Deep Research
Bioreason Pro
(Ghr-deep-research-bioreason-sft.md)
Bioreason Pro
(Ghr-deep-research-bioreason-sft.md)Thinking Trace
I begin with the InterPro architecture. The N-terminal extracellular region contains IPR015152 (Growth hormone/erythropoietin receptor, ligand binding, residues 46–131), which is the canonical cytokine-binding module of class I hematopoietin receptors. This segment is embedded within immunoglobulin-like and fibronectin type III folds: IPR013783 (Immunoglobulin-like fold, residues 50–149) and IPR003961 (Fibronectin type III, residues 149–250), with the corresponding superfamily annotations IPR036116 (Fibronectin type III superfamily, residues 50–148 and 149–252) and IPR013783 (Immunoglobulin-like fold, residues 150–250). This extracellular module arrangement is characteristic of ligand-gated receptors that capture a specific peptide hormone at the cell surface. Immediately downstream, IPR003528 (Long hematopoietin receptor, single chain, conserved site, residues 171–249) marks the conserved signature of long-chain cytokine receptors, which include the WSXWS-related motif and the proline-rich Box1/Box2 regions that organize intracellular kinase docking. The ectodomain also includes IPR025871 (Growth hormone-binding protein, residues 317–616), indicating that a proteolytically released soluble ectodomain (GHRP) circulates and binds growth hormone with high affinity. Together with the stated topology (single-pass type I membrane protein), this architecture causes ligand-dependent receptor dimerization/rotation at the plasma membrane and positions a cytoplasmic tail to recruit tyrosine kinases and SH2-containing effectors.
From this domain layout, the molecular function follows. The ligand-binding cytokine receptor module confers growth hormone receptor activity (GO:0004903). The conserved cytoplasmic tail motifs in long hematopoietin receptors create phosphotyrosine docking sites for SH2 domains, establishing SH2 domain binding (GO:0042169). The same tail recruits non-receptor protein tyrosine kinases, consistent with protein kinase binding (GO:0019901), and engages regulatory phosphatases that tune signaling amplitude, consistent with protein phosphatase binding (GO:0019903). The presence of a cleavable ectodomain and the membrane-anchored form together explain how signaling is initiated by hormone-induced conformational changes that juxtapose cytoplasmic Box1/Box2 motifs to bind and activate associated kinases.
These molecular activities drive specific biological processes. Ligand-induced activation of the receptor’s associated tyrosine kinase triggers peptidyl-tyrosine phosphorylation (GO:0018108) on the receptor and STATs, initiating the growth hormone receptor signaling pathway via JAK-STAT (GO:0060397) and its positive regulation (GO:0060357). Phosphorylated STATs translocate to the nucleus to modulate transcription, while parallel recruitment of SHC/GRB2-SOS elicits activation of protein kinase activity (GO:0032147) and positive regulation of MAP kinase activity (GO:0043406). These signaling axes underlie organismal and tissue-level outcomes: positive regulation of cell differentiation (GO:0045597), cartilage development involved in endochondral bone morphogenesis (GO:0060351), and broader anatomical structure morphogenesis (GO:0009887). Because growth hormone integrates endocrine cues, the receptor participates in hormone-mediated signaling (GO:0009755) and cellular responses to growth hormone (GO:0071378), insulin (GO:0032869), interleukin-1 (GO:0071347), morphine (GO:0071316), and glucocorticoid (GO:0051384), reflecting crosstalk with metabolic and inflammatory pathways. Neurotrophic outputs of GH/IGF signaling contribute to negative regulation of neuron death (GO:1901215). System-level modulation of metabolism and behavior aligns with response to food (GO:0032094) and response to gravity (GO:0009629), consistent with GH’s role in fluid balance and posture reflexes. The receptor’s signaling also intersects with vascular and renal physiology, explaining its involvement in regulating renal sodium excretion and blood pressure.
The cellular component is dictated by the architecture and processing. The single-pass type I configuration places the full-length receptor at the plasma membrane (GO:0005886), where ligand binding occurs. Proteolytic shedding yields the secreted growth hormone-binding protein in the extracellular space (GO:0005615), which buffers hormone availability and shapes signaling gradients. Activated complexes form cytoplasmic signaling hubs (GO:0005737) that culminate in nuclear signaling (GO:0005634) via STAT translocation. Expression in neurons situates the receptor in the neuronal cell body (GO:0043025), and GH/IGF signaling can influence mitochondrial function (GO:0005739), consistent with observed associations with oxidative metabolism.
Mechanistically, hormone binding to the extracellular cytokine receptor module stabilizes an active dimer that reorients the cytoplasmic tails, enabling the conserved site (IPR003528) to bind and activate associated tyrosine kinases. The receptor then undergoes peptidyl-tyrosine phosphorylation, creating SH2 docking sites for STAT5A/STAT5B, which are phosphorylated and translocate to the nucleus to drive transcriptional programs for growth, differentiation, and metabolic adaptation. Parallel recruitment of SHC/GRB2-SOS activates MAPK, while phosphatases and SOCS proteins (e.g., SOCS2) bind to attenuate signaling. The secreted ectodomain (GHRP) modulates hormone bioavailability and receptor occupancy, shaping the amplitude and duration of signaling.
