Deep Research
Bioreason Pro
(lon-8-deep-research-bioreason-sft.md)
Bioreason Pro
(lon-8-deep-research-bioreason-sft.md)Thinking Trace
I begin with the domain architecture. The sequence contains IPR057449 (BPTI-like, C-terminal domain, residues 77–151). The BPTI/Kunitz fold is a compact, disulfide-rich inhibitory module classically used to block serine protease active sites. Its C-terminal placement implies an N-terminal signal peptide and pro-region preceding the Kunitz core, a layout that typically routes the protein into the secretory pathway and positions the inhibitory domain in the extracellular milieu. The disulfide-stabilized Kunitz scaffold is optimized for high-affinity, reversible protein–protein interactions rather than catalysis, and in extracellular matrices it often tunes proteolysis by restraining endopeptidases that would otherwise remodel matrix components.
This architecture causally supports a molecular function as an extracellular matrix structural constituent (GO:0005201). A secreted, disulfide-rich module can intercalate into matrix assemblies and provide mechanical stability and controlled viscoelasticity by modulating both direct binding to matrix proteins and the proteolytic turnover of those proteins. The Kunitz fold’s inhibitory geometry explains how the protein could regulate matrix remodeling enzymes, thereby preserving specific collagenous and glycoprotein architectures.
From this molecular role, the biological processes follow. By stabilizing and patterning the extracellular scaffold, the protein contributes to shaping specialized structures in the male tail, consistent with nematode male tail tip morphogenesis (GO:0045138). The same structural tuning can limit excessive body elongation by constraining extracellular matrix expansion and by signaling through a cell-surface receptor to temper cell proliferation, aligning with negative regulation of multicellular organism growth (GO:0040015). Mechanistically, a Kunitz-domain protein tethered to the matrix can both physically reinforce the cuticle and dampen protease-driven loosening, while its receptor engagement on hypodermal cells feeds back to restrain growth programs.
The cellular context is dictated by secretion and matrix integration. The Kunitz domain’s extracellular design, together with the protein’s role in morphogenesis and growth restraint, places it in the basement membrane (GO:0005604). Localization to the hypodermal basement membrane allows it to contact both apical extracellular matrix components and cell-surface receptors, enabling it to influence both matrix assembly and cell proliferation.
A coherent mechanistic model emerges: the protein is secreted by hypodermal cells into the basement membrane, where its BPTI-like domain binds and inhibits matrix-remodeling proteases, thereby stabilizing collagens and glycoproteins. In parallel, it engages a cell-surface receptor to trigger signaling that restrains body size and modulates proliferation. The observed partners—Col_cuticle_N domain-containing protein, ZP domain-containing protein, and cuticle collagen lon-3—fit this model as direct matrix ligands or co-scaffolds. The Kunitz module likely binds these extracellular components to form a protective network, while receptor engagement on adjacent cells transduces a growth-limiting signal. This dual action explains how the protein shapes the male tail tip and prevents overgrowth while remaining dispensable for the general integrity of the cuticle.
## Functional Summary
A secreted, basement-membrane-associated extracellular matrix protein in C. elegans that uses a disulfide-rich inhibitory module to stabilize and pattern the matrix while tempering proteolytic remodeling. By binding cuticle and glycoprotein components and engaging a cell-surface receptor, it shapes the male tail tip and limits body elongation by dampening proliferation. Despite reinforcing local extracellular architecture, it is not essential for overall cuticle integrity.
## UniProt Summary
Involved in male tail morphogenesis. Negatively regulates body size, probably by modulating cell proliferation through interaction with one or more cell surface receptors. Not required for the structural integrity of the cuticle.
