Deep Research
Falcon
(ASF1-deep-research-falcon.md)
Falcon
(ASF1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: ASF1 (CIA1/YJL115W; UniProt P32447) in Saccharomyces cerevisiae (S288c)
1) Target identity verification (critical disambiguation)
The target is Saccharomyces cerevisiae Asf1, encoded by ASF1 (synonym CIA1; locus YJL115W) and annotated in UniProt as P32447, a member of the conserved ASF1 family and a histone H3–H4 chaperone. The evidence used below consistently refers to yeast Asf1/Asf1p in chromatin assembly and histone-modification pathways (e.g., HIR, CAF-1, Rtt106, Rtt109), matching the UniProt-provided identity and domain/function expectations. (kim2024structureofthe pages 29-30, hiraga2008histoneh3lysine pages 4-5)
2) Key concepts and definitions (current understanding)
2.1 Histone chaperone (definition in this context)
In yeast, Asf1 is a histone chaperone that binds histones H3–H4 and participates in nucleosome assembly in multiple DNA-templated processes, including DNA replication and transcription-associated chromatin remodeling. (hiraga2008histoneh3lysine pages 4-5)
2.2 Molecular function of Asf1
Core chaperone activity: Asf1 binds H3–H4 dimers and shields the surface needed to form a stable (H3–H4)2 tetramer, thereby controlling histone oligomerization state during handling/transport and handoff. Although the key mechanistic description is discussed in a conserved (yeast-comparative) context, it is explicitly connected to budding yeast functional biology (including H3K56 acetylation) in the cited work. (ding2025theorthologof pages 1-2)
Structural/interaction basis: The conserved N-terminal region forms a globular core with acidic patches that contact histone H3 (C-terminal region) and can also bind H4, consistent with Asf1’s role as an H3–H4 chaperone. (breuer2024histonebindingof pages 1-2)
3) Primary pathways and biological processes in budding yeast
3.1 Replication-coupled chromatin assembly via the H3K56ac pathway (Asf1 → Rtt109 → CAF-1/Rtt106)
A central experimentally grounded pathway in budding yeast is the Asf1-dependent acetylation of histone H3 lysine 56 (H3K56ac) by the acetyltransferase Rtt109, followed by transfer of acetylated H3–H4 to deposition factors.
Mechanistic model (handoff cascade):
- Asf1 binds newly synthesized H3–H4 (described as occurring in the cytoplasm in one mechanistic account), promotes nuclear import/availability, and presents H3–H4 to Rtt109 for acetylation of H3K56. (kattarmell2012regulationofhistone pages 10-17)
- After H3K56 acetylation, H3–H4 are transferred to downstream histone chaperones, including CAF-1 (replication/repair-coupled deposition) and Rtt106 (another H3–H4 chaperone involved in chromatin assembly). (dannah2024novelinsightsinto pages 67-72, kattarmell2012regulationofhistone pages 10-17)
- Asf1 is described as solely required for H3K56ac in yeast cells in the Dannah thesis excerpts, and loss of Asf1 markedly decreases Rtt109 activity and H3K56ac. (dannah2024novelinsightsinto pages 56-60)
CAF-1 physical link: CAF-1 is reported to accept H3–H4 via direct interaction between CAF-1 subunit Cac2 and Asf1 in one mechanistic account. (kattarmell2012regulationofhistone pages 10-17)
3.2 Replication-independent H3–H4 deposition and transcriptional chromatin regulation (Asf1 with HIR/Rtt106)
A distinct major axis of Asf1 biology is replication-independent deposition and promoter-associated chromatin regulation.
A 2024 Molecular Cell study supports that Asf1 works together with the HIR complex (and with Rtt106) to mediate replication-independent H3–H4 deposition and maintain promoter fidelity, connecting Asf1 to transcriptional regulation through chromatin assembly/disassembly dynamics. (kim2024structureofthe pages 29-30)
3.3 Histone gene transcription regulation (cell-cycle coupling)
In yeast-focused statements summarized in a 2024 preprint, Asf1 is described as participating in histone gene transcriptional activation in S phase and transcriptional repression outside S phase in combination with Hir1 (a yeast counterpart of metazoan HIRA pathway components). (mendiratta2024regulationofreplicative pages 1-6)
4) Subcellular localization: where Asf1 acts
4.1 Nuclear localization and nuclear import determinants
A 2024 thesis focused on chromatin assembly metabolism in S. cerevisiae reports that Asf1 contains a functional classical nuclear localization signal (cNLS) in its highly acidic C-terminal tail, and that removal of this motif makes Asf1 fully cytoplasmic, indicating the motif is required for nuclear localization. (dannah2024novelinsightsinto pages 166-170)
4.2 Localization-function coupling (link to H3K56ac)
The same thesis reports that deleting the Asf1 C-terminal cNLS reduces H3K56 acetylation when VPS75 is present and abolishes H3K56 acetylation when VPS75 is absent, connecting nuclear localization competence to full H3K56ac pathway output. (dannah2024novelinsightsinto pages 166-170)
5) Quantitative phenotypes and statistics from experimental studies
5.1 Chromosome positioning/telomere localization (quantitative)
A key quantitative phenotype connecting Asf1 to replication-associated chromatin state transmission is telomere peripheral localization.
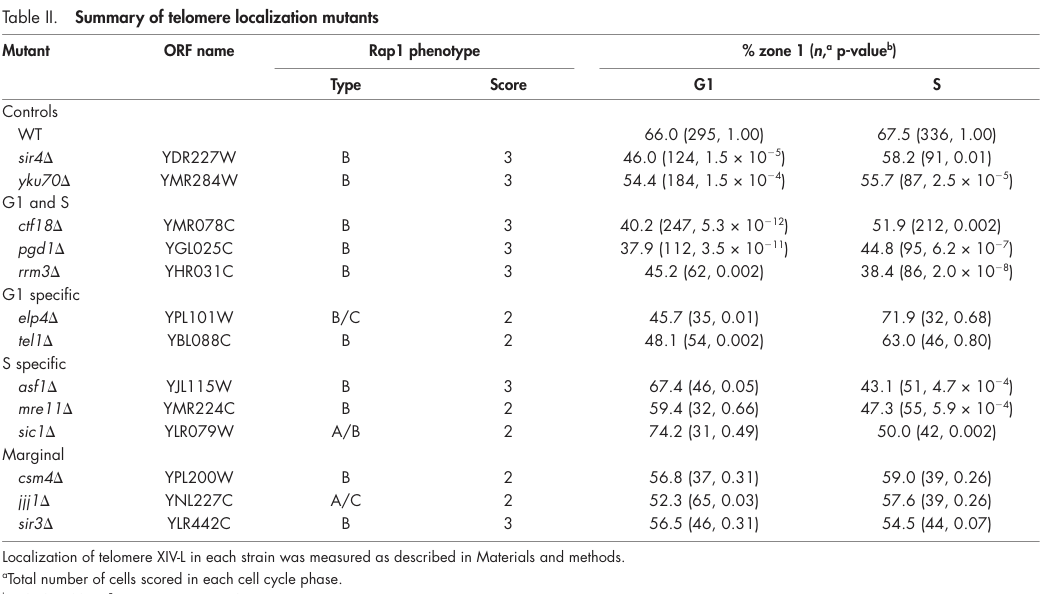
In a telomere XIV-L positioning assay, WT cells show ~66.0% peripheral localization (zone 1) in G1 and 67.5% in S phase. In contrast, an asf1 mutant shows 67.4% in G1 (n=46; p=0.05 vs WT) but drops to 43.1% in S phase (n=51; p=4.7×10−4 vs WT), demonstrating a statistically significant S-phase-specific defect. (hiraga2008histoneh3lysine pages 4-5, hiraga2008histoneh3lysine media eba6b4e4)
The same study supports that proper chromosome positioning depends on a regulated H3K56 acetylation/deacetylation cycle, since both non-acetylatable (H3K56R) and acetyl-mimic (H3K56Q) mutations strongly disrupt telomere localization. (hiraga2008histoneh3lysine pages 5-7)
5.2 Additional chromatin domain positioning phenotype
Asf1 is also required for perinuclear localization of the ETC6 chromatin domain during interphase, and this positioning depends on Rtt109/H3K56 acetylation, linking Asf1-mediated histone modification to higher-order chromosomal organization. (hiraga2008histoneh3lysine pages 7-8)
6) Recent developments (prioritizing 2023–2024)
6.1 2024 Molecular Cell: mechanistic/structural advance for HIR-linked deposition
Recent structural and functional work on the HIR histone chaperone complex provides updated mechanistic support for how Asf1 interfaces with replication-independent H3–H4 deposition and transcription-associated promoter fidelity pathways in yeast. (Kim et al., 2024; publication July 2024; https://doi.org/10.1016/j.molcel.2024.05.031) (kim2024structureofthe pages 29-30)
6.2 2024 thesis: yeast-specific nuclear localization mechanism for Asf1