These inferences suggest specific interaction partners. The cytoplasmic tail’s kinase-docking motifs predict association with tyrosine-protein kinase JAK2 as the primary activator, with possible context-dependent engagement of JAK3. The SH2 docking sites recruit STAT5A and STAT5B. Negative feedback arises via SOCS2 binding. The ligand is somatotropin (growth hormone), and downstream endocrine coupling predicts functional interaction with insulin-like growth factor I. Crosstalk with prolactin and prolactin-6A1 is plausible through shared JAK/STAT machinery and overlapping transcriptional programs. Generic “tyrosine-protein kinase” entries reflect additional non-receptor kinases that can transiently bind during signaling bursts.
## Functional Summary
A single-pass cell-surface cytokine receptor that binds growth hormone through an extracellular ligand-binding module and, upon ligand-induced dimerization, activates an associated tyrosine kinase to drive tyrosine phosphorylation and STAT-mediated transcription. This signaling promotes growth, differentiation, and metabolic adaptation, with parallel engagement of MAP kinase pathways and crosstalk with insulin, inflammatory, and stress-responsive hormones. The receptor signals from the plasma membrane with transient cytoplasmic complexes and nuclear translocation of activated transcription factors, while a proteolytically released soluble ectodomain circulates as a high-affinity growth hormone-binding protein that modulates hormone availability and signaling range. Its activity contributes to renal sodium excretion and blood pressure control and extends to neuronal and mitochondrial contexts where it supports cell survival and metabolic tuning.
## UniProt Summary
Receptor for pituitary gland growth hormone involved in regulating postnatal body growth. On ligand binding, couples to, and activates the JAK2/STAT5 pathway (By similarity). Involved in regulating renal sodium excretion and blood pressure.
## InterPro Domains
- IPR015152: Growth hormone/erythropoietin receptor, ligand binding (domain) [46-131]
- IPR013783: Immunoglobulin-like fold (homologous_superfamily) [50-149]
- IPR036116: Fibronectin type III superfamily (homologous_superfamily) [50-148]
- IPR003961: Fibronectin type III (domain) [149-250]
- IPR036116: Fibronectin type III superfamily (homologous_superfamily) [149-252]
- IPR013783: Immunoglobulin-like fold (homologous_superfamily) [150-250]
- IPR003961: Fibronectin type III (domain) [151-254]
- IPR003528: Long hematopoietin receptor, single chain, conserved site (conserved_site) [171-249]
- IPR025871: Growth hormone-binding protein (domain) [317-616]
## GO Term Predictions
### Molecular Function
### Biological Process
### Cellular Component
Falcon
(Ghr-deep-research-falcon.md)
Falcon
(Ghr-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: Rat Ghr (Growth hormone receptor; UniProt P16310) functional annotation
Scope and identity verification (mandatory)
The target gene symbol Ghr in Rattus norvegicus corresponds to the canonical growth hormone receptor (GHR), a class I/type I cytokine receptor family single-pass transmembrane receptor. Structural descriptions match UniProt P16310 expectations: an extracellular GH-binding domain built from two fibronectin type III (FNIII)-like modules, a single transmembrane helix, and an intracellular region containing Box1 and Box2 motifs that couple to cytoplasmic kinases (gun2022investigationofmonoclonal pages 5-9, gun2022investigationofmonoclonal media b3c4ed24, gun2022investigationofmonoclonal media 19db3114). The same receptor system also yields a soluble growth hormone-binding protein (GHBP) corresponding to the receptor extracellular domain (gun2022investigationofmonoclonal pages 9-13, list2001growthhormonereceptorbinding pages 1-3).
1) Key concepts and definitions (current understanding)
1.1 What GHR is (molecular function)
GHR is a cell-surface receptor whose primary function is to bind circulating growth hormone (GH) and transduce that extracellular hormonal signal into intracellular phosphorylation cascades and gene regulation, notably including induction of hepatic IGF-1 and many other GH-responsive genes (cartersu2016growthhormonesignaling pages 1-5, dehkhoda2018thegrowthhormone pages 1-2).
1.2 Domain architecture and conserved motifs
A widely used structural model partitions GHR into an extracellular domain (ECD), a transmembrane domain (TMD), and an intracellular domain (ICD). One detailed schematic assigns approximate boundaries ECD ~19–262, TMD ~263–288, and ICD ~289–638, and highlights the intracellular Box1 motif (proximal, proline-rich) and Box2 region (gun2022investigationofmonoclonal pages 5-9, gun2022investigationofmonoclonal media 19db3114). The ECD can also be represented as a soluble GHBP form (gun2022investigationofmonoclonal pages 9-13, gun2022investigationofmonoclonal media 19db3114).
1.3 GH-binding protein (GHBP)
GHBP is a soluble binding partner for GH in circulation that corresponds to the receptor ECD. It binds GH with receptor-like affinity and is treated genetically as a product of the same GHR/GHBP gene system in rodent knockout studies (gun2022investigationofmonoclonal pages 9-13, list2001growthhormonereceptorbinding pages 1-3). Functionally, GHBP is described as influencing GH bioavailability and transport in blood (ortiz2014…impactof pages 46-51, gun2022investigationofmonoclonal pages 9-13).
2) Mechanism of receptor activation and signaling pathways
2.1 Preformed dimers and 1:2 ligand:receptor stoichiometry
Modern mechanistic models emphasize that GHR exists as a preformed homodimer at the cell surface (and can assemble in the ER) rather than relying primarily on ligand-induced dimerization (dehkhoda2018thegrowthhormone pages 1-2, dehkhoda2018thegrowthhormone pages 2-5). GH engages the receptor with 1 GH : 2 GHR stoichiometry (equivalently 2:1 receptor:ligand), binding “site 1” and “site 2” across the two receptor chains (wojcik2018postreceptorinhibitorsof pages 1-3, dehkhoda2018thegrowthhormone pages 2-5).
