Deep Research
Falcon
(PHD1-deep-research-falcon.md)
Falcon
(PHD1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Functional Annotation of Saccharomyces cerevisiae PHD1 (UniProt P36093; gene PHD1; locus YKL043W)
1) Target identity verification (mandatory)
The research target is PHD1 from Saccharomyces cerevisiae (S288c background; systematic locus YKL043W), encoding Phd1, a transcription factor implicated in filamentous differentiation programs (pseudohyphal and invasive growth). Multiple primary studies explicitly name PHD1/Phd1 in S. cerevisiae and characterize it as a filamentation transcription factor that regulates FLO11/MUC1 and other differentiation regulators, matching the UniProt context of a putative transcription factor rather than an enzyme or unrelated “PHD” domain protein. (raithatha2012cdk8regulatesstability pages 1-2, pan2000sok2regulatesyeast pages 1-2)
2) Key concepts and definitions (current understanding)
2.1 Filamentous growth in budding yeast
Saccharomyces cerevisiae can switch from unicellular budding to multicellular, elongated growth forms under nutrient limitation. Two commonly discussed forms are:
- Pseudohyphal growth (classically in diploids), characterized by elongated cells that remain attached and form filaments.
- Invasive growth (classically in haploids), involving agar invasion and adhesion.
Both phenotypes require coordinated control of morphology, polarity, and especially cell-surface adhesion, with FLO11/MUC1 a central effector gene encoding a surface glycoprotein (flocculin). (pan2000sok2regulatesyeast pages 1-2, kumar2021thecomplexgenetic pages 5-7)
2.2 What PHD1/Phd1 is (molecular function)
Phd1 is a transcription factor positioned as a high-level regulator (“master regulator”) of filamentous differentiation gene expression programs, rather than a catalytic enzyme or transporter. Its biological role is therefore best defined by the transcriptional programs it activates/represses and by how its activity is controlled by upstream signaling and protein stability. (raithatha2012cdk8regulatesstability pages 1-2)
3) Core functional annotation: processes, targets, pathways, and mechanism
3.1 Primary biological process: regulation of filamentous differentiation and adhesion via FLO11
A central experimentally supported role for Phd1 is regulation of cell–cell/cell–surface adhesion and filamentation through FLO11:
- In a transcription-factor cascade context, Pan & Heitman (2000) report that Phd1 and Ash1 regulate expression of the cell surface protein Flo11, and that both factors are largely required for filamentation in a sok2 hyperfilamentous background. (Publication date: 2000-11; URL: https://doi.org/10.1128/MCB.20.22.8364-8372.2000) (pan2000sok2regulatesyeast pages 1-2)
- A later authoritative synthesis (Annual Review of Genetics) places Phd1 in a Sok2→Phd1/Ash1/Swi5 transcription-factor cascade capable of regulating FLO11 “independently of the PKA and MAPK pathways,” framing Phd1 as part of a layered regulatory architecture converging on FLO11. (Publication date: 2021-11; URL: https://doi.org/10.1146/annurev-genet-071719-020249) (kumar2021thecomplexgenetic pages 5-7)
3.2 Genetic/phenotypic evidence for PHD1 as a differentiation driver
Multiple lines of genetic evidence support Phd1 as a potent differentiation regulator:
- Overexpression of PHD1 strongly promotes pseudohyphal growth even on nitrogen-rich medium; in Pan & Heitman (2000), PHD1 overexpression can suppress pseudohyphal defects of tpk2 and ste12 mutants, indicating Phd1 can drive downstream differentiation programs even when canonical upstream regulators are compromised. (pan2000sok2regulatesyeast pages 1-2)
- Conversely, phd1/phd1 mutants did not show obvious pseudohyphal defects under the conditions emphasized in that study, suggesting background/condition dependence and redundancy among differentiation regulators. (pan2000sok2regulatesyeast pages 1-2)
- Raithatha et al. (2012) also describe Phd1 as a “master regulator” of filamentation and report that PHD1 expression is induced under nitrogen starvation and Phd1 accumulates under differentiating conditions, consistent with a role in initiating/maintaining the program. (Publication date: 2012-02; URL: https://doi.org/10.1128/MCB.05420-11) (raithatha2012cdk8regulatesstability pages 1-2)
3.3 Upstream signaling and transcriptional inputs: MAPK/PKA/Snf1 context and the Sok2 axis
Phd1 sits within a broader network in which nutrient sensing feeds into transcriptional regulators controlling FLO11 and morphology:
- The broader differentiation system is described as being regulated by MAPK (Kss1), Ras–cAMP–PKA, and Snf1/AMPK signaling; this context is explicitly discussed alongside PHD1 regulation in Raithatha et al. (2012). (raithatha2012cdk8regulatesstability pages 1-2)
- Pan & Heitman (2000) place Phd1 in relation to the cAMP/PKA and MAPK systems and suggest Phd1 may act distinct from those pathways in some contexts, consistent with multiple partially independent inputs to FLO11 regulation. (pan2000sok2regulatesyeast pages 1-2)
- Kumar (2021) highlights that Sok2 is thought to act downstream of cAMP/PKA but can activate a TF cascade (including Phd1) that regulates FLO11 “independently” of direct PKA/MAPK control, emphasizing network modularity and redundancy. (kumar2021thecomplexgenetic pages 5-7)
3.4 Post-translational regulation and mechanism: Cdk8-dependent turnover of Phd1
A major mechanistic advance in understanding Phd1 is that its activity is controlled not only transcriptionally but also by protein stability:
- Raithatha et al. (2012) provide experimental evidence that Phd1 is intrinsically unstable and that its degradation is initiated via Cdk8-dependent phosphorylation (Cdk8 is a kinase associated with the RNA polymerase II Mediator complex). (raithatha2012cdk8regulatesstability pages 1-2)
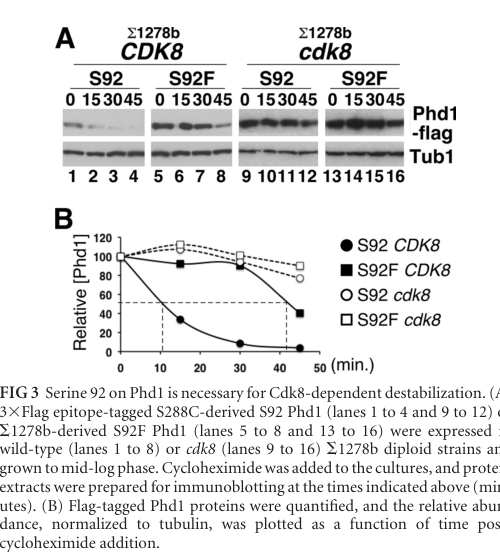
- Quantitative stability measurements by cycloheximide-chase show a short half-life (~10–15 min) in rich medium, and strong stabilization in conditions where Cdk8 activity is absent or reduced. (raithatha2012cdk8regulatesstability pages 6-7)
- Under nitrogen limitation, Phd1 becomes progressively stabilized: half-life increases to ~40 min after 2 h in nitrogen-limiting SLAD and to >45 min after 4 h, consistent with a differentiation-triggered stabilization mechanism. (raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability media d01b8851)
- In cdk8Δ, Phd1 is strongly stabilized (>45 min), consistent with Cdk8 acting as a negative regulator through turnover. (raithatha2012cdk8regulatesstability pages 2-3)
- A notable genetics–biochemistry bridge is a natural polymorphism in a differentiating strain background (1278b) that removes a candidate Cdk8 phosphosite (S92F), stabilizing Phd1 and enhancing filamentation, supporting causality between phosphorylation site, stability, and phenotype. (raithatha2012cdk8regulatesstability pages 1-2, raithatha2012cdk8regulatesstability pages 6-7)
Visual evidence: cycloheximide-chase panels and the regulatory model were retrieved from Raithatha et al. (2012), including stability curves and a pathway model summarizing how nutrient limitation reduces Cdk8 activity to permit accumulation of Phd1 and other transcription factors. (raithatha2012cdk8regulatesstability media d01b8851, raithatha2012cdk8regulatesstability media c487618b)
4) Subcellular localization (what is known vs. evidence gaps)
Direct experimental localization evidence (e.g., GFP microscopy or cell fractionation explicitly stating nuclear localization) was not found in the retrieved excerpts. However:
- Raithatha et al. (2012) used FLAG-tagged Phd1 for biochemical detection and treat Phd1 as a DNA-binding transcriptional regulator; nuclear function is therefore strongly implied but not directly demonstrated in the retrieved text segments. (raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability pages 3-4)
Accordingly, the most defensible annotation from the retrieved evidence is:
- Cellular compartment (evidence level): nuclear localization is inferred from transcription factor function and Mediator/Cdk8 context, but direct localization assays were not available in the gathered evidence. (raithatha2012cdk8regulatesstability pages 1-2)
5) Recent developments (prioritizing 2023–2024)
Direct 2023–2024 mechanistic papers specifically dissecting Phd1 regulation were not retrieved by the search tools. However, one 2024 primary study provides contemporary functional context:
5.1 PHD1 in structured colony morphology (2024 RNA-seq + genetics)
Cromie et al. (2024) examined gene-expression programs underlying “ruffled/structured” colony morphology and performed deletion tests in a structured-colony background (F13). They report:
- PHD1 (YKL043W) is in a gene-expression cluster whose expression correlates positively with colony structure.
