Deep Research
Falcon
(CHD1-deep-research-falcon.md)
Falcon
(CHD1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Functional Annotation of Saccharomyces cerevisiae CHD1 (UniProt P32657; gene CHD1/YER164W)
Executive summary
Saccharomyces cerevisiae CHD1 encodes Chd1, a conserved SNF2-family ATP-dependent chromatin remodeller that uses ATP hydrolysis to translocate nucleosomal DNA and thereby slide and space nucleosomes, particularly over actively transcribed gene bodies. Yeast Chd1 functions in transcription-coupled chromatin reassembly/organization, suppressing cryptic internal transcription and helping maintain proper histone-modification domains near 5′ gene ends; its action is coordinated with elongation factors and histone chaperones (notably Paf1C/Rtf1 and FACT). Mechanistic understanding has been sharpened by structural biology (cryo-EM) and by 2023–2024 biophysical and sequencing-based assays that resolve stepwise and context-dependent sliding behaviors. (lee2017theatpdependentchromatin pages 1-2, sundaramoorthy2018structureofthe pages 1-5, park2023bidirectionalnucleosomesliding pages 1-2, murawska2011chdchromatinremodelers pages 6-7)
Verification of correct gene/protein identity (disambiguation)
The evidence base summarized here is explicitly focused on yeast Chd1 and matches the UniProt target: S. cerevisiae Chd1 is described as a chromodomain-helicase-DNA-binding remodeler with tandem chromodomains, a bilobal Snf2-like ATPase motor, and a C-terminal DNA-binding module (SANT/SLIDE), consistent with UniProt P32657 annotations and the SNF2/RAD54 helicase-family assignment. The cryo-EM structure is specifically reported for Chd1 from the yeast Saccharomyces cerevisiae bound to a nucleosome. (sundaramoorthy2018structureofthe pages 1-5)
1) Key concepts and definitions (current understanding)
ATP-dependent chromatin remodeller (SNF2-family)
ATP-dependent chromatin remodellers are molecular motors that hydrolyse ATP to alter nucleosome position/structure and thereby regulate DNA accessibility. In yeast, Chd1 is classed among remodelers that can slide nucleosomes and generate regular spacing in nucleosomal arrays. (park2023bidirectionalnucleosomesliding pages 1-2, prajapati2020interplayamongatpdependent pages 1-3)
Nucleosome sliding vs spacing vs phasing
- Sliding: movement of a nucleosome along DNA, typically driven by a remodeller’s ATPase motor.
- Spacing: establishment of a characteristic nucleosome repeat length (distance between nucleosomes).
- Phasing: alignment of nucleosomes relative to a genomic “barrier” (e.g., promoter-associated complexes), producing regularly positioned +1, +2, +3… nucleosomes across a population.
A yeast-centric synthesis identifies CHD1 as one of the main enzymes shaping spacing, with distinct spacing outcomes compared with ISWI-family remodelers. (prajapati2020interplayamongatpdependent pages 5-8)
2) Molecular function and mechanism of Chd1 in budding yeast
Primary biochemical activity: ATP hydrolysis–driven nucleosome remodelling
Yeast Chd1 is a monomeric, helicase-type ATPase chromatin remodeller that engages nucleosomal DNA with its ATPase motor at superhelix location 2 (SHL2), ~20 bp from the dyad, and shifts DNA around the histone core through a stepwise translocation cycle, repositioning nucleosomes along DNA. (park2023bidirectionalnucleosomesliding pages 1-2)
Structural mechanism on the nucleosome (cryo-EM / structural biochemistry)
A cryo-EM structure of S. cerevisiae Chd1 bound to a nucleosome indicates that Chd1 can detach/unwrap ~two turns of nucleosomal DNA and binds in a catalytically poised configuration. The SANT/SLIDE DNA-binding region contacts detached/linker DNA while the ATPase engages at SHL2; chromodomain movements are linked to ATPase closure and catalysis. (sundaramoorthy2018structureofthe pages 1-5)
Complementary structural/biochemical analysis supports the importance of linker DNA engagement: Chd1 remodels in a state where its DNA-binding domain is positioned on linker DNA, and efficient repositioning depends on this DNA-binding region. (sundaramoorthy2018structureofthe pages 16-19)
Substrate context and specificity
Chd1 acts on nucleosomes (DNA wrapped on histone octamers) and is sensitive to extranucleosomal DNA context.
Quantitative and mechanistic observations include:
- Reported repositioning range: Chd1 can reposition nucleosomes 23–39 bp into a 54 bp linker (contextualized relative to ISW1a/ISW2). (sundaramoorthy2018structureofthe pages 16-19)
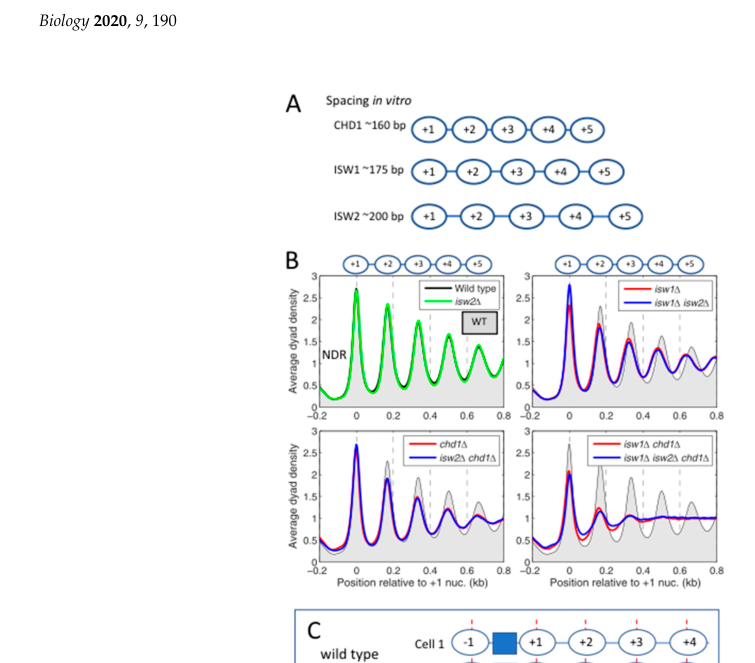
- In vitro array spacing: CHD1 establishes ~160 bp spacing, shorter than ISW1/INO80 (~175 bp) and ISW2 (~200 bp). (prajapati2020interplayamongatpdependent pages 5-8, prajapati2020interplayamongatpdependent media 42544443)
Sequence dependence and bidirectional sliding (2023 mechanistic advance)
A 2023 high-resolution sliding study using both Widom 601 and a natural S. cerevisiae +1 nucleosome sequence (SWH1) shows that although Chd1 action at one SHL2 site is unidirectional, nucleosome symmetry allows back-and-forth (bi-directional) sliding when the enzyme acts on either side. The work also shows that DNA perturbations (poly(dA:dT), mismatches, single-nucleotide insertions) become preferentially positioned about one helical turn outside SHL2, and modeling indicates strong phasing can favor ~10 bp shifts. (park2023bidirectionalnucleosomesliding pages 1-2)
3) Biological roles in yeast: transcription-coupled chromatin organization
Localization: gene bodies/coding regions (not promoters)
Yeast Chd1 is associated with transcribed gene bodies rather than promoters and is enriched over coding regions of active genes. (murawska2011chdchromatinremodelers pages 6-7)
Recruitment and interaction network in elongation
Chd1 recruitment is linked to transcription elongation machinery:
- Genome-wide analyses indicate the PAF1 complex (Paf1C) is a key determinant of Chd1 recruitment to active genes, and Chd1 occupancy concords with RNAPII Ser5-phosphorylated patterns (an early elongation-associated form). (lee2017theatpdependentchromatin pages 1-2, lee2017theatpdependentchromatin pages 2-3)
- Spt4 (DSIF component) modulates recruitment; spt4Δ increases Chd1 binding near transcription start sites, consistent with negative modulation of 5′ recruitment. (lee2017theatpdependentchromatin pages 2-3)
- Rtf1 (Paf1C subunit) is implicated in Chd1 recruitment by earlier gene-level evidence and review synthesis. (murawska2011chdchromatinremodelers pages 6-7)
Direct interaction evidence (preprint; interpret cautiously): a 2024/2025 bioRxiv preprint maps a direct Rtf1–Chd1 interaction: a short N-terminal region of Rtf1 (aa 1–30) interacts with the Chd1 CHCT domain, supported by yeast two-hybrid mapping and alanine-scan disruption of key residues. The authors propose this interaction helps distribute Chd1 across transcribed genes and influences nucleosome positioning and cryptic transcription. (tripplehorn2025adirectinteraction pages 23-26, tripplehorn2025adirectinteraction pages 1-5)
Upstream coupling to DSIF is supported by a peer-reviewed mechanistic study showing that in S. cerevisiae a domain of Rtf1 directly interacts with the Spt5 CTR and is required for proper Paf1C recruitment to active genes, consistent with an indirect path by which DSIF→Paf1C could shape Chd1 positioning. (mayekar2013therecruitmentof pages 1-2)
Functional cooperation/competition with other remodelers
Yeast chromatin organization reflects interplay among remodelers:
- ISW1 and CHD1 are described as major nucleosome-spacing enzymes that can compete to set nucleosome spacing; loss of both produces major disruption partly due to close-packed dinucleosomes. (prajapati2020interplayamongatpdependent pages 1-3)
- Genetic/functional synthesis indicates cryptic transcription and altered nucleosome spacing are exacerbated in chd1 isw1 double mutants, consistent with partially redundant roles in maintaining gene-body chromatin integrity. (murawska2011chdchromatinremodelers pages 6-7)
4) Effects on histone modification domains and RNA processing (quantitative genome-wide findings)
H3K4me3/H3K36me3 boundary maintenance
A genome-wide study reports that loss of CHD1 yields significantly aberrant H3K4me3/H3K36me3 patterns across ~half of the yeast genome, predominantly within ~1 kb of transcription start sites, with reciprocal changes between promoter-proximal (+1–+3) and more distal (+4–+6) nucleosomes. (lee2017theatpdependentchromatin pages 6-9)
Mechanistically, structural work suggests that Chd1-induced partial DNA unwrapping could alter display/accessibility of histone tail epitopes and thereby influence writer/eraser/reader enzymes, consistent with observed mark-boundary effects. (sundaramoorthy2018structureofthe pages 16-19)
Splicing/intron retention effects
CHD1 deletion is associated with altered co-transcriptional RNA processing:
- In one RNA-seq analysis, 35 introns were significantly affected, with 28/35 (80%) showing lower intron retention (improved splicing) in chd1Δ. (lee2017theatpdependentchromatin pages 6-9)
