Deep Research
Falcon
(RCO1-deep-research-falcon.md)
Falcon
(RCO1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Functional annotation research report: RCO1 (UniProt Q04779; YMR075W) in Saccharomyces cerevisiae (S288c)
0. Identity verification (critical)
Primary literature explicitly identifies S. cerevisiae Rco1 as UniProt Q04779 and as a PHD-domain protein that is a unique, defining subunit of the Rpd3S histone deacetylase complex (distinct from the promoter-focused Rpd3L complex). (ruan2016homodimericphddomaincontaining pages 3-4, drouin2010dsifandrna pages 1-2, li2023structureofhistone pages 1-3)
1. Key concepts, definitions, and current understanding
1.1 What RCO1 encodes (and what it does not)
RCO1 encodes Rco1, a non-catalytic chromatin regulatory protein. It is not itself a histone deacetylase; rather, it is a reader/targeting and regulatory subunit required for proper activity and chromatin engagement of the Rpd3S (Rpd3 small) HDAC complex. (li2023structureofhistone pages 1-3, mcdaniel2016combinatorialhistonereadout pages 2-4)
1.2 Rpd3S pathway context (Set2–H3K36 methylation and transcription elongation)
Rpd3S is a histone deacetylase complex that operates co-transcriptionally across coding regions (ORFs/gene bodies) to remove histone acetylation “in the wake” of RNA polymerase II (Pol II), thereby suppressing cryptic (spurious intragenic) transcription initiation. Rco1 is one of the Rpd3S-specific subunits underlying this gene-body surveillance function. (drouin2010dsifandrna pages 1-2, govind2010phosphorylatedpolii pages 1-2)
A central targeting axis for Rpd3S is the Set2 → H3K36 methylation pathway: Set2 deposits H3K36 methylation during elongation, which is read by Eaf3, while Rco1 contributes complementary histone-tail engagement and intra-complex regulation. (govind2010phosphorylatedpolii pages 1-2, lee2018combinatorialgeneticcontrol pages 1-2)
1.3 Reader domains in Rco1: PHD fingers and combinatorial histone readout
Rco1 contains two plant homeodomain (PHD) zinc-finger modules (PHD1 and PHD2). Biochemical evidence indicates that both PHD1 and PHD2 bind the extreme N-terminus of histone H3 (H3 1–20) and that H3K4 trimethylation reduces binding—a mechanism that helps restrict Rpd3S away from promoter nucleosomes enriched in H3K4me3. (mcdaniel2016combinatorialhistonereadout pages 2-4)
More recent structural interpretations emphasize combinatorial readout: Rco1 preferentially reads unmodified H3K4 while Eaf3 reads H3K36me3, together directing productive nucleosome engagement and affecting which histone tails/lysines are positioned for deacetylation. (guan2023diversemodesof pages 1-2, zhang2023structuralbasisfor pages 1-2)
1.4 Core molecular interactions and complex membership
Rpd3S is classically described as a five-subunit complex (Rpd3, Sin3, Ume1, Eaf3, Rco1). Mechanistic work shows Rco1 is a critical interaction hub: Rco1 forms key contacts with Eaf3 (via a Sin3 interaction domain/SID interacting with Eaf3’s MRG domain) and is required for full Rpd3S nucleosome engagement and cryptic transcription suppression. (ruan2015nucleosomecontacttriggers pages 3-5, ruan2016homodimericphddomaincontaining pages 1-2)
2. Biological function, processes, and localization
2.1 Primary biological role: suppressing cryptic intragenic transcription
Rpd3S functions across transcribed ORFs to deacetylate histones after Pol II passage and thereby prevents cryptic transcription initiation within genes. Rco1 is an essential Rpd3S subunit for this process; perturbing Rco1 regions needed for nucleosome engagement causes cryptic transcription phenotypes even when overall complex integrity is maintained. (drouin2010dsifandrna pages 1-2, ruan2016homodimericphddomaincontaining pages 6-8)
2.2 Chromatin and nuclear localization
Functional fractionation and chromatin association assays show wild-type Rco1 is predominantly chromatin-associated, consistent with its role in Rpd3S recruitment/engagement at gene bodies. Genome-scale localization approaches place Rpd3S/Rco1 primarily within coding regions at active genes. (mcdaniel2016combinatorialhistonereadout pages 2-4, drouin2010dsifandrna pages 1-2)
2.3 Recruitment mechanisms: integration of histone marks and transcription machinery
Although histone mark readout is central (Eaf3–H3K36me and Rco1–H3 N-terminus/H3K4me0), recruitment is also coordinated by Pol II-associated factors. ChIP–Chip evidence indicates Rpd3S binds active ORFs but is not recruited to every active gene; importantly, recruitment can be Set2-independent in some contexts, whereas complexes recruited without H3K36 methylation can be inactive, suggesting methylation contributes to activation and/or productive substrate engagement. (drouin2010dsifandrna pages 1-2)
Recruitment and/or stabilization of Rpd3S at transcribed loci is additionally linked to Pol II CTD phosphorylation (Kin28, Ctk1) and DSIF (Spt4/5). (drouin2010dsifandrna pages 1-2, govind2010phosphorylatedpolii pages 1-2)
3. Recent developments (prioritizing 2023–2024)
3.1 2023: Structural resolution of Rpd3S–nucleosome engagement and Rco1’s placement
A set of 2023 cryo-EM studies transformed understanding of Rpd3S targeting and catalysis by resolving how the holoenzyme binds nucleosomes.
-
Stoichiometry and architecture. Multiple structures indicate Rpd3S contains two Eaf3–Rco1 modules arranged around Rpd3/Sin3, with two copies of Rco1 and two copies of Eaf3. This duplicated module architecture supports multivalent nucleosome engagement and helps explain preference for dinucleosomal substrates. (li2023structureofhistone pages 1-3, markert2023structureofthea pages 1-3)
-
Mechanistic nucleosome contacts. Cryo-EM structures show multivalent engagement of histone tails, nucleosomal DNA, and linker DNA. Rco1 is positioned to contribute to nucleosome binding and to help route/position the H3 N-terminal tail for catalysis. (zhang2023structuralbasisfor pages 1-2, li2023structureofhistone pages 1-3)
-
Context-dependent catalytic modes. Structural/biochemical work indicates H3K36me3-guided docking can orient Rpd3S toward H4-tail deacetylation in one mode, while a combinatorial readout involving Rco1 (unmodified H3K4) and Eaf3 (H3K36me3) can direct H3-tail deacetylation with site selectivity (e.g., sparing “registered” H3K9ac in a particular mode). (guan2023diversemodesof pages 1-2)
3.2 2024: Expert commentary and emerging mechanistic extensions (Rco1 IDRs and traveling with Pol II)
A 2024 expert commentary synthesizing 2023 structures highlights Rco1 as a specificity determinant: its PHD engagement of unmodified H3K4 and DNA contacts contribute to gene-body targeting and help rationalize how Rpd3S distinguishes promoter versus coding-region chromatin. (carrozza2024rpd3smeetsthe pages 1-2)
A 2024 preprint extends recruitment/allostery models by proposing an important role for an Rco1 N-terminal intrinsically disordered region (IDR) in Pol II association: mutations in a basic K/R cluster reduce association with the Pol II CTD without disrupting nucleosome recognition, and a minimal CTD-binding module involving Rco1-PHD1 and Eaf3-CHD is described. (li2024intrinsicallydisorderedregions pages 1-4)
4. Mechanistic model for Rco1 function (integrated synthesis)
4.1 Multivalent chromatin reading and allosteric cooperation with Eaf3
Rco1’s PHD domains contribute to nucleosome engagement by binding the H3 N-terminus; meanwhile, Rco1’s SID/interaction region binds Eaf3’s MRG domain and can allosterically enhance Eaf3 chromodomain engagement with H3K36-methylated peptides, supporting a cooperative model where “reader” domains are weak alone but strengthened in the assembled complex. (ruan2015nucleosomecontacttriggers pages 3-5, ruan2016homodimericphddomaincontaining pages 1-2)
