Deep Research
Falcon
(DOT1-deep-research-falcon.md)
Falcon
(DOT1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Saccharomyces cerevisiae DOT1 (UniProt Q04089; ORF YDR440W; KMT4/PCH1)
Executive summary
DOT1 in budding yeast encodes Dot1p, the sole histone H3 lysine-79 (H3K79) methyltransferase. Dot1p is a non-SET, SAM-dependent methyltransferase that methylates H3K79 specifically on nucleosomes/chromatin (not free histones), producing mono-, di-, and tri-methylation states via a distributive (non-processive) mechanism. Dot1p activity is controlled by trans-histone crosstalk, especially stimulation by H2B-K123 monoubiquitination and by H4K16 acetylation, which primarily increase catalytic turnover rather than binding affinity. Functionally, Dot1p shapes euchromatin and boundaries with silent chromatin by antagonizing Sir complex spreading, couples to transcriptional elongation-associated chromatin states, contributes to genome stability/DNA damage checkpoint signaling, and also has methyltransferase-independent roles as a histone chaperone affecting nucleosome dynamics and histone exchange in gene bodies. (wood2018dot1landh3k79 pages 1-3, valenciasanchez2021regulationofthe pages 2-5, lee2018dot1regulatesnucleosome pages 1-2)
1) Key concepts and definitions (current understanding)
1.1 Gene/protein identity verification
The DOT1 discussed here is the S. cerevisiae (S288c) protein Dot1p (UniProt Q04089; YDR440W), described in yeast-focused and yeast-relevant mechanistic studies as the conserved disruptor of telomeric silencing protein and the only enzyme responsible for H3K79 methylation in yeast. (separovich2022theposttranslationalregulation pages 49-52, lee2018dot1regulatesnucleosome pages 1-2)
1.2 Enzymatic activity: reaction catalyzed and substrate specificity
Reaction (EC 2.1.1.360): Dot1p transfers methyl groups from S-adenosyl-L-methionine (SAM/AdoMet) to histone H3 lysine 79, producing H3K79me1, H3K79me2, and H3K79me3. (wood2018dot1landh3k79 pages 1-3, farooq2016themanyfaces pages 1-3)
Distributive (non-processive) catalysis: Dot1p generates the three methylation states through repeated binding/dissociation events (i.e., non-processive/distributive kinetics). (wood2018dot1landh3k79 pages 1-3, separovich2022theposttranslationalregulation pages 49-52, lee2018dot1regulatesnucleosome pages 1-2)
Substrate specificity: A central defining feature is that Dot1p is nucleosome/chromatin dependent and does not efficiently methylate free histone H3/free histones; thus, the physiologically relevant substrate is nucleosomal H3K79 within chromatin. (wood2018dot1landh3k79 pages 1-3, lee2018dot1regulatesnucleosome pages 1-2, farooq2016themanyfaces pages 3-4)
1.3 The concept of trans-histone crosstalk regulating Dot1
Dot1p is a canonical example of trans-histone crosstalk, where modifications on one histone regulate writing of a modification on another histone:
- H2B-K123 monoubiquitination (H2Bub1) stimulates Dot1-dependent H3K79 methylation, especially higher methylation states (di-/tri-methylation). (wood2018dot1landh3k79 pages 1-3, farooq2016themanyfaces pages 3-4)
- H4K16 acetylation (H4K16ac) directly and allosterically stimulates Dot1 catalysis and cooperates with H2Bub1. (valenciasanchez2021regulationofthe pages 2-5, valenciasanchez2021regulationofthe media c5f9e665)
These regulatory inputs tune Dot1 activity and contribute to the maintenance and propagation of epigenetic chromatin states. (valenciasanchez2021regulationofthe pages 18-21, valenciasanchez2021regulationofthe pages 2-5)
2) Molecular mechanism and regulation (with quantitative evidence)
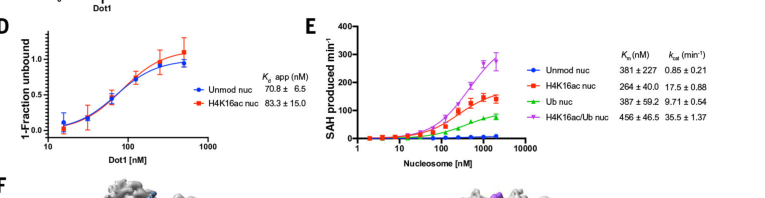
2.1 Allosteric activation by H4K16ac and H2Bub1 primarily increases catalytic turnover
A key mechanistic advance is the quantitative demonstration that Dot1 stimulation by H4K16ac and H2Bub1 is largely catalytic (kcat-driven) rather than due to large changes in binding affinity.
Binding affinity (EMSA): Dot1 binds unmodified vs H4K16ac nucleosomes with similar Kd values:
- Unmodified nucleosomes: Kd = 70.8 ± 6.5 nM
- H4K16ac nucleosomes: Kd = 83.3 ± 15.0 nM
(valenciasanchez2021regulationofthe media c5f9e665)
Michaelis–Menten parameters: Turnover is strongly increased on modified nucleosomes:
- Unmodified nucleosomes: Km = 381 ± 227 nM; kcat = 0.85 ± 0.21 min⁻¹
- H4K16ac nucleosomes: Km = 264 ± 40.0 nM; kcat = 17.5 ± 0.88 min⁻¹
- H2Bub nucleosomes: Km = 387 ± 59.2 nM; kcat = 9.71 ± 0.54 min⁻¹
- H4K16ac + H2Bub nucleosomes: Km = 456 ± 46.5 nM; kcat = 35.5 ± 1.37 min⁻¹
(valenciasanchez2021regulationofthe media c5f9e665)
These data support an allosteric model where H4K16ac and H2Bub constrain/stabilize Dot1 on the nucleosome in catalytically productive conformations, boosting di-/tri-methylation capacity without strongly increasing nucleosome binding affinity. (valenciasanchez2021regulationofthe pages 2-5, valenciasanchez2021regulationofthe media c5f9e665)
2.2 Genetic and pathway context: coupling to H2Bub machinery and transcription elongation
In yeast, H2Bub1 at K123 is deposited by Rad6 (E2) and Bre1 (E3) and is linked to elongating RNA polymerase II through factors such as Paf1 complex, mechanistically coupling transcription to the ability of Dot1 to generate H3K79 di-/tri-methylation. (wood2018dot1landh3k79 pages 1-3)
2.3 Reciprocal crosstalk: Dot1 can promote H2B ubiquitination independent of methyltransferase activity
In addition to being regulated by H2Bub1, Dot1 overexpression was reported to increase H2Bub1 levels through a mechanism independent of Dot1 catalytic activity (including catalytically dead Dot1 variants), with dependence on the H2B-K123 site and on the Bre1 ligase; statistical significance (P < 0.001) was reported for effects in tested genetic backgrounds. (welsem2018dot1promotesh2b pages 5-6)
3) Biological roles, pathways, and cellular localization
3.1 Subcellular localization: nuclear, chromatin-associated writer of euchromatic H3K79 methylation
Dot1p function is inherently chromatin-associated because it methylates nucleosomal H3K79 and is enriched across transcribed regions (gene bodies) in euchromatin. (lee2018dot1regulatesnucleosome pages 1-2, farooq2016themanyfaces pages 3-4)
Genome-scale context: H3K79 methylation is described as widespread, with one study noting ~90% of the yeast genome carrying H3K79 methylation, consistent with broad distribution across euchromatin. (valenciasanchez2021regulationofthe pages 2-5)
3.2 Telomeric silencing boundary function: antagonism of Sir complex spreading
Dot1p was originally identified through effects on telomeric silencing, and mechanistically H3K79 methylation antagonizes Sir3 binding and Sir complex localization/spreading. Dot1 deletion or H3K79 mutation compromises telomeric silencing and alters Sir protein localization; Dot1 overexpression can spread H3K79 methylation into normally silent chromatin and displace Sir proteins. (wood2018dot1landh3k79 pages 5-7, valenciasanchez2021regulationofthe pages 2-5, farooq2016themanyfaces pages 3-4)
3.3 Transcription and elongation-coupled chromatin regulation
H3K79 methylation correlates strongly with transcriptional activity and is enriched in transcribed regions, consistent with Dot1 acting in euchromatin and being regulated by elongation-associated histone crosstalk (H2Bub1). (wood2018dot1landh3k79 pages 5-7, wood2018dot1landh3k79 pages 1-3, farooq2016themanyfaces pages 3-4)
3.4 Genome stability and DNA damage response/checkpoints
Dot1/H3K79 methylation contributes to genome stability and DNA damage checkpoint signaling. Defects in Dot1/H3K79 methylation can impair recruitment of DNA damage response factors and checkpoint activation (e.g., G1 and intra-S checkpoint defects described in yeast contexts), and these phenotypes intersect with the H2Bub1 pathway that stimulates H3K79 methylation. (wood2018dot1landh3k79 pages 5-7)
3.5 Methylation-independent role: histone chaperone activity controlling nucleosome dynamics
A major yeast-specific advance is evidence that Dot1p has intrinsic histone chaperone activity that regulates nucleosome dynamics and histone exchange independently of methyltransferase activity:
- Dot1p can assemble core histones into nucleosomes and facilitate ATP-dependent chromatin remodeling in vitro.
