Deep Research
Falcon
(SWI1-deep-research-falcon.md)
Falcon
(SWI1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Saccharomyces cerevisiae SWI1 (UniProt P09547; YPL016W; ADR6/GAM3)
Scope and gene/protein identity verification (mandatory)
The target protein is Swi1, a core non-catalytic subunit of the yeast SWI/SNF ATP-dependent chromatin remodeling complex, encoded by SWI1/YPL016W and historically referred to as ADR6/GAM3 in genetic literature; this identity matches the UniProt description provided and is consistently described as a SWI/SNF subunit in modern work. (wendegatz2024transcriptionalactivationdomains pages 1-2, wang1994geneticinteractionsbetween pages 1-2)
1) Key concepts and definitions (current understanding)
SWI/SNF chromatin remodeling
SWI/SNF is an ATP-dependent chromatin remodeler that repositions and/or evicts nucleosomes to modulate access to DNA. In yeast, SWI/SNF remodeling at promoters is linked to formation/maintenance of nucleosome-depleted regions that facilitate transcription-factor and general transcription machinery access. (wendegatz2024transcriptionalactivationdomains pages 1-2)
Modular architecture: where Swi1 fits
A contemporary mechanistic framework divides SWI/SNF-family remodelers into (i) a motor module (ATPase; in yeast Snf2), (ii) an ARP (actin-related protein) module, and (iii) a substrate recruitment module (SRM) that contributes targeting/engagement with nucleosomal features. (chen2023mechanismofaction pages 1-2, he2021structureofthe pages 1-3)
Within this organization, Swi1 functions as an auxiliary recruitment/structural subunit rather than the catalytic ATPase: it helps position/organize recruitment elements and provides interfaces for activator- and DNA-associated contacts. (chen2023mechanismofaction pages 1-2, he2021structureofthe pages 1-3)
ARID domain (AT-rich interaction domain)
Swi1 contains an ARID (AT-rich interaction) domain, a conserved DNA-binding fold found across eukaryotes and frequently associated with transcriptional regulation/chromatin remodeling proteins. In the yeast Swi1 subunit, an ARID region has been experimentally mapped and reported to bind DNA non-specifically. (wendegatz2024transcriptionalactivationdomains pages 14-15, wendegatz2024transcriptionalactivationdomains pages 5-6)
2) Recent developments and latest research (prioritized 2023–2024)
2.1. Activator-binding interfaces in Swi1 (2024 primary study)
A 2024 study directly tested interactions between transcriptional activation domains (TADs) and yeast chromatin remodeler subunits, showing that Swi1 (along with other SWI/SNF subunits) can bind activator TADs, including the lipid-metabolism activator Ino2. (wendegatz2024transcriptionalactivationdomains pages 1-2, wendegatz2024transcriptionalactivationdomains pages 2-3)
Critically, this work refines domain-level functional annotation within Swi1: it reports the ARID domain spans ~aa 405–506, and maps an activator-interacting region in a broader internal segment (aa 329–657). Truncation mapping indicated that a fragment aa 428–606 retained efficient binding to an Ino2 activation domain, whereas a shorter fragment aa 329–531 (which includes the ARID) did not, implying that ARID is not sufficient for activator binding and that residues ~507–606 likely contribute a core activator-binding surface. (wendegatz2024transcriptionalactivationdomains pages 5-6)
2.2. Functional modularity and mechanistic framing (2023 review)
A 2023 review synthesizes modern structural/biochemical understanding of SWI/SNF-family mechanisms and emphasizes that yeast SWI/SNF comprises an Snf2 ATPase plus auxiliary subunits that govern targeting and nucleosome engagement; it also reports an estimate that yeast SWI/SNF regulates expression of ~5% of yeast genes. (chen2023mechanismofaction pages 1-2)
2.3. Authoritative perspective on centrality of Swi1 (2024 review)
A 2024 review that discusses SWI/SNF complexes across species characterizes Swi1 (yeast) as centrally located within SWI/SNF architecture, functioning as a “molecular nexus” in the complex body organization described by cryo-EM studies. (sailik2024openingandchanging pages 2-3)
3) Molecular function, domains, and interaction partners (functional annotation)
3.1. Primary molecular function of Swi1
Swi1’s primary function is structural and regulatory within the SWI/SNF complex, contributing to:
1) substrate recruitment (positioning near nucleosomal DNA and helping target SWI/SNF to promoters), and
2) coactivator/activator interfaces that enable transcription-factor-dependent SWI/SNF recruitment. (wendegatz2024transcriptionalactivationdomains pages 1-2, he2021structureofthe pages 1-3)
Swi1 is not itself described as an enzyme catalyzing a specific reaction; the ATP hydrolysis and mechanical remodeling is carried by the SWI/SNF ATPase (Snf2/Swi2 in yeast nomenclature). (chen2023mechanismofaction pages 1-2)
3.2. Domain architecture supported by experimental evidence
ARID domain: Swi1 contains an ARID (AT-rich interaction) domain mapped to aa ~405–506, reported to bind DNA non-specifically. (wendegatz2024transcriptionalactivationdomains pages 5-6)
Activator-binding region: Swi1’s internal region aa 329–657 is experimentally implicated in activator interactions, and mapping suggests a core region around aa 507–606 is important for Ino2 TAD binding (distinct from the ARID itself). (wendegatz2024transcriptionalactivationdomains pages 5-6)
3.3. Key interaction partners and interfaces
Transcriptional activators/TADs: Swi1 binds activation domains from multiple activators (historic and newly tested), and in 2024 it was shown to bind the Ino2 activation domains relevant to phospholipid gene expression. (wendegatz2024transcriptionalactivationdomains pages 2-3, wendegatz2024transcriptionalactivationdomains pages 1-2)
SWI/SNF subunits: Structural work indicates that the N-termini of Swi1 and Snf5 interact and that Swi1 contributes to the SRM DNA-binding lobe together with Snf6, Snf12, and Snf5 N-terminus. (he2021structureofthe pages 1-3)
4) Localization: where Swi1 acts in the cell
4.1. Nuclear/chromatin site of action
Functionally, SWI/SNF is a promoter-acting chromatin remodeler, and structural work directly visualizes SWI/SNF bound to a nucleosome, demonstrating its chromatin-associated mode of action. (he2021structureofthe pages 1-3, wendegatz2024transcriptionalactivationdomains pages 1-2)
4.2. Structural positioning relative to nucleosome and DNA
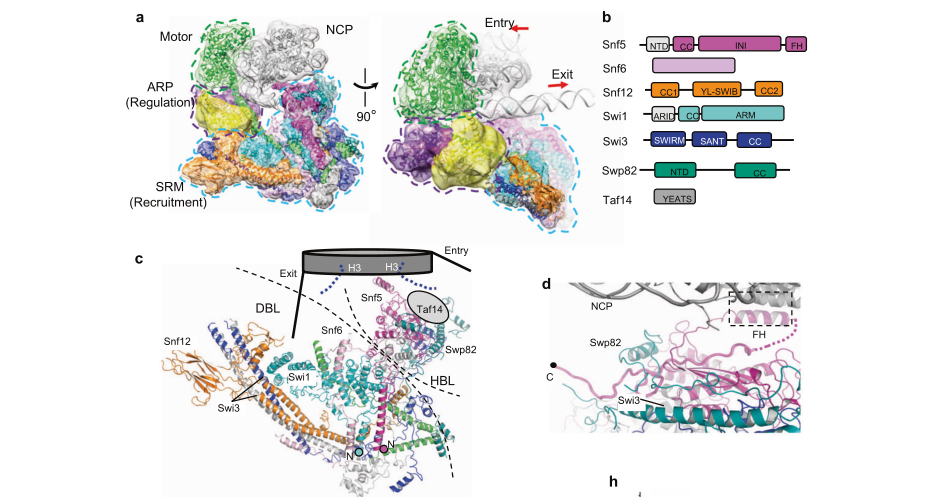
In the cryo-EM structure of yeast SWI/SNF bound to a mononucleosome, Swi1 (construct residues 251–1336) contributes to the substrate recruitment module (SRM) and specifically to the DNA-binding lobe (DBL) located near the exit DNA; Swi1 is proposed to help bind nucleosomal DNA either directly (via ARID) or indirectly (via coactivators). (he2021structureofthe pages 1-3)
A figure from this primary structure paper depicts the overall architecture/modules of nucleosome-bound SWI/SNF and the placement of Swi1 within the SRM/DBL in relation to the nucleosome. (he2021structureofthe media 010f0c81)
5) Biological processes and pathways linked to SWI1/SWI/SNF function
5.1. Transcriptional activation programs (classic genetics)
Genetic evidence places SWI1 within the SWI/SNF machinery required for transcriptional activation at specific loci. For example, SWI1 was reported to be required for expression of the a-specific gene STE6, positioning SWI1/SWI/SNF within mating-type gene regulation networks. (wang1994geneticinteractionsbetween pages 1-2)
5.2. Metabolic and stress-responsive gene regulation (modern framing)
Recent work emphasizes SWI/SNF’s general role in promoter remodeling and inducible gene activation programs. A 2024 mechanistic study connects SWI/SNF (including Swi1 interactions) to activator-dependent expression of phospholipid biosynthetic genes through Ino2 activation domains. (wendegatz2024transcriptionalactivationdomains pages 1-2)