4.2 Exclusion from promoters via H3K4 methylation sensitivity
H3K4me3 reduces binding of both Rco1 PHD domains to the H3 N-terminus, providing a plausible molecular basis for preferential Rpd3S action in gene bodies rather than H3K4me3-rich promoters. (mcdaniel2016combinatorialhistonereadout pages 2-4, lee2018combinatorialgeneticcontrol pages 1-2)
4.3 Structural routing of H3 tail and substrate selection
High-resolution structures show that engagement of unmodified H3K4 (via Rco1) can immobilize portions of the H3 tail and influence which lysines can access the catalytic pocket, yielding structural rationales for lysine-specific deacetylation patterns. (zhang2023structuralbasisfor pages 1-2)
5. Applications and real-world implementations
5.1 Yeast as an implementation platform for epigenetic mechanism discovery
Rco1/Rpd3S has become a model system for understanding histone mark-guided HDAC targeting, including multivalent recognition, reader-domain allostery, and elongation-coupled chromatin restoration. Modern cryo-EM reconstructions of Rpd3S–nucleosome complexes (2023) provide atomic frameworks that are now used to interpret conserved SIN3/HDAC complex behavior in other eukaryotes. (li2023structureofhistone pages 1-3, zhang2023structuralbasisfor pages 1-2)
5.2 Tooling and experimental implementations
- Structure-guided mutagenesis. 2023 structures map Rco1 regions that contact H3 tail/DNA and define residues/segments for functional perturbation; these are used to test cryptic transcription suppression and chromatin binding. (zhang2023structuralbasisfor pages 1-2, dong2023structuralbasisof pages 7-8)
- Mechanistic coupling to Pol II. Recruitment models incorporating CTD phosphorylation and DSIF have practical implications for designing experiments that decouple “recruitment” from “activation” of HDAC function at transcribed chromatin. (drouin2010dsifandrna pages 1-2, govind2010phosphorylatedpolii pages 1-2)
6. Expert opinion and authoritative analysis
The 2024 commentary emphasizes that Rpd3S achieves specificity through multivalent contacts (Rco1, Eaf3, Sin3) and that multiple structural “states” imply conformational plasticity relevant to substrate selection and chromatin context, with Rco1 contributing to gene-body targeting via PHD readout of H3K4me0 and DNA contacts. (carrozza2024rpd3smeetsthe pages 1-2)
7. Statistics and quantitative data highlights
- Complex size: Rpd3S is reported as ~410 kDa and contains two copies of Rco1, both required for full function. (ruan2016homodimericphddomaincontaining pages 2-3)
- Cryo-EM resolutions (recent): 2.8 Å for the complete Rpd3S–nucleosome complex (Nature Communications, Dec 2023). (markert2023structureofthea pages 1-3)
- Cryo-EM resolutions (additional 2023 structures): 3.1 Å (overall) / 3.0 Å (focused) for Rpd3S–mono-nucleosome; 10.5 Å for a flexible di-nucleosome reconstruction (Nature Structural & Molecular Biology, Oct 2023). (li2023structureofhistone pages 1-3)
- Linker DNA preference: Rpd3S binding shows sensitivity to linker length with a modest preference for ~30–40 bp linkers, and preference for di-nucleosome over mono-nucleosome in vitro. (li2023structureofhistone pages 1-3)
- Histone-tail affinity: Rco1 PHD1 binds unmodified H3(1–10) with Kd ~39 µM (ITC); PHD2 showed no detectable H3(1–10) reader activity in that assay context (Research Square preprint, May 2023). (li2023dynamicandmultivalent pages 7-10)
- Lysine preference (commentary summary): Rpd3S deacetylates H3K23 and H3K14 more efficiently than H3K9, H3K18, and H3K27, indicating lysine-level selectivity likely shaped by structural positioning and engagement mode (Cell Research commentary, Nov 2024). (carrozza2024rpd3smeetsthe pages 2-2)
8. Visual evidence (key structural panels)
The following figure panels (cropped) visually support the statements about (i) overall Rpd3S–nucleosome architecture and (ii) Rco1 PHD engagement of the H3 N-terminus: (guan2023diversemodesof media a0d7f516, guan2023diversemodesof media 8facf5db, guan2023diversemodesof media 8448b558).
9. Evidence gaps and limitations (what was not fully retrievable here)
Some potentially important quantitative elements—particularly global genome-wide recruitment percentages, many binding constants for full Rpd3S–nucleosome/CTD interactions, and numerical cryptic transcription effect sizes—were not available in the accessible excerpts used for this report. Where possible, mechanistic conclusions were supported by structural resolutions, explicitly reported Kd values (for PHD1–H3), and qualitative but experimentally grounded phenotypes. (drouin2010dsifandrna pages 1-2, li2023dynamicandmultivalent pages 7-10)
Summary table
| Category | Evidence-backed details | Key citations (with year) | URL |
|---|---|---|---|
| Verified identity | RCO1/YMR075W/UniProt Q04779 in Saccharomyces cerevisiae is the Rco1 subunit of the Rpd3S histone deacetylase complex; primary literature describes it as a PHD zinc-finger/plant homeodomain-containing chromatin regulator, not an enzyme on its own. | Li 2023 (li2023structureofhistone pages 1-3); Ruan 2016 (ruan2016homodimericphddomaincontaining pages 3-4, ruan2016homodimericphddomaincontaining pages 2-3) | https://doi.org/10.1038/s41594-023-01121-5; https://doi.org/10.1074/jbc.m115.703637 |
| Molecular function within Rpd3S | Rco1 helps target and activate Rpd3S on transcribed chromatin so the complex can deacetylate histones in coding regions after Pol II passage, maintaining hypoacetylated gene bodies and suppressing cryptic intragenic transcription. It acts together with Eaf3, Sin3, Rpd3, and Ume1. | Drouin 2010 (drouin2010dsifandrna pages 1-2); Govind 2010 (govind2010phosphorylatedpolii pages 1-2); Li 2023 (li2023structureofhistone pages 1-3) | https://doi.org/10.1371/journal.pgen.1001173; https://doi.org/10.1016/j.molcel.2010.07.003; https://doi.org/10.1038/s41594-023-01121-5 |
| Domains / reader activities | Rco1 contains two PHD fingers (PHD1, PHD2). These domains bind the extreme H3 N-terminus (H3 1–20) and preferentially read unmodified H3K4 (H3K4me0); H3K4me3 reduces binding, helping exclude Rpd3S from promoter chromatin. Rco1 also contains a SID/MRG-interacting region that connects functionally to Eaf3. | McDaniel 2016 (mcdaniel2016combinatorialhistonereadout pages 2-4, mcdaniel2016combinatorialhistonereadout pages 4-6); Guan 2023 (guan2023diversemodesof pages 1-2); Zhang 2023 (zhang2023structuralbasisfor pages 1-2) | https://doi.org/10.1074/jbc.m116.720193; https://doi.org/10.1038/s41586-023-06349-1; https://doi.org/10.1038/s41422-023-00884-2 |
| Binding partners / complex architecture | Core partners are Rpd3, Sin3, Ume1, and Eaf3. Structural studies show two copies of Rco1 and two copies of Eaf3 organized around catalytic Rpd3; Rco1 forms Eaf3–Rco1 heterodimers and can homodimerize, making it a major interaction hub in Rpd3S. | Markert 2023 (markert2023structureofthea pages 1-3); Li 2023 (li2023structureofhistone pages 1-3); Ruan 2016 (ruan2016homodimericphddomaincontaining pages 3-4, ruan2016homodimericphddomaincontaining pages 1-2) | https://doi.org/10.1038/s41467-023-43968-8; https://doi.org/10.1038/s41594-023-01121-5; https://doi.org/10.1074/jbc.m115.703637 |
| Recruitment determinants | Recruitment/function is coupled to Set2-dependent H3K36 methylation via Eaf3 chromodomain recognition, while Rco1 contributes H3 tail readout and nucleosome engagement. Rpd3S recruitment to active ORFs is also coordinated by RNAPII CTD phosphorylation (especially Ser5P, with S2P contribution) and DSIF/Spt4-Spt5. Recent work further suggests an Rco1 IDR contributes to Pol II CTD association. | Drouin 2010 (drouin2010dsifandrna pages 1-2); Govind 2010 (govind2010phosphorylatedpolii pages 1-2); Li 2024 preprint (li2024intrinsicallydisorderedregions pages 1-4) | https://doi.org/10.1371/journal.pgen.1001173; https://doi.org/10.1016/j.molcel.2010.07.003; https://doi.org/10.21203/rs.3.rs-4000909/v1 |