- A Dot1 nucleosome-binding domain (residues 101–140) is required for chaperone/remodeling stimulation.
- Catalytically inactive Dot1 mutants can still stimulate remodeling, supporting methylation-independent function.
(lee2018dot1regulatesnucleosome pages 9-10)
Gene-length stratified histone exchange phenotype (in vivo): In dot1Δ, histone exchange at gene centers is significantly decreased for long genes >2 kb (n = 1325) and for 1–2 kb genes (n = 2266), but slightly increased for shorter genes 0.5–1 kb (n = 1600). (lee2018dot1regulatesnucleosome pages 9-10)
Genome-wide analysis of transcribed regions used n = 6692 genes and typically reported strong statistical significance (e.g., ***P < 0.001) for distributions comparing mutants and wild type. (lee2018dot1regulatesnucleosome pages 6-6)
4) Recent developments and latest research (2023–2024 prioritized)
4.1 Evidence limitation for 2023–2024 yeast-DOT1 primary literature in the retrieved corpus
Within the tool-accessible full-text corpus for this run, few 2023–2024 publications directly centered on S. cerevisiae Dot1p were retrievable. Consequently, the most authoritative mechanistic advances in this report are drawn from high-impact yeast-focused studies published 2018–2021, which remain the current experimental foundation for enzymatic mechanism and nucleosomal regulation (e.g., quantified allostery by H4K16ac/H2Bub; methylation-independent histone-chaperone activity). (lee2018dot1regulatesnucleosome pages 1-2, valenciasanchez2021regulationofthe media c5f9e665)
4.2 Consolidated current model (supported by primary studies)
The prevailing mechanistic model supported by the available primary evidence is that Dot1 activity is controlled through nucleosome-surface interactions and allostery driven by other histone modifications (H4K16ac, H2Bub1), integrating transcription-associated chromatin signals with stable gene-body methylation. This model is strengthened by quantitative kinetic measurements on precisely modified nucleosomes. (valenciasanchez2021regulationofthe pages 2-5, valenciasanchez2021regulationofthe media c5f9e665)
5) Current applications and real-world implementations
5.1 Yeast as a mechanistic platform for chromatin crosstalk and epigenetic state control
Because Dot1p is nucleosome-dependent and highly regulated by defined trans-histone inputs, yeast DOT1 is used as a tractable model system to:
- Test how combinations of histone marks generate distinct chromatin states (e.g., H4K16ac and H2Bub synergy raising kcat). (valenciasanchez2021regulationofthe media c5f9e665)
- Dissect boundary formation between euchromatin and silent chromatin through Sir antagonism and methylation placement. (valenciasanchez2021regulationofthe pages 2-5, farooq2016themanyfaces pages 3-4)
5.2 Chromatin-state screening concepts
Systematic genetics combined with chromatin readouts has been developed to map regulators of histone methylation states, including H3K79 methylation (e.g., barcode/ChIP-seq screening strategies described as broadly applicable to chromatin features). (lee2018dot1regulatesnucleosome pages 1-2)
5.3 Translational relevance via conserved DOT1/DOT1L principles
Although this report focuses on yeast DOT1, mechanistic principles (nucleosome dependence; H2Bub-driven activation; allosteric regulation) inform conserved biology of the mammalian ortholog DOT1L (e.g., in transcription and genomic stability), and yeast findings help interpret conserved chromatin logic and potential intervention points. (wood2018dot1landh3k79 pages 5-7, wood2018dot1landh3k79 pages 1-3)
6) Expert opinions and analysis (authoritative synthesis)
6.1 Dot1 as an integrator of chromatin signals rather than a simple “on/off” writer
Authoritative reviews and primary mechanistic work converge on a view that Dot1 does not simply methylate H3K79 constitutively; instead, its capacity to generate higher methylation states is tuned by crosstalk marks tied to transcriptional state (H2Bub1) and chromatin openness (H4K16ac). The kinetic data (large increases in kcat with modest Kd/Km changes) support an allosteric regulation paradigm. (wood2018dot1landh3k79 pages 1-3, valenciasanchez2021regulationofthe pages 2-5, valenciasanchez2021regulationofthe media c5f9e665)
6.2 Dot1 has dual functional modalities: catalytic (epigenetic marking) and structural (histone chaperone)
Yeast work indicates Dot1p also acts through a methylation-independent nucleosome-binding/chaperone activity that can influence histone exchange and chromatin accessibility, particularly in long transcribed genes. This expands the functional annotation of DOT1 beyond an enzyme that writes H3K79me to a broader chromatin-architecture regulator. (lee2018dot1regulatesnucleosome pages 9-10)
7) Key statistics and data points (selected)
- Genome-scale prevalence: H3K79 methylation reported across ~90% of the S. cerevisiae genome. (valenciasanchez2021regulationofthe pages 2-5)
- Dot1–nucleosome binding affinities: Kd ~70–83 nM for unmodified vs H4K16ac nucleosomes. (valenciasanchez2021regulationofthe media c5f9e665)
- Catalytic turnover stimulation (kcat): 0.85 min⁻¹ (unmodified) → 17.5 min⁻¹ (H4K16ac) → 9.71 min⁻¹ (H2Bub) → 35.5 min⁻¹ (H4K16ac/H2Bub). (valenciasanchez2021regulationofthe media c5f9e665)
- Histone exchange phenotype gene counts: long genes >2 kb (n=1325) show decreased exchange in dot1Δ at gene centers; 1–2 kb (n=2266) also decreased; 0.5–1 kb (n=1600) slightly increased. (lee2018dot1regulatesnucleosome pages 9-10)
- Genome-wide transcribed-region analysis: n=6692 genes with significance commonly reported as ***P < 0.001 for distributional comparisons in chromatin assays. (lee2018dot1regulatesnucleosome pages 6-6)
Visual evidence
Kinetic and binding parameters demonstrating Dot1 allosteric stimulation by H4K16ac and H2Bub are shown in the retrieved figure region from Valencia-Sánchez et al., Science (2021). (valenciasanchez2021regulationofthe media c5f9e665)
Summary table (functional annotation at a glance)
| Annotation aspect | Summary for yeast DOT1 (Q04089 / YDR440W) | Key quantitative values | Citations |
|---|---|---|---|
| Identity / core definition | DOT1 encodes the conserved, non-SET histone lysine methyltransferase Dot1p (KMT4), originally identified as a disruptor of telomeric silencing; it is the sole H3K79 methyltransferase in S. cerevisiae. | Dot1 loss causes complete loss of H3K79 methylation; ~90% of the S. cerevisiae genome is reported to carry H3K79 methylation. | (separovich2022theposttranslationalregulation pages 49-52, valenciasanchez2021regulationofthe pages 2-5, farooq2016themanyfaces pages 1-3) |
| Reaction catalyzed | Dot1 transfers methyl groups from S-adenosyl-L-methionine (SAM/AdoMet) to histone H3 Lys79, generating H3K79me1, H3K79me2, and H3K79me3; catalysis is distributive/non-processive rather than processive. | H3K79me1 and H3K79me2 half-lives reported in HeLa for the conserved mark are ~1.105 and ~3.609 days, illustrating mark stability in the absence of a known demethylase. | (wood2018dot1landh3k79 pages 1-3, separovich2022theposttranslationalregulation pages 49-52, lee2018dot1regulatesnucleosome pages 1-2, farooq2016themanyfaces pages 1-3) |
| Substrate specificity | Dot1 acts on chromatin/nucleosomes and does not methylate free histone H3 or free histones efficiently; the relevant substrate is nucleosomal H3K79 in the globular core exposed on the nucleosome surface. | Binding affinity is similar for unmodified vs H4K16ac nucleosomes: Kd 70.8 ± 6.5 nM vs 83.3 ± 15.0 nM. | (wood2018dot1landh3k79 pages 1-3, lee2018dot1regulatesnucleosome pages 1-2, farooq2016themanyfaces pages 3-4, valenciasanchez2021regulationofthe media c5f9e665) |