2.2 JAK2 coupling and activation (core biochemical mechanism)
GHR lacks intrinsic kinase activity; instead its intracellular Box1 (and Box2) region is central for recruiting/coupling to JAK2, which is the principal JAK associated with GHR (gun2022investigationofmonoclonal pages 5-9, dehkhoda2018thegrowthhormone pages 1-2). Upon GH binding and receptor conformational rearrangement, the two receptor-associated JAK2 molecules undergo trans-phosphorylation, which initiates downstream signaling (dehkhoda2018thegrowthhormone pages 2-5, ortiz2014…impactof pages 46-51).
2.3 Major downstream pathways
Evidence from mechanistic reviews supports three major branches:
- JAK2 → STAT pathway: JAK2 phosphorylates receptor tyrosines and activates STAT5a/STAT5b (dominant), as well as STAT1 and STAT3, which dimerize and translocate to the nucleus to regulate transcription (cartersu2016growthhormonesignaling pages 5-8, cartersu2016growthhormonesignaling pages 1-5, dehkhoda2018thegrowthhormone pages 1-2).
- MAPK/ERK pathway: GH/GHR signaling can activate ERK1/2 through Shc/Grb2/SOS/Ras/Raf/MEK signaling (cartersu2016growthhormonesignaling pages 5-8, cartersu2016growthhormonesignaling pages 1-5, ortiz2014…impactof pages 46-51).
- PI3K/Akt pathway: GH can drive IRS1/2 phosphorylation and PI3K/Akt activation, providing metabolic signaling outputs (cartersu2016growthhormonesignaling pages 5-8, cartersu2016growthhormonesignaling pages 1-5, ortiz2014…impactof pages 46-51).
2.4 Alternative signaling branch (LYN/ERK) and pathway modularity
Recent rodent genetic work emphasizes that the receptor can engage distinct signaling branches. In particular, GHR signaling can be partitioned into canonical JAK2–STAT5 versus an alternative LYN–ERK1/2 pathway; mutations in Box1 can prevent JAK2 activation while preserving LYN activity (chhabra2024therolesof pages 1-2, chhabra2024therolesofa pages 1-2). This supports a view of GHR as a modular signaling platform whose outputs depend on intracellular motif integrity and cellular context (chhabra2024therolesof pages 1-2, chhabra2024therolesofa pages 1-2).
3) Subcellular localization and trafficking
GHR is a single-pass plasma-membrane receptor (gun2022investigationofmonoclonal pages 5-9, gun2022investigationofmonoclonal media b3c4ed24). Dimer formation can occur during biosynthesis (ER) before surface delivery (dehkhoda2018thegrowthhormone pages 2-5). Following activation, receptor complexes can internalize; one review notes evidence consistent with continued JAK2 association after internalization, supporting the possibility of endosomal signaling (wojcik2018postreceptorinhibitorsof pages 1-3).
A 2023 bioRxiv/2024-indexed preprint used super-resolution microscopy to quantify plasma-membrane dynamics of GHR and prolactin receptor (PRLR) in breast cancer cells, reporting ligand-dependent loss of surface GHR and implicating Box1/JAK2 coupling in cross-receptor regulation of surface availability (chen2024arolefor pages 1-3). Although not a rat study, this represents a recent mechanistic advance relevant to GHR trafficking concepts.
4) Negative regulation and homeostatic control (expert synthesis)
Multiple authoritative sources converge on the view that GH signaling must be tightly constrained to avoid pathological outcomes. Negative regulation occurs at several levels:
- SOCS/CIS feedback: STAT5 induces SOCS2 expression, and SOCS2 can bind phosphorylated GHR tyrosines (e.g., Tyr487, Tyr595 noted in one review) and recruit an E3 ubiquitin ligase complex that promotes GHR ubiquitination, internalization, and lysosomal degradation (fernandezperez2016growthhormonereceptor pages 4-7). SOCS1/3 and CIS can also inhibit signaling (fernandezperez2016growthhormonereceptor pages 4-7, wojcik2018postreceptorinhibitorsof pages 1-3).
- PIAS proteins: PIAS family proteins are listed as post-receptor inhibitors of JAK/STAT outputs (wojcik2018postreceptorinhibitorsof pages 1-3).
- Protein tyrosine phosphatases (PTPs): PTP1B and other phosphatases (e.g., SHP1/2) dephosphorylate pathway components and restrict signal duration (fernandezperez2016growthhormonereceptor pages 4-7, wojcik2018postreceptorinhibitorsof pages 1-3).
Kinetic measurements summarized in one review indicate that GH-induced JAK2–STAT5b activation can be transient (maximal within ~30 minutes) followed by a refractory period of ~3–4 hours unless GH is removed, consistent with strong negative feedback and receptor downregulation processes (fernandezperez2016growthhormonereceptor pages 4-7).
5) Recent developments and latest research (prioritizing 2023–2024)
5.1 2024: Genetic uncoupling of signaling branches links JAK2/STAT5 to lifespan and cancer
A 2024 Endocrinology paper (advance access) used a panel of GHR mutant mice to separate signaling outputs (STAT5-deficient mutants, Box1 mutants, and full knockouts) and related them to lifespan and cancer incidence. In the reported results, Box1 mutant males (retaining Lyn activation) had a median lifespan of 1016 days compared with 890 days for Ghr−/− males; in females, GhrBox1−/− had median lifespan 970 days vs 911 days in knockouts (chhabra2024therolesofa pages 1-2). This work provides updated evidence that different intracellular signaling arms downstream of GHR can have separable organismal consequences (chhabra2024therolesofa pages 1-2).