## InterPro Domains
- IPR057449: BPTI-like, C-terminal domain (domain) [77-151]
## GO Term Predictions
### Molecular Function
### Biological Process
### Cellular Component
Falcon
(lon-8-deep-research-falcon.md)
Falcon
(lon-8-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research report: Caenorhabditis elegans lon-8 (UniProt G5EGH7; ORF Y59A8B.20)
0) Scope, identity verification, and ambiguity check
The literature retrieved and analyzed here consistently uses lon-8 to refer to the C. elegans locus Y59A8B.20, originally defined by Soete, Betist, and Korswagen (2007). In that primary description, Y59A8B.20 was renamed lon-8 based on long-body (Lon) and male tail phenotypes, and the gene was shown to encode a small secreted precursor protein (signal peptide-bearing) expressed in the hypodermis. (soete2007regulationofcaenorhabditis pages 1-2)
Within the retrieved corpus, no alternative “lon-8” gene in another organism was found to be conflated with the C. elegans gene; downstream mentions of LON-8 occur in the context of apical extracellular matrices (aECM)/cuticle and male rays. (soete2007regulationofcaenorhabditis pages 8-9, ragle2025multiscalepatterningof pages 12-15)
Important limitation: Although the user provided UniProt/InterPro domain context (BPTI_nem/BPTI_C_nem) for UniProt G5EGH7, those database pages were not directly retrievable with the current tools, and no domain-focused primary papers were retrieved that explicitly connect those domain identifiers to LON-8. Therefore, mechanistic statements about “BPTI/Kunitz-like protease inhibitor activity” are treated as hypotheses rather than evidence-based claims in this report.
1) Key concepts and definitions (current understanding)
1.1 What is lon-8?
lon-8 encodes LON-8, a small (~162 aa) protein with an N-terminal signal peptide, consistent with a secreted protein/peptide produced by the hypodermis (epidermis). (soete2007regulationofcaenorhabditis pages 1-2)
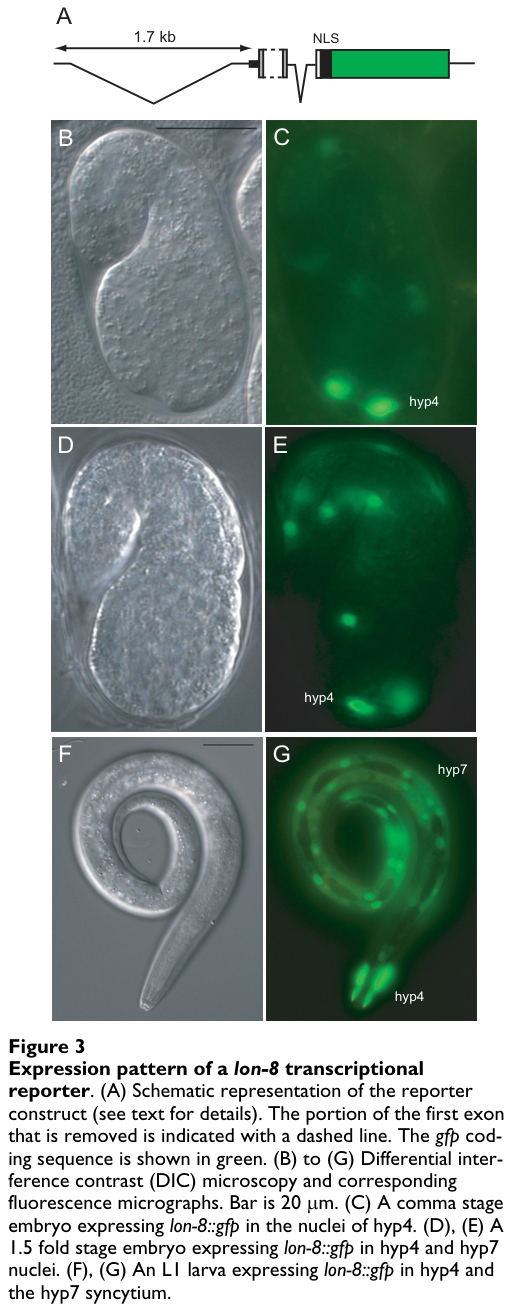
In reporter assays, inclusion of the N-terminal signal peptide caused GFP to appear secreted/diffuse; clearer cell-associated reporter localization required deleting the signal peptide from the fusion construct, consistent with secretion of the native protein. (soete2007regulationofcaenorhabditis pages 4-6)
1.2 Biological context: cuticle and apical extracellular matrix (aECM)
The C. elegans cuticle is an apical extracellular matrix (aECM) secreted by the hypodermis and specialized epithelia. In an aECM-focused review, LON-8 is discussed among proteins important for building male rays, placing it within the broader conceptual framework of aECM components shaping epithelial structures. (soete2007regulationofcaenorhabditis pages 1-2)
1.3 Phenotype terms used in this gene’s characterization
- Lon (long body size): Animals are longer than wild-type.
