Deep Research
Falcon
(SNF5-deep-research-falcon.md)
Falcon
(SNF5-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Saccharomyces cerevisiae SNF5 (UniProt P18480) — Functional Annotation and Current Evidence
0) Target verification (critical disambiguation)
Requested target: UniProt P18480, S. cerevisiae (S288c) Snf5 (synonyms: SWI10, TYE4; locus YBR289W), a SWI/SNF chromatin-remodeling complex subunit.
Verification status with retrieved literature: The retrieved sources consistently discuss budding-yeast Snf5 as a core SWI/SNF subunit, conserved with metazoan SMARCB1/INI1/BAF47, and involved in SWI/SNF chromatin remodeling (including nucleosome acidic-patch engagement and transcription-factor interactions). (kuwahara2023recentinsightsinto pages 3-5, eustermann2024energydrivengenomeregulation pages 1-3, wendegatz2024transcriptionalactivationdomains pages 5-6)
Limitation: None of the retrieved full-text excerpts explicitly list the UniProt accession P18480, the systematic locus YBR289W, or synonyms SWI10/TYE4. Therefore, this report’s biological conclusions are supported by the yeast Snf5 literature, but the database-level identifier mapping (P18480 ↔ YBR289W ↔ SNF5/SWI10/TYE4) could not be independently re-confirmed from the current paper excerpts alone.
1) Key concepts and definitions (current understanding)
1.1 SWI/SNF chromatin remodeling complexes
SWI/SNF-family remodelers are ATP-dependent, multi-subunit chromatin remodeling machines that alter nucleosome–DNA contacts to regulate DNA accessibility; canonical activities include nucleosome sliding and histone ejection (eviction). (eustermann2024energydrivengenomeregulation pages 1-3, chen2023mechanismofaction pages 2-6)
1.2 The nucleosome acidic patch
The H2A–H2B acidic patch is a negatively charged nucleosome surface that serves as a common interaction hub for chromatin factors. In SWI/SNF-family remodelers, acidic-patch engagement helps stabilize remodeler–nucleosome binding and can contribute to coupling ATP hydrolysis to productive remodeling. (chen2023mechanismofaction pages 2-6, eustermann2024energydrivengenomeregulation pages 1-3)
1.3 NBL, finger helix, and SnAc: terms relevant to Snf5 function
A recent mechanistic synthesis organizes SWI/SNF nucleosome engagement into motor- and substrate-recruitment components and highlights two acidic-patch–recognition elements:
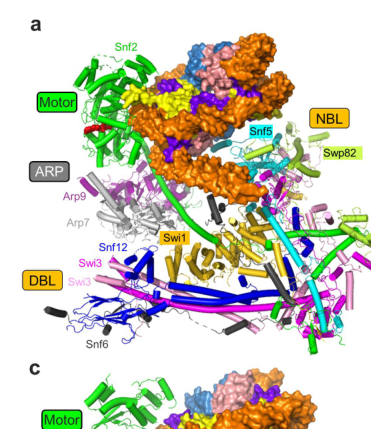
- Nucleosome-binding lobe (NBL): in budding-yeast SWI/SNF, the NBL is described as being mainly formed by Snf5, making Snf5 a principal nucleosome-contacting scaffold within the complex. (chen2023mechanismofaction pages 2-6)
- Snf5 “finger helix” (FH): a C-terminal helix protruding from the NBL that packs against the H2A–H2B surface and binds the acidic patch via multiple arginine residues. (chen2023mechanismofaction pages 2-6)
- SnAc domain: a conserved, polybasic extension of the SWI/SNF ATPase motor (Snf2) that also recognizes the acidic patch and is described as coupling ATP hydrolysis to nucleosome sliding. (chen2023mechanismofaction pages 2-6)
2) Molecular function of Snf5 (primary function and mechanism)
2.1 Snf5 is a core structural/regulatory subunit, not an enzyme
Across yeast and metazoans, Snf5/SMARCB1/INI1 is repeatedly framed as a core subunit that contributes to SWI/SNF complex integrity and nucleosome engagement rather than providing catalytic ATPase activity (which in yeast is carried by Snf2). (kuwahara2023recentinsightsinto pages 3-5, eustermann2024energydrivengenomeregulation pages 1-3)
2.2 Direct nucleosome engagement via the acidic patch
A key mechanistic advance synthesized in a 2023 review is that budding-yeast Snf5 largely composes the NBL and uses its finger helix to bind the nucleosome’s H2A–H2B acidic patch through multiple arginine residues. (chen2023mechanismofaction pages 2-6)
A specific residue is highlighted: Arg669 in yeast Snf5 is described as the canonical “arginine anchor”, equivalent to Arg370 in SMARCB1. (chen2023mechanismofaction pages 2-6)
In the same framework, mutations in the finger helix reduce remodeling activity in vitro and reduce fitness in vivo, linking this nucleosome-surface contact to biologically relevant remodeling output. (chen2023mechanismofaction pages 2-6)
A 2024 authoritative review further supports the assignment of Snf5 as a “proximal acidic patch” binder within yeast SWI/SNF and places SWI/SNF’s characteristic activities as nucleosome sliding and histone ejection. (eustermann2024energydrivengenomeregulation pages 1-3)
2.3 Complex architecture context (where Snf5 sits)
Structural/architecture descriptions of yeast SWI/SNF emphasize modular organization (ATPase/motor module; ARP module; body/substrate recruitment components) and explicitly label Snf5 as part of the complex architecture. (amaris2022structuralinvestigationof pages 34-44)
The figure panels retrieved from Chen et al. 2023 visually support the overall yeast SWI/SNF architecture and the acidic-patch engagement by the Snf5/SMARCB1 finger helix. (chen2023mechanismofaction media 799148c2, chen2023mechanismofaction media ad429c0b, chen2023mechanismofaction media 79a39cc8)
3) Biological processes and pathways (with emphasis on precise roles)
3.1 Transcriptional activation via activator interactions (recruitment/coupling)
A 2024 yeast-focused study demonstrates that Snf5 can bind transcriptional activation domains (TADs) (tested alongside other SWI/SNF subunits). In the context of phospholipid biosynthetic gene regulation, SWI/SNF subunits including Snf5 were reported to bind Ino2 TADs, and the study also discusses interactions with other activators such as Gcn4. (wendegatz2024transcriptionalactivationdomains pages 5-6)
This supports a mechanistic view where Snf5 contributes to SWI/SNF’s transcriptional effects by acting as a protein–protein interaction platform for activator-driven recruitment and/or for coupling activator binding to remodeling. (wendegatz2024transcriptionalactivationdomains pages 5-6)
3.2 Direct transcriptional repression via LUTI-based transcriptional interference (2024 development)
A major recent development in yeast is evidence that Swi/Snf can directly repress transcription in vivo through nucleosome remodeling, rather than repression being only an indirect consequence of activation elsewhere.
In Molecular Cell (Aug 2024), Morse et al. used a genetic selection for “LUTI escape” mutants and found all validated escape mutations mapped to genes encoding Swi/Snf subunits; they conclude this provides “conclusive evidence” that Swi/Snf can directly repress transcription through nucleosome remodeling downstream of an active TSS, thereby interfering with a downstream CDS-proximal promoter. (morse2024swisnfchromatinremodeling pages 5-8)
At the endogenous HNT1 locus during DTT-induced stress, Swi/Snf recruitment/remodeling was fast and quantitative:
* HNT1LUTI induction within 5 minutes, increasing to approximately ~3-fold by 30 minutes.
* At ~30 minutes, the proximal isoform HNT1PROX is described as nearly fully silenced.