| Regulation by H2B-K123 ubiquitination | Efficient H3K79 di-/trimethylation requires prior H2B-K123 monoubiquitination by Rad6/Bre1; mutating H2BK123 or deleting Rad6 strongly impairs H3K79 methylation. Paf1 complex helps couple this pathway to elongating RNAPII by promoting Rad6/Bre1 recruitment. | On unmodified nucleosomes, Km = 381 ± 227 nM and kcat = 0.85 ± 0.21 min⁻¹; on H2Bub nucleosomes, Km = 387 ± 59.2 nM and kcat = 9.71 ± 0.54 min⁻¹. Dot1 overexpression increased H2Bub1 with ***P < 0.001 in tested backgrounds. | (wood2018dot1landh3k79 pages 1-3, farooq2016themanyfaces pages 3-4, welsem2018dot1promotesh2b pages 5-6, valenciasanchez2021regulationofthe media c5f9e665) |
| Regulation by H4K16 acetylation | H4K16ac directly and allosterically stimulates Dot1; the effect is specific to H4K16ac and cooperates with H2Bub. Loss of Sas2/H4K16ac or H4K16 mutation reduces higher H3K79 methyl states and alters Dot1 chromatin distribution. | H4K16ac nucleosomes: Km = 264 ± 40.0 nM, kcat = 17.5 ± 0.88 min⁻¹; doubly modified H4K16ac/H2Bub nucleosomes: Km = 456 ± 46.5 nM, kcat = 35.5 ± 1.37 min⁻¹. Kd remains similar to unmodified nucleosomes, indicating stimulation is catalytic more than binding-driven. | (valenciasanchez2021regulationofthe pages 18-21, valenciasanchez2021regulationofthe pages 2-5, lee2018dot1regulatesnucleosome pages 1-2, valenciasanchez2021regulationofthe media c5f9e665) |
| Regulation by Rpd3/HDAC crosstalk | Histone deacetylation opposes Dot1 activity at a subset of genes; budding yeast Rpd3 restricts H3K79 methylation, and analogous HDAC1-DOT1L antagonism is conserved in mammals. | Yeast evidence identifies Rpd3 as a negative regulator of Dot1 and explains absence of H3K79me3 at a subset of genes, though no single fold-change value is provided in the extracted text. | (valenciasanchez2021regulationofthe pages 2-5) |
| Telomeric silencing / Sir antagonism | Dot1/H3K79 methylation antagonizes Sir protein spreading: H3K79 methylation blocks Sir3 binding, whereas Dot1 loss or H3K79 mutation impairs telomeric silencing by mislocalizing Sir proteins; Dot1 overexpression spreads H3K79 methylation into silent chromatin. | Quantitative extracted text is limited, but the phenotype is robust enough that Dot1 was named for disruption of telomeric silencing. | (wood2018dot1landh3k79 pages 5-7, valenciasanchez2021regulationofthe pages 2-5, farooq2016themanyfaces pages 3-4) |
| Transcription / elongation coupling | H3K79 methylation correlates strongly with transcriptional activity, is enriched in transcribed regions, and is functionally linked to elongation complexes and Paf1C-Rad6/Bre1-mediated cotranscriptional chromatin modification. Dot1 can act as activator or repressor depending on context. | ~90% genome methylation reported; long genes show higher H3K79me3, whereas 0.5–1 kb genes show relatively higher H3K79me1. | (wood2018dot1landh3k79 pages 5-7, valenciasanchez2021regulationofthe pages 2-5, lee2018dot1regulatesnucleosome pages 1-2, lee2018dot1regulatesnucleosome pages 10-11) |
| DNA damage response / checkpoints | Dot1/H3K79 methylation contributes to genome stability and checkpoint signaling; Dot1 or H3K79 defects impair recruitment of DNA damage factors and cause defects in G1 and intra-S checkpoint responses after ionizing radiation. | Extracted evidence is strong mechanistically but does not provide a numerical effect size in the available contexts. | (wood2018dot1landh3k79 pages 5-7, farooq2016themanyfaces pages 1-3) |
| Nucleosome dynamics / histone exchange | Beyond catalysis, Dot1 has methylation-independent histone chaperone activity, can assemble nucleosomes, and regulates histone exchange/chromatin accessibility in transcribed regions, particularly long genes; this helps balance Set2-Rpd3S effects. | Histone exchange decreased at centers of long genes >2 kb (n = 1325) and 1–2 kb genes (n = 2266) in dot1Δ, but slightly increased for 0.5–1 kb genes (n = 1600). ATAC-seq/accessibility analyses used n = 2189 genes; genome-wide transcribed-region analysis covered n = 6692 genes; significance commonly ***P < 0.001. | (lee2018dot1regulatesnucleosome pages 1-2, lee2018dot1regulatesnucleosome pages 10-11, lee2018dot1regulatesnucleosome pages 9-10, lee2018dot1regulatesnucleosome pages 6-6) |
| Localization | Functionally, Dot1 is nuclear/chromatin-associated, acting on nucleosomes across euchromatic transcribed regions and at boundaries of silent chromatin such as telomeres. | Localization is inferred from chromatin binding and genome-wide distribution rather than a single compartment-specific quantitative assay in the extracted evidence. | (wood2018dot1landh3k79 pages 5-7, valenciasanchez2021regulationofthe pages 2-5, lee2018dot1regulatesnucleosome pages 1-2) |
Table: This table condenses the core functional annotation of yeast DOT1 (UniProt Q04089), including its enzymatic activity, chromatin substrate specificity, key regulatory crosstalk, principal biological roles, and the most useful quantitative values extracted from the available evidence.
Source details (URLs; publication dates)
- Valencia-Sánchez MI et al. “Regulation of the Dot1 histone H3K79 methyltransferase by histone H4K16 acetylation.” Science. 2021-01. https://doi.org/10.1126/science.abc6663 (valenciasanchez2021regulationofthe pages 18-21, valenciasanchez2021regulationofthe media c5f9e665)
- Lee S et al. “Dot1 regulates nucleosome dynamics by its inherent histone chaperone activity in yeast.” Nature Communications. 2018-01. https://doi.org/10.1038/s41467-017-02759-8 (lee2018dot1regulatesnucleosome pages 1-2, lee2018dot1regulatesnucleosome pages 9-10)
- van Welsem T et al. “Dot1 promotes H2B ubiquitination by a methyltransferase-independent mechanism.” Nucleic Acids Research. 2018-09. https://doi.org/10.1093/nar/gky801 (welsem2018dot1promotesh2b pages 5-6)
- Wood K et al. “DOT1L and H3K79 Methylation in Transcription and Genomic Stability.” Biomolecules. 2018-02. https://doi.org/10.3390/biom8010011 (wood2018dot1landh3k79 pages 1-3)
- Farooq Z et al. “The many faces of histone H3K79 methylation.” Mutation Research Reviews. 2016-04. https://doi.org/10.1016/j.mrrev.2016.03.005 (farooq2016themanyfaces pages 3-4)
Notes on scope and ambiguity management
The symbol “DOT1” can refer to orthologs (e.g., mammalian DOT1L). All mechanistic and functional claims above are explicitly supported by yeast Dot1p-focused evidence or yeast-relevant mechanistic work, and cross-species statements are framed as conserved principles rather than as yeast-specific claims. (wood2018dot1landh3k79 pages 1-3, lee2018dot1regulatesnucleosome pages 1-2)
References
-
(wood2018dot1landh3k79 pages 1-3): Katherine Wood, Michael Tellier, and Shona Murphy. Dot1l and h3k79 methylation in transcription and genomic stability. Biomolecules, 8:11, Feb 2018. URL: https://doi.org/10.3390/biom8010011, doi:10.3390/biom8010011. This article has 249 citations.
-
(valenciasanchez2021regulationofthe pages 2-5): Marco Igor Valencia-Sánchez, Pablo De Ioannes, Miao Wang, David M. Truong, Rachel Lee, Jean-Paul Armache, Jef D. Boeke, and Karim-Jean Armache. Regulation of the dot1 histone h3k79 methyltransferase by histone h4k16 acetylation. Science, Jan 2021. URL: https://doi.org/10.1126/science.abc6663, doi:10.1126/science.abc6663. This article has 114 citations and is from a highest quality peer-reviewed journal.
-
(lee2018dot1regulatesnucleosome pages 1-2): Soyun Lee, Seunghee Oh, Kwiwan Jeong, Hyelim Jo, Yoonjung Choi, Hogyu David Seo, Minhoo Kim, Joonho Choe, Chang Seob Kwon, and Daeyoup Lee. Dot1 regulates nucleosome dynamics by its inherent histone chaperone activity in yeast. Nature Communications, Jan 2018. URL: https://doi.org/10.1038/s41467-017-02759-8, doi:10.1038/s41467-017-02759-8. This article has 55 citations and is from a highest quality peer-reviewed journal.