- Yet, phd1Δ had little effect on colony morphology at day 5 (colonies still displayed an outer structured zone and inner smooth zone), whereas flo11Δ or msb2Δ produced fully smooth colonies. (Publication date: 2024-09; URL: https://doi.org/10.1371/journal.pone.0311061) (cromie2024spatiotemporalpatternsof pages 21-23)
Interpretation: This supports the view that PHD1 participates in a broader colony-structure/filamentation-associated transcriptional state, but may be nonessential or redundant in at least some genetic backgrounds and environmental regimes—consistent with older observations that loss of PHD1 alone can show modest phenotypes while overexpression is strongly pro-filamentation. (pan2000sok2regulatesyeast pages 1-2, cromie2024spatiotemporalpatternsof pages 21-23)
6) Current applications and real-world implementations
6.1 Synthetic biology: externally controlled induction of pseudohyphal growth
A clear real-world implementation of PHD1 knowledge is in synthetic gene regulation:
- Pothoulakis & Ellis (2018) engineered synthetic regulatory systems to control expression of native PHD1 and FLO8, demonstrating that externally controlled induction of these transcription factors can trigger pseudohyphal growth in both diploid and haploid strains, including in rich media. This establishes PHD1 as an actionable “handle” to program multicellular morphology in yeast. (Publication date: 2018-01; URL: https://doi.org/10.1038/s42003-017-0008-0) (pothoulakis2018syntheticgeneregulation pages 1-2)
Application relevance: controllable filamentation can be used as a chassis capability for engineered surface adhesion, structured multicellular assemblies, and programmable colony architectures, leveraging natural differentiation modules. (pothoulakis2018syntheticgeneregulation pages 1-2)
7) Expert opinions and authoritative synthesis
- Kumar (2021) emphasizes that pseudohyphal growth is governed by a complex genetic basis and multilayered regulatory control, with multiple pathways converging on FLO11 and a large gene set enabling morphological transition. Within this framework, the Sok2→Phd1/Ash1/Swi5 cascade represents one route to FLO11 regulation that can bypass canonical PKA/MAPK control, illustrating the field’s view that filamentation is controlled by redundant, context-dependent regulatory modules. (kumar2021thecomplexgenetic pages 5-7)
- Raithatha et al. (2012) contributes an expert mechanistic interpretation that controlling differentiation can hinge on changing the stability of labile transcription factors (including Phd1) by down-regulating Cdk8 activity under nitrogen limitation, highlighting protein turnover as a regulatory “gate” for cell-fate transitions. (raithatha2012cdk8regulatesstability pages 8-9)
8) Key statistics and quantitative data (from recent/primary studies)
- Phd1 half-life in rich medium: ~10–15 min (cycloheximide chase). (raithatha2012cdk8regulatesstability pages 6-7)
- Phd1 half-life after nitrogen limitation: ~40 min after 2 h in SLAD; >45 min after 4 h in SLAD. (raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability media d01b8851)
- Phd1 half-life in cdk8Δ: >45 min (stabilized). (raithatha2012cdk8regulatesstability pages 2-3)
- 2024 deletion phenotype: in the F13 colony-structure system, phd1Δ showed little effect on the presence of structured outer zones at day 5, unlike flo11Δ (structure abolished). (cromie2024spatiotemporalpatternsof pages 21-23)
Evidence summary table
The following table consolidates the supported claims, evidence types, and quantitative data extracted from the retrieved corpus.
| Claim/annotation | Evidence type | Key experimental details (strain/condition/assay) | Quantitative/statistical data | Primary source (authors year journal) | DOI URL |
|---|---|---|---|---|---|
| Phd1 is a transcription factor and a master regulator of filamentous differentiation in S. cerevisiae | Genetics; regulatory analysis | Primary study summarized Phd1 as regulating expression of most other differentiation TFs; overproduction induced filamentation; evidence integrated with nitrogen-starvation differentiation assays (raithatha2012cdk8regulatesstability pages 1-2, raithatha2012cdk8regulatesstability pages 2-2) | Overexpression sufficient to induce filamentation; no exact fold reported in gathered evidence (raithatha2012cdk8regulatesstability pages 1-2, raithatha2012cdk8regulatesstability pages 2-2) | Raithatha et al. 2012, Molecular and Cellular Biology (raithatha2012cdk8regulatesstability pages 1-2, raithatha2012cdk8regulatesstability pages 2-2) | https://doi.org/10.1128/MCB.05420-11 |
| Primary molecular function is transcriptional regulation of pseudohyphal/invasive growth programs rather than enzymatic catalysis | Genetics; phenotype suppression | Overexpression of PHD1 strongly promoted pseudohyphal growth even on nitrogen-rich medium and could suppress pseudohyphal defects of tpk2 and ste12 mutants (pan2000sok2regulatesyeast pages 1-2) | Strong pseudohyphal growth on rich medium upon overexpression; phd1/phd1 mutants alone showed no obvious pseudohyphal defect in that assay (pan2000sok2regulatesyeast pages 1-2) | Pan & Heitman 2000, Molecular and Cellular Biology (pan2000sok2regulatesyeast pages 1-2) | https://doi.org/10.1128/MCB.20.22.8364-8372.2000 |
| Phd1 positively regulates FLO11/MUC1, a key cell-surface flocculin required for filamentation/invasion | Genetics; transcriptional cascade | Genome-array/Northern analyses in sok2/sok2 hyperfilamentous mutants placed PHD1 in a cascade with Ash1 controlling FLO11 expression and cell-cell adhesion (pan2000sok2regulatesyeast pages 1-2); review synthesis places Phd1 in Sok2→Phd1/Ash1/Swi5→FLO11 regulation (kumar2021thecomplexgenetic pages 5-7) | In sok2 background, Phd1 and Ash1 were largely required for filamentation; exact expression fold not given in gathered excerpts (pan2000sok2regulatesyeast pages 1-2, kumar2021thecomplexgenetic pages 5-7) | Pan & Heitman 2000, MCB; Kumar 2021, Annual Review of Genetics (pan2000sok2regulatesyeast pages 1-2, kumar2021thecomplexgenetic pages 5-7) | https://doi.org/10.1128/MCB.20.22.8364-8372.2000; https://doi.org/10.1146/annurev-genet-071719-020249 |
| Phd1 functions in pseudohyphal growth and haploid invasive growth pathways | Genetics; phenotypic analysis | Ste12 and PHD1 were required for haploid invasive growth; PHD1 induction/stabilization tracked with nitrogen starvation, a canonical pseudohyphal trigger (raithatha2012cdk8regulatesstability pages 8-9, raithatha2012cdk8regulatesstability pages 1-2) | Nitrogen starvation increased PHD1 mRNA and stabilized protein; invasive-growth requirement shown genetically but without exact percentages in gathered text (raithatha2012cdk8regulatesstability pages 8-9, raithatha2012cdk8regulatesstability pages 1-2) | Raithatha et al. 2012, MCB (raithatha2012cdk8regulatesstability pages 8-9, raithatha2012cdk8regulatesstability pages 1-2) | https://doi.org/10.1128/MCB.05420-11 |
| Upstream pathway context includes Ras2/cAMP-PKA, MAPK/Kss1, and Snf1; Phd1 acts within or alongside this network | Review synthesis; genetics | Filamentous differentiation broadly regulated by MAPK (Kss1), Ras-cAMP-PKA, and Snf1-AMPK; Sok2 is thought to act downstream of cAMP/PKA, while the Sok2→Phd1/Ash1/Swi5 cascade can regulate FLO11 independently of direct PKA/MAPK control (raithatha2012cdk8regulatesstability pages 1-2, pan2000sok2regulatesyeast pages 1-2, kumar2021thecomplexgenetic pages 5-7) | Pathway placement is qualitative in gathered evidence; no direct kinetic constants reported (raithatha2012cdk8regulatesstability pages 1-2, pan2000sok2regulatesyeast pages 1-2, kumar2021thecomplexgenetic pages 5-7) | Raithatha et al. 2012, MCB; Pan & Heitman 2000, MCB; Kumar 2021, Annu Rev Genet (raithatha2012cdk8regulatesstability pages 1-2, pan2000sok2regulatesyeast pages 1-2, kumar2021thecomplexgenetic pages 5-7) | https://doi.org/10.1128/MCB.05420-11; https://doi.org/10.1128/MCB.20.22.8364-8372.2000; https://doi.org/10.1146/annurev-genet-071719-020249 |
