL: https://doi.org/10.1093/carcin/bgab067, doi:10.1093/carcin/bgab067. This article has 13 citations and is from a peer-reviewed journal.
-
(bravoplaza2022tetheringbyuso1 pages 20-23): Ignacio Bravo-Plaza, Víctor G. Tagua, Herbert N. Arst, Ana Alonso, Mario Pinar, Begoña Monterroso, Antonio Galindo, and Miguel Á. Peñalva. Tethering by uso1 is dispensable: the uso1 monomeric globular head domain interacts with snares to maintain viability. bioRxiv, Dec 2022. URL: https://doi.org/10.1101/2022.11.30.518472, doi:10.1101/2022.11.30.518472. This article has 0 citations.
-
(yoon2021uso1isoformsdifferentially pages 13-13): Sarah Yoon, Ji-Hye Choi, Masaud Shah, So Mee Kwon, Jieun Yang, Young Nyun Park, Hee-Jung Wang, and Hyun Goo Woo. Uso1 isoforms differentially promote liver cancer progression by dysregulating the er-golgi network. Carcinogenesis, 42:1208-1220, Jul 2021. URL: https://doi.org/10.1093/carcin/bgab067, doi:10.1093/carcin/bgab067. This article has 13 citations and is from a peer-reviewed journal.
-
(li2025uso1coordinatescentriolar pages 13-14): Xinyi Li, Peiyi Lin, Zaikuan Zhang, Runzhi Wang, Jing Cai, Xiaosong Feng, Zhihong Jiang, Shengming Xu, and Yajun Xie. Uso1 coordinates centriolar satellites to regulate male germ cell proliferation and cell cycle progression. International Journal of Molecular Sciences, 26:4274, Apr 2025. URL: https://doi.org/10.3390/ijms26094274, doi:10.3390/ijms26094274. This article has 0 citations.
Artifacts
Citations
- yakunin2023characterisingthefactors pages 36-39
- yakunin2025thegolgivesicle pages 1-5
- cubuk2024phosphoproteomicprofilingof pages 1-2
- maruntelu2024thegolgiapparatus pages 1-2
- chen2022atypicallegionellagtpase pages 1-2
- yakunin2025thegolgivesicle pages 11-14
- heo2020crystalstructuresof pages 1-2
- https://doi.org/10.1038/s41598-020-66480-1,
- https://doi.org/10.17863/cam.93885,
- https://doi.org/10.1101/2025.10.16.682774,
- https://doi.org/10.7554/elife.85079,
- https://doi.org/10.1126/sciadv.add7945,
- https://doi.org/10.1101/2022.11.30.518472,
- https://doi.org/10.3390/ijms25074120,
- https://doi.org/10.1186/s13075-024-03351-4,
- https://doi.org/10.1093/carcin/bgab067,
- https://doi.org/10.7554/elife.85079
- https://doi.org/10.1186/s13075-024-03351-4
- https://doi.org/10.3390/ijms25074120
- https://doi.org/10.1126/sciadv.add7945
- https://doi.org/10.1093/carcin/bgab067
- https://doi.org/10.17863/cam.93885
- https://doi.org/10.3390/ijms26094274,
OpenAI
(USO1-deep-research-openai.md)
OpenAI
(USO1-deep-research-openai.md)Overview of USO1 and its Protein Product (p115)
USO1 (HGNC symbol for “USO1 vesicle transport factor”) is a human gene encoding the protein commonly known as p115, or General Vesicular Transport Factor p115 (research.bioinformatics.udel.edu). This protein was originally identified in yeast (Uso1p) and is highly conserved in eukaryotes (pubmed.ncbi.nlm.nih.gov). Human p115 is a large coiled-coil peripheral membrane protein of approximately 959 amino acids (~115 kDa for the homodimer) (pmc.ncbi.nlm.nih.gov) (www.ncbi.nlm.nih.gov). It operates as a homodimer and is often described as having a “myosin-shaped” architecture, with an N-terminal globular head domain, a long coiled-coil rod domain that mediates dimerization, and a short acidic C-terminal tail (www.ncbi.nlm.nih.gov). USO1 is ubiquitously expressed across tissues (e.g. high in thyroid and testis), reflecting its fundamental role in secretory trafficking (www.ncbi.nlm.nih.gov). Major known aliases for USO1 include VDP (vesicle docking protein), TAP (transcytosis-associated protein), and “vesicle docking protein homolog (yeast),” underscoring its role in vesicle tethering and its conservation from yeast to humans (www.genecards.org).
Key Concept – Membrane Tethering: p115 is a vesicle tethering factor that facilitates the docking of transport vesicles to their target membranes within the early secretory pathway (pmc.ncbi.nlm.nih.gov). Unlike enzymes, p115 does not catalyze a chemical reaction; rather, it functions as a structural adaptor or scaffold that physically links membranes to promote vesicle fusion. The current understanding is that p115 plays a central role in ER-to-Golgi and intra-Golgi transport, capturing incoming vesicles and holding them near the Golgi membrane until SNARE proteins can mediate fusion (www.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In practical terms, USO1/p115 is essential for maintaining efficient protein and lipid trafficking from the endoplasmic reticulum (ER) to the Golgi apparatus and between Golgi cisternae. Its activity is critical for normal cellular secretory function, and loss of p115 leads to severe trafficking defects and disorganization of Golgi structure (pmc.ncbi.nlm.nih.gov). In the sections below, we delve into the structure, localization, primary function, regulatory mechanisms, and pathways involving USO1, and we highlight recent research (2020–2024) on its roles and significance. Citations to primary literature and authoritative reviews are provided throughout to support each statement.
Structure and Localization of p115
Domain Organization: The p115 protein consists of distinct domains that underlie its function. The N-terminal portion (~1–300 amino acids) contains armadillo/HEAT repeat motifs (an armadillo-fold) (www.frontiersin.org) (www.ncbi.nlm.nih.gov). This globular head domain is conserved but was long of unknown function; newer evidence suggests it mediates interactions beyond vesicle tethering (e.g. binding to cytosolic factors) (www.ncbi.nlm.nih.gov). The central region of p115 is a lengthy coiled-coil that forms an elongated rod; this region self-associates into an anti-parallel homodimer, effectively creating a dumbbell-shaped dimer with two globular heads and a rod-like tail (www.ncbi.nlm.nih.gov). The coiled-coil is subdivided into at least four segments (CC1–CC4) based on sequence homology. Importantly, specific coiled-coil segments mediate binding to other proteins: for example, the first coiled-coil (CC1) and the fourth coiled-coil (CC4) of p115 can bind to specific ER–Golgi SNARE proteins (pubmed.ncbi.nlm.nih.gov). The extreme C-terminal tail (~30 amino acids) of p115 is highly acidic and is crucial for tethering activity (www.ncbi.nlm.nih.gov). This tail region is responsible for binding the target membrane “golgin” proteins (such as GM130) and vesicle-associated coiled-coil proteins (such as giantin), as described below (www.ncbi.nlm.nih.gov).
Subcellular Localization: USO1-encoded p115 localizes to the cytoplasmic face of membranes in the early secretory pathway. During interphase, p115 cycles between the cytosol and the Golgi apparatus (www.ncbi.nlm.nih.gov). It is predominantly found at the cis-Golgi network and ER–Golgi intermediate compartment (ERGIC), including on vesicular carriers that shuttle between ER and Golgi (www.frontiersin.org). Because p115 lacks any transmembrane region, it associates peripherally: in its active, dephosphorylated state, p115 binds to Golgi membranes, whereas phosphorylation causes it to dissociate back into the cytosol (www.ncbi.nlm.nih.gov). High-resolution microscopy studies have shown p115 concentrated at Golgi rims and vesicle clustering sites near ER exit sites (www.frontiersin.org). In fact, small GTPases of the Rab family help target p115 to these locations: the Rab1 GTPase (previously called Ras-associated protein 1) specifically recruits p115 to ER-derived vesicles (COPII-coated vesicles) at ER exit sites (www.ncbi.nlm.nih.gov). This recruitment localizes p115 to sites where nascent vesicles bud from the ER, positioning it to capture those vesicles upon their arrival at the Golgi. p115 is thus often found at the interface of ER exit sites and the cis-Golgi.
Notably, p115’s distribution changes during the cell cycle. In mitosis, the Golgi apparatus disassembles and p115 disperses; interestingly, a fraction of p115 relocalizes to the spindle poles during mitosis (www.frontiersin.org). This mitotic relocalization is mediated by the N-terminal armadillo domain of p115 binding to γ-tubulin complexes, which are key microtubule nucleation factors (www.frontiersin.org). Through this interaction, p115 contributes to non-centrosomal microtubule organizing centers (MTOCs) at the Golgi in interphase, and it associates with centrosomal structures in mitosis (www.frontiersin.org). In summary, under normal interphase conditions p115 is primarily a Golgi-localized peripheral membrane protein (with rapid on-and-off membrane cycling), whereas during mitosis it transiently adopts a spindle pole localization, reflecting its regulatory interactions with the cytoskeleton.
Role in Vesicle Tethering and Membrane Trafficking
The primary function of USO1/p115 is to act as a vesicle tethering factor in the early secretory pathway. Tethering factors ensure that transport vesicles are correctly docked to their target membrane before SNARE-mediated fusion occurs. p115 is considered a “general” vesicle transport factor because it is required at multiple steps of membrane traffic, especially ER-to-Golgi transport and intra-Golgi transport (www.frontiersin.org) (pmc.ncbi.nlm.nih.gov). It does not have enzymatic activity or cargo specificity; instead, it recognizes membrane-bound partner proteins to form a physical bridge between a vesicle and the target compartment.
Mechanism of Tethering: Classic studies in mammalian cells and yeast revealed that p115 bridges vesicles to the Golgi by simultaneously binding factors on both membranes. On the Golgi side, p115 interacts with GM130 (Golgin-95/GOLGA2), a coiled-coil Golgi matrix protein anchored to the cis-Golgi membrane (pmc.ncbi.nlm.nih.gov). On the vesicle side, p115 binds to giantin (GOLGB1), a long coiled-coil protein that can be carried by Golgi-derived vesicles (pmc.ncbi.nlm.nih.gov). This trimeric complex – GM130–p115–giantin – forms a tethering bridge that holds the vesicle at the Golgi surface (pmc.ncbi.nlm.nih.gov). Sönnichsen et al. first demonstrated in 1998 that p115 and these golgins form a complex required for Golgi vesicle docking (pmc.ncbi.nlm.nih.gov). The acidic tail of p115 is critical for these interactions: it directly binds a coiled-coil region of giantin on the vesicle and also binds GM130 on the Golgi membrane (www.ncbi.nlm.nih.gov). By connecting giantin and GM130, p115 helps maintain the structural integrity of the Golgi and ensures vesicles (particularly COPI-coated vesicles) are tethered in place for fusion (www.ncbi.nlm.nih.gov). Indeed, biochemical assays have confirmed that p115 can simultaneously bind GM130 and giantin, forming a “tentacle”-like link between two membranes (pmc.ncbi.nlm.nih.gov).
In the context of anterograde ER-to-Golgi transport, p115 collaborates with Rab1 and other factors. Rab1 in its GTP-bound form localizes to COPII vesicles budding from the ER and recruits p115 onto these vesicles (www.ncbi.nlm.nih.gov). Once the vesicle arrives at the Golgi vicinity, p115 (on the vesicle) can engage GM130 (on the cis-Golgi), thereby tethering the vesicle at the Golgi membrane (www.ncbi.nlm.nih.gov). This mechanism effectively “programs” budding COPII vesicles for fusion with Golgi by pre-loading them with the tethering machinery (reactome.org) (www.ncbi.nlm.nih.gov). Moyer et al. (2001) showed that a Rab1–p115–GM130 complex is required for efficient tethering of ER-derived vesicles to the cis-Golgi (reactome.org). Consistently, depletion or inactivation of p115 causes accumulation of cargo in ER-derived vesicles that fail to fuse, as observed both in cell culture and whole organisms (pubmed.ncbi.nlm.nih.gov). For example, C. elegans embryos lacking p115 activity accumulate secretory yolk proteins in the body cavity and show ER/Golgi retention of cargo receptors, phenotypes consistent with blocked vesicle fusion (pubmed.ncbi.nlm.nih.gov).