5.2 2023–2024 preprint: Nanoscale membrane organization and receptor crosstalk (GHR–PRLR)
A bioRxiv preprint (posted Sept 5, 2023; indexed here as 2024) applied dSTORM to quantify GHR and PRLR clustering and colocalization on the plasma membrane after GH or PRL exposure. It reports that PRL can induce loss of surface GHR in cells co-expressing PRLR, and concludes Box1/JAK2 coupling is crucial for one receptor’s ligand activation affecting the other’s surface availability (chen2024arolefor pages 1-3). While focused on human cancer cells, this is a recent methodological and mechanistic development relevant to receptor trafficking and signaling integration.
6) Current applications and real-world implementations
6.1 Endocrine physiology and clinical parallels inferred from rodent genetics
Rodent GHR/GHBP knockout models remain central real-world implementations for functional annotation because they create a defined loss-of-function state. A detailed 3-year update reports strong growth impairment and endocrine changes in GHR/BP knockout mice: body weight ~45% of wild type at 4 weeks and ~40% of wild-type maximum weight, with undetectable serum IGF-I, plus reproductive and hormonal alterations (list2001growthhormonereceptorbinding pages 1-3). These phenotypes parallel the conceptual framework of GH insensitivity syndromes and demonstrate the centrality of the GH→GHR→IGF axis in growth and metabolism (list2001growthhormonereceptorbinding pages 1-3).
6.2 Metabolic and hepatic disease modeling via tissue-specific disruption
A mechanistic review compiles tissue-specific mouse models indicating that hepatic disruption of GH/GHR/IGF signaling can produce large decreases in circulating IGF-1 (e.g., ~80% reduction with hepatocyte-specific igf1 deficiency, and >90% reduction in an Alb-cre liver-specific GHR deletion context) together with metabolic phenotypes such as liver steatosis and insulin resistance (dehkhoda2018thegrowthhormone pages 14-15). These models are widely used to dissect endocrine versus local GH actions and to model fatty liver and insulin resistance mechanisms downstream of altered GH signaling (dehkhoda2018thegrowthhormone pages 14-15).
6.3 Developmental endocrinology in rat: GH insensitivity early in life
A rat in vivo study (Sprague–Dawley pups) administered rat GH 2 mg/kg intravenously and measured signaling responses in liver. Despite JAK2 and STAT5 protein expression, JAK2/STAT5 phosphorylation was absent at postnatal day 1 and 4 after GH stimulation; STAT3/STAT1 activation was also not detected in newborn stages, and ERK1/2 activation emerged later (4 days onward) (ruonan2018growthhormonedid pages 1-2). This provides a rat-specific implementation illustrating how Ghr signaling output depends on developmental stage and hepatocyte competence (ruonan2018growthhormonedid pages 1-2).
7) Relevant statistics and data (selected quantitative highlights)
- GHR/BP knockout mouse growth: ~45% of WT weight at 4 weeks; ~40% of WT maximum weight; serum IGF-I undetectable (list2001growthhormonereceptorbinding pages 1-3).
- Reproductive metrics (mouse GHR/BP knockout): pregnancy rate 88% (control matings) vs 53% when males are knockout (P < 0.001); vaginal opening 35.7 ± 0.2 days vs 28.6 ± 0.6 days (P < 0.001) (list2001growthhormonereceptorbinding pages 1-3).
- Hepatic IGF axis perturbation (mouse): circulating IGF-1 reduced ~80% in hepatocyte-specific igf1 deficiency; >90% reduction (context: circulating IGF-1) with liver-specific GHR deletion (Alb-cre), with metabolic sequelae (dehkhoda2018thegrowthhormone pages 14-15).
- Negative regulation phenotype: SOCS2−/− mice ~40% larger (fernandezperez2016growthhormonereceptor pages 4-7).
- Signal kinetics: GH-induced STAT5b activation maximal within ~30 min, refractory ~3–4 h (fernandezperez2016growthhormonereceptor pages 4-7).
- 2024 lifespan medians (mouse GHR mutants): Box1 mutant males 1016 days vs Ghr−/− males 890 days; Box1 mutant females 970 days vs Ghr−/− females 911 days (chhabra2024therolesofa pages 1-2).
8) Expert interpretation (authoritative synthesis)
Collectively, high-citation mechanistic reviews and targeted genetic studies support a consistent model: rat Ghr encodes a non-enzymatic cytokine receptor whose central biochemical role is to organize and activate JAK2 at the plasma membrane in response to GH binding, driving transcriptional programs via STAT5 and complementary metabolic and proliferative programs via ERK and PI3K/Akt (cartersu2016growthhormonesignaling pages 1-5, dehkhoda2018thegrowthhormone pages 1-2, dehkhoda2018thegrowthhormone pages 2-5). The field’s “current understanding” has shifted away from simple ligand-induced dimerization toward preformed dimers activated by conformational rearrangement, with transmembrane and intracellular geometry changes mediating JAK2 activation (dehkhoda2018thegrowthhormone pages 2-5, ortiz2014…impactof pages 46-51).