| PHD1 is directly repressed by Sok2 in rich medium | Genetics; transcriptional regulation | In rich medium, Sok2 acted as a negative regulator of filamentation and directly/indirectly repressed the transcription-factor cascade including PHD1; PHD1 was induced in sok2/sok2 hyperfilamentous mutants (raithatha2012cdk8regulatesstability pages 8-9, pan2000sok2regulatesyeast pages 1-2) | PHD1 induction observed in sok2/sok2 mutants; exact fold change not given in gathered excerpts (raithatha2012cdk8regulatesstability pages 8-9, pan2000sok2regulatesyeast pages 1-2) | Pan & Heitman 2000, MCB; Raithatha et al. 2012, MCB (raithatha2012cdk8regulatesstability pages 8-9, pan2000sok2regulatesyeast pages 1-2) | https://doi.org/10.1128/MCB.20.22.8364-8372.2000; https://doi.org/10.1128/MCB.05420-11 |
| PHD1 expression is partially dependent on Ste12, and Ste12 binds the PHD1 promoter in vitro | Promoter binding; genetics | DNase I footprinting/promoter analysis showed Ste12 binds sites on the PHD1 promoter in vitro; in vivo promoter occupancy also implicated Tec1, Flo8, Sok2, and Phd1 itself (raithatha2012cdk8regulatesstability pages 8-9, raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability pages 2-3) | Qualitative promoter-binding evidence; no occupancy percentages in gathered excerpts (raithatha2012cdk8regulatesstability pages 8-9, raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability pages 2-3) | Raithatha et al. 2012, MCB (raithatha2012cdk8regulatesstability pages 8-9, raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability pages 2-3) | https://doi.org/10.1128/MCB.05420-11 |
| Phd1 positively autoregulates its own expression under nitrogen limitation | Promoter binding; regulatory model | Promoter occupancy and regulatory model indicate Phd1 can bind/activate its own promoter, forming a positive-feedback loop during differentiation (raithatha2012cdk8regulatesstability pages 8-9, raithatha2012cdk8regulatesstability pages 6-7) | Positive feedback described qualitatively; no exact fold increase reported in gathered excerpts (raithatha2012cdk8regulatesstability pages 8-9, raithatha2012cdk8regulatesstability pages 6-7) | Raithatha et al. 2012, MCB (raithatha2012cdk8regulatesstability pages 8-9, raithatha2012cdk8regulatesstability pages 6-7) | https://doi.org/10.1128/MCB.05420-11 |
| Cdk8 negatively regulates Phd1 by phosphorylation-dependent destabilization | Protein stability assay; in vitro kinase/phosphopeptide analysis | Cycloheximide-chase, 32Pi metabolic labeling, immunoprecipitation, in vitro kinase assays, and phosphopeptide analysis showed Cdk8-dependent phosphorylation promotes Phd1 degradation (raithatha2012cdk8regulatesstability pages 1-2, raithatha2012cdk8regulatesstability pages 2-2, raithatha2012cdk8regulatesstability pages 2-3) | Phd1 half-life was reported as ~10–15 min in wild type/rich medium and >45 min in cdk8Δ cells (raithatha2012cdk8regulatesstability pages 1-2, raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability pages 2-3) | Raithatha et al. 2012, MCB (raithatha2012cdk8regulatesstability pages 1-2, raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability pages 2-3) | https://doi.org/10.1128/MCB.05420-11 |
| Natural polymorphism at the Cdk8 phosphosite stabilizes Phd1 and enhances filamentation | Protein stability assay; allele comparison | Comparison of PHD1 alleles (S92 versus S92F) showed that the 1278b strain carries S92F, removing a Cdk8 phosphorylation site and stabilizing Phd1 (raithatha2012cdk8regulatesstability pages 1-2, raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability pages 2-2) | Wild-type S92 Phd1 half-life ~15 min in rich medium; stabilized variant and/or cdk8Δ backgrounds showed >45 min stability (raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability pages 2-2) | Raithatha et al. 2012, MCB (raithatha2012cdk8regulatesstability pages 1-2, raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability pages 2-2) | https://doi.org/10.1128/MCB.05420-11 |
| Nitrogen starvation stabilizes Phd1 protein and increases PHD1 transcript, promoting differentiation | Protein stability assay; transcript analysis | Cells shifted to SLAD/nitrogen-limiting medium showed increased PHD1 mRNA and marked stabilization of Phd1 protein; model proposes loss of Cdk8 activity under these conditions (raithatha2012cdk8regulatesstability pages 8-9, raithatha2012cdk8regulatesstability pages 1-2, raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability media d01b8851) | Half-life increased from ~15 min in SD to ~40 min after 2 h in SLAD and >45 min after 4 h in SLAD (raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability media d01b8851) | Raithatha et al. 2012, MCB (raithatha2012cdk8regulatesstability pages 8-9, raithatha2012cdk8regulatesstability pages 1-2, raithatha2012cdk8regulatesstability pages 6-7, raithatha2012cdk8regulatesstability media d01b8851) | https://doi.org/10.1128/MCB.05420-11 |
| Forced expression of PHD1 is sufficient to induce pseudohyphal growth synthetically, even in rich media and in haploids | Synthetic induction | Synthetic gene-regulation circuits controlling native PHD1 and FLO8 expression triggered pseudohyphal growth in diploid and haploid cells in rich media; used as an engineered, externally inducible differentiation system (pothoulakis2018syntheticgeneregulation pages 1-2) | Phenotype induction in both diploid and haploid strains under non-starvation conditions; exact percentages not given in gathered excerpt (pothoulakis2018syntheticgeneregulation pages 1-2) | Pothoulakis & Ellis 2018, Communications Biology (pothoulakis2018syntheticgeneregulation pages 1-2) | https://doi.org/10.1038/s42003-017-0008-0 |
| Recent 2024 colony-morphology data support association of PHD1 expression with structured colony regions, but deletion may be background-dependent and nonessential in F13 | RNA-seq/cluster; deletion phenotype | In structured-colony strain F13, PHD1/YKL043W clustered with genes whose expression correlated positively with colony structure (cluster I9); however, phd1Δ retained central smooth plus outer structured zones at day 5, unlike flo11Δ or msb2Δ (cromie2024spatiotemporalpatternsof pages 21-23) | phd1Δ had little effect on F13 morphology, whereas flo11Δ and msb2Δ remained fully smooth at day 5 (cromie2024spatiotemporalpatternsof pages 21-23) | Cromie et al. 2024, PLOS ONE (cromie2024spatiotemporalpatternsof pages 21-23) | https://doi.org/10.1371/journal.pone.0311061 |
| Subcellular localization is expected to be nuclear for a transcription factor, but no direct Phd1 localization evidence was identified in the gathered sources | Evidence gap / inference | Gathered primary and review sources discuss Phd1 as a transcription factor regulating promoters and differentiation programs, but none of the retrieved evidence IDs reported direct GFP/immunolocalization for Phd1 (raithatha2012cdk8regulatesstability pages 8-9, pan2000sok2regulatesyeast pages 1-2) | No direct localization measurement available in gathered evidence (raithatha2012cdk8regulatesstability pages 8-9, pan2000sok2regulatesyeast pages 1-2) | Evidence gap within retrieved literature (raithatha2012cdk8regulatesstability pages 8-9, pan2000sok2regulatesyeast pages 1-2) | N/A |
Table: This table summarizes experimentally supported functional annotations for Saccharomyces cerevisiae PHD1/Phd1 using only the gathered evidence contexts. It highlights molecular function, pathway placement, regulatory inputs, phenotypes, and the current evidence gap for direct localization data.