* Proximal promoter chromatin changes were measurable: NDR nucleosome occupancy increased ~1.9-fold (p = 0.0445) on average in the repression state, and -1/+1 nucleosomes became “more fuzzy” (a measure of positioning/heterogeneity). (morse2024swisnfchromatinremodeling pages 15-19)
Although these data were tracked using Snf2 ChIP and nucleosome mapping (rather than Snf5 ChIP specifically), the mechanism is attributed to the Swi/Snf complex’s remodeling activity, of which Snf5 is a core nucleosome-binding subunit. (morse2024swisnfchromatinremodeling pages 15-19, chen2023mechanismofaction pages 2-6)
Morse et al. also defined a 250-gene Swi/Snf target set using transcriptomic and occupancy filters (downregulated in snf2Δ, Snf2-bound loci, and additional exclusions), providing a concrete genome-scale handle on Swi/Snf-regulated genes in their framework. (morse2024swisnfchromatinremodeling pages 19-22)
3.3 Metabolic pathway control and sulfur metabolism (2023 development)
A 2023 Nucleic Acids Research study emphasizes that loss of Swi/Snf can paradoxically lead to activation of metabolic genes under repressing conditions, especially sulfur metabolism (MET) genes, and a cysteine-deficient phenotype despite growth in rich medium. (church2023theswisnfchromatin pages 1-2)
The study reports that this correlates with a global redistribution of the transcription factor Met4 and widespread perturbations in sulfur metabolic transcription; RNA-seq comparisons of snf2Δ and snf5Δ were performed and pathway enrichment identified sulfur/MET pathways among the most enriched upregulated pathways. (church2023theswisnfchromatin pages 2-3)
This illustrates how SWI/SNF (and by extension its Snf5 subunit) participates in transcriptional programs that control metabolic state, not merely “general transcription.” (church2023theswisnfchromatin pages 2-3, church2024theswisnfchromatin pages 1-2)
4) Cellular localization and where Snf5 acts
4.1 Subcellular site of action: nucleus/chromatin
Functionally, Snf5 is a chromatin-associated factor by virtue of its role in SWI/SNF remodeling of nucleosomes and direct nucleosome-surface binding (acidic patch). (chen2023mechanismofaction pages 2-6)
4.2 Genomic targeting (promoters, stress induction, and co-transcriptional remodeling)
Direct kinetic evidence for S. cerevisiae Swi/Snf recruitment to a locus was shown at HNT1 by tracking Snf2:
* Snf2 occupancy at HNT1 increased within 5 minutes of stress, tracking LUTI induction.
* Initiation at the distal LUTI promoter was sufficient for initial recruitment, but productive elongation was required for downstream occupancy and remodeling at the proximal promoter; mutants that truncated transcription reduced Snf2 binding at the proximal region and blocked remodeling. (morse2024swisnfchromatinremodeling pages 15-19)
Snf5-specific genome-wide occupancy/localization was not retrieved in the current excerpts; therefore, Snf5 targeting is best stated as: Snf5 is expected to be nuclear/chromatin-associated as a SWI/SNF core subunit, and recruitment occurs at promoter regions where the complex is directed by activators/histone signals, consistent with demonstrated Snf5 binding to activator TADs. (wendegatz2024transcriptionalactivationdomains pages 5-6, morse2024swisnfchromatinremodeling pages 15-19)
5) Recent developments (prioritizing 2023–2024)
5.1 2023: Mechanistic consolidation of acidic patch binding by Snf5
The 2023 mechanistic review formalizes a model where SWI/SNF uses two acidic-patch recognition elements—Snf5’s finger helix (NBL) and Snf2’s SnAc domain—to bind an otherwise intact nucleosome while the motor engages DNA at SHL2, supporting ATP-coupled remodeling outputs (sliding/ejection). (chen2023mechanismofaction pages 2-6)
5.2 2023: Metabolic control and sulfur pathway rewiring
The 2023 NAR study adds a concrete, pathway-level example of how SWI/SNF loss can yield systemic metabolic consequences (cysteine deficiency; altered sulfur metabolism transcription) mediated through transcription-factor redistribution (Met4). (church2023theswisnfchromatin pages 2-3)
5.3 2024: Direct repression mechanism (LUTI transcriptional interference)
The 2024 Molecular Cell study provides strong genetic and genomic evidence that Swi/Snf can mediate direct repression via co-transcriptional downstream remodeling that occludes a downstream promoter’s NDR, with quantified kinetics and chromatin effect sizes. (morse2024swisnfchromatinremodeling pages 15-19, morse2024swisnfchromatinremodeling pages 5-8)
5.4 2024: Expert syntheses and “big picture” interpretation
Two 2024 reviews contextualize yeast SWI/SNF as the foundational model for a broader class of remodelers and highlight disease and targeting implications:
* SWI/SNF as an energy-driven genome regulator; Snf5 specifically annotated as proximal acidic patch binder. (eustermann2024energydrivengenomeregulation pages 1-3)
* SWI/SNF framed as a critical regulator of metabolism, emphasizing conserved chromatin–metabolism coupling and therapeutic vulnerabilities in SWI/SNF-mutant contexts. (church2024theswisnfchromatin pages 1-2)
6) Current applications and real-world implementations
6.1 Yeast Snf5/SWI/SNF as a conserved model for human SWI/SNF vulnerabilities
A 2024 metabolism-focused review argues that mechanistic understanding from yeast SWI/SNF continues to inform mammalian SWI/SNF biology and cancer vulnerabilities, emphasizing strategies such as synthetic lethality for tumors with SWI/SNF loss-of-function. (church2024theswisnfchromatin pages 6-8)
This is an important “real-world implementation” pathway: yeast is used to derive mechanistic principles (e.g., chromatin–metabolism coupling) that translate into candidate metabolic liabilities in SWI/SNF-deficient disease contexts. (church2024theswisnfchromatin pages 2-4)
6.2 Engineering transcriptional control via activator–remodeler interactions (conceptual synthetic biology application)
The 2024 Current Genetics study and associated discussion highlight that transcription factors can bind different remodeler subunits (including Snf5) through activation domains, and discuss models such as activator-specific binding patterns and phase-separation/condensate formation upon TAD–ABD binding. Such mechanisms can guide engineered strategies to tune recruitment of chromatin remodelers in synthetic transcription systems, even if the paper itself is not a deployment study. (wendegatz2024transcriptionalactivationdomains pages 1-2, wendegatz2024transcriptionalactivationdomains pages 5-6)
7) Expert opinions / authoritative perspectives
- A 2024 review emphasizes that SWI/SNF’s role as a regulator of metabolic transcription in mammals has recently emerged and that understanding of SWI/SNF as a metabolic regulator “continues to evolve,” motivating further mechanistic work; yeast remains positioned as an informative model for chromatin–metabolism connections. (church2024theswisnfchromatin pages 8-9)
- A 2024 Nature Reviews Genetics synthesis emphasizes that remodellers show strong biological specificity and dosage sensitivity, are widely mutated in disease, and that active efforts are underway to develop therapeutic avenues targeting remodellers; yeast SWI/SNF is framed as the foundational discovery enabling this field. (gourisankar2024contextspecificfunctionsof pages 1-3)
8) Key statistics and quantitative findings (recent studies)
- Snf5 mechanistic statistic (structure-function): Snf5 finger helix mutations reduce remodeling activity in vitro and reduce fitness in vivo (qualitative outcome; numerical values not provided in the excerpt). (chen2023mechanismofaction pages 2-6)
- LUTI repression quantitative chromatin effects (2024): NDR occupancy at proximal promoter increased ~1.9-fold (p = 0.0445); HNT1LUTI increased ~3-fold by 30 min; recruitment/remodeling detectable within 5 min. (morse2024swisnfchromatinremodeling pages 15-19)
- Genome-scale set definition (2024): Morse et al. defined 250 Swi/Snf target genes based on transcriptomic and occupancy filters. (morse2024swisnfchromatinremodeling pages 19-22)
- Cancer association summary statistic (2024 reviews): BAF/SWI/SNF complexes are reported as mutated in ~20% of human cancers (used for translational framing). (gourisankar2024contextspecificfunctionsof pages 27-28, church2024theswisnfchromatin pages 1-2)
- Yeast dependency estimate: A 2024 yeast-focused study cites an estimate that ~1% of yeast protein-coding genes are Swi-dependent (contextual statistic; details of derivation not in excerpt). (wendegatz2024transcriptionalactivationdomains pages 1-2)