- Ram-like male ray defects: Male tail sensory rays exhibit abnormal morphology. lon-8 mutants show extensive ray deformations without the ray fate/misspecification typical of some DBL-1/Sma/Mab pathway mutants. (soete2007regulationofcaenorhabditis pages 9-11, soete2007regulationofcaenorhabditis pages 6-8)
2) Experimentally supported functions of LON-8
2.1 Primary function: regulation of post-embryonic body size (larval elongation/early adult growth)
Soete et al. (2007; BMC Developmental Biology, published 2007-03, DOI 10.1186/1471-213X-7-20, URL https://doi.org/10.1186/1471-213x-7-20) showed that reducing lon-8 activity increases body length (Lon phenotype). (soete2007regulationofcaenorhabditis pages 1-2, soete2007regulationofcaenorhabditis pages 2-4)
Quantitative examples from their assays include:
* RNAi against lon-8 increased body length from 1.3 ± 0.08 mm (control) to 1.4 ± 0.08 mm (n = 75). (soete2007regulationofcaenorhabditis pages 1-2)
* lon-8 mutants were described as ~10% longer than wild type, smaller effect than lon-1 (~30%) or lon-3 (~25%), with effects mainly during larval and early adult stages. (soete2007regulationofcaenorhabditis pages 8-9)
Mechanistically, they tested whether size increase reflected more epidermal nuclei or higher epidermal ploidy and found no strong support:
* Hypodermal nuclei counts: 21 ± 1 (n=20), no change reported. (soete2007regulationofcaenorhabditis pages 2-4)
* Seam cell counts: 16–17 seam cell nuclei (n=32), no change reported. (soete2007regulationofcaenorhabditis pages 2-4)
* Hypodermal ploidy: 7.9 ± 1.6 (N2) vs 8.2 ± 2.4 (lon-8(hu188); n≈10), not detectably changed. (soete2007regulationofcaenorhabditis pages 2-4)
These data support a model in which LON-8 influences body shape/length via extracellular matrix (cuticle) properties and/or cell size, rather than by increasing epidermal cell number or nuclear DNA content. (soete2007regulationofcaenorhabditis pages 2-4, soete2007regulationofcaenorhabditis pages 9-11)
2.2 Role in male tail development (ray morphology)
A distinguishing aspect of lon-8 among “lon” genes is its effect on male tail/ray morphogenesis. lon-8 deletion mutants hu187 and hu188 show widespread, variable ray defects (Ram-like), with rays 5 and 6 most severely affected; males remain capable of mating. (soete2007regulationofcaenorhabditis pages 6-8)
The phenotype is visually documented in Soete et al. figures (wild-type vs lon-8 mutant male tail). (soete2007regulationofcaenorhabditis media 8670cd25, soete2007regulationofcaenorhabditis media ae92ca98)
3) Expression pattern and inferred site of action
3.1 Hypodermal expression throughout development
lon-8 is strongly expressed in hypodermal syncytia (notably hyp4 and hyp7) from embryonic stages (comma stage) through larval stages and adulthood based on reporter analysis. (soete2007regulationofcaenorhabditis pages 4-6)
A cropped figure panel shows hypodermal reporter expression in an L1 larva (hyp7 and hyp4 nuclei). (soete2007regulationofcaenorhabditis media 8670cd25)
3.2 Male tail hypodermis surrounding developing rays
lon-8::gfp reporter expression is seen in specific nuclei contributing to the ventral male tail hypodermis surrounding developing rays (cell identities enumerated in the original paper). (soete2007regulationofcaenorhabditis pages 8-9, soete2007regulationofcaenorhabditis pages 6-8)