-
(separovich2022theposttranslationalregulation pages 49-52): The post-translational regulation of histone methylation enzymes by upstream signalling pathways This article has 0 citations.
-
(farooq2016themanyfaces pages 1-3): Zeenat Farooq, Shahid Banday, Tej K. Pandita, and Mohammad Altaf. The many faces of histone h3k79 methylation. Mutation research. Reviews in mutation research, 768:46-52, Apr 2016. URL: https://doi.org/10.1016/j.mrrev.2016.03.005, doi:10.1016/j.mrrev.2016.03.005. This article has 210 citations.
-
(farooq2016themanyfaces pages 3-4): Zeenat Farooq, Shahid Banday, Tej K. Pandita, and Mohammad Altaf. The many faces of histone h3k79 methylation. Mutation research. Reviews in mutation research, 768:46-52, Apr 2016. URL: https://doi.org/10.1016/j.mrrev.2016.03.005, doi:10.1016/j.mrrev.2016.03.005. This article has 210 citations.
-
(valenciasanchez2021regulationofthe media c5f9e665): Marco Igor Valencia-Sánchez, Pablo De Ioannes, Miao Wang, David M. Truong, Rachel Lee, Jean-Paul Armache, Jef D. Boeke, and Karim-Jean Armache. Regulation of the dot1 histone h3k79 methyltransferase by histone h4k16 acetylation. Science, Jan 2021. URL: https://doi.org/10.1126/science.abc6663, doi:10.1126/science.abc6663. This article has 114 citations and is from a highest quality peer-reviewed journal.
-
(valenciasanchez2021regulationofthe pages 18-21): Marco Igor Valencia-Sánchez, Pablo De Ioannes, Miao Wang, David M. Truong, Rachel Lee, Jean-Paul Armache, Jef D. Boeke, and Karim-Jean Armache. Regulation of the dot1 histone h3k79 methyltransferase by histone h4k16 acetylation. Science, Jan 2021. URL: https://doi.org/10.1126/science.abc6663, doi:10.1126/science.abc6663. This article has 114 citations and is from a highest quality peer-reviewed journal.
-
(welsem2018dot1promotesh2b pages 5-6): Tibor van Welsem, Tessy Korthout, Reggy Ekkebus, Dominique Morais, Thom M Molenaar, Kirsten van Harten, Deepani W Poramba-Liyanage, Su Ming Sun, Tineke L Lenstra, Rohith Srivas, Trey Ideker, Frank C P Holstege, Haico van Attikum, Farid El Oualid, Huib Ovaa, Iris J E Stulemeijer, Hanneke Vlaming, and Fred van Leeuwen. Dot1 promotes h2b ubiquitination by a methyltransferase-independent mechanism. Nucleic Acids Research, 46:11251-11261, Sep 2018. URL: https://doi.org/10.1093/nar/gky801, doi:10.1093/nar/gky801. This article has 45 citations and is from a highest quality peer-reviewed journal.
-
(wood2018dot1landh3k79 pages 5-7): Katherine Wood, Michael Tellier, and Shona Murphy. Dot1l and h3k79 methylation in transcription and genomic stability. Biomolecules, 8:11, Feb 2018. URL: https://doi.org/10.3390/biom8010011, doi:10.3390/biom8010011. This article has 249 citations.
-
(lee2018dot1regulatesnucleosome pages 9-10): Soyun Lee, Seunghee Oh, Kwiwan Jeong, Hyelim Jo, Yoonjung Choi, Hogyu David Seo, Minhoo Kim, Joonho Choe, Chang Seob Kwon, and Daeyoup Lee. Dot1 regulates nucleosome dynamics by its inherent histone chaperone activity in yeast. Nature Communications, Jan 2018. URL: https://doi.org/10.1038/s41467-017-02759-8, doi:10.1038/s41467-017-02759-8. This article has 55 citations and is from a highest quality peer-reviewed journal.
-
(lee2018dot1regulatesnucleosome pages 6-6): Soyun Lee, Seunghee Oh, Kwiwan Jeong, Hyelim Jo, Yoonjung Choi, Hogyu David Seo, Minhoo Kim, Joonho Choe, Chang Seob Kwon, and Daeyoup Lee. Dot1 regulates nucleosome dynamics by its inherent histone chaperone activity in yeast. Nature Communications, Jan 2018. URL: https://doi.org/10.1038/s41467-017-02759-8, doi:10.1038/s41467-017-02759-8. This article has 55 citations and is from a highest quality peer-reviewed journal.
-
(lee2018dot1regulatesnucleosome pages 10-11): Soyun Lee, Seunghee Oh, Kwiwan Jeong, Hyelim Jo, Yoonjung Choi, Hogyu David Seo, Minhoo Kim, Joonho Choe, Chang Seob Kwon, and Daeyoup Lee. Dot1 regulates nucleosome dynamics by its inherent histone chaperone activity in yeast. Nature Communications, Jan 2018. URL: https://doi.org/10.1038/s41467-017-02759-8, doi:10.1038/s41467-017-02759-8. This article has 55 citations and is from a highest quality peer-reviewed journal.
Artifacts
Citations
- valenciasanchez2021regulationofthe pages 2-5
- farooq2016themanyfaces pages 3-4
- separovich2022theposttranslationalregulation pages 49-52
- farooq2016themanyfaces pages 1-3
- valenciasanchez2021regulationofthe pages 18-21
- https://doi.org/10.1126/science.abc6663
- https://doi.org/10.1038/s41467-017-02759-8
- https://doi.org/10.1093/nar/gky801
- https://doi.org/10.3390/biom8010011
- https://doi.org/10.1016/j.mrrev.2016.03.005
- https://doi.org/10.3390/biom8010011,
- https://doi.org/10.1126/science.abc6663,
- https://doi.org/10.1038/s41467-017-02759-8,
- https://doi.org/10.1016/j.mrrev.2016.03.005,
- https://doi.org/10.1093/nar/gky801,
Perplexity
(DOT1-deep-research-perplexity.md)
Perplexity
(DOT1-deep-research-perplexity.md)DOT1: The Conserved Histone H3K79 Methyltransferase and Its Diverse Functions in Saccharomyces cerevisiae
The DOT1 gene in Saccharomyces cerevisiae encodes a highly conserved histone methyltransferase that catalyzes the mono-, di-, and trimethylation of histone H3 at lysine 79 (H3K79), a modification located uniquely within the globular domain of the histone rather than in the N-terminal tail where most histone modifications occur[1][7]. Originally identified through a genetic screen as a disruptor of telomeric silencing, DOT1 (also known as KMT4 or PCH1) has emerged as a multifunctional protein essential for maintaining genome stability, regulating transcription, controlling cell cycle progression, and ensuring proper meiotic division[1][10][32]. The protein belongs to the class I-like SAM-binding methyltransferase superfamily and possesses a unique non-SET domain structure that distinguishes it from other histone lysine methyltransferases, making it the founding member of the evolutionarily conserved Dot1 family found across eukaryotes[7][9][44]. Through its catalytic activity and multiple protein-protein interactions, DOT1 orchestrates complex chromatin remodeling events that affect fundamental cellular processes, with dysregulation of this enzyme and its human ortholog DOT1L implicated in various human pathologies including leukemias and developmental disorders.
Molecular Structure and Enzymatic Properties
Unique Structural Organization of the Dot1 Catalytic Domain
The Dot1 protein presents a distinctive structural architecture that sets it apart from other histone methyltransferases, particularly in its lack of a conserved SET domain typically found in histone lysine methyltransferases[9][44]. Instead, the yeast Dot1 protein, which is 582 amino acids in length, contains an elongated catalytic core composed of an N-terminal helical domain and a seven-stranded catalytic domain that harbors the binding site for S-adenosyl-L-methionine (SAM), the universal methyl donor[44]. Structural studies at 2.19 Ångstrom resolution revealed that this core catalytic region maintains an extended SAM conformation that distinguishes it from the folded conformation found in SET domain histone methyltransferases, suggesting fundamentally different catalytic mechanisms despite achieving the same biochemical outcome[44]. The active site pocket is lined with conserved hydrophobic residues that contribute to the specificity of substrate recognition and catalysis, and this catalytic core is highly conserved among Dot1 homologues across diverse organisms, with the core domain located at the C-terminus in yeast but repositioned to the N-terminus in mammals, Caenorhabditis elegans, Drosophila, and Anopheles gambiae[44]. Beyond the catalytic core, the yeast Dot1 protein contains a unique N-terminal region characterized by high lysine content, with at least two consensus monopartite nuclear localization signals that direct the protein to the nucleus[21].