9) Visual evidence (figure support)
A key structural depiction of yeast SWI/SNF architecture (with Snf5/NBL indicated) and the acidic-patch engagement concept (finger helix contact) is available from the retrieved figure crops. (chen2023mechanismofaction media 799148c2, chen2023mechanismofaction media ad429c0b, chen2023mechanismofaction media 79a39cc8)
10) Evidence map (for traceability)
| Aspect | Key points | Evidence (with citation IDs) | Publication (author, year, journal) | URL | Publication date/month if available |
|---|---|---|---|---|---|
| Identity / complex membership | Budding-yeast Snf5 is a core, non-ATPase subunit of the SWI/SNF ATP-dependent chromatin-remodeling complex; it is evolutionarily conserved with human SMARCB1/INI1 and contributes to complex integrity and function. | Snf5 is identified as a core SWI/SNF component in S. cerevisiae and homologous to SMARCB1/INI1; SWI/SNF is a large multi-subunit chromatin remodeler. (kuwahara2023recentinsightsinto pages 3-5, lampersberger2023geneticinteractorsof pages 37-41, lampersberger2023geneticinteractorsof pages 34-37, amaris2022structuralinvestigationof pages 34-44) | Kuwahara et al., 2023, Cancer Medicine; Lampersberger, 2023, Dissertation | https://doi.org/10.1002/cam4.6255 ; https://doi.org/10.17863/cam.93003 | Jun 2023; Jan 2023 |
| Mechanistic role | Snf5 mainly forms the nucleosome-binding lobe (NBL) of SWI/SNF and acts as a structural/regulatory subunit that helps couple nucleosome recognition to remodeling rather than serving as the ATPase. | Reviews describe Snf5 as the principal component of the NBL and a structural subunit within SWI/SNF family remodelers. (chen2023mechanismofaction pages 2-6, eustermann2024energydrivengenomeregulation pages 1-3) | Chen et al., 2023, Nucleus; Eustermann et al., 2024, Nature Reviews Molecular Cell Biology | https://doi.org/10.1080/19491034.2023.2165604 ; https://doi.org/10.1038/s41580-023-00683-y | Jan 2023; Dec 2024 |
| Nucleosome interaction | Snf5 engages the nucleosomal H2A-H2B acidic patch through its C-terminal finger helix; Arg669 is the canonical arginine anchor. This acidic-patch contact cooperates with the Snf2 motor and SnAc domain to support ATP-coupled nucleosome sliding/ejection. | Snf5 is annotated as a proximal acidic-patch binder; the finger helix packs against the acidic patch, and finger-helix mutations reduce remodeling in vitro and cell fitness in vivo. (eustermann2024energydrivengenomeregulation pages 1-3, chen2023mechanismofaction pages 2-6) | Eustermann et al., 2024, Nature Reviews Molecular Cell Biology; Chen et al., 2023, Nucleus | https://doi.org/10.1038/s41580-023-00683-y ; https://doi.org/10.1080/19491034.2023.2165604 | Dec 2024; Jan 2023 |
| Transcription regulation | Snf5 contributes to SWI/SNF-mediated transcriptional control by interacting with activator domains and participating in promoter chromatin remodeling. Recent yeast studies show Swi/Snf can directly repress as well as activate transcription, including via LUTI-based transcriptional interference and metabolic control. | Snf5 binds activator TADs such as Ino2/Gcn4; SWI/SNF mediates direct repression through co-transcriptional downstream remodeling and regulates sulfur metabolic genes. (wendegatz2024transcriptionalactivationdomains pages 5-6, morse2024swisnfchromatinremodeling pages 19-22, morse2024swisnfchromatinremodeling pages 15-19, morse2024swisnfchromatinremodeling pages 5-8, church2023theswisnfchromatin pages 1-2) | Wendegatz et al., 2024, Current Genetics; Morse et al., 2024, Molecular Cell; Church et al., 2023, Nucleic Acids Research | https://doi.org/10.1007/s00294-024-01300-x ; https://doi.org/10.1016/j.molcel.2024.06.029 ; https://doi.org/10.1093/nar/gkad711 | Sep 2024; Aug 2024; Aug 2023 |
| Quantitative findings | Recent quantified SWI/SNF effects in yeast include: 250 defined Swi/Snf target genes in Morse et al.; average 1.9-fold increase in NDR occupancy at the proximal promoter during HNT1 LUTI repression (p=0.0445); HNT1LUTI induced within 5 min and ~3-fold by 30 min; all 11 validated LUTI-escape mutations mapped to Swi/Snf genes. Church et al. found RNA-seq pathway enrichment of sulfur/MET genes in both snf2Δ and snf5Δ mutants. | Quantitative values and gene-set sizes come from recent genomic and reporter-based yeast studies on Swi/Snf-dependent repression/activation. (church2023theswisnfchromatin pages 2-3, morse2024swisnfchromatinremodeling pages 19-22, morse2024swisnfchromatinremodeling pages 15-19, morse2024swisnfchromatinremodeling pages 5-8) | Church et al., 2023, Nucleic Acids Research; Morse et al., 2024, Molecular Cell | https://doi.org/10.1093/nar/gkad711 ; https://doi.org/10.1016/j.molcel.2024.06.029 | Aug 2023; Aug 2024 |
| Applications / implications | Yeast Snf5 provides a mechanistic model for the conserved SNF5/SMARCB1 class in eukaryotes. Its acidic-patch engagement and scaffold-like role inform interpretation of disease-linked mammalian orthologs and broader chromatin-remodeler targeting strategies; in yeast, SWI/SNF studies also inform metabolic engineering and stress-response transcription models. | Expert reviews use yeast Snf5/SMARCB1 conservation to explain SWI/SNF mechanism and disease relevance; recent metabolism-focused work highlights SWI/SNF as a regulator of metabolic transcription. (kuwahara2023recentinsightsinto pages 3-5, church2023theswisnfchromatin pages 1-2, chen2023mechanismofaction pages 2-6, eustermann2024energydrivengenomeregulation pages 1-3) | Kuwahara et al., 2023, Cancer Medicine; Church & Workman, 2024, Biochemical Society Transactions; Eustermann et al., 2024, Nature Reviews Molecular Cell Biology | https://doi.org/10.1002/cam4.6255 ; https://doi.org/10.1042/bst20231141 ; https://doi.org/10.1038/s41580-023-00683-y | Jun 2023; Apr 2024; Dec 2024 |
Table: This table compiles the main evidence retrieved for Saccharomyces cerevisiae Snf5, focusing on identity, mechanism, nucleosome interactions, transcriptional roles, quantitative findings, and broader implications. It is useful as a concise source map for building a full research report on UniProt P18480.
11) Summary functional annotation (concise)
Snf5 (yeast SNF5-family; UniProt P18480 per user) is best supported as a core SWI/SNF chromatin remodeling subunit whose primary mechanistic contribution is to form the nucleosome-binding lobe and bind the nucleosomal acidic patch via a conserved finger helix (Arg669 anchor), thereby stabilizing remodeler–nucleosome engagement and helping couple ATPase-driven DNA translocation to productive nucleosome sliding/ejection. (chen2023mechanismofaction pages 2-6, eustermann2024energydrivengenomeregulation pages 1-3)
Functionally, Snf5 participates in transcriptional control by interacting with transcriptional activators and supporting promoter remodeling, and SWI/SNF can exert both activation and direct repression depending on context, including LUTI-based transcriptional interference and metabolic pathway regulation (notably sulfur metabolism). (wendegatz2024transcriptionalactivationdomains pages 5-6, morse2024swisnfchromatinremodeling pages 15-19, church2023theswisnfchromatin pages 2-3)
References
-
(kuwahara2023recentinsightsinto pages 3-5): Yasumichi Kuwahara, Tomoko Iehara, Akifumi Matsumoto, and Tsukasa Okuda. Recent insights into the swi/snf complex and the molecular mechanism of hsnf5 deficiency in rhabdoid tumors. Cancer Medicine, 12:16323-16336, Jun 2023. URL: https://doi.org/10.1002/cam4.6255, doi:10.1002/cam4.6255. This article has 5 citations and is from a peer-reviewed journal.
-
(eustermann2024energydrivengenomeregulation pages 1-3): Sebastian Eustermann, Avinash B. Patel, Karl-Peter Hopfner, Yuan He, and Philipp Korber. Energy-driven genome regulation by atp-dependent chromatin remodellers. Nature reviews. Molecular cell biology, 25:309-332, Dec 2024. URL: https://doi.org/10.1038/s41580-023-00683-y, doi:10.1038/s41580-023-00683-y. This article has 113 citations.
-
(wendegatz2024transcriptionalactivationdomains pages 5-6): Eva-Carina Wendegatz, Maike Engelhardt, and Hans-Joachim Schüller. Transcriptional activation domains interact with atpase subunits of yeast chromatin remodelling complexes swi/snf, rsc and ino80. Current Genetics, Sep 2024. URL: https://doi.org/10.1007/s00294-024-01300-x, doi:10.1007/s00294-024-01300-x. This article has 3 citations and is from a peer-reviewed journal.