- Reanalysis with deeper RNA-seq indicated 94% of introns show lower intron retention in chd1Δ, with stronger effects in ribosomal protein genes where Chd1 is highly enriched. (lee2017theatpdependentchromatin pages 6-9)
These data support a model in which Chd1 influences elongation-coupled chromatin state and thereby indirectly impacts splicing efficiency. (lee2017theatpdependentchromatin pages 6-9)
5) Regulation by histone modifications and nucleosome features
H2B ubiquitination
Structural/biochemical work indicates that H2B K120 (yeast K123) ubiquitination can stimulate Chd1 activity ~2-fold in vitro and is enriched in coding regions. The same work discusses an estimated ~10% occupancy of the transiently unwrapped nucleosomal DNA state (baseline), and proposes that ubiquitin repositioning upon Chd1 binding stabilizes this unwrapped configuration, potentially enhancing DNA accessibility. (sundaramoorthy2018structureofthe pages 16-19)
H3K36me3 context
Mimicking H3K36 trimethylation (via alkylation) modestly stimulates Chd1 activity, but the structural state captured did not show a stable direct interaction of the H3K36 region with the chromodomains, suggesting either weak/transient recognition or a requirement for alternative conformations. (sundaramoorthy2018structureofthe pages 16-19)
6) Recent developments and expert synthesis (prioritizing 2023–2024)
2023: High-resolution mapping of remodeler-driven sliding outcomes
The 2023 Slide-seq study provides a modern mechanistic picture of how Chd1 integrates intrinsic DNA features and ATP-driven remodeling, including a concrete explanation for how a nominally unidirectional motor can yield bi-directional outcomes through nucleosome symmetry and repeated engagement of SHL2 sites. (park2023bidirectionalnucleosomesliding pages 1-2)
2024: Mechanistic consensus emerging from cryo-EM + single-molecule work (general; cross-species)
Two 2024 reviews summarize the field-wide shift toward a consensus “inside-the-box” translocation mechanism:
- A 2024 Biochemical Society Transactions review emphasizes that cryo-EM and single-molecule methods have only recently brought nucleosome-sliding mechanisms into focus and argues that CHD-family enzymes share a central remodeling mechanism with other remodelers, while auxiliary domains create family-specific behaviors. (reid2024howdoeschd4 pages 1-2)
- The same review synthesizes a stepwise model in which nucleotide-state changes in the RecA-like ATPase lobes produce incremental tracking-strand shifts and burst-like DNA release, and notes that comparable asymmetric stepping behaviors have been observed for multiple remodelers including CHD1 in related work (general, not yeast-specific). (reid2024howdoeschd4 pages 5-6)
- A 2024 biophysics-focused review highlights how cryo-EM and single-molecule FRET studies help explain apparent directional bias in remodeler action (including Chd1) as an emergent property of motor architecture plus substrate asymmetry (e.g., differential unwrapping propensity of the Widom 601 sequence). (klempahn2024chromatinremodelersa pages 2-5)
A high-level 2024 Nature Reviews Genetics synthesis (mammalian-focused) further underscores that structural/biochemical studies are increasingly defining how remodelers contact nucleosomes and other regulators; although not yeast-specific, it supports the broader interpretive framework for how remodeler specificity emerges from interaction surfaces and dynamic localization. (gourisankar2024contextspecificfunctionsof pages 1-3)
7) Current applications and real-world implementations
Yeast as a mechanistic model system
The most established “real-world implementation” of yeast CHD1 research is its use as a model system for chromatin remodeling mechanisms and for interpreting high-throughput chromatin maps. A yeast-focused review emphasizes yeast’s advantages (small genome, tractable mutants) for chromatin organization studies, including nucleosome spacing/phasing and remodeler competition models. (prajapati2020interplayamongatpdependent pages 5-8)
Practical implications for genomic assays and synthetic biology contexts (evidence-limited)
Chd1-dependent spacing/organization is directly relevant to:
- designing and interpreting MNase-seq/chemical mapping and nucleosome-repeat-length analyses (e.g., distinguishing remodeler-set spacing regimes), and
- conceptual engineering of transcriptional programs where chromatin reassembly and cryptic transcription suppression matter.
However, the retrieved literature does not establish a specific industrial/clinical deployment of yeast CHD1 itself; its application is primarily as a foundational mechanistic tool and model. (prajapati2020interplayamongatpdependent pages 5-8, murawska2011chdchromatinremodelers pages 6-7)
Evidence-mapped summary table
The following table compiles key functional claims with quantitative values and source types.
| Functional aspect | Key findings | Evidence type | Primary source(s) with year, DOI URL |
|---|---|---|---|
| Target identity / domain architecture | The literature matches Saccharomyces cerevisiae Chd1, the ATP-dependent chromatin remodeler encoded by CHD1/YER164W. Core architecture reported for yeast Chd1: tandem chromodomains, bilobal Snf2-family ATPase motor, and C-terminal SANT/SLIDE DNA-binding region; reviews also describe CHD-family flanking regulatory domains. This aligns with UniProt P32657 annotation as a SNF2/RAD54-family chromodomain helicase DNA-binding protein. (sundaramoorthy2018structureofthe pages 1-5, murawska2011chdchromatinremodelers pages 6-7) | Structural, review | Farnung et al., 2017, Nature, https://doi.org/10.1038/nature24046; Murawska & Brehm, 2011, Transcription, https://doi.org/10.4161/trns.2.6.17840 |
| Primary biochemical activity | Chd1 is an ATP-dependent chromatin remodeler that repositions/slides nucleosomes and spaces nucleosomal arrays. Chd1 acts on nucleosomal DNA at SHL2 (~20 bp from the dyad) and can assemble/reposition nucleosomes into evenly spaced arrays. (park2023bidirectionalnucleosomesliding pages 1-2) | Biochemical, mechanistic | Park et al., 2023, Nucleic Acids Research, https://doi.org/10.1093/nar/gkad738 |
| In vitro spacing specificity | In comparative in vitro spacing assays, CHD1 establishes the shortest average nucleosome spacing (~160 bp), compared with ~175 bp for ISW1/INO80 and ~200 bp for ISW2. Yeast average in vivo spacing is ~165 bp. (prajapati2020interplayamongatpdependent pages 5-8, prajapati2020interplayamongatpdependent media 42544443) | Review of primary spacing assays | Prajapati et al., 2020, Biology, https://doi.org/10.3390/biology9080190 |
| Structural mechanism on nucleosomes | Cryo-EM of S. cerevisiae Chd1 bound to the nucleosome shows the ATPase at SHL +2, the SANT/SLIDE DNA-binding region contacting linker DNA near SHL −7, and Chd1 detaching/unwrapping two turns of DNA from the histone octamer. This provides a direct structural explanation for ATP-coupled remodeling. (sundaramoorthy2018structureofthe pages 1-5) | Structural | Farnung et al., 2017, Nature, https://doi.org/10.1038/nature24046; Sundaramoorthy et al., 2018, eLife, https://doi.org/10.1101/290874 |
| Repositioning range / linker dependence | Text summarizing yeast Chd1 remodeling reports that Chd1 can reposition nucleosomes 23–39 bp into a 54 bp linker, similar to ISW1a/ISW2, supporting active engagement of extranucleosomal linker DNA by the DNA-binding domain. (sundaramoorthy2018structureofthe pages 16-19) | Structural discussion with prior biochemical reference | Sundaramoorthy et al., 2018, eLife, https://doi.org/10.1101/290874 |
| Sliding directionality | Chd1 translocates DNA unidirectionally toward the dyad from a single SHL2 site, but because nucleosomes have two symmetric SHL2 sites, the net outcome can be back-and-forth/bidirectional sliding. DNA perturbations tend to localize about one helical turn (~10 bp) outside SHL2, and strong phasing can favor ~10 bp shifts. (park2023bidirectionalnucleosomesliding pages 1-2) | Biophysical, sequencing-based mechanistic study | Park et al., 2023, Nucleic Acids Research, https://doi.org/10.1093/nar/gkad738 |
| DNA-sequence sensitivity / substrate preference | On the Widom 601 sequence, Chd1 preferentially shifts the dyad toward the TA-poor side; introducing long poly(dA:dT) tracts on that side reverses the preferred sliding direction. Similar principles were observed using the natural S. cerevisiae SWH1 +1 nucleosome sequence. (park2023bidirectionalnucleosomesliding pages 1-2) | Biophysical, substrate preference | Park et al., 2023, Nucleic Acids Research, https://doi.org/10.1093/nar/gkad738 |
| Cellular localization / chromatin context | Yeast Chd1 is enriched in the nucleus on transcribed gene bodies/coding regions, rather than promoters, and is associated with active transcription. It tracks especially well with highly transcribed genes and with RNAPII Ser5-phosphorylated occupancy patterns. (murawska2011chdchromatinremodelers pages 6-7, lee2017theatpdependentchromatin pages 1-2) | Genome-wide, review | Lee et al., 2017, Nucleic Acids Research, https://doi.org/10.1093/nar/gkx636; Murawska & Brehm, 2011, Transcription, https://doi.org/10.4161/trns.2.6.17840 |