The flexible, positively charged region at the C-terminus of the catalytic domain plays a critical role in nucleosome binding and enzymatic activity, distinguishing Dot1's approach to substrate recognition from other histone-modifying enzymes[9]. Cryo-EM structural studies examining Dot1 bound to nucleosomes with various histone modifications revealed that Dot1 can adopt multiple conformational states on the nucleosome surface, with distinct catalytic and non-catalytic conformations[18][22]. When bound to unmodified nucleosomes, the enzyme samples both conformations, but allosteric stimulation by H4K16 acetylation or H2B ubiquitination restricts and stabilizes the active catalytic conformation, leading to increased methylation rates[18][22]. This conformational selectivity mechanism explains how Dot1 integrates signals from other histone modifications to regulate its own activity with remarkable precision.
S-Adenosylmethionine Binding and Cofactor Requirements
As a SAM-dependent methyltransferase, Dot1 uses SAM as its methyl donor, with structural studies demonstrating a SAM binding pocket formed by conserved motifs characteristic of class I methyltransferases[25][44]. The interaction between Dot1 and SAM involves conserved sequence motifs scattered throughout the catalytic core, including motifs that directly contact the carboxypropyl moiety, ribose hydroxyls, and nitrogenous base of SAM, as well as motifs associated with the catalytic pocket[25][44]. The reaction mechanism proceeds through nucleophilic attack on the active methyl group in SAM, inducing cleavage of the carbon-sulfur bond and transfer of the methyl group to the lysine residue, producing S-adenosylhomocysteine (SAH) as the byproduct[44]. Unlike some SAM-dependent enzymes, Dot1 does not require additional metal cofactors for its catalytic activity, relying instead on the intrinsic chemistry of the SAM cofactor and the precise geometry of its active site to facilitate methyl transfer to histone substrates.
Primary Enzymatic Function and Substrate Specificity
The Unique Position of H3K79 as a Catalytic Target
The most distinctive feature of Dot1's enzymatic specificity lies in its exclusive methylation of histone H3 at lysine 79, a position located within loop 1 of the globular core domain of histone H3 rather than in the N-terminal tail where the vast majority of histone modifications occur[1][7][9][46]. This glycine-rich loop region containing K79 is exposed on the surface of the nucleosome where it remains solvent-accessible, positioned at the interface between the H3/H4 tetramer and the H2A/H2B dimer[1][7]. The solvent-exposed nature of K79 within the nucleosomal context permits access by the Dot1 enzyme despite the relatively constrained geometry of the nucleosome structure. The unique position of H3K79 within the globular domain rather than in the histone tail explains why Dot1 has evolved specialized recognition mechanisms distinct from other histone methyltransferases that typically target tail-embedded lysine residues. This location places H3K79 methylation in close proximity to key protein-DNA interaction surfaces and nucleosome-nucleosome contact points, positioning it as a modification capable of influencing higher-order chromatin structure and nucleosome dynamics.
Nucleosomal Substrate Preference and Chromatin-Specificity
A critical and distinctive feature of Dot1 enzymatic activity is its strong preference for nucleosomal substrates compared to free histones or recombinant H3 protein[1][7][9][27]. Biochemical characterization demonstrated that Dot1 exhibits essentially no activity on histone H3 in solution or free histone octamers but efficiently methylates H3K79 when histone H3 is incorporated into the nucleosomal context[1][27]. This nucleosomal specificity suggests that Dot1 recognizes multiple features of the nucleosome beyond simply the presence of the K79 lysine residue, requiring proper nucleosome assembly and chromatin structure for substrate recognition and catalysis[1]. The preferential activity on nucleosomal substrates indicates that Dot1 has evolved to respond to and modify chromatin in its native context, allowing for integration of signals from other nucleosomal features and modifications that can influence substrate accessibility and catalytic efficiency[18][22].
Non-Processive Mono-, Di-, and Trimethylation Mechanism
Distinct from many other histone methyltransferases that catalyze processive addition of multiple methyl groups in a single enzymatic engagement, Dot1 operates through a distributive mechanism, sequentially converting monomethylated H3K79 to dimethylated and subsequently to trimethylated H3K79 through multiple cycles of enzyme-substrate binding and product release[1][27][54]. In yeast, approximately 90% of total histone H3 bears H3K79 methylation, with the distribution heavily weighted toward the monomethylated form, followed by dimethylated and trace amounts of trimethylated H3K79[1][2]. In vitro studies using purified enzyme and nucleosome substrates revealed that Dot1 catalyzes all three methylation states at H3K79 in a non-processive manner, with monomethylation occurring more readily than the higher methylation states[54]. The requirements for progression through methylation states vary, with H2B ubiquitination providing particular support for the conversion from monomethylation to dimethylation and trimethylation states[1][27].
Regulation of Dot1 Catalytic Activity Through Histone Modifications
Cross-Talk with H2B Ubiquitination
The catalytic activity of Dot1 is subject to sophisticated regulation through interactions with other histone modifications, particularly histone H2B ubiquitination at lysine 123 (H2B-K123ub)[1][10][27]. The Rad6/Bre1 ubiquitin conjugating system catalyzes monoubiquitination of H2B at K123, and this modification functions as a critical regulatory signal that enhances Dot1-mediated H3K79 methylation[1][27][33]. Studies demonstrated that deletion of RAD6 or mutation of H2B-K123 severely impaired H3K79 dimethylation and trimethylation without substantially affecting monomethylation, indicating that H2B ubiquitination serves as a particularly important enhancer of the higher methylation states[1][33]. The molecular mechanism underlying this cross-talk involves multiple, potentially redundant pathways: ubiquitinated H2B can indirectly recruit Dot1 to methylate H3K79 through bridging factors, Dot1 can interact directly with ubiquitin through a lysine-rich region (amino acids 101-140) within its N-terminus that is also required for nucleosome binding, and H2B ubiquitination may cause chromatin structure changes to facilitate Dot1-mediated H3K79 methylation[1][27].
H4K16 Acetylation as an Allosteric Regulator
More recent structural and biochemical studies revealed that acetylation of histone H4 at lysine 16 (H4K16ac), a hallmark of decondensed, transcriptionally permissive chromatin, serves as a potent allosteric regulator that directly stimulates the catalytic activity of Dot1[18][22]. Cryo-EM structural analysis of Dot1 bound to nucleosomes with H4K16ac revealed that this modification restricts the conformational sampling space of Dot1, stabilizing it predominantly in a catalytic conformation that increases enzymatic activity approximately twofold compared to control substrates bearing only H2B ubiquitination[18][22]. When both H4K16ac and H2B ubiquitination are present on the same nucleosome, the stimulatory effects are synergistic, with doubly modified nucleosomes showing even greater stimulation of Dot1 activity than singly modified substrates[18][22]. Detailed kinetic analysis revealed that both H4K16ac and H2B ubiquitination increase the catalytic rate constant (kcat) of the methylation reaction while leaving the Michaelis constant (Km) essentially unchanged, indicating allosteric mechanism of activation that improves catalytic efficiency without altering substrate binding[18][22].
The mechanistic basis for H4K16ac stimulation involves direct contacts between the acetylated H4 N-terminus and critical residues in the Dot1 active site, specifically residues H347 and H355 of Dot1[18][22]. In unacetylated nucleosomes, these positively charged histidine residues may repel the positively charged, protonated amino group of unacetylated H4K16, destabilizing productive Dot1 conformations and reducing catalytic efficiency[18][22]. Acetylation of H4K16 neutralizes the positive charge, favoring stabilizing interactions that maintain the Dot1 active site in a catalytically competent conformation, thereby increasing the probability of productive methyl transfer reactions[18][22]. This elegant regulatory mechanism allows Dot1 to sense the acetylation status of chromatin and preferentially methylate H3K79 in transcriptionally active regions marked by H4K16 acetylation.
Competition with Sir3 for H4 Binding
In the context of telomeric heterochromatin formation, Dot1 activity is regulated through competitive interactions with the Sir3 heterochromatin protein for a limited binding site on the histone H4 N-terminal tail[1][26][29]. The basic patch region of H4 comprising residues R17H18R19 serves as a critical binding interface for both Dot1 and Sir3, and both proteins require this basic patch region for their respective functions[26][29]. A charge-based interaction between the acidic patch on Dot1 (comprising residues EDVDE) and the basic patch of H4 is essential for Dot1 binding and subsequent H3K79 methylation[26][29]. Sir3 and Dot1 compete for this same binding site, with the equilibrium between their binding determining whether the chromatin adopts a silenced or active state[1][26][29]. Acetylation of H4K16 disrupts Sir3 binding to the H4 tail while simultaneously enhancing Dot1 binding and catalytic activity, creating a bistable switch mechanism that coordinates the mutually exclusive binding of these two regulatory proteins[1][18][22][29].