A 2024 thesis provides a localization-centric update, identifying a yeast-specific cNLS within the acidic Asf1 C-terminal tail and experimentally linking it to nuclear localization and full H3K56ac capacity. (Dannah, 2024; publication Feb 2024; https://doi.org/10.32920/25233562.v1) (dannah2024novelinsightsinto pages 166-170)
6.3 2024 preprint: linking histone chaperones to histone RNA/histone supply logic (contextual)
A 2024 preprint in a metazoan context frames ASF1 as a factor coupling chromatin assembly to histone supply and includes yeast-context statements that Asf1 participates in cell-cycle control of histone gene transcription with Hir1. While this preprint is not yeast primary research, it reflects current synthesis linking Asf1 to histone dosage/homeostasis frameworks. (Mendiratta et al., 2024; https://doi.org/10.1101/2022.11.30.518476) (mendiratta2024regulationofreplicative pages 1-6)
7) Current applications and real-world implementations
7.1 Yeast Asf1 as a mechanistic model for conserved chromatin assembly pathways
Yeast Asf1 is used as a model system to dissect histone-handling logic (H3–H4 chaperoning, modification, and handoff) that is broadly conserved across eukaryotes. The mechanistic features—particularly Asf1-dependent H3K56 acetylation and its impact on genome stability and chromatin state transmission—make yeast Asf1 a practical platform for studying replication-coupled chromatin assembly and epigenetic inheritance mechanisms. (hiraga2008histoneh3lysine pages 4-5, kattarmell2012regulationofhistone pages 10-17)
7.2 Functional readouts used in practice
Chromosome positioning assays (telomere/ETC6 perinuclear localization) provide tractable, quantitative readouts of Asf1/Rtt109/H3K56ac pathway function in vivo, and can be used to probe chromatin state inheritance across the cell cycle. (hiraga2008histoneh3lysine pages 4-5, hiraga2008histoneh3lysine pages 7-8, hiraga2008histoneh3lysine media eba6b4e4)
8) Expert synthesis and interpretive analysis (from authoritative sources in context)
8.1 Conceptual integration: Asf1 as a “hub” linking histone handling to chromatin transactions
Across the evidence, Asf1 emerges as a hub factor that: (i) binds H3–H4 to manage oligomerization and prevent inappropriate interactions; (ii) enables Rtt109-mediated H3K56 acetylation on newly synthesized histones; and (iii) supports histone handoff to deposition factors (CAF-1/Rtt106) for chromatin assembly during replication/repair, while also supporting HIR-mediated replication-independent deposition important for promoter fidelity and silencing. (kim2024structureofthe pages 29-30, dannah2024novelinsightsinto pages 67-72, kattarmell2012regulationofhistone pages 10-17)
8.2 Localization as regulatory control
The 2024 localization work implies that yeast has evolved a C-tail-embedded cNLS in Asf1 that can control nuclear availability and thereby modulate pathway output (H3K56ac), suggesting that localization is not merely passive but can be a point of regulation connecting transport to chromatin assembly competence. (dannah2024novelinsightsinto pages 166-170)
Evidence map (compact)
The following evidence map summarizes major claims, with sources, URLs, and citation IDs.
| Category | Specific claim | Evidence (short) | Key source (with year, venue) | URL | Citation ID(s) |
|---|---|---|---|---|---|
| Molecular function | Asf1 is the budding-yeast ASF1-family histone chaperone that binds H3–H4 dimers and shields the H3 tetramerization surface. | Conserved chaperone activity; binds H3–H4 and prevents inappropriate tetramer formation before handoff. | Ding et al., 2025, Nucleic Acids Research | https://doi.org/10.1093/nar/gkaf036 | (ding2025theorthologof pages 1-2) |
| Molecular function | The N-terminal core of ASF1 contains conserved acidic patches contacting histone H3 and also binds H4. | Review excerpt notes N-terminal 155 aa globular core; binds H3–H4 in vitro and in vivo and can bind each histone individually. | Breuer et al., 2024, mBio | https://doi.org/10.1128/mbio.02896-23 | (breuer2024histonebindingof pages 1-2) |
| Biological process | Asf1 functions in replication-coupled chromatin assembly and chromatin remodeling during transcription. | Hiraga excerpt states Asf1 assembles H3–H4 into nucleosomes and functions in DNA replication and transcription-coupled remodeling. | Hiraga et al., 2008, Journal of Cell Biology | https://doi.org/10.1083/jcb.200806065 | (hiraga2008histoneh3lysine pages 4-5) |
| Pathway/module | Canonical budding-yeast handoff model: Asf1 binds newly synthesized H3–H4, presents them to Rtt109 for H3K56 acetylation, then acetylated dimers are transferred to CAF-1 and/or Rtt106 for deposition. | Dannah states H3K56ac is solely reliant on Asf1 and Asf1 stimulates Rtt109; Kattar-Mell states Asf1-bound cytoplasmic H3–H4 dimers are imported, acetylated by Rtt109, then transferred to downstream chaperones including CAF-1. | Dannah, 2024, thesis; Kattar-Mell, 2012, thesis | https://doi.org/10.32920/25233562.v1 | (dannah2024novelinsightsinto pages 56-60, kattarmell2012regulationofhistone pages 10-17) |
| Key interactions | Asf1 functionally cooperates with the HIR complex and Rtt106 in replication-independent H3–H4 deposition and promoter fidelity. | Kim 2024 excerpt states Asf1 works with HIR and Rtt106 for replication-independent deposition and promoter fidelity; linked to heterochromatic silencing. | Kim et al., 2024, Molecular Cell | https://doi.org/10.1016/j.molcel.2024.05.031 | (kim2024structureofthe pages 29-30) |
| Key interactions | Asf1 interacts with HIR/Hir1 in histone gene regulation outside S phase and contributes to histone-gene activation in S phase. | Mendiratta excerpt states yeast Asf1 activates histone gene transcription in S phase and represses it outside S phase in combination with Hir1. | Mendiratta et al., 2024, bioRxiv | https://doi.org/10.1101/2022.11.30.518476 | (mendiratta2024regulationofreplicative pages 1-6) |
| Localization | Asf1 contains a functional classical NLS in its highly acidic C-terminal tail that is required for nuclear localization. | Dannah reports deleting the C-tail cNLS makes Asf1 fully cytoplasmic, independent of Vps75. | Dannah, 2024, thesis | https://doi.org/10.32920/25233562.v1 | (dannah2024novelinsightsinto pages 166-170) |
| Localization | The Asf1 C-terminal cNLS is required for full H3K56 acetylation and for interactions with Rad53 and Hir1. | Dannah reports reduced H3K56ac when the cNLS is deleted in VPS75+ cells and abolition of H3K56ac in vps75Δ background; also required for Rad53/Hir1 interaction. | Dannah, 2024, thesis | https://doi.org/10.32920/25233562.v1 | (dannah2024novelinsightsinto pages 166-170) |
| Pathway/module | H3K56 acetylation is a nuclear S-phase mark on newly synthesized H3, enabling efficient handoff of histones to CAF-1 and Rtt106 during DNA replication and repair. | Dannah notes nuclear H3K56ac during S phase and role in transfer to CAF-1/Rtt106; Hst3/Hst4 remove the mark in G2/M. | Dannah, 2024, thesis | https://doi.org/10.32920/25233562.v1 | (dannah2024novelinsightsinto pages 67-72) |