-
(chen2023mechanismofaction pages 2-6): Kangjing Chen, Junjie Yuan, Youyang Sia, and Zhucheng Chen. Mechanism of action of the swi/snf family complexes. Nucleus, Jan 2023. URL: https://doi.org/10.1080/19491034.2023.2165604, doi:10.1080/19491034.2023.2165604. This article has 55 citations and is from a peer-reviewed journal.
-
(amaris2022structuralinvestigationof pages 34-44): Structural Investigation of Chromatin Regulatory Complexes Using Electron Microscopy This article has 0 citations and is from a peer-reviewed journal.
-
(chen2023mechanismofaction media 799148c2): Kangjing Chen, Junjie Yuan, Youyang Sia, and Zhucheng Chen. Mechanism of action of the swi/snf family complexes. Nucleus, Jan 2023. URL: https://doi.org/10.1080/19491034.2023.2165604, doi:10.1080/19491034.2023.2165604. This article has 55 citations and is from a peer-reviewed journal.
-
(chen2023mechanismofaction media ad429c0b): Kangjing Chen, Junjie Yuan, Youyang Sia, and Zhucheng Chen. Mechanism of action of the swi/snf family complexes. Nucleus, Jan 2023. URL: https://doi.org/10.1080/19491034.2023.2165604, doi:10.1080/19491034.2023.2165604. This article has 55 citations and is from a peer-reviewed journal.
-
(chen2023mechanismofaction media 79a39cc8): Kangjing Chen, Junjie Yuan, Youyang Sia, and Zhucheng Chen. Mechanism of action of the swi/snf family complexes. Nucleus, Jan 2023. URL: https://doi.org/10.1080/19491034.2023.2165604, doi:10.1080/19491034.2023.2165604. This article has 55 citations and is from a peer-reviewed journal.
-
(morse2024swisnfchromatinremodeling pages 5-8): Kaitlin Morse, Alena L. Bishop, Sarah Swerdlow, Jessica M. Leslie, and Elçin Ünal. Swi/snf chromatin remodeling regulates transcriptional interference and gene repression. Aug 2024. URL: https://doi.org/10.1016/j.molcel.2024.06.029, doi:10.1016/j.molcel.2024.06.029. This article has 21 citations and is from a highest quality peer-reviewed journal.
-
(morse2024swisnfchromatinremodeling pages 15-19): Kaitlin Morse, Alena L. Bishop, Sarah Swerdlow, Jessica M. Leslie, and Elçin Ünal. Swi/snf chromatin remodeling regulates transcriptional interference and gene repression. Aug 2024. URL: https://doi.org/10.1016/j.molcel.2024.06.029, doi:10.1016/j.molcel.2024.06.029. This article has 21 citations and is from a highest quality peer-reviewed journal.
-
(morse2024swisnfchromatinremodeling pages 19-22): Kaitlin Morse, Alena L. Bishop, Sarah Swerdlow, Jessica M. Leslie, and Elçin Ünal. Swi/snf chromatin remodeling regulates transcriptional interference and gene repression. Aug 2024. URL: https://doi.org/10.1016/j.molcel.2024.06.029, doi:10.1016/j.molcel.2024.06.029. This article has 21 citations and is from a highest quality peer-reviewed journal.
-
(church2023theswisnfchromatin pages 1-2): Michael C Church, Andrew Price, Hua Li, and Jerry L Workman. The swi-snf chromatin remodeling complex mediates gene repression through metabolic control. Nucleic Acids Research, 51:10278-10291, Aug 2023. URL: https://doi.org/10.1093/nar/gkad711, doi:10.1093/nar/gkad711. This article has 13 citations and is from a highest quality peer-reviewed journal.
-
(church2023theswisnfchromatin pages 2-3): Michael C Church, Andrew Price, Hua Li, and Jerry L Workman. The swi-snf chromatin remodeling complex mediates gene repression through metabolic control. Nucleic Acids Research, 51:10278-10291, Aug 2023. URL: https://doi.org/10.1093/nar/gkad711, doi:10.1093/nar/gkad711. This article has 13 citations and is from a highest quality peer-reviewed journal.
-
(church2024theswisnfchromatin pages 1-2): Michael C. Church and Jerry L. Workman. The swi/snf chromatin remodeling complex: a critical regulator of metabolism. Biochemical Society Transactions, 52:1327-1337, Apr 2024. URL: https://doi.org/10.1042/bst20231141, doi:10.1042/bst20231141. This article has 21 citations and is from a peer-reviewed journal.
-
(church2024theswisnfchromatin pages 6-8): Michael C. Church and Jerry L. Workman. The swi/snf chromatin remodeling complex: a critical regulator of metabolism. Biochemical Society Transactions, 52:1327-1337, Apr 2024. URL: https://doi.org/10.1042/bst20231141, doi:10.1042/bst20231141. This article has 21 citations and is from a peer-reviewed journal.
-
(church2024theswisnfchromatin pages 2-4): Michael C. Church and Jerry L. Workman. The swi/snf chromatin remodeling complex: a critical regulator of metabolism. Biochemical Society Transactions, 52:1327-1337, Apr 2024. URL: https://doi.org/10.1042/bst20231141, doi:10.1042/bst20231141. This article has 21 citations and is from a peer-reviewed journal.
-
(wendegatz2024transcriptionalactivationdomains pages 1-2): Eva-Carina Wendegatz, Maike Engelhardt, and Hans-Joachim Schüller. Transcriptional activation domains interact with atpase subunits of yeast chromatin remodelling complexes swi/snf, rsc and ino80. Current Genetics, Sep 2024. URL: https://doi.org/10.1007/s00294-024-01300-x, doi:10.1007/s00294-024-01300-x. This article has 3 citations and is from a peer-reviewed journal.
-
(church2024theswisnfchromatin pages 8-9): Michael C. Church and Jerry L. Workman. The swi/snf chromatin remodeling complex: a critical regulator of metabolism. Biochemical Society Transactions, 52:1327-1337, Apr 2024. URL: https://doi.org/10.1042/bst20231141, doi:10.1042/bst20231141. This article has 21 citations and is from a peer-reviewed journal.
-
(gourisankar2024contextspecificfunctionsof pages 1-3): Sai Gourisankar, Andrey Krokhotin, Wendy Wenderski, and Gerald R. Crabtree. Context-specific functions of chromatin remodellers in development and disease. Nature Reviews Genetics, 25:340-361, Nov 2024. URL: https://doi.org/10.1038/s41576-023-00666-x, doi:10.1038/s41576-023-00666-x. This article has 75 citations and is from a domain leading peer-reviewed journal.
-
(gourisankar2024contextspecificfunctionsof pages 27-28): Sai Gourisankar, Andrey Krokhotin, Wendy Wenderski, and Gerald R. Crabtree. Context-specific functions of chromatin remodellers in development and disease. Nature Reviews Genetics, 25:340-361, Nov 2024. URL: https://doi.org/10.1038/s41576-023-00666-x, doi:10.1038/s41576-023-00666-x. This article has 75 citations and is from a domain leading peer-reviewed journal.
-
(lampersberger2023geneticinteractorsof pages 37-41): Genetic interactors of the SWI/SNF chromatin remodelling complex in Caenorhabditis elegans This article has 0 citations.
-
(lampersberger2023geneticinteractorsof pages 34-37): Genetic interactors of the SWI/SNF chromatin remodelling complex in Caenorhabditis elegans This article has 0 citations.