References (with publication dates and URLs)
- Pan X, Heitman J. 2000-11. Sok2 regulates yeast pseudohyphal differentiation via a transcription factor cascade that regulates cell-cell adhesion. Molecular and Cellular Biology. https://doi.org/10.1128/MCB.20.22.8364-8372.2000 (pan2000sok2regulatesyeast pages 1-2)
- Raithatha S, Su T-C, Lourenco P, Goto S, Sadowski I. 2012-02. Cdk8 regulates stability of the transcription factor Phd1 to control pseudohyphal differentiation of Saccharomyces cerevisiae. Molecular and Cellular Biology. https://doi.org/10.1128/MCB.05420-11 (raithatha2012cdk8regulatesstability pages 1-2)
- Pothoulakis G, Ellis T. 2018-01. Synthetic gene regulation for independent external induction of the Saccharomyces cerevisiae pseudohyphal growth phenotype. Communications Biology. https://doi.org/10.1038/s42003-017-0008-0 (pothoulakis2018syntheticgeneregulation pages 1-2)
- Kumar A. 2021-11. The Complex Genetic Basis and Multilayered Regulatory Control of Yeast Pseudohyphal Growth. Annual Review of Genetics. https://doi.org/10.1146/annurev-genet-071719-020249 (kumar2021thecomplexgenetic pages 5-7)
- Cromie GA, Tan Z, Hays M, Sirr A, Dudley AM. 2024-09. Spatiotemporal patterns of gene expression during development of a complex colony morphology. PLOS ONE. https://doi.org/10.1371/journal.pone.0311061 (cromie2024spatiotemporalpatternsof pages 21-23)
Limitations of this report (evidence gaps)
- Direct subcellular localization of Phd1 (e.g., microscopy of Phd1-GFP or nuclear fractionation) was not identified in the retrieved text evidence, so nuclear localization is stated as inferred rather than directly demonstrated here. (raithatha2012cdk8regulatesstability pages 1-2)
- While the prompt requested prioritization of 2023–2024 sources, tool-based retrieval yielded limited Phd1-specific mechanistic literature in those years; the 2024 colony-morphology study provides the most direct recent primary evidence involving PHD1. (cromie2024spatiotemporalpatternsof pages 21-23)
References
-
(raithatha2012cdk8regulatesstability pages 1-2): Sheetal Raithatha, Ting-Cheng Su, Pedro Lourenco, Susan Goto, and Ivan Sadowski. Cdk8 regulates stability of the transcription factor phd1 to control pseudohyphal differentiation of saccharomyces cerevisiae. Molecular and Cellular Biology, 32:664-674, Feb 2012. URL: https://doi.org/10.1128/mcb.05420-11, doi:10.1128/mcb.05420-11. This article has 59 citations and is from a domain leading peer-reviewed journal.
-
(pan2000sok2regulatesyeast pages 1-2): Xuewen Pan and Joseph Heitman. Sok2 regulates yeast pseudohyphal differentiation via a transcription factor cascade that regulates cell-cell adhesion. Molecular and Cellular Biology, 20:8364-8372, Nov 2000. URL: https://doi.org/10.1128/mcb.20.22.8364-8372.2000, doi:10.1128/mcb.20.22.8364-8372.2000. This article has 164 citations and is from a domain leading peer-reviewed journal.
-
(kumar2021thecomplexgenetic pages 5-7): Anuj Kumar. The complex genetic basis and multilayered regulatory control of yeast pseudohyphal growth. Nov 2021. URL: https://doi.org/10.1146/annurev-genet-071719-020249, doi:10.1146/annurev-genet-071719-020249. This article has 41 citations and is from a domain leading peer-reviewed journal.
-
(raithatha2012cdk8regulatesstability pages 6-7): Sheetal Raithatha, Ting-Cheng Su, Pedro Lourenco, Susan Goto, and Ivan Sadowski. Cdk8 regulates stability of the transcription factor phd1 to control pseudohyphal differentiation of saccharomyces cerevisiae. Molecular and Cellular Biology, 32:664-674, Feb 2012. URL: https://doi.org/10.1128/mcb.05420-11, doi:10.1128/mcb.05420-11. This article has 59 citations and is from a domain leading peer-reviewed journal.
-
(raithatha2012cdk8regulatesstability media d01b8851): Sheetal Raithatha, Ting-Cheng Su, Pedro Lourenco, Susan Goto, and Ivan Sadowski. Cdk8 regulates stability of the transcription factor phd1 to control pseudohyphal differentiation of saccharomyces cerevisiae. Molecular and Cellular Biology, 32:664-674, Feb 2012. URL: https://doi.org/10.1128/mcb.05420-11, doi:10.1128/mcb.05420-11. This article has 59 citations and is from a domain leading peer-reviewed journal.
-
(raithatha2012cdk8regulatesstability pages 2-3): Sheetal Raithatha, Ting-Cheng Su, Pedro Lourenco, Susan Goto, and Ivan Sadowski. Cdk8 regulates stability of the transcription factor phd1 to control pseudohyphal differentiation of saccharomyces cerevisiae. Molecular and Cellular Biology, 32:664-674, Feb 2012. URL: https://doi.org/10.1128/mcb.05420-11, doi:10.1128/mcb.05420-11. This article has 59 citations and is from a domain leading peer-reviewed journal.
-
(raithatha2012cdk8regulatesstability media c487618b): Sheetal Raithatha, Ting-Cheng Su, Pedro Lourenco, Susan Goto, and Ivan Sadowski. Cdk8 regulates stability of the transcription factor phd1 to control pseudohyphal differentiation of saccharomyces cerevisiae. Molecular and Cellular Biology, 32:664-674, Feb 2012. URL: https://doi.org/10.1128/mcb.05420-11, doi:10.1128/mcb.05420-11. This article has 59 citations and is from a domain leading peer-reviewed journal.
-
(raithatha2012cdk8regulatesstability pages 3-4): Sheetal Raithatha, Ting-Cheng Su, Pedro Lourenco, Susan Goto, and Ivan Sadowski. Cdk8 regulates stability of the transcription factor phd1 to control pseudohyphal differentiation of saccharomyces cerevisiae. Molecular and Cellular Biology, 32:664-674, Feb 2012. URL: https://doi.org/10.1128/mcb.05420-11, doi:10.1128/mcb.05420-11. This article has 59 citations and is from a domain leading peer-reviewed journal.
-
(cromie2024spatiotemporalpatternsof pages 21-23): Gareth A. Cromie, Zhihao Tan, Michelle Hays, Amy Sirr, and Aimée M. Dudley. Spatiotemporal patterns of gene expression during development of a complex colony morphology. PLOS ONE, Sep 2024. URL: https://doi.org/10.1371/journal.pone.0311061, doi:10.1371/journal.pone.0311061. This article has 3 citations and is from a peer-reviewed journal.
-
(pothoulakis2018syntheticgeneregulation pages 1-2): Georgios Pothoulakis and Tom Ellis. Synthetic gene regulation for independent external induction of the saccharomyces cerevisiae pseudohyphal growth phenotype. Communications Biology, Jan 2018. URL: https://doi.org/10.1038/s42003-017-0008-0, doi:10.1038/s42003-017-0008-0. This article has 19 citations and is from a peer-reviewed journal.
-
(raithatha2012cdk8regulatesstability pages 8-9): Sheetal Raithatha, Ting-Cheng Su, Pedro Lourenco, Susan Goto, and Ivan Sadowski. Cdk8 regulates stability of the transcription factor phd1 to control pseudohyphal differentiation of saccharomyces cerevisiae. Molecular and Cellular Biology, 32:664-674, Feb 2012. URL: https://doi.org/10.1128/mcb.05420-11, doi:10.1128/mcb.05420-11. This article has 59 citations and is from a domain leading peer-reviewed journal.
-
(raithatha2012cdk8regulatesstability pages 2-2): Sheetal Raithatha, Ting-Cheng Su, Pedro Lourenco, Susan Goto, and Ivan Sadowski. Cdk8 regulates stability of the transcription factor phd1 to control pseudohyphal differentiation of saccharomyces cerevisiae. Molecular and Cellular Biology, 32:664-674, Feb 2012. URL: https://doi.org/10.1128/mcb.05420-11, doi:10.1128/mcb.05420-11. This article has 59 citations and is from a domain leading peer-reviewed journal.