Corresponding figure panels show reporter expression in male tail hypodermal cells, supporting a local hypodermal source for the secreted product relevant to ray morphogenesis. (soete2007regulationofcaenorhabditis media ae92ca98)
4) Genetic pathway placement and interactions
4.1 Relationship to DBL-1/Sma/Mab (TGF-β) pathway
Multiple lines of evidence suggest lon-8 is not a direct transcriptional target of the DBL-1/Sma/Mab pathway and functions in parallel/independently for body size control:
* lon-8 expression was not detectably altered in dbl-1 or sma-6 mutants in reporter and/or transcript assays. (soete2007regulationofcaenorhabditis pages 4-6, soete2007regulationofcaenorhabditis pages 6-8)
* lon-8 does not suppress the strong Sma phenotype of dbl-1 or sma-6 mutants; double mutants remain strongly Sma. (soete2007regulationofcaenorhabditis pages 4-6)
However, dbl-1 can genetically suppress the lon-8 body-size phenotype, consistent with partially convergent control of growth through distinct effectors. (soete2007regulationofcaenorhabditis pages 8-9)
4.2 Interactions with cuticle/collagen-modifying enzymes dpy-11 and dpy-18
The most informative genetic interactions reported are with dpy-11 and dpy-18, which encode enzymes involved in cuticle collagen modification. RNAi of either gene completely suppresses the lon-8 Lon phenotype, implying lon-8’s effect on body size is mediated through cuticle/collagen-related processes. (soete2007regulationofcaenorhabditis pages 1-2, soete2007regulationofcaenorhabditis pages 6-8)
This interaction is consistent with LON-8 acting as a secreted cuticle/aECM factor that modulates cuticle organization, the accessibility of collagen-modifying enzyme substrates, or the mechanical properties of the extracellular matrix that determine body length. (soete2007regulationofcaenorhabditis pages 9-11)
5) Recent developments and latest research (prioritizing 2023–2024)
5.1 Availability of recent lon-8-focused studies
A targeted search for 2023–2024 primary studies specifically dissecting lon-8 did not yield relevant lon-8-focused mechanistic papers in the retrieved set. This likely reflects that lon-8 remains a comparatively under-studied, small secreted factor with a limited direct literature footprint.
5.2 Most recent authoritative context available in this corpus: aECM review and newer tagging resources
Two more recent sources in the retrieved corpus are useful:
-
aECM review context (2020): Cohen & Sundaram (2020; Journal of Developmental Biology, published 2020-10, DOI 10.3390/jdb8040023, URL https://doi.org/10.3390/jdb8040023) place LON-8 among proteins important for building male rays within the conceptual framework of apical extracellular matrices. (soete2007regulationofcaenorhabditis pages 1-2)
-
Endogenous tagging resource (preprint 2025): Ragle et al. (2025; bioRxiv, published 2025-05, DOI 10.1101/2025.05.14.653803, URL https://doi.org/10.1101/2025.05.14.653803) report systematic endogenous tagging of aECM components including LON-8::mNG, describing LON-8 as a secreted protein implicated in cuticle morphology. They further report localization to cuticle structures including the male tail fan, and note atypical FRAP recovery for LON-8::mNG in the L4 vulva relative to most knock-ins. (ragle2025multiscalepatterningof pages 12-15)
Although this preprint is outside 2024, it represents the most recent methodological “real-world implementation” in the retrieved corpus that directly enables improved functional annotation of LON-8.