Across yeast SWI/SNF literature, SWI/SNF defects are classically associated with failures in promoter induction/derepression at loci such as HO and SUC2 and impaired utilization of certain carbon sources (e.g., sucrose), consistent with a role in environmentally responsive transcription. (wendegatz2024transcriptionalactivationdomains pages 1-2)
6) Relevant statistics and data (recent and authoritative)
6.1. Fraction of genes regulated
A 2023 review reports an estimate that yeast SWI/SNF regulates expression of approximately ~5% of yeast genes. (chen2023mechanismofaction pages 1-2)
Consistent with this magnitude, the nucleosome-bound structure paper reiterates that SWI/SNF regulates ~5% of genes in yeast. (he2021structureofthe pages 1-3)
Separately, a 2024 paper cites microarray evidence that expression of ~1% of yeast protein-coding genes was significantly SWI-dependent (as reported in the cited microarray comparison). (wendegatz2024transcriptionalactivationdomains pages 1-2)
6.2. Quantitative structural and domain mapping data
Key quantitative structural/domain data for Swi1-supported annotation include:
- Swi1 residues 251–1336 used in cryo-EM reconstruction of nucleosome-bound SWI/SNF. (he2021structureofthe pages 1-3)
- Overall cryo-EM resolution ~6.9 Å, with higher local resolutions reported at the nucleosome and SRM. (he2021structureofthe pages 1-3)
- ARID domain ~aa 405–506, and an internal activator-interaction region aa 329–657 with a likely core around aa 507–606 for Ino2 TAD binding. (wendegatz2024transcriptionalactivationdomains pages 5-6)
7) Current applications and real-world implementations
Yeast SWI1/SWI/SNF is primarily used as a model system for mechanistic understanding of ATP-dependent chromatin remodeling and transcription-factor-dependent recruitment, including domain-level mapping of activator-binding interfaces. (wendegatz2024transcriptionalactivationdomains pages 1-2, he2021structureofthe pages 1-3)
Within the retrieved corpus, direct industrial or clinical implementations specifically targeting yeast SWI1 were not identified; however, cross-species reviews explicitly connect SWI/SNF structural/mechanistic insights (including yeast archetype studies) to broader biomedical contexts such as understanding SWI/SNF dysfunction in disease and therapeutic development in other organisms. (sailik2024openingandchanging pages 2-3)
8) Expert opinions and synthesis (authoritative interpretation)
Collectively, the most strongly supported functional interpretation is that Swi1 is a central architectural and targeting subunit within yeast SWI/SNF: it helps form the SRM recruitment surface near nucleosomal exit DNA and provides interfaces that bind transcriptional activation domains, thereby enabling promoter-specific recruitment and efficient remodeling by the Snf2 ATPase. This integrated view is supported by the convergence of (i) direct nucleosome-bound SWI/SNF structure defining Swi1’s physical placement and module membership, and (ii) 2024 biochemical interaction mapping showing Swi1 contains separable DNA- (ARID) and activator-binding functions. (he2021structureofthe pages 1-3, wendegatz2024transcriptionalactivationdomains pages 5-6, wendegatz2024transcriptionalactivationdomains pages 1-2)
Evidence summary table
The following evidence map summarizes the main supported annotations and quantitative points for Swi1/P09547.
| Category | Key points | Best supporting sources (citation IDs) | Publication year(s) and URL(s) when available |
|---|---|---|---|
| Identity | SWI1 in this report matches the Saccharomyces cerevisiae SWI/SNF chromatin-remodeling complex subunit encoded by YPL016W, also called ADR6/GAM3; it is a non-catalytic auxiliary subunit of the yeast SWI/SNF complex rather than the ATPase motor. | (wendegatz2024transcriptionalactivationdomains pages 1-2, wang1994geneticinteractionsbetween pages 1-2, chen2023mechanismofaction pages 1-2) | 2024 https://doi.org/10.1007/s00294-024-01300-x; 1994 https://doi.org/10.1007/BF00297274; 2023 https://doi.org/10.1080/19491034.2023.2165604 |
| Domains | Swi1 contains an ARID/AT-rich interaction domain spanning approximately aa 405-506; this domain has been reported to bind DNA non-specifically. An internal region aa 329-657 interacts with activators, and mapping suggests aa 507-606 contributes a core activator-binding region distinct from the ARID itself. | (wendegatz2024transcriptionalactivationdomains pages 14-15, wendegatz2024transcriptionalactivationdomains pages 5-6) | 2024 https://doi.org/10.1007/s00294-024-01300-x |
| Complex architecture | In cryo-EM reconstructions, Swi1 is a central structural subunit or “molecular nexus.” Its N-terminus contributes to the substrate recruitment module (SRM), specifically the DNA-binding lobe (DBL) near nucleosomal exit DNA. The DBL includes Snf6, Snf12, and the N-termini of Swi1 and Snf5. A truncated construct containing Swi1 residues 251-1336 was used for structure determination. | (he2021structureofthe pages 1-3, sailik2024openingandchanging pages 2-3, he2021structureofthe media 010f0c81) | 2021 https://doi.org/10.1038/s41421-021-00262-5; 2024 https://doi.org/10.1098/rsob.240039 |
| Recruitment & activator binding | Swi1 functions as a coactivator interface that helps recruit SWI/SNF to promoters through transcriptional activators. It has documented interactions with activation domains from Gcn4, VP16, Hap4, and in 2024 work with Ino2; GST pull-down mapping showed Swi1 fragments containing aa 428-606 bound Ino2 TAD1 efficiently, whereas aa 329-531 did not, implying the ARID alone is insufficient for activator binding. | (wendegatz2024transcriptionalactivationdomains pages 5-6, wendegatz2024transcriptionalactivationdomains pages 2-3, wendegatz2024transcriptionalactivationdomains pages 1-2) | 2024 https://doi.org/10.1007/s00294-024-01300-x |
| Biological processes | As part of SWI/SNF, Swi1 supports ATP-dependent chromatin remodeling that opens promoter chromatin and helps generate nucleosome-depleted regions for transcription initiation. Classic genetic evidence links SWI1 to activation of genes such as STE6 and to broader regulation of carbon-catabolite-responsive programs and other SWI/SNF-dependent genes. Yeast SWI/SNF overall has been estimated to regulate about 5% of genes. | (wendegatz2024transcriptionalactivationdomains pages 1-2, wang1994geneticinteractionsbetween pages 1-2, chen2023mechanismofaction pages 1-2) | 2024 https://doi.org/10.1007/s00294-024-01300-x; 1994 https://doi.org/10.1007/BF00297274; 2023 https://doi.org/10.1080/19491034.2023.2165604 |
| Localization & site of action | Swi1 acts in the nucleus, associated with the SWI/SNF complex on chromatin/nucleosomes. Structurally it is positioned in the recruitment portion of the complex adjacent to nucleosomal exit DNA, and its N-terminus is proposed to help contact DNA directly or indirectly via transcription coactivators. | (chen2023mechanismofaction pages 1-2, he2021structureofthe pages 1-3, he2021structureofthe media 010f0c81) | 2023 https://doi.org/10.1080/19491034.2023.2165604; 2021 https://doi.org/10.1038/s41421-021-00262-5 |
| Quantitative data | Quantitative details currently supported by available evidence include: yeast SWI/SNF has ~12 subunits; regulates about ~5% of yeast genes; cryo-EM map resolution was 6.9 Å overall, with local resolution around ~3.6 Å for the SRM and ~3.1 Å near the nucleosome-facing region; Swi1 ARID is aa 405-506; activator-interacting region aa 329-657; efficient Ino2-TAD1-binding fragment aa 428-606. | (wendegatz2024transcriptionalactivationdomains pages 5-6, wendegatz2024transcriptionalactivationdomains pages 1-2, chen2023mechanismofaction pages 1-2, he2021structureofthe pages 1-3) | 2024 https://doi.org/10.1007/s00294-024-01300-x; 2023 https://doi.org/10.1080/19491034.2023.2165604; 2021 https://doi.org/10.1038/s41421-021-00262-5 |
Table: This table compiles the currently supported functional annotation for Saccharomyces cerevisiae Swi1/P09547, including identity verification, domain organization, structural role in SWI/SNF, recruitment mechanisms, biological roles, localization, and quantitative findings. It is useful as a compact evidence map before writing the full narrative report.