More recent (2023–2024) work emphasizes that GHR outputs are branch-specific and context dependent: Box1/JAK2/STAT5 signaling can be separated from Src-family-kinase signaling with distinct organismal consequences, and receptor trafficking/availability can be dynamically regulated and integrated with related cytokine receptors (chhabra2024therolesofa pages 1-2, chen2024arolefor pages 1-3). For rat-specific annotation, developmental data indicate that the presence of GHR pathway proteins alone is not sufficient—newborn hepatocytes can show functional GH insensitivity at the level of phosphorylation activation (ruonan2018growthhormonedid pages 1-2).
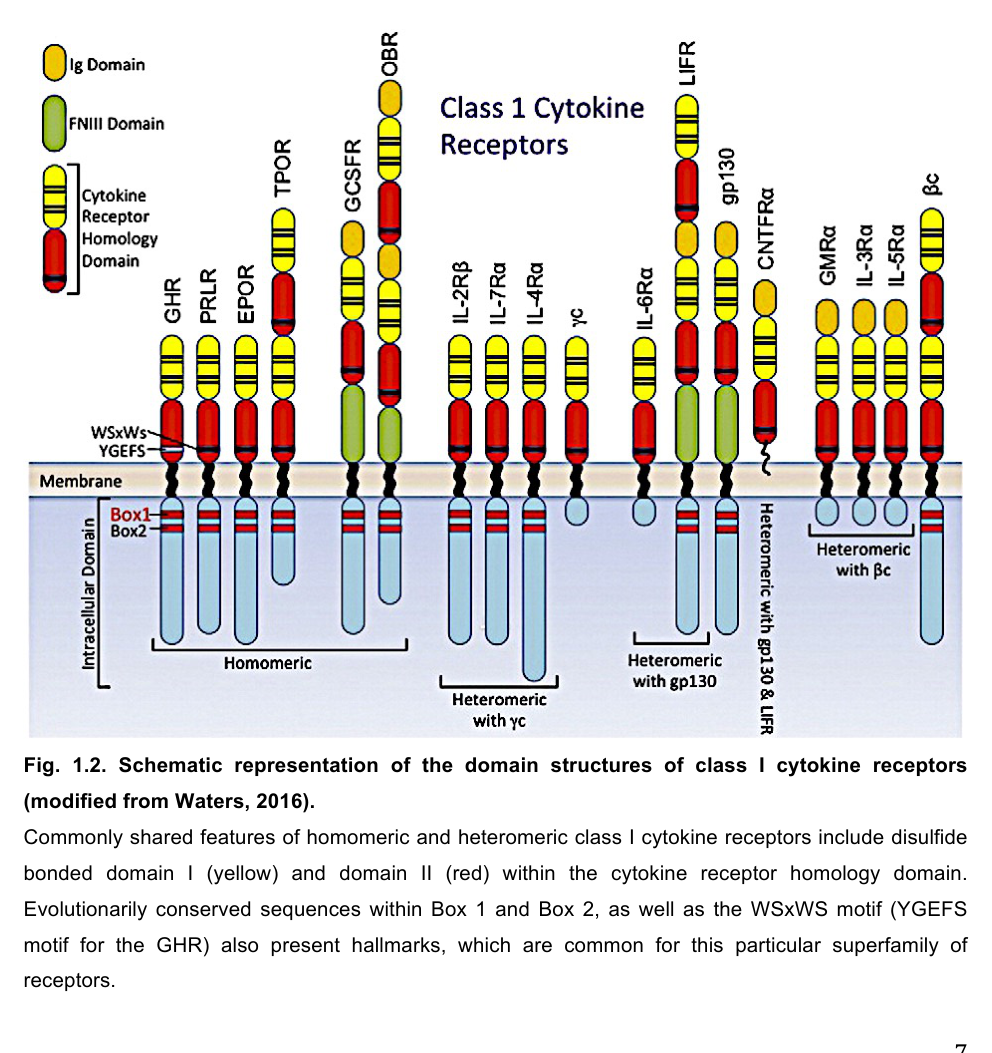
Visual evidence (domain architecture)
A representative schematic of GHR/class I cytokine receptor domain architecture—highlighting FNIII extracellular modules, single transmembrane region, and intracellular Box motifs, plus annotation related to GHBP generation—was retrieved from a GHR-focused source (gun2022investigationofmonoclonal media b3c4ed24, gun2022investigationofmonoclonal media 19db3114).