Artifacts
Citations
- kumar2021thecomplexgenetic pages 5-7
- cromie2024spatiotemporalpatternsof pages 21-23
- pothoulakis2018syntheticgeneregulation pages 1-2
- https://doi.org/10.1128/MCB.20.22.8364-8372.2000
- https://doi.org/10.1146/annurev-genet-071719-020249
- https://doi.org/10.1128/MCB.05420-11
- https://doi.org/10.1371/journal.pone.0311061
- https://doi.org/10.1038/s42003-017-0008-0
- https://doi.org/10.1128/MCB.20.22.8364-8372.2000;
- https://doi.org/10.1128/MCB.05420-11;
- https://doi.org/10.1128/mcb.05420-11,
- https://doi.org/10.1128/mcb.20.22.8364-8372.2000,
- https://doi.org/10.1146/annurev-genet-071719-020249,
- https://doi.org/10.1371/journal.pone.0311061,
- https://doi.org/10.1038/s42003-017-0008-0,
Perplexity
(PHD1-deep-research-perplexity.md)
Perplexity
(PHD1-deep-research-perplexity.md)PHD1: A Master Transcriptional Regulator of Pseudohyphal Growth and Morphogenesis in Saccharomyces cerevisiae
Executive Summary
The PHD1 gene (YKL043W, UniProt: P36093) in Saccharomyces cerevisiae encodes a putative transcription factor that belongs to the APSES family of fungal-specific transcriptional regulators, characterized by a conserved basic helix-loop-helix (bHLH) DNA-binding domain[4][10]. As a master regulator of developmental processes, Phd1 functions as a transcriptional activator that dramatically enhances pseudohyphal growth when overexpressed, even under nutrient-rich conditions[1][11], and serves as a key regulatory hub that coordinates complex morphogenetic signaling networks in response to nitrogen starvation[11][38]. The protein physically interacts with the Tup1-Cyc8 complex and recruits this corepressor complex to its target genes, demonstrating a sophisticated mechanism of transcriptional regulation[7][17]. Phd1 is an inherently unstable protein whose degradation is initiated through phosphorylation by Cdk8 of the RNA polymerase II mediator complex, with stabilization of the protein occurring during nutrient limitation to initiate differentiation[20][23]. Through its regulation of extensive transcriptional networks involving over 200 genes and its ability to induce filamentous growth independently of major signaling cascades, PHD1 represents a crucial control point for fungal development and cellular differentiation.
Protein Structure and Molecular Architecture
Basic Helix-Loop-Helix Domain Architecture
Phd1 belongs to the basic helix-loop-helix (bHLH) transcription factor family, a large and diverse class of dimerizing transcription factors found throughout the eukaryotic kingdom[39][42]. The bHLH structural motif consists of two α-helices connected by a loop, with one helix containing basic amino acid residues that facilitate DNA binding to consensus sequences[39][42]. The conserved bHLH domain functions as the core DNA-binding module, allowing Phd1 to recognize and bind specific DNA sequences at target gene promoters. As a member of the APSES subfamily of bHLH proteins, Phd1 shares a highly conserved DNA-binding domain approximately 100 residues in length with other APSES family members including Efg1, Efh1, and Sok2 in various fungal species[3][9]. The APSES domain nomenclature derives from its presence in four characterized regulatory proteins: Asm1 (from Neurospora crassa), Phd1 (from Saccharomyces cerevisiae), Sok2 (from Saccharomyces cerevisiae), and Efg1 and Stuа (from Candida species)[3].
The basic region of the Phd1 protein contains positively charged residues that interact with the major groove of target DNA sequences, while the helix-loop-helix motif mediates protein-protein interactions necessary for dimerization[39][42]. Studies using protein binding microarrays and crystallographic analysis have demonstrated that bHLH proteins typically recognize E-box consensus sequences with the canonical form CACGTG (palindromic) or related variants[25][39]. For Phd1 specifically, structural analysis indicates binding to E-box motifs, consistent with its classification as a member of the bHLH family, though the precise recognition sequences may show variation depending on the specific target promoter and cofactors involved in complex formation[25].
APSES Domain and Phylogenetic Classification
The APSES domain itself comprises the entire DNA-binding region and contains the bHLH subdomain with characteristic structural features[3][9][28]. Phylogenetic analysis of APSES proteins across fungal species has classified them into four major groups (A, B, C, and D) based on sequence relatedness of the APSES domain[28]. Phd1 falls within the Group C-I classification in hemiascomycetes like Saccharomyces and Candida species, along with Efg1, Efh1, and Sok2[28]. This classification indicates evolutionary conservation and functional relationships among these proteins in regulating filamentous versus yeast growth. The presence of the APSES domain represents a distinctive feature of fungal transcription factors, as this domain is specific to fungi and not found in other eukaryotic transcription factors, suggesting that APSES proteins arose through specialized evolution to regulate uniquely fungal developmental processes[28].
Post-Translational Modification Sites
Phd1 contains identifiable phosphorylation sites that modulate its activity and stability. Analysis of the protein sequence indicates potential phosphorylation targets for protein kinase A (PKA) and other kinases[3], and experimental evidence demonstrates that Cdk8, a kinase component of the RNA polymerase II mediator complex, phosphorylates Phd1 to target it for degradation[20][23]. The naturally filamenting Σ1278b strain of Saccharomyces cerevisiae possesses a sequence polymorphism that eliminates one Cdk8 phosphorylation site, thereby stabilizing the Phd1 protein and contributing to enhanced differentiation in this strain background[20][23]. This finding highlights how relatively subtle sequence variations can have profound effects on protein stability and phenotypic outcomes related to fungal development.
Role in Pseudohyphal Growth and Morphogenesis
Induction of Pseudohyphal Development
Phd1 serves as a powerful inducer of pseudohyphal growth, the filamentous growth form adopted by Saccharomyces cerevisiae diploid cells in response to nitrogen starvation[1][11]. Overexpression of the PHD1 gene dramatically enhances pseudohyphal growth even under nutrient-rich medium conditions, producing long chains of elongated cells characteristic of the filamentous state[1][11]. This capacity for ectopic induction of filamentous growth makes Phd1 a key node in the regulatory network controlling developmental transitions. Phenotypically, strains overexpressing PHD1 exhibit enhanced agar invasion, the ability to penetrate growth substrates, and the formation of pseudohyphal cell chains even when grown in high nitrogen liquid media[11][38]. These observations demonstrate that Phd1 protein levels represent a sufficient and, in many contexts, necessary condition for driving the pseudohyphal developmental program.
Comparison with Regulatory Counterpart Sok2
The regulatory specificity of Phd1 becomes particularly evident when compared with Sok2, a closely related APSES family member that functions as a repressor of pseudohyphal differentiation[9][21]. While Phd1 activates pseudohyphal growth, Sok2 represses this developmental program, and these proteins exhibit opposing regulatory functions despite sharing substantial sequence similarity and the conserved APSES domain[9][21]. The finding that overexpression of PHD1 in Saccharomyces cerevisiae produces effects equivalent to deletion of SOK2 further supports this antagonistic relationship[9]. This functional divergence between closely related transcription factors demonstrates that small differences in protein sequence or domain organization can result in opposite regulatory outcomes. The observation that Sok2 normally represses expression of the PHD1 gene itself establishes a regulatory circuit where Sok2 inhibits filamentation by suppressing expression of its transcriptional antagonist[21]. Through deletion of SOK2, the PHD1 gene becomes upregulated, enhancing the filamentation pathway and accelerating pseudohyphal growth to levels comparable to PHD1 overexpression[21].
Morphological Consequences and Cell Shape Regulation
Beyond effects on filament formation itself, Phd1 directly influences cell morphology and shape during pseudohyphal development[21]. Overexpression of PHD1 promotes cell elongation, producing the characteristic elongated cell morphology of pseudohyphae, whereas deletion of PHD1 impairs cell elongation[21]. This suggests that Phd1 regulates not only genes required for filament formation and cell-cell adhesion but also genes controlling cell cycle progression and cell shape determination. The ability of Phd1 to independently regulate cell elongation independently of its effects on filament formation indicates that this transcription factor coordinates multiple aspects of morphogenesis through its diverse target gene repertoire.
Transcriptional Regulatory Networks and Target Gene Identification
FLO11 as a Critical Downstream Target
The FLO11 gene represents one of the most important direct targets of Phd1 regulation[11][38][41]. FLO11 encodes a cell wall protein critical for cell-cell adhesion and agar invasion, and its expression is absolutely required for both pseudohyphae formation and invasive growth in Saccharomyces cerevisiae[41]. The promoter of FLO11 contains a consensus binding sequence for Ste12 and Tec1, and genome-wide chromatin immunoprecipitation studies have demonstrated that Phd1 also binds directly to the FLO11 promoter region[11][38]. Interestingly, Phd1 can induce filamentation through both FLO11-dependent and FLO11-independent mechanisms[13], indicating that this transcription factor regulates multiple distinct pathways controlling filamentous growth. When FLO11 is deleted, overexpression of PHD1 can still restore pseudohyphal growth (though not invasive growth), demonstrating that Phd1 has additional targets beyond FLO11 that contribute to the filamentous phenotype[21]. This functional redundancy provides robustness to the developmental program and allows Phd1 to coordinate complex morphogenetic responses through multiple parallel regulatory pathways.