| Recruitment by elongation machinery | Recruitment is strongly linked to transcription elongation factors. Chd1 association decreases in Rtf1/PAF1C mutants, supporting recruitment by PAF1C; Spt4 can oppose/promote redistribution near 5′ ends depending on context. Chd1 also interacts functionally with FACT (Spt16-Pob3) and transcription elongation machinery. (murawska2011chdchromatinremodelers pages 6-7, lee2017theatpdependentchromatin pages 1-2, lee2017theatpdependentchromatin pages 2-3) | Genome-wide, genetic, review | Lee et al., 2017, Nucleic Acids Research, https://doi.org/10.1093/nar/gkx636; Murawska & Brehm, 2011, Transcription, https://doi.org/10.4161/trns.2.6.17840 |
| Histone mark recognition / chromodomain function | In budding yeast, Chd1 contributes to maintaining H3K4me3/H3K36me3 domain boundaries, but Lee et al. report yeast Chd1 does not directly bind H3K4me3 the way human CHD1 can. A H3K36me3 mimic modestly stimulates Chd1 activity, but the structural study did not support a stable direct H3K36-tail interaction in the captured state. (lee2017theatpdependentchromatin pages 1-2, sundaramoorthy2018structureofthe pages 16-19) | Genome-wide, structural/biochemical | Lee et al., 2017, Nucleic Acids Research, https://doi.org/10.1093/nar/gkx636; Sundaramoorthy et al., 2018, eLife, https://doi.org/10.1101/290874 |
| Modulation by H2B ubiquitination | H2BK120/K123 ubiquitination stimulates Chd1 activity ~2-fold in vitro. In the Chd1-bound unwrapped nucleosome, ubiquitin is repositioned toward DNA; the unwrapped nucleosome state is estimated to have ~10% occupancy in the absence of stabilizing factors, and ubiquitin is proposed to stabilize this transiently unwrapped state. Chd1 mutants also show reduced H2B ubiquitination. (sundaramoorthy2018structureofthe pages 1-5, sundaramoorthy2018structureofthe pages 16-19) | Structural, biochemical | Sundaramoorthy et al., 2018, eLife, https://doi.org/10.1101/290874 |
| Role in transcription-coupled chromatin restoration | Chd1 is thought to help re-establish nucleosome organization after RNA polymerase II passage, maintaining chromatin structure over coding regions and preventing inappropriate exposure of internal promoter-like DNA. (lee2017theatpdependentchromatin pages 1-2, park2023bidirectionalnucleosomesliding pages 1-2, murawska2011chdchromatinremodelers pages 6-7) | Genome-wide, mechanistic review | Lee et al., 2017, Nucleic Acids Research, https://doi.org/10.1093/nar/gkx636; Murawska & Brehm, 2011, Transcription, https://doi.org/10.4161/trns.2.6.17840 |
| Cryptic transcription suppression | chd1 mutants show initiation from cryptic internal promoters; this defect is strongly enhanced in chd1 isw1 double mutants, indicating partially redundant roles of Chd1 and Isw1 in preserving coding-region chromatin integrity. (murawska2011chdchromatinremodelers pages 6-7, prajapati2020interplayamongatpdependent pages 1-3) | Genetic, review | Murawska & Brehm, 2011, Transcription, https://doi.org/10.4161/trns.2.6.17840; Prajapati et al., 2020, Biology, https://doi.org/10.3390/biology9080190 |
| Nucleosome organization phenotypes in mutants | Loss of both ISW1 and CHD1 causes major chromatin disruption and formation of close-packed dinucleosomes. chd1Δ alone reduces phasing beyond the +1 nucleosome, whereas double loss causes much stronger disorganization, especially on highly transcribed genes. (tripplehorn2025adirectinteraction pages 1-5, prajapati2020interplayamongatpdependent pages 1-3) | Genetic, review | Prajapati et al., 2020, Biology, https://doi.org/10.3390/biology9080190 |
| Effects on histone modification domains | Loss of CHD1 causes widespread, reciprocal disruption of H3K4me3 and H3K36me3 near the 5′ ends of genes, concentrated within ~1 kb of the TSS and affecting approximately half of the yeast genome. (lee2017theatpdependentchromatin pages 2-3, lee2017theatpdependentchromatin pages 6-9) | Genome-wide ChIP-seq / RNA-seq | Lee et al., 2017, Nucleic Acids Research, https://doi.org/10.1093/nar/gkx636 |
| Effects on intron retention / splicing-linked transcription | RNA-seq showed 35 introns significantly affected in chd1Δ, with 28/35 (80%) showing lower intron retention (improved splicing). Reanalysis with deeper RNA-seq found 94% of introns had lower retention in chd1Δ, with the effect especially clear in ribosomal protein genes where Chd1 is enriched. (lee2017theatpdependentchromatin pages 6-9) | Genome-wide RNA-seq | Lee et al., 2017, Nucleic Acids Research, https://doi.org/10.1093/nar/gkx636 |
| Relationship with FACT / other partners | Chd1 functionally interacts with FACT (Spt16-Pob3), PAF1 complex/Rtf1, and Spt4-Spt5 elongation factors; these interactions place Chd1 in the transcription-coupled chromatin reassembly pathway rather than acting primarily by changing RNAPII processivity directly. (lee2017theatpdependentchromatin pages 1-2, sundaramoorthy2018structureofthe pages 1-5, murawska2011chdchromatinremodelers pages 6-7) | Genome-wide, structural context, review | Lee et al., 2017, Nucleic Acids Research, https://doi.org/10.1093/nar/gkx636; Sundaramoorthy et al., 2018, eLife, https://doi.org/10.1101/290874; Murawska & Brehm, 2011, Transcription, https://doi.org/10.4161/trns.2.6.17840 |
Table: This table summarizes functional annotation evidence for Saccharomyces cerevisiae CHD1 (UniProt P32657), including biochemical activity, structural mechanism, recruitment, localization, mutant phenotypes, and key quantitative findings. It is useful as a compact evidence map linking specific claims to source types and DOI-resolved references.
Visual evidence: remodeler-specific nucleosome spacing
Prajapati et al. (2020) Figure 4 summarizes how different yeast remodelers generate distinct nucleosome spacing in vitro, including the short spacing attributed to CHD1.
(prajapati2020interplayamongatpdependent media 42544443)
Limitations and evidence quality notes
- Several key yeast CHD1 functional conclusions are anchored in peer-reviewed primary studies and reviews (e.g., structural and genome-wide functional work). (sundaramoorthy2018structureofthe pages 1-5, lee2017theatpdependentchromatin pages 6-9, murawska2011chdchromatinremodelers pages 6-7)
- The most specific direct Rtf1–Chd1 interaction mapping available in this evidence set is from a bioRxiv preprint (2024/2025); conclusions drawn from it should be considered provisional until peer-reviewed publication, but it is mechanistically consistent with earlier recruitment models. (tripplehorn2025adirectinteraction pages 23-26, tripplehorn2025adirectinteraction pages 1-5)
Key references (with dates and URLs)
- Park S. et al. “Bi-directional nucleosome sliding by the Chd1 chromatin remodeler…” Published online 20 Sep 2023. Nucleic Acids Research. https://doi.org/10.1093/nar/gkad738 (park2023bidirectionalnucleosomesliding pages 1-2)
- Klempahn S. et al. “Chromatin remodelers: a concise introduction for biophysicists.” Jun 2024. Biophysical Reviews. https://doi.org/10.1007/s12551-024-01199-1 (klempahn2024chromatinremodelersa pages 2-5)
- Reid X.J. et al. “How does CHD4 slide nucleosomes?” 2 Sep 2024. Biochemical Society Transactions. https://doi.org/10.1042/BST20230070 (reid2024howdoeschd4 pages 1-2)
- Lee Y. et al. “The ATP-dependent chromatin remodeler Chd1…” Jul 2017. Nucleic Acids Research. https://doi.org/10.1093/nar/gkx636 (lee2017theatpdependentchromatin pages 1-2)
- Sundaramoorthy R. et al. “Structure of the chromatin remodelling enzyme Chd1 bound to a ubiquitinylated nucleosome.” Posted 30 Mar 2018 (preprint version in this corpus; eLife-associated). https://doi.org/10.1101/290874 (sundaramoorthy2018structureofthe pages 16-19)
- Prajapati H.K. et al. “Interplay among ATP-Dependent Chromatin Remodelers Determines Chromatin Organisation in Yeast.” Jul 2020. Biology. https://doi.org/10.3390/biology9080190 (prajapati2020interplayamongatpdependent pages 5-8)
- Murawska M. & Brehm A. “CHD chromatin remodelers and the transcription cycle.” Nov 2011. Transcription. https://doi.org/10.4161/trns.2.6.17840 (murawska2011chdchromatinremodelers pages 6-7)
- Mayekar M.K. et al. “The Recruitment of the S. cerevisiae Paf1 Complex…” Aug 2013. Molecular and Cellular Biology. https://doi.org/10.1128/mcb.00270-13 (mayekar2013therecruitmentof pages 1-2)
References
-
(lee2017theatpdependentchromatin pages 1-2): Yaelim Lee, Daechan Park, and Vishwanath R. Iyer. The atp-dependent chromatin remodeler chd1 is recruited by transcription elongation factors and maintains h3k4me3/h3k36me3 domains at actively transcribed and spliced genes. Nucleic Acids Research, 45:8646-8646, Jul 2017. URL: https://doi.org/10.1093/nar/gkx636, doi:10.1093/nar/gkx636. This article has 70 citations and is from a highest quality peer-reviewed journal.