| Phenotypes & quantitative data | asf1Δ causes an S-phase-specific telomere localization defect. | Table/assay values: WT telomere XIV-L peripheral localization (zone 1) 66.0% in G1 and 67.5% in S; asf1 mutant 67.4% in G1 (n=46, p=0.05 vs WT) and 43.1% in S (n=51, p=4.7×10^-4 vs WT). | Hiraga et al., 2008, Journal of Cell Biology | https://doi.org/10.1083/jcb.200806065 | (hiraga2008histoneh3lysine pages 4-5, hiraga2008histoneh3lysine media eba6b4e4) |
| Phenotypes & quantitative data | Regulated H3K56 acetylation/deacetylation is required for chromosome positioning. | rtt109Δ shows severe telomere localization defects; both H3K56R and H3K56Q nearly abolish proper telomere localization in G1 and S. | Hiraga et al., 2008, Journal of Cell Biology | https://doi.org/10.1083/jcb.200806065 | (hiraga2008histoneh3lysine pages 5-7) |
| Biological process | Asf1 is required for perinuclear localization of the ETC6 chromatin domain across interphase. | asf1 deletion randomizes ETC6 positioning; rtt109 mutants also lose peripheral positioning, linking Asf1-mediated H3K56ac to higher-order chromosome organization. | Hiraga et al., 2008, Journal of Cell Biology | https://doi.org/10.1083/jcb.200806065 | (hiraga2008histoneh3lysine pages 7-8) |
| Key interactions | Asf1 is required for efficient Rtt109 activity and H3K56ac; Vps75 supports Rtt109 but cannot fully replace Asf1. | Dannah: loss of Asf1 markedly diminishes Rtt109 activity/H3K56ac; Hiraga: asf1 vps75 double mutant phenocopies strong localization defects. | Dannah, 2024, thesis; Hiraga et al., 2008, J Cell Biol | https://doi.org/10.32920/25233562.v1; https://doi.org/10.1083/jcb.200806065 | (dannah2024novelinsightsinto pages 56-60, hiraga2008histoneh3lysine pages 4-5) |
| Recent (2023-2024) developments | New structural work strengthens the model that Asf1 engages the HIR complex in replication-independent chromatin assembly. | Kim 2024 provides structural/biochemical evidence for Asf1 binding to HIR-related machinery and supports replication-independent H3–H4 deposition and promoter fidelity roles. | Kim et al., 2024, Molecular Cell | https://doi.org/10.1016/j.molcel.2024.05.031 | (kim2024structureofthe pages 29-30, kim2024structureofthe pages 28-29) |
| Recent (2023-2024) developments | New localization work identifies a yeast-specific Asf1 cNLS in the acidic tail and links localization directly to H3K56ac competence. | Dannah 2024 reports C-tail cNLS, cytoplasmic mislocalization upon deletion, and H3K56ac defects tied to the motif. | Dannah, 2024, thesis | https://doi.org/10.32920/25233562.v1 | (dannah2024novelinsightsinto pages 166-170) |
| Recent (2023-2024) developments | Current cross-species work reiterates that budding-yeast Asf1 promotes H3K56ac yet is dispensable for viability/growth in S. cerevisiae. | Ding 2025 contrasts S. cerevisiae with fission yeast, noting Asf1 is dispensable for growth in budding yeast while retaining conserved chaperone/H3K56ac functions. | Ding et al., 2025, Nucleic Acids Research | https://doi.org/10.1093/nar/gkaf036 | (ding2025theorthologof pages 1-2) |
Table: This table summarizes experimentally supported functional annotation for Saccharomyces cerevisiae Asf1 (UniProt P32447), emphasizing molecular function, pathways, localization, interactions, phenotypes, and recent 2023–2024 developments. It is useful as a compact evidence map for narrative gene-function reporting.
Visual evidence (figure/table excerpt)
Quantitative telomere localization differences between WT and asf1 mutants (G1 vs S phase), supporting an S-phase-specific chromosome positioning defect linked to Asf1/H3K56ac pathway function, are shown in the cropped figure/table regions from Hiraga et al. 2008. (hiraga2008histoneh3lysine media eba6b4e4, hiraga2008histoneh3lysine media bcd2be39)
Limitations of the present evidence set
- Some mechanistic statements about cytoplasmic binding/import and CAF-1 interaction derive from theses rather than peer-reviewed journal articles in the provided corpus; they are included with clear attribution and should ideally be cross-validated with additional primary papers beyond the current retrieval set. (dannah2024novelinsightsinto pages 166-170, kattarmell2012regulationofhistone pages 10-17)
- The retrieved 2023–2024 peer-reviewed yeast literature in this run is strong for HIR-complex/replication-independent deposition (Kim 2024) and for localization (Dannah 2024 thesis), but more 2023–2024 yeast primary studies specifically centered on Asf1 (e.g., direct biochemical reconstitution of yeast Asf1–Rtt109–Vps75–CAF-1 handoff) were not captured in the current tool retrieval.
References
-
(kim2024structureofthe pages 29-30): Hee Jong Kim, Mary R. Szurgot, Trevor van Eeuwen, M. Daniel Ricketts, Pratik Basnet, Athena L. Zhang, Austin Vogt, Samah Sharmin, Craig D. Kaplan, Benjamin A. Garcia, Ronen Marmorstein, and Kenji Murakami. Structure of the hir histone chaperone complex. Molecular Cell, 84:2601-2617.e12, Jul 2024. URL: https://doi.org/10.1016/j.molcel.2024.05.031, doi:10.1016/j.molcel.2024.05.031. This article has 14 citations and is from a highest quality peer-reviewed journal.
-
(hiraga2008histoneh3lysine pages 4-5): Shin-ichiro Hiraga, Sotirios Botsios, and Anne D. Donaldson. Histone h3 lysine 56 acetylation by rtt109 is crucial for chromosome positioning. The Journal of Cell Biology, 183:641-651, Nov 2008. URL: https://doi.org/10.1083/jcb.200806065, doi:10.1083/jcb.200806065. This article has 48 citations.
-
(ding2025theorthologof pages 1-2): Yan Ding, Jun Li, He-Li Jiang, Fang Suo, Guang-Can Shao, Xiao-Ran Zhang, Meng-Qiu Dong, Chao-Pei Liu, Rui-Ming Xu, and Li-Lin Du. The ortholog of human dnajc9 promotes histone h3–h4 degradation and is counteracted by asf1 in fission yeast. Nucleic Acids Research, Jan 2025. URL: https://doi.org/10.1093/nar/gkaf036, doi:10.1093/nar/gkaf036. This article has 2 citations and is from a highest quality peer-reviewed journal.
-
(breuer2024histonebindingof pages 1-2): Jan Breuer, Tobias Busche, Jörn Kalinowski, and Minou Nowrousian. Histone binding of asf1 is required for fruiting body development but not for genome stability in the filamentous fungus sordaria macrospora. mBio, Jan 2024. URL: https://doi.org/10.1128/mbio.02896-23, doi:10.1128/mbio.02896-23. This article has 6 citations and is from a domain leading peer-reviewed journal.
-
(kattarmell2012regulationofhistone pages 10-17): SJ Kattar-Mell. Regulation of histone h3k56 acetylation by the histone h2a-h2b acidic patch. Unknown journal, 2012.
-
(dannah2024novelinsightsinto pages 67-72): Nora Saud Dannah. Novel insights into nuclear localization of proteins crucial for chromatin assembly metabolism in saccharomyces cerevisiae. Unknown journal, Feb 2024. URL: https://doi.org/10.32920/25233562.v1, doi:10.32920/25233562.v1.