Summary table
| Aspect | Key points | Best supporting citations | Source details |
|---|---|---|---|
| Identity / domains | Rat Ghr (UniProt P16310) matches the canonical growth hormone receptor (GHR), a class I/type I cytokine receptor. Architecture includes an extracellular ligand-binding region with two FNIII-like modules, a single transmembrane helix, and an intracellular domain with Box1 and Box2 motifs important for JAK coupling. A domain schematic also identifies ECD ~19–262, TMD ~263–288, ICD ~289–638 in a conserved GHR framework. | (gun2022investigationofmonoclonal pages 5-9, gun2022investigationofmonoclonal media b3c4ed24, gun2022investigationofmonoclonal media 19db3114) | Gün, 2022, DOI: https://doi.org/10.17185/duepublico/46490 |
| Ligand / stoichiometry | The ligand is growth hormone (GH). Structural and mechanistic sources support 1 GH : 2 GHR binding, with site 1 binding one receptor first and site 2 engaging the second receptor. Reviews also describe the receptor:ligand complex as 2:1 GHR:GH. | (gun2022investigationofmonoclonal pages 13-18, wojcik2018postreceptorinhibitorsof pages 1-3, dehkhoda2018thegrowthhormone pages 2-5) | Dehkhoda, 2018, DOI: https://doi.org/10.3389/fendo.2018.00035; Wójcik, 2018, DOI: https://doi.org/10.3390/ijms19071843 |
| Activation mechanism | Older models proposed ligand-induced dimerization, but current understanding favors preformed GHR homodimers at the cell surface and even in the ER. GH binding induces a conformational rearrangement of the dimer rather than creating the dimer de novo. This reorientation separates/releases intracellular restraints so JAK2 molecules trans-phosphorylate and initiate signaling. | (dehkhoda2018thegrowthhormone pages 1-2, dehkhoda2018thegrowthhormone pages 2-5, ortiz2014…impactof pages 46-51) | Dehkhoda, 2018, DOI: https://doi.org/10.3389/fendo.2018.00035; Ortiz, 2014, no DOI in evidence |
| Key pathways | The dominant signaling output is JAK2 → STAT5a/STAT5b, with additional activation of STAT1 and STAT3. GHR also signals through MAPK/ERK via Shc/Grb2/SOS/Ras/Raf/MEK and through IRS-PI3K-Akt pathways. These pathways link GHR to growth, metabolism, and transcriptional regulation including IGF-1 production. | (cartersu2016growthhormonesignaling pages 5-8, cartersu2016growthhormonesignaling pages 1-5, dehkhoda2018thegrowthhormone pages 1-2, ortiz2014…impactof pages 46-51) | Carter-Su, 2016, DOI: https://doi.org/10.1016/j.ghir.2015.09.002; Dehkhoda, 2018, DOI: https://doi.org/10.3389/fendo.2018.00035 |
| Negative regulation | Multiple post-receptor brakes constrain signaling: SOCS2, SOCS1, SOCS3, CIS, PIAS, PTP1B, PTP-H1, SHP1, SHP2, and SIRPα1. SOCS2 is especially important: it binds phosphorylated GHR, helps recruit an E3 ubiquitin ligase complex, and promotes GHR internalization/degradation. Signaling is transient, with maximal activation around ~30 min and a refractory period of ~3–4 h in one review. | (fernandezperez2016growthhormonereceptor pages 4-7, wojcik2018postreceptorinhibitorsof pages 1-3) | Fernández-Pérez, 2016, DOI: https://doi.org/10.5772/64606; Wójcik, 2018, DOI: https://doi.org/10.3390/ijms19071843 |
| GH-binding protein (GHBP) | A soluble GH-binding protein (GHBP) corresponds to the extracellular domain of GHR and binds GH with receptor-like affinity. Literature and knockout genetics treat GHR and GHBP as products of the same gene system. GHBP modulates GH bioavailability in circulation, and schematic work highlights extracellular-domain shedding / soluble release. | (gun2022investigationofmonoclonal pages 9-13, list2001growthhormonereceptorbinding pages 1-3, gun2022investigationofmonoclonal pages 5-9) | List, 2001, DOI: https://doi.org/10.1006/mgme.2001.3164; Gün, 2022, DOI: https://doi.org/10.17185/duepublico/46490 |
| Localization / trafficking | GHR is primarily a plasma-membrane single-pass receptor, but preformed dimers are reported to assemble in the endoplasmic reticulum before surface expression. JAK2 can remain associated during internalization, suggesting some endosomal signaling. 2024 work also indicates ligand-dependent changes in surface availability and crosstalk with PRLR, with Box1/JAK2 coupling affecting receptor trafficking behavior. | (dehkhoda2018thegrowthhormone pages 2-5, wojcik2018postreceptorinhibitorsof pages 1-3, chhabra2024therolesof pages 1-2) | Wójcik, 2018, DOI: https://doi.org/10.3390/ijms19071843; Chen, 2024, DOI: https://doi.org/10.1101/2023.09.01.555812 |
| Rodent phenotypes / quantitative data | Rodent genetics show that loss of JAK2/STAT5 signaling strongly impairs postnatal growth. Liver-focused disruption of GH-GHR signaling can reduce circulating IGF-1 by ~80% or >90% depending on model, with metabolic effects including steatosis and insulin resistance. SOCS2 knockout mice are ~40% larger, underscoring negative regulation. GHR/BP knockout models also show undetectable serum IGF-I and elevated GH. | (dehkhoda2018thegrowthhormone pages 14-15, fernandezperez2016growthhormonereceptor pages 4-7, list2001growthhormonereceptorbinding pages 1-3) | Dehkhoda, 2018, DOI: https://doi.org/10.3389/fendo.2018.00035; Fernández-Pérez, 2016, DOI: https://doi.org/10.5772/64606; List, 2001, DOI: https://doi.org/10.1006/mgme.2001.3164 |
| Rat-specific / rodent signaling observations | In rat liver hepatocytes during early life, one study reported GH did not activate expected intracellular mediators, highlighting developmental context dependence of GHR responsiveness. Reviews also note that male rat GH secretion is pulsatile, shaping STAT5b activation differently from more continuous female patterns. Thus, rat Ghr function is conserved, but pathway output is strongly conditioned by age and endocrine context. | (fernandezperez2016growthhormonereceptor pages 4-7) | Fernández-Pérez, 2016, DOI: https://doi.org/10.5772/64606 |
| Recent 2024 developments | 2024 work refines GHR biology in two ways: (1) branch-specific signaling studies indicate Box1/JAK2/STAT5 and LYN/ERK outputs can be genetically uncoupled, with STAT5 being the key growth-driving branch; (2) super-resolution / membrane studies suggest JAK2 and Box1 are important for reciprocal surface regulation of GHR and PRLR. These studies update trafficking/crosstalk and pathway specificity rather than overturning the core GHR model. | (chhabra2024therolesof pages 1-2, chhabra2024therolesof pages 2-3) | Chhabra, 2024, DOI unavailable in evidence; Chen, 2024, DOI: https://doi.org/10.1101/2023.09.01.555812 |
Table: This table summarizes the verified identity, structure, signaling, regulation, localization, and phenotype evidence for rat Ghr/GHR. It is useful as a compact functional-annotation reference with direct context-ID support for each major claim.