Regulatory Hub Function and Master Regulator Status
Comprehensive chromatin immunoprecipitation studies combined with DNA microarray analysis (ChIP-chip) have mapped the global binding targets of Phd1 across the yeast genome[11][29][38]. These studies identified Phd1 as a major regulatory hub in the pseudohyphal growth network, with the protein binding to the promoters of numerous genes involved in filament formation and development[11][38]. A particularly striking finding is that the promoter of PHD1 itself is bound by all six major transcription factors involved in pseudohyphal growth regulation (Ste12, Tec1, Sok2, Phd1, Flo8, and Mga1), identifying PHD1 as a key target hub receiving input from multiple regulatory pathways[29][38]. This architecture suggests that PHD1 serves as an integrator of diverse signals controlling filamentous development.
When subjected to expression microarray analysis following overexpression, Phd1 induces approximately 214 genes with at least a twofold change in expression[11][38]. These overexpression-regulated genes show high correlation with genes regulated during normal pseudohyphal growth in response to nitrogen starvation, indicating that Phd1 activation triggers a transcriptional program largely equivalent to the natural developmental response[11][38]. Remarkably, overexpression of Phd1 induces 20-69% of the genes normally regulated during pseudohyphal growth (depending on scoring methodology), demonstrating that this single transcription factor is sufficient to initiate a substantial portion of the developmental transcriptional cascade[38]. The enrichment analysis of genes regulated by Phd1 overexpression revealed major functional categories associated with metabolism, particularly carboxylic acid metabolism, consistent with the metabolic remodeling that accompanies fungal development[38].
Other Transcriptional Targets and Regulatory Cascade
Beyond FLO11, Phd1 regulates expression of numerous other transcription factors involved in filamentous growth, including the genes encoding Tec1, Eed1, and Crz1[3]. This regulatory architecture creates a transcriptional cascade in which Phd1 activates expression of additional transcription factors that, in turn, regulate downstream genes required for the morphogenetic transition. The binding of Phd1 to intergenic regions often spans large distances, with some promoter regions showing Phd1 binding extending over multiple kilobases and containing multiple peaks of occupancy[6], suggesting complex and multimodal interactions with target promoters. This extensive binding pattern indicates that Phd1 may regulate target genes through multiple independent regulatory elements or through cooperative interactions with other transcription factors.
Regulatory Signaling Pathways and Integration of Environmental Signals
cAMP-Protein Kinase A Pathway
The cAMP-dependent protein kinase A (PKA) pathway represents a major regulatory input controlling pseudohyphal growth, and evidence indicates that this pathway regulates multiple components of the pseudohyphal transcriptional network including Phd1[8][15][16]. In Saccharomyces cerevisiae, PKA comprises a single regulatory subunit (Bcy1) and three catalytic subunits (Tpk1, Tpk2, and Tpk3)[15][16]. The Tpk2 catalytic subunit plays a unique positive role in pseudohyphal differentiation and activates filamentous growth through phosphorylation of multiple downstream targets, including transcription factors such as Flo8, Sfl1, and Phd1[8][16]. While genetic evidence suggests that Phd1 functions downstream of PKA signaling[21], the mechanisms by which PKA-dependent phosphorylation modulates Phd1 activity remain partially characterized. The upregulation of PHD1 expression via the Tpk1 branch of the cAMP-PKA pathway during glucose starvation provides evidence that PKA signaling controls Phd1 at the transcriptional level[8].
Mitogen-Activated Protein Kinase Cascade
The Kss1/Fus3 mitogen-activated protein kinase (MAPK) cascade represents another major signaling pathway controlling filamentous growth[35][41][43]. This pathway, also known as the pheromone response pathway, involves the kinases Ste11, Ste7, and Kss1, along with the transcription factors Ste12 and Tec1[35][41]. While genetic interactions have been documented between elements of this MAPK cascade and PHD1, the exact relationship differs from the PKA pathway. Specifically, genetic interactions with elements of the MAPK cascade support an activator function of Phd1, but overexpression of PHD1 can suppress some filamentation defects even in MAPK pathway mutants[21]. This suggests that Phd1 functions in both MAPK-dependent and MAPK-independent regulatory networks, allowing it to coordinate filamentous development through multiple parallel signaling routes.
Nutrient Sensing and Nitrogen Starvation Response
Nitrogen starvation represents the primary environmental trigger for pseudohyphal differentiation in diploid yeast, and multiple studies have examined how the cell senses nitrogen limitation and activates developmental responses[37][40][43]. The GPA2 gene encodes a heterotrimeric G protein alpha subunit that plays a critical role in nitrogen sensing and regulation of pseudohyphal growth[43]. Deletion of GPA2 results in defective pseudohyphal growth, while a constitutively active allele of GPA2 stimulates filamentation even on nitrogen-rich media, demonstrating the importance of this signaling component for detecting nutrient limitation[43]. The mechanisms by which GPA2 signaling regulates Phd1 involve modulation of cAMP levels, connecting nutrient sensing to the PKA pathway[43].
The unfolded protein response (UPR) pathway has emerged as an unexpected regulator of pseudohyphal growth in response to nitrogen starvation[37]. The UPR is normally activated when accumulation of unfolded proteins in the endoplasmic reticulum triggers the endonuclease activity of Ire1, leading to splicing of the HAC1 mRNA and production of the transcription factor Hac1p[37]. In nitrogen-rich environments characterized by high translation rates, the UPR is active and Hac1p functions to repress both pseudohyphal growth and meiosis[37]. During nitrogen starvation, translation rates decline and the UPR becomes inactive, allowing pseudohyphal development and meiosis to proceed[37]. This mechanism provides a link between protein synthesis capacity and developmental decision-making, ensuring that the energetically expensive process of filamentation only occurs when nutrient limitation imposes constraints on protein production.
Protein Stability Regulation and Post-Translational Control
Cdk8-Dependent Phosphorylation and Degradation
One of the most significant recent findings regarding Phd1 function concerns its regulation by the Cdk8 kinase component of the RNA polymerase II mediator complex[20][23]. Cdk8 is part of the mediator kinase module, which associates with the main mediator complex and influences transcription of multiple genes. Research has demonstrated that Phd1 is an inherently unstable protein, with degradation initiated through phosphorylation by Cdk8[20][23]. The naturally filamenting Σ1278b laboratory strain of Saccharomyces cerevisiae contains a sequence polymorphism in the PHD1 gene that eliminates a Cdk8 phosphorylation site, resulting in stabilization of the Phd1 protein and contributing to the enhanced pseudohyphal growth characteristic of this strain[20][23].
During nitrogen starvation, the cAMP-PKA pathway is activated, and this pathway responds to nutrient limitation by reducing Cdk8 activity[20][23]. Inhibition of Cdk8 activity during nutrient starvation results in stabilization of multiple transcription factors including Phd1, allowing their accumulation and activation of genes necessary for the filamentous response[20][23]. In contrast, under nutrient-rich conditions when Cdk8 activity is high, Phd1 is phosphorylated and rapidly degraded, preventing inappropriate activation of filamentous growth genes. This mechanism provides an elegant nutrient-sensing system where kinase activity changes in response to metabolic state directly control the stability of key developmental regulators.
Interaction with the Tup1-Cyc8 Corepressor Complex
Phd1 physically interacts with the Tup1-Cyc8 complex, a pair of proteins that function as global corepressors of gene expression[7][17]. This interaction allows Phd1 to recruit Tup1 to its target promoters, modulating transcriptional activity[7]. The Cyc8-Tup1 complex is recruited to multiple gene promoters controlled by Phd1 and functions to establish or maintain organized chromatin structures that regulate gene expression[14]. The complex interacts with histone deacetylases including Hda1 and Rpd3, which coordinate histone deacetylation and nucleosome stabilization[14]. At genes like FLO1, the Cyc8-Tup1 complex occupies a DNase I hypersensitive site within an ordered array of strongly positioned nucleosomes, and deletion of cyc8 results in histone hyperacetylation and gene activation[14]. This indicates that Phd1-mediated recruitment of Tup1-Cyc8 may function to either activate or repress target genes depending on the promoter context and other bound regulatory factors.
Position within Broader Regulatory Networks
Integration with Other Transcription Factors
Phd1 functions as part of a larger transcriptional regulatory network controlling pseudohyphal growth, and its activity is coordinated with that of multiple other transcription factors[11][29][38]. The transcription factors Ste12, Tec1, Sok2, Flo8, and Mga1 all interact with Phd1-regulated promoters and contribute to control of downstream genes[11][29][38]. Chromatin immunoprecipitation studies have revealed extensive combinatorial binding, with large numbers of promoter regions bound by multiple factors simultaneously[11][29][38]. Particularly striking is the finding that 20 intergenic regions, including those upstream of genes such as FLO11, PHD1, MGA1, and HMS1, are bound by all six major regulatory factors[29][38]. This suggests that the transcriptional control of these key genes involves integration of inputs from multiple signal transduction pathways.