SNARE Complex Assembly: Beyond simply holding vesicles near the Golgi, p115 actively contributes to the next step: assembling the SNARE complex that drives membrane fusion. p115 has been shown to bind directly to multiple ER–Golgi SNARE proteins via its coiled-coil domains (pubmed.ncbi.nlm.nih.gov). Specifically, the CC1 region of p115 has a weak homology to SNARE motifs and can bind to SNAREs on the target membrane (t-SNAREs like syntaxin 5) (reactome.org), while another region (CC4) can bind vesicle SNAREs (pubmed.ncbi.nlm.nih.gov). This multivalent binding may help align the SNAREs from vesicle (v-SNAREs) and Golgi (t-SNAREs) into the proper complex (sometimes termed a “cis-SNARE complex” when formed prior to membrane fusion) (www.ncbi.nlm.nih.gov). Shorter et al. (2002) demonstrated that p115 catalyzes SNAREpin assembly after the initial tethering: p115 first brings Golgi golgins together, and then facilitates the pairing of SNARE proteins, thereby accelerating vesicle fusion (reactome.org). In essence, p115 acts as a platform that coordinates tethering with the molecular fusion machinery. The outcome is efficient docking and fusion of incoming vesicles, delivering their cargo into the Golgi.
Pathways and Processes: The activity of p115 is indispensable for multiple transport steps:
- ER to Golgi transport: p115 is critical for forward trafficking of proteins from the ER. Genetic interference (siRNA or temperature-sensitive mutations) leads to a blockade of ER export and collapse of Golgi function (www.frontiersin.org). In mammalian cells, p115 is required for the formation of the Golgi ribbon structure from ER-Golgi intermediate elements, as it helps tether vesicular and tubular intermediates during Golgi assembly (pubmed.ncbi.nlm.nih.gov).
- Intra-Golgi and retrograde transport: p115’s interaction with COPI vesicles suggests it also functions in retrograde trafficking (Golgi-to-ER or intra-Golgi movement). It is required for inter-cisternal Golgi transport – the exchange of material between Golgi cisternae to maintain Golgi homeostasis (www.ncbi.nlm.nih.gov). For example, in cell-free systems p115 is necessary for transferring cargo between successive Golgi compartments (www.ncbi.nlm.nih.gov).
- Golgi biogenesis and maintenance: Because p115 tethers membranes, it contributes to the overall integrity of the Golgi. Silencing p115 in cultured cells causes the Golgi to fragment or disperse into vesicles (pmc.ncbi.nlm.nih.gov). Conversely, during post-mitotic Golgi reassembly, p115 is one of the key factors (along with GM130 and GRASP65) that relink Golgi fragments into a unified ribbon (www.frontiersin.org). These roles place p115 at the heart of Golgi morphogenesis.
Experimentally, the essential nature of USO1/p115 is well documented. In mice, a knockout of USO1 leads to early embryonic lethality, with embryos failing shortly after the blastocyst stage (around E3–E8.5) (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). The few cells observed in those mutants showed disrupted Golgi architecture, underscoring how vital p115-mediated tethering is for cell viability (pmc.ncbi.nlm.nih.gov). Even in conditional or partial knockdowns, cells often exhibit Golgi disorganization, secretion defects, and growth arrest (pmc.ncbi.nlm.nih.gov). Thus, the consensus from functional studies is that p115 is indispensable for secretory trafficking and Golgi structure across eukaryotes.
Regulation and Interaction Partners of p115
Recruitment by Rab GTPases: A key regulator of p115 is Rab1, a small GTP-binding protein that operates at ER–Golgi interfaces. Active Rab1 (Rab1–GTP) on ER-derived vesicles serves as a signal to recruit p115 from the cytosol to the vesicle membrane (www.ncbi.nlm.nih.gov). This recruitment is part of a larger “tethering complex”: Rab1–GTP binds not only p115 but also Golgi-side factors like GM130 and the golgin tethering complex GRASP65 (GORASP1) (reactome.org) (reactome.org). Reactome pathway reconstructions show a specific step where Rab1:GTP binds USO1 (p115) and GM130/GRASP65 at the Golgi membrane, initiating the tethering of an incoming vesicle (reactome.org). The dependence on Rab1 explains why loss of Rab1 function phenocopies p115 loss – both result in ER-to-Golgi traffic jams. Other Rabs might also interact with p115 (for instance, Rab2 has been implicated in Golgi trafficking), but Rab1 is the best-characterized direct recruiter (reactome.org).
Phosphorylation Cycling: p115’s association with membranes is controlled by phosphorylation. During interphase, p115 is largely dephosphorylated and stays bound to Golgi membranes; when phosphorylated (on specific serine/threonine sites), it releases from membranes into the cytosol (www.ncbi.nlm.nih.gov). One well-known regulatory event occurs in mitosis: the mitotic kinase CDK1/cyclin B phosphorylates GM130 on serine-25, which in turn prevents p115 from binding GM130 (reactome.org). This disrupts the p115–GM130 tethering and contributes to Golgi disassembly at prophase, a normal step in cell division. p115 itself is also a phosphoprotein; casein kinase II and other kinases can phosphorylate p115 and modulate its activity (www.frontiersin.org). For instance, studies indicate that p115’s ability to bind GM130 and giantin is phosphorylation-dependent (www.frontiersin.org). Brandon et al. (2003) showed that p115 phosphorylation status changes its membrane affinity (www.frontiersin.org). Thus, cells regulate p115 by kinase signaling to toggle its tethering function on or off as needed (e.g., switching off tethering during mitosis, or possibly modulating tethering during signaling events).
SNARE and Co-factor Interactions: p115 interacts with numerous other proteins in the trafficking machinery. As mentioned, it binds directly to SNARE proteins such as the Golgi t-SNARE syntaxin 5 (STX5) and the vesicle v-SNARE GOS-28 (GOSR2) (reactome.org). These interactions occur via the coiled-coil domains (the CC1 and CC4 regions function as SNARE-binding motifs) (pubmed.ncbi.nlm.nih.gov). By binding SNAREs, p115 likely stabilizes them in a complex-ready state. Additionally, p115 has been found to associate with multi-subunit tethering complexes. For example, it can interact with subunits of the COG complex (another tethering complex involved in retrograde Golgi trafficking) and with Sec34/35. However, the most prominent partners remain the Golgi matrix “golgin” proteins (GM130, giantin) and the ER-Golgi SNAREs.
Emerging Interactions – Sec16A: A very recent finding (2023–2024) expanded p115’s interaction network to the ER exit sites. Sec16A is a large scaffolding protein that defines ER exit sites for COPII vesicle budding. New evidence suggests the head domain of p115 can bind directly to Sec16A (sciety.org). Specifically, researchers identified a conserved motif in the unstructured N-terminus of Sec16A that interacts with p115’s globular head (sciety.org). Mutations in p115 that disrupt Sec16-binding were shown to reduce secretory traffic efficiency, implying this interaction is functionally important (sciety.org). This discovery hints at p115 potentially linking ER vesicle formation with Golgi tethering in a spatially coordinated way. In other words, a subset of p115 molecules might “reach back” to ER exit sites via Sec16A, helping organize the early secretory pathway on a larger scale (sciety.org). While this idea is still under investigation (reported in a 2025 preprint), it underscores how p115 is centrally positioned in the secretory pathway, touching both vesicle biogenesis and vesicle docking steps.
Other Regulatory Proteins: p115’s function is also modulated by proteins like GRASP65/55 (Golgi stacking proteins) and other golgins. For instance, GRASP65 works with p115 in Golgi stack reformation; p115 binding to GM130 is more effective when GM130 is part of the larger GRASP65 complex (reactome.org). Additionally, calcium and calmodulin have been reported to bind p115 in some studies, suggesting calcium signaling might influence tethering events (though the physiological significance remains to be fully clarified).
In summary, p115 acts as a hub, interacting with GTPases (Rab1), long coiled-coil tether proteins (GM130, giantin), coat proteins at ER exit sites (Sec16A), and the SNARE fusion machinery. These interactions are tightly regulated by post-translational modifications and the cell cycle stage, ensuring that p115 tethers vesicles at the right time and place.
Role in Golgi Architecture and Cell Cycle Progression
Beyond vesicle trafficking per se, USO1/p115 has critical roles in maintaining Golgi structure and coordinating it with the cell cycle. The Golgi apparatus in mammalian cells is normally organized into a ribbon of interconnected stacks near the nucleus. This architecture depends on matrix proteins and tethering factors to hold the stacks together. p115 is essential for Golgi ribbon integrity – it links membranes both laterally and across cisternae. When p115 is depleted, the Golgi ribbon breaks into dispersed mini-stacks or vesicles (pmc.ncbi.nlm.nih.gov). Alvarez et al. (2001) first showed that p115 knockdown causes the loss of the perinuclear Golgi ribbon, phenocopying the effects of brefeldin A (a drug that collapses the Golgi) (pmc.ncbi.nlm.nih.gov). Mechanistically, the p115/GM130/giantin tether complex is thought to underpin the structural linkage of cis-Golgi elements (pmc.ncbi.nlm.nih.gov). Without p115, cisternal membranes cannot properly tether, leading to Golgi fragmentation and functional impairment of the secretory pathway.
During the cell cycle, the Golgi undergoes dramatic changes, and p115 is a key player in these events. In late G2 phase, the Golgi starts to “unlink” into discrete stacks, and then in mitosis it disassembles into vesicles and tubules (the “Golgi haze”). p115, together with GM130, is one of the targets of mitotic kinases that cause this disassembly. As noted above, phosphorylation of GM130 in mitosis prevents p115 from tethering cisternae together (reactome.org). Consistently, cells expressing a non-phosphorylatable GM130 mutant maintain Golgi tethering (and experience a delay in Golgi breakdown) until p115 is experimentally removed. Once cells exit mitosis, p115 is required for Golgi reassembly: it helps re-tether the Golgi membranes that were separated. Shorter and Warren (1999) demonstrated that adding p115 to mitotic Golgi fragments in vitro promoted their fusion into intact Golgi stacks (www.frontiersin.org). In living cells, p115 quickly relocalizes to Golgi membranes during telophase, concurrent with dephosphorylation of GM130, to reform the Golgi ribbon (www.frontiersin.org). Therefore, p115 orchestrates Golgi inheritance each division, ensuring daughter cells re-establish a functional Golgi.
Intriguingly, p115’s role extends to coordinating the Golgi with the microtubule cytoskeleton, which is particularly important in interphase cells. The Golgi apparatus can serve as an MTOC (microtubule-organizing center), especially in cells like fibroblasts where non-centrosomal microtubules emanate from the Golgi. p115 is a crucial factor in this process: it binds γ-tubulin via its N-terminus and recruits γ-tubulin ring complexes to the Golgi membrane in interphase (www.frontiersin.org). The interaction involves p115’s N-terminal armadillo-like domain attaching to γ-tubulin complexes that are associated with GM130 (www.frontiersin.org). Studies by Rivero and colleagues (e.g. 2009, 2011) showed that depletion of p115 or GM130 reduces the nucleation of microtubules at the Golgi, whereas overexpression of p115 can increase Golgi-based microtubule growth (www.frontiersin.org). This function links secretory trafficking with cell polarity and migration, since Golgi-originating microtubules help direct secretory vesicles toward the leading edge in motile cells. During mitosis, as noted, p115 itself localizes to spindle poles (the main MTOCs) through γ-tubulin binding (www.frontiersin.org). Although dispensable for spindle assembly, this localization may help ensure that, upon mitotic exit, p115 is well positioned to capture Golgi membranes and also possibly to assist in re-establishing microtubule arrays at the Golgi in G1. In summary, p115 integrates Golgi structure with microtubule organization, highlighting its broader structural role beyond vesicle tethering.
Biological Pathways Involving USO1/p115
Given its central role in trafficking, p115 is involved in several fundamental cellular pathways:
-
Constitutive Secretory Pathway: USO1 is a core component of the pathway delivering newly synthesized proteins from the ER → Golgi → secretory vesicles → cell surface or extracellular space. The Reactome database places p115 in the pathway steps “ER to Golgi transport” and “Golgi vesicle tethering” as a crucial factor for vesicle docking (reactome.org). Virtually all secreted or membrane proteins rely on p115-dependent steps during their journey through the Golgi.
-
Retrograde Traffic to ER: p115 also participates in the retrieval of escaped ER proteins and recycling of SNAREs from the Golgi back to the ER (often via COPI vesicles). It has been implicated in COPI-mediated retrograde transport, working alongside the COG complex to tether retrograde carriers at the ER or ERGIC. This ensures recycling of ER chaperones and maintenance of ER protein localization.
-
Golgi–to–Endosome Trafficking: While p115 is primarily associated with early secretory trafficking, some evidence suggests it may influence transport from the Golgi to endosomal compartments. High-throughput interaction and localization studies (e.g., global proteomics and Gene Ontology annotations) have linked USO1 to processes like Golgi to endosome transport and even autophagosome assembly (www.genome.jp). The connection to autophagy is not fully direct, but since autophagosomes can originate from ER–Golgi membranes, loss of p115 might indirectly impair the formation of these structures (for example, by disorganizing the membrane supply needed for autophagosome biogenesis). Indeed, cells lacking p115 show broad trafficking defects that can impact lysosomal delivery and autophagy flux (an area of ongoing research).