6) Current applications and real-world implementations
6.1 As a genetic handle on cuticle-dependent morphogenesis
lon-8 mutants provide a tractable genotype to study how secreted epidermal factors influence:
* Whole-body size/shape (growth/elongation) (soete2007regulationofcaenorhabditis pages 1-2)
* Male tail ray morphogenesis and aECM patterning (soete2007regulationofcaenorhabditis pages 6-8)
Because body length changes are modest (~10%) and not due to obvious hypodermal cell number/ploidy changes, lon-8 can be used to probe ECM mechanics/assembly in a relatively specific manner compared to broader growth pathways. (soete2007regulationofcaenorhabditis pages 2-4)
6.2 Imaging and localization resources
Two complementary “implementation” styles are demonstrated:
* Transcriptional reporter lon-8::gfp (extrachromosomal), used to map expression in hypodermis and male tail hypodermis. (soete2007regulationofcaenorhabditis pages 11-12, soete2007regulationofcaenorhabditis media ae92ca98)
* Endogenous knock-in LON-8::mNG (Ragle et al.), which can support higher-fidelity localization dynamics and comparative aECM mapping across tissues (e.g., male tail fan, alae). (ragle2025multiscalepatterningof pages 12-15)
7) Expert opinions / analysis grounded in authoritative sources
7.1 What authoritative sources imply about mechanism
The strongest interpretation supported by experimental genetics is that LON-8 is a secreted hypodermal factor acting on the cuticle/aECM, influencing body length and specialized cuticular structures (male rays). This is supported by its signal peptide, hypodermal expression, and genetic suppression by collagen-modifying enzyme knockdowns (dpy-11, dpy-18). (soete2007regulationofcaenorhabditis pages 1-2, soete2007regulationofcaenorhabditis pages 6-8)
A reasonable working model is that LON-8 modulates cuticle composition or architecture, thereby affecting organismal elongation and the mechanical extension/morphogenesis of male rays. The epistasis/suppression pattern is consistent with a mechanism involving collagen post-translational modification pathways, although the biochemical mode of action remains undefined in the retrieved literature. (soete2007regulationofcaenorhabditis pages 9-11)
7.2 Domain-based hypotheses (explicitly labeled as hypotheses)
The user-provided UniProt context indicates BPTI-like nematode domains (BPTI_nem/BPTI_C_nem). If correct, this could suggest LON-8 is structurally related to small cysteine-rich inhibitors (e.g., protease inhibitors). However, no retrieved primary sources directly connect LON-8 to protease inhibition biochemistry, nor were substrates or enzymatic reactions reported. Therefore, any “protease inhibitor” mechanism should be treated as speculative until supported by biochemical assays or domain-confirming sources.
8) Key quantitative/statistical results (from primary studies)
Selected quantitative statistics from Soete et al. (2007):
* Body length (RNAi): 1.3 ± 0.08 mm control vs 1.4 ± 0.08 mm lon-8 RNAi (n=75). (soete2007regulationofcaenorhabditis pages 1-2)
* Hypodermal nuclei: 21 ± 1 (n=20), no change reported for lon-8 mutants. (soete2007regulationofcaenorhabditis pages 2-4)
* Seam cell nuclei: 16–17 (n=32), no change reported. (soete2007regulationofcaenorhabditis pages 2-4)
* Hypodermal ploidy: 7.9 ± 1.6 (N2) vs 8.2 ± 2.4 (lon-8(hu188); n≈10). (soete2007regulationofcaenorhabditis pages 2-4)
9) Evidence map (summary table)
The following table consolidates evidence claims, experimental types, quantitative values, and sources.
| Claim/annotation | Evidence type (genetics/expression/quantitative/resource) | Key details (numbers, tissues, alleles, interactions) | Source (author year, venue, DOI/URL) |
|---|---|---|---|
| Identity mapping: lon-8 = Y59A8B.20 in C. elegans; encodes a small secreted precursor | genetics/expression | ORF Y59A8B.20 named lon-8; predicted 162 aa protein with N-terminal signal peptide; no TM/lipid anchor noted; one deletion allele (hu187) yields alternative transcript predicted to lack the signal peptide (soete2007regulationofcaenorhabditis pages 1-2, soete2007regulationofcaenorhabditis pages 2-4) | Soete et al. 2007, BMC Developmental Biology; DOI: https://doi.org/10.1186/1471-213X-7-20 |
| Primary function: regulator of body size/larval elongation | genetics/quantitative | RNAi against Y59A8B.20 increased body length from 1.3 ± 0.08 mm to 1.4 ± 0.08 mm (n = 75); deletion alleles lon-8(hu187) and lon-8(hu188) produce clear Lon phenotype; mutants are reported as about ~10% longer than wild type (soete2007regulationofcaenorhabditis pages 1-2, soete2007regulationofcaenorhabditis pages 8-9) | Soete et al. 2007, BMC Developmental Biology; DOI: https://doi.org/10.1186/1471-213X-7-20 |