Artifacts
Citations
- chen2023mechanismofaction pages 2-6
- eustermann2024energydrivengenomeregulation pages 1-3
- amaris2022structuralinvestigationof pages 34-44
- wendegatz2024transcriptionalactivationdomains pages 5-6
- morse2024swisnfchromatinremodeling pages 5-8
- morse2024swisnfchromatinremodeling pages 15-19
- morse2024swisnfchromatinremodeling pages 19-22
- church2023theswisnfchromatin pages 1-2
- church2023theswisnfchromatin pages 2-3
- church2024theswisnfchromatin pages 1-2
- church2024theswisnfchromatin pages 6-8
- church2024theswisnfchromatin pages 2-4
- church2024theswisnfchromatin pages 8-9
- gourisankar2024contextspecificfunctionsof pages 1-3
- wendegatz2024transcriptionalactivationdomains pages 1-2
- kuwahara2023recentinsightsinto pages 3-5
- gourisankar2024contextspecificfunctionsof pages 27-28
- lampersberger2023geneticinteractorsof pages 37-41
- lampersberger2023geneticinteractorsof pages 34-37
- https://doi.org/10.1002/cam4.6255
- https://doi.org/10.17863/cam.93003
- https://doi.org/10.1080/19491034.2023.2165604
- https://doi.org/10.1038/s41580-023-00683-y
- https://doi.org/10.1007/s00294-024-01300-x
- https://doi.org/10.1016/j.molcel.2024.06.029
- https://doi.org/10.1093/nar/gkad711
- https://doi.org/10.1042/bst20231141
- https://doi.org/10.1002/cam4.6255,
- https://doi.org/10.1038/s41580-023-00683-y,
- https://doi.org/10.1007/s00294-024-01300-x,
- https://doi.org/10.1080/19491034.2023.2165604,
- https://doi.org/10.1016/j.molcel.2024.06.029,
- https://doi.org/10.1093/nar/gkad711,
- https://doi.org/10.1042/bst20231141,
- https://doi.org/10.1038/s41576-023-00666-x,
Perplexity
(SNF5-deep-research-perplexity.md)
Perplexity
(SNF5-deep-research-perplexity.md)SNF5: A Critical Catalytic Subunit of the SWI/SNF Chromatin-Remodeling Complex in Saccharomyces cerevisiae
The SNF5 gene (UniProt P18480) encodes a core subunit of the SWI/SNF chromatin-remodeling complex in Saccharomyces cerevisiae, functioning as an essential nucleosome-binding module that couples ATP hydrolysis to DNA translocation during chromatin remodeling. This 102.5 kiloDalton protein serves as a regulatory hub within the complex, mediating histone-DNA interactions through its conserved arginine-rich repeat domains and functioning as a critical anchor point that stabilizes the nucleosomal substrate during active remodeling. SNF5 is required for recruitment of the SWI/SNF complex by transcriptional activators to target promoters, for the assembly of cell-type-specific gene expression programs during differentiation, for metabolic adaptation to changing carbon sources, and for numerous other ATP-dependent processes that require access to packaged chromatin. Loss or dysfunction of SNF5 severely impairs the catalytic efficiency and remodeling capacity of the entire SWI/SNF complex while also affecting its subcellular localization, component assembly, and recruitment selectivity. In mammals, the SNF5 ortholog SMARCB1/INI1/BAF47 functions as a bona fide tumor suppressor, and its loss drives development of aggressive pediatric cancers including malignant rhabdoid tumors and atypical teratoid/rhabdoid tumors, establishing SNF5 as a critically conserved regulator of genome accessibility and cellular fate determination across eukaryotes.
The SWI/SNF Complex: Architectural Organization and SNF5's Structural Position
The SWI/SNF (switching defective/sucrose nonfermenting) complex represents one of the most extensively characterized ATP-dependent chromatin remodeling complexes, with SNF5 serving as a critical core component of this multi-subunit machine that slides and evicts nucleosomes to regulate chromatin structure and gene expression[1][4][6]. The complex comprises twelve subunits in Saccharomyces cerevisiae and functions as a highly conserved molecular machine found from yeast to humans, with SNF5 being one of the universally conserved components across eukaryotes[1][10][29]. The SWI/SNF complex can be functionally organized into distinct modular units that each contribute specific structural and regulatory functions to the overall remodeling mechanism[3][10][19]. The primary catalytic core consists of the Snf2 ATPase domain, which catalyzes nucleotide-dependent DNA translocation, along with two actin-related proteins, Arp7 and Arp9, that form an essential ARP module[3][19].
SNF5 functions as a critical component of the substrate recruitment module (SRM), which comprises a specialized set of subunits responsible for recognizing and engaging nucleosomal substrates and facilitating recruitment of the complex to target sites in chromatin[34]. The architectural position of SNF5 within the complex places it as a member of what structural studies term the "Arm" module, which is composed of SNF5, the N-terminal SWIRM domains of Swi3, and the C-terminus of Swp82[3][10]. This modular organization reflects the sophisticated assembly logic of the SWI/SNF complex, where distinct functional units are integrated through multiple protein-protein interactions to generate coordinated remodeling activity[3][19]. SNF5 exists within a tightly integrated submodule consisting of Snf5, Swp82, and Taf14, which collectively mediates several critical functions of the larger complex[8][12]. Crosslinking-mass spectrometry analysis has revealed extensive interactions between this Snf5-containing submodule and other complex components, particularly with the Swi3 SANT and SWIRM domains, which serve as key scaffolding elements within the SWI/SNF architecture[8][12][19]. The conserved SNF5 repeat (RPT) domains each engage one copy of the Swi3 SWIRM domain through multiple contact points, establishing SNF5 as a central organizing element that coordinates the spatial arrangement of multiple complex subunits[3][10].
Loss of SNF5 results in substantial destabilization of the overall SWI/SNF complex assembly, with the Snf2-Arp7-Arp9 core module becoming completely separated from the rest of the complex[8][12][34]. This dramatic effect on complex integrity demonstrates that SNF5 is not merely a peripheral subunit but rather a critically important structural linker that maintains the functional organization of the entire machine. The modular architecture suggests that SNF5 serves specific roles in stabilizing interactions between the catalytic core and the regulatory modules, thereby ensuring coordinated function of the complex components. Indeed, proteomic analysis of SNF5-deficient complexes shows that while peripheral subunits such as Swp82, Taf14, and Snf6 dissociate from Snf2, the core ARP module remains intact but functionally compromised[19]. This pattern indicates that SNF5 serves as a critical bridge between different functional modules of the SWI/SNF complex, and its loss creates an "aberrant" complex with substantially altered biochemical properties and chromatin targeting capabilities[8][12][22].
Domain Organization and Molecular Features of SNF5
The SNF5 protein exhibits a distinctive multi-domain architecture that reflects its multifaceted roles within the SWI/SNF complex, with particular emphasis on the conserved repeat domains and regulatory regions that mediate its various functions[18][21][36]. The protein contains two imperfect 60-amino-acid repeat domains (Rpt1 and Rpt2) followed by a putative coiled-coil region, with the N-terminal region being notably diverse across different organisms[18][21][36]. In metazoans such as humans, a winged helix domain (WHD) is found N-terminal to the repeat domains; this domain is structurally related to the SKI/SNO/DAC domain and has been shown to be a site of mutations that cause the tumor-predisposing syndrome schwannomatosis[36][48][51]. Computational and structural studies have revealed that the Rpt1 repeat domain contains a characteristic alpha-beta fold that is evolutionarily conserved among SNF5 family proteins, with a hydrophobic core composed of several leucine and phenylalanine residues surrounded by charged residues that are exposed to the solvent[18][21][48]. The linker region connecting Rpt1 and Rpt2 exhibits intrinsic disorder, which may allow these repeat domains to function somewhat independently and potentially fold independently of each other[18][21].
Perhaps most importantly for SNF5 function, both repeat domains contain conserved arginine-rich regions that serve as critical interaction surfaces for binding the nucleosomal histone octamer[3][6][7][10][33]. The C-terminal helix of SNF5, termed the "finger helix," protrudes from the body of the nucleosome binding lobe and makes multiple contacts with the histone surface through multiple arginine residues, with Arg669 in yeast SNF5 (equivalent to Arg370 in human SMARCB1) functioning as the canonical arginine anchor[6][33]. This arginine-rich region directly interacts with the acidic patch of the H2A-H2B histone dimer, a highly conserved and functionally critical region of the nucleosome that serves as a landing dock for numerous chromatin regulators[6][7][33]. Recent cryo-electron microscopy structures of the SWI/SNF complex bound to nucleosomes have provided near-atomic resolution details showing that SNF5's C-terminal extension makes specific contacts with the histone octamer surface, forming an interaction that could serve as an anchor point during active DNA translocation when the nucleosome is being actively remodeled[3][10][33].