| Cellular localization | Rco1 is predominantly chromatin-associated and localizes to active ORFs / gene bodies rather than promoters, consistent with Rpd3S function in transcribed coding regions. Fractionation studies place wild-type Rco1 mainly in the chromatin fraction. | Drouin 2010 (drouin2010dsifandrna pages 1-2); McDaniel 2016 (mcdaniel2016combinatorialhistonereadout pages 2-4); McDaniel thesis 2016 (stephen2016expandingthefunction pages 86-91) | https://doi.org/10.1371/journal.pgen.1001173; https://doi.org/10.1074/jbc.m116.720193; https://doi.org/10.17615/shj3-8048 |
| Mechanistic insight from 2023–2024 studies | 2023 cryo-EM studies show multivalent nucleosome engagement: Rco1, Eaf3, and Sin3 contact H3K36me3-marked nucleosomes, nucleosomal DNA, and linker DNA. Rco1 PHD1 helps place the H3 tail toward the catalytic center, while alternate engagement modes explain context-dependent H3/H4 deacetylation. A 2024 commentary interprets Rco1 as a specificity determinant for gene-body chromatin and lysine selection. | Guan 2023 (guan2023diversemodesof pages 1-2, guan2023diversemodesof media a0d7f516); Li 2023 (li2023structureofhistone pages 1-3); Carrozza & Workman 2024 (carrozza2024rpd3smeetsthe pages 1-2) | https://doi.org/10.1038/s41586-023-06349-1; https://doi.org/10.1038/s41594-023-01121-5; https://doi.org/10.1038/s41422-023-00899-9 |
| Phenotypes / functional readouts | rco1Δ or PHD-disrupting mutants impair nucleosome/chromatin binding and cause cryptic transcription phenotypes in reporter assays; truncations or mutations in key Rco1 regions compromise Rpd3S function even when complex assembly is retained. Genetic studies also show Rco1-mediated Rpd3S is antagonized by H3K4 methylation and promoted by H3K36 methylation. | Ruan 2016 (ruan2016homodimericphddomaincontaining pages 6-8); McDaniel 2016 (mcdaniel2016combinatorialhistonereadout pages 2-4, stephen2016expandingthefunction pages 86-91); Lee 2018 (lee2018combinatorialgeneticcontrol pages 1-2) | https://doi.org/10.1074/jbc.m115.703637; https://doi.org/10.1074/jbc.m116.720193; https://doi.org/10.1534/g3.118.200589 |
| Key quantitative metrics | Reported values include ~410 kDa for Rpd3S, 2× Rco1 + 2× Eaf3 stoichiometry in 2023 structures, and cryo-EM resolutions of 2.8 Å (Markert 2023), 3.1/3.0 Å with 10.5 Å di-nucleosome map (Li 2023), and 3.5 Å (Dong 2023). Rpd3S shows a modest preference for 30–40 bp linker DNA and di-nucleosomes. One measured affinity from structural/biochemical work: Rco1 PHD1 binds H3(1–10) with Kd ~39 µM; PHD2 showed no detectable H3(1–10) reader activity in that assay. | Markert 2023 (markert2023structureofthea pages 1-3); Li 2023 (li2023structureofhistone pages 1-3); Dong 2023 (dong2023structuralbasisof pages 1-2); Ruan 2016 (ruan2016homodimericphddomaincontaining pages 2-3); Li/Guan preprint 2023 (li2023dynamicandmultivalent pages 7-10) | https://doi.org/10.1038/s41467-023-43968-8; https://doi.org/10.1038/s41594-023-01121-5; https://doi.org/10.1038/s41422-023-00869-1; https://doi.org/10.1074/jbc.m115.703637; https://doi.org/10.21203/rs.3.rs-2060494/v1 |
| Enzymatic/substrate specificity context | Rco1 is not the catalytic HDAC; it is a reader/targeting and regulatory subunit. Recent structural commentary notes Rpd3S deacetylates H3K23 and H3K14 more efficiently than H3K9, H3K18, and H3K27, and some structures place H3K18 or H3/H4 tails near the active site depending on nucleosome engagement mode. | Carrozza & Workman 2024 (carrozza2024rpd3smeetsthe pages 2-2); Guan 2023 (guan2023diversemodesof pages 1-2, guan2023diversemodesof pages 5-6) | https://doi.org/10.1038/s41422-023-00899-9; https://doi.org/10.1038/s41586-023-06349-1 |
Table: This table compiles evidence-backed functional annotation for Saccharomyces cerevisiae RCO1/Q04779, emphasizing its role as the PHD-containing targeting subunit of the Rpd3S histone deacetylase complex. It combines recent 2023-2024 structural advances with foundational mechanistic studies and includes URLs for rapid source checking.
Most recent/high-priority references used (publication date; URL):
* Guan et al., Nature (Jul 2023): https://doi.org/10.1038/s41586-023-06349-1 (guan2023diversemodesof pages 1-2)
* Li et al., Nat Struct Mol Biol (Oct 2023): https://doi.org/10.1038/s41594-023-01121-5 (li2023structureofhistone pages 1-3)
* Dong et al., Cell Research (Sep 2023): https://doi.org/10.1038/s41422-023-00869-1 (dong2023structuralbasisof pages 1-2)
* Zhang et al., Cell Research (Oct 2023): https://doi.org/10.1038/s41422-023-00884-2 (zhang2023structuralbasisfor pages 1-2)
* Markert et al., Nat Communications (Dec 2023): https://doi.org/10.1038/s41467-023-43968-8 (markert2023structureofthea pages 1-3)
* Carrozza & Workman, Cell Research (Nov 2024 commentary): https://doi.org/10.1038/s41422-023-00899-9 (carrozza2024rpd3smeetsthe pages 1-2)
References
-
(ruan2016homodimericphddomaincontaining pages 3-4): Chun Ruan, Haochen Cui, Chul-Hwan Lee, Sheng Li, and Bing Li. Homodimeric phd domain-containing rco1 subunit constitutes a critical interaction hub within the rpd3s histone deacetylase complex. Journal of Biological Chemistry, 291:5428-5438, Mar 2016. URL: https://doi.org/10.1074/jbc.m115.703637, doi:10.1074/jbc.m115.703637. This article has 27 citations and is from a domain leading peer-reviewed journal.
-
(drouin2010dsifandrna pages 1-2): Simon Drouin, Louise Laramée, Pierre-Étienne Jacques, Audrey Forest, Maxime Bergeron, and François Robert. Dsif and rna polymerase ii ctd phosphorylation coordinate the recruitment of rpd3s to actively transcribed genes. PLoS Genetics, 6:e1001173, Oct 2010. URL: https://doi.org/10.1371/journal.pgen.1001173, doi:10.1371/journal.pgen.1001173. This article has 180 citations and is from a domain leading peer-reviewed journal.
-
(li2023structureofhistone pages 1-3): Wulong Li, Hengjun Cui, Zhimin Lu, and Haibo Wang. Structure of histone deacetylase complex rpd3s bound to nucleosome. Nature structural & molecular biology, 30:1893-1901, Oct 2023. URL: https://doi.org/10.1038/s41594-023-01121-5, doi:10.1038/s41594-023-01121-5. This article has 18 citations and is from a highest quality peer-reviewed journal.
-
(mcdaniel2016combinatorialhistonereadout pages 2-4): Stephen L. McDaniel, Jennifer E. Fligor, Chun Ruan, Haochen Cui, Joseph B. Bridgers, Julia V. DiFiore, Angela H. Guo, Bing Li, and Brian D. Strahl. Combinatorial histone readout by the dual plant homeodomain (phd) fingers of rco1 mediates rpd3s chromatin recruitment and the maintenance of transcriptional fidelity. Journal of Biological Chemistry, 291:14796-14802, Jul 2016. URL: https://doi.org/10.1074/jbc.m116.720193, doi:10.1074/jbc.m116.720193. This article has 32 citations and is from a domain leading peer-reviewed journal.
-
(govind2010phosphorylatedpolii pages 1-2): Chhabi K. Govind, Hongfang Qiu, Daniel S. Ginsburg, Chun Ruan, Kimberly Hofmeyer, Cuihua Hu, Venkatesh Swaminathan, Jerry L. Workman, Bing Li, and Alan G. Hinnebusch. Phosphorylated pol ii ctd recruits multiple hdacs, including rpd3c(s), for methylation-dependent deacetylation of orf nucleosomes. Molecular cell, 39:234-246, Jul 2010. URL: https://doi.org/10.1016/j.molcel.2010.07.003, doi:10.1016/j.molcel.2010.07.003. This article has 273 citations and is from a highest quality peer-reviewed journal.
-
(lee2018combinatorialgeneticcontrol pages 1-2): Kwan Yin Lee, Mathieu Ranger, and Marc D. Meneghini. Combinatorial genetic control of rpd3s through histone h3k4 and h3k36 methylation in budding yeast. G3: Genes|Genomes|Genetics, 8:3411-3420, Jul 2018. URL: https://doi.org/10.1534/g3.118.200589, doi:10.1534/g3.118.200589. This article has 20 citations.