-
(sundaramoorthy2018structureofthe pages 1-5): Ramasubramanian Sundaramoorthy, Amanda L. Hughes, Hassane El-Mkami, David Norman, and Tom Owen-Hughes. Structure of the chromatin remodelling enzyme chd1 bound to a ubiquitinylated nucleosome. eLife, Mar 2018. URL: https://doi.org/10.1101/290874, doi:10.1101/290874. This article has 100 citations and is from a domain leading peer-reviewed journal.
-
(park2023bidirectionalnucleosomesliding pages 1-2): Sangwoo Park, Giovanni B Brandani, Taekjip Ha, and Gregory D Bowman. Bi-directional nucleosome sliding by the chd1 chromatin remodeler integrates intrinsic sequence-dependent and atp-dependent nucleosome positioning. Nucleic Acids Research, 51:10326-10343, Sep 2023. URL: https://doi.org/10.1093/nar/gkad738, doi:10.1093/nar/gkad738. This article has 23 citations and is from a highest quality peer-reviewed journal.
-
(murawska2011chdchromatinremodelers pages 6-7): Magdalena Murawska and Alexander Brehm. Chd chromatin remodelers and the transcription cycle. Transcription, 2:244-253, Nov 2011. URL: https://doi.org/10.4161/trns.2.6.17840, doi:10.4161/trns.2.6.17840. This article has 162 citations and is from a peer-reviewed journal.
-
(prajapati2020interplayamongatpdependent pages 1-3): Hemant K. Prajapati, Josefina Ocampo, and David J. Clark. Interplay among atp-dependent chromatin remodelers determines chromatin organisation in yeast. Biology, 9:190, Jul 2020. URL: https://doi.org/10.3390/biology9080190, doi:10.3390/biology9080190. This article has 43 citations.
-
(prajapati2020interplayamongatpdependent pages 5-8): Hemant K. Prajapati, Josefina Ocampo, and David J. Clark. Interplay among atp-dependent chromatin remodelers determines chromatin organisation in yeast. Biology, 9:190, Jul 2020. URL: https://doi.org/10.3390/biology9080190, doi:10.3390/biology9080190. This article has 43 citations.
-
(sundaramoorthy2018structureofthe pages 16-19): Ramasubramanian Sundaramoorthy, Amanda L. Hughes, Hassane El-Mkami, David Norman, and Tom Owen-Hughes. Structure of the chromatin remodelling enzyme chd1 bound to a ubiquitinylated nucleosome. eLife, Mar 2018. URL: https://doi.org/10.1101/290874, doi:10.1101/290874. This article has 100 citations and is from a domain leading peer-reviewed journal.
-
(prajapati2020interplayamongatpdependent media 42544443): Hemant K. Prajapati, Josefina Ocampo, and David J. Clark. Interplay among atp-dependent chromatin remodelers determines chromatin organisation in yeast. Biology, 9:190, Jul 2020. URL: https://doi.org/10.3390/biology9080190, doi:10.3390/biology9080190. This article has 43 citations.
-
(lee2017theatpdependentchromatin pages 2-3): Yaelim Lee, Daechan Park, and Vishwanath R. Iyer. The atp-dependent chromatin remodeler chd1 is recruited by transcription elongation factors and maintains h3k4me3/h3k36me3 domains at actively transcribed and spliced genes. Nucleic Acids Research, 45:8646-8646, Jul 2017. URL: https://doi.org/10.1093/nar/gkx636, doi:10.1093/nar/gkx636. This article has 70 citations and is from a highest quality peer-reviewed journal.
-
(tripplehorn2025adirectinteraction pages 23-26): Sarah A. Tripplehorn, Margaret K. Shirra, Santana M. Lardo, Hannah G. Marvil, Sarah J. Hainer, and Karen M. Arndt. A direct interaction between the chd1 chct domain and rtf1 controls chd1 distribution and nucleosome positioning on active genes. bioRxiv, Dec 2025. URL: https://doi.org/10.1101/2024.12.06.627179, doi:10.1101/2024.12.06.627179. This article has 7 citations.
-
(tripplehorn2025adirectinteraction pages 1-5): Sarah A. Tripplehorn, Margaret K. Shirra, Santana M. Lardo, Hannah G. Marvil, Sarah J. Hainer, and Karen M. Arndt. A direct interaction between the chd1 chct domain and rtf1 controls chd1 distribution and nucleosome positioning on active genes. bioRxiv, Dec 2025. URL: https://doi.org/10.1101/2024.12.06.627179, doi:10.1101/2024.12.06.627179. This article has 7 citations.
-
(mayekar2013therecruitmentof pages 1-2): Manasi K. Mayekar, Richard G. Gardner, and Karen M. Arndt. The recruitment of the saccharomyces cerevisiae paf1 complex to active genes requires a domain of rtf1 that directly interacts with the spt4-spt5 complex. Molecular and Cellular Biology, 33:3259-3273, Aug 2013. URL: https://doi.org/10.1128/mcb.00270-13, doi:10.1128/mcb.00270-13. This article has 87 citations and is from a domain leading peer-reviewed journal.
-
(lee2017theatpdependentchromatin pages 6-9): Yaelim Lee, Daechan Park, and Vishwanath R. Iyer. The atp-dependent chromatin remodeler chd1 is recruited by transcription elongation factors and maintains h3k4me3/h3k36me3 domains at actively transcribed and spliced genes. Nucleic Acids Research, 45:8646-8646, Jul 2017. URL: https://doi.org/10.1093/nar/gkx636, doi:10.1093/nar/gkx636. This article has 70 citations and is from a highest quality peer-reviewed journal.
-
(reid2024howdoeschd4 pages 1-2): Xavier J. Reid, Yichen Zhong, and Joel P. Mackay. How does chd4 slide nucleosomes? Biochemical Society Transactions, 52:1995-2008, Sep 2024. URL: https://doi.org/10.1042/bst20230070, doi:10.1042/bst20230070. This article has 9 citations and is from a peer-reviewed journal.
-
(reid2024howdoeschd4 pages 5-6): Xavier J. Reid, Yichen Zhong, and Joel P. Mackay. How does chd4 slide nucleosomes? Biochemical Society Transactions, 52:1995-2008, Sep 2024. URL: https://doi.org/10.1042/bst20230070, doi:10.1042/bst20230070. This article has 9 citations and is from a peer-reviewed journal.
-
(klempahn2024chromatinremodelersa pages 2-5): Sophie Klempahn, Helmut Schiessel, and Ralf Blossey. Chromatin remodelers: a concise introduction for biophysicists. Biophysical reviews, 16 3:357-363, Jun 2024. URL: https://doi.org/10.1007/s12551-024-01199-1, doi:10.1007/s12551-024-01199-1. This article has 2 citations and is from a peer-reviewed journal.
-
(gourisankar2024contextspecificfunctionsof pages 1-3): Sai Gourisankar, Andrey Krokhotin, Wendy Wenderski, and Gerald R. Crabtree. Context-specific functions of chromatin remodellers in development and disease. Nature Reviews Genetics, 25:340-361, Nov 2024. URL: https://doi.org/10.1038/s41576-023-00666-x, doi:10.1038/s41576-023-00666-x. This article has 75 citations and is from a domain leading peer-reviewed journal.