Phd1 and the closely related Mga1 protein represent the two most highly connected regulatory hubs in the pseudohyphal growth network[29][38]. These two proteins have the most extensive interactions with other transcription factors and bind to a large number of target promoters. The promoters of both PHD1 and MGA1 are bound by all six major pseudohyphal regulatory factors (Ste12, Tec1, Sok2, Phd1, Flo8, and Mga1), identifying them as key integration points where multiple regulatory signals converge[29][38]. The extraordinary redundancy in binding at these promoters suggests that Phd1 and Mga1 are positioned as master regulators that integrate multiple distinct signaling pathways to control the overall decision to undergo filamentous growth.
Feedback Regulation and Self-Regulation
In addition to regulating numerous downstream genes, Phd1 exhibits feedback regulation through binding to its own promoter[6]. The binding of transcription factors to their own promoters provides a mechanism for positive or negative feedback control of gene expression and can contribute to the establishment of stable expression states or the creation of bistable switches[6]. Efg1, the Candida homolog of Phd1, also binds to its own promoter and negatively regulates its own expression[3], suggesting that self-regulation of expression may be a conserved feature of APSES family proteins.
Evolutionary Conservation and Cross-Kingdom Homology
APSES Family Conservation Across Fungi
The APSES family of transcription factors shows remarkable conservation across diverse fungal species, indicating ancient evolutionary origins and fundamental importance for fungal development[28]. In the yeast Saccharomyces cerevisiae, the APSES proteins include Phd1, Sok2, Efg1, and Efh1[28]. In Candida albicans, a pathogenic fungus closely related to Saccharomyces, the major APSES proteins are Efg1, Efh1, Phd1, and Sok2[3][6]. In filamentous fungi such as Aspergillus nidulans and Neurospora crassa, the APSES proteins include StuA and Asm1[28]. The high degree of sequence conservation within the APSES domain across these diverse species suggests that this domain has been under strong purifying selection and that the core regulatory functions are likely conserved[28].
The fact that Efg1 from Candida albicans exhibits both repressor and activator functions when expressed heterologously in Saccharomyces cerevisiae, in contrast to the primarily activator function of Phd1, suggests that sequence divergence between family members leads to subtle differences in regulatory specificity[9]. The conserved APSES domain does not determine regulatory specificity, as proteins with nearly identical DNA-binding domains can have opposite effects on pseudohyphal growth[9]. This implies that regulatory specificity arises from differences in additional domains, cofactor interactions, or position-dependent effects in the nucleotide sequence.
Mechanisms of Functional Divergence
The existence of multiple APSES proteins with opposed regulatory functions in a single organism raises important questions about mechanisms of functional divergence[9]. Despite sharing a conserved bHLH motif, Phd1 and Sok2 have evolved to activate and repress pseudohyphal growth, respectively. The mechanisms underlying this functional divergence likely involve differences in protein-protein interactions, modifications of DNA-binding specificity, and recruitment of distinct corepressor or coactivator complexes. The finding that these proteins show genetic interactions with MAPK pathway components provides evidence that regulatory specificity involves differential connection to upstream signaling pathways[9].
Cellular Localization and Subcellular Function
Nuclear Localization and Nuclear Import
As a transcription factor, Phd1 functions in the nucleus where it binds to DNA and activates or represses target genes[7][10]. The presence of basic amino acid residues in the DNA-binding domain is characteristic of nuclear proteins, and the basic domain serves dual roles in facilitating both nuclear accumulation and sequence-specific DNA binding[39][42]. While the specific nuclear import machinery used by Phd1 has not been extensively characterized, the presence of conserved basic residues suggests that Phd1 likely utilizes the importin-α/β pathway for nuclear import, which recognizes classical nuclear localization signals (NLS) typically composed of clusters of basic amino acid residues.
Temporal Dynamics of Nuclear Accumulation
For the related APSES protein Efg1 in Candida albicans, detailed analysis of subcellular localization during developmental transitions has shown that Efg1 is present in the nuclei of yeast cells under normal growth conditions and also upon hyphal induction[3]. Interestingly, following hyphal induction, both EFG1 expression and nuclear levels of Efg1 protein drop dramatically, suggesting that Efg1 may be important for the initial transition to hyphae but not for continued hyphal growth in serum or Spider media[3]. Furthermore, overexpression of EFG1 in the presence of serum prevents the formation of true hyphae and instead produces yeast and pseudohyphae cells, indicating that precise modulation of Efg1 levels is critical for proper morphogenesis[3]. These observations suggest that transient, elevated levels of Phd1/Efg1 are required to initiate the developmental transition, while sustained high levels of expression may actually prevent continued development. This implies that the regulatory networks controlling APSES protein expression levels themselves represent important control points for fungal morphogenesis.
Comparison with Related Fungal APSES Proteins
Efg1 in Candida albicans and Pathogenic Fungi
Efg1 in Candida albicans, the most extensively characterized APSES protein in pathogenic fungi, shares approximately 65% amino acid identity with Phd1 within the conserved APSES domain[3]. Despite this high degree of sequence conservation, Efg1 regulates different developmental processes than Phd1 in Candida, controlling the yeast-to-hyphal transition, generation of chlamydospores, and white-opaque switching in addition to aspects of filamentous growth[3][9]. The observation that Efg1 exhibits both activator and repressor functions when heterologously expressed in Saccharomyces cerevisiae, whereas Phd1 primarily functions as an activator, indicates that regulatory specificity has diverged between these proteins[9].
Efg1 also regulates different aspects of cell biology beyond morphogenesis, including cell wall synthesis, metabolism, and cellular differentiation[3]. Genes regulated by Efg1 include those involved in cell wall synthesis, metabolism, and cellular differentiation, and Efg1 functions downstream of protein kinase A in yeast-hyphal transitions[3]. In contrast, EFG1 expression levels demonstrate considerable cell-to-cell variability in standard growth media, reflecting heterogeneity in the host population[3]. This variability in expression levels across individual cells may contribute to phenotypic heterogeneity and the ability of Candida populations to maintain multiple cellular states simultaneously.
WdStuA in Wangiella dermatitidis
The APSES protein WdStuA from the polymorphic fungal pathogen Wangiella dermatitidis shows structural and functional conservation with Phd1[12][32]. The WdSTUA gene encodes a protein of 627 amino acids with an APSES DNA-binding domain consisting of residues 157-234[12]. Analysis of the ORF showed that this protein shares significant identity with APSES transcription factors from other fungi, including 48-50% identity with AnStuA, AfStuA, and PmStuA from filamentous fungi[12]. Deletion of WdSTUA in Wangiella dermatitidis produces convoluted instead of normal smooth colony surface growth, and reduces aerial hyphal growth and conidiation, demonstrating the importance of this APSES protein for fungal development in this pathogenic species[32]. The conservation of structure and function across such divergent fungal species indicates that APSES proteins represent an ancient and fundamental class of developmental regulators in fungi.
Mechanisms of Coordinate Regulation
Multi-Input Integration at Target Promoters
The extensive combinatorial binding observed at pseudohyphal genes reflects a design principle where individual target promoters receive regulatory input from multiple transcription factors[11][29][38]. This architecture provides several advantages for developmental regulation. First, it allows integration of multiple independent signals, creating AND-gate logic where multiple conditions must be satisfied to activate genes. Second, it provides robustness through redundancy, such that loss of individual regulatory factors may be partially compensated by remaining factors. Third, it allows for graded responses where varying levels of multiple transcription factors produce distinct levels of gene expression.
The promoter of FLO11, a critical target gene in the pseudohyphal growth network, exemplifies this multi-input integration[8][11][29][38][41]. The FLO11 promoter contains binding sites for Ste12, Tec1, and other transcription factors, and is under control of multiple signaling cascades including the cAMP-PKA pathway, the Fus3/Kss1 MAPK cascade, the Snf1 pathway, the general amino acid control system, and the target of rapamycin (TOR) network[8][41]. This extensive regulation ensures that FLO11 expression is activated only when appropriate environmental and metabolic conditions are met for the initiation of filamentous growth.
Kinase Networks and Localization-Based Regulation
Recent analysis of yeast kinome reveals that multiple protein kinases undergo striking subcellular relocalization during filamentous growth[19]. Six cytoplasmic kinases including Bcy1, Fus3, Ksp1, Kss1, Sks1, and Tpk2 translocate predominantly to the nucleus during filamentous growth, representing an overlooked regulatory component of the filamentous growth response[19]. These kinases form an interdependent, localization-based regulatory network where deletion of individual kinases disrupts nuclear translocation of other kinases[19]. This suggests that the subcellular localization and concentration of kinases represent important regulatory parameters controlling the filamentous growth response, in addition to kinase activity itself.