-
(guan2023diversemodesof pages 1-2): Haipeng Guan, Pei Wang, Pei Zhang, Chun Ruan, Yutian Ou, Bo Peng, Xiangdong Zheng, Jianlin Lei, Bing Li, Chuangye Yan, and Haitao Li. Diverse modes of h3k36me3-guided nucleosomal deacetylation by rpd3s. Nature, 620:669-675, Jul 2023. URL: https://doi.org/10.1038/s41586-023-06349-1, doi:10.1038/s41586-023-06349-1. This article has 40 citations and is from a highest quality peer-reviewed journal.
-
(zhang2023structuralbasisfor pages 1-2): Yueyue Zhang, Mengxue Xu, Po Wang, Jiahui Zhou, Guangxian Wang, Shuailong Han, Gang Cai, and Xuejuan Wang. Structural basis for nucleosome binding and catalysis by the yeast rpd3s/hdac holoenzyme. Cell Research, 33:971-974, Oct 2023. URL: https://doi.org/10.1038/s41422-023-00884-2, doi:10.1038/s41422-023-00884-2. This article has 12 citations and is from a domain leading peer-reviewed journal.
-
(ruan2015nucleosomecontacttriggers pages 3-5): Chun Ruan, Chul-Hwan Lee, Haochen Cui, Sheng Li, and Bing Li. Nucleosome contact triggers conformational changes of rpd3s driving high-affinity h3k36me nucleosome engagement. Cell reports, 10 2:204-15, Jan 2015. URL: https://doi.org/10.1016/j.celrep.2014.12.027, doi:10.1016/j.celrep.2014.12.027. This article has 55 citations and is from a highest quality peer-reviewed journal.
-
(ruan2016homodimericphddomaincontaining pages 1-2): Chun Ruan, Haochen Cui, Chul-Hwan Lee, Sheng Li, and Bing Li. Homodimeric phd domain-containing rco1 subunit constitutes a critical interaction hub within the rpd3s histone deacetylase complex. Journal of Biological Chemistry, 291:5428-5438, Mar 2016. URL: https://doi.org/10.1074/jbc.m115.703637, doi:10.1074/jbc.m115.703637. This article has 27 citations and is from a domain leading peer-reviewed journal.
-
(ruan2016homodimericphddomaincontaining pages 6-8): Chun Ruan, Haochen Cui, Chul-Hwan Lee, Sheng Li, and Bing Li. Homodimeric phd domain-containing rco1 subunit constitutes a critical interaction hub within the rpd3s histone deacetylase complex. Journal of Biological Chemistry, 291:5428-5438, Mar 2016. URL: https://doi.org/10.1074/jbc.m115.703637, doi:10.1074/jbc.m115.703637. This article has 27 citations and is from a domain leading peer-reviewed journal.
-
(markert2023structureofthea pages 1-3): Jonathan W. Markert, Seychelle M. Vos, and Lucas Farnung. Structure of the complete saccharomyces cerevisiae rpd3s-nucleosome complex. Nature Communications, Dec 2023. URL: https://doi.org/10.1038/s41467-023-43968-8, doi:10.1038/s41467-023-43968-8. This article has 10 citations and is from a highest quality peer-reviewed journal.
-
(carrozza2024rpd3smeetsthe pages 1-2): Michael J. Carrozza and Jerry L. Workman. Rpd3s meets the nucleosome. Cell research, 34:1-2, Nov 2024. URL: https://doi.org/10.1038/s41422-023-00899-9, doi:10.1038/s41422-023-00899-9. This article has 2 citations and is from a domain leading peer-reviewed journal.
-
(li2024intrinsicallydisorderedregions pages 1-4): Bing Li, Yixuan Pan, Meiyang Liu, Chun Ruan, Mengyuan Peng, Min Hao, Qi Zhang, Jingdong Xue, Ningzhe Li, Haipeng Guan, Pei Wang, Mingqian Hu, Haitao Li, Wenjuan Wang, Juan Song, Yanhua Yao, and Yimin Lao. Intrinsically disordered regions steer the function coordination of the traveling chromatin modifier during pol ii elongation. Unknown journal, Mar 2024. URL: https://doi.org/10.21203/rs.3.rs-4000909/v1, doi:10.21203/rs.3.rs-4000909/v1.
-
(dong2023structuralbasisof pages 7-8): Shuqi Dong, Huadong Li, Meilin Wang, Nadia Rasheed, Binqian Zou, Xijie Gao, Jiali Guan, Weijie Li, Jiale Zhang, Chi Wang, Ningkun Zhou, Xue Shi, Mei Li, Min Zhou, Junfeng Huang, He Li, Ying Zhang, Koon Ho Wong, Xiaofei Zhang, William Chong Hang Chao, and Jun He. Structural basis of nucleosome deacetylation and dna linker tightening by rpd3s histone deacetylase complex. Cell Research, 33:790-801, Sep 2023. URL: https://doi.org/10.1038/s41422-023-00869-1, doi:10.1038/s41422-023-00869-1. This article has 12 citations and is from a domain leading peer-reviewed journal.
-
(ruan2016homodimericphddomaincontaining pages 2-3): Chun Ruan, Haochen Cui, Chul-Hwan Lee, Sheng Li, and Bing Li. Homodimeric phd domain-containing rco1 subunit constitutes a critical interaction hub within the rpd3s histone deacetylase complex. Journal of Biological Chemistry, 291:5428-5438, Mar 2016. URL: https://doi.org/10.1074/jbc.m115.703637, doi:10.1074/jbc.m115.703637. This article has 27 citations and is from a domain leading peer-reviewed journal.
-
(li2023dynamicandmultivalent pages 7-10): Haitao Li, Haipeng Guan, Pei Wang, Pei Zhang, Chun Ruan, Yutian Ou, Bo Peng, Xiangdong Zheng, Jianlin Lei, Bing Li, and Chuangye Yan. Dynamic and multivalent engagement determines context-dependent nucleosomal deacetylation by the rpd3s complex. Unknown journal, May 2023. URL: https://doi.org/10.21203/rs.3.rs-2060494/v1, doi:10.21203/rs.3.rs-2060494/v1.
-
(carrozza2024rpd3smeetsthe pages 2-2): Michael J. Carrozza and Jerry L. Workman. Rpd3s meets the nucleosome. Cell research, 34:1-2, Nov 2024. URL: https://doi.org/10.1038/s41422-023-00899-9, doi:10.1038/s41422-023-00899-9. This article has 2 citations and is from a domain leading peer-reviewed journal.
-
(guan2023diversemodesof media a0d7f516): Haipeng Guan, Pei Wang, Pei Zhang, Chun Ruan, Yutian Ou, Bo Peng, Xiangdong Zheng, Jianlin Lei, Bing Li, Chuangye Yan, and Haitao Li. Diverse modes of h3k36me3-guided nucleosomal deacetylation by rpd3s. Nature, 620:669-675, Jul 2023. URL: https://doi.org/10.1038/s41586-023-06349-1, doi:10.1038/s41586-023-06349-1. This article has 40 citations and is from a highest quality peer-reviewed journal.
-
(guan2023diversemodesof media 8facf5db): Haipeng Guan, Pei Wang, Pei Zhang, Chun Ruan, Yutian Ou, Bo Peng, Xiangdong Zheng, Jianlin Lei, Bing Li, Chuangye Yan, and Haitao Li. Diverse modes of h3k36me3-guided nucleosomal deacetylation by rpd3s. Nature, 620:669-675, Jul 2023. URL: https://doi.org/10.1038/s41586-023-06349-1, doi:10.1038/s41586-023-06349-1. This article has 40 citations and is from a highest quality peer-reviewed journal.
-
(guan2023diversemodesof media 8448b558): Haipeng Guan, Pei Wang, Pei Zhang, Chun Ruan, Yutian Ou, Bo Peng, Xiangdong Zheng, Jianlin Lei, Bing Li, Chuangye Yan, and Haitao Li. Diverse modes of h3k36me3-guided nucleosomal deacetylation by rpd3s. Nature, 620:669-675, Jul 2023. URL: https://doi.org/10.1038/s41586-023-06349-1, doi:10.1038/s41586-023-06349-1. This article has 40 citations and is from a highest quality peer-reviewed journal.