| Cellular localization / site of action: hypodermis-derived, likely extracellular/cuticular | expression | Strong reporter expression in hypodermal syncytium, especially hyp4 and hyp7; clear cellular localization required deletion of signal peptide from reporter because native N-terminus could cause secretion/diffusion of GFP; supports extracellular secretion from epidermis (soete2007regulationofcaenorhabditis pages 1-2, soete2007regulationofcaenorhabditis pages 4-6, soete2007regulationofcaenorhabditis media 8670cd25) | Soete et al. 2007, BMC Developmental Biology; DOI: https://doi.org/10.1186/1471-213X-7-20 |
| Male tail / ray morphogenesis role | genetics/expression | lon-8(hu187) and lon-8(hu188) males show Ram-like defects; all rays abnormal, with rays 5 and 6 most severe; rays can appear clumped as an amorphous mass; males remain able to mate; reporter expressed in ventral male tail hypodermis around developing rays, including V6.pppaa, T.aa, R6.p, T.apapa, R7.p, R8.p, R9.p (soete2007regulationofcaenorhabditis pages 6-8, soete2007regulationofcaenorhabditis pages 8-9, soete2007regulationofcaenorhabditis media 8670cd25) | Soete et al. 2007, BMC Developmental Biology; DOI: https://doi.org/10.1186/1471-213X-7-20 |
| Pathway placement: acts outside direct transcriptional control of DBL-1/Sma/Mab (TGF-β) | genetics/expression | lon-8 expression not detectably changed in dbl-1 or sma-6 mutants; dbl-1 suppresses the lon-8 body-size phenotype, but data support action in parallel/independent rather than as a direct DBL-1 transcriptional target (soete2007regulationofcaenorhabditis pages 8-9, soete2007regulationofcaenorhabditis pages 4-6, soete2007regulationofcaenorhabditis pages 6-8) | Soete et al. 2007, BMC Developmental Biology; DOI: https://doi.org/10.1186/1471-213X-7-20 |
| Cuticle/aECM association via collagen-modifying enzymes | genetics | dpy-11(RNAi) and dpy-18(RNAi) completely suppress the lon-8 body-size phenotype; these genes encode cuticle collagen modifying enzymes, implicating LON-8 in cuticle composition/assembly or regulation of cuticle-associated targets (soete2007regulationofcaenorhabditis pages 1-2, soete2007regulationofcaenorhabditis pages 9-11, soete2007regulationofcaenorhabditis pages 6-8) | Soete et al. 2007, BMC Developmental Biology; DOI: https://doi.org/10.1186/1471-213X-7-20 |
| Mechanistic constraint: Lon phenotype is not explained by increased hypodermal cell number or ploidy | quantitative | Hypodermal nuclei counts unchanged (21 ± 1, n = 20); seam cell nuclei unchanged (16–17, n = 32); hypodermal ploidy not detectably changed (7.9 ± 1.6 in N2 vs 8.2 ± 2.4 in lon-8(hu188), n ≈ 10); supports a model involving cell size / matrix properties rather than extra cells (soete2007regulationofcaenorhabditis pages 2-4) | Soete et al. 2007, BMC Developmental Biology; DOI: https://doi.org/10.1186/1471-213X-7-20 |
| Evolutionary context: conserved in rhabditid nematodes | expression/quantitative | Homologs reported across Rhabditida; N-termini are divergent but consistently predicted to be signal peptides, reinforcing conserved secretion/extracellular localization (soete2007regulationofcaenorhabditis pages 9-11, soete2007regulationofcaenorhabditis pages 8-9) | Soete et al. 2007, BMC Developmental Biology; DOI: https://doi.org/10.1186/1471-213X-7-20 |
| Review-level annotation: secreted aECM/cuticle factor important for male rays | resource | Review on C. elegans apical extracellular matrices lists LON-8 among proteins important for building male rays, describing it as a short, secreted peptide/protein in the ray/cuticle context (soete2007regulationofcaenorhabditis pages 1-2) | Cohen & Sundaram 2020, Journal of Developmental Biology; DOI: https://doi.org/10.3390/jdb8040023 |
| Recent resource / real-world implementation: endogenous fluorescent tagging | resource | Systematic aECM study generated an endogenous lon-8::mNG knock-in, explicitly describing LON-8 as a novel secreted protein implicated in body cuticle morphology; useful for in vivo localization and functional follow-up (ragle2025multiscalepatterningof pages 9-12) | Ragle et al. 2025, bioRxiv; DOI: https://doi.org/10.1101/2025.05.14.653803 |
Table: This table summarizes the strongest functional annotation evidence for C. elegans lon-8/Y59A8B.20, including secretion, hypodermal expression, body-size and male-tail phenotypes, pathway relationships, and recent tagging resources. It is useful as a compact evidence map linking specific claims to quantitative results and source citations.