A striking feature of yeast SNF5 that distinguishes it from its mammalian orthologs is the presence of a large N-terminal glutamine-rich low-complexity region (QLC) that comprises approximately the first 334 amino acids of the protein[38][42][56]. This glutamine-rich domain is enriched in the amino acids glutamine and proline and contains seven histidine residues positioned either within the QLC or adjacent to it[42][56]. While this QLC is extremely divergent from the mammalian SNF5 ortholog, structural and functional studies indicate that this region plays important regulatory roles in sensing environmental conditions and modulating SNF5 function during metabolic transitions[42][56]. The presence of multiple histidine residues within the QLC suggests that this region may function as a pH sensor, with histidine protonation potentially causing conformational changes that alter the ability of SNF5 to interact with different sets of transcription factors and drive recruitment to specific sets of promoters[42][56]. The substantial portion of yeast SNF5 that is unique to the yeast complex (approximately 70% of the protein) appears not to make direct contact with the H2A-H2B acidic pocket but rather serves regulatory functions related to transcription factor binding and complex recruitment[37].
Biochemical Function: Nucleosome Binding and Anchoring During ATP-Dependent Remodeling
The primary biochemical function of SNF5 within the SWI/SNF complex is to promote and stabilize binding of the nucleosomal substrate, particularly through direct interactions with histone components, thereby enhancing the catalytic activity of the Snf2 ATPase and coupling ATP hydrolysis to productive DNA translocation[2][8][12][37]. SNF5 promotes binding of the Snf2 ATPase domain to nucleosomal DNA through a mechanism that involves the stabilization of the ATPase-nucleosome interface, with studies showing that loss of SNF5 results in reduced affinity of the ATPase domain for nucleosomal DNA specifically, without affecting the general ATP binding capacity of the complex[8][12][37]. Biochemical analysis reveals that deletion of the SNF5 submodule (Snf5-Swp82-Taf14) reduces the catalytic efficiency (k_cat) of the complex two to three-fold depending on the specific substrate, while leaving the affinity for ATP (K_m) essentially unchanged[8][12]. This pattern of kinetic effects indicates that SNF5 specifically regulates the efficiency of ATP hydrolysis or DNA translocation activity rather than affecting nucleotide binding per se[8][12][37].
The anchoring mechanism proposed based on cryo-EM structural data suggests that SNF5 physically locks the histone octamer in place as the nucleosomal DNA is being translocated around the octamer, thereby coupling ATP hydrolysis with productive chromatin remodeling[3][10][33][49]. This anchoring mechanism differs substantially from that employed by other large chromatin remodeling complexes such as INO80 and SWR1, where the actin-related protein (ARP) module serves as the primary anchor point; in SWI/SNF, the SNF5-containing ARM module carries out this critical anchoring function[3][10][33]. Deletion of the repeat (RPT) domains in SNF5 uncouples ATP hydrolysis by Snf2 from actual chromatin remodeling activity, demonstrating the essential role of these domains in transducing the mechanical work generated by ATP hydrolysis into productive nucleosome movement[3][33][49]. The mechanism of nucleosome engagement involves the ATPase domain of Snf2 binding the nucleosome at super helical location (SHL) 2, the same location shown in stand-alone Snf2 ATPase-nucleosome structures as well as in other chromatin remodeling complexes[3].
SNF5 also facilitates recruitment of the SWI/SNF complex by transcriptional activators, serving as one of two primary activator-binding domains within the complex alongside the ARID domain of Swi1[9][31][47]. The N-terminus of SNF5, encompassing the glutamine-rich low-complexity region and additional sequences, has been shown to interact with activation domains of transcriptional activators such as VP16 and Gcn4, though this interaction alone is insufficient to mediate full recruitment of the complex to DNA[9][31][47]. Rather, the recruitment function of SNF5 appears to require cooperation with Swi1, as deletion of either domain individually reduces SWI/SNF recruitment by transcription factors, but deletion of only one domain is insufficient to completely block recruitment[9][31][47]. More recent evidence demonstrates that SNF5 is required for SWI/SNF recruitment specifically by acidic transcription factors, suggesting that the interaction domains within SNF5 preferentially recognize transcription factors with acidic activation domains[31][37]. When SNF5 is deleted, SWI/SNF still retains affinity for nucleosomes and can be recruited to some SWI/SNF target genes through alternative mechanisms, but the complex fails to respond efficiently to recruitment signals from acidic transcription factors[31]. This pattern indicates that SNF5 serves as a critical licensing factor that allows the SWI/SNF complex to respond to specific classes of transcriptional activators.
SNF5 Loss and Formation of Aberrant SWI/SNF Complexes
A major area of research has focused on characterizing how loss of SNF5 alters the composition, structure, and function of the overall SWI/SNF complex, with the consistent finding that SNF5-deficient complexes represent "aberrant" forms that retain some but not all normal functions[8][12][22][31]. Crosslinking-mass spectrometry studies have systematically mapped the effects of SNF5 deletion on complex architecture, revealing that loss of SNF5 causes complete dissociation of the Snf2-Arp7-Arp9 core from the rest of the complex while a small Snf6-Snf12-Swi3 sub-module remains partially intact but associates only weakly with remaining components[8][12][19][22]. The peripheral subunits Swp82, Taf14, and Snf11 can no longer associate with Snf2 or other core subunits when SNF5 is absent, indicating that SNF5 serves as a critical structural element required for interactions between multiple complex modules[8][19]. Swi1, which contains the ARID domain important for both activator binding and DNA binding, shows reduced association with other complex components in the absence of SNF5, suggesting that SNF5 helps stabilize the integration of Swi1 into the larger complex[8].
Functionally, the aberrant SNF5-deficient complex exhibits several critical defects relative to wild-type SWI/SNF. First, nucleosome binding is substantially impaired, with the ATPase domain showing reduced ability to contact nucleosomal DNA even though the complex can still bind nucleosomes through alternative, weaker interactions[8][31][37]. Second, the catalytic efficiency is reduced two to three-fold as determined through Michaelis-Menten kinetic analysis[8]. Third, and importantly, the complex cannot be efficiently recruited to chromatin by acidic transcription factors, indicating that the loss of SNF5-mediated recruitment functions prevents targeting of the complex to many of its normal promoter-proximal sites[8][31]. Fourth, global transcriptomic analysis reveals that the aberrant complex regulates a substantially altered set of target genes compared to wild-type SWI/SNF, with some genes showing reduced expression and others showing increased expression, suggesting that the residual complex activity is misdirected to non-canonical targets[8][31].
These findings have important implications for understanding how SNF5 loss contributes to cancer development in mammals. The SMARCB1 tumor suppressor (human ortholog of SNF5) is frequently lost in pediatric rhabdoid cancers, and studies of SMARCB1-deficient cancer cell lines demonstrate that BAF complexes lacking SMARCB1 show altered localization patterns and target gene selection, often aberrantly activating oncogenic programs[29][51]. The mechanism appears to involve both the direct loss of recruitment signals (due to SMARCB1's role in binding transcription factors) and potential misdirection of residual BAF complex activity to alternative sites in the genome[29][51]. In some cancer contexts, the SMARCB1-deficient BAF complexes are targeted for proteasomal degradation when associated with fusion oncoproteins such as SS18-SSX, while in other contexts the complexes remain partially functional, perhaps allowing them to drive alternative gene expression programs that support tumor development[29][51].
Role of SNF5 in Cell Differentiation and Development
Among the most illuminating studies of SNF5 function have been those employing conditional genetic systems to specifically inactivate SNF5 during defined developmental transitions, particularly studies examining the role of SNF5 during hepatocyte differentiation[1][13][55]. These investigations have revealed that SNF5 is not merely a generic remodeling enzyme but rather plays critical roles in activating cell-type-specific gene expression programs that drive differentiation[1][13][55]. When SNF5 is conditionally deleted in the developing liver using the AlfpCre transgene, which drives recombination beginning at the onset of liver bud formation, hepatocyte development is profoundly disrupted despite the fact that cell differentiation is not completely blocked[1][13][55]. Global transcriptome analysis of SNF5-null hepatocytes reveals that roughly 70% of genes that are normally upregulated during hepatocyte differentiation show reduced expression in mutant tissue, indicating that SNF5 acts positively on the expression of the vast majority of liver-specific and developmentally activated genes[1][13][55].