Key sources (with dates and URLs)
- Wendegatz EC, Engelhardt M, Schüller H-J. Current Genetics (Sep 2024). “Transcriptional activation domains interact with ATPase subunits of yeast chromatin remodelling complexes SWI/SNF, RSC and INO80.” https://doi.org/10.1007/s00294-024-01300-x (wendegatz2024transcriptionalactivationdomains pages 1-2)
- Chen K, Yuan J, Sia Y, Chen Z. Nucleus (Jan 2023). “Mechanism of action of the SWI/SNF family complexes.” https://doi.org/10.1080/19491034.2023.2165604 (chen2023mechanismofaction pages 1-2)
- Abu Sailik F, Emerald BS, Ansari SA. Open Biology (Oct 2024). “Opening and changing: mammalian SWI/SNF complexes in organ development and carcinogenesis.” https://doi.org/10.1098/rsob.240039 (sailik2024openingandchanging pages 2-3)
- He Z, Chen K, Ye Y, Chen Z. Cell Discovery (Apr 2021). “Structure of the SWI/SNF complex bound to the nucleosome and insights into the functional modularity.” https://doi.org/10.1038/s41421-021-00262-5 (he2021structureofthe pages 1-3)
- Wang H, Reynolds-Hager L, Stillman DJ. Mol Gen Genet (Dec 1994). “Genetic interactions between SIN3 mutations and the Saccharomyces cerevisiae transcriptional activators encoded by MCM1, STE12, and SWI1.” https://doi.org/10.1007/BF00297274 (wang1994geneticinteractionsbetween pages 1-2)
References
-
(wendegatz2024transcriptionalactivationdomains pages 1-2): Eva-Carina Wendegatz, Maike Engelhardt, and Hans-Joachim Schüller. Transcriptional activation domains interact with atpase subunits of yeast chromatin remodelling complexes swi/snf, rsc and ino80. Current Genetics, Sep 2024. URL: https://doi.org/10.1007/s00294-024-01300-x, doi:10.1007/s00294-024-01300-x. This article has 3 citations and is from a peer-reviewed journal.
-
(wang1994geneticinteractionsbetween pages 1-2): H. Wang, L. Reynolds-Hager, and D. J. Stillman. Genetic interactions between sin3 mutations and the saccharomyces cerevisiae transcriptional activators encoded by mcm1, ste12, and swi1. Molecular and General Genetics MGG, 245:675-685, Dec 1994. URL: https://doi.org/10.1007/bf00297274, doi:10.1007/bf00297274. This article has 22 citations.
-
(chen2023mechanismofaction pages 1-2): Kangjing Chen, Junjie Yuan, Youyang Sia, and Zhucheng Chen. Mechanism of action of the swi/snf family complexes. Nucleus, Jan 2023. URL: https://doi.org/10.1080/19491034.2023.2165604, doi:10.1080/19491034.2023.2165604. This article has 55 citations and is from a peer-reviewed journal.
-
(he2021structureofthe pages 1-3): Zhenyu He, Kangjing Chen, Youpi Ye, and Zhucheng Chen. Structure of the swi/snf complex bound to the nucleosome and insights into the functional modularity. Cell Discovery, Apr 2021. URL: https://doi.org/10.1038/s41421-021-00262-5, doi:10.1038/s41421-021-00262-5. This article has 36 citations and is from a peer-reviewed journal.
-
(wendegatz2024transcriptionalactivationdomains pages 14-15): Eva-Carina Wendegatz, Maike Engelhardt, and Hans-Joachim Schüller. Transcriptional activation domains interact with atpase subunits of yeast chromatin remodelling complexes swi/snf, rsc and ino80. Current Genetics, Sep 2024. URL: https://doi.org/10.1007/s00294-024-01300-x, doi:10.1007/s00294-024-01300-x. This article has 3 citations and is from a peer-reviewed journal.
-
(wendegatz2024transcriptionalactivationdomains pages 5-6): Eva-Carina Wendegatz, Maike Engelhardt, and Hans-Joachim Schüller. Transcriptional activation domains interact with atpase subunits of yeast chromatin remodelling complexes swi/snf, rsc and ino80. Current Genetics, Sep 2024. URL: https://doi.org/10.1007/s00294-024-01300-x, doi:10.1007/s00294-024-01300-x. This article has 3 citations and is from a peer-reviewed journal.
-
(wendegatz2024transcriptionalactivationdomains pages 2-3): Eva-Carina Wendegatz, Maike Engelhardt, and Hans-Joachim Schüller. Transcriptional activation domains interact with atpase subunits of yeast chromatin remodelling complexes swi/snf, rsc and ino80. Current Genetics, Sep 2024. URL: https://doi.org/10.1007/s00294-024-01300-x, doi:10.1007/s00294-024-01300-x. This article has 3 citations and is from a peer-reviewed journal.
-
(sailik2024openingandchanging pages 2-3): Fadia Abu Sailik, Bright Starling Emerald, and Suraiya Anjum Ansari. Opening and changing: mammalian swi/snf complexes in organ development and carcinogenesis. Open Biology, Oct 2024. URL: https://doi.org/10.1098/rsob.240039, doi:10.1098/rsob.240039. This article has 5 citations and is from a peer-reviewed journal.
-
(he2021structureofthe media 010f0c81): Zhenyu He, Kangjing Chen, Youpi Ye, and Zhucheng Chen. Structure of the swi/snf complex bound to the nucleosome and insights into the functional modularity. Cell Discovery, Apr 2021. URL: https://doi.org/10.1038/s41421-021-00262-5, doi:10.1038/s41421-021-00262-5. This article has 36 citations and is from a peer-reviewed journal.