Broader Biological Context and Phenotypic Consequences
Agar Invasion and Invasive Growth
Beyond pseudohyphal growth, Phd1 also regulates invasive growth, the ability of yeast cells to penetrate solid growth substrates[11][35][38]. The mechanism of agar invasion involves penetration of individual cells into the agar matrix beneath a yeast colony. Haploid cells exhibit invasive growth on rich media through a process involving filament formation and agar penetration, both of which depend on elements of the MAP kinase cascade[35]. The genetic analysis reveals that invasive growth and filament formation, while often occurring together, are in fact distinct cellular processes that can be regulated independently[35]. Both diploid pseudohyphal growth and haploid invasive growth require the transcriptional activator Ste12, but diploid pseudohyphal growth additionally requires elements of the cAMP-PKA pathway including Phd1[35][38].
Overexpression of Phd1 stimulates agar invasion, producing strains with enhanced ability to penetrate growth substrates[11][38]. The invasiveness phenotype correlates with increased expression of FLO11 and other genes required for cell-cell and cell-matrix adhesion. The role of Phd1 in invasive growth extends beyond its function in pseudohyphae formation, as Phd1 can induce some aspects of invasiveness independently of effects on filament morphology[13][21]. This indicates that Phd1 coordinates multiple distinct cellular processes contributing to the invasive growth phenotype.
Virulence and Pathogenic Implications
In Candida albicans, the APSES protein Efg1 is required for virulence in animal models of infection[6]. Strains carrying deletions of EFG1 fail to undergo the yeast-to-hyphal transition under most conditions and show reduced virulence[6][12]. The ability to form filaments appears to be important for fungal pathogenesis, as the switch to hyphal growth correlates with invasive behavior and tissue penetration. The conservation of APSES function across Saccharomyces cerevisiae and pathogenic fungi suggests that insights into Phd1 function in baker's yeast may provide valuable information relevant to understanding virulence mechanisms in human fungal pathogens.
Cell Wall Integrity and Stress Responses
Beyond morphological effects, Phd1 appears to influence cell wall integrity and cellular stress responses[6]. In Candida parapsilosis, deletion of EFG1 confers sensitivity to SDS and caspofungin (a β-glucan synthesis inhibitor) and reduces growth on calcofluor white, a dye that binds to cell wall components[6]. Both smooth and concentric cells lacking Efg1 are more susceptible to cell wall stress, with the phenotype of smooth cells being particularly dramatic[6]. Efg1 is known to be a major regulator of cell wall genes in Candida albicans, and this regulatory function appears to be conserved across APSES proteins[6]. This suggests that Phd1 coordinately regulates genes involved in cell wall synthesis and remodeling, ensuring that the cell wall is appropriately modified to accommodate the morphological changes accompanying filamentous growth.
Integration of Transcriptional and Post-Translational Regulation
Convergence of Multiple Regulatory Mechanisms
The control of Phd1 exemplifies sophisticated integration of multiple regulatory mechanisms operating at distinct levels. At the transcriptional level, PHD1 gene expression is upregulated in response to nitrogen starvation and controlled by the cAMP-PKA pathway through the Tpk1 kinase[8][20][21]. At the post-translational level, Phd1 protein stability is regulated by Cdk8-dependent phosphorylation, with high Cdk8 activity promoting Phd1 degradation and low Cdk8 activity during nutrient starvation allowing Phd1 accumulation[20][23]. Additionally, Phd1 activity is modulated through its interaction with the Tup1-Cyc8 corepressor complex and its recruitment of additional cofactors to target promoters[7][17].
This multi-level regulation provides redundancy and ensures robust control of the developmental decision. If transcriptional upregulation of PHD1 alone were the only regulatory mechanism, cells might fail to respond appropriately if transcription factors were unavailable or if their synthesis was impaired. The addition of post-translational control through protein stability regulation provides a secondary checkpoint that allows rapid adjustment of Phd1 protein levels in response to changing metabolic conditions. The recruitment of specific cofactors to target promoters provides a third level of regulation, allowing the same transcription factor to activate different sets of genes depending on available cofactors and chromatin state.
Conclusion
PHD1 encodes a master transcriptional regulator that plays a central role in controlling pseudohyphal growth and morphogenesis in Saccharomyces cerevisiae[1][11][20][38]. As a member of the APSES family of fungal transcription factors, Phd1 contains a conserved basic helix-loop-helix DNA-binding domain that recognizes E-box-like consensus sequences in target promoters[4][9][10][39]. The protein serves as a major regulatory hub that integrates input from multiple signal transduction pathways including the cAMP-PKA pathway, the MAPK cascade, and nutrient sensing mechanisms to coordinate the elaborate transcriptional and morphological changes associated with filamentous growth[8][11][29][38][43]. Phd1 directly regulates hundreds of genes including the critical FLO11 cell adhesion gene and numerous other transcription factors that form regulatory cascades controlling filamentous development[11][38]. As an inherently unstable protein degraded through Cdk8-dependent phosphorylation, Phd1 provides a sensitive molecular sensor of nutrient availability that allows rapid activation of developmental programs when nutrients become limiting[20][23].
The position of Phd1 as a master regulator controlling complex developmental responses highlights the importance of transcription factor regulation in fungal development and suggests that similar hierarchical regulatory architectures may operate in other eukaryotic developmental systems[11][38]. The conservation of APSES proteins across diverse fungal species, from laboratory strains of Saccharomyces cerevisiae to human pathogenic fungi such as Candida albicans and Wangiella dermatitidis, indicates that this regulatory system has ancient evolutionary origins and remains fundamental for fungal development[28][32]. Further investigation of Phd1 function and its interactions with upstream signaling pathways and downstream target genes will provide valuable insights into the fundamental principles governing cell differentiation and morphogenesis in fungi and potentially inform strategies for controlling pathogenic fungal morphogenesis and virulence.
Citations
- https://pubmed.ncbi.nlm.nih.gov/8114741/
- https://pubmed.ncbi.nlm.nih.gov/15247232/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8980467/
- https://www.yeastgenome.org/locus/S000001526
- https://www.tgsmith.com/PDFs/PHD_abstract_2011.pdf
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3912905/
- https://thebiogrid.org/34089/summary/saccharomyces-cerevisiae/phd1.html
- https://journals.asm.org/doi/10.1128/mbio.00427-15
- https://pmc.ncbi.nlm.nih.gov/articles/PMC452574/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1369046/
- https://journals.asm.org/doi/10.1128/ec.00037-07
- https://onlinelibrary.wiley.com/doi/full/10.1046/j.1365-2958.2003.03358.x
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4316177/
- https://portlandpress.com/biochemj/article/462/3/567/47800/Regulation-of-PKA-activity-by-an
- https://pmc.ncbi.nlm.nih.gov/articles/PMC133872/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3266601/
- https://www.molbiolcell.org/doi/10.1091/mbc.e07-11-1199
- https://pubmed.ncbi.nlm.nih.gov/22124158/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC102143/
- https://pubmed.ncbi.nlm.nih.gov/17485461/
- https://pubmed.ncbi.nlm.nih.gov/11046133/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3334620/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1489142/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6454542/
- https://genesdev.cshlp.org/content/20/4/435
- https://febs.onlinelibrary.wiley.com/doi/10.1111/j.1742-4658.2010.07728.x
- https://core.ac.uk/download/pdf/4398885.pdf
- https://pmc.ncbi.nlm.nih.gov/articles/PMC358570/
- https://dspace.mit.edu/handle/1721.1/33506
- https://genesdev.cshlp.org/content/8/24/2974.full.pdf
- https://pmc.ncbi.nlm.nih.gov/articles/PMC317105/
- https://en.wikipedia.org/wiki/Basic_helix%E2%80%93loop%E2%80%93helix
- https://cfatg.org/en/publis_scientifiques/the-mitochondrial-phosphatidylserine-decarboxylase-psd1-is-involved-in-nitrogen-starvation-induced-mitophagy-in-yeast/
- https://www.molbiolcell.org/doi/10.1091/mbc.9.1.161
- https://pmc.ncbi.nlm.nih.gov/articles/PMC463060/
- https://pubmed.ncbi.nlm.nih.gov/9384580/
- https://pubmed.ncbi.nlm.nih.gov/9832522/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC12091097/
- https://frontend.qa.yeastgenome.org/locus/S000001526/literature
- https://academic.oup.com/genetics/article-abstract/150/4/1443/6034467
- https://yeastgenome.org/reference/S000137016