10) Visual evidence excerpts
Soete et al. (2007) figure crops support (i) hypodermal expression and (ii) male tail ray morphological defects in lon-8 mutants. (soete2007regulationofcaenorhabditis media 8670cd25, soete2007regulationofcaenorhabditis media ae92ca98)
11) Conclusions and research gaps
Conclusions (supported): lon-8/Y59A8B.20 encodes a small secreted hypodermal protein that functions in the extracellular cuticle/aECM to regulate larval elongation/body size and male ray morphology, acting largely independently of DBL-1/Sma/Mab transcriptional control but genetically interacting with collagen-modifying enzymes dpy-11 and dpy-18. (soete2007regulationofcaenorhabditis pages 1-2, soete2007regulationofcaenorhabditis pages 6-8)
Major gaps (not resolved by retrieved literature):
* Direct biochemical function (binding partners, substrates, enzymatic targets) and whether LON-8 has protease-inhibitor activity.
* Domain confirmation and structure–function mapping tied to BPTI_nem/BPTI_C_nem annotations.
* Updated (2023–2024) mechanistic studies of lon-8 specifically.
Actionable next steps enabled by recent resources: Endogenous LON-8::mNG strains and systematic aECM tagging approaches should facilitate tissue-resolved localization, dynamics (e.g., FRAP), and genetic interaction mapping to uncover mechanism and pathway integration. (ragle2025multiscalepatterningof pages 12-15)
References
-
(soete2007regulationofcaenorhabditis pages 1-2): Gwen Soete, Marco C Betist, and Hendrik C Korswagen. Regulation of caenorhabditis elegans body size and male tail development by the novel gene lon-8. BMC Developmental Biology, 7:20-20, Mar 2007. URL: https://doi.org/10.1186/1471-213x-7-20, doi:10.1186/1471-213x-7-20. This article has 24 citations and is from a peer-reviewed journal.
-
(soete2007regulationofcaenorhabditis pages 8-9): Gwen Soete, Marco C Betist, and Hendrik C Korswagen. Regulation of caenorhabditis elegans body size and male tail development by the novel gene lon-8. BMC Developmental Biology, 7:20-20, Mar 2007. URL: https://doi.org/10.1186/1471-213x-7-20, doi:10.1186/1471-213x-7-20. This article has 24 citations and is from a peer-reviewed journal.
-
(ragle2025multiscalepatterningof pages 12-15): James Matthew Ragle, Murugesan Pooranachithra, Guinevere E. Ashley, Emma Cadena, Batia Blank, Katelyn Kang, Cincy Chen, Aditree R. Bhowmick, Soraya H. Mercado, Tabatha E. Wells, John C. Clancy, Andrew D. Chisholm, and Jordan D. Ward. Multiscale patterning of a model apical extracellular matrix revealed by systematic endogenous protein tagging. BioRxiv, May 2025. URL: https://doi.org/10.1101/2025.05.14.653803, doi:10.1101/2025.05.14.653803. This article has 10 citations.
-
(soete2007regulationofcaenorhabditis pages 4-6): Gwen Soete, Marco C Betist, and Hendrik C Korswagen. Regulation of caenorhabditis elegans body size and male tail development by the novel gene lon-8. BMC Developmental Biology, 7:20-20, Mar 2007. URL: https://doi.org/10.1186/1471-213x-7-20, doi:10.1186/1471-213x-7-20. This article has 24 citations and is from a peer-reviewed journal.