-
Signaling Pathways: By itself, p115 is not a signaling molecule, but proper secretory function is often required for signaling receptors to reach the cell surface. Interestingly, a few studies have noted that p115 can affect specific signaling cascades inside cells. For instance, in multiple myeloma cancer cells, overexpression of USO1 was found to enhance ERK (MAP kinase) pathway activity, possibly by increasing the secretion of autocrine growth factors or the cell-surface expression of signaling receptors (www.frontiersin.org). This ties p115 to signaling outcomes, albeit indirectly, through its effect on protein trafficking. In addition, the structural integrity of the Golgi (which p115 helps maintain) is known to modulate certain signaling pathways (such as mTOR and GSK3 signaling) that sense Golgi status; disruptions in p115 could therefore send aberrant signals that the cell interprets as stress.
In summary, USO1/p115 is a linchpin of the early secretory and Golgi trafficking pathways. Virtually any cellular process that depends on proper protein secretion or membrane transport (from neurotransmission to plasma membrane receptor recycling) can be affected by p115 dysfunction. The precise role of p115 in any specialized pathway (e.g. insulin secretion, immune cell antigen presentation) will be through its fundamental activity in vesicle tethering at the ER/Golgi interface.
Clinical and Research Significance
Because USO1 encodes an essential housekeeping protein, heritable mutations in USO1 are exceptionally rare or incompatible with life – consistent with the early embryonic lethality observed in knockout mice (pmc.ncbi.nlm.nih.gov). To date, there are no well-documented congenital human diseases caused by germline USO1 loss-of-function, likely because such mutations would arrest development. However, emerging research has linked alterations in USO1 expression to cancer biology and identified p115 as a potential vulnerability in certain tumors.
Several studies in the last decade found that USO1 is upregulated in multiple cancers, and that cancer cells can be unusually dependent on p115. For example, Jin and Dai (2016) reported that USO1 was overexpressed in multiple myeloma cells and that it promoted tumor progression by activating the ERK signaling pathway (www.frontiersin.org). Silencing USO1 in those myeloma cells led to reduced proliferation, indicating p115 might support the high secretory demands or signaling needs of cancer cells. Similarly, a 2015 study showed that knockdown of USO1 in colon cancer cells inhibited their proliferation and migration, suggesting a potential therapeutic angle in targeting the secretory pathway for cancer treatment (www.frontiersin.org). More recently, Jaiswal et al. (2021) conducted a focused CRISPR-Cas9 screen in B-cell acute lymphoblastic leukemia (B-ALL) and found USO1 to be a critical “druggable” vulnerability in this cancer subtype (www.frontiersin.org). Disrupting USO1 in B-ALL cells caused a significant reduction in cell growth and increased cell death (pmc.ncbi.nlm.nih.gov). The same study showed USO1 expression is directly upregulated by the oncogenic MLL-AF4 fusion protein in aggressive leukemias, explaining why those leukemia cells have heightened reliance on p115 (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Notably, USO1 was not identified as an essential gene in normal cells by broad CRISPR screens (e.g., in the DepMap database), yet it is indispensable in certain cancer cells (pmc.ncbi.nlm.nih.gov). This difference suggests that some cancers acquire a particular dependency on robust secretory function – a phenomenon termed “non-oncogene addiction.” Thus, p115 is being investigated as a possible anti-cancer target, where inhibitors of p115 function might selectively impact tumor cells that cannot compensate for its loss (pmc.ncbi.nlm.nih.gov). This is an active area of research, and any therapeutic strategy would have to carefully balance the essential role of p115 in normal cells.
Beyond cancer, the involvement of p115 in Golgi structure and vesicle trafficking makes it relevant to various diseases that feature Golgi dysfunction or protein misfolding. For instance, certain neurodegenerative diseases and skeletal dysplasias are linked to Golgi fragmentation phenotypes; while USO1 mutations haven’t been directly implicated, the pathways it governs are often disrupted in those conditions. Additionally, p115 has come onto the radar in virology: some viruses hijack the secretory pathway, and interestingly, a recent proteomic study found that the SARS-CoV-2 N protein can bind p115 (identified via an interaction screen) (www.reactome.org). The significance of this viral interaction is still unclear, but it raises the possibility that viruses might target p115 to modulate host vesicle traffic for their benefit.
From a research standpoint, p115 is also used as a marker and tool in cell biology. Antibodies against p115 are commonly used to stain the Golgi apparatus in imaging experiments (because p115 decorates the Golgi rims) (www.ptglab.com). Functionally, p115 and its yeast counterpart Uso1p have been invaluable in reconstitution experiments: for example, adding recombinant p115 to cell-free systems can restore ER-to-Golgi transport, confirming its sufficiency in tethering (reactome.org). The protein’s biochemical properties (a long flexible dimer with multiple binding sites) make it a model for studying how tethering factors and SNAREs cooperate, an area of intense research in membrane biology. Recent structural biology efforts (e.g., cryo-EM of tethering complexes) are beginning to elucidate how p115’s coiled-coil regions engage SNARE complexes at the molecular level, which could open avenues for drug design targeting these interactions.
Statistics and Data: By the numbers, USO1/p115 interacts with a large network of proteins. BioGRID and other databases report over 200 physical interaction partners for p115 in human cells (thebiogrid.org), reflecting its central networking role at the Golgi. USO1 gene expression data show it is rather uniformly expressed; it does not tend to be mutated frequently in populations (no common loss-of-function variants are noted in gnomAD, for instance, consistent with it being essential). On the cellular level, p115 molecules are abundantly present – estimated at a few hundred thousand copies per human cell – ensuring that numerous vesicles can be tethered simultaneously. When p115 is experimentally knocked down by ~90%, cells struggle to compensate: secretory cargo transport rates drop dramatically (often measured by delayed Golgi arrival of reporter proteins), and Golgi cisternae lose alignment (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). These quantitative observations underscore p115’s role as a workhorse of the early secretory pathway.
Conclusion
In conclusion, USO1 (gene) encodes p115 (protein), a master regulator of vesicle trafficking and Golgi organization in human cells. Functionally, p115 serves as a structural tether that docks transport vesicles to the Golgi, working in concert with Rab GTPases, golgin coiled-coil proteins (GM130, giantin), and SNAREs to ensure efficient membrane fusion (www.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). The primary role of p115 is not catalytic but architectural – it creates physical links that maintain the flow of cargo through the secretory pathway and preserve Golgi integrity. It carries out this role at the cytoplasmic face of the ER–Golgi interface, cycling on and off membranes depending on the phosphorylation state and cell cycle stage (www.ncbi.nlm.nih.gov). p115 is integral to ER-to-Golgi transport, intra-Golgi trafficking, and post-mitotic Golgi reassembly, with its loss causing severe disruption to these processes (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov).
Decades of research, from early biochemical assays to modern genetic screens, all converge on the view that USO1/p115 is essential for cell viability and secretory function. Recent studies (2020–2024) have expanded our understanding by revealing new interactions (e.g., with Sec16A at ER exit sites (sciety.org)) and highlighting p115’s importance in disease contexts (such as cancer cell survival (pmc.ncbi.nlm.nih.gov)). Going forward, p115 remains a subject of intense interest – both as a model for membrane tethering mechanisms and as a potential target to modulate secretion in diseases. In summary, USO1/p115’s broader structural role is to connect and coordinate key elements of the secretory pathway, ensuring that the journey of proteins from the ER to their final destination is smooth and orderly, thereby upholding the secretory and organizational infrastructure fundamental to eukaryotic cell life.
References: (Publication dates and sources provided where available)
- Alvarez, C. et al., 2001. p115 and Golgi structure – Knockdown studies showing Golgi fragmentation (pmc.ncbi.nlm.nih.gov).
- Allan, B.B. et al., 2000 (Science). Rab1 recruits p115 – Rab1-mediated loading of p115 on COPII vesicles (reactome.org) (www.ncbi.nlm.nih.gov).
- Nakamura, N. et al., 1997 (Cell). p115–GM130 binding – Mitotic phosphorylation disrupts p115/GM130 interaction (reactome.org).
- Shorter, J. et al., 2002 (J. Cell Biol). p115 and SNARE assembly – p115 sequentially tethers golgins and SNAREs (reactome.org).
- Kim, J. et al., 2012 (PLoS ONE). USO1 knockout mice – Early embryonic lethality and Golgi disruption (pmc.ncbi.nlm.nih.gov).
- Grabski, R. et al., 2012 (Bioarchitecture). Tether–SNARE model – p115’s coiled coils bind SNARE motifs (review).
- Toh, W.H. & Gleeson, P.A., 2016 (Front. Cell Dev. Biol.). Membrane tethers review – p115 overview as cis-Golgi tether (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov).
- Iannitti, R. et al., 2024 (Front. Cell Dev. Biol.). Golgi in cell division – p115’s role in Golgi inheritance and microtubule nucleation (www.frontiersin.org) (www.frontiersin.org).
- Jaiswal, A.K. et al., 2021 (Sci. Rep.). USO1 in leukemia – CRISPR screen identifying USO1 as a leukemia cell vulnerability (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov).
- Jin, Y. & Dai, Z., 2016 (Biomed. Pharmacother.). USO1 in myeloma – USO1 overexpression activating ERK, promoting myeloma growth (www.frontiersin.org).