-
(mcdaniel2016combinatorialhistonereadout pages 4-6): Stephen L. McDaniel, Jennifer E. Fligor, Chun Ruan, Haochen Cui, Joseph B. Bridgers, Julia V. DiFiore, Angela H. Guo, Bing Li, and Brian D. Strahl. Combinatorial histone readout by the dual plant homeodomain (phd) fingers of rco1 mediates rpd3s chromatin recruitment and the maintenance of transcriptional fidelity. Journal of Biological Chemistry, 291:14796-14802, Jul 2016. URL: https://doi.org/10.1074/jbc.m116.720193, doi:10.1074/jbc.m116.720193. This article has 32 citations and is from a domain leading peer-reviewed journal.
-
(stephen2016expandingthefunction pages 86-91): Stephen McDaniel. Expanding the function of histone h3 lysine 36 methylation in saccharomyces cerevisiae. Text, 2016. URL: https://doi.org/10.17615/shj3-8048, doi:10.17615/shj3-8048. This article has 0 citations and is from a peer-reviewed journal.
-
(dong2023structuralbasisof pages 1-2): Shuqi Dong, Huadong Li, Meilin Wang, Nadia Rasheed, Binqian Zou, Xijie Gao, Jiali Guan, Weijie Li, Jiale Zhang, Chi Wang, Ningkun Zhou, Xue Shi, Mei Li, Min Zhou, Junfeng Huang, He Li, Ying Zhang, Koon Ho Wong, Xiaofei Zhang, William Chong Hang Chao, and Jun He. Structural basis of nucleosome deacetylation and dna linker tightening by rpd3s histone deacetylase complex. Cell Research, 33:790-801, Sep 2023. URL: https://doi.org/10.1038/s41422-023-00869-1, doi:10.1038/s41422-023-00869-1. This article has 12 citations and is from a domain leading peer-reviewed journal.
-
(guan2023diversemodesof pages 5-6): Haipeng Guan, Pei Wang, Pei Zhang, Chun Ruan, Yutian Ou, Bo Peng, Xiangdong Zheng, Jianlin Lei, Bing Li, Chuangye Yan, and Haitao Li. Diverse modes of h3k36me3-guided nucleosomal deacetylation by rpd3s. Nature, 620:669-675, Jul 2023. URL: https://doi.org/10.1038/s41586-023-06349-1, doi:10.1038/s41586-023-06349-1. This article has 40 citations and is from a highest quality peer-reviewed journal.
Artifacts
Citations
- mcdaniel2016combinatorialhistonereadout pages 2-4
- drouin2010dsifandrna pages 1-2
- guan2023diversemodesof pages 1-2
- li2024intrinsicallydisorderedregions pages 1-4
- zhang2023structuralbasisfor pages 1-2
- ruan2016homodimericphddomaincontaining pages 2-3
- markert2023structureofthea pages 1-3
- li2023structureofhistone pages 1-3
- li2023dynamicandmultivalent pages 7-10
- govind2010phosphorylatedpolii pages 1-2
- stephen2016expandingthefunction pages 86-91
- ruan2016homodimericphddomaincontaining pages 6-8
- lee2018combinatorialgeneticcontrol pages 1-2
- dong2023structuralbasisof pages 1-2
- ruan2016homodimericphddomaincontaining pages 3-4
- ruan2015nucleosomecontacttriggers pages 3-5
- ruan2016homodimericphddomaincontaining pages 1-2
- dong2023structuralbasisof pages 7-8
- mcdaniel2016combinatorialhistonereadout pages 4-6
- guan2023diversemodesof pages 5-6
- https://doi.org/10.1038/s41594-023-01121-5;
- https://doi.org/10.1074/jbc.m115.703637
- https://doi.org/10.1371/journal.pgen.1001173;
- https://doi.org/10.1016/j.molcel.2010.07.003;
- https://doi.org/10.1038/s41594-023-01121-5
- https://doi.org/10.1074/jbc.m116.720193;
- https://doi.org/10.1038/s41586-023-06349-1;
- https://doi.org/10.1038/s41422-023-00884-2
- https://doi.org/10.1038/s41467-023-43968-8;
- https://doi.org/10.21203/rs.3.rs-4000909/v1
- https://doi.org/10.17615/shj3-8048
- https://doi.org/10.1038/s41422-023-00899-9
- https://doi.org/10.1074/jbc.m115.703637;
- https://doi.org/10.1534/g3.118.200589
- https://doi.org/10.1038/s41422-023-00869-1;
- https://doi.org/10.21203/rs.3.rs-2060494/v1
- https://doi.org/10.1038/s41422-023-00899-9;
- https://doi.org/10.1038/s41586-023-06349-1
- https://doi.org/10.1038/s41422-023-00869-1
- https://doi.org/10.1038/s41467-023-43968-8
- https://doi.org/10.1074/jbc.m115.703637,
- https://doi.org/10.1371/journal.pgen.1001173,
- https://doi.org/10.1038/s41594-023-01121-5,
- https://doi.org/10.1074/jbc.m116.720193,
- https://doi.org/10.1016/j.molcel.2010.07.003,
- https://doi.org/10.1534/g3.118.200589,
- https://doi.org/10.1038/s41586-023-06349-1,
- https://doi.org/10.1038/s41422-023-00884-2,
- https://doi.org/10.1016/j.celrep.2014.12.027,
- https://doi.org/10.1038/s41467-023-43968-8,
- https://doi.org/10.1038/s41422-023-00899-9,
- https://doi.org/10.21203/rs.3.rs-4000909/v1,
- https://doi.org/10.1038/s41422-023-00869-1,
- https://doi.org/10.21203/rs.3.rs-2060494/v1,
- https://doi.org/10.17615/shj3-8048,
Perplexity
(RCO1-deep-research-perplexity.md)
Perplexity
(RCO1-deep-research-perplexity.md)RCO1: A Critical Regulatory Subunit of the Rpd3S Histone Deacetylase Complex in Saccharomyces cerevisiae
RCO1 (YMR075W) encodes a 684-amino acid transcriptional regulatory protein in the model organism Saccharomyces cerevisiae that functions as an essential subunit of the Rpd3S histone deacetylase (HDAC) complex[2][4]. This protein plays a pivotal role in maintaining proper chromatin structure and suppressing spurious transcription within gene bodies through its dual capacity to recognize histone post-translational modifications and to facilitate histone deacetylation by the catalytic Rpd3 subunit. Recent structural and biochemical analyses have revealed that Rco1 exists as a homodimer within the Rpd3S complex, with both copies contributing essential functions that cannot be compensated by a single copy[13][32]. The protein contains several critical domains including a plant homeobox (PHD) domain and a Sin3-interacting domain (SID), which together orchestrate the recognition of nucleosomes methylated at histone H3 lysine 36 (H3K36me) and facilitate proper positioning of the deacetylase for substrate targeting[7][14][17]. Through its interactions with the chromodomain-containing protein Eaf3, the scaffold protein Sin3, and the catalytic Rpd3 subunit, Rco1 enables the Rpd3S complex to deacetylate histone H3 and H4 tails specifically in the context of transcribed genes, thereby maintaining chromatin integrity and preventing the initiation of cryptic transcripts from internal promoters[16][25][29][40]. This comprehensive analysis examines the structure, function, molecular mechanisms, and biological significance of RCO1, drawing on recent structural biology studies, biochemical analyses, and genomic evidence to elucidate how this protein contributes to fundamental processes of chromatin regulation and transcriptional control in eukaryotes.
Overview of RCO1 Protein Structure and Domain Architecture
The RCO1 gene encodes a transcriptional regulatory protein that serves multiple architectural and recognition functions within the Rpd3S complex[2][4][8][23]. The protein spans 684 amino acids and contains several highly conserved functional domains that have been preserved throughout eukaryotic evolution[2][34]. The most prominent of these domains is the plant homeobox (PHD) domain, classified as a zinc finger motif that belongs to the FYVE/PHD zinc finger superfamily (IPR011011)[31][34]. This PHD domain functions as a module for recognizing post-translational modifications on histone proteins, though with some important caveats regarding its specificity that have emerged from recent structural and biochemical studies[14][17]. Beyond the PHD domain, Rco1 contains a Sin3-interacting domain (SID) that mediates critical protein-protein interactions within the complex and contributes to allosteric regulation of the complex's histone binding properties[14][40][37]. The structural analyses have revealed that the SID domain not only serves to anchor Eaf3 within the complex but also allosterically activates the chromodomain of Eaf3, thereby enhancing its specificity for H3K36-methylated histones[40][37].