Artifacts
Citations
- wendegatz2024transcriptionalactivationdomains pages 1-2
- wendegatz2024transcriptionalactivationdomains pages 5-6
- chen2023mechanismofaction pages 1-2
- sailik2024openingandchanging pages 2-3
- he2021structureofthe pages 1-3
- wang1994geneticinteractionsbetween pages 1-2
- wendegatz2024transcriptionalactivationdomains pages 14-15
- wendegatz2024transcriptionalactivationdomains pages 2-3
- https://doi.org/10.1007/s00294-024-01300-x;
- https://doi.org/10.1007/BF00297274;
- https://doi.org/10.1080/19491034.2023.2165604
- https://doi.org/10.1007/s00294-024-01300-x
- https://doi.org/10.1038/s41421-021-00262-5;
- https://doi.org/10.1098/rsob.240039
- https://doi.org/10.1080/19491034.2023.2165604;
- https://doi.org/10.1038/s41421-021-00262-5
- https://doi.org/10.1007/BF00297274
- https://doi.org/10.1007/s00294-024-01300-x,
- https://doi.org/10.1007/bf00297274,
- https://doi.org/10.1080/19491034.2023.2165604,
- https://doi.org/10.1038/s41421-021-00262-5,
- https://doi.org/10.1098/rsob.240039,
Perplexity
(SWI1-deep-research-perplexity.md)
Perplexity
(SWI1-deep-research-perplexity.md)SWI1: A Critical Subunit of the SWI/SNF Chromatin Remodeling Complex in Saccharomyces cerevisiae
Executive Summary
SWI1 (also known as ADR6, GAM3, YPL016W) is a key subunit of the SWI/SNF ATP-dependent chromatin remodeling complex in the budding yeast Saccharomyces cerevisiae, where it serves as a regulatory component of one of the most extensively studied chromatin remodeling machines in eukaryotic cells[1][2]. The protein contains an AT-rich interaction domain (ARID) at its C-terminus and an asparagine/glutamine-rich prion domain at its N-terminus, structural features that determine both its normal biochemical function and its capacity to adopt an alternative conformational state as a prion[1][3]. Beyond its role in nucleosome remodeling and transcriptional regulation, SWI1 has emerged as a fascinating model system for understanding protein conformational diseases, as the prionogenic form [SWI+] exhibits inheritable phenotypic changes relevant to understanding epigenetic inheritance[1][4]. This report comprehensively examines SWI1's molecular structure, biochemical functions within the SWI/SNF complex, its role in regulating gene expression through chromatin remodeling, its interactions with transcriptional regulators, and the novel biology of its prion state, while emphasizing experimental evidence and providing specific mechanistic insights into how this protein contributes to fundamental cellular processes.
SWI1 Protein Structure and Domain Organization
Amino-Terminal Prion Domain and Asparagine-Rich Region
The SWI1 protein displays a tripartite structural organization that reflects its dual roles in chromatin remodeling and protein-protein interactions[1][3]. The amino-terminal region of SWI1 is notably enriched in asparagine residues, forming what is known as the prion determination domain (PrD), which has become the focus of extensive research into how proteins can adopt alternative conformational states[1][3]. More specifically, the extreme amino-terminus containing residues 1-38 consists of an exceptionally asparagine-rich region that is simultaneously glutamine-free, and this minimal region alone proves sufficient to co-aggregate with full-length SWI1 in cells carrying the [SWI+] prion conformation, maintain the prion fold independently, and transmit the prion conformational back to full-length Swi1[1]. This extraordinary property demonstrates that conformational information can be encoded in extremely short protein sequences, challenging traditional notions of the minimum length required for prion propagation[1]. The asparagine-rich nature of this region appears critical for the formation of amyloid-like fibrils that characterize the prion state, as recombinant N-terminal fragments of SWI1 have been shown to form amyloid fibrils in vitro with infectivity confirmed through fibril transformation experiments[1].
The middle Q region of SWI1, spanning a significant portion of the central protein sequence, is enriched in glutamine residues and appears to mediate specific protein interactions, potentially including those involving an actin-binding domain[1]. The structural organization of this region suggests roles beyond simple substrate binding, potentially facilitating the integration of SWI1 into multi-subunit complexes and mediating responses to cellular signals[1]. Interestingly, expression of constructs containing both the Q and C regions (QC) demonstrate a greater ability to rescue the growth defects observed in swi1Δ deletion mutants on non-glucose carbon sources compared to expression of the C region alone, indicating that the Q region contributes functionally to normal SWI1 activity even though it is not absolutely essential[1].
The C-Terminal ARID Domain and Functional Core
The carboxyl-terminal region of SWI1 contains the functional domain of the protein and is enriched in neither asparagine nor glutamine residues[1]. This region houses the ARID (AT-rich interaction domain), a structurally distinct helix-turn-helix motif-based DNA-binding domain that is found in numerous chromatin-associated proteins throughout eukaryotes[7][16]. The SWI1 ARID is highly unusual among known ARID family members in that it exhibits markedly weak DNA-binding affinity compared to its human counterparts p270 and ARID1B, as well as the prototypical ARID protein Dri from Drosophila[7][16][39]. This weak DNA-binding property arises from specific sequence variations in the major groove interaction site, particularly in Loop 2 and Helix 5 regions[7][16][39]. SWI1 uniquely lacks basic residues between the beginning of Loop 2 and an invariant tyrosine, and instead contains acidic residues in this critical region—a feature present in no other known ARID protein[7][16][39]. This combination of missing basic residues and the presence of negative charges from acidic amino acids substantially reduces the affinity of SWI1 for DNA, and site-directed mutagenesis experiments demonstrate that introducing basic residues into these positions can substantially improve DNA binding[7].
Despite this apparently disadvantageous weak DNA-binding property, the SWI1 ARID domain proves functionally important in the context of the intact SWI/SNF complex. The C-terminal region containing the ARID domain is sufficient to restore SWI1 function in yeast carrying deletion mutations or prion conformations, demonstrating its essential role in the complex's biological activity[1]. Furthermore, the structure of the SWI/SNF complex bound to nucleosomes reveals that the N-termini of Swi1 and Snf5 interact with each other and may bind to nucleosomal DNA either through the ARID domain directly or indirectly through recruitment of transcriptional coactivators, positioning this domain within a DNA-binding lobe that participates in nucleosome recognition[13]. The weak DNA-binding affinity of the isolated ARID domain appears to be a feature rather than a flaw, as it likely prevents non-specific DNA binding that could interfere with the complex's regulated targeting to specific genomic loci through transcriptional regulators.
The SWI/SNF Chromatin Remodeling Complex: Architecture and Function
Complex Composition and Modular Organization
The SWI/SNF chromatin remodeling complex in S. cerevisiae comprises 12 subunits that work together to remodel chromatin structure through ATP-dependent mechanisms[9][12]. The complex contains Snf2 (also known as Swi2), which serves as the catalytic motor protein, along with eleven auxiliary subunits including Swi1, Swi3, Snf5, Snf6, Snf12, Arp7, Arp9, Snf11, Swp82, and Taf14[9][12]. Functional and structural analyses have revealed that this complex possesses a tripartite modular architecture consisting of three functionally distinct modules: the motor module, the regulatory actin-related protein (ARP) module, and the substrate recruitment module (SRM)[9][12][32]. The motor module, centered on the Snf2 ATPase, provides the catalytic activity that drives nucleosome remodeling through ATP hydrolysis[12][32]. The regulatory ARP module, composed of actin and actin-related proteins, functions to regulate motor activity and stabilize the overall complex structure[9][12]. The substrate recruitment module, which is substantially larger and more complex, performs the critical function of integrating various chromatin clues—including transcription factor binding sites and histone post-translational modifications—to direct the motor activity to appropriate genomic locations[9][12][32].
Mass spectrometric analysis has revealed that the SWI/SNF complex exhibits functional modularity, with distinct subcomplexes that can be isolated and characterized[9]. These analyses identified four functionally distinct modules within the yeast complex: an Arp module, a catalytic module, a Snf5/Swi3 regulatory module, and a Swi1 module[9]. This modularity provides insights into how different subunits contribute to complex assembly and function, suggesting that some subunits maintain stable interactions with each other while others associate more transiently[9]. The identification of a discrete "Swi1 module" highlights the importance of this subunit as an organizing principle within the larger complex, potentially serving as a nucleation site for recruitment of other auxiliary subunits.