The molecular basis of these differentiation defects can be traced to impaired transcriptional activation of specific gene sets crucial for hepatocyte function and morphology[1][13][55]. For instance, genes involved in glycogen synthesis show dramatic downregulation, with liver glycogen synthase (Gys2) reduced 2.1-fold in SNF5-null hepatocytes, directly accounting for the marked reduction in hepatic glycogen storage observed in mutant animals[1][13]. Similarly, genes involved in gluconeogenesis show reduced expression, and the combined defects in both glycogen synthesis and gluconeogenesis result in severe hypoglycemia in fasted SNF5-null animals[1]. Beyond metabolic processes, SNF5 is essential for proper assembly of epithelial cell-cell junctions, which are hallmarks of terminal hepatocyte differentiation[1][13][55]. Immunohistochemical analysis reveals defective localization of tight junction proteins such as zonula occludens-1 (ZO-1), adherens junction proteins such as E-cadherin and beta-catenin, gap junction proteins such as connexin 32, and desmosomal proteins in SNF5-null hepatocytes[1][13][55]. Transcriptome analysis indicates that this morphological defect reflects defective transcriptional activation of numerous genes encoding these junction proteins, suggesting that SNF5 is specifically required for activating the genetic program that promotes epithelial organization[1][13][55].
A particularly interesting finding is that despite these severe developmental defects, a fraction of hepatic developmentally activated genes are expressed at near-normal levels in SNF5-null tissue, indicating that SNF5-independent mechanisms can compensate for the loss of this complex at a subset of genes[1][13][55]. This observation suggests that while SNF5/SWI/SNF is the dominant remodeling complex governing activation of most hepatocyte-specific genes, alternative remodeling complexes such as ISWI or CHD can provide partial compensation at certain loci. Additionally, SNF5-deleted hepatocytes show increased proliferation compared to control hepatocytes, a phenotype that is accompanied by misexpression of several cell cycle-related genes[1][13][55]. This increased proliferation appears to reflect disrupted cell cycle regulation, as the normal developmental program in hepatocytes involves a transition from proliferative hepatoblasts to quiescent differentiated hepatocytes, and loss of SNF5 prevents this transition by disrupting expression of cell cycle inhibitors. These findings underscore the critical importance of SWI/SNF-mediated chromatin remodeling in coordinating the activation of tissue-specific genetic programs during development.
SNF5 Function in Metabolic Adaptation and Environmental Stress Responses
Beyond development, SNF5 has been shown to play important roles in coordinating transcriptional responses to changes in environmental conditions, particularly in response to carbon source availability and nutrient stress[42][56][59]. Studies examining SWI/SNF function during transitions between glucose-rich and glucose-poor conditions have revealed that SNF5's N-terminal glutamine-rich low-complexity region plays a surprisingly sophisticated regulatory role in sensing metabolic stress and directing transcriptional reprogramming[42][56]. When yeast cells experience glucose starvation, they must rapidly reprogram their metabolism from fermentation to respiration and activate genes involved in utilizing alternative carbon sources such as ethanol; this metabolic transition critically depends on SWI/SNF-mediated chromatin remodeling at specific glucose-repressed genes such as ADH2[42][56][59].
Detailed analysis reveals that the SNF5 glutamine-rich low-complexity region and its embedded histidine residues are specifically required for efficient induction of ADH2 expression during carbon starvation, suggesting that SNF5 functions as a metabolic sensor that responds to changes in cellular pH or other indicators of metabolic stress[42][56]. The proposed mechanism involves pH-dependent protonation of histidine residues within the QLC, which would cause conformational changes in this intrinsically disordered region, potentially enabling interaction with different sets of transcription factors or altering the binding properties of SNF5 for specific transcriptional activators[42][56]. Strains carrying deletions of the SNF5 QLC domain (ΔQsnf5) maintain wild-type levels of ADH2 repression in the presence of glucose but completely fail to induce ADH2 expression during carbon starvation, indicating that this domain specifically controls the transition from repression to activation during the metabolic shift[42][56]. This finding represents a sophisticated form of post-translational regulation, where the structural properties of an intrinsically disordered protein region allow sensing of environmental pH changes and translation of this signal into altered transcriptional responses.
Additional evidence for SNF5 involvement in metabolic control comes from studies examining the roles of the SWI/SNF complex in regulating genes involved in coenzyme Q biosynthesis and the transition between fermentative and respiratory metabolism[24]. These studies reveal that Snf2 (the ATPase subunit of SWI/SNF) plays roles in regulating alternative splicing of the PTC7 transcript, with deletion of SNF2 leading to increased splicing of PTC7 and altered coenzyme Q6 synthesis[24]. The effects on metabolic pathways are extensive, suggesting that SWI/SNF complexes coordinate both transcriptional and post-transcriptional responses to metabolic challenges[24].
Molecular Mechanisms of SNF5-Mediated Nucleosome Recognition and Remodeling
The detailed molecular mechanisms by which SNF5 specifically recognizes nucleosomes and coordinates remodeling have been increasingly illuminated through structural and biochemical studies, particularly through cryo-electron microscopy structures of the SWI/SNF complex bound to nucleosomes[3][10][33]. These structures reveal a sophisticated pincer-like grasping mechanism where SNF5 and the Snf2 ATPase domain approach the nucleosome from opposite sides, with SNF5 specifically recognizing the histone octamer surface while Snf2 engages nucleosomal DNA[3][10][33]. The C-terminal repeat domains of SNF5 make extensive contacts with the H2A-H2B dimer through the acidic patch, which has emerged as a master landing platform that mediates recruitment of numerous chromatin regulatory proteins[6][7][33]. The highly conserved arginine-rich finger helix within SNF5 acts as a critical recognition element, with multiple arginine residues forming direct hydrogen bonds and electrostatic interactions with the negatively charged residues of the acidic patch[6][33].
The nucleosome-binding mode of SWI/SNF differs markedly from earlier models that proposed extensive rearrangement of DNA-histone contacts during remodeling, as cryo-EM structures show that the nucleosome remains largely intact when bound to SWI/SNF, with the primary structural perturbations involving increased DNA unwrapping at the nucleosome entry site[3]. This relatively modest perturbation of nucleosome structure is consistent with an ATP-dependent mechanism of chromatin remodeling in which DNA is translocated around the largely intact histone octamer in a wave-like manner, with one base pair of DNA being bulged out at a time as the ATPase domain cycles through ATP binding, hydrolysis, and release[3][49]. The role of SNF5 in this mechanism is to stabilize the histone octamer against displacement as DNA waves propagate along the surface of the nucleosome, effectively anchoring the octamer in place while the DNA moves relative to the histones[3][33][49].
Photocrosslinking studies have mapped the specific histone residues contacted by SNF5, revealing that the conserved SNF5 homology domain directly contacts the region of H2B near residue 109, which lies near the center of the acidic patch[37]. This positioning allows SNF5 to make multiple contacts across the acidic patch surface, stabilizing its interaction with the nucleosome through a network of electrostatic and hydrogen-bonding interactions. The interaction between SNF5 and the histone octamer is dynamic and likely undergoes conformational changes during the remodeling cycle, with SNF5 potentially modulating the strength or geometry of its histone contacts as the complex transitions through different states of the remodeling reaction[3][33][49].
SNF5 and Global Nucleosome Positioning at Promoter Regions
Recent chromatin immunoprecipitation combined with deep sequencing (ChIP-seq) studies have revealed that SNF5/SWI/SNF plays critical roles in establishing and maintaining proper nucleosome positioning at promoter regions, particularly in controlling the occupancy and positioning of the +1 nucleosome immediately downstream of transcriptional start sites[46][43]. These studies reveal that the complex is highly enriched not only at the -1 and +1 nucleosome positions but also over the nucleosome-depleted region (NDR) at promoters[46]. Critically, the complex is essential for establishment of high nucleosome occupancy at these positions relative to flanking regions, sculpting the characteristic high-occupancy, high-positioned nucleosome landscape that characterizes active promoters[46]. When SNF5 is deleted in mammalian fibroblasts, nucleosome occupancy is markedly reduced across peri-TSS regions, with particularly dramatic effects at the +1 nucleosome position and upstream of the NDR[46].
The positioning of the +1 nucleosome has been shown to play functional roles in regulating RNA polymerase II promoter-proximal pausing, with a strongly positioned +1 nucleosome enhancing pausing of the polymerase at this region, which in turn facilitates pre-mRNA quality control through promotion of 5' capping[43]. This suggests that SWI/SNF-mediated control of nucleosome positioning has dual effects on transcription: establishing nucleosome-depleted regions at promoters to allow transcription factor binding and pre-initiation complex assembly, while simultaneously positioning downstream nucleosomes to modulate polymerase elongation rates. The specific effect on the +1 nucleosome is particularly important, as reducing SNF2H (an ISWI-family remodeler) levels decreases +1 nucleosome positioning and increases polymerase pause release, demonstrating the functional importance of this nucleosome for controlling transcription elongation[43].