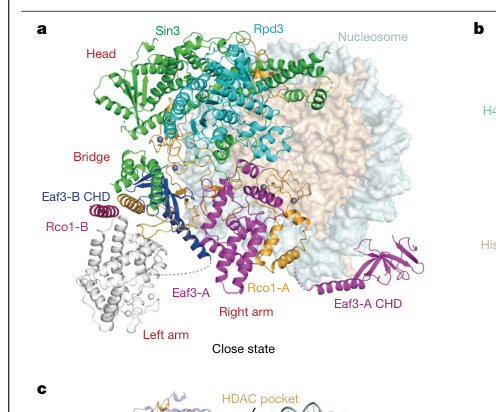
Recent cryo-EM structures determined at high resolution have provided unprecedented insight into the precise three-dimensional architecture of Rco1 within the Rpd3S complex[7][29][41][49]. The protein forms an extended scaffold structure that makes extensive contacts with multiple subunits of the complex, including direct interactions with the catalytic Rpd3 deacetylase, the Sin3 scaffold protein, and the Eaf3 chromodomain protein[7][29][41]. The N-terminal region of Rco1, spanning residues 33-66, binds directly to the Sin3 base to form an elongated scaffold that serves as a structural foundation for the entire complex[7][29][41]. This N-terminal region is connected to the PHD-SID domain (residues 260-374), which contains both the plant homeobox domain and the Sin3-interacting domain[7][29][41]. Additionally, Rco1 contains a conserved acidic patch-interacting motif (AIM) located at residues 377-397 that undergoes conformational changes upon nucleosome contact to facilitate interactions with the histone H2A-H2B dimer acidic patch on the nucleosome surface[7][29][41]. The C-terminal region of Rco1 consists of a long helix that interacts with the equivalent helical segment of the second copy of Rco1 within the complex, maintaining the homodimeric architecture essential for Rpd3S function[7][29][41][32].
The Rpd3S Complex Architecture and Rco1's Role as a Central Hub
The Rpd3S histone deacetylase complex represents one of two major enzymatic forms of the Rpd3 HDAC, the other being the larger Rpd3L complex[7][8][11][29]. The five-subunit Rpd3S complex contains unique subunits that are not found in Rpd3L, most notably Rco1, along with three core subunits (Rpd3, Sin3, and Ume1) that are shared between both complexes, and an Eaf3 subunit that is also a component of the NuA4 histone acetyltransferase complex[7][11][29]. Remarkably, biochemical and genetic studies have established that the Rpd3S complex contains two copies of the Rco1 protein, and both copies are essential for full functionality of the complex[13][32][37][44]. This discovery was unexpected and has important implications for understanding the evolutionary origin and functional capacity of the complex. The finding that Rpd3S in budding yeast contains two copies of Rco1 is consistent with observations that the fission yeast orthologue of Rpd3S contains two related PHD domain-containing subunits, Cph1 and Cph2, both of which are Rco1 orthologs, suggesting that the presence of two copies of this protein may represent a conserved feature across different fungal species[13][32][44].
The structural organization of the complex places Sin3 as the major scaffold subunit, directly linking the catalytic Rpd3 to the Rco1/Eaf3 heterodimer[7][13][29][32]. Within this architecture, Rco1 functions as a critical interaction hub that coordinates multiple layers of complex assembly and function[13][32]. The first copy of Rco1 (designated Rco1A in structural studies) makes the most extensive contacts with the other complex components, while the second copy (Rco1B) is integrated into the complex through helical interactions with the C-terminal region of Rco1A and through its interaction with Eaf3[7][29][41]. The interaction between Rco1 and Eaf3 is particularly critical, as Rco1 is required for the incorporation of Eaf3 into the complex, whereas omission of Eaf3 only minimally impacts the association of Rco1 with the remaining complex subunits[13][32][37]. This hierarchical dependence suggests that Rco1 plays a scaffolding role that is essential for building the complete nucleosome recognition module of the complex. The SID domain of Rco1 directly binds to the MRG domain of Eaf3, forming one of the two "MRG-PHD arms" that are characteristic features of the Rpd3S complex structure[7][29][41].
Chromatin Recognition Through H3K36me and Multivalent Interactions
The Rpd3S complex achieves remarkable specificity in recognizing methylated nucleosomes through a coordinated interaction between two of its subunits: Eaf3 and Rco1[14][17][25][40]. The coding regions of actively transcribed genes in Saccharomyces cerevisiae are highly marked with histone H3 lysine 36 trimethylation (H3K36me3), which is catalyzed by the Set2 methyltransferase that travels with RNA polymerase II during transcription elongation[25][49][58]. This H3K36me3 mark is specifically recognized by the chromodomain of Eaf3, which can bind to H3K36 monomethylated (H3K36me1), dimethylated (H3K36me2), and trimethylated (H3K36me3) substrates[14][17]. However, the chromodomain of Eaf3 exhibits relatively low affinity for histone peptides in isolation and shows limited specificity, binding with millimolar-range affinity that would be insufficient for robust in vivo recognition[14][17]. This apparent paradox is resolved through the allosteric activation mechanism provided by the PHD domain of Rco1. The PHD domain of Rco1 enhances the overall affinity of the Rpd3S complex for H3K36-methylated nucleosomes through a modification-independent nucleosomal binding mechanism[14][17][25][37]. This enhancement occurs through interactions with nucleosomal DNA and with histone proteins at sites distinct from the H3K36 methylation recognition site itself[7][14][17][29].
The mechanism by which Rco1 allosterically activates Eaf3 has been revealed through extensive biochemical and structural studies[14][40][37]. The Sin3-interacting domain of Rco1 binds directly to the MRG domain of Eaf3 and allosterically stimulates the binding of Eaf3's chromodomain to H3K36-methylated peptides, increasing both the affinity and specificity of recognition[14][40][37]. This allosteric activation suggests that the ability to read post-translational modifications on histones can be a crucial regulatory step for chromatin-modifying enzymes, allowing for temporal and spatial control of enzymatic activity. The PHD domain of Rco1 itself has multiple roles in nucleosome recognition. First, it directly contacts nucleosomal DNA and histone proteins at multiple sites, including the H3 α1L1 elbow region, one of the emerging hotspots for chromatin protein binding that is shared by many different chromatin-associated factors[7][22][29][33]. Second, the PHD domain interacts with the extra-nucleosomal DNA linkers flanking the nucleosome, utilizing these interactions to help guide the complex into proper positioning relative to the histone octamer[7][29]. Third, the conserved acidic patch-interacting motif (AIM) of Rco1 participates in interactions with the nucleosomal acidic patch, a surface composed of residues from histones H2A and H2B that serves as a docking interface for many chromatin proteins[7][29][41].
Functional Dissection of Rco1 Domains Reveals Multiple Essential Functions
The development of functional assays to test the importance of different regions of Rco1 has revealed surprising complexity in the protein's contributions to Rpd3S function[13][32][37]. When the Sin3-interacting domain (SID) is deleted from only one of the two copies of Rco1 within the complex, the nucleosome binding ability of Rpd3S is completely abolished without disrupting the integrity of the complex[13][32]. This indicates that both functional copies of the SID domain are required for robust nucleosome engagement. In contrast, deletion of the PHD domain from only one copy of Rco1 modestly reduces the binding of Rpd3S to mononucleosomes and has little effect on dinucleosome binding[13][32], suggesting that there is some redundancy in PHD domain function between the two copies. However, the combined deletion of both SID regions or loss of both PHD domains results in complete loss of nucleosome binding activity, confirming that each domain from each copy contributes to the overall nucleosome recognition capacity of the complex[13][32].
Beyond the known chromatin recognition interfaces, the N-terminal and C-terminal regions of Rco1 also play critical roles in Rpd3S function[13][32][44]. Truncation mutants that lack either the N-terminal region (residues 1-32) or portions of the C-terminal region (beyond residue 480) cause a marked cryptic transcription phenotype even when these mutant proteins are expressed at levels comparable to wild-type Rco1 and do not disrupt the interaction of Eaf3 with the complex[13][32][44]. This unexpected finding indicates that the N-terminal and C-terminal regions of Rco1 contribute to Rpd3S function through mechanisms beyond simple structural scaffolding. These regions may contribute to proper positioning of the Rpd3 catalytic subunit relative to histone substrates, facilitate conformational changes upon nucleosome contact, or enable interactions with other chromatin factors that enhance Rpd3S activity in vivo. The fact that all tested truncated Rco1 mutants could still dimerize with full-length Rco1, suggesting that Rco1 dimerization is mediated through multiple regions of the protein rather than a single localized interface[13][32][44], further underscores the complexity of Rco1's structural contributions to the complex.