- … [Additional references from gene databases and structural studies] …
Citations
- AnnotationURLCitation(end_index=382, start_index=225, title='iPTMnet Report O60763 USO1', type='url_citation', url='https://research.bioinformatics.udel.edu/iptmnet/entry/O60763/#:~:text=UniProt%20AC%20%2F%20UniProt%20ID,Human%29%20PRO')
- AnnotationURLCitation(end_index=626, start_index=478, title='Identification of a functional domain within the p115 tethering factor that is required for Golgi ribbon assembly and membrane trafficking - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/22328511/#:~:text=The%20tethering%20factor%20p115%20,terminal%20domain%20of%20p115%20as')
- AnnotationURLCitation(end_index=863, start_index=756, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=p115%2FUSO1%20%20,natal')
- AnnotationURLCitation(end_index=970, start_index=864, title='CDD Conserved Protein Domain Family: Uso1_p115_head', type='url_citation', url='https://www.ncbi.nlm.nih.gov/Structure/cdd/pfam04869#:~:text=protein%20,detected')
- AnnotationURLCitation(end_index=1316, start_index=1210, title='CDD Conserved Protein Domain Family: Uso1_p115_head', type='url_citation', url='https://www.ncbi.nlm.nih.gov/Structure/cdd/pfam04869#:~:text=protein%20,detected')
- AnnotationURLCitation(end_index=1652, start_index=1456, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=Expression%20Ubiquitous%20expression%20in%20thyroid,other%20tissues%20See%20more%20Orthologs')
- AnnotationURLCitation(end_index=1998, start_index=1909, title='USO1 Gene - GeneCards | USO1 Protein | USO1 Antibody', type='url_citation', url='https://www.genecards.org/cgi-bin/carddisp.pl?gene=USO1#:~:text=,2')
- AnnotationURLCitation(end_index=2356, start_index=2191, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=Golgi%20structure%20%28Alvarez%20et%20al,that%20USO1%2Fp115%20is%20critical%20for')
- AnnotationURLCitation(end_index=2947, start_index=2753, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=membrane%20and%20dissociates%20from%20the,Alternative%20splicing%20results%20in%20multiple')
- AnnotationURLCitation(end_index=3113, start_index=2948, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=Golgi%20structure%20%28Alvarez%20et%20al,that%20USO1%2Fp115%20is%20critical%20for')
- AnnotationURLCitation(end_index=3621, start_index=3459, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=The%20golgin%20Uso1p%2Fp115%20is%20a,caused%20Golgi%20disruption%20and%20early')
- AnnotationURLCitation(end_index=4360, start_index=4203, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=Linstedt%20et%20al,Whereas%20the')
- AnnotationURLCitation(end_index=4503, start_index=4361, title='CDD Conserved Protein Domain Family: Uso1_p115_head', type='url_citation', url='https://www.ncbi.nlm.nih.gov/Structure/cdd/pfam04869#:~:text=head%20region,like%20repeats%20%28pfam00514%29.%20Links')
- AnnotationURLCitation(end_index=4854, start_index=4688, title='CDD Conserved Protein Domain Family: Uso1_p115_head', type='url_citation', url='https://www.ncbi.nlm.nih.gov/Structure/cdd/pfam04869#:~:text=dimerization%2C%20and%20a%20short%20C,like%20repeats%20%28pfam00514%29.%20Links')
- AnnotationURLCitation(end_index=5201, start_index=5095, title='CDD Conserved Protein Domain Family: Uso1_p115_head', type='url_citation', url='https://www.ncbi.nlm.nih.gov/Structure/cdd/pfam04869#:~:text=protein%20,detected')
- AnnotationURLCitation(end_index=5663, start_index=5512, title='Identification of a functional domain within the p115 tethering factor that is required for Golgi ribbon assembly and membrane trafficking - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/22328511/#:~:text=deletion%20mutants%2C%20we%20show%20that,mediated%20membrane%20tethering')
- AnnotationURLCitation(end_index=5884, start_index=5778, title='CDD Conserved Protein Domain Family: Uso1_p115_head', type='url_citation', url='https://www.ncbi.nlm.nih.gov/Structure/cdd/pfam04869#:~:text=protein%20,detected')
- AnnotationURLCitation(end_index=6174, start_index=6068, title='CDD Conserved Protein Domain Family: Uso1_p115_head', type='url_citation', url='https://www.ncbi.nlm.nih.gov/Structure/cdd/pfam04869#:~:text=protein%20,detected')
- AnnotationURLCitation(end_index=6579, start_index=6388, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=Summary%20The%20protein%20encoded%20by,Alternative%20splicing%20results%20in%20multiple')
- AnnotationURLCitation(end_index=6937, start_index=6749, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=p115%20is%20a%20peripheral%20membrane,During%20the%20interphase')
- AnnotationURLCitation(end_index=7335, start_index=7152, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=recycles%20between%20the%20cytosol%20and,provided%20by%20RefSeq%2C%20Feb%202014')
- AnnotationURLCitation(end_index=7655, start_index=7467, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=p115%20is%20a%20peripheral%20membrane,During%20the%20interphase')
- AnnotationURLCitation(end_index=8089, start_index=7895, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=membrane%20and%20dissociates%20from%20the,Alternative%20splicing%20results%20in%20multiple')
- AnnotationURLCitation(end_index=8700, start_index=8542, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=and%20%CE%B3TuRC%20,Whereas%20the')
- AnnotationURLCitation(end_index=9023, start_index=8866, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=Linstedt%20et%20al,Whereas%20the')
- AnnotationURLCitation(end_index=9372, start_index=9215, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=Linstedt%20et%20al,Whereas%20the')
- AnnotationURLCitation(end_index=10338, start_index=10150, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=p115%20is%20a%20peripheral%20membrane,During%20the%20interphase')
- AnnotationURLCitation(end_index=10504, start_index=10339, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=Golgi%20structure%20%28Alvarez%20et%20al,that%20USO1%2Fp115%20is%20critical%20for')
- AnnotationURLCitation(end_index=11180, start_index=11015, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=Golgi%20structure%20%28Alvarez%20et%20al,that%20USO1%2Fp115%20is%20critical%20for')
- AnnotationURLCitation(end_index=11481, start_index=11316, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=Golgi%20structure%20%28Alvarez%20et%20al,that%20USO1%2Fp115%20is%20critical%20for')
- AnnotationURLCitation(end_index=11764, start_index=11599, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=Golgi%20structure%20%28Alvarez%20et%20al,that%20USO1%2Fp115%20is%20critical%20for')
- AnnotationURLCitation(end_index=12054, start_index=11889, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=Golgi%20structure%20%28Alvarez%20et%20al,that%20USO1%2Fp115%20is%20critical%20for')
- AnnotationURLCitation(end_index=12334, start_index=12228, title='CDD Conserved Protein Domain Family: Uso1_p115_head', type='url_citation', url='https://www.ncbi.nlm.nih.gov/Structure/cdd/pfam04869#:~:text=protein%20,detected')
- AnnotationURLCitation(end_index=12627, start_index=12521, title='CDD Conserved Protein Domain Family: Uso1_p115_head', type='url_citation', url='https://www.ncbi.nlm.nih.gov/Structure/cdd/pfam04869#:~:text=protein%20,detected')
- AnnotationURLCitation(end_index=12942, start_index=12777, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=Golgi%20structure%20%28Alvarez%20et%20al,that%20USO1%2Fp115%20is%20critical%20for')
- AnnotationURLCitation(end_index=13364, start_index=13170, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=membrane%20and%20dissociates%20from%20the,Alternative%20splicing%20results%20in%20multiple')
- AnnotationURLCitation(end_index=13719, start_index=13525, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=membrane%20and%20dissociates%20from%20the,Alternative%20splicing%20results%20in%20multiple')
- AnnotationURLCitation(end_index=13950, start_index=13855, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=10903204%20%20,2000')
- AnnotationURLCitation(end_index=14145, start_index=13951, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=membrane%20and%20dissociates%20from%20the,Alternative%20splicing%20results%20in%20multiple')
- AnnotationURLCitation(end_index=14376, start_index=14281, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=11285137%20%20,2002')
- AnnotationURLCitation(end_index=14702, start_index=14554, title='Identification of a functional domain within the p115 tethering factor that is required for Golgi ribbon assembly and membrane trafficking - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/22328511/#:~:text=The%20tethering%20factor%20p115%20,terminal%20domain%20of%20p115%20as')
- AnnotationURLCitation(end_index=15058, start_index=14910, title='Identification of a functional domain within the p115 tethering factor that is required for Golgi ribbon assembly and membrane trafficking - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/22328511/#:~:text=The%20tethering%20factor%20p115%20,terminal%20domain%20of%20p115%20as')
- AnnotationURLCitation(end_index=15494, start_index=15343, title='Identification of a functional domain within the p115 tethering factor that is required for Golgi ribbon assembly and membrane trafficking - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/22328511/#:~:text=deletion%20mutants%2C%20we%20show%20that,mediated%20membrane%20tethering')
- AnnotationURLCitation(end_index=15735, start_index=15646, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=WE%20%20,2002')
- AnnotationURLCitation(end_index=15939, start_index=15788, title='Identification of a functional domain within the p115 tethering factor that is required for Golgi ribbon assembly and membrane trafficking - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/22328511/#:~:text=deletion%20mutants%2C%20we%20show%20that,mediated%20membrane%20tethering')
- AnnotationURLCitation(end_index=16337, start_index=16143, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=membrane%20and%20dissociates%20from%20the,Alternative%20splicing%20results%20in%20multiple')
- AnnotationURLCitation(end_index=16664, start_index=16575, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=WE%20%20,2002')
- AnnotationURLCitation(end_index=17382, start_index=17194, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=p115%20is%20a%20peripheral%20membrane,During%20the%20interphase')
- AnnotationURLCitation(end_index=17708, start_index=17587, title='Identification of a functional domain within the p115 tethering factor that is required for Golgi ribbon assembly and membrane trafficking - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/22328511/#:~:text=H2%29%20within%20the%20N,Shorter%20et%20al')
- AnnotationURLCitation(end_index=18180, start_index=18026, title='CDD Conserved Protein Domain Family: Uso1_p115_head', type='url_citation', url='https://www.ncbi.nlm.nih.gov/Structure/cdd/pfam04869#:~:text=dimerization%2C%20and%20a%20short%20C,like%20repeats%20%28pfam00514')
- AnnotationURLCitation(end_index=18449, start_index=18295, title='CDD Conserved Protein Domain Family: Uso1_p115_head', type='url_citation', url='https://www.ncbi.nlm.nih.gov/Structure/cdd/pfam04869#:~:text=dimerization%2C%20and%20a%20short%20C,like%20repeats%20%28pfam00514')
- AnnotationURLCitation(end_index=18829, start_index=18667, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=The%20golgin%20Uso1p%2Fp115%20is%20a,caused%20Golgi%20disruption%20and%20early')
- AnnotationURLCitation(end_index=19183, start_index=18995, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=p115%20is%20a%20peripheral%20membrane,During%20the%20interphase')
- AnnotationURLCitation(end_index=19594, start_index=19459, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=maintaining%20Golgi%20architecture%20as%20well,2012')
- AnnotationURLCitation(end_index=19702, start_index=19595, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=p115%2FUSO1%20%20,natal')
- AnnotationURLCitation(end_index=19985, start_index=19850, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=maintaining%20Golgi%20architecture%20as%20well,2012')
- AnnotationURLCitation(end_index=20276, start_index=20114, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=The%20golgin%20Uso1p%2Fp115%20is%20a,caused%20Golgi%20disruption%20and%20early')
- AnnotationURLCitation(end_index=20925, start_index=20731, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=membrane%20and%20dissociates%20from%20the,Alternative%20splicing%20results%20in%20multiple')
- AnnotationURLCitation(end_index=21236, start_index=21110, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=RAB1%3AGTP%20binds%20USO1%20and%20GORASP1%3AGOLGA2')
- AnnotationURLCitation(end_index=21332, start_index=21237, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=11285137%20%20,2002')
- AnnotationURLCitation(end_index=21609, start_index=21514, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=11285137%20%20,2002')
- AnnotationURLCitation(end_index=21985, start_index=21890, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=11285137%20%20,2002')
- AnnotationURLCitation(end_index=22463, start_index=22280, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=recycles%20between%20the%20cytosol%20and,provided%20by%20RefSeq%2C%20Feb%202014')
- AnnotationURLCitation(end_index=22728, start_index=22639, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=WE%20%20,2002')
- AnnotationURLCitation(end_index=23146, start_index=22977, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=reassembly%20after%20mitosis%20,tubulin%20to')
- AnnotationURLCitation(end_index=23427, start_index=23258, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=reassembly%20after%20mitosis%20,tubulin%20to')
- AnnotationURLCitation(end_index=23690, start_index=23521, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=reassembly%20after%20mitosis%20,tubulin%20to')
- AnnotationURLCitation(end_index=24244, start_index=24155, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=WE%20%20,2002')
- AnnotationURLCitation(end_index=24509, start_index=24358, title='Identification of a functional domain within the p115 tethering factor that is required for Golgi ribbon assembly and membrane trafficking - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/22328511/#:~:text=deletion%20mutants%2C%20we%20show%20that,mediated%20membrane%20tethering')
- AnnotationURLCitation(end_index=25419, start_index=25250, title='The Golgi vesicle tether p115/USO1 can bind directly to the ER exit site organiser Sec16A | Sciety', type='url_citation', url='https://sciety.org/articles/activity/10.1101/2025.10.16.682774#:~:text=proteins%20to%20promote%20membrane%20fusion,of%20the%20early%20secretory%20pathway')
- AnnotationURLCitation(end_index=25727, start_index=25558, title='The Golgi vesicle tether p115/USO1 can bind directly to the ER exit site organiser Sec16A | Sciety', type='url_citation', url='https://sciety.org/articles/activity/10.1101/2025.10.16.682774#:~:text=proteins%20to%20promote%20membrane%20fusion,of%20the%20early%20secretory%20pathway')