-
(soete2007regulationofcaenorhabditis pages 9-11): Gwen Soete, Marco C Betist, and Hendrik C Korswagen. Regulation of caenorhabditis elegans body size and male tail development by the novel gene lon-8. BMC Developmental Biology, 7:20-20, Mar 2007. URL: https://doi.org/10.1186/1471-213x-7-20, doi:10.1186/1471-213x-7-20. This article has 24 citations and is from a peer-reviewed journal.
-
(soete2007regulationofcaenorhabditis pages 6-8): Gwen Soete, Marco C Betist, and Hendrik C Korswagen. Regulation of caenorhabditis elegans body size and male tail development by the novel gene lon-8. BMC Developmental Biology, 7:20-20, Mar 2007. URL: https://doi.org/10.1186/1471-213x-7-20, doi:10.1186/1471-213x-7-20. This article has 24 citations and is from a peer-reviewed journal.
-
(soete2007regulationofcaenorhabditis pages 2-4): Gwen Soete, Marco C Betist, and Hendrik C Korswagen. Regulation of caenorhabditis elegans body size and male tail development by the novel gene lon-8. BMC Developmental Biology, 7:20-20, Mar 2007. URL: https://doi.org/10.1186/1471-213x-7-20, doi:10.1186/1471-213x-7-20. This article has 24 citations and is from a peer-reviewed journal.
-
(soete2007regulationofcaenorhabditis media 8670cd25): Gwen Soete, Marco C Betist, and Hendrik C Korswagen. Regulation of caenorhabditis elegans body size and male tail development by the novel gene lon-8. BMC Developmental Biology, 7:20-20, Mar 2007. URL: https://doi.org/10.1186/1471-213x-7-20, doi:10.1186/1471-213x-7-20. This article has 24 citations and is from a peer-reviewed journal.
-
(soete2007regulationofcaenorhabditis media ae92ca98): Gwen Soete, Marco C Betist, and Hendrik C Korswagen. Regulation of caenorhabditis elegans body size and male tail development by the novel gene lon-8. BMC Developmental Biology, 7:20-20, Mar 2007. URL: https://doi.org/10.1186/1471-213x-7-20, doi:10.1186/1471-213x-7-20. This article has 24 citations and is from a peer-reviewed journal.
-
(soete2007regulationofcaenorhabditis pages 11-12): Gwen Soete, Marco C Betist, and Hendrik C Korswagen. Regulation of caenorhabditis elegans body size and male tail development by the novel gene lon-8. BMC Developmental Biology, 7:20-20, Mar 2007. URL: https://doi.org/10.1186/1471-213x-7-20, doi:10.1186/1471-213x-7-20. This article has 24 citations and is from a peer-reviewed journal.
-
(ragle2025multiscalepatterningof pages 9-12): James Matthew Ragle, Murugesan Pooranachithra, Guinevere E. Ashley, Emma Cadena, Batia Blank, Katelyn Kang, Cincy Chen, Aditree R. Bhowmick, Soraya H. Mercado, Tabatha E. Wells, John C. Clancy, Andrew D. Chisholm, and Jordan D. Ward. Multiscale patterning of a model apical extracellular matrix revealed by systematic endogenous protein tagging. BioRxiv, May 2025. URL: https://doi.org/10.1101/2025.05.14.653803, doi:10.1101/2025.05.14.653803. This article has 10 citations.
Artifacts
Citations
- soete2007regulationofcaenorhabditis pages 1-2
- soete2007regulationofcaenorhabditis pages 4-6
- soete2007regulationofcaenorhabditis pages 8-9
- soete2007regulationofcaenorhabditis pages 2-4
- soete2007regulationofcaenorhabditis pages 6-8
- soete2007regulationofcaenorhabditis pages 9-11
- ragle2025multiscalepatterningof pages 12-15
- ragle2025multiscalepatterningof pages 9-12
- soete2007regulationofcaenorhabditis pages 11-12
- https://doi.org/10.1186/1471-213x-7-20
- https://doi.org/10.3390/jdb8040023
- https://doi.org/10.1101/2025.05.14.653803
- https://doi.org/10.1186/1471-213X-7-20
- https://doi.org/10.1186/1471-213x-7-20,
- https://doi.org/10.1101/2025.05.14.653803,