Nucleosome Deacetylation and Cryptic Transcription Suppression
The primary biological function of the Rpd3S complex, which depends critically on proper Rco1 function, is to suppress the initiation of cryptic transcripts that would otherwise initiate from internal promoters within the bodies of actively transcribed genes[16][25][29][40][49][58]. The mechanism by which this suppression occurs involves the coordinated action of Set2 methylation and Rpd3S deacetylation to maintain a hypoacetylated chromatin state over gene bodies[25][58]. Set2 is an RNA polymerase II-associated histone methyltransferase that catalyzes the co-transcriptional methylation of H3K36, generating mono-, di-, and trimethylated H3K36 in a degree-dependent manner as the polymerase moves through the gene body[25][58]. These H3K36me marks are recognized by the Rpd3S complex, which is recruited to the coding region through interactions with the phosphorylated C-terminal domain (CTD) of RNA polymerase II[25][58]. Once recruited, the Rpd3 deacetylase removes acetyl groups from histone lysine residues, maintaining a hypoacetylated state that is incompatible with the recruitment of transcription initiation machinery to internal promoter regions[25][29][49][58].
Cryo-EM structures of Rpd3S bound to nucleosome core particles have revealed multiple distinct functional states of the complex that shed light on how it achieves efficient deacetylation while also reorganizing chromatin structure[7][29][49]. In the H3/H4 deacetylation state, the active site of the Rpd3 subunit is positioned to access acetylated lysine residues on the N-terminal tails of H3 and H4 histones[7][29][49]. The complex utilizes a conserved basic surface on Sin3 to navigate through nucleosomal DNA, guided by its interactions with H3K36 methylation and the extra-nucleosomal DNA linkers[7][29][49]. In addition to its canonical deacetylase activity, Rpd3S also functions as an HDAC-independent chromatin stabilizer that prevents nucleosome eviction by the chromatin remodeler RSC, a function that is distinct from its deacetylase activity[7][29][49]. After deacetylating nucleosome substrates, Rpd3S enters an alternative deacetylation state in which it can sample histone tails at different positions relative to the nucleosome, and finally transitions to a linker-tightening state in which it reconfigures the extra-nucleosomal DNA linkers[7][29][49]. In this linker-tightening state, the exit DNA linker bends more sharply (with an angle of 37 degrees compared to 27 degrees in the H3K9 deacetylation state), effectively compacting the chromatin structure[7][29][49].
Role of Core Promoter Strength in Regulating Cryptic Transcription
An important recent discovery regarding Rpd3S function is that the activity of internal cryptic promoters within gene bodies is strongly repressed by high core promoter activity but becomes activated when core promoter activity is downregulated[16][30]. This finding suggests a dynamic interplay between the main promoter of a gene and internal cryptic promoters, with strong core promoter activity directly repressing internal cryptic initiation even in mutants lacking Set2 or Rco1[16][30]. The molecular basis for this repression by high core promoter activity appears to involve competitive recruitment of general transcription factors. When the core promoter is very strong, it preferentially recruits transcription factors and maintains high levels of transcription initiation machinery at the main promoter, thereby limiting the availability of these factors for binding to internal cryptic promoters[16][30]. When core promoter activity is weakened, internal cryptic promoters become more accessible to transcription factors and can be activated by the removal of Rpd3S-mediated chromatin repression. This finding has important implications for understanding how different genes are differentially vulnerable to cryptic transcription activation in response to mutations in transcription elongation factors. Genes like STE11, PCA1, and FLO8, which have intrinsically weak promoters, are highly sensitive to cryptic transcription activation when Set2 or Rco1 are deleted[16][30]. In contrast, genes with strong core promoters show minimal cryptic transcription even in the absence of functional Rpd3S[16][30].
Interaction with Linker Histone Hho1 for Gene Silencing
Recent structural studies have revealed that Rpd3S does not work in isolation but rather acts in concert with the linker histone Hho1 (the Saccharomyces cerevisiae ortholog of mammalian H1) to promote gene silencing[7][29][49]. The cryo-EM structure of an Rpd3S-nucleosome-Hho1 complex shows that Hho1 binds to the nucleosome at the canonical dyad binding site where linker histones typically bind[7][29][49]. This binding of Hho1 occurs in the context of Rpd3S-mediated nucleosome deacetylation and involves reconfiguration of the extra-nucleosomal DNA linkers[7][29][49]. The combined results suggest a transition from Rpd3S-mediated cryptic transcription repression to Hho1-mediated chromatin compaction, potentially providing a mechanism by which transient Rpd3S deacetylation activity can be converted into stable repressive chromatin structure through subsequent Hho1 binding and higher-order chromatin compaction[7][29][49]. The hho1Δrpd3Δ double mutant results in additive derepression of early meiotic gene transcription, implying that Rpd3 works together with Hho1 to stabilize the repressive chromatin structure established by Rpd3L and Rpd3S, suggesting that this partnership between the deacetylase complex and linker histones extends to meiotic gene regulation[7][29][49].
Evolutionary Conservation of Rco1 Across Eukaryotes
Phylogenetic analyses have revealed that Rco1 orthologs are widely conserved throughout eukaryotic organisms, though with important variations in the number of copies present in different species[13][32][44]. All species within the budding yeast branch of fungal evolution carry only one RCO1 ortholog gene, yet the Rpd3S complex in these organisms contains two copies of the Rco1 protein, indicating that the homodimer is assembled from a single gene through expression of multiple copies that associate during complex assembly[13][32][44]. In contrast, the fission yeast branch appears to have undergone a gene duplication event, resulting in two genes encoding the Rco1 counterpart, designated cph1 and cph2, with one of them containing a shorter N-terminal region and a less-conserved PHD domain[13][32][44][47]. This evolutionary divergence suggests that Rpd3S complexes in different fungal species might contain either two copies of a single Rco1 protein or two closely related homologous proteins that have emerged through gene duplication and divergence[13][32][44]. In fission yeast, deletion of cph1 alone is sufficient to cause total Rpd3S defects, suggesting that both functional copies of the Rco1 counterpart are essential under physiological conditions[13][32][44]. The conservation of this homodimeric architecture across different fungal lineages, despite the different evolutionary routes (duplication within a single gene versus duplication of the gene itself) that have led to it, underscores the fundamental importance of the two-copy requirement for Rpd3S function.
Beyond fungal organisms, homologs of Rco1 have been identified in higher eukaryotes, where they may function in related but more complex chromatin-modifying complexes. The human ortholog of Rco1 is known as MRG15 (MRG15 histone binding protein), which contains similar PHD domain architecture and participates in multiple chromatin-modifying complexes including Sin3-containing histone deacetylase complexes analogous to Rpd3S[15][40]. The evolutionary conservation of Rco1/MRG15 function across eukaryotes, combined with the conservation of the core Rpd3/HDAC1 catalytic activity and Sin3 scaffold architecture, indicates that the Set2-Rpd3S pathway and its regulatory mechanisms represent fundamental mechanisms for controlling transcription and chromatin structure that have been preserved throughout eukaryotic evolution.
Regulatory Interactions with Chromatin Remodeling Complexes
The function of Rco1-containing Rpd3S complex is integrated with that of chromatin remodeling enzymes that regulate nucleosome positioning and histone exchange. Studies have shown that the ISWI family remodeling enzyme Isw1 and the Chd1 remodeler both facilitate Rpd3S activity by spacing nucleosomes 30-40 base pairs apart, thereby allowing Rpd3S to bridge two adjacent nucleosomes[56]. The ability of Rpd3S to bind to two nucleosomes simultaneously through a dinucleosomal binding mode is a crucial feature of its activity, allowing it to target both H3K36me nucleosomes and their neighbors for deacetylation, enabling efficient stabilization of nucleosomes even when chromatin domains are not completely saturated by the methylation mark[56]. The RSC chromatin remodeling complex, in contrast, appears to antagonize Rpd3S function by promoting nucleosome remodeling and histone variant exchange, activities that are inhibited by the HDAC-independent chromatin stabilization functions of Rpd3S[7][56].
Additionally, the FACT complex (Facilitates Chromatin Transcription) interacts functionally with Rpd3S to regulate the assembly and disassembly of nucleosomes during transcription. The Spt6 protein, which is associated with FACT-like activities, works together with Rpd3S to repress cryptic transcription, and mutations in SPT6 produce cryptic transcription phenotypes very similar to those observed in rco1Δ and set2Δ mutants[16][30]. This functional overlap suggests that the maintenance of proper chromatin structure during transcription requires the coordinated action of nucleosome assembly factors, chromatin remodeling complexes, and histone-modifying enzymes, with Rco1-containing Rpd3S playing a central coordinating role.