Artifacts
Citations
- sundaramoorthy2018structureofthe pages 1-5
- prajapati2020interplayamongatpdependent pages 5-8
- park2023bidirectionalnucleosomesliding pages 1-2
- sundaramoorthy2018structureofthe pages 16-19
- murawska2011chdchromatinremodelers pages 6-7
- lee2017theatpdependentchromatin pages 2-3
- mayekar2013therecruitmentof pages 1-2
- prajapati2020interplayamongatpdependent pages 1-3
- lee2017theatpdependentchromatin pages 6-9
- klempahn2024chromatinremodelersa pages 2-5
- gourisankar2024contextspecificfunctionsof pages 1-3
- lee2017theatpdependentchromatin pages 1-2
- tripplehorn2025adirectinteraction pages 23-26
- tripplehorn2025adirectinteraction pages 1-5
- https://doi.org/10.1038/nature24046;
- https://doi.org/10.4161/trns.2.6.17840
- https://doi.org/10.1093/nar/gkad738
- https://doi.org/10.3390/biology9080190
- https://doi.org/10.1101/290874
- https://doi.org/10.1093/nar/gkx636;
- https://doi.org/10.4161/trns.2.6.17840;
- https://doi.org/10.1093/nar/gkx636
- https://doi.org/10.1101/290874;
- https://doi.org/10.1007/s12551-024-01199-1
- https://doi.org/10.1042/BST20230070
- https://doi.org/10.1128/mcb.00270-13
- https://doi.org/10.1093/nar/gkx636,
- https://doi.org/10.1101/290874,
- https://doi.org/10.1093/nar/gkad738,
- https://doi.org/10.4161/trns.2.6.17840,
- https://doi.org/10.3390/biology9080190,
- https://doi.org/10.1101/2024.12.06.627179,
- https://doi.org/10.1128/mcb.00270-13,
- https://doi.org/10.1042/bst20230070,
- https://doi.org/10.1007/s12551-024-01199-1,
- https://doi.org/10.1038/s41576-023-00666-x,
Perplexity
(CHD1-deep-research-perplexity.md)
Perplexity
(CHD1-deep-research-perplexity.md)Comprehensive Functional Annotation of CHD1: An ATP-Dependent Chromatin Remodeling Factor in Saccharomyces cerevisiae
The chromodomain-helicase DNA-binding protein 1 (CHD1) represents a critical ATP-dependent chromatin remodeling enzyme that functions as a monomeric protein in Saccharomyces cerevisiae, distinguished by its dual chromodomains, an SNF2-related ATPase catalytic core, and a C-terminal DNA-binding domain comprising SANT and SLIDE motifs[1][9]. This enzyme has emerged as a central regulator of chromatin architecture during transcription elongation, maintaining proper nucleosome positioning and spacing while facilitating the passage of RNA polymerase II through nucleosomal obstacles. The discovery of high-resolution cryo-electron microscopy structures between 2017 and 2025 has provided unprecedented molecular insight into how CHD1 engages nucleosomes and executes its remodeling activities, revealing that the protein uses ATP hydrolysis to selectively unwrap nucleosomal DNA and translocate the histone octamer along the DNA template[7][10]. Beyond transcription, CHD1 participates in homologous recombination-based DNA repair, assists in establishing proper heterochromatin structure, and maintains boundaries between distinct histone modification domains that regulate gene expression patterns. The functional specialization of CHD1 demonstrates remarkable evolutionary conservation across eukaryotic organisms, with humans expressing nine CHD family members that diverged into distinct subfamilies with partially overlapping yet specialized functions.
Protein Architecture and Structural Organization
Domains and Their Arrangement
CHD1 is constructed from several evolutionarily conserved functional domains arranged in a specific three-dimensional architecture that enables its multifaceted regulatory functions[6][9][14]. The protein spans approximately 1,274 amino acids in yeast and contains its defining structural elements arranged from the N-terminus to C-terminus as follows: a pair of tandem chromodomains at the N-terminus, a central SNF2-related helicase domain containing conserved ATP-binding and catalytic motifs, a helicase C domain, and a C-terminal DNA-binding domain[9][31]. The double chromodomains, which are characteristic of all CHD proteins, serve as regulatory platforms that mediate interactions with chromatin components and influence the overall conformational state of the enzyme[14]. These chromodomains, while present in yeast CHD1, do not bind to methylated histone H3K4 as they do in human CHD1, representing a notable divergence in substrate recognition mechanisms between yeast and mammalian orthologues[12][31].
The catalytic ATPase domain is remarkably conserved across all SNF2-family chromatin remodelers and consists of two tandem RecA-like folds containing seven conserved helicase-related sequence motifs that classify the enzyme within Superfamily 2 (SF2) of helicase-like proteins[17][39]. However, unlike true helicases, CHD1 and its SNF2-family relatives cannot separate nucleic acid strands in the manner characteristic of classical helicases; rather, they couple ATP hydrolysis to mechanical rearrangement of nucleosomal structure[17]. The two lobes of the ATPase domain undergo a characteristic ratcheting motion during the catalytic cycle that drives DNA translocation, with the lobes containing additional helices and structural protrusions not found in single-strand translocases, enabling the simultaneous engagement with both DNA strands of the nucleosomal DNA[42][58].
The C-terminal DNA-binding domain revealed through crystallographic studies contains structural homology to SANT and SLIDE domains previously identified in ISWI remodeling enzymes[6][17]. These domains function as a DNA-binding module that engages extranucleosomal DNA flanking the nucleosome core particle, with the SLIDE domain particularly important for nucleosome spacing activity[6][56]. The SANT domain provides histone-binding capacity found in ISWI remodelers, though CHD1 exhibits relatively minimal direct contacts with histone proteins and instead relies predominantly on DNA interactions to achieve nucleosome repositioning[34][58]. Between the ATPase motor and DNA-binding domain lies a polypeptide segment containing residues critical for coupling nucleosome-stimulated ATP hydrolysis to productive nucleosome sliding, as demonstrated through systematic mutagenesis studies[8].
Structural Conformational Changes Upon Nucleosome Binding
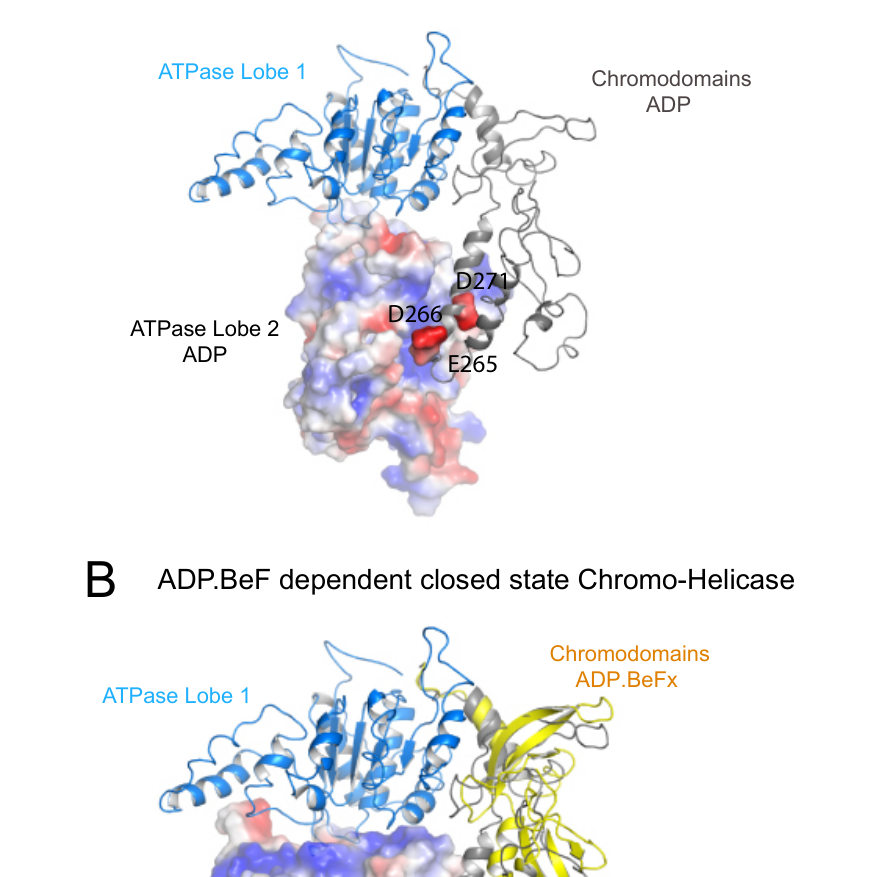
The cryo-EM structures of CHD1 bound to nucleosomes have revealed dramatic conformational reorganization that occurs as the enzyme transitions from its solution state to an active, nucleosome-engaged state[7][10][15][24]. In the apo state before nucleosome engagement, the tandem chromodomains present a relatively open configuration that physically impedes DNA binding to the ATPase domains, effectively maintaining the enzyme in an auto-inhibited state[24]. Upon nucleosome binding in the presence of ATP or non-hydrolyzable ATP analogues, the chromodomains undergo an approximately 18-degree rotation relative to their orientation in crystal structures of the isolated catalytic core[10][15]. This repositioning orients chromodomain I toward the nucleosomal DNA at superhelical location -1 (SHL-1), enabling productive engagement with the nucleosome surface[10][34].
The DNA-binding domain similarly undergoes substantial repositioning during this transition, moving from a peripheral position in the apo state to a configuration where the SANT and SLIDE domains bind tightly to the first 10 base pairs of linker DNA immediately adjacent to the nucleosome core particle[24][55]. This engagement of extranucleosomal DNA is functionally critical, as studies utilizing artificially constrained linker DNA demonstrate that nucleosome sliding is directed toward the longer linker when choices are available, and the DNA-binding domain affinity for extranucleosomal DNA constitutes the primary determinant of remodeling directionality[56]. Importantly, the conformational changes observed upon nucleosome binding are mechanistically connected, with the repositioning of chromodomains linked through the central ATPase domains to enable closure of the ATPase catalytic pocket, bringing conserved residues into alignment for ATP hydrolysis[34][42][58].