-
(dannah2024novelinsightsinto pages 56-60): Nora Saud Dannah. Novel insights into nuclear localization of proteins crucial for chromatin assembly metabolism in saccharomyces cerevisiae. Unknown journal, Feb 2024. URL: https://doi.org/10.32920/25233562.v1, doi:10.32920/25233562.v1.
-
(mendiratta2024regulationofreplicative pages 1-6): Shweta Mendiratta, Dominique Ray-Gallet, Alberto Gatto, Sébastien Lemaire, Maciej A. Kerlin, Antoine Coulon, and Geneviève Almouzni. Regulation of replicative histone rna metabolism by the histone chaperone asf1. bioRxiv, Nov 2024. URL: https://doi.org/10.1101/2022.11.30.518476, doi:10.1101/2022.11.30.518476. This article has 10 citations.
-
(dannah2024novelinsightsinto pages 166-170): Nora Saud Dannah. Novel insights into nuclear localization of proteins crucial for chromatin assembly metabolism in saccharomyces cerevisiae. Unknown journal, Feb 2024. URL: https://doi.org/10.32920/25233562.v1, doi:10.32920/25233562.v1.
-
(hiraga2008histoneh3lysine media eba6b4e4): Shin-ichiro Hiraga, Sotirios Botsios, and Anne D. Donaldson. Histone h3 lysine 56 acetylation by rtt109 is crucial for chromosome positioning. The Journal of Cell Biology, 183:641-651, Nov 2008. URL: https://doi.org/10.1083/jcb.200806065, doi:10.1083/jcb.200806065. This article has 48 citations.
-
(hiraga2008histoneh3lysine pages 5-7): Shin-ichiro Hiraga, Sotirios Botsios, and Anne D. Donaldson. Histone h3 lysine 56 acetylation by rtt109 is crucial for chromosome positioning. The Journal of Cell Biology, 183:641-651, Nov 2008. URL: https://doi.org/10.1083/jcb.200806065, doi:10.1083/jcb.200806065. This article has 48 citations.
-
(hiraga2008histoneh3lysine pages 7-8): Shin-ichiro Hiraga, Sotirios Botsios, and Anne D. Donaldson. Histone h3 lysine 56 acetylation by rtt109 is crucial for chromosome positioning. The Journal of Cell Biology, 183:641-651, Nov 2008. URL: https://doi.org/10.1083/jcb.200806065, doi:10.1083/jcb.200806065. This article has 48 citations.
-
(kim2024structureofthe pages 28-29): Hee Jong Kim, Mary R. Szurgot, Trevor van Eeuwen, M. Daniel Ricketts, Pratik Basnet, Athena L. Zhang, Austin Vogt, Samah Sharmin, Craig D. Kaplan, Benjamin A. Garcia, Ronen Marmorstein, and Kenji Murakami. Structure of the hir histone chaperone complex. Molecular Cell, 84:2601-2617.e12, Jul 2024. URL: https://doi.org/10.1016/j.molcel.2024.05.031, doi:10.1016/j.molcel.2024.05.031. This article has 14 citations and is from a highest quality peer-reviewed journal.
-
(hiraga2008histoneh3lysine media bcd2be39): Shin-ichiro Hiraga, Sotirios Botsios, and Anne D. Donaldson. Histone h3 lysine 56 acetylation by rtt109 is crucial for chromosome positioning. The Journal of Cell Biology, 183:641-651, Nov 2008. URL: https://doi.org/10.1083/jcb.200806065, doi:10.1083/jcb.200806065. This article has 48 citations.
Artifacts
Citations
- ding2025theorthologof pages 1-2
- breuer2024histonebindingof pages 1-2
- kattarmell2012regulationofhistone pages 10-17
- dannah2024novelinsightsinto pages 56-60
- kim2024structureofthe pages 29-30
- mendiratta2024regulationofreplicative pages 1-6
- dannah2024novelinsightsinto pages 166-170
- dannah2024novelinsightsinto pages 67-72
- kim2024structureofthe pages 28-29
- https://doi.org/10.1016/j.molcel.2024.05.031
- https://doi.org/10.32920/25233562.v1
- https://doi.org/10.1101/2022.11.30.518476
- https://doi.org/10.1093/nar/gkaf036
- https://doi.org/10.1128/mbio.02896-23
- https://doi.org/10.1083/jcb.200806065
- https://doi.org/10.32920/25233562.v1;
- https://doi.org/10.1016/j.molcel.2024.05.031,
- https://doi.org/10.1083/jcb.200806065,
- https://doi.org/10.1093/nar/gkaf036,
- https://doi.org/10.1128/mbio.02896-23,
- https://doi.org/10.32920/25233562.v1,
- https://doi.org/10.1101/2022.11.30.518476,
Perplexity
(ASF1-deep-research-perplexity.md)
Perplexity
(ASF1-deep-research-perplexity.md)ASF1: A Multifunctional Histone H3-H4 Chaperone in Saccharomyces cerevisiae
ASF1 (anti-silencing function protein 1), encoded by the ASF1 gene (UniProt P32447) in Saccharomyces cerevisiae, is a highly conserved histone chaperone that serves as a central hub in chromatin assembly, disassembly, and remodeling pathways. This protein plays critical roles in nucleosome assembly coupled to DNA replication, replication-independent histone deposition, transcriptional regulation, and the cellular response to DNA damage. ASF1 functions through its specific interaction with histone H3-H4 dimers and its capacity to transfer these histones to downstream assembly factors including chromatin assembly factor 1 (CAF-1) and the HIRA complex. The protein's diverse cellular functions are enabled by its multiple protein-protein interaction interfaces and its regulation by post-translational modifications and checkpoint kinases. This report synthesizes current understanding of ASF1 structure, biochemical mechanisms, and biological roles in coordinating chromatin dynamics with DNA metabolism.
Structural Features and Protein Architecture
The N-Terminal Immunoglobulin-Like Fold Domain
ASF1 comprises a highly conserved N-terminal domain and a divergent C-terminal tail that are functionally distinct. The N-terminal domain, consisting of approximately 156 amino acids, adopts an immunoglobulin-like (Ig-fold) structure characterized by a β-sandwich architecture with three β-sheets topped by two short α-helices[6][26][37]. This core structural domain is responsible for histone binding and interactions with other histone chaperones and checkpoint proteins. The crystal structure of yeast ASF1 bound to the C-terminal helix of histone H3 reveals that the histone-binding surface forms a highly conserved hydrophobic groove surrounded by charged residues[6][26]. The binding pocket accommodates the C-terminal helix of histone H3 (residues 121-134 in yeast) in a configuration that presents both conserved and yeast-specific residues critical for histone recognition[26]. Notably, the histone H3 C-terminal helix adopts a helical conformation when bound to ASF1, even though this peptide is unstructured in isolation[6].
The geometry of the ASF1-histone interaction reveals that ASF1 binds to histones H3-H4 in a manner that physically blocks the H3-H3 dimerization interface found in the histone octamer[26][37]. This structural feature explains ASF1's ability to prevent formation of H3-H4 tetramers and maintain histones in the dimeric state. The interaction interface extends beyond the histone tail peptide, with ASF1 making critical contacts with both the C-terminal tail of histone H3 and the α2 and α3 helices of the histone fold domain of both H3 and H4[37]. Unexpectedly, the binding of ASF1 to H3-H4 causes a dramatic conformational change in the C-terminal tail of histone H4, which unfolds from its nucleosomal position and adds a β-strand to the ASF1 β-sheet sandwich[37]. This conformational remodeling of histone H4 is functionally important for subsequent enzymatic modifications and transfer to downstream chaperones[37].
The C-Terminal Tail and Protein-Protein Interactions
In contrast to the highly conserved N-terminal core, the C-terminal region of ASF1 is nonconserved between species and largely unstructured[2]. However, despite its apparent lack of structural organization, this C-terminal tail plays important roles in regulating protein-protein interactions. The C-terminal region contributes to interactions with both histone H3-H4 and the CAF-1 complex[14][17], and it also serves as the primary binding site for the DNA damage checkpoint kinase Rad53[6][12]. Mutations in the C-terminus of ASF1 can enhance histone silencing abilities while reducing the affinity for Rad53, suggesting that the unstructured tail acts to stabilize protein-protein interactions and that the balance between different ASF1 partners may be regulated through this domain[1]. The presence of an acidic cluster near the C-terminus has been identified as responsible for modulation of protein kinase CK2 activity, revealing an unexpected regulatory role for this region[21].