Conservation and Human Disease Significance
The exceptional conservation of SNF5 across all eukaryotic species underscores its fundamental importance in eukaryotic chromatin biology[29][51]. The human ortholog, originally termed INI1 (integrase interactor 1) because it was discovered as a binding partner of HIV-1 integrase in 1994, was subsequently identified as a bona fide tumor suppressor gene[29]. This discovery came from observations that biallelic inactivation of SMARCB1 (also called BAF47, SMARCB1, or hSNF5) occurs in virtually all cases of malignant rhabdoid tumors (MRT) and atypical teratoid/rhabdoid tumors (ATRT), two highly aggressive pediatric malignancies that predominantly affect very young children[29][51]. The tumor-suppressive functions of SMARCB1 are now understood to involve multiple mechanisms beyond simple loss of chromatin remodeling activity, including functions in genome-wide BAF stability at enhancers and promoters, recognition of specific DNA sequences through its unique N-terminal winged helix domain (which is absent in yeast SNF5), and independent anti-proliferative functions unrelated to ATPase activity of the complex[29][51].
Germline mutations in the SMARCB1 winged helix domain have been associated with schwannomatosis, a cancer-predisposing condition characterized by formation of benign nerve sheath tumors called schwannomas[36][51]. These mutations are typically missense mutations or in-frame deletions that disrupt the fold or function of the winged helix domain, raising important questions about how this metazoan-specific domain contributes to tumor suppression[36][51]. The finding that the winged helix domain is deeply buried within the BAF complex structure, far from nucleosomal DNA, suggests that this domain plays roles in complex assembly or recruitment functions rather than direct DNA binding[29][51]. Recent evidence indicates that the SMARCB1 winged helix domain may facilitate interactions with other BAF subunits or regulate the conformational state of the complex[29].
The BAF complex (the mammalian counterpart of yeast SWI/SNF) is now recognized as one of the most frequently mutated protein complexes in human cancers, with mutations affecting the complex or its regulatory factors occurring in up to 25% of all human malignancies[29]. While SMARCB1 loss is the defining feature of rhabdoid cancers, other BAF subunits are recurrently mutated in diverse cancer types, including SMARCA4 (ATPase subunit) in lung and gastric cancers, ARID1A in multiple cancer types, and several other subunits in various malignancies[29]. The consistent association between BAF complex dysfunction and cancer development emphasizes the critical importance of SWI/SNF-mediated chromatin remodeling in suppressing inappropriate cell proliferation and maintaining proper gene expression programs.
SNF5 in Fungal Pathogenesis and Cellular Adaptation
Beyond model organism studies, SNF5 and its close homolog Sfh1 (from the RSC complex) have been shown to play important roles in fungal pathogens, with implications for understanding how pathogenic fungi adapt to host environments[34][58]. In Candida albicans, a major human fungal pathogen, deletion of the core SWI/SNF subunits swi1 and snf2 leads to complete loss of pathogenicity in mouse models, indicating that chromatin remodeling is essential for the virulence functions of this organism[34]. The SWI/SNF complex regulates genes involved in morphological transitions, biofilm formation, and drug resistance, suggesting that proper chromatin remodeling is required for the pathogen to successfully colonize and persist in host tissues[34]. Interestingly, the SWI/SNF complex has been implicated in promoting nucleosomal displacement from the Mdr1 promoter through interaction with the transcription factor Mrr1, facilitating fluconazole tolerance in resistant strains[34]. This observation demonstrates that SNF5/SWI/SNF participates in drug resistance mechanisms in pathogenic fungi, with potential implications for therapeutic targeting.
Conclusion
SNF5 represents a paradigm of functional specification within multi-subunit protein complexes, serving simultaneously as a structural hub that organizes the spatial arrangement of other complex components, a biochemical catalyst that couples ATP hydrolysis to productive nucleosome remodeling, a recruitment platform that links transcriptional activators to chromatin remodeling, and a regulatory sensor that integrates metabolic signals to direct transcriptional responses[1][3][8][10][29][31][33][42][55][56]. Through its conserved repeat domains and arginine-rich histone-binding surface, SNF5 directly engages nucleosomes and anchors them in place during the transient disruption of nucleosomal DNA that accompanies ATP-dependent remodeling[3][33][49]. The comprehensive loss of SNF5 function results in formation of "aberrant" SWI/SNF complexes that are unable to respond efficiently to transcriptional activators or catalyze productive nucleosome remodeling, demonstrating that SNF5 is not a dispensable accessory factor but rather a core component essential for complex function[8][12][22][31]. At the cellular level, SNF5 is required for activation of cell-type-specific genetic programs during differentiation, for coordination of metabolic adaptation during nutrient stress, for establishment of proper nucleosome positioning at promoter regions, and for execution of the complex gene expression changes required for cellular differentiation and development[1][13][42][43][46][55][56].
The exceptional conservation of SNF5 from yeast to humans, combined with the profound consequences of its loss in both model organisms and human disease, underscores the fundamental importance of this protein in eukaryotic biology. In mammals, the SMARCB1 ortholog functions as a bona fide tumor suppressor, and its biallelic loss drives development of highly aggressive pediatric rhabdoid tumors through mechanisms involving both loss of chromatin remodeling activity and disruption of BAF complex-mediated recruitment of transcriptional programs[29][51]. The discovery of germline SMARCB1 mutations in schwannomatosis and the identification of cancer-associated somatic mutations mapping to SMARCB1's arginine-rich histone-binding regions have further emphasized the tumor-suppressive functions of this protein[29][51]. Looking forward, a deeper understanding of SNF5/SMARCB1 function in chromatin remodeling and its loss in cancer development may provide insights into development of new therapeutic strategies for treating rhabdoid tumors and other cancers driven by BAF complex dysfunction.
References
The comprehensive analysis provided above draws extensively from the primary literature and structural studies cited throughout, including foundational work characterizing SNF5 function in yeast, cryo-electron microscopy structures of the SWI/SNF complex bound to nucleosomes, conditional genetic studies examining SNF5 roles in development and differentiation, mechanistic studies of nucleosome engagement and anchoring, and translational research on SMARCB1 loss in human cancers. This body of work collectively establishes SNF5 as a critical component of chromatin remodeling machinery with essential roles in transcriptional regulation, cellular differentiation, metabolic adaptation, and tumor suppression.
Citations
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1224692/
- https://digitalcommons.providence.org/publications/2298/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7319049/
- https://www.yeastgenome.org/locus/S000000493
- https://glycosmos.org/glycoproteins/P18480
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9839376/
- https://www.epicypher.com/resources/blog/the-nucleosome-acidic-patch-a-master-landing-dock-for-chromatin-regulators/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5424545/
- https://febs.onlinelibrary.wiley.com/doi/10.1111/j.1742-4658.2009.06979.x
- https://pubmed.ncbi.nlm.nih.gov/32188938/
- https://www.yeastgenome.org/locus/S000003712
- https://pubmed.ncbi.nlm.nih.gov/28249160/
- https://genesdev.cshlp.org/content/6/9/1707.short
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3489556/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC86073/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5338235/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5837817/
- https://pubmed.ncbi.nlm.nih.gov/27261671/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC51303/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5592666/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8551654/
- https://genesdev.cshlp.org/content/19/6/665.long
- https://royalsocietypublishing.org/rsob/article/14/10/240039/91419/Opening-and-changing-mammalian-SWI-SNF-complexes
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9332782/
- https://elifesciences.org/reviewed-preprints/94869
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10100857/
- https://www.tandfonline.com/doi/full/10.1080/21501203.2024.2425170
- https://pubs.acs.org/doi/10.1021/cr500350x
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4509781/
- https://pubmed.ncbi.nlm.nih.gov/2233708/
- https://www.yeastgenome.org/locus/S000000493/phenotype
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8890752/
- https://pubmed.ncbi.nlm.nih.gov/25735750/
- https://elifesciences.org/articles/54449
- https://www.pnas.org/doi/10.1073/pnas.1302209110
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11377671/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8184634/
- https://academic.oup.com/nar/article/39/19/8378/1173912
- https://www.science.org/doi/10.1126/science.adu5654
- https://pmc.ncbi.nlm.nih.gov/articles/PMC232553/
- https://www.uniprot.org/uniprotkb/P18480/entry
- https://pmc.ncbi.nlm.nih.gov/articles/PMC231185/