Catalytic Mechanism and Nucleosome Remodeling Activity
ATP Hydrolysis-Coupled DNA Unwrapping
The mechanism of nucleosome remodeling by CHD1 involves a sophisticated coupling of ATP hydrolysis to mechanical manipulation of the nucleosomal DNA structure[7]. High-resolution cryo-EM structures obtained at 4.8 Ångstrom resolution with transition state-mimicking compounds such as ADP·BeF₃ have captured CHD1 in a state poised for catalysis, revealing that two complete turns of DNA at superhelical locations -5 to -7 become detached from the histone octamer surface as the enzyme engages the nucleosome[7]. This DNA unwrapping fundamentally alters the trajectory of extranucleosomal DNA by approximately 60 degrees and disrupts DNA interactions with histone proteins H2A, H2B, and H3 that normally stabilize the wrapped configuration[7][53].
The unwrapping of nucleosomal DNA at the entry side of the nucleosome is nucleotide-dependent, requiring the binding of ATP or ATP analogues to proceed[7][15][53]. Biochemical experiments demonstrate that ATP binding to yeast CHD1 induces transient unwrapping of DNA on the exit side of the nucleosome, with this unwrapping specifically facilitating nucleosome translocation[50]. The process differs fundamentally from histone octamer ejection or histone variant exchange, as the histone octamer itself remains intact during CHD1-mediated remodeling and adopts only minimal conformational changes in comparison to remodeling complexes such as ACF[7]. This preservation of histone octamer integrity underscores CHD1's role as a nucleosome spacing enzyme rather than a nucleosome disruption or assembly factor.
Ratcheting Mechanism and DNA Translocation
The mechanism driving nucleosome repositioning involves a ratcheting cycle of the two lobes of the ATPase domain that moves along the DNA template in a directional manner[7][42]. The first DNA gyre at the entry side becomes engaged by the ATPase motor at superhelical location (SHL) +2, where motifs within lobe one contact one DNA strand while motifs within lobe two contact the opposing strand[42][58]. This bifurcated engagement with both strands enables the ATPase lobes to grip the DNA and drive its translocation toward the nucleosome dyad through conformational changes driven by ATP hydrolysis[42][58]. In contrast to the related Snf2 ATPase structure, CHD1 contacts DNA at specific motifs (motifs II and III within lobe one) that contact the opposite DNA strand, and these motifs undergo a similar ratcheting motion that would be anticipated to drive nucleosomal DNA movement[42].
The histone H4 tail plays an essential regulatory role in this catalytic process, with deletion or mutation of the H4 N-terminal tail substantially reducing both nucleosome sliding activity and ATPase activity[15][18]. Specifically, conserved acidic residues D729 and E669 in CHD1 contact the H4 tail, and these contacts are conserved across multiple classes of remodeling enzymes, suggesting a fundamental mechanistic principle[15]. The repositioning of ATPase lobe II upon ATP binding enables critical contacts with the histone H3 alpha-one helix and the histone H4 tail, stabilizing the nucleosome-bound enzyme in a catalytically competent state[15][34][58]. Furthermore, the interaction between the SANT and SLIDE DNA-binding domains with linker DNA is mechanistically coupled through the chromodomains to the ATP-dependent closure of the ATPase motor domains, providing molecular connectivity between linker DNA availability and motor domain activity that may regulate the rate of nucleosome movement[34][59].
Nucleosome Spacing and Periodic Array Formation
CHD1 possesses a characteristic activity to generate and maintain regularly spaced nucleosomal arrays with relatively constant spacing independent of nucleosome density, a property termed "clamping activity"[28]. In purified in vitro systems, CHD1 can convert irregularly spaced polynucleosomes into evenly spaced regular arrays with near-physiological repeat lengths of approximately 160-166 base pairs in S. cerevisiae[28][44]. This spacing activity is achieved through the protein's ability to slide nucleosomes and position them at defined intervals, distinct from the stochastic nucleosome deposition that occurs in the absence of active remodeling[25]. The SANT and SLIDE DNA-binding domains appear particularly important for this spacing function, as their removal substantially impairs the ability of CHD1 to generate ordered arrays, though complete loss of these domains does not entirely eliminate spacing activity[8][44].
Surprisingly, CHD1 generates constant nucleosome spacing even when the nucleosome density is experimentally reduced through removal of histone octamers, countering the long-held assumption that nucleosome spacing is simply determined by the proportion of histones relative to DNA[28]. This observation suggests that CHD1 maintains nucleosome spacing through an active mechanism involving protein-protein interactions between nucleosomes or indirect interactions mediated through the remodeler itself[28]. The mechanism may involve a "protein ruler" function where specific domain contacts with multiple nucleosomes simultaneously establish the spacing interval, though the precise molecular details remain to be fully elucidated. In genomic contexts, CHD1 maintains nucleosomal arrays with shorter spacing (approximately 159 base pairs in yeast) compared to the longer spacing established by ISW1 remodelers (approximately 175 base pairs), and these two remodeling enzymes compete on most genes to establish the final spacing observed in wild-type cells[25][60].
Role in Transcription Elongation and Chromatin Dynamics
Interactions with Elongation Factors and RNA Polymerase II Complex
CHD1 functions as an integral component of the transcription elongation machinery in S. cerevisiae through its direct physical and genetic interactions with multiple RNA polymerase II-associated factors[2][26][33][43]. The initial identification of CHD1 in two-hybrid screens with Rtf1, a component of the Paf1 complex (Paf1C) that travels with RNA polymerase II and regulates transcription elongation, established a functional connection between chromatin remodeling and transcription elongation[2][26]. Subsequently, co-immunoprecipitation studies confirmed that CHD1 interacts with critical elongation factors including the Spt4-Spt5 heterodimer (known as DSIF in mammals) and the Spt16-Pob3 complex (known as FACT in mammals), both of which function to facilitate RNA polymerase II passage through nucleosomal barriers[2][26][33].
Genetic analyses provide complementary evidence for CHD1's role in transcription elongation, as deletion of CHD1 suppresses cold-sensitive mutations in the elongation factor Spt5, and this suppression is shared with mutations affecting the Paf1 complex and RNA polymerase II itself[2][26]. Furthermore, a direct interaction has been identified between an N-terminal region of Rtf1 and the CHCT domain (a conserved domain within CHD1 about which little is known functionally) as being both necessary and sufficient for physical association, with mutations disrupting this interaction resulting in accumulation of CHD1 at the 5' ends of CHD1-occupied genes and increased cryptic transcription[13]. The conservation of this Rtf1-CHD1 interaction mechanism in mammalian systems, where homologous regions within mouse RTF1 interact with the CHCT domains of both mouse CHD1 and CHD2, suggests an evolutionarily conserved mechanism for coupling CHD family remodelers to transcription elongation complexes[13].
Nucleosome Repositioning During Transcription
Genome-wide chromatin immunoprecipitation studies demonstrate that CHD1 specifically localizes to the coding regions of actively transcribed genes, where it associates with the nucleosomes that must be traversed by RNA polymerase II during transcription elongation[2][26][43]. Importantly, CHD1 occupancy on gene bodies is partially dependent on Rtf1, the transcription elongation factor, suggesting that the elongation machinery actively recruits or retains CHD1 at transcribed chromatin regions[2][26]. The positioning of CHD1 on transcribed regions appears distinct from its role at promoters and transcription start sites, indicating that CHD1 carries out specialized functions during the elongation phase rather than transcription initiation.
A key function of CHD1 during transcription elongation involves re-establishing proper nucleosome positioning in the wake of RNA polymerase II passage[13][37]. As RNA polymerase II transcribes through chromatin, the transcription process inherently disrupts nucleosomal structure, causing nucleosomes to be pushed forward along the DNA template and potentially leading to displacement or disassembly of histone octamers if not actively managed[13][37]. CHD1 works coordinately with the FACT histone chaperone complex to resolve nucleosomal disruptions caused by transcription, with CHD1 promoting the translocation of FACT molecules from one nucleosome to the next along actively transcribed genes, enabling processive maintenance of chromatin integrity during transcription elongation[37][40]. The ATPase activity of CHD1 is critical for determining FACT distribution along genes, as catalytically inactive mutants of CHD1 phenocopy the effect of CHD1 deletion on FACT localization[37].
Effects on Histone Dynamics and Modification Patterns
Genome-wide analyses of histone turnover reveal that CHD1 exerts significant influence over the dynamics of histone H3 replacement, with the most prominent effects occurring at the 3' ends of genes rather than at promoters[51]. The effects of CHD1 on histone H3 turnover show gene-length dependence, with Chd1's influence on both 5' end and 3' end H3 turnover being greatest at unusually long genes and highly transcribed genes[51]. Mechanistically, CHD1 appears to stabilize nucleosomes that have been perturbed by transcription, preventing excessive turnover while simultaneously permitting the controlled replacement of histone variants and modified histones necessary for proper gene expression[51]. The influence of CHD1 on histone dynamics extends to changes in histone modification patterns genome-wide, with H3K36me3 in particular shifting its distribution pattern in concert with altered H3 replacement patterns in CHD1-deficient cells[51].
Nucleosome Organization and Chromatin Architecture
Suppression of Cryptic Transcription Through Nucleosome Positioning
One of the most striking consequences of CHD1 deletion is a dramatic increase in aberrant or cryptic transcription emanating from internal promoter sequences within coding regions[5][27]. This cryptic transcription arises not from reduced nucleosome occupancy genome-wide, as nucleosome abundance remains relatively constant in CHD1-deficient cells, but rather from loss of regular nucleosome positioning and phasing[5][27]. Specifically, nucleosome mapping studies in both S. cerevisiae and S. pombe reveal that CHD1 (or its fission yeast orthologues Hrp1 and Hrp3) is responsible for generating and maintaining transcription start site-aligned nucleosomal arrays throughout the genome with characteristic well-positioned nucleosomes immediately flanking nucleosome-free regions at promoters[27][44].