Structural Conservation and Evolutionary Relationships
The ASF1 N-terminal domain is remarkably conserved throughout eukaryotic evolution, with orthologous proteins present in yeast, plants, flies, and mammals[2][6]. Human cells express two non-allelic ASF1 isoforms, ASF1a and ASF1b, which share 70% sequence identity in their first 156 amino acids but diverge significantly in their C-terminal regions[3][31]. The conserved N-terminal domain is sufficient for many ASF1 functions in vivo, as truncated versions of ASF1 lacking the C-terminus retain the ability to bind histones and participate in nucleosome assembly[3][26]. Structural comparison of ASF1 with other proteins reveals an Ig-fold architecture that is also present in the YEATS domain of Yaf9, a component of histone variant deposition complexes, suggesting that this fold may be a general platform for protein-histone interactions[29]. The conservation of ASF1 structure and function across diverse organisms highlights the fundamental importance of this histone chaperone in chromatin regulation.
Stoichiometry and Biochemistry of the ASF1-H3-H4 Complex
Binding of ASF1 to H3-H4 Dimers Rather than Tetramers
A fundamental aspect of ASF1 function is its specific binding to H3-H4 heterodimers rather than H3-H4 heterotetramers. Early biochemical studies established through gel filtration, amino acid analysis, reversed-phase chromatography, and analytical ultracentrifugation that a single molecule of ASF1 binds to an H3-H4 heterodimer in a 1:1:1 stoichiometry[2]. This finding was surprising because the first step in nucleosome formation was traditionally assumed to be the deposition of a histone H3-H4 heterotetramer onto DNA. The discovery that ASF1 binds only to H3-H4 dimers and blocks formation of the H3-H4 heterotetramer through occlusion of the H3-H3 dimerization interface suggested that nucleosome assembly proceeds through a two-step mechanism involving sequential deposition of two H3-H4 heterodimers[2]. The theoretical molecular mass of a 1:1:1 ASF1-H3-H4 complex is approximately 42 kDa, and sedimentation equilibrium analysis confirmed this stoichiometry under multiple experimental conditions[2].
The ASF1-H3-H4 complex exists as a major soluble pool of newly synthesized histones in cells[31]. The interaction between ASF1 and H3-H4 is remarkably tight, with binding affinities in the nanomolar range (approximately 2 nM)[14]. This high-affinity interaction ensures that newly synthesized H3-H4 dimers are efficiently captured by ASF1 and protected from aggregation or inappropriate interactions. The binding of ASF1 to H3-H4 is mediated through specific contacts between the N-terminal core of ASF1 and both histones, with the histone-binding surface forming a conserved hydrophobic groove[6]. Studies using NMR chemical shift mapping and mutagenesis identified a surface on ASF1 that binds the C-terminal helix of histone H3, and mutations within this site can dramatically reduce or abolish H3-H4 binding, resulting in defects in chromatin assembly and transcriptional silencing[6][40].
Regulation of ASF1-H3-H4 Interactions by Other Proteins
The ASF1-H3-H4 interaction is not static but rather can be influenced by other histone chaperones and regulatory proteins. Interestingly, NASP (histone-binding protein) can compete with ASF1 for binding to H3 monomers, and NASP and ASF1 can share an H3-H4 dimer in a conformation compatible with both proteins binding simultaneously[31]. The HAT1 histone acetyltransferase complex, which acetylates histones H4 on lysines 5 and 12, can form complexes with ASF1 and H3-H4, suggesting that histone modification occurs while histones are in complex with ASF1[31][60]. The protein CDAN1 (codanin-1) competes with histone H3-H4 for ASF1 binding in human cells, potentially regulating the availability of ASF1 for different downstream pathways[31]. These regulatory mechanisms indicate that the ASF1-H3-H4 complex is a dynamic hub in the histone chaperoning network rather than a static end product.
Role in Replication-Coupled Nucleosome Assembly
Interaction with CAF-1 and Transfer of H3-H4 Dimers
ASF1 functions synergistically with chromatin assembly factor 1 (CAF-1) during replication-coupled chromatin assembly. CAF-1 is recruited to active replication forks through its interaction with the processivity clamp PCNA (proliferating cell nuclear antigen)[31]. The transfer of H3-H4 from ASF1 to CAF-1 is a critical step in the assembly pathway, and ASF1 interacts directly with CAF-1 through the B-domain of CAF-1's p60 subunit[14][31]. Surprisingly, biophysical studies have shown that although H3-H4 enhances the interaction between ASF1 and CAF-1, the resulting CAF-1-H3-H4 complex is mutually exclusive of ASF1, indicating that CAF-1 functions independently of ASF1 after histone acquisition[14]. The binding of H3-H4 dimers to CAF-1 differs from ASF1 binding in that a single CAF-1 complex can bind multiple H3-H4 dimers, ultimately promoting formation of (H3-H4)₂ tetramers on DNA[14].
Studies with purified proteins demonstrate that ASF1 can transfer H3-H4 dimers to CAF-1 without forming a stable higher-order complex containing both histone chaperones[5]. The replication fork protein MCM2 (minichromosome maintenance protein 2) also interacts with ASF1-H3-H4, and the crystal structure of this complex reveals a 1:1:1:1 stoichiometry indicating that dimeric H3-H4 is propagated at replication forks[5]. The interaction of ASF1 with MCM2 occurs at the unstructured N-terminal region of MCM2 and is compatible with simultaneous binding of H3-H4, suggesting that ASF1 may facilitate histone transfer from the replicative helicase to downstream assembly factors[5][31]. The directionality of the histone deposition reaction is provided by histone tetramerization, which exhibits higher binding affinity for DNA than the ASF1-H3-H4 complex or CAF-1 alone, effectively pulling histones forward through the assembly pathway[5].
Role of ASF1 in Maintaining Replication Fork Integrity
Beyond its role in histone deposition, ASF1 directly interacts with the replication machinery and maintains the stability of the replication fork. RFC (replication factor C), the heteropentameric ATPase that loads PCNA onto DNA, directly binds to ASF1 and can recruit ASF1 to DNA in vitro[7][32]. In budding yeast cells lacking ASF1, the amounts of several DNA replication proteins including RFC subunits, PCNA, and DNA polymerase ε are reduced at stalled replication forks, while DNA polymerase α accumulates to higher than normal levels[7][32]. The ASF1-RFC interaction is reversible and appears to be regulated by nucleotide binding, with addition of nucleotides reducing the interaction by approximately 50%[7]. This reversible interaction suggests that ASF1 may regulate replication forks through transient contacts that dynamically respond to the state of the replication machinery[7].
The importance of ASF1 for replication fork stability is highlighted by genetic studies showing that cells lacking ASF1 are highly sensitive to replication fork-stalling drugs such as hydroxyurea (HU) and camptothecin (CPT)[33]. These sensitivities are not shared by cells lacking CAF-1 or HIRA complexes, indicating that ASF1 has unique functions in maintaining fork integrity that are distinct from its role in histone deposition. The physical interaction between ASF1 and RFC ensures that histone chaperone activity is coupled to DNA replication and that replication fork stability is maintained through proper chromatin assembly. Studies examining the kinetics of DNA synthesis have revealed that nucleosome assembly is required for normal fork progression and PCNA unloading, with blocking of histone transfer from ASF1 to CAF-1 causing significant slowdown of replication fork movement[57].