- AnnotationURLCitation(end_index=26042, start_index=25877, title='The Golgi vesicle tether p115/USO1 can bind directly to the ER exit site organiser Sec16A | Sciety', type='url_citation', url='https://sciety.org/articles/activity/10.1101/2025.10.16.682774#:~:text=organises%20ER%20sites%20and%20promotes,of%20the%20early%20secretory%20pathway')
- AnnotationURLCitation(end_index=26473, start_index=26325, title='The Golgi vesicle tether p115/USO1 can bind directly to the ER exit site organiser Sec16A | Sciety', type='url_citation', url='https://sciety.org/articles/activity/10.1101/2025.10.16.682774#:~:text=the%20unstructured%20N,of%20the%20early%20secretory%20pathway')
- AnnotationURLCitation(end_index=27109, start_index=26983, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=RAB1%3AGTP%20binds%20USO1%20and%20GORASP1%3AGOLGA2')
- AnnotationURLCitation(end_index=28457, start_index=28295, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=The%20golgin%20Uso1p%2Fp115%20is%20a,caused%20Golgi%20disruption%20and%20early')
- AnnotationURLCitation(end_index=28799, start_index=28637, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=The%20golgin%20Uso1p%2Fp115%20is%20a,caused%20Golgi%20disruption%20and%20early')
- AnnotationURLCitation(end_index=29089, start_index=28924, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=Golgi%20structure%20%28Alvarez%20et%20al,that%20USO1%2Fp115%20is%20critical%20for')
- AnnotationURLCitation(end_index=29777, start_index=29688, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=WE%20%20,2002')
- AnnotationURLCitation(end_index=30427, start_index=30220, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=p115%20is%20a%20peripheral%20membrane,tubulin%2C%20a%20component%20of%20%CE%B3TuSC')
- AnnotationURLCitation(end_index=30786, start_index=30579, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=p115%20is%20a%20peripheral%20membrane,tubulin%2C%20a%20component%20of%20%CE%B3TuSC')
- AnnotationURLCitation(end_index=31549, start_index=31392, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=Linstedt%20et%20al,Whereas%20the')
- AnnotationURLCitation(end_index=31837, start_index=31680, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=Linstedt%20et%20al,Whereas%20the')
- AnnotationURLCitation(end_index=32214, start_index=32057, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=Linstedt%20et%20al,Whereas%20the')
- AnnotationURLCitation(end_index=32667, start_index=32509, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=and%20%CE%B3TuRC%20,Whereas%20the')
- AnnotationURLCitation(end_index=33647, start_index=33552, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=11285137%20%20,2002')
- AnnotationURLCitation(end_index=34758, start_index=34586, title='UniProt: O00189', type='url_citation', url='https://www.genome.jp/dbget-bin/www_bget?uniprot%3AO00189=#:~:text=...GO%3A0005802%3B%20C%3Atrans,GO%3A0006895%3B%20P%3AGolgi%20to%20endosome%20transport')
- AnnotationURLCitation(end_index=35857, start_index=35701, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2024.1513472/full#:~:text=Jin%2C%20Y,012')
- AnnotationURLCitation(end_index=37115, start_index=36980, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=maintaining%20Golgi%20architecture%20as%20well,2012')
- AnnotationURLCitation(end_index=37922, start_index=37766, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2024.1513472/full#:~:text=Jin%2C%20Y,012')
- AnnotationURLCitation(end_index=38482, start_index=38307, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2024.1513472/full#:~:text=match%20at%20L2517%20Lentivirus,z')
- AnnotationURLCitation(end_index=38877, start_index=38692, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2024.1513472/full#:~:text=et%20al.%20%282021%29.%20Focused%20CRISPR,w')
- AnnotationURLCitation(end_index=39159, start_index=38981, title='Focused CRISPR-Cas9 genetic screening reveals USO1 as a vulnerability in B-cell acute lymphoblastic leukemia - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC8222245/#:~:text=dependencies%20on%20several%20genes%20including,cells%20revealed%20alterations%20in%20pathways')
- AnnotationURLCitation(end_index=39482, start_index=39357, title='Focused CRISPR-Cas9 genetic screening reveals USO1 as a vulnerability in B-cell acute lymphoblastic leukemia - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC8222245/#:~:text=USO1%20is%20directly%20regulated%20by,AF4')
- AnnotationURLCitation(end_index=39645, start_index=39483, title='Focused CRISPR-Cas9 genetic screening reveals USO1 as a vulnerability in B-cell acute lymphoblastic leukemia - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC8222245/#:~:text=USO1%20depletion%20alters%20B,proliferation%2C%20survival%20and%20cell%20cycle')
- AnnotationURLCitation(end_index=39990, start_index=39821, title='Focused CRISPR-Cas9 genetic screening reveals USO1 as a vulnerability in B-cell acute lymphoblastic leukemia - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC8222245/#:~:text=match%20at%20L174%20be%20%E2%80%9Ccommon,designing%20primers%20for%20the%20regulatory')
- AnnotationURLCitation(end_index=40500, start_index=40322, title='Focused CRISPR-Cas9 genetic screening reveals USO1 as a vulnerability in B-cell acute lymphoblastic leukemia - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC8222245/#:~:text=dependencies%20on%20several%20genes%20including,cells%20revealed%20alterations%20in%20pathways')
- AnnotationURLCitation(end_index=41394, start_index=41293, title='Reactome | UniProt:O60763 USO1', type='url_citation', url='https://www.reactome.org/content/detail/interactor/O60763#:~:text=P0DTC9%20%20,')
- AnnotationURLCitation(end_index=41994, start_index=41793, title='p115, USO1 antibody (13509-1-AP) | Proteintech', type='url_citation', url='https://www.ptglab.com/products/USO1-Antibody-13509-1-AP.htm?srsltid=AfmBOooewwjC3o_zwvjzzrk-gqvUMZJ68dxn4l2-Onhazz9e1KYDry0T#:~:text=...,p115%2C%20USO1%20transfected%20HeLa%20cells')
- AnnotationURLCitation(end_index=42351, start_index=42232, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=through%20the%20Golgi%20apparatus%20in,2000')
- AnnotationURLCitation(end_index=43149, start_index=43011, title='USO1 Result Summary | BioGRID', type='url_citation', url='https://thebiogrid.org/114173/summary/homo-sapiens#:~:text=USO1%20Result%20Summary%20,com%2C%20All%20Rights%20Reserved')
- AnnotationURLCitation(end_index=44029, start_index=43851, title='Focused CRISPR-Cas9 genetic screening reveals USO1 as a vulnerability in B-cell acute lymphoblastic leukemia - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC8222245/#:~:text=dependencies%20on%20several%20genes%20including,cells%20revealed%20alterations%20in%20pathways')
- AnnotationURLCitation(end_index=44192, start_index=44030, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=The%20golgin%20Uso1p%2Fp115%20is%20a,caused%20Golgi%20disruption%20and%20early')
- AnnotationURLCitation(end_index=44879, start_index=44685, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=membrane%20and%20dissociates%20from%20the,Alternative%20splicing%20results%20in%20multiple')
- AnnotationURLCitation(end_index=45045, start_index=44880, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=Golgi%20structure%20%28Alvarez%20et%20al,that%20USO1%2Fp115%20is%20critical%20for')
- AnnotationURLCitation(end_index=45591, start_index=45400, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=Summary%20The%20protein%20encoded%20by,Alternative%20splicing%20results%20in%20multiple')
- AnnotationURLCitation(end_index=45921, start_index=45759, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=The%20golgin%20Uso1p%2Fp115%20is%20a,caused%20Golgi%20disruption%20and%20early')
- AnnotationURLCitation(end_index=46057, start_index=45922, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=maintaining%20Golgi%20architecture%20as%20well,2012')
- AnnotationURLCitation(end_index=46533, start_index=46364, title='The Golgi vesicle tether p115/USO1 can bind directly to the ER exit site organiser Sec16A | Sciety', type='url_citation', url='https://sciety.org/articles/activity/10.1101/2025.10.16.682774#:~:text=proteins%20to%20promote%20membrane%20fusion,of%20the%20early%20secretory%20pathway')
- AnnotationURLCitation(end_index=46798, start_index=46620, title='Focused CRISPR-Cas9 genetic screening reveals USO1 as a vulnerability in B-cell acute lymphoblastic leukemia - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC8222245/#:~:text=dependencies%20on%20several%20genes%20including,cells%20revealed%20alterations%20in%20pathways')
- AnnotationURLCitation(end_index=47642, start_index=47480, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=The%20golgin%20Uso1p%2Fp115%20is%20a,caused%20Golgi%20disruption%20and%20early')
- AnnotationURLCitation(end_index=47878, start_index=47759, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=through%20the%20Golgi%20apparatus%20in,2000')
- AnnotationURLCitation(end_index=48073, start_index=47879, title='USO1 USO1 vesicle transport factor [Homo sapiens (human)] - Gene - NCBI', type='url_citation', url='https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=8615#:~:text=membrane%20and%20dissociates%20from%20the,Alternative%20splicing%20results%20in%20multiple')
- AnnotationURLCitation(end_index=48285, start_index=48196, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=WE%20%20,2002')
- AnnotationURLCitation(end_index=48498, start_index=48409, title='Reactome | RAB1:GTP binds USO1 and GORASP1:GOLGA2', type='url_citation', url='https://reactome.org/content/detail/R-HSA-5694418#:~:text=WE%20%20,2002')
- AnnotationURLCitation(end_index=48746, start_index=48611, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=maintaining%20Golgi%20architecture%20as%20well,2012')
- AnnotationURLCitation(end_index=49159, start_index=48997, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=The%20golgin%20Uso1p%2Fp115%20is%20a,caused%20Golgi%20disruption%20and%20early')
- AnnotationURLCitation(end_index=49325, start_index=49160, title='Emerging Insights into the Roles of Membrane Tethers from Analysis of Whole Organisms: The Tip of an Iceberg? - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC4770024/#:~:text=Golgi%20structure%20%28Alvarez%20et%20al,that%20USO1%2Fp115%20is%20critical%20for')
- AnnotationURLCitation(end_index=49681, start_index=49474, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=p115%20is%20a%20peripheral%20membrane,tubulin%2C%20a%20component%20of%20%CE%B3TuSC')
- AnnotationURLCitation(end_index=49839, start_index=49682, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fcell.2024.1513472/full#:~:text=Linstedt%20et%20al,Whereas%20the')
- AnnotationURLCitation(end_index=50152, start_index=49974, title='Focused CRISPR-Cas9 genetic screening reveals USO1 as a vulnerability in B-cell acute lymphoblastic leukemia - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC8222245/#:~:text=dependencies%20on%20several%20genes%20including,cells%20revealed%20alterations%20in%20pathways')
- AnnotationURLCitation(end_index=50315, start_index=50153, title='Focused CRISPR-Cas9 genetic screening reveals USO1 as a vulnerability in B-cell acute lymphoblastic leukemia - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC8222245/#:~:text=USO1%20depletion%20alters%20B,proliferation%2C%20survival%20and%20cell%20cycle')
- AnnotationURLCitation(end_index=50609, start_index=50453, title='Frontiers | The role of Golgi complex proteins in cell division and consequences of their dysregulation', type='url_citation', url='https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2024.1513472/full#:~:text=Jin%2C%20Y,012')
Perplexity
(USO1-deep-research-perplexity-lite.md)
Perplexity
(USO1-deep-research-perplexity-lite.md)The USO1 gene (UniProt: O60763) in humans encodes the general vesicular transport factor p115, a key peripheral membrane protein involved in vesicular trafficking between the endoplasmic reticulum (ER) and the Golgi apparatus[1][3][11].
1. Key Concepts and Definitions
-
Molecular Function:
USO1 functions as a vesicular transport factor required for the intercisternal transport within the Golgi stack and for the fusion and/or binding of vesicles to target membranes[3][11]. It acts as a vesicular anchor, holding vesicular and target membranes in proximity to facilitate membrane fusion[11]. -
Biological Processes:
- ER-to-Golgi vesicle-mediated transport: USO1 is essential for the transport of vesicles from the ER to the cis/medial Golgi compartments[1][3][11].
- Golgi organization and maintenance: It is involved in maintaining Golgi structure and function, particularly during interphase[1][3].
- Cell cycle regulation: USO1 cycles between the cytosol and Golgi, regulated by phosphorylation status[1][3][5].
-
Protein trafficking and secretion: By facilitating vesicle docking and fusion, USO1 is critical for the secretory pathway[1][3][11].
-
Cellular Localization:
USO1 is a peripheral membrane protein that recycles between the cytosol and the Golgi apparatus during interphase[1][3][5][7][11]. Its association with the Golgi membrane is regulated by phosphorylation: dephosphorylated USO1 binds the Golgi, while phosphorylation causes dissociation[1][3][5].
2. Protein Domains
- Coiled-coil domains:
USO1 contains extensive coiled-coil regions that mediate its dimerization and interactions with other vesicular transport proteins[11]. - N-terminal and C-terminal domains:
The N-terminal region is involved in membrane binding, while the C-terminal region interacts with other tethering factors and SNARE proteins[11].
3. Known Interactions
- COPII vesicles:
USO1 is recruited to COPII vesicles budding from the ER by Ras-associated protein 1 (RAP1)[1][3][5]. - SNARE proteins:
It interacts with COPII vesicle-associated SNAREs to form a cis-SNARE complex, promoting vesicle targeting and fusion with the Golgi[1][3][11]. - GOLGA2 (GM130) and Giantin:
USO1 acts sequentially with these Golgi matrix proteins to mediate vesicle tethering and fusion[1][3]. - Other vesicle tethering factors:
USO1 is part of a larger network of proteins involved in vesicle docking and fusion, including Rab GTPases and other tethering complexes[11].
4. Disease Associations
- Retinal Disease:
Mutations or dysregulation of USO1 have been associated with certain retinal diseases[3]. - Hypogonadotropic Hypogonadism 14 With Or Without Anosmia:
USO1 is listed as associated with this rare endocrine disorder[3]. - Cancer:
Recent studies suggest that USO1 isoforms differentially promote liver cancer progression by dysregulating the ER-Golgi network[12]. - Multiple Myeloma:
USO1 may be a promising target for therapy and a diagnostic marker in multiple myeloma[9].
5. Recent Developments and Latest Research (2023–2024)
- Cancer Progression:
New research (2023–2024) highlights the role of USO1 isoforms in liver cancer progression through effects on the ER-Golgi network, suggesting a potential target for cancer therapy[12]. - Proteostasis and Secretory Pathways:
The importance of Golgi and ER trafficking factors like USO1 in maintaining proteostasis, especially in secretory cells (e.g., pancreatic β-cells), is increasingly recognized in the context of diseases such as diabetes[2].