In CHD1-deficient cells, the loss of regularly positioned nucleosomes in gene coding regions presumably creates transiently accessible regions of chromatin that expose cryptic transcription initiation sites normally occluded by properly positioned nucleosomes[27]. Supporting this model, the addition of other mutations that affect nucleosome occupancy or histone modification patterns generates cryptic transcription through distinct mechanisms: deletion of the FACT complex subunit Pob3 reduces nucleosome occupancy without affecting nucleosome positioning, while deletion of the histone deacetylase Alp13 does not substantially alter nucleosome positioning or occupancy but increases histone acetylation that may recruit other remodeling factors[27]. These observations indicate that cryptic transcription can result from multiple distinct perturbations to chromatin structure, but when caused specifically by CHD1 deletion, it reflects defects in nucleosome positioning rather than occupancy.
Nucleosome-Free Regions and TSS Organization
The nucleosome-free region (NFR) typically observed at promoters, flanked by well-positioned +1 and -1 nucleosomes, requires coordinated action of multiple nucleosome remodeling enzymes in addition to sequence features[44][54]. In S. pombe, the two CHD-family remodelers Hrp1 and Hrp3 are individually responsible for generating nucleosome spacing in in vitro assays and are essential for maintaining genic nucleosomal arrays in vivo, yet neither single deletion causes complete loss of NFR formation at promoters[44]. This suggests that while CHD family remodelers are important for maintaining nucleosome arrays within genes, other mechanisms (potentially involving sequence-determined nucleosome positioning and other remodeling factors) contribute to NFR generation and maintenance at promoters. However, the combined deletion of both hrp1 and hrp3 severely compromises nucleosome positioning throughout the genome, demonstrating the essential collective role of CHD-family remodelers in establishing nucleosomal organization[44].
The positioning of nucleosomes relative to transcription start sites appears to influence transcriptional regulation independently of promoter accessibility, as experimental manipulation of the inter-nucleosome distance at specific heterochromatic loci reveals an oscillatory relationship between inter-nucleosome-free region distance and silencing efficiency[30]. This observation indicates that nucleosome positioning per se, beyond simple presence or absence of nucleosomes, contributes to the establishment and maintenance of repressive chromatin states, and proper nucleosome positioning may be necessary for robust gene silencing[30].
Interactions with Other Chromatin Remodeling Complexes
Redundancy and Competition with ISW1 and ISW2
CHD1 functions within a larger network of ATP-dependent chromatin remodeling activities that frequently exhibit functional redundancy yet maintain specialized roles in specific genomic contexts[5][25][27][45][60]. While individual deletion of CHD1 causes only subtle transcriptional changes and weak phenotypes, combined deletion of CHD1 with genes encoding other nucleosome spacing remodelers produces synthetic growth defects and severe defects in nucleosome positioning[5][14][27][45]. Specifically, yeast strains bearing combined deletions of chd1, isw1, and isw2 exhibit temperature sensitivity for growth and widespread disruption of nucleosome positioning, demonstrating substantial functional overlap among these spacing remodelers[5][14].
Genome-wide nucleosome sequencing studies reveal that CHD1 and ISW1 actively compete to set nucleosome spacing on most genes, with CHD1 establishing shorter spacing (approximately 159 base pairs) and ISW1 establishing longer spacing (approximately 175 base pairs)[25][60]. On heavily transcribed genes, nucleosome spacing is often extreme (either very short or very long), and these genes are specifically depleted of linker histone H1, which typically binds to longer spacing arrays to promote higher-order chromatin folding[25][60]. The hypothesis emerges that CHD1-specified short spacing evicts H1 and causes chromatin unfolding suitable for active transcription, whereas ISW1-specified longer spacing allows H1 binding and promotes chromatin condensation appropriate for more regulated transcription or inactive chromatin[25][60].
The competition between CHD1 and ISW1 appears dynamic across the genome, with the two remodelers potentially operating sequentially or concurrently on individual genes[25][60]. This dynamic competition provides a mechanism for fine-tuning nucleosome spacing in response to transcriptional activity and developmental cues. ISW2, another spacing remodeler, plays a more limited role restricted primarily to transcriptionally inactive genes, where it generates the longest spacing observed at any class of genes, suggesting a specialized function in establishing chromatin structure at silenced loci[60].
Cooperativity with SWI/SNF Complex
Genetic interactions between CHD1 and components of the SWI/SNF chromatin remodeling complex, which catalyzes nucleosome disruption rather than spacing, reveal additional layers of functional cooperation and potential antagonism[14][45][48]. Synthetic-lethal screening identified that combined mutations in CHD1 and SWI/SNF genes result in inviability, and similar temperature-sensitive synthetic growth defects arise from combined deletion of chd1 with mutations in the ISWI genes ISW1 and ISW2[14][45][48]. These genetic interactions strongly suggest that CHD1 and SWI/SNF maintain partially redundant functions in cell growth and nuclear organization, consistent with their distinct catalytic activities (nucleosome spacing versus disruption) operating toward the common goal of maintaining functional chromatin structure.
Importantly, the relationship between CHD1 and remodeling complexes is not simply one of redundancy but involves complex coordination of activities in specific biological contexts. For instance, CHD1 and FACT histone chaperone have been reported to act in opposition in regulating DNA replication, as chd1 and set2 mutations can suppress mutations in FACT components that cause sensitivity to hydroxyurea and other replication stresses[38]. This opposition appears to involve Chd1 negatively regulating DNA replication in a manner that differs from FACT's function in promoting replication fork progression[38].
Functions Beyond Transcription Elongation
DNA Repair and Homologous Recombination
Beyond its well-established roles in transcription and nucleosome organization, CHD1 participates in homologous recombination-based DNA double-strand break (DSB) repair through mechanisms that involve chromatin remodeling and chromatin opening at damage sites[19][22]. Both yeast and human CHD1 are recruited to chromatin adjacent to DSBs, where they promote the opening of chromatin and facilitate the recruitment of DNA repair factors[9][19][22]. Specifically, CHD1 promotes DSB repair through its effects on DNA end resection, the process by which the 5'-terminated DNA strands surrounding a DSB are degraded to generate single-stranded DNA overhangs necessary for homologous recombination[19][22].
In S. cerevisiae, CHD1 is required for efficient recruitment of the MRX complex (Mre11-Rad50-Xrs2) and the nuclease Exo1 to DSB ends, and deletion or catalytic inactivation of CHD1 reduces histone occupancy near DSB ends and substantially impairs DSB repair by homologous recombination[19]. The requirement for CHD1 ATPase activity demonstrates that the chromatin remodeling function, rather than structural roles of the protein, drives its contribution to DNA repair[19]. In human cells, targeted disruption of CHD1 leads to defects in early double-strand break signaling and homologous recombination, resulting in hypersensitivity to ionizing radiation as well as PARP and PTEN inhibition[22]. Furthermore, human CHD1 contains a unique N-terminal inhibitory region that represses the DNA-binding, ATPase, and chromatin assembly activities of the protein, and CHD1 lacking this N-terminal region is more active in rescuing defects in γH2AX formation and CtIP recruitment in CHD1-knockout cells[22].
The role of CHD1 in DNA repair appears conserved among the CHD protein family, as other family members including CHD2, CHD3, CHD4, and CHD7 have also been implicated in the cellular response to DNA damage, though with specialized roles in either homologous recombination or non-homologous end joining pathways[19]. Mutations in human CHD7 associated with CHARGE syndrome occur specifically in the SLIDE domain, the DNA-binding domain shared between Chd1 and ISWI remodelers[6], suggesting that defects in chromatin remodeling activity contribute to the disease phenotype.
Heterochromatin Assembly and Silencing
Chromodomain-containing proteins in general play important roles in heterochromatin assembly and transcriptional silencing, exemplified by the well-characterized role of the heterochromatin protein HP1 and the fission yeast protein Chp1[3][9]. While the yeast CHD1 chromodomains do not bind methylated histone H3K4 as occurs in mammalian CHD1 and other chromodomain proteins, CHD1 nonetheless participates in proper chromatin organization within heterochromatic domains and at telomeres[9]. The maintenance of nucleosome positioning in heterochromatic regions contributes to the stability of silenced chromatin states, as demonstrated by experiments altering the inter-nucleosome distance at heterochromatic loci, which causes corresponding oscillations in silencing efficiency[30].
Genomic Localization and Target Gene Selection
Association with Actively Transcribed Chromatin
Genome-wide chromatin immunoprecipitation followed by microarray analysis has established that yeast CHD1 specifically localizes to the coding regions of actively transcribed genes, with enrichment particularly pronounced within the transcribed regions rather than at promoters or transcription termination sites[2][26][43]. This localization pattern is consistent with CHD1's primary role in maintaining nucleosome positioning during transcription elongation rather than in transcription initiation. The association of CHD1 with transcribed chromatin appears partially dependent on Rtf1 and other components of the transcription elongation machinery, suggesting that active transcription and associated recruitment by elongation factors drive CHD1 targeting to specific genomic loci[2][26].