Role in Replication-Independent Histone Deposition
Interaction with the HIR Complex and H3.3 Variant Deposition
ASF1 also functions in replication-independent (RI) chromatin assembly through interaction with the HIRA (histone repression A factor) complex. The HIRA complex is composed of Hir1, Hir2, Hir3, and Hpc2 proteins and specifically deposits histone H3.3, a variant that differs from canonical H3.1 at only four amino acid positions but is incorporated throughout the cell cycle independently of DNA replication[19][45]. ASF1 co-purifies with all four components of the HIR complex and together with HIRA deposits H3.3 onto DNA in a replication-independent manner[19][45]. The interaction between ASF1 and HIRA is mediated through the B-domain of HIRA (residues 439-475), which binds to the hydrophobic cleft of ASF1's N-terminal domain[48]. This binding interface is specific to ASF1a but not ASF1b in humans, potentially explaining functional specialization between these isoforms in mammals[48].
Replication-independent chromatin assembly by the HIR complex and ASF1 occurs at transcriptionally active regions and after histone eviction during transcriptional activation, allowing replacement of nucleosomes that have been displaced by transcription factors and transcription machinery[19][45]. The HIR-ASF1 pathway is functionally important for reestablishment of histone acetylation patterns after DNA replication and for maintaining proper chromatin structure at promoters after histone deposition. Genetic studies have shown that deletion of ASF1 in the absence of CAF-1 and HIRA causes synthetic growth defects and loss of silencing at heterochromatic loci, indicating that these histone deposition pathways have overlapping but distinct functions[19][45]. The use of different histone variants (H3.1 via CAF-1 during replication, H3.3 via HIRA independent of replication) may allow cells to distinguish and regulate chromatin assembled at different times and under different circumstances.
Role in Histone H3 Lysine 56 Acetylation and S-Phase Chromatin Assembly
The ASF1-Rtt109 Complex and K56ac Deposition
A particularly important function of ASF1 is its role in facilitating acetylation of histone H3 at lysine 56 (H3 K56ac), a modification that is essential for proper nucleosome assembly during S phase and for maintaining genome stability in the presence of replication stress. The histone acetyltransferase (HAT) Rtt109 catalyzes H3 K56 acetylation and requires either ASF1 or Vps75 as a histone chaperone partner for enzymatic activity[33][55]. The crystal structure of the Rtt109-ASF1-H3-H4 complex reveals that ASF1 is essential for H3 K56 acetylation through stabilization of histone H4's C-terminal tail, allowing proper substrate positioning for the Rtt109 active site[55]. Unexpectedly, unwinding of the N-terminal helix of histone H3 (where K56 is normally located) also occurs during acetylation, and this conformational rearrangement is facilitated by the ASF1-H3-H4 interaction[55].
Cells lacking ASF1 show dramatically reduced levels of H3 K56ac and display sensitivity to replication fork-stalling drugs and DNA-damaging agents, phenotypes that can be largely suppressed by an H3 K56Q mutation that mimics acetylation[33]. This genetic evidence demonstrates that the role of ASF1 in genome stability during S phase is largely mediated through its function in H3 K56 acetylation. The modification of H3 K56 weakens the electrostatic interaction between the positively charged histone H3 and the negatively charged DNA, creating a more "open" chromatin environment that allows easier access for remodeling and repair factors[33]. During replication stress, newly synthesized histone H3 interacting with ASF1 increases substantially, supporting a model in which ASF1 functions as a histone buffer that donates H3-H4 to assembly factors[3]. The high level of H3 K56ac in S-phase cells (approximately five-fold higher than in asynchronous populations) indicates that this modification is specifically enriched on newly synthesized histones immediately following their incorporation into chromatin[33].
H3 K56ac in Transcriptional Regulation and Chromatin Dynamics
In addition to its role in replication, H3 K56ac functions in transcriptional regulation by facilitating chromatin disassembly at promoters during gene activation. At the PHO5 promoter, transcriptional induction is accompanied by increased levels of H3 K56ac on the promoter region, and mutations that prevent K56 acetylation (such as in rtt109Δ or asf1Δ strains) result in delayed chromatin disassembly and delayed transcriptional activation[58]. Conversely, chromatin reassembly during transcriptional repression is accompanied by decreased H3 K56ac levels, suggesting that this modification actively drives chromatin toward a disassembled state[58]. The role of H3 K56ac in promoting chromatin disassembly may involve displacement of parental histones from DNA, as the acetylation of this residue on free H3 in complex with ASF1 and incorporation into chromatin drives disassembly of preexisting nucleosomes[58].
Transcriptional Regulation Functions
Nucleosome Disassembly at Promoters and Transcriptional Activation
ASF1 plays a critical role in nucleosome disassembly at promoters during transcriptional activation. Early studies showed that chromatin disassembly mediated by ASF1 is essential for transcriptional activation of the yeast PHO5 and PHO8 genes[1][8]. Subsequent research revealed that this function occurs through multiple mechanisms. At the PHO5 promoter, nucleosome displacement occurs in waves both along the length of the promoter and during the cell cycle, with ASF1 being required specifically for nucleosome eviction at upstream regulatory regions (URS2) but not at the upstream activating sequence URS1[18]. Different chromatin modifiers are required for chromatin disassembly at different promoter regions, with Swi/Snf being required at URS1, while FACT (facilitates chromatin transcription) and ASF1 are both required at URS2[18].
The mechanism by which ASF1 promotes nucleosome disassembly likely involves multiple functions. ASF1 binds to the C-terminal region of histone H3 in free histones, and this interaction cannot occur within the context of an intact nucleosome[26]. Therefore, ASF1-mediated nucleosome disassembly presumably involves displacement of H3-H4 from nucleosomal DNA followed by capture of the displaced H3-H4 by ASF1. The binding of ASF1 to free H3-H4 physically prevents reformation of H3-H4 tetramers, allowing newly synthesized H3-H4 (particularly acetylated forms) to be incorporated while old histones are retained in ASF1 complexes. Chromatin disassembly at promoters occurs concurrently with histone acetylation, and the acetylation of newly incorporated H3 K56 may destabilize downstream nucleosomes, creating a positive feedback loop for progressive chromatin opening[18].
Involvement in Transcriptional Silencing and Gene Repression
Although ASF1 is most commonly recognized for its role in nucleosome assembly and transcriptional activation, the protein also functions in transcriptional silencing, particularly at heterochromatic loci including telomeres and the silent mating-type loci (HML and HMR). Mutations in ASF1 enhance transcriptional silencing at these loci by affecting histone deposition and Sir protein recruitment[1]. Dominant mutations in the C-terminus of ASF1 (such as 152I and 185T) result in enhanced silencing that overcomes the requirement for CAF-1 in repressing silent loci[1]. These silencing-enhancing mutations reduce the affinity of ASF1 for histone H3-H4, apparently shifting the balance toward the HIR-Asf1 pathway relative to CAF-1 binding[1]. The enhanced silencing function correlates with restoration of histone H3 occupancy and Sir2 and Sir4 recruitment at silent loci in CAF-1-deficient strains[1].
In Drosophila, ASF1 has been shown to interact with the Su(H)/HAIRLESS DNA-binding protein complex and to be recruited to specific Notch target loci, where it promotes gene silencing rather than activation[30]. This observation suggests that ASF1 can function in repression of specific genes when recruited through interactions with sequence-specific DNA-binding proteins. The SAS-I complex, which includes Sas2, Sas4, and Sas5 proteins and links histone acetylation to silencing, physically interacts with both Cac1 and ASF1[13][16]. These findings indicate that ASF1 participates in multiple chromatin modification pathways that can promote either transcriptional activation or repression depending on its partner proteins and the histone modifications present.
Regulation of Histone Gene Expression
ASF1 participates in the regulation of histone gene expression, which must be carefully controlled to match the increased demand for histones during S phase. The HIR complex, composed of Hir1, Hir2, Hir3, and Hpc2, functions with ASF1 to repress histone gene transcription outside of S phase[51]. However, depletion of ASF1 from cells has little effect on the levels of histone transcripts during the cell cycle, suggesting that the HIR complex and associated ASF1 do not have redundant functions with other transcriptional regulators at histone promoters[51]. The NegI region of histone promoters mediates HIR-dependent repression, and deletion of this region derepresses histone gene expression, indicating that the HIR complex acts as a direct transcriptional repressor[51]. The primary regulators of histone gene expression during S phase appear to be Spt10 and the SBF transcription factor, which together drive the major peak of histone mRNA synthesis in sync with DNA replication.