Role in Transcriptional Regulation and Gene Silencing
Distribution of H3K79 Methylation Across the Genome
Genome-wide analyses demonstrate that H3K79 methylation is predominantly associated with actively transcribed chromatin regions, with approximately 90% of the yeast genome bearing H3K79 methylation marks, primarily in euchromatic regions[1][8][30][48]. H3K79me1, me2, and me3 are found predominantly within the coding regions of actively expressed genes and correlate strongly with transcriptional activity levels[1][32][49]. In contrast, H3K79 methylation is virtually absent from heterochromatic domains including telomeric repeats, silent mating-type loci (HML and HMR), and ribosomal DNA regions, where repressive chromatin structure is established and maintained by Sir proteins[8][16][48]. This mutually exclusive spatial distribution reflects the opposing functions of H3K79 methylation, which promotes an open euchromatic state, and the Sir proteins, which establish and maintain repressive heterochromatic structure.
Heterochromatin Boundary Function and Prevention of Sir Protein Spreading
While H3K79 methylation is abundant in euchromatic regions, its presence prevents non-specific interactions between heterochromatin-forming Sir proteins (Sir2, Sir3, and Sir4) and euchromatic nucleosomes, thereby promoting the proper compartmentalization of chromatin structure and restricting Sir protein accumulation to authentic heterochromatic domains[1][8][20][30][48]. Loss of Dot1 activity leads to mislocalization and spreading of Sir proteins beyond their normal heterochromatic boundaries, extending silencing to normally transcriptionally active regions and causing inappropriate gene repression[1][8][20][30]. Conversely, overexpression of Dot1 spreads H3K79 methylation into normally silent heterochromatic regions, displacing Sir proteins and causing derepression of normally silenced genes[1][27][30][48]. This observation demonstrates that Dot1 and H3K79 methylation function indirectly in promoting heterochromatin formation by creating an inhospitable environment for Sir protein binding in euchromatin rather than by directly activating gene expression[30][48].
Transcriptional Elongation and Association with RNA Polymerase II Complexes
Genome-wide localization studies revealed strong enrichment of H3K79 methylation within the coding regions of actively transcribed genes, particularly in the body of genes undergoing active transcription, suggesting a specific role for DOT1 in transcriptional elongation[1][27][32][49]. The Paf1 complex (Paf1C), a multi-subunit protein complex that associates with elongating RNA polymerase II (Pol II) throughout gene bodies, interacts with and regulates the methyltransferase activity of Dot1[1][32][37][40]. This interaction between Paf1C and Dot1 positions the methyltransferase at sites of active transcription elongation, where it deposits H3K79 methylation marks that facilitate continued productive Pol II elongation[1][37][40]. Additionally, human DOT1L purifies in association with various Pol II elongation complexes, and overexpression of MLL fusion proteins that aberrantly recruit DOT1L results in elevated H3K79 methylation at target genes accompanied by increased transcription elongation and expression[1][49].
Methylation-Independent and Methylation-Dependent Derepression Functions
Targeted recruitment studies using tethered Dot1 revealed that the protein possesses two distinct derepression mechanisms, one dependent on its methyltransferase activity toward H3K79 and another independent of catalytic activity[8][24][48]. The methylation-independent derepression mechanism requires the N-terminal domain of Dot1 and involves chromatin relocalization away from the repressive nuclear periphery, with the histone acetyltransferase Gcn5 appearing to mediate this relocalization function[8][24][48]. Even catalytically inactive Dot1 proteins retain the ability to reposition heterochromatic telomeres from the nuclear periphery to the interior, suggesting that Dot1 binding itself, independent of H3K79 methylation, can alter local chromatin organization and accessibility[8][24]. The methylation-dependent derepression mechanism involves direct antagonism of Sir protein binding through locally elevated H3K79 methylation that prevents Sir complex assembly and function[8][24][48]. These dual mechanisms allow Dot1 to exert both short-range effects through H3K79 methylation and long-range effects through chromatin remodeling and repositioning.
Role in DNA Damage Response and Checkpoint Control
Recruitment of 53BP1 and Rad9 to DNA Double-Strand Breaks
The first direct evidence linking DOT1-mediated H3K79 methylation to DNA damage repair came from discovery that the tandem tudor domain of the human 53BP1 protein, the vertebrate ortholog of budding yeast Rad9, specifically recognizes and binds to methylated H3K79[1][10][31][32][38]. The tudor domain of 53BP1 functions as a methyl-lysine reader domain that directly contacts H3K79me2 and H3K79me3 at sites of DNA double-strand breaks (DSBs), with this interaction being essential for efficient recruitment of 53BP1 to DSB sites[1][31][32]. Mutation of the tudor domain or blocking H3K79 methylation through either mutation of H3K79 to arginine or knockdown of DOT1L expression significantly impaired recruitment of 53BP1 to ionizing radiation-induced foci, demonstrating the functional importance of this interaction in vivo[1][10]. The yeast Rad9 protein similarly recognizes methylated H3K79 via its tudor domain, and this interaction is critical for Rad9 recruitment to DSBs and subsequent activation of the Rad53 checkpoint kinase[10][17][34][38].
G1 and Intra-S Phase Checkpoint Control
Budding yeast cells bearing deletion mutations of DOT1 display striking defects in both the G1 and intra-S phase DNA damage checkpoints in response to ionizing radiation[1][10][38]. When wild-type yeast cells synchronized in G1 are treated with ionizing radiation, they undergo a G1 checkpoint delay that prevents entry into S phase, allowing time for DNA damage repair before the cell commits to DNA replication[1][38]. In contrast, DOT1 deletion mutants fail to display this checkpoint delay, progressing through Start and into S phase despite unrepaired DNA damage, resulting in markedly increased accumulation of mutations[1][10][38]. Similarly, wild-type cells irradiated during S phase exhibit a checkpoint response that slows DNA replication progression (intra-S phase checkpoint), while DOT1 mutants complete S phase within normal timeframes regardless of radiation exposure[1][38]. These checkpoint defects are not due to accumulation of additional mutations in checkpoint genes, as DOT1 mutants show normal expression of classical checkpoint proteins like Rad17 and Mec1[1][38].
Critical evidence that DOT1's checkpoint function requires its methyltransferase activity came from complementation experiments in which catalytically inactive dot1-Gly401Arg mutant failed to restore checkpoint function despite being expressed at normal levels, whereas wild-type DOT1 completely restored the checkpoint response[1][38]. Furthermore, yeast expressing a H3K79R histone mutation that prevents methylation exhibited checkpoint defects similar to DOT1 deletion mutants, definitively establishing that H3K79 methylation, rather than some other function of the DOT1 protein, is required for checkpoint control[1][38]. The mechanism appears to involve H3K79me-dependent recruitment of Rad9 to the damage sites, as both Rad9 and DOT1 mutations produce similar checkpoint phenotypes and act in the same pathway[1][10][38].
Homologous Recombination and DNA Repair
Beyond its role in checkpoint arrest, Dot1-mediated H3K79 methylation promotes DNA repair through homologous recombination (HR) mechanisms[1][10][32]. DOT1 and H2B ubiquitination are required for efficient repair of ionizing radiation-induced DSBs via HR, with DOT1 deletion mutants showing reduced capacity to repair DSBs using the HR pathway[1][32]. The mechanism involves Dot1-dependent H3K79 methylation facilitating recruitment of repair factors to DSB sites and potentially promoting the resection of DSB ends, a critical early step in HR that generates the 3' single-stranded DNA overhangs required for homologous recombination[1][32]. Both H3K79 methylation and Rad9 recruitment appear to play integral roles in regulating the extent of DSB resection to prevent excessive ssDNA production that would interfere with HR efficiency[1][32].
Ultraviolet Damage Response and Nucleotide Excision Repair
Beyond DSB repair, DOT1-mediated H3K79 methylation plays a critical role in repairing ultraviolet (UV)-induced DNA damage through multiple pathways[1][5][32]. Loss of H3K79 methylation results in hypersensitivity to UV radiation, with cells lacking Dot1 activity showing substantially reduced survival following UV exposure[5][32][49]. The mechanisms by which H3K79 methylation contributes to UV damage repair are multifaceted, involving support of nucleotide excision repair (NER), recombination repair (RR), and post-replication repair (PRR) pathways, all three of which are compromised when H3K79 methylation is eliminated[1][32]. The direct involvement of Dot1 in NER is supported by the observation that UV survival patterns in dot1Δ strains are similar to rad1Δ strains (deficient in NER), and double dot1Δ rad1Δ mutants show no additional hypersensitivity beyond either single mutant, suggesting DOT1 participates in NER-mediated repair[1][32].