In contrast to human CHD1, which preferentially recognizes and binds histone H3K4me3 through its chromodomains as a mechanism for recruitment to active gene promoters, yeast Chd1 chromodomains do not directly recognize methylated histone H3K4[9][12][31]. Instead, yeast CHD1 appears to be recruited to genes through its interaction with the Paf1 complex and other elongation factors that physically associate with RNA polymerase II during transcription[2][26][43]. This divergence between yeast and mammalian CHD1 mechanisms for genomic targeting suggests that while the catalytic and regulatory logic of CHD family proteins has been conserved, the specific mechanisms for recruitment to genomic targets have been subject to evolutionary modification.
Biochemical Substrate Specificity
While CHD1 does not display obvious sequence-specific DNA-binding preferences, as its DNA-binding domain engages extranucleosomal DNA in a sequence-nonspecific manner, the enzyme does exhibit specificity for certain nucleosome substrate features[56]. CHD1 preferentially binds to and slides nucleosomes that possess longer extranucleosomal DNA (linker DNA), with this binding preference biasing the remodeler to shift histone octamers toward longer linker DNA, promoting nucleosome centering on short DNA fragments[56]. This preference for nucleosomes with longer linker DNA is shared with the related ISWI spacing remodelers, suggesting a conserved mechanism for directing nucleosome positioning that optimizes for nucleosome centering on small DNA molecules and regular spacing on genomic templates.
The nucleosome substrate specificity of CHD1 extends to histone modifications, as studies comparing nucleosome binding and remodeling activity on substrates with or without specific histone modifications reveal that H3K36 methylation, particularly the di- and tri-methylated forms that accumulate in actively transcribed gene bodies, influences CHD1 recruitment and activity[5][25][51]. Furthermore, the histone H4 acetylation state influences CHD1 activity at specific genomic loci, suggesting that both histone tail modifications and nucleosome density contribute to determining where and when CHD1 acts within the genome[5].
Protein Regulation and Structural Plasticity
Autoinhibitory Domains and Activation Mechanisms
The discovery that truncated forms of CHD1 lacking the first 57-88 amino acid residues or the entire N-terminal 133 amino acids show hyperactivity compared to full-length CHD1 indicates that N-terminal regions normally restrain the enzymatic activity of the protein through autoinhibitory mechanisms[24][55]. These hyperactive deletions also show reduced association between the ATPase motor and DNA-binding domain, suggesting that a more flexible linkage between these functional modules facilitates productive engagement with nucleosomes, while certain N-terminal residues maintain an autoinhibited state through interactions that maintain rigid positioning of the DNA-binding domain[24][55]. The fact that both enhanced activity mutants specifically reduce association between the ATPase and DNA-binding domain suggests that a less constrained positioning allows these domains to find optimal orientations for productive catalysis[24][55].
In human CHD1, an autoinhibitory N-terminal region distinct from the serine-rich region represses DNA-binding, ATP hydrolysis, and chromatin assembly and remodeling activities in vitro, though this region appears largely absent or divergent in yeast and Drosophila CHD1 proteins[22]. The repressive region of human CHD1 at the N-terminus is not conserved between human CHD1 and the closely related human CHD2, suggesting that the two closely related family members have evolved differential regulatory mechanisms for controlling their activity[22]. The identification of such autoinhibitory mechanisms suggests that CHD family remodelers may be subject to positive and negative regulation in response to cellular signals or post-translational modifications, though specific activators and inhibitors of CHD1 have not yet been definitively identified.
Nucleosome Substrate-Dependent Regulation
The regulation of CHD1 activity is intimately connected to the structural properties of its nucleosome substrates, as the linker DNA length available for interaction with the DNA-binding domain directly influences the rate of nucleosome movement[34][59]. The molecular connectivity between the positioning of the DNA-binding domain via the chromodomains to the ATPase catalytic pocket implies that linker DNA length directly regulates CHD1 activity, with longer linker DNA enabling more efficient closure of the ATPase motor domains and nucleotide binding[34][59]. This substrate-dependent regulation provides an elegant mechanism by which the spacing of nucleosomes genome-wide, which exhibits an average linker length of approximately 19 base pairs in S. cerevisiae, directly determines CHD1 catalytic efficiency and thus influence which genes CHD1 acts upon[34][59].
Summary of Molecular Functions and Mechanisms
The comprehensive picture of CHD1 function that emerges from structural, biochemical, and genomic studies reveals a highly specialized ATP-dependent chromatin remodeling enzyme whose primary catalytic function involves nucleosome sliding and spacing[1][2][5][7][8]. CHD1 executes this remodeling through a sophisticated molecular mechanism involving ATP-dependent ratcheting motion of ATPase lobes that translocates nucleosomal DNA toward the histone octamer dyad, coupled with substrate DNA-binding domains that engage extranucleosomal DNA to direct sliding toward longer linker DNA and promote nucleosome centering[7][34][42][56]. The enzyme operates predominantly as a monomeric protein and functions as a nucleosome-spacing enzyme rather than a nucleosome disruption or assembly factor, generating and maintaining regularly spaced nucleosomal arrays with constant spacing independent of nucleosome density[28][44].
Within the cell, CHD1 localizes to nuclei where it associates with transcribed chromatin through recruitment by transcription elongation factors including Rtf1 of the Paf1 complex and works coordinately with FACT histone chaperone to maintain nucleosome positioning and chromatin integrity as RNA polymerase II traverses the transcribed regions of genes[2][26][37][40]. Beyond transcription, CHD1 participates in homologous recombination-based DSB repair by promoting chromatin opening and the recruitment of nucleotide resection machinery[19][22]. The evolutionary conservation of CHD1 from yeast to humans, combined with the specialization of nine distinct CHD family members in mammals, underscores the fundamental importance of this remodeling activity for proper nuclear organization and gene regulation across eukaryotic organisms.
Conclusion
CHD1 stands as a paradigmatic example of an ATP-dependent chromatin remodeling factor whose sophisticated molecular architecture and dynamic regulatory mechanisms enable precise control of nucleosome positioning and spacing throughout the yeast genome. The monomeric architecture of yeast CHD1, contrasting with the multi-subunit complexes formed by other remodeling enzymes, provides a simplified yet comprehensive model system for understanding chromatin remodeling principles that apply broadly to all SNF2-family chromatin remodelers. The cryo-electron microscopy structures determined between 2017 and 2025 have provided unprecedented molecular detail into how CHD1 engages nucleosomes through the coordinated positioning of its functional domains to achieve DNA unwrapping and translocation, establishing mechanistic principles likely applicable to the broader family of related enzymes.
The functional diversity of CHD1 extends far beyond simple nucleosome repositioning to encompass roles in transcription elongation, homologous recombination DNA repair, heterochromatin maintenance, and epigenetic regulation through its effects on histone dynamics and modification patterns[1][9][13][19][22][51]. The remarkable evolutionary conservation of CHD protein domains and mechanisms across eukaryotic phyla, coupled with the specialization of mammalian CHD family members into distinct subfamilies with overlapping yet specialized functions, reflects the fundamental necessity of precise chromatin regulation for proper nuclear organization and gene expression in complex organisms. Future investigations into post-translational modification of CHD1, additional protein-protein interactions, and mechanisms regulating the autoinhibitory domains will likely reveal additional layers of regulation controlling when and where this crucial chromatin remodeling enzyme acts within the nuclear environment. The clinical significance of CHD1 mutations in human disease, particularly in prostate cancer development where CHD1 is mutated or deleted in approximately 10% of cases, underscores the importance of understanding this protein's cellular functions and the consequences of its dysregulation in pathological contexts.
Citations
- https://www.yeastgenome.org/locus/S000000966
- https://pmc.ncbi.nlm.nih.gov/articles/PMC154471/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC522800/
- https://www.ncbi.nlm.nih.gov/gene/856911
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3560298/
- https://pdbj.org/mine/summary/2XB0
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5697743/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3243530/
- https://en.wikipedia.org/wiki/CHD1
- https://elifesciences.org/articles/35720
- https://www.uniprot.org/uniprotkb/P32657/entry
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1421377/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC12390759/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1899158/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6118821/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4162295/
- https://royalsocietypublishing.org/rstb/article/372/1731/20160284/23281/The-tale-of-a-tail-histone-H4-acetylation-and-the
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8462745/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC230505/
- https://academic.oup.com/nar/article/46/8/3891/4909984
- https://elifesciences.org/articles/22510
- https://www.science.org/doi/10.1126/sciadv.adw7108
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3492713/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4387221/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2099615/
- https://www.pnas.org/doi/10.1073/pnas.2004111117
- https://elifesciences.org/articles/35720.pdf
- https://pubmed.ncbi.nlm.nih.gov/33174727/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9149603/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2248355/
- https://www.tandfonline.com/doi/full/10.1080/21541264.2022.2069995
- https://genesdev.cshlp.org/content/12/18/2956
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4230344/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3512388/
- https://febs.onlinelibrary.wiley.com/doi/10.1002/2211-5463.13226
- https://elifesciences.org/articles/01270
- https://pmc.ncbi.nlm.nih.gov/articles/PMC384354/
- https://www.rcsb.org/structure/2dy7
- https://journals.plos.org/plosgenetics/article?id=10.1371%2Fjournal.pgen.1002811
- https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2014.00223/full
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3232923/
- https://academic.oup.com/nar/article/38/5/1441/3112508
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4889916/