Role in DNA Damage Response and Checkpoint Signaling
Interaction with the Rad53 Checkpoint Kinase
ASF1 physically interacts with the DNA damage checkpoint kinase Rad53 (mammalian Chk2 homolog) and plays a crucial role in the cellular response to DNA damage. The Rad53 kinase is activated by phosphorylation in response to DNA damage and replication blocks, and Rad53 binding to ASF1 inhibits the histone deposition activity of ASF1 in vitro, providing a mechanism to couple chromatin assembly to checkpoint signaling[12]. The interaction between Rad53 and ASF1 is mediated through the C-terminal tail of ASF1, and this interaction is regulated by Mec1-dependent phosphorylation of Rad53[42]. Following DNA damage, phosphorylated Rad53 dissociates from ASF1, allowing ASF1 to resume its histone deposition function. When DNA repair is complete and the checkpoint is turned off, Rad53 is dephosphorylated and reassociates with ASF1[42].
The biological significance of ASF1-Rad53 interaction is highlighted by the discovery that deletion of ASF1 reduces cell viability when multiple DNA double-strand breaks (DSBs) are induced[9][56]. In cells suffering two DSBs, deletion of ASF1 reduces viability from approximately 70% in wild-type cells to 40%, and the defect correlates with delayed dephosphorylation of Rad53[56]. The role of ASF1 in facilitating Rad53 dephosphorylation is both histone-chaperone dependent and independent, as dissociation of histone H3 from ASF1 is required for efficient recovery from the DNA damage checkpoint[9]. Significantly, transient reduction of Rad53 levels in ASF1-deficient cells experiencing DSBs restores viability to near wild-type levels, demonstrating that the recovery defect of asf1Δ cells is due entirely to a failure to properly deactivate the checkpoint[56].
Regulation by the Mec1-Rad53-Dun1 Checkpoint Pathway
The Mec1 and Dun1 checkpoint kinases regulate the ASF1-Rad53 interaction and therefore control the activity of the ASF1-HIR complex at heterochromatic loci. Deletion of the upstream kinase Mec1 alone does not cause major defects in telomeric silencing, but in cells lacking both Mec1 and CAF-1, silencing is dramatically reduced[42]. This phenotype can be reversed by deletion of Rad53, suggesting that Mec1 normally antagonizes Rad53-mediated inhibition of the ASF1-HIR complex[42]. The degree of suppression of silencing defects by various rad53 alleles correlates with their effects on ASF1 binding, further supporting the model that Mec1 and Dun1 checkpoint kinases regulate chromatin assembly through modulation of the Rad53-ASF1 interaction[42].
Protein Interactions and Regulatory Complexes
The Multifunctional Protein-Protein Interaction Surface of ASF1
ASF1 possesses multiple protein interaction surfaces that allow it to function as a central hub in chromatin regulation. The N-terminal core of ASF1 contains on one face a binding site for the C-terminal helix of histone H3-H4, and on the opposite face a β-hairpin-like "B-domain" interaction surface that binds to CAF-1, HIRA, and Rad53[14][48]. This arrangement allows ASF1 to potentially interact simultaneously with histones and histone deposition factors on one face while binding checkpoint proteins on the other face. The C-terminal tail of ASF1 contributes additional interactions with both H3-H4 and CAF-1, and these interactions can be disrupted by mutations that enhance silencing, suggesting that the balance between ASF1 partner proteins can be shifted through C-terminal mutations[1][14][17].
The interaction of ASF1 with Rad53 occurs at the B-domain interaction surface and can be inhibited by phosphorylation of Rad53[42]. In contrast, the interaction between ASF1 and H3-H4 occurs on the same hydrophobic groove used for CAF-1 binding, yet NASP can compete with ASF1 for this site, and other chaperones can form complexes with ASF1 and H3-H4 simultaneously[31]. These observations suggest that protein-protein interactions at the ASF1 N-terminal core are highly dynamic and subject to regulation by post-translational modifications, changes in protein concentration, and protein-protein interactions.
Interaction with Transcription-Associated Chromatin Remodelers
ASF1 interacts with multiple chromatin remodeling and transcription-associated complexes. Studies in Drosophila demonstrated that ASF1 directly associates with the Brahma (SWI/SNF) chromatin-remodeling complex[30], and biochemical studies have shown that ASF1 interacts with FACT (facilitates chromatin transcription), a complex that reorganizes nucleosomes without hydrolyzing ATP[18]. At the PHO5 promoter, ASF1 and FACT are both required for nucleosome disassembly at specific promoter regions and for subsequent coactivator recruitment[18]. The involvement of ASF1 with SWI/SNF and FACT suggests that ASF1 works in concert with chromatin-remodeling machinery to promote nucleosome displacement and facilitate transcription factor access to DNA.
ASF1 also interacts with the SAS-I complex (comprising Sas2, Sas4, and Sas5), which links histone acetylation to the establishment of transcriptional silencing[13][16]. The SAS-I complex interacts with Cac1 (and indirectly with CAF-1) and with ASF1, and genetic studies suggest overlapping functions of Cac1 and ASF1 that are mediated by the effect of Sas2 on histone acetylation[13]. These findings indicate that ASF1 functions as a connection point between histone deposition machinery and the enzymes that modify histones, allowing coordination of histone incorporation with histone tail modifications.
Localization and Cell Cycle Regulation
Nuclear and Cytoplasmic Localization
ASF1 exhibits cell cycle-dependent changes in its nuclear and cytoplasmic localization. The protein has been shown to have a cell cycle-specific nuclear and cytoplasmic localization pattern, with localization sensitive to Triton extraction, indicating that much of the protein in the nucleus is associated with chromatin or protein complexes[27]. In mammalian cells, ASF1a displays homogeneous distribution throughout the nucleus without major changes upon UV irradiation, while ASF1b is associated with cellular proliferation and may have cell cycle-dependent localization[3][27]. Following DNA damage, CAF-1 p60 shows increased Triton-resistant fraction, indicating association with chromatin or stable protein complexes, suggesting that ASF1 may similarly relocate or associate with damaged DNA.
The localization of ASF1 is coordinated with histone H3 deposition, and time-course analysis of H3 incorporation shows colocalization with replication foci one hour after S phase entry[3]. By two hours after S phase entry, H3 incorporation shows only partial overlap with replication forks, suggesting that the majority of newly synthesized H3 has been incorporated into chromatin[3]. ASF1 localization may also be regulated by interaction with its binding partners, as formation of ASF1-Rad53 complexes or ASF1-H3-H4 complexes could alter the nuclear localization and stability of the protein.
Evolutionary Conservation and Orthologous Proteins
Conservation Across Eukaryotes
ASF1 is one of the most highly conserved histone chaperones, with orthologous proteins present throughout eukaryotes, from unicellular yeast to mammals and plants. The N-terminal immunoglobulin-like fold domain is nearly identical across species, with the conservation extending to the specific histone H3-H4 binding interface[2][6][22]. This exceptional evolutionary conservation underscores the fundamental importance of ASF1 function in chromatin regulation. Human cells express two ASF1 isoforms, ASF1a and ASF1b, which diverged from a common ancestor and show functional specialization: ASF1a is expressed constitutively and participates in both replication-dependent and replication-independent pathways, while ASF1b is preferentially associated with cellular proliferation[3][48].
In protists such as Plasmodium falciparum (the causative agent of malaria), ASF1 has been structurally and functionally characterized and shown to deposit H3-H4 dimers onto DNA template to form disomes, revealing that the core function of ASF1 as an H3-H4 chaperone is conserved even in this distantly related organism[22]. In trypanosomes such as Trypanosoma brucei, multiple ASF1 proteins exist with differential localization and cell cycle-dependent nuclear translocation, suggesting expanded functions in these organisms with unusual chromatin organization[24]. The conservation of ASF1 function across such evolutionarily distant organisms indicates that this histone chaperone represents a fundamental and ancient mechanism of chromatin regulation.