The Substrate Recruitment Module and SWI1's Role
Within the SWI/SNF complex, the substrate recruitment module is further delineated into distinct lobes that serve different functions in recognizing nucleosomal substrates[12][13][32]. The DNA-binding lobe (DBL) contains SWI1 along with other subunits including Snf6, Snf5, and Snf12, and these proteins directly or indirectly bind to the DNA components of nucleosomes[13][32]. The histone-binding lobe (HBL) contains elements that recognize histone tail modifications and the acidic patch of the histone octamer, particularly through the finger helix of Snf5[13]. The nucleosome-binding lobe (NBL) serves additional roles in nucleosome recognition and positioning[13]. The precise positioning of SWI1 within the DNA-binding lobe, as revealed by recent cryo-EM structures of the SWI/SNF-nucleosome complex at 6.9 Å overall resolution with local resolution improvements to 3.1 Å and 3.6 Å in critical regions, demonstrates that the N-terminal regions of both Swi1 and Snf5 interact with each other at the interface between the DNA-binding and catalytic modules[13]. This positioning places SWI1 in close proximity to the exit DNA of the nucleosome, suggesting that this subunit participates in recognizing linker DNA and potentially facilitating the recruitment of transcription factors to remodeled chromatin[13].
SWI1 DNA-Binding Properties and Their Functional Implications
Characterization of Weak DNA Binding
The DNA-binding properties of SWI1 represent a unique case study in understanding how evolutionary divergence of protein function can occur within conserved protein families[7][16][39]. When the DNA-binding affinity of SWI1 is directly compared to that of its human counterparts p270 (also called ARID1A) and ARID1B, as well as other ARID family members, SWI1 exhibits markedly lower affinity for DNA in quantitative binding assays[7][16][39]. In particular, while p270 and ARID1B show the same high-affinity binding characteristic of the prototypical ARID protein Dri from Drosophila, SWI1 binds DNA with substantially reduced affinity[7][16][39]. Most of the DNA bound by SWI1 in gel-shift assays is released by salt washing at 100 mM, and no evidence of sequence selectivity is observed, contrasting sharply with the behavior of sequence-specific ARID proteins[7][39]. This weak DNA-binding behavior has been demonstrated to be an intrinsic feature of the SWI1 ARID sequence rather than an artifact of experimental conditions, as site-directed mutagenesis studies show that introducing human ARID1A residues into the critical Loop 2 and Helix 5 regions substantially enhances DNA-binding affinity[7][39].
The structural basis for this weak DNA-binding property has been elucidated through detailed sequence comparisons and mutagenesis experiments[7][16][39]. The SWI1 ARID is particularly unusual in that Loop 2, the region predicted to participate in major groove contacts based on structures of other ARID proteins, is shorter in SWI1 than in other ARID family members and entirely lacks basic residues between its beginning and an invariant tyrosine residue[7][16][39]. Instead, this region contains acidic amino acids that would electrostatically disfavor DNA binding[7][16][39]. When four residues corresponding to missing loop residues are reintroduced into human p270 in the context of appropriate basic residues, the resulting chimera (p270.L2.TFT) still does not acquire sequence specificity, suggesting that the overall arrangement and flexibility of these regions, rather than individual amino acid substitutions, determines the DNA-binding properties[39]. The presence of negatively charged acidic residues at what should be a major groove contact site represents an unusual evolutionary divergence from other ARID family members and appears to actively suppress non-specific DNA binding in the isolated domain.
SWI1 DNA Binding Within the Complex Context
Despite the weak DNA-binding affinity of SWI1 in isolation, the intact SWI/SNF complex binds DNA with high affinity, indicating that other subunits contribute substantially to the DNA-binding capability of the complete enzyme[7][39]. In early UV crosslinking experiments, three components of the SWI/SNF complex were found to crosslink to naked DNA: SWI1 and two other components (p68 and p78), whose DNA-binding properties have not been further characterized[7][39]. In subsequent studies, several other members of the complex were found to crosslink with nucleosomal DNA, suggesting that multiple subunits contribute to DNA binding in the intact complex[7][39]. The interaction of yeast SWI/SNF complexes with naked DNA is distamycin-sensitive, indicating that binding occurs through minor groove interactions rather than through the major groove contacts typically used by ARID domains[7][23][39]. This observation is entirely consistent with the conclusion that the major groove contact region is not functionally conserved in the SWI1 ARID, as the complex apparently employs minor groove contacts through other subunits for non-specific DNA binding[7][23][39]. When the complex is bound to nucleosomes, however, it is not displaced by distamycin, indicating that other stabilizing interactions occur and that the nucleosomal context provides additional binding specificity through histone-protein contacts[7][23][39].
The functional role of the ARID domain in SWI1 therefore appears to extend beyond simple DNA binding. Deletion of the ARID region does not substantially impair the ability of yeast to grow in conditions requiring functional SWI/SNF or the ability of the isolated complex to remodel nucleosomes in vitro[7][16][39][49]. This observation suggests that while the ARID domain is present in all known SWI/SNF complexes across eukaryotes and likely has physiological importance, particularly in higher eukaryotes with more complex regulatory requirements, the weak DNA-binding affinity of the yeast domain may reflect a specialization for a different function, such as recruitment through protein-protein interactions with transcriptional regulators rather than through direct sequence-specific DNA binding[7][16][39].
Recruitment of SWI/SNF to Promoters and Target Genes
Transcription Factor-Dependent Recruitment
SWI/SNF is not recruited to all promoters in yeast but rather to a subset of genes requiring chromatin remodeling for optimal expression, and considerable evidence demonstrates that this targeting is achieved through direct or indirect interactions with gene-specific transcriptional activators[14][37][44]. Classical studies established that SWI/SNF can be recruited from yeast nuclear extracts by a DNA-bound transcriptional activator, such as Gal4 containing an acidic activation domain, without requiring an obligatory association with RNA polymerase II holoenzyme or general transcription factors[14][37]. When immobilized templates containing activator binding sites are incubated in nuclear extracts under conditions where preinitiation complex components are not sequestered, SWI/SNF subunits (such as Swi3 and Snf5) are efficiently recruited to the template by activators, and this recruitment is dependent on a functional activation domain but not on promoter sequences or TATA-binding protein[14][37]. Furthermore, the recruitment of SWI/SNF requires greater than 53 base pairs of DNA adjacent to the Gal4 binding site, suggesting that SWI/SNF does not interact stably with activators in solution but rather is targeted by DNA-bound activators[14][37].
At the GAL1 promoter specifically, both Swi1 and Snf5 subunits are efficiently recruited to the upstream activating sequence upon galactose induction, but this recruitment requires the integrity of the Mediator complex, TAFII subunits, and RNA polymerase II itself[44]. The observation that Swi/Snf recruitment at GAL1 does not require acetylation of chromatin by Gcn5 or the presence of SAGA complex components, despite the known role of histone acetylation in SWI/SNF recruitment at other genes, indicates that different genes employ distinct recruitment mechanisms[44]. Artificial tethering of the Mediator to GAL1 through a Gal4-Gal11 fusion protein is sufficient to recruit both Swi/Snf and SAGA complexes, demonstrating that Mediator interactions are sufficient for SWI/SNF targeting[44]. The fact that disruption of Mediator, TAFII subunits, or RNA polymerase II dramatically reduces SWI/SNF recruitment while SAGA recruitment is only modestly affected suggests that SWI/SNF has a particularly stringent requirement for Mediator-dependent recruitment at this locus[44].