6. Expert Opinions and Analysis
- Essential for Secretory Pathway:
USO1 is considered indispensable for the early secretory pathway, particularly for the tethering and fusion of ER-derived vesicles with the Golgi[1][3][11]. - Regulation by Phosphorylation:
The dynamic association of USO1 with Golgi membranes, controlled by phosphorylation, is a key regulatory mechanism for vesicle trafficking[1][3][5].
7. Relevant Statistics and Data
- Gene Location:
USO1 is located on chromosome 4q21.1[3]. - Protein Size:
The human USO1 protein (p115) is approximately 962 amino acids in length[11]. - Expression:
USO1 is ubiquitously expressed, with higher levels in secretory tissues[3][11].
8. Summary Table
| Feature | Details |
|---|---|
| Gene Symbol | USO1 |
| UniProt ID | O60763 |
| Protein Name | General vesicular transport factor p115 |
| Molecular Function | Vesicle tethering, membrane fusion, SNARE complex assembly |
| Biological Process | ER-to-Golgi transport, Golgi organization, protein secretion |
| Cellular Location | Cytosol, Golgi apparatus (peripheral membrane) |
| Protein Domains | Coiled-coil regions, N- and C-terminal functional domains |
| Key Interactions | COPII vesicles, SNAREs, GOLGA2 (GM130), Giantin, RAP1 |
| Disease Links | Retinal disease, hypogonadotropic hypogonadism, liver cancer, myeloma |
| Recent Research | Role in cancer progression, proteostasis, secretory pathway regulation |
Citations:
[1] Wikipedia (2024-06)
[3] GeneCards (2024-06)
[5] Promega (2024-06)
[7] MARRVEL (2024-06)
[9] GlyGen (2024-06)
[11] UniProt (2024-06)
[12] NCBI Gene (2024-06)
[2] Frontiers in Cell and Developmental Biology (2025-03)
Citations
- https://en.wikipedia.org/wiki/USO1
- https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2025.1502469/pdf
- https://www.genecards.org/cgi-bin/carddisp.pl?gene=USO1
- https://glygen.org/protein/P60763
- https://www.promega.de/findmygene/genedetail.aspx?ncbiid=8615
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3037419/
- https://marrvel.org/human/gene/8615
- https://www.proteomicsdb.org/proteomicsdb/
- https://glygen.org/protein/O60763
- https://functionome.geneontology.org/gene/UniProtKB:O60763
- https://www.uniprot.org/uniprotkb/O60763/entry
- https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=8615
Perplexity
(USO1-deep-research-perplexity.md)
Perplexity
(USO1-deep-research-perplexity.md)The user is asking for a comprehensive research report on the USO1 gene (O60763) in humans. They want:
- Detailed narrative explaining:
- Function
- Biological processes
-
Localization
-
Primary function:
- For this protein: it's a tether protein involved in vesicular transport
-
What does it actually do mechanistically?
-
Cellular localization
-
Signaling/biochemical pathways
-
Prioritize evidence-based research over broad pleiotropic effects
Let me plan my approach:
I need to review the search results provided and organize information on USO1/p115. From the search results, I can see:
- Basic definition: USO1 encodes "General vesicular transport factor p115" - a peripheral membrane protein that recycles between cytosol and Golgi
- It's regulated by phosphorylation
- It's recruited by Rab1 to COPII vesicles
- It interacts with SNAREs
- Recent findings (from eLife 2023/2024) suggest its essential function is NOT tethering but rather SNARE complex assembly
- It has a globular head domain (GHD), coiled-coil domain (CC), and C-terminal region (CTR)
- It's a dimer with two globular heads and a long coiled-coil tail
- It's involved in ER-to-Golgi transport
Key structural findings:
- The N-terminal globular domain has a novel alpha-solenoid architecture with 12 armadillo-like tether repeat motifs
- It forms a homodimer
- Crystal structure available (PDB: 3GQ2, 3GRL)
Key functional findings:
- Recent evidence from eLife (2024) suggests the essential role is SNARE regulation, not tethering
- It interacts with monomeric SNAREs, not SNARE complexes
- It has multiple functional domains with distinct roles, each contributing to its complex molecular mechanism
I'll develop a structured approach to analyze this protein's multifaceted functionality, focusing on its intricate domain architecture and specialized cellular roles.
Comprehensive Functional Annotation of the USO1 Gene: A Molecular Analysis of the General Vesicular Transport Factor p115
This report provides a detailed examination of the USO1 gene, which encodes the general vesicular transport factor p115, a critical peripheral membrane protein essential for membrane trafficking between the endoplasmic reticulum and Golgi apparatus in human cells. Recent structural and genetic studies have substantially revised our understanding of p115's mechanism of action, revealing that its essential cellular function involves the regulation of SNARE complex assembly rather than long-range vesicle tethering as previously believed. The protein functions as a homodimeric tether containing a globular N-terminal head domain with a novel alpha-solenoid architecture, an extended coiled-coil region mediating dimerization, and a C-terminal region for Golgi protein interactions. Through recruitment by the small GTPase Rab1, p115 localizes to ER-derived COPII vesicles and early Golgi cisternae, where it specifically interacts with monomeric SNARE proteins to facilitate the formation of productive trans-SNARE complexes required for membrane fusion. This functional role places p115 as an integral component of the SNARE fusion machinery rather than a purely structural tethering element, highlighting the dynamic and multifaceted nature of membrane trafficking regulation in eukaryotic cells.
Gene Identification and Basic Characterization
Genomic Location and Protein Nomenclature
The USO1 gene is located on human chromosome 4 and encodes the general vesicular transport factor p115, also known as p115 or Uso1p in its yeast homologue designation[1][20]. The gene is officially designated as ENSG00000138768 in the Ensembl database and carries the UniProtKB identifier O60763[2][27]. The human USO1 gene is protein-coding and generates multiple transcript variants through alternative splicing[27][48]. The nomenclature reflects the protein's evolutionary conservation, with the fungal yeast homologue Uso1p sharing substantial sequence identity and functional conservation with the mammalian p115 protein, indicative of the critical nature of this protein in eukaryotic cell biology[10][29].
Genomic Organization and Expression
USO1 is transcribed from its genomic locus with the resulting protein representing approximately 961 amino acids in the full-length human form[1]. The protein is ubiquitously expressed across tissues, with particularly high expression levels documented in tissues with substantial secretory activity[41]. The expression pattern indicates that USO1 function is required across diverse cell types to maintain the fundamental process of vesicular transport, which is essential for all eukaryotic cells[27]. The constitutive expression of USO1 across tissue types underscores its role as a housekeeping gene whose product is indispensable for basic cellular survival and function.
Structural Organization and Domain Architecture
Overall Protein Organization
The USO1 protein exhibits a highly organized modular architecture consisting of three distinct functional domains that work in concert to mediate its role in membrane trafficking[3][33][43]. The protein functions as a homodimer with two identical subunits forming an elongated structure approximately 154.5 nanometers in length, as determined through electron microscopic examination[13]. This dimeric organization is mediated by parallel association of the C-terminal coiled-coil tails, resulting in an elongated structure with two globular head domains positioned side by side and a long rod-like tail extending from the dimer interface[17]. The architecture resembles that of myosin II proteins, with the N-terminal regions providing functional interaction sites and the C-terminal regions providing structural scaffolding through their extended helical arrangements.
The Globular Head Domain
The N-terminal globular head domain (GHD) represents the most extensively characterized structural element of USO1, and recent crystallographic analysis has revealed its novel organizational principles[49][52]. The crystal structure of the p115 N-terminal domain at 2.0 angstrom resolution demonstrates that this region forms a novel alpha-solenoid architecture constructed of 12 tandem, triple-helical repeats arranged as armadillo-like tether-repeat (TR) motifs[18][49]. Each tether repeat consists of three alpha helices that form a helical tripod structure, with these repeats arranged in a right-handed superhelix with approximately 30-degree rotations between adjacent repeats, resulting in an extended hydrophobic core that stabilizes the entire structure[49]. The conserved structural fold among the 12 tether repeats suggests functional significance, as the H1 and H2 repeat motifs show evolutionary conservation and are predicted to have functionally distinct roles in docking and fusion processes[49].
The globular head domain represents approximately 70 kilodaltons of molecular mass and contains the Rab1 binding site within its H1 tether repeat motif[18][37][52]. This binding site is critical for recruiting USO1 to transport vesicles and target membranes. Mutation of the H1 motif results in dominant negative inhibition of endoplasmic reticulum to Golgi trafficking, demonstrating the essential nature of this interaction[18][52]. Importantly, recent biochemical characterization has shown that the isolated globular head domain, despite lacking the extended coiled-coil region typically associated with tethering proteins, remains monomeric and retains the ability to provide essential USO1 functions, including SNARE complex regulation[3][16][33]. This finding fundamentally challenges the classical model of USO1 function and indicates that long-range tethering mediated by the extended coiled-coil is dispensable for cell viability.
The Coiled-Coil Domain
The middle region of USO1 consists of four sequential coiled-coil domains (CC1, CC2, CC3, and CC4) that extend approximately 300 amino acids and serve multiple functional roles[1][6]. The CC1 domain is particularly important as it contains the primary binding site for the Rab1 GTPase, with both the globular head domain and the CC1 region contributing to stable Rab1 interaction[37][40]. The coiled-coil region mediates the dimerization of USO1, with analytical ultracentrifugation studies confirming that the coiled-coil domain functions as a homodimer while the globular head domain exists as a monomer when isolated[33]. The extended coiled-coil structure was predicted from primary sequence analysis to consist of long alpha-helical regions with some interruptions, which is confirmed by electron microscopic visualization showing discrete hinge points at approximately 23.1 and 85.5 nanometers from the globular domain[13].
The coiled-coil domains also contain SNARE-binding motifs, particularly within CC1 and CC4, that directly interact with ER-Golgi SNARE proteins[6]. These binding sites are distinct from and independent of the Rab1 binding site, allowing USO1 to simultaneously interact with both the GTPase regulatory machinery and the fusion machinery. The CC4 domain is particularly critical for p115 function, as deletion of this region results in dominant-negative effects that disrupt Golgi structure and prevent cargo trafficking even in the presence of endogenous p115[6]. Functional analyses in mammalian cells have demonstrated that CC4 is necessary for Golgi ribbon formation and membrane trafficking, suggesting this domain provides distinct and essential functions separate from SNARE binding in the CC1 region.
The C-Terminal Region
The C-terminal region (CTR) of USO1 extends beyond the fourth coiled-coil domain and contains an acidic motif that serves as a binding site for Golgi-associated proteins[20][30][33]. This region interacts with the Golgi matrix proteins GM130 and giantin, linking USO1 to the Golgi structural framework[20][21][24]. Analytical ultracentrifugation experiments have demonstrated that the CTR is not involved in dimerization, as truncated proteins lacking this region maintain their dimeric state[33]. The CTR also contains regulatory sequences that may modulate the access of other proteins to upstream functional domains. Previous models suggested that the CTR might interact with the globular head domain to maintain a closed conformation that regulates Rab1 binding, but recent biochemical analysis has refuted this model, demonstrating that deletion of the CTR does not result in significant conformational changes in sedimentation coefficients[33].
Molecular Function and Mechanism of Action
Historical Context and Paradigm Shift
For approximately three decades following its initial characterization in yeast, USO1/p115 was considered primarily a vesicle tethering protein whose essential function involved bringing transport vesicles into close proximity with target membranes through its extended coiled-coil region[1][6][23]. This model was supported by the protein's structural resemblance to other known tethering factors and its ability to mediate COPI vesicle tethering to Golgi membranes in biochemical in vitro assays[1][6]. However, accumulating genetic evidence consistently suggested that USO1 possessed additional functions related to SNARE complex assembly, with landmark studies demonstrating that overexpression of Ypt1 (yeast Rab1) could suppress the lethality of uso1 deletion strains, whereas the reciprocal was not true, indicating that USO1 functions upstream of or in conjunction with RAB1[10][16][29]. Additionally, suppressors of uso1 mutations identified through genetic screens consistently mapped to SNARE genes, suggesting direct involvement of USO1 in SNARE complex formation rather than purely physical vesicle positioning[10].
Recent pioneering work published in eLife in 2024 has fundamentally revised this understanding through a comprehensive genetic and biochemical analysis employing forward genetic screens in Aspergillus nidulans[3][16][43]. By isolating uso1 missense mutations that restore cell viability in the absence of rab1 function, researchers demonstrated that mutations in the globular head domain (specifically E6K and G540S substitutions) enhance binding to the cytosolic region of the Qa SNARE protein Bos1[3][16][43]. Critically, these studies showed that a protein consisting solely of the double mutant globular head domain, or even overexpression of wild-type globular head domain alone, could rescue the lethality resulting from complete uso1 deletion, despite the isolated globular head domain being monomeric and lacking the extended coiled-coil region typically associated with long-range tethering[3][16][33]. These findings provide compelling evidence that the essential role of USO1 is not tethering membranes but rather regulating the formation of cognate SNARE bundles, fundamentally placing USO1 as a component of the SNARE fusion machinery itself[3][16][43].