Source URLs and publication dates (where available in retrieved texts)
- Carter-Su et al., “Growth hormone signaling pathways,” Growth Hormone & IGF Research, Jun 2016. https://doi.org/10.1016/j.ghir.2015.09.002 (cartersu2016growthhormonesignaling pages 1-5)
- Dehkhoda et al., “The growth hormone receptor: mechanism of receptor activation, cell signaling, and physiological aspects,” Frontiers in Endocrinology, Feb 2018. https://doi.org/10.3389/fendo.2018.00035 (dehkhoda2018thegrowthhormone pages 1-2)
- Wójcik et al., “Post-receptor inhibitors of the GHR-JAK2-STAT pathway…,” Int. J. Mol. Sci., Jun 2018. https://doi.org/10.3390/ijms19071843 (wojcik2018postreceptorinhibitorsof pages 1-3)
- Li et al., “Growth Hormone Did Not Activate… in Rats’ Liver… Early Life Period,” Int. J. Endocrinol. Metab., Published online Jun 23, 2018. https://doi.org/10.5812/ijem.61385 (ruonan2018growthhormonedid pages 1-2)
- List et al., “GHR/BP knockout mice: a 3-year update,” Molecular Genetics and Metabolism, May 2001. https://doi.org/10.1006/mgme.2001.3164 (list2001growthhormonereceptorbinding pages 1-3)
- Chhabra et al., “Roles of growth hormone–dependent JAK-STAT5 and Lyn kinase signaling…,” Endocrinology (advance access), Oct 2024. (URL shown in text) https://academic.oup.com/endo/advance-article/doi/10.1210/endocr/bqae136/7815814 (chhabra2024therolesofa pages 1-2)
- Chen et al., “A role for JAK2 in mediating cell surface GHR-PRLR interaction,” bioRxiv preprint posted Sep 5, 2023. https://doi.org/10.1101/2023.09.01.555812 (chen2024arolefor pages 1-3)
References
-
(gun2022investigationofmonoclonal pages 5-9): Investigation of monoclonal antibodies generated against the growth hormone receptor on growth hormone signaling This article has 0 citations and is from a peer-reviewed journal.
-
(gun2022investigationofmonoclonal media b3c4ed24): Investigation of monoclonal antibodies generated against the growth hormone receptor on growth hormone signaling This article has 0 citations and is from a peer-reviewed journal.
-
(gun2022investigationofmonoclonal media 19db3114): Investigation of monoclonal antibodies generated against the growth hormone receptor on growth hormone signaling This article has 0 citations and is from a peer-reviewed journal.
-
(gun2022investigationofmonoclonal pages 9-13): Investigation of monoclonal antibodies generated against the growth hormone receptor on growth hormone signaling This article has 0 citations and is from a peer-reviewed journal.
-
(list2001growthhormonereceptorbinding pages 1-3): Edward O. List, Karen T. Coschigano, and John J. Kopchick. Growth hormone receptor/binding protein (ghr/bp) knockout mice: a 3-year update. Molecular genetics and metabolism, 73 1:1-10, May 2001. URL: https://doi.org/10.1006/mgme.2001.3164, doi:10.1006/mgme.2001.3164. This article has 34 citations and is from a peer-reviewed journal.
-
(cartersu2016growthhormonesignaling pages 1-5): Christin Carter-Su, Jessica Schwartz, and Lawrence S. Argetsinger. Growth hormone signaling pathways. Growth hormone & IGF research : official journal of the Growth Hormone Research Society and the International IGF Research Society, 28:11-5, Jun 2016. URL: https://doi.org/10.1016/j.ghir.2015.09.002, doi:10.1016/j.ghir.2015.09.002. This article has 168 citations.
-
(dehkhoda2018thegrowthhormone pages 1-2): Farhad Dehkhoda, Christine M. M. Lee, Johan Medina, and Andrew J. Brooks. The growth hormone receptor: mechanism of receptor activation, cell signaling, and physiological aspects. Frontiers in Endocrinology, Feb 2018. URL: https://doi.org/10.3389/fendo.2018.00035, doi:10.3389/fendo.2018.00035. This article has 379 citations.
-
(ortiz2014…impactof pages 46-51): S Duran Ortiz. … impact of growth hormone on angiogenesis and other cellular pathways in subcutaneous and epididymal adipose tissue from wild type and bovine growth hormone …. Unknown journal, 2014.
-
(dehkhoda2018thegrowthhormone pages 2-5): Farhad Dehkhoda, Christine M. M. Lee, Johan Medina, and Andrew J. Brooks. The growth hormone receptor: mechanism of receptor activation, cell signaling, and physiological aspects. Frontiers in Endocrinology, Feb 2018. URL: https://doi.org/10.3389/fendo.2018.00035, doi:10.3389/fendo.2018.00035. This article has 379 citations.
-
(wojcik2018postreceptorinhibitorsof pages 1-3): Maciej Wójcik, Agata Krawczyńska, Hanna Antushevich, and Andrzej Przemysław Herman. Post-receptor inhibitors of the ghr-jak2-stat pathway in the growth hormone signal transduction. International Journal of Molecular Sciences, 19:1843, Jun 2018. URL: https://doi.org/10.3390/ijms19071843, doi:10.3390/ijms19071843. This article has 41 citations.