Histone Acetylation and SWI/SNF Function
The relationship between histone acetylation and SWI/SNF activity is complex and context-dependent, with different genes employing different requirements for acetylation in SWI/SNF targeting and nucleosome remodeling[21][24][44][45]. At the HO promoter, histone acetylation by Gcn5 is required for SWI/SNF binding, and the Ash1 repressor protein apparently functions to inhibit SWI/SNF binding through effects on acetylation status[21]. In contrast, at the GAL1 locus, histone acetylation by Gcn5 is not essential for SWI/SNF recruitment[44]. At the SUC2 gene, which is induced through glucose starvation and uses metabolic signals to regulate expression, both Gcn5 and Esa1 histone acetyltransferases enhance the binding of SWI/SNF to the promoter, with Gcn5 being recruited concurrently with SWI/SNF while Esa1 associates constitutively with the promoter[45]. The Snf1 kinase, which is activated in response to glucose starvation, regulates the association of SWI/SNF and Gcn5 with the SUC2 promoter, linking nutrient-sensing pathways to chromatin remodeling[45].
The mechanism by which SAGA-acetylated nucleosomes are preferentially displaced by SWI/SNF has been characterized through biochemical studies[24]. When nucleosome arrays are acetylated by the SAGA complex in the presence of bound transcription activators, SWI/SNF treatment selectively displaces acetylated histones from the array in an ATP-dependent manner, preferentially removing SAGA-acetylated histones relative to total histones[24]. The Swi2/Snf2 bromodomain, an acetyl-lysine binding module located at the C-terminus of the motor subunit, plays a role in this displacement activity[24]. These observations suggest that targeted histone acetylation by SAGA predisposes promoter nucleosomes for displacement by SWI/SNF, creating a two-step mechanism for chromatin remodeling in which acetylation is followed by physical nucleosome movement[24].
SWI1 Protein Interactions and Complex Assembly
Interaction with Other SWI/SNF Subunits
SWI1 makes multiple critical interactions with other subunits that are essential for complex assembly and function[9][13]. The N-termini of SWI1 and Snf5 interact with each other at the interface between the DNA-binding lobe and the catalytic motor module[13]. This interaction appears to be central to proper positioning of both subunits within the substrate recruitment module[13]. Mass spectrometric analysis of complexes purified from subunit deletion strains reveals that SWI1 associates most strongly with certain other subunits while maintaining only weak associations with others, indicative of a modular architecture in which SWI1 forms part of a discrete functional unit[9]. The interaction of SWI1 with the motor subunit Snf2 is mediated through multiple contact points, with the NTD and preHSA domains of Snf2 interacting with targeting subunits including SWI1[12].
The stability and composition of the SWI/SNF complex depends on the presence of specific subunits, as demonstrated by analysis of deletion mutants[9]. In swi1 deletion strains, the overall integrity of the complex is maintained but its targeting and activity are substantially compromised[9]. Conversely, deletion of certain other subunits leads to reduced occupancy of the catalytic Snf2 subunit at target genes, suggesting that auxiliary subunits regulate the functional activity of the motor[9]. The modular nature of the complex means that different combinations of subunits can associate to form partially functional subcomplexes with different properties and functions[9].
Interaction with Transcriptional Activators
SWI/SNF interacts with numerous transcriptional activators beyond Gal4, including Rlm1, which responds to cell wall stress[20]. The interaction between Rlm1 and SWI/SNF occurs in a stress-dependent manner, and this physical interaction directs SWI/SNF to cell wall integrity pathway-responsive genes[20]. Purified SWI/SNF complex clearly interacts with Rlm1 in vitro, and the DNA-binding domain of Rlm1 is not required for this association, indicating that the interaction occurs through other domains[20]. The SWI/SNF complex also associates with c-MYC, a prototypical transcriptional activator, and mutations in SNF5 or BRG1 abolish the ability of c-MYC to activate transcription[20]. These various activator-SWI/SNF interactions demonstrate that the complex serves as a general platform for recruitment to genes activated by diverse transcriptional regulators, with specificity in recruitment likely determined by the particular activators bound to promoter elements.
The Prion Form of SWI1: [SWI+] and Epigenetic Inheritance
Discovery and Characterization of [SWI+]
One of the most remarkable discoveries in the study of SWI1 is that the protein can undergo a conformational change to form a prion, known as [SWI+], which exhibits inheritable phenotypic properties without changes in DNA sequence[1][3][4][30][43]. This prion represents one of several endogenous yeast prions that function as epigenetic elements transmitted as altered protein conformations and manifested as heritable phenotypic traits[1]. The discovery that Swi1 overexpression could induce [PSI+], another yeast prion, through prion-prion interactions led researchers to hypothesize that Swi1 itself might be capable of forming a prion[1]. By co-overexpressing SWI1 and SUP35 constructs, researchers generated [PSI+] cells that were further screened for [SWI+] by scoring their ability to use raffinose as a carbon source[1].
The characterization of [SWI+] revealed that the prionization of Swi1 leads to a variety of phenotypes reflecting disrupted function of the SWI/SNF complex[1][30][43]. The primary phenotype of [SWI+] cells is poor growth on non-glucose carbon sources including raffinose, galactose, glycerol, and sucrose[1][30]. [SWI+] cells grow at a significantly slower rate on raffinose media compared to [swi−] cells, providing a partial loss-of-function phenotype[1][30]. Additionally, [SWI+] cells display abolishment of multicellular features, including loss of yeast flocculation and the ability to form multicellular aggregates[1][30]. These phenotypic changes have implications for understanding [SWI+] as either detrimental or potentially beneficial to yeast under certain environmental circumstances[1].
Prion Domain Structure and Transmission
The prion domain of SWI1 consists of the asparagine-rich N-terminal region, which fulfills the definition of a prion determination domain in containing regions enriched in polar residues such as glutamines and asparagines[1][3][43][46]. The minimum prion domain was identified through systematic truncation experiments, which revealed that the extreme amino-terminal of Swi1 acts as a major determinant for [SWI+][1][43]. Importantly, a small region (amino acids 1-38 or even as short as 1-32) could transmit [SWI+] back to full-length Swi1 when provided in trans, and these minimal regions could co-aggregate with full-length Swi1 in [SWI+] cells, maintain the prion fold in the absence of full-length protein, and transmit the prion conformation back to full-length protein[1][43]. The extreme asparagine-richness of the Swi1 1-38 region, combined with its glutamine-free composition, makes it unique among characterized protein regions[1].
Recent bioinformatics approaches have predicted and confirmed the existence of an additional amyloid-forming region within the Swi1 prion domain (residues 239-259) that lies beyond the extensively characterized amino-terminal region[1]. It remains to be determined whether this additional region is independently prionogenic or whether it contributes to [SWI+] formation and propagation events[1]. The identification of multiple prion-forming regions within a single protein domain suggests that prion formation may involve cooperative interactions between distinct structural elements[1].
Recombinant N-region fragments of Swi1 have been demonstrated to form amyloid fibrils in vitro, and these fibrils show infectivity as measured through fibril transformation experiments, providing biochemical evidence that the N region contains the prion-determining sequences[1][43]. The formation of these fibrils likely involves the conversion of normally soluble protein into an ordered, β-sheet-rich amyloid structure that can template the conformational change of newly synthesized protein molecules[1][43].
Phenotypic Consequences of [SWI+]
The phenotypic effects of [SWI+] prionization reflect the normal functions of Swi1 within the SWI/SNF complex[1][30][43]. Prionization of Swi1 appears to lead to loss or substantial reduction of function, as [SWI+] cells exhibit many of the same defects observed in swi1Δ deletion mutants, including inability to utilize non-glucose carbon sources efficiently[1][30][43]. The regulation of flocculin (FLO) genes by SWI/SNF, and specifically by Swi1, provides a molecular mechanism for [SWI+] to abolish multicellular features[1][30]. In the presence of [SWI+], FLO gene transcription is severely reduced or abolished, and interestingly, other FLO gene transcriptional activators such as Mss11 and Sap30 co-aggregate with Swi1 and are titrated away by [SWI+], causing complete abolishment rather than mere reduction of multicellular features[1][30]. This elegant regulatory scheme allows [SWI+] to coordinate the expression of multiple genes involved in multicellular growth through sequestration of their transcriptional regulators.