Role in Transcription Elongation and RNA Polymerase II Regulation
Beyond its function in cryptic transcription suppression, the Rpd3S complex, through its dependence on Rco1 function, plays broader roles in regulating transcription elongation and RNA polymerase II activity. Nascent transcript sequencing (NET-seq) studies, which map the density of transcriptionally active RNA polymerases at nucleotide resolution, have revealed that deletion of rco1 increases antisense transcription originating from antisense promoters positioned opposite the 3' ends of genes[51]. These antisense transcripts have the same transcription start sites and the same lengths in rco1Δ strains as in wild-type strains, indicating that Rco1 acts at the initiation stage of antisense transcription and does not affect transcription termination[51]. This finding suggests that the primary function of the Rpd3S histone deacetylase complex, which critically depends on Rco1, is to enforce promoter directionality by preventing transcription initiation from promoters on both strands of the DNA within transcribed regions[51]. The fact that deletion of EAF3, another subunit of Rpd3S, mimics the increases in antisense transcription seen in the rco1Δ data confirms that the entire Rpd3S complex is required for this directional control function[51].
Contribution to Heterochromatin Maintenance and Silencing
While Rpd3S is primarily known for its function in suppressing cryptic transcription within gene bodies, the larger Rpd3L complex (which shares core subunits with Rpd3S but contains different accessory subunits and does not contain Rco1) plays distinct roles in heterochromatin formation and silencing at repetitive elements and silent loci[39]. However, Rpd3L and Rpd3S work together in some contexts to maintain proper chromatin structure. The Rpd3L complex is responsible for anti-silencing functions at heterochromatic loci, particularly silencers and boundaries between euchromatic and heterochromatic regions[39]. Deletion of RPD3L-specific subunits like PHO23, RXT2, and SDS3 enhances silencing at the silent mating type loci and telomeres, whereas deletion of RCO1 or EAF3, which are specific to Rpd3S, has little effect on silencing at these locations[39]. This functional specialization between Rpd3L and Rpd3S is consistent with their different subcellular and genomic localizations, with Rpd3S being recruited preferentially to coding regions through interactions with H3K36me and the elongating RNA polymerase II, while Rpd3L is recruited to promoters and regulatory regions through interactions with H3K4me3[29][39].
Implications for Understanding Chromatin Regulatory Complexity
The study of Rco1 and its role within the Rpd3S complex has contributed significantly to our understanding of how cells achieve precision and specificity in chromatin regulation despite the apparent simplicity of the histone modifications and chromatin-associated proteins involved. The Rco1 protein exemplifies several important principles of chromatin regulation that have emerged from recent structural and biochemical studies. First, the use of multivalent interactions involving multiple contact points between a chromatin-modifying enzyme and its nucleosomal substrate enables the achievement of high specificity and affinity even when individual interaction surfaces would have low affinity or specificity in isolation[7][14][17][22][33]. Second, the use of allosteric regulation, in which binding of one ligand to a protein alters the binding properties of distant sites, allows chromatin complexes to integrate multiple types of information (such as histone modifications and DNA sequences) to make regulatory decisions[14][40][37]. Third, the presence of multiple copies of key regulatory subunits within a single complex, as exemplified by the homodimeric Rco1 architecture within Rpd3S, provides redundancy and flexibility while maintaining the capacity for complex regulation[13][32][44].
The Rco1-containing Rpd3S complex also illustrates how chromatin-modifying enzymes are not isolated molecular machines but rather function as integral parts of larger networks of chromatin regulators that work together to coordinate DNA accessibility, transcription factor recruitment, and gene expression. The Set2-Rpd3S pathway integrates with nucleosome assembly and remodeling machinery, RNA processing factors, and other transcription regulatory proteins to ensure that the passage of RNA polymerase II through a gene body results in the proper combination of histone modifications, chromatin remodeling, and histone exchange necessary to maintain transcriptional fidelity while also preparing chromatin for potential future rounds of transcription or for entry into alternative chromatin states.
Conclusion: Rco1 as a Central Regulator of Chromatin Structure and Gene Expression
The RCO1 gene product represents a critical regulatory node in the control of chromatin structure and transcriptional fidelity in Saccharomyces cerevisiae. As an essential component of the Rpd3S histone deacetylase complex, Rco1 serves multiple functions that extend far beyond simple structural scaffolding. The protein contains multiple functional domains that collectively enable the Rpd3S complex to recognize H3K36-methylated nucleosomes with high specificity and affinity, to deacetylate histone tails in a position-specific manner, to suppress the initiation of cryptic transcripts from internal gene body promoters, and to coordinate with other chromatin-modifying enzymes to maintain proper chromatin structure throughout the transcribed regions of active genes. The homodimeric architecture of Rco1 within the Rpd3S complex, with both copies contributing essential functions, represents an elegant solution to the challenge of achieving the complex three-dimensional positioning and flexibility required for the enzyme to access histone tails projecting in different directions while also maintaining interaction with nucleosomal DNA and linker DNA regions. Recent structural biology studies have provided atomic-resolution models of Rco1 in complex with nucleosomal substrates, revealing multiple distinct functional states and clarifying the mechanisms by which allosteric regulation enables the integration of histone modification signals with chromatin recognition and enzyme activity.
The evolutionary conservation of Rco1 orthologs and of the Rpd3S complex architecture across eukaryotes indicates that the mechanisms and principles governing Rco1 function represent fundamental solutions to the problem of maintaining transcriptional fidelity and chromatin integrity that have been preserved throughout the evolution of eukaryotic organisms. The functional studies demonstrating that both copies of Rco1 are required for full Rpd3S activity, combined with findings that the N-terminal, central, and C-terminal regions of the protein each contribute distinct functions, suggest that Rco1 represents a highly optimized protein whose structure and function have been refined through millions of years of evolution to achieve the precise balance between transcriptional activation and repression necessary for proper cellular function. Future studies employing advanced structural biology techniques, quantitative proteomic analyses, and single-molecule biochemical assays will likely continue to reveal additional layers of regulation and function associated with Rco1 and its partners in the Rpd3S complex, potentially uncovering new therapeutic targets for diseases in which chromatin regulation is dysregulated.
Citations
- https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=855097
- https://www.uniprot.org/citations/17142463
- https://thecellvision.org/cyclopsgenelist/?sort=-aliases
- https://www.uniprot.org/uniprotkb/Q04779/entry
- https://ibdc.dbtindia.gov.in/isda/protein_detail/Q04779/
- https://dspace.library.uu.nl/bitstream/1874/254111/1/lenstra.pdf
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10542350/
- https://www.yeastgenome.org/locus/S000004680
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8045735/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC206603/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4777872/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4359074/
- https://portlandpress.com/biochemj/article/475/24/3921/49835/Co-repressor-co-activator-and-general
- https://academic.oup.com/nar/article/49/14/8097/6329575
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2582589/
- https://www.yeastgenome.org/complex/S000218002/literature
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4834262/
- https://elifesciences.org/articles/78944
- https://rupress.org/jcb/article/212/6/633/38510/Strategies-to-regulate-transcription-factor
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8648869/
- https://www.yeastgenome.org/locus/S000004680/go
- https://genesdev.cshlp.org/content/20/6/660
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3767924/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2944879/
- https://www.tandfonline.com/doi/full/10.1080/21541264.2016.1246076
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8373055/
- https://www.ebi.ac.uk/interpro/entry/interpro/IPR011011
- https://maayanlab.cloud/Harmonizome/gene_set/Zinc+finger,+RING$slash$FYVE$slash$PHD-type/InterPro+Predicted+Protein+Domain+Annotations
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3068686/
- https://academic.oup.com/nar/article/53/14/gkaf734/8219896
- https://elifesciences.org/articles/94869
- https://academic.oup.com/nar/article/37/11/3699/1089232
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2781707/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10531175/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9769939/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11704871/
- https://www.pombase.org/gene/SPAC2F7.07c
- https://onlinelibrary.wiley.com/doi/10.1111/brv.12605
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6212844/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3880149/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3320549/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4004355/
- https://www.yeastgenome.org/locus/S000004680/expression
- https://genesdev.cshlp.org/content/28/10/1029.full
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4099280/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3001064/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5854953/
- https://d-scholarship.pitt.edu/13326/1/Hainer_2012_1.pdf
- https://journals.plos.org/plosgenetics/article?id=10.1371%2Fjournal.pgen.1000478