-
(cartersu2016growthhormonesignaling pages 5-8): Christin Carter-Su, Jessica Schwartz, and Lawrence S. Argetsinger. Growth hormone signaling pathways. Growth hormone & IGF research : official journal of the Growth Hormone Research Society and the International IGF Research Society, 28:11-5, Jun 2016. URL: https://doi.org/10.1016/j.ghir.2015.09.002, doi:10.1016/j.ghir.2015.09.002. This article has 168 citations.
-
(chhabra2024therolesof pages 1-2): Y Chhabra, H Bielefeldt-Ohmann, and TL Brooks. The roles of growth hormone dependent jak-stat5 and lyn kinase signalling in 1. Unknown journal, 2024.
-
(chhabra2024therolesofa pages 1-2): Y Chhabra, H Bielefeldt-Ohmann, and TL Brooks. The roles of growth hormone dependent jak-stat5 and lyn kinase signalling in 1. Unknown journal, 2024.
-
(chen2024arolefor pages 1-3): Chen Chen, Jing Jiang, Tejeshwar C. Rao, Ying Liu, Tatiana T. Marquez Lago, Stuart J. Frank, and André Leier. A role for jak2 in mediating cell surface ghr-prlr interaction. bioRxiv, Dec 2024. URL: https://doi.org/10.1101/2023.09.01.555812, doi:10.1101/2023.09.01.555812. This article has 0 citations.
-
(fernandezperez2016growthhormonereceptor pages 4-7): Leandro Fernández-Pérez, Amilcar Flores-Morales, Borja Guerra, Juan C. Díaz-Chico, and Diego Iglesias-Gato. Growth hormone receptor signaling pathways and its negative regulation by socs2. Restricted Growth - Clinical, Genetic and Molecular Aspects, Oct 2016. URL: https://doi.org/10.5772/64606, doi:10.5772/64606. This article has 5 citations.
-
(dehkhoda2018thegrowthhormone pages 14-15): Farhad Dehkhoda, Christine M. M. Lee, Johan Medina, and Andrew J. Brooks. The growth hormone receptor: mechanism of receptor activation, cell signaling, and physiological aspects. Frontiers in Endocrinology, Feb 2018. URL: https://doi.org/10.3389/fendo.2018.00035, doi:10.3389/fendo.2018.00035. This article has 379 citations.
-
(ruonan2018growthhormonedid pages 1-2): LRN Li RuoNan, HP Hong Pan, and LHN Lan HaiNan. Growth hormone did not activate its intracellular signaling molecules in rats’ liver hepatocytes during early life period. International Journal of Endocrinology and Metabolism, Jun 2018. URL: https://doi.org/10.5812/ijem.61385, doi:10.5812/ijem.61385. This article has 0 citations.
-
(gun2022investigationofmonoclonal pages 13-18): Investigation of monoclonal antibodies generated against the growth hormone receptor on growth hormone signaling This article has 0 citations and is from a peer-reviewed journal.
-
(chhabra2024therolesof pages 2-3): Y Chhabra, H Bielefeldt-Ohmann, and TL Brooks. The roles of growth hormone dependent jak-stat5 and lyn kinase signalling in 1. Unknown journal, 2024.
Artifacts
Citations
- dehkhoda2018thegrowthhormone pages 2-5
- wojcik2018postreceptorinhibitorsof pages 1-3
- chen2024arolefor pages 1-3
- fernandezperez2016growthhormonereceptor pages 4-7
- chhabra2024therolesofa pages 1-2
- list2001growthhormonereceptorbinding pages 1-3
- dehkhoda2018thegrowthhormone pages 14-15
- ruonan2018growthhormonedid pages 1-2
- cartersu2016growthhormonesignaling pages 1-5
- dehkhoda2018thegrowthhormone pages 1-2
- gun2022investigationofmonoclonal pages 5-9
- gun2022investigationofmonoclonal pages 9-13
- cartersu2016growthhormonesignaling pages 5-8
- chhabra2024therolesof pages 1-2
- gun2022investigationofmonoclonal pages 13-18
- chhabra2024therolesof pages 2-3
- https://doi.org/10.17185/duepublico/46490
- https://doi.org/10.3389/fendo.2018.00035;
- https://doi.org/10.3390/ijms19071843
- https://doi.org/10.1016/j.ghir.2015.09.002;
- https://doi.org/10.3389/fendo.2018.00035
- https://doi.org/10.5772/64606;
- https://doi.org/10.1006/mgme.2001.3164;
- https://doi.org/10.3390/ijms19071843;
- https://doi.org/10.1101/2023.09.01.555812
- https://doi.org/10.1006/mgme.2001.3164
- https://doi.org/10.5772/64606
- https://doi.org/10.1016/j.ghir.2015.09.002
- https://doi.org/10.5812/ijem.61385
- https://academic.oup.com/endo/advance-article/doi/10.1210/endocr/bqae136/7815814
- https://doi.org/10.1006/mgme.2001.3164,
- https://doi.org/10.1016/j.ghir.2015.09.002,
- https://doi.org/10.3389/fendo.2018.00035,
- https://doi.org/10.3390/ijms19071843,
- https://doi.org/10.1101/2023.09.01.555812,
- https://doi.org/10.5772/64606,
- https://doi.org/10.5812/ijem.61385,