The increased conversion frequency of [SWI+] compared to other yeast prions such as [PSI+] suggests that the ability to gain and lose this prion state rapidly may provide yeast with flexibility and capacity to adapt to environmental changes[1][30]. The loss of multicellular features through [SWI+] formation allows yeast cells to transition from a multicellular growth form to a unicellular form capable of rapid migration and multiplication[1][30]. Recent research has demonstrated that the loss of multicellular features through [SWI+] enables enhanced mobility of yeast cells, suggesting an adaptive advantage for prion formation under certain environmental conditions[1][30].
Maintenance and Propagation of [SWI+]
The maintenance and propagation of [SWI+] depend critically on the Hsp40/Hsp70 chaperone system, which is required for converting newly synthesized protein into the prion conformation[1][30]. The Sse1 and Sse2 heat shock proteins (members of the Hsp70 family) play sensitive roles in [SWI+] maintenance and propagation, with overexpression of Sse1 hindering curing of the prion via guanidinium chloride treatment[1][30]. This dependence on the Hsp40/Hsp70 system is particularly stringent for [SWI+] compared to other yeast prions, and this sensitivity may vary strain-to-strain[1][30]. The dominant negative Ssa1-21 variant, when expressed, manages to cure [SWI+] in all observed colonies, indicating that even partial disruption of the Hsp70 system can eliminate prion propagation[1][30]. This unique sensitivity of [SWI+] to disruptions of the chaperone system may allow for more rapid adaptation to environmental changes through rapid loss of the prion state[1][30].
SWI1 Function in Metabolic and Mating Type Regulation
Carbon Source Utilization and Metabolic Gene Regulation
The original discovery of the SWI/SNF complex was made through genetic screens for mutations affecting the expression of genes involved in carbon source metabolism, particularly sucrose fermentation and the metabolism of alternative carbon sources[26]. SWI1 is essential for transcription of genes required for utilizing non-glucose carbon sources, including those encoding invertase (SUC2) and inositol metabolizing enzymes[1][15][26]. The regulation of these genes represents a paradigmatic example of how SWI/SNF-mediated chromatin remodeling contributes to cellular adaptation to changing nutrient conditions[2][36][50].
The relationship between chromatin remodeling and metabolism has been extensively characterized, with recent studies demonstrating that the SWI/SNF complex serves as a critical bridge between metabolic signals and transcriptional responses[2][36][50]. The mechanism by which SWI/SNF activates transcription of metabolic genes involves modifying promoter chromatin structure and facilitating activator binding, as demonstrated through studies of the GAL1 and SUC2 promoters[2][36][50]. Both ATP and acetyl-CoA, key metabolic cofactors, are required for the ATPase activity of SWI/SNF and for histone acetylation, respectively[36][50]. This direct connection between metabolic status and chromatin remodeling activity suggests that SWI/SNF can sense cellular metabolic state through the availability of these cofactors[36][50]. During starvation, dynamic deacetylation of histones and reallocation of acetyl-CoA away from chromatin modifications could affect SWI/SNF activity both through removing the acetylated histone marks that the complex recognizes via its bromodomain and through reducing the ATP levels required for nucleosome remodeling[36][50].
Mating Type Switching and Cellular Memory
SWI1 is essential for proper regulation of the HO endonuclease gene, which catalyzes the programmed recombination event that switches yeast mating type[1][15][26]. Mating type switching requires SWI/SNF-mediated chromatin remodeling at the HO promoter to allow expression of the endonuclease that catalyzes the mating type locus switching reaction[1][26]. The prionization of Swi1 in the form of [SWI+] leads to a likely decrease in HO expression and reduced mating-type switching frequency[1][30]. Remarkably, [SWI+] spores demonstrate an increased rate of out-crossing (mating of mother cells with non-daughter cells) compared to [swi−] spores, suggesting that loss of the ability to perform mating-type switching creates a scenario in which cells must seek mates outside the mother-daughter relationship[1][30]. This increased out-crossing effectively increases genetic variation in the population and reduces inbreeding, potentially providing an evolutionary advantage to prion-containing cells in diverse environments[1][30]. This phenomenon illustrates how a protein conformational disease can have population-level effects on sexual reproduction and genetic variation in yeast.
SWI1 Function in Stress Response and Gene Regulation
Heat Shock and Stress Response Gene Activation
SWI/SNF plays critical roles in the transcriptional response to cellular stress, including heat shock, and SWI1 as a component of this complex participates in both the activation and repression of stress-responsive genes[56][59]. The relationship between nucleosome occupancy, histone acetylation, and gene expression differs between activation and repression during heat shock, with strongly upregulated genes showing reduction in nucleosome occupancy and higher histone H4 acetylation at their promoters[56]. The promoters of repressed genes show an inverse effect, with decrease in histone H4 acetylation being more pronounced than increase in nucleosome occupancy[56]. During the rapid heat shock response, reduction in nucleosome occupancy relative to histone acetylation at activated genes is stronger than that seen for slower stationary-phase stress responses, suggesting that rapid clearance of nucleosomes from promoters is particularly important for transcriptional activation following heat shock[56].
In snf2Δ deletion mutants, the histone H4 occupancy changes observed in wild-type cells following heat shock are substantially altered, with an initial increase in histone H4 occupancy 5 seconds after heat shock that is not seen in wild-type cells[56]. This observation, replicated across three independent biological replicates and hundreds of genes at multiple timepoints, demonstrates that although the histone H4 occupancy changes were small, they were consistent and indicate that SWI/SNF is required for appropriate nucleosome dynamics during the heat shock response[56]. The SWI/SNF complex is required for normal activation as well as repression of genes during heat shock, and loss of SWI/SNF delays chromatin remodeling in response to stress conditions[56].
Cell Wall Integrity Pathway and Rlm1-SWI/SNF Interactions
The cell wall integrity (CWI) pathway represents another important regulatory context in which SWI1 functions through the SWI/SNF complex[20]. The Slt2/Mpk1 mitogen-activated protein kinase (MAPK) of the CWI pathway coordinates the transcriptional program triggered by cell wall stress, primarily through the transcription factor Rlm1[20]. SWI/SNF plays a critical role in orchestrating this transcriptional response, with swi/snf mutants showing drastically reduced expression of cell wall stress-responsive genes and hypersensitivity to cell wall-interfering compounds[20]. Upon cell wall stress, binding of RNA polymerase II to the promoters of cell wall stress-responsive genes depends on Rlm1, Slt2, and SWI/SNF, and Rlm1 physically interacts with SWI/SNF to direct its association to target promoters[20]. Nucleosome displacement at cell wall integrity-responsive genes such as MLP1/KDX1 occurs in a manner dependent on the SWI/SNF complex, and recruitment of SWI/SNF depends on both MAPK-dependent phosphorylation of Rlm1 and the presence of active SWI/SNF[20]. These findings identify SWI/SNF as a key element of the CWI MAPK pathway that mediates the chromatin remodeling necessary for adequate transcriptional response to cell wall stress[20].
SWI1 Interactions with Broader Cellular Machinery
SWI/SNF and RNA Polymerase I Transcription
While SWI/SNF is primarily studied in the context of RNA polymerase II transcription, emerging evidence indicates that this complex also plays important roles in transcription by RNA polymerase I[48]. The SWI/SNF complex influences transcription of ribosomal RNA genes by the most robust transcription machinery in proliferating cells, with deletion of SNF5 or SNF6 being lethal in strains impaired for transcription initiation and elongation by Pol I[48]. Chromatin immunoprecipitation analysis shows that several subunits of SWI/SNF associate with the ribosomal DNA (rDNA) locus, including Snf2, Snf5, and Snf6[48]. Deletion of SNF6 specifically leads to approximately 2.5-fold less Pol I transcription compared to wild-type cells, and this reduction appears to reflect a role for SWI/SNF in Pol I transcription elongation[48]. These observations suggest that SWI/SNF may directly affect rDNA chromatin structure to facilitate Pol I transcription elongation[48].