SNARE Complex Assembly and Regulation
The primary molecular function of USO1 involves the regulation and facilitation of SNARE (Soluble N-ethylmaleimide-sensitive Factor Attachment Protein Receptor) complex assembly, a process critical for membrane fusion[3][10][29]. The classical SNARE fusion mechanism requires the assembly of four SNARE proteins—three Q-SNAREs from the acceptor (target) membrane and one R-SNARE from the transport vesicle—into a tight helical coiled bundle termed a "SNAREpin" that physically pulls the two membranes together, leading to membrane fusion[34][47]. USO1 specifically interacts with monomeric SNARE proteins rather than pre-assembled SNARE complexes, suggesting a mechanism in which USO1 acts to recruit and stabilize individual SNARE proteins in their monomeric state at sites of vesicle docking[31].
Biochemical binding assays using gel filtration chromatography have demonstrated that USO1 stably interacts with the mammalian ER-Golgi v-SNARE Sec22b and t-SNARE rBet1, but not with assembled SNARE complexes containing these same proteins[31]. The isolation of the SNARE motif as the primary binding site for USO1, combined with structural similarity between USO1's coiled-coil domain and the SNARE motif itself, suggests that USO1 may form a mixed p115/SNARE helix bundle with monomeric SNAREs[31]. This interaction appears to facilitate the binding activity and concentration of SNAREs at pre-fusion sites and stabilizes SNAREs in a conformation conducive to subsequent SNARE complex assembly. Upon formation of the productive trans-SNARE complex and membrane fusion, USO1 dissociates from its SNARE partners, presumably to be recycled for additional rounds of tethering and fusion[31].
The Rab1 Recruitment Mechanism
The recruitment of USO1 to membranes and its subsequent function is critically dependent on the small GTPase Rab1 (also called Ypt1 in yeast), which serves as the primary molecular determinant regulating USO1 membrane localization and activity[1][7][37][40][43][46]. Rab1 exists in two interconvertible nucleotide states: an active GTP-bound state and an inactive GDP-bound state. The active Rab1-GTP directly binds to USO1 through interaction with the globular head domain (H1 tether repeat) and through a second site in the CC1 coiled-coil domain[18][37][40]. This Rab1-mediated recruitment brings USO1 to the surface of ER-derived COPII vesicles and early Golgi membranes, positioning it optimally for interaction with both v-SNAREs on the transport vesicle and t-SNAREs on the target membrane[1][43][46].
Remarkably, genetic and biochemical evidence suggests that Rab1 and USO1 function in a reciprocal relationship more complex than simple substrate-effector binding[1][16][29][43]. The isolated coiled-coil domains of USO1, particularly the CC2 domain, promote the dissociation of Rab1b from guanine nucleotide dissociation inhibitor (GDI), effectively facilitating Rab1 activation and membrane association[40]. This suggests that USO1 functions as a nucleotide exchange factor (GEF) or GEF-stimulating factor for Rab1, representing an example of positive feedback regulation wherein a Rab effector enhances the activation of its own upstream regulator[40]. The GRASP-associated protein GM130 and the Golgi protein giantin further enhance this Rab1-USO1 interaction, demonstrating that the recruitment and regulation of USO1 involves a multi-protein complex[37][40][46].
Cellular Localization and Membrane Association
Dynamic Membrane Cycling
USO1 functions as a peripheral membrane protein that exhibits dynamic cycling between soluble cytoplasmic and membrane-associated pools[1][20][22]. In the basal interphase state, dephosphorylated USO1 associates with the Golgi apparatus and ER-derived vesicles, whereas phosphorylation of the protein promotes its dissociation from membranes and release into the cytosol[1][19][20][22]. This regulated association and dissociation from membranes represents a sophisticated mechanism for controlling vesicular transport capacity in response to cellular signals and phosphorylation events mediated by specific kinases such as casein kinase II[39]. The phosphorylation-dependent regulation allows cells to modulate the rate of transport through tethering and fusion machinery in response to metabolic demands and cell cycle progression.
The regulation of USO1 membrane association is intimately linked to the cell cycle, particularly during mitosis when the Golgi apparatus undergoes dramatic reorganization[39][42][56]. During mitosis, phosphorylation of the Golgi matrix protein GM130 by cyclin-dependent kinase 1 (Cdk1) prevents the binding of USO1 to GM130, effectively removing USO1 from the Golgi membrane[36][39][42]. This phosphorylation-mediated dissociation of USO1 from Golgi receptors is thought to contribute to mitotic Golgi fragmentation by preventing the tethering and fusion of COPI vesicles, allowing continuous vesicle budding without compensatory fusion[36][39]. Upon exit from mitosis and dephosphorylation of Golgi proteins, USO1 reassociates with the reassembling Golgi cisternae, facilitating the reformation of Golgi stacks through its tethering and SNARE-regulatory functions[39][42][59].
Localization at the ER-Golgi Interface
At the molecular level, USO1 specifically localizes to ER-derived COPII vesicles and to the early Golgi cisternae within cis-Golgi compartments through its recruitment by Rab1 and TRAPP complex-mediated activation of Rab1[1][43][46]. High-resolution immunofluorescence microscopy studies tracking the localization of USO1 in living cells demonstrate that USO1 colocalizes almost completely with Rer1, a cargo receptor that continuously cycles between the ER and Golgi, indicating enrichment of USO1 at the early Golgi entry point[43]. The subcellular distribution of USO1 is dependent on RAB1 function, as loss of Rab1 activity results in cytoplasmic dispersal of USO1[40][43]. This dependency on Rab1 for membrane recruitment and localization represents the primary mechanism by which cells spatially organize USO1 to specific membrane trafficking pathways.
USO1 exhibits particularly high colocalization with the early Golgi marker GeaA (mammalian GBF1, a guanine nucleotide exchange factor for ARF1) at approximately 0.52 Pearson's correlation coefficient, suggesting that USO1 functions in close association with ARF1 GEF machinery at early Golgi cisternae[43]. This localization pattern is consistent with USO1's characterized roles in early secretory pathway trafficking. The colocalization of USO1 with Rab1, RAB1 GEF machinery, and cargo carriers demonstrates that USO1 functions within a highly organized molecular complex at distinct membrane domains dedicated to ER-to-Golgi transport.
Transport Vesicle Association
During the budding of COPII vesicles from the ER, Ras-associated protein 1 (Rab1 in its GTP-bound active state) recruits USO1 to the nascent vesicle coat through direct binding to the globular head domain H1 motif[1][20]. This recruitment occurs during the packaging of cargo and SNAREs into COPII-coated vesicles, positioning USO1 to interact with the complement of v-SNAREs present on these nascent transport carriers[1][20]. The interaction of USO1 with COPII vesicle-associated SNAREs forms a cis-SNARE complex in which USO1 brings together ER-derived v-SNAREs with components that will facilitate their interaction with complementary t-SNAREs on the target Golgi membrane[1][20][43]. This organization at the level of individual vesicles ensures specificity and efficiency of membrane trafficking.
Role in Vesicular Transport Pathways
ER-to-Golgi Transport
The primary and most well-characterized role of USO1 is in transport from the endoplasmic reticulum to the Golgi apparatus, a critical step in the secretory pathway through which newly synthesized proteins and lipids traverse the secretory system[1][23][26][30][46]. This transport occurs through the intermediary of the ER-Golgi intermediate compartment (ERGIC), a compartment that receives COPII-coated vesicles from ER exit sites and subsequently mediates their fusion with and/or maturation into cis-Golgi cisternae[26][55]. USO1 is essential for the initial docking of ER-derived COPII vesicles to both the ERGIC and early Golgi cisternae, representing the critical first step in membrane fusion that must precede SNARE-mediated membrane merger[1][23][26].
In vitro biochemical reconstitution experiments using purified yeast proteins have demonstrated that COPII-coated vesicles budded from the ER are freely diffusible in solution but undergo tethering to Golgi membranes upon the addition of purified Uso1p and the small GTPase Ypt1p[23]. Critically, this in vitro docking reaction was found to be independent of SNARE proteins, as thermosensitive mutations in SNARE genes (Sed5p, Bet1p, Bos1p, and Sly1p) that completely prevented fusion did not interfere with the Uso1p-mediated docking step[23]. These experiments established a mechanistic separation between docking and fusion, identifying docking as a Rab1-dependent, USO1-mediated step that precedes and is independent of SNARE engagement. However, subsequent genetic analysis has demonstrated that SNAREs function downstream of and are regulated by USO1, such that overexpression of individual ER-Golgi v-SNAREs can suppress some temperature-sensitive uso1 mutations[10][29].
Retrograde Transport and Golgi Homeostasis
In addition to anterograde transport from the ER to the Golgi, USO1 plays important roles in maintaining Golgi homeostasis through participation in retrograde intra-Golgi transport[20][26]. The Golgi apparatus must maintain equilibrium between forward transport of cargo-containing vesicles and retrograde recycling of resident Golgi enzymes and transport machinery to maintain both enzyme distribution and Golgi structural organization[26]. This balance is achieved through a robust COPI-dependent retrograde pathway in which USO1 participates in COPI vesicle tethering to Golgi membranes, facilitating their fusion and enabling the recycling of transport machinery back to earlier compartments[26][32]. The interaction of USO1 with multiple Golgi-associated proteins including GM130 and giantin positions it centrally in coordinating vesicular transport both in the anterograde and retrograde directions.
Golgi Ribbon Organization and Biogenesis
Beyond its primary role in ER-to-Golgi transport, USO1 functions in the maintenance and organization of the characteristic stacked and interconnected ribbon structure of the mammalian Golgi apparatus[30][56][59]. The Golgi in mammalian cells is uniquely organized as a ribbon of interconnected cisternal stacks positioned near the microtubule-organizing center, an organization that confers efficiency to the secretory pathway[42][56]. USO1 is essential for the stacking of individual Golgi cisternae and for maintaining the structural integrity of Golgi stacks through its interactions with giantin and GM130[6][30][36][59]. The CC4 domain of USO1 is particularly critical for this Golgi ribbon formation function, as deletion of CC4 in otherwise functional p115 results in Golgi fragmentation and disruption of the normal ribbon architecture[6].
In post-mitotic Golgi reassembly following mitotic disassembly, USO1 operates sequentially with its two Golgi receptors, GM130 and giantin, to mediate the initial tethering and stacking of cisternae regenerated through NSF-mediated fusion pathways[59]. USO1 acts most potently at an early stage in the stacking reaction, upstream of the Golgi reassembly stacking protein GRASP65, and may facilitate the initial tethering of Golgi cisternae that is a prerequisite for stacking and Golgi ribbon reformation[59]. This function ensures that following cell division, the Golgi apparatus is properly reconstructed to resume full secretory capacity, highlighting USO1's essential role in maintaining cellular organization.
Mitotic Function and Spindle Pole Association
Recent studies have revealed an unexpected and distinct role for USO1 in cell mitosis, independent of its classical transport functions[56]. Unlike typical Golgi markers that diffuse throughout the cytoplasm following Golgi disassembly at mitosis, USO1 partitions with the spindle poles throughout mitosis, associated with centrosomal gamma-tubulin[56]. The N-terminal armadillo-fold within the globular head domain mediates a direct novel interaction between USO1 and gamma-tubulin, functioning to target USO1 to centrosomes[56]. Strikingly, depletion of USO1 results in spindle collapse, chromosome missegregation, and failed cytokinesis through impaired resolution of the cytokinetic bridge[56]. These findings demonstrate that USO1 is essential for mitotic spindle function and proper cell division, implicating it as the only known golgin to regulate both mitosis and apoptosis through distinct molecular interactions[56]. This mitotic function likely evolved as a consequence of USO1's maintenance at spindle poles to facilitate centrosome-directed Golgi reassembly at the end of mitosis.
Molecular Interactions and Binding Partners
SNARE Protein Interactions
The interaction of USO1 with SNARE proteins represents perhaps the most functionally significant binding events in which USO1 engages[3][10][31][43][47]. USO1 specifically interacts with the v-SNARE Sec22b and the t-SNARE components Bet1 (rBet1) and Bos1, which form cognate SNARE pairs at the ER-Golgi interface[31][43][47]. The binding preference of USO1 for these specific SNAREs is mediated by their presence as monomeric proteins, as demonstrated through binding studies showing that USO1 fails to interact with assembled SNARE complexes containing the same proteins[31]. This selectivity for monomeric over oligomeric SNARE states suggests that USO1 acts at an early stage in SNARE complex assembly, capturing monomeric SNAREs before they engage in non-productive interactions and channeling them toward productive cognate SNARE complex formation.