Regulation of Translesion Synthesis
In addition to promoting canonical DNA repair pathways, Dot1 negatively regulates translesion synthesis (TLS), a mutagenic repair pathway that allows DNA polymerase progression past unrepaired lesions at the cost of introducing mutations[1][32][35]. Dot1 inhibits the operation of error-prone polymerases like Polζ/Rev1 that catalyze TLS, thereby maintaining genome integrity and preventing accumulation of mutations upon DNA damage[1][32][35]. When DNA damage cannot be repaired through conventional mechanisms, TLS becomes necessary for cell survival, and Dot1-mediated inhibition of TLS helps prioritize canonical repair pathways over mutagenic TLS before resorting to the less accurate repair approach[1][32][35]. This represents an important quality control mechanism that protects genome stability by suppressing mutagenic repair pathways in favor of high-fidelity alternatives when possible.
Role in Meiotic Checkpoint Control and Chromosome Segregation
The Pachytene Checkpoint and DOT1 Function
During meiotic cell division, Dot1 plays an essential role in the pachytene checkpoint, a surveillance mechanism that arrests meiotic prophase I when chromosome synapsis and recombination are defective[1][14][21][41][50]. The pachytene checkpoint monitors proper synapsis of homologous chromosomes via the synaptonemal complex and completion of recombination to prevent premature meiotic progression and aneuploidy[1][14][21]. In yeast strains bearing mutations in genes required for synaptonemal complex formation (such as ZIP1) or meiotic recombination (such as DMC1), the pachytene checkpoint normally prevents exit from meiotic prophase I, maintaining cell cycle arrest until synapsis and recombination defects are resolved[1][14][21]. However, in DOT1 deletion mutants, the ZIP1 and DMC1 mutants inappropriately progress through meiosis and sporulation despite persistent defects in chromosome synapsis and recombination, resulting in the formation of inviable spores with fragmented chromosomes[1][14][21][41].
The checkpoint arrest function of Dot1 correlates with its role in maintaining proper nuclear localization and concentration of checkpoint proteins, particularly Pch2 and Sir2[1][14][21][41]. In wild-type meiotic cells, Pch2 (a nucleolar protein) and Sir2 accumulate in the nucleolus during pachytene, and this localization depends on proper Sir protein function and Dot1 activity[1][14][21]. Disruption of DOT1 leads to mislocalization and loss of nucleolar concentration of both Pch2 and Sir2, causing failure of the checkpoint and inappropriate resumption of meiotic division[1][21][41]. Additionally, Sir3, normally a telomeric heterochromatin protein, becomes delocalized from telomeres in dot1Δ mutants, suggesting that Dot1's chromatin-organizing functions affecting Sir protein distribution are critical for both telomeric silencing and meiotic checkpoint control[1][14][21][41].
Regulation of Meiotic Double-Strand Break Repair
Beyond its checkpoint function, Dot1 regulates the choice of DNA repair pathway for meiotic double-strand breaks (DSBs)[1][14][41][50]. During normal meiosis, meiotic DSBs are repaired using the homologous recombination pathway, with homologous chromosomes serving as the primary repair template, ensuring proper chromosome segregation and genetic recombination[1][14][21]. However, in the absence of DMC1 protein (required for homologous recombination), meiotic DSBs can be repaired through an alternative pathway utilizing sister chromatid recombination, resulting in loss of genetic material and aneuploidy[1][14][21][41]. Dot1 actively blocks this error-prone sister chromatid recombination pathway through a Rad54-dependent mechanism, preventing inappropriate use of sister chromatids as repair templates when proper homologous recombination is available[1][14][21][41].
When DOT1 is deleted in DMC1 mutants, the meiotic DSBs are repaired using the sister chromatid pathway, and this repair proceeds relatively efficiently, allowing cell cycle progression and sporulation[1][14][41]. However, spore viability is dramatically reduced, as the spores produced contain significant chromosomal abnormalities and fragmented DNA due to aberrant recombination between sister chromatids rather than homologous chromosomes[1][14][41]. This demonstrates that Dot1 serves as a critical regulator of repair pathway choice during meiosis, ensuring that homologous recombination is the primary mechanism for resolving meiotic DSBs.
Cell Cycle-Regulated Expression and Dynamic H3K79 Methylation Patterns During Meiosis
The expression of the DOT1 gene is cell cycle-regulated, with the promoter containing a cell cycle box element recognized by the MBF transcription factor, and microarray analyses showing that DOT1 transcript levels peak during G1 phase of the mitotic cell cycle[1][21][41]. In contrast to the G1-phase peak of DOT1 transcript abundance in mitotic cells, the global level of H3K79me2 begins to increase during S phase, suggesting that the methylation mark accumulates as the enzyme deposits it on newly replicated chromatin[1][32]. During meiotic prophase I, DOT1L localization and H3K79 methylation exhibit distinctive spatio-temporal dynamics, with very low levels of DOT1L and H3K79me2/me3 at the leptonema stage (early meiotic prophase) that increase progressively through zygonema and reach dramatic elevations at pachynema and diplonema[19][55]. The H3K79me3 mark exhibits particularly interesting localization patterns, becoming strongly enriched in the heterochromatic centromeric regions at pachynema, with roughly sevenfold accumulation relative to euchromatic chromatin, and additionally concentrating in the sex body (the transcriptionally silent XY bivalent) during late prophase I[19][55].
Cellular Localization and Subnuclear Distribution
Nuclear Localization and Nucleolar Accumulation
The DOT1 protein contains at least two consensus monopartite nuclear localization signals within its N-terminal lysine-rich domain, directing the protein to the nucleus where it carries out its enzymatic functions[21][41]. Studies using fluorescently tagged Dot1-GFP fusion proteins expressed from yeast plasmids revealed that Dot1 localizes predominantly to the nucleus, with distinctive subcellular distribution patterns in vegetative versus meiotic cells[21][41]. In vegetative mitotic cells, Dot1-GFP signal is often enriched toward one side of the nucleus in a region that stains faintly with DAPI (DNA stain) and corresponds to the nucleolus, suggesting accumulation at this subnuclear site[21][41]. This nucleolar enrichment is evident even when Dot1-GFP is expressed from low-copy plasmids, indicating that the nucleolar localization is not an artifact of protein overexpression but rather represents a genuine subcellular accumulation pattern[21][41].
In meiotic cells during the early and middle stages of meiotic prophase I, Dot1 localization is more uniformly nuclear without pronounced nucleolar enrichment, until late in meiosis during spore formation when nucleolar accumulation again becomes evident[19][21][41]. Surface spreading of meiotic chromosomes combined with immunofluorescence using anti-HA antibodies recognizing Dot1-HA fusion proteins revealed two distinct patterns of Dot1 localization in vegetative cell nuclei: in approximately 29% of nuclei, Dot1 is distributed relatively homogeneously throughout chromatin, while in the remaining nuclei, Dot1 is clearly enriched in the nucleolar region[21][41]. These observations suggest that Dot1 localizes to both nuclear compartments and, in particular, accumulates at the nucleolus where it may interact with nucleolar proteins and chromatin, including the ribosomal DNA (rDNA) regions that are subject to Sir-mediated silencing.
Chromatin Binding and Histone Interactions
Dot1 demonstrates a strong preference for binding to chromatin in the context of intact nucleosomes rather than to DNA or free histones, consistent with its requirement for nucleosomal context for enzymatic activity[1][9][26][29]. The protein forms extensive contacts with multiple histone proteins and regions of the nucleosome during binding, with particular importance of the histone H4 N-terminal tail basic patch region (comprising residues R17, H18, and R19) for both binding and catalytic activity[20][26][29]. In vitro binding assays demonstrated that Dot1 can directly interact with histone H4 peptides encompassing the basic patch region through an acidic patch on Dot1 (EDVDE), establishing a charge-based interaction critical for nucleosome recognition[20][26][29]. The interaction between Dot1 and the H4 basic patch is modulated by histone modifications and other binding proteins, including the Sir3 heterochromatin protein that competes for the same binding site[1][20][26][29].
While the histone H4 tail contributes substantially to Dot1 catalytic activity, nucleosomes lacking histone N-terminal tails (prepared by trypsin digestion to remove tail domains) retain significant Dot1 binding capacity, albeit with somewhat reduced binding compared to intact nucleosomes[26][29]. This observation indicates that Dot1 nucleosome binding involves multiple contact points beyond the H4 tail, including interactions with the histone octamer core and possibly with DNA[9][26][29]. The flexible positively charged region at the C-terminus of the Dot1 catalytic domain is particularly important for nucleosome binding and enzymatic activity, suggesting that this region makes critical contacts with nucleosomal features necessary for substrate engagement[9].