Regulatory Mechanisms and Post-Translational Modifications
Phosphorylation and Kinase Regulation
ASF1 is subject to phosphorylation by multiple kinases that regulate its function. In Drosophila, the Tousled-like kinase (TLK) phosphorylates ASF1, and the two components cooperate to control chromatin replication in vivo[39]. The phosphorylation of ASF1 by TLK or related kinases may modulate its interaction with histones or downstream chaperones, coupling checkpoint signaling to chromatin assembly. The interaction between Rad53 and ASF1 is regulated by Rad53 phosphorylation, with phosphorylated Rad53 failing to bind ASF1[42]. Conversely, the Dun1 kinase, a downstream target of Rad53, positively regulates the Rad53-ASF1 interaction in vivo[42], suggesting a mechanism in which Dun1 phosphorylation of Rad53 or ASF1 enhances their interaction and coupling of the DNA damage response to chromatin assembly.
Protein Kinase CK2 Modulation
The acidic cluster near the C-terminus of ASF1 (residues 170-193) interacts with both catalytic subunits of protein kinase CK2 (CK2α and CK2α') and modulates their activity[21]. ASF1 exhibits differential effects on the two CK2 catalytic subunits, activating CK2α at low concentrations while inhibiting it at higher concentrations, while consistently inhibiting CK2α'[21]. This bipartite regulation of CK2 represents an unexpected function of ASF1 outside of histone chaperoning and may reflect a mechanism by which ASF1 coordinates protein phosphorylation with chromatin assembly during the cell cycle.
Functional Integration and Cellular Roles
ASF1 as a Histone Buffering Hub
ASF1 functions as a central hub and "histone sink" in the cytoplasm that buffers the soluble pool of newly synthesized histone H3-H4[3][14][31]. During normal growth, ASF1 forms stable complexes with H3-H4 that protect the histones from aggregation and inappropriate interactions. During replication stress, the amount of newly synthesized histone H3 interacting with human ASF1 (hASF1) increases dramatically[3], supporting a model in which ASF1 serves as a buffer that donates H3-H4 to CAF-1 and HIRA, which are more directly involved in depositing histones onto DNA. This buffering function allows cells to maintain soluble pools of histones that can be rapidly delivered to replication forks or transcriptionally active regions as needed.
The thermodynamic properties of ASF1-H3-H4 interaction support this buffering role, as the ASF1-H3-H4 complex represents a stable intermediate with defined binding affinity that allows kinetic control of histone deposition through regulated transfer to downstream chaperones[5][17]. The formation of (H3-H4)₂ tetramers on DNA exhibits higher binding affinity (approximately 1 nM) than ASF1 binding (approximately 2 nM)[5], creating a thermodynamic gradient that drives histone transfer from ASF1 to downstream assembly factors and ultimately to DNA, while maintaining specificity through protein-protein interactions.
Genome-Wide Mapping of ASF1 Binding Sites
Genome-wide chromatin immunoprecipitation and sequencing (ChIP-seq) studies have revealed that yeast ASF1 associates with 590 unique targets throughout the genome[44][47]. ASF1 shows high association with heterochromatic centromeres and telomeres, suggesting roles in condensin loading and heterochromatin formation[47]. ASF1 also associates with genes transcribed by RNA polymerase III, where it facilitates replication-independent histone H3 exchange[47]. These findings indicate that ASF1 functions broadly throughout the genome in both constitutively expressed loci and at heterochromatic regions, consistent with its roles in both nucleosome assembly and transcriptional regulation.
Conclusion
ASF1 represents a paradigm of a multifunctional histone chaperone that coordinates multiple cellular processes through its specific interaction with histone H3-H4 dimers and its multiple protein-protein interaction surfaces. The protein binds histone H3-H4 dimers with high affinity and prevents formation of H3-H4 tetramers, serving as a central hub that accepts newly synthesized histones from nascent peptide synthesis and delivers them to downstream assembly factors. Through interactions with CAF-1, ASF1 promotes replication-coupled nucleosome assembly, maintaining genome stability and allowing rapid chromatin assembly in the wake of DNA replication. Through interactions with HIRA, ASF1 facilitates replication-independent deposition of histone H3.3 at transcriptionally active regions and after histone displacement. Through its involvement in histone H3 lysine 56 acetylation, ASF1 functions as an essential component of the machinery that marks newly synthesized histones and facilitates their integration into chromatin while creating "open" chromatin environments accessible to transcription machinery and repair factors. Through its interaction with the Rad53 checkpoint kinase, ASF1 couples chromatin assembly to DNA damage surveillance, ensuring that checkpoint signaling is properly extinguished when damage is repaired. Through its participation in nucleosome disassembly at promoters, ASF1 facilitates transcriptional activation while also supporting transcriptional repression through interactions with silencing complexes. The remarkable evolutionary conservation of ASF1 structure and function, combined with its extensive network of protein-protein interactions and regulatory connections to checkpoint and transcriptional regulatory pathways, establishes ASF1 as a central organizer of chromatin dynamics that integrates histone metabolism with DNA replication, repair, and transcription. Future studies examining the dynamic assembly and disassembly of ASF1 complexes in living cells and the molecular basis of its pathway selectivity will further illuminate how this ancient histone chaperone achieves such remarkable functional versatility[1][2][3][5][6][7][8][9][14][18][19][20][26][27][31][33][37][42][45][48][55][56].
Citations
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1526541/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4445473/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2423286/
- https://www.yeastgenome.org/locus/s000003651
- https://elifesciences.org/articles/23474
- https://www.pnas.org/doi/10.1073/pnas.0500149102
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1142559/
- https://pubmed.ncbi.nlm.nih.gov/15175160/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4888841/
- https://www.tandfonline.com/doi/abs/10.1128/mcb.00981-07
- https://pubmed.ncbi.nlm.nih.gov/11331602/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC312838/
- https://academic.oup.com/nar/article/40/22/11229/1148739
- https://www.pnas.org/doi/10.1073/pnas.0511050103
- https://genesdev.cshlp.org/content/15/23/3169.full
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3620686/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2767235/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2819815/
- https://www.pnas.org/doi/10.1073/pnas.0601676103
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9779303/
- https://portlandpress.com/biochemj/article/478/5/1117/227682/Structural-insights-into-histone-chaperone-Asf1
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3035992/
- https://academic.oup.com/nar/article/42/5/2906/1051901
- https://www.pnas.org/doi/10.1073/pnas.142627899
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1762009/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1084056/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3970516/
- https://www.pnas.org/doi/10.1073/pnas.0906539106
- https://www.sdbonline.org/sites/fly/genebrief/asf1.htm
- https://portlandpress.com/essaysbiochem/article/63/1/29/174/The-histone-chaperoning-pathway-from-ribosome-to
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3666882/
- https://genesdev.cshlp.org/content/19/11/1365.full.pdf
- https://pubmed.ncbi.nlm.nih.gov/15664198/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2981792/
- https://www.yeastgenome.org/reference/S000079460
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3044965/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1456847/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4167031/
- https://pubmed.ncbi.nlm.nih.gov/25264624/
- https://genesdev.cshlp.org/content/15/23/3155.long
- https://journals.plos.org/plosone/article?id=10.1371%2Fjournal.pone.0108652
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2933817/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1456080/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4448624/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3338271/
- https://elifesciences.org/articles/22799
- https://febs.onlinelibrary.wiley.com/doi/10.1111/j.1742-4658.2009.07502.x
- https://pubmed.ncbi.nlm.nih.gov/21190944/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6103222/
- https://rupress.org/jcb/article/204/1/29/37486/New-histone-supply-regulates-replication-fork
- https://www.pnas.org/doi/10.1073/pnas.0800057105
- https://genesdev.cshlp.org/content/15/9/1061.full
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7307201/