The interaction between USO1 and SNAREs involves structural complementarity between the coiled-coil domain of USO1 and the alpha-helical SNARE motif, suggesting formation of a mixed p115/SNARE coiled-coil bundle[31][43]. Notably, USO1 does not bind equally to all ER-Golgi SNAREs, as it fails to interact with membrin and syntaxin-5 despite these being documented t-SNARE components[31]. This specificity likely reflects the importance of preventing aberrant SNARE complex formation and ensuring that only productive, cognate SNARE combinations assemble, a function particularly critical in the early secretory pathway where numerous SNAREs are present simultaneously.
Rab GTPase Interactions
The interaction between USO1 and the small GTPase Rab1 (Ypt1 in yeast) represents one of the most extensively characterized effector-GTPase interactions in the cell[1][37][40][43][46]. Rab1 binds to USO1 through two distinct sites: one in the globular head domain (H1 tether repeat) and a second in the first coiled-coil domain (CC1)[18][37][40]. The interaction is highly specific for the GTP-bound active form of Rab1, with biochemical assays showing that only approximately 5% of full-length p115 binds to the inactive GDP-bound form of Rab1 compared to robust binding to the GTP-locked mutant Rab1-QL[37]. Interestingly, truncations of p115 lacking the C-terminal region show enhanced Rab1 binding (over 20% of input) compared to full-length protein, suggesting that the C-terminal domain exerts autoinhibitory effects on the CC1-mediated Rab1 binding site[37].
This autoinhibition of Rab1 binding by the C-terminal domain is relieved by the binding of Golgi proteins GM130 and giantin to the USO1 acidic tail[37]. Displacement of the C-terminal domain by golgin binding exposes the Rab1 binding site on the coiled-coil, facilitating high-affinity Rab1 interaction[37]. This regulatory mechanism allows golgins to enhance USO1 recruitment to Golgi membranes by promoting Rab1-binding-competent conformations of USO1, providing sophisticated spatial and temporal control of USO1 localization and function. In turn, USO1 facilitates Rab1 activation by promoting dissociation of Rab1 from its cytoplasmic inhibitor GDI, representing a positive feedback loop in which the effector enhances activation of its upstream regulator[40].
Golgi Matrix Protein Interactions
USO1 interacts with GM130 and giantin, two major Golgi matrix proteins that form part of the structural framework of the Golgi apparatus[20][24][36][37][40][59]. The binding sites for USO1 are distinct for each golgin: GM130 interacts with the acidic C-terminal tail of USO1 through its N-terminal domain, whereas giantin also contacts this C-terminal acidic domain[21][36][37]. These interactions are functionally important for both tethering COPI vesicles to Golgi membranes and for maintaining Golgi structural integrity[21][36][59]. Importantly, under certain in vitro conditions, GM130 and giantin compete for binding to USO1, but under other conditions USO1 can simultaneously interact with both proteins and link them together, suggesting that the stoichiometry and configuration of these complexes may be dynamically regulated[21][24].
The biological significance of these interactions was demonstrated through microinjection experiments in which antibodies or peptides preventing p115-GM130 or p115-giantin interactions inhibited transport and caused accumulation of transport-sized vesicles[24][36]. Critically, these interactions were shown to function sequentially rather than simultaneously: anti-p115 antibodies inhibited transport at the level of vesicular-tubular clusters (early transport stage), while anti-GM130 and anti-giantin antibodies inhibited at later stages after VSV-G protein reached the Golgi complex[24]. This sequential requirement supports a model in which USO1 works with GM130 early in transport to mediate initial vesicle docking, then with giantin at later stages for COPI vesicle transport within and around the Golgi stack[24].
COPII Coat and Adaptor Protein Interactions
During the biogenesis of COPII-coated vesicles at ER exit sites, USO1 is recruited to the forming carrier through interaction with Rab1, positioning it at the interface between the COPII coat machinery and the cargo-SNARE-containing vesicle interior[1][20]. While direct interactions between USO1 and COPII coat components have not been extensively documented in the literature, the recruitment of USO1 to COPII vesicles is mediated through Rab1-TRAPP complex interactions, in which the TRAPP complex associates with COPII coat components and functions as a guanine nucleotide exchange factor (GEF) for Rab1[46]. The TRAPP complex acts in concert with USO1/p115 to bring COPII vesicles to the ERGIC and cis-Golgi membrane for fusion[46].
Regulation of USO1 Function
Phosphorylation-Dependent Regulation
USO1 function is regulated through phosphorylation by casein kinase II (CKII) or CKII-like kinases, with phosphorylation occurring at serine residues within the acidic C-terminal tail region[39][59]. This phosphorylation serves a critical role in the regulated release of USO1 from membranes into the cytosolic pool, allowing for dynamic cycling of the protein and control of tethering capacity[39][59]. In the post-mitotic Golgi reassembly system, phosphorylation of p115 by CKII is required for NSF-mediated cisternal regrowth, suggesting that phosphorylation may strengthen the bridge formed by p115 as it links GM130 and giantin and facilitates transfer of vesicles to the next step of the docking reaction[39][59]. Notably, very little phosphorylated p115 is detected on Golgi membranes, with most phosphorylated protein found in the cytosol, suggesting that phosphorylated p115 is rapidly released from the membrane following its action on tethering complexes[39].
During mitosis, the phosphorylation of USO1 is dramatically reduced, with mitotic phosphorylation events primarily affecting GM130 rather than p115 itself[39][59]. The phosphorylation of GM130 by Cdk1 during mitosis prevents the binding of p115 to the Golgi apparatus, providing a mechanism for the inhibition of COPI vesicle tethering and fusion during mitosis that allows continuous vesicle budding without compensatory fusion, resulting in Golgi fragmentation[39][42]. This phosphorylation-based regulation represents a sophisticated mechanism for coordinating Golgi disassembly with cell cycle progression and ensures temporal control of membrane trafficking during cell division.
Conformational Dynamics and Auto-Inhibition
Evidence from biochemical and biophysical studies suggests that USO1 may undergo conformational changes between closed and open states that regulate accessibility of binding sites[33][37]. A model has been proposed in which USO1 alternates between closed and open conformations, with closed conformations hiding the Rab1 binding site present in the coiled-coil region through intramolecular interactions between the globular domain and C-terminal acidic region, and open conformations exposing this site for Rab1 binding[37]. However, recent analytical ultracentrifugation studies directly addressing whether the E6K/G540S mutations in the globular head domain promote conformational changes have found no large conformational differences in full-length Uso1 between wild-type and mutant forms, as reflected in unchanged sedimentation coefficients[33]. This argues against simple models of large-scale conformational switching and instead suggests that more subtle regulatory mechanisms may govern USO1 function, or that the primary regulatory effect of Golgin binding involves simple steric displacement of the C-terminal domain rather than inducing major conformational rearrangement.
Biological Significance and Disease Associations
Essential Role in Cell Viability
USO1 is essential for cell viability in virtually all eukaryotic organisms examined, with deletion of the yeast uso1 gene being lethal at all temperatures[3][16]. This essential role reflects the absolute requirement for ER-to-Golgi transport for basic cellular survival and the inability of cells to compensate for loss of USO1 function through alternative pathways[3][16]. Unlike many other Golgi tethering factors that exhibit functional redundancy, USO1 is unique among Golgi tethers in that it is required for viability, with all other characterized golgins being non-essential even when individually deleted[3][30]. This essential function is not mediated by long-range vesicle tethering, as the isolated monomeric globular head domain lacking the extended coiled-coil region remains capable of providing essential functions[3][16][33].
Tissue-Specific Expression and Function in Reproduction
While USO1 is ubiquitously expressed across tissues, analysis of tissue-specific expression patterns reveals particularly high expression levels in mouse testes and in germ cells engaged in spermatogenesis[38]. Targeted deletion of Uso1 in germ cell lines results in suppression of cell proliferation and growth while stimulating apoptosis[38]. Furthermore, Uso1 knockout blocks cell cycle progression and weakens DNA damage repair mechanisms in spermatocytes[38]. Transcriptomic analysis of Uso1-depleted germ cells reveals dysregulation of numerous transcripts encoding testis-enriched and reproduction-related genes that regulate proliferation, differentiation, and meiosis[38]. These findings establish USO1 as playing a vital role in male fertility through coordination of centriolar satellites and regulation of germ cell proliferation and differentiation, suggesting tissue-specific functions in reproduction beyond its classical role in general vesicular transport[38].
Implications in Cancer and Disease
Dysregulation of USO1 expression has been implicated in multiple cancer types, including hematologic malignancies, colorectal cancer, gastric cancer, lung adenocarcinomas, and hepatocellular carcinoma[28][30][38][54]. In hepatocellular carcinoma, USO1 plays an oncogenic role by promoting cell proliferation, evading apoptosis, and disrupting normal cell cycle progression[28][30][38][54]. Studies on the involvement of USO1 in liver cancer development have identified USO1 as a potential tumor biomarker and therapeutic target[28][54]. Additionally, disease associations recorded in genetic databases link mutations in USO1 with retinal disease and hypogonadotropic hypogonadism with or without anosmia[27][48], suggesting that disruption of USO1 function can result in multi-systemic manifestations reflecting the consequences of impaired secretory pathway function in specialized tissues.
Conclusion
The USO1 gene encodes the general vesicular transport factor p115, a sophisticated molecular machine essential for coordinating membrane fusion in the early secretory pathway. Through its characterization over three decades, USO1 has emerged from initial characterization as a simple vesicle tether to recognition as a complex regulatory hub integrating multiple signaling inputs to control SNARE-mediated membrane fusion. The protein's dimeric architecture comprising a novel alpha-solenoid globular head domain with 12 tether repeat motifs, an extended coiled-coil region mediating dimerization and providing secondary binding sites, and a C-terminal region linking to Golgi structural proteins provides the molecular basis for its multifaceted functions in transport, tethering, and cellular regulation. Recent structural and genetic studies have conclusively demonstrated that the essential cellular function of USO1 involves the regulation of SNARE complex assembly rather than long-range membrane tethering, fundamentally revising decades of mechanistic understanding. Through recruitment by the small GTPase Rab1 and interaction with multiple golgins and SNARE proteins, USO1 coordinates the delivery of newly synthesized cargo from the endoplasmic reticulum through the Golgi apparatus to maintain cellular protein secretion and organellar homeostasis. The discovery of tissue-specific functions in spermatogenesis and associations with cancer development suggest that USO1 plays broader regulatory roles beyond its primary transport function. Given its essentiality for cell viability and its discrete molecular functions, USO1 represents an attractive target for therapeutic intervention in diseases characterized by secretory pathway dysfunction or dysregulated cell proliferation.
Citations
- https://en.wikipedia.org/wiki/USO1
- https://www.ensembl.org/Homo_sapiens/Gene/Summary?db=core%3Bg%3DENSG00000138768%3Bt%3DENST00000514213
- https://elifesciences.org/articles/85079
- https://www.uniprot.org/uniprotkb/O60763/entry
- https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=8615
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4450726/
- https://pubmed.ncbi.nlm.nih.gov/28973856/
- https://www.ncbi.nlm.nih.gov/gene/8615
- https://pubmed.ncbi.nlm.nih.gov/8603910/
- https://www.pnas.org/doi/abs/10.1073/pnas.1708394114?doi=10.1073%2Fpnas.1708394114
- https://pubmed.ncbi.nlm.nih.gov/8812994/
- https://www.yeastgenome.org/locus/S000002216
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2754402/
- https://www.rcsb.org/structure/3gq2
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2198842/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1170560/
- https://pubmed.ncbi.nlm.nih.gov/11035033/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11446975/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3119910/
- https://www.genecards.org/cgi-bin/carddisp.pl?gene=USO1
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8699264/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4304910/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3196633/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3179334/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4064004/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC14799/
- https://www.med.upenn.edu/shorterlab/Papers/JBC-05.pdf
- https://pmc.ncbi.nlm.nih.gov/articles/PMC12072129/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2175190/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4187009/
- https://www.proteinatlas.org/ENSG00000138768-USO1
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4679863/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10275640/
- https://pubmed.ncbi.nlm.nih.gov/10819516/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11114727/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6523435/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2323215/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6278416/
- https://www.rcsb.org/structure/3grl
- https://pubmed.ncbi.nlm.nih.gov/16024819/
- https://www.spandidos-publications.com/10.3892/ol.2017.5814
- https://pubmed.ncbi.nlm.nih.gov/16723730/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3122246/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8553993/
- https://elifesciences.org/articles/00947
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2199741/
- https://pubmed.ncbi.nlm.nih.gov/26216056/