Evolutionary Conservation and Human DOT1L Ortholog
Conservation Across Eukaryotes and Functional Homology
The DOT1 gene is evolutionarily conserved across eukaryotes, with homologous genes identified in humans (DOT1L, also called KMT4), plants, insects, protists, and other microorganisms[1][4][6]. The human DOT1L protein shares approximately 70% sequence identity with the yeast Dot1 enzyme and retains the characteristic class I methyltransferase fold and SAM-binding domain[1][6]. Importantly, human DOT1L can functionally substitute for yeast Dot1 protein when targeted to yeast telomeres, producing similar derepression of heterochromatin, demonstrating remarkable functional conservation despite the evolutionary divergence between yeast and mammals[1][8][48]. The conservation of both sequence and function across such evolutionary distance underscores the fundamental importance of this enzymatic activity and the diverse processes it regulates.
DOT1L in Mammalian Development and Human Disease
In mammals, DOT1L is essential for normal embryonic development, with knockout studies in mice revealing that loss of DOT1L function results in embryonic lethality with pronounced defects in yolk sac angiogenesis[2]. DOT1L-deficient embryonic stem cells show global loss of H3K79 methylation accompanied by reduced levels of heterochromatic marks at centromeres and telomeres, changes that are accompanied by aneuploidy, telomere elongation, and severe proliferation defects[2]. The mammalian DOT1L has been particularly intensively studied in the context of mixed-lineage leukemia (MLL) translocations, where fusion proteins containing MLL are aberrantly recruited to target genes with DOT1L, resulting in elevated H3K79 methylation and leukemogenic gene expression[1][6][27][32][49]. Small molecule inhibitors of DOT1L catalytic activity have been developed as potential therapeutic agents for MLL-rearranged leukemias, with clinical trials demonstrating efficacy in preventing the proliferation of leukemic cells bearing MLL translocations[6].
DOT1L also plays important roles in mammalian spermatogenesis, where it acts as a transcriptional activator for genes responsible for the histone-to-protamine transition during sperm maturation[6]. Additionally, DOT1L-mediated H3K79 methylation participates in V(D)J recombination and somatic hypermutation in B lymphocytes, supporting the high-rate transcription necessary for these specialized processes in immune cells[49][51].
Integration and Synthesis of DOT1 Functions
Cross-Talk Between Telomeric Silencing and Transcriptional Activation
A central paradox in understanding DOT1 function is that H3K79 methylation occurs predominantly in transcriptionally active euchromatin, yet loss of Dot1 activity severely impairs telomeric silencing, suggesting that Dot1's role in silencing is indirect rather than through direct activation of silencing machinery[1][8][30][48]. Current evidence indicates that Dot1 affects telomeric silencing primarily through creating an euchromatic environment (marked by H3K79 methylation) that is inhospitable to Sir protein binding and function[1][8][30][48]. This interpretation is strongly supported by the observation that Sir proteins actively prevent methylation of H3K79 by Dot1, establishing a mutually antagonistic relationship between the two silencing mechanisms[1][8][30]. Overexpression of Dot1, which extends H3K79 methylation into heterochromatic domains, disrupts Sir protein function and causes derepression of normally silent genes, while loss of Dot1 allows Sir proteins to extend their influence into euchromatic regions[1][8][30][48].
The Role of Multiple Regulatory Mechanisms in Preventing Ectopic Silencing
The multi-layered regulation of Dot1 activity through H2B ubiquitination, H4K16 acetylation, and Sir protein competition appears designed to ensure that H3K79 methylation is deposited exclusively in the euchromatic context where it is compatible with active transcription and prevents ectopic heterochromatin formation[1][18][22][26][27][29][30]. H4K16 acetylation, which broadly marks transcriptionally active chromatin, directly stimulates Dot1 catalytic activity, ensuring that H3K79 methylation is enriched in regions poised for transcription[1][18][22][27]. H2B ubiquitination, which is coupled to transcription elongation, provides additional support for Dot1 activity, further linking H3K79 methylation to active transcription[1][27][37][40]. The competition between Dot1 and Sir3 for H4 tail binding creates a bistable switch that establishes clear boundaries between euchromatic and heterochromatic domains[1][26][29][30]. This multifaceted regulation ensures genomic stability by maintaining appropriate chromatin domain organization and preventing the inappropriate invasion of heterochromatin into transcribed regions.
Cell Cycle Regulation and Coordination with Replication
The cell cycle-regulated expression of DOT1, with peak transcript levels at G1 phase, coordinates DOT1 protein production with the early G1 phase when decisions about cell cycle progression and checkpoint responses are made[1][21][32][41][56]. However, the global H3K79 methylation mark begins accumulating during S phase, suggesting that DOT1 protein synthesized at G1 is subsequently deployed during DNA replication to mark newly synthesized chromatin with H3K79 methylation[1][32][56]. This temporal coordination between DOT1 expression and H3K79 methylation accumulation couples the expression of the methyltransferase enzyme with subsequent chromatin modification during replication, ensuring that newly replicated histones are promptly modified to establish appropriate euchromatic states[1][32][56]. The requirement for Dot1 in both G1 and intra-S phase checkpoint control further underscores the importance of this enzyme for maintaining genome stability throughout the cell cycle.
Conclusion
The DOT1 gene of Saccharomyces cerevisiae encodes a highly specialized histone methyltransferase that catalyzes the mono-, di-, and trimethylation of histone H3 at lysine 79, a modification uniquely positioned within the globular domain of the histone rather than in histone tails[1][7][9]. Through its unique structural organization lacking the SET domain found in other histone lysine methyltransferases, DOT1 achieves nucleosomal substrate specificity and responsive regulation through allosteric mechanisms triggered by H4K16 acetylation and H2B ubiquitination[1][9][18][22]. The biological functions of DOT1-mediated H3K79 methylation encompass transcriptional regulation, where it marks active genes and prevents Sir protein ectopic binding to euchromatic chromatin; DNA damage response, where it enables recruitment of the 53BP1/Rad9 checkpoint proteins to DSBs through methylated H3K79 recognition by tudor domains; checkpoint control in both mitotic and meiotic cell cycles, where it regulates progression decisions and pathway choice for DNA repair; and meiotic recombination, where it ensures homologous chromosome pairing and proper use of homologous recombination for meiotic DSB repair[1][10][14][21][32][38][41].
The sophistication of DOT1 regulation through multiple histone modifications and the multifunctional nature of its enzymatic products highlight the central importance of this modification in cellular processes ranging from transcriptional control to genome maintenance. The evolutionary conservation of DOT1 across eukaryotes and the critical roles of its mammalian ortholog DOT1L in normal development and cellular differentiation, coupled with its aberrant activity in human leukemias, underscore its fundamental biological significance. Future research into the precise mechanisms by which H3K79 methylation coordinates with other histone modifications and protein factors to regulate distinct cellular processes, as well as the structural basis for differential responses of Dot1 to its various allosteric regulators, will continue to enhance our understanding of chromatin biology and epigenetic gene regulation.
Citations
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3134078/
- https://journals.plos.org/plosgenetics/article?id=10.1371%2Fjournal.pgen.1000190
- https://pubmed.ncbi.nlm.nih.gov/12097318/
- https://www.yeastgenome.org/locus/S000002848
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4041417/
- https://en.wikipedia.org/wiki/DOT1L
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4889126/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3038881/
- https://pubmed.ncbi.nlm.nih.gov/12628190/
- https://www.uniprot.org/uniprotkb/Q8TEK3/entry
- https://www.molbiolcell.org/doi/10.1091/mbc.11.10.3601
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2724485/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3085244/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2691753/
- https://www.science.org/doi/10.1126/science.abc6663
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3969405/
- https://genesdev.cshlp.org/content/21/16/2018.full
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5770421/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7982431/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1948857/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5871980/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1282579/
- https://genesdev.cshlp.org/content/early/2007/08/03/gad.1560607
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2643802/
- https://genesdev.cshlp.org/content/25/13/1345.full
- https://pubmed.ncbi.nlm.nih.gov/12167634/
- https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2021.635383/full
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2475726/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2678028/
- https://www.pnas.org/doi/10.1073/pnas.1904324116
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1265753/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3979909/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3296560/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2688786/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6280759/
- https://febs.onlinelibrary.wiley.com/doi/10.1111/febs.17062
- https://pmc.ncbi.nlm.nih.gov/articles/PMC15018/
- https://www.pnas.org/doi/10.1073/pnas.2104013118
- https://www.molbiolcell.org/doi/abs/10.1091/mbc.11.10.3601
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4650758/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10036116/
- https://www.frontiersin.org/journals/genetics/articles/10.3389/fgene.2022.828086/full
- https://academic.oup.com/nar/article/51/5/2117/7009124