SWI/SNF and RNA Polymerase II Promoter-Proximal Pausing
Recent studies have demonstrated that mammalian SWI/SNF subunits, including the human orthologs of Swi1 (ARID1A and ARID1B), play roles in regulating RNA polymerase II promoter-proximal pausing, a widespread regulatory mechanism controlling gene expression[51][54]. While this has been more extensively characterized in mammalian cells, the conservation of SWI/SNF architecture and function suggests that similar mechanisms may operate in yeast[51][54]. Promoter-proximal pausing allows polymerase II to be poised for rapid activation in response to regulatory signals, and SWI/SNF family proteins appear to influence the transition between paused and actively elongating states[51][54].
Evolutionary Conservation and Mammalian Orthologs
ARID1A and ARID1B as Human SWI1 Orthologs
The SWI/SNF complex is highly conserved across eukaryotes, from yeast to humans, with remarkable preservation of both structure and function despite hundreds of millions of years of evolution[2][35][51][54]. The yeast SWI1 subunit has two closely related human orthologs: p270 (also called ARID1A) and ARID1B (also called BAF180)[51][54]. These human ARID-containing subunits are conserved throughout metazoans and are expressed across most human cells and tissues[51][54]. Both ARID1A and ARID1B encode for the two largest, mutually exclusive subunits of the mammalian SWI/SNF complex (known as BAF or PBAF depending on composition), meaning that a given complex contains either ARID1A or ARID1B but not both[51][54].
Mutations in ARID1A and especially ARID1B are associated with a wide array of developmental disorders and cancers, suggesting that these subunits are implicated in maintaining cell identity and determining cell fate[51][54]. Genetic studies in mammalian systems have revealed that ARID1A and ARID1B are conserved in function as well as sequence, playing roles in the regulation of transcription factor binding, histone modifications, and the integration of metabolic signals with gene expression[51][54]. Unlike the weak DNA-binding property of yeast SWI1, the human ARID1A and ARID1B proteins retain relatively high-affinity DNA binding, reflecting a divergence in function between the yeast and mammalian orthologs[7][16][39].
Conservation of Complex Architecture and Function
The overall architecture of the SWI/SNF complex has been conserved from yeast to mammals with remarkable fidelity[2][9][12][32][35]. The tripartite modular organization consisting of motor, regulatory, and substrate recruitment modules is present in all characterized SWI/SNF family complexes, from the yeast SWI/SNF and RSC complexes to the mammalian BAF and PBAF complexes[12][32]. The motor subunits (Snf2 in yeast, Brg1 and Brm in mammals) retain their catalytic properties and overall domain architecture despite sequence divergence[12][32]. The regulatory ARP modules are similarly conserved, with actin and actin-related proteins serving roles in stabilizing and regulating motor activity across all complexes[12][32].
The substrate recruitment modules display the greatest diversity across different complexes, with mammalian complexes containing additional subunits not present in yeast[12][32][35]. However, the functional principle remains constant: auxiliary subunits recognize various chromatin features, including transcription factor binding sites, histone modifications, and DNA sequences, to direct the motor activity to appropriate genomic locations[12][32][35]. This conservation of core function alongside divergence in specific components reflects how evolution has modified a fundamental cellular machine to accommodate the more complex transcriptional regulatory demands of multicellular organisms[12][32][35].
Cellular Localization and In Vivo Function
Nuclear Localization and Chromatin Association
SWI1 is a nuclear protein that functions as a component of the SWI/SNF chromatin remodeling complex[1][8][18][22]. The protein is located in the nucleus throughout the cell cycle and associates with chromatin in a manner that depends on the cellular context and the genes being regulated[1]. At specific target genes, SWI1 and other SWI/SNF components are recruited in response to activator binding and appropriate cellular signals, with occupancy patterns reflecting the dynamics of transcriptional regulation[1][8]. The complex can associate transiently with chromatin at promoters that require remodeling for activation, or maintain more stable associations at genes subject to continuous regulation[1].
In the context of fission yeast (Schizosaccharomyces pombe), a Swi1 ortholog with related functions has been identified as a component of replication fork protection complexes, where Swi1 is required for programmed pausing of replication forks near the mat1 locus and for activation of the replication checkpoint kinase Cds1[19]. This dual function in both transcriptional regulation and DNA replication checkpoint control represents an interesting example of how related proteins with sequence conservation can acquire specialized functions in different cellular contexts[19]. The Swi1 ortholog copurifies with Swi3, and this Swi1-Swi3 complex associates with chromatin specifically in S phase, indicating temporal regulation of complex function[19].
Metabolic Integration and Post-translational Modification
SWI1 Acetylation and Metabolic Regulation
Recent studies have revealed that the SWI1 subunit itself undergoes post-translational modification that affects SWI/SNF function in response to metabolic status[36][50]. The yeast SWI/SNF complex itself is acetylated, and this acetylation affects its function in response to stress[36][50]. The Snf2 ATPase subunit is a target of the methyltransferase Hmt1, and loss of Hmt1 catalytic activity correlates with reduced Snf2-mediated promoter nucleosome remodeling[36][50]. Since the cofactor required for methylation by Hmt1 is a direct product of sulfur metabolism, this finding suggests an intriguing link between sulfur starvation and SWI/SNF function[36][50]. These observations underscore the complexity of the intersection between chromatin remodeling and metabolism, highlighting how metabolic status can be directly communicated to chromatin remodeling complexes through post-translational modification of complex subunits[36][50].
SWI1, like the Snf5 subunit, possesses a glutamine and asparagine-rich N-terminus that may be subject to conformational changes in response to pH alterations[36][50]. Changes in intracellular pH may impact the conformation of these glutamine-rich regions, presenting opportunities for novel protein interactions and directly linking nutritional status to transcription via intracellular conditions[36][50]. This molecular sensing mechanism allows SWI/SNF to integrate multiple layers of cellular information—from transcription factor binding to metabolic cofactor availability to pH changes—into appropriate transcriptional responses[36][50].
Biological Significance and Cellular Context
Gene Expression Control and the "5% Rule"
The SWI/SNF complex in yeast regulates the expression of approximately 5% of the yeast genome[12][32][48]. This relatively modest fraction of affected genes reflects the fact that SWI/SNF is not required for the expression of most constitutively expressed genes, but rather plays a critical role in the transcriptional regulation of genes activated in response to specific developmental or environmental signals[12][32][48]. The genes controlled by SWI/SNF include those involved in carbon source metabolism, mating-type switching, response to stress conditions, cell wall integrity, and various other adaptive responses[1][12][26][32][48]. The requirement for SWI/SNF at specific genes is gene-specific and dependent on the particular promoter context, chromatin structure, and transcriptional regulators involved[9].
Modulation of Chromatin Structure and Transcriptional Accessibility
The fundamental biochemical role of SWI1 within the SWI/SNF complex is to participate in modulating chromatin structure through ATP-dependent nucleosome remodeling[2][12][32][36][50]. The complex can slide, eject, or restructure nucleosomes, thereby altering the accessibility of DNA to transcription factors and RNA polymerase machinery[2][12][32]. This nucleosome remodeling is not a random event but rather a highly regulated process directed to specific genomic locations by interactions between SWI/SNF and sequence-specific transcription factors, by recognition of specific histone modifications, and by integration of signals reflecting cellular metabolic and developmental status[2][12][32][36][50].
Conclusion
SWI1 represents a multifunctional protein that exemplifies how cellular regulation is achieved through the integration of protein function, conformational dynamics, metabolic signaling, and epigenetic mechanisms. As a key component of the SWI/SNF chromatin remodeling complex, SWI1 contributes to the regulated expression of genes involved in fundamental cellular processes including carbon source utilization, mating-type switching, stress response, and DNA replication[1][2][12][26][32][36]. The weakness of SWI1's DNA-binding domain, seemingly disadvantageous compared to its human orthologs, appears to be a specialized adaptation that prevents non-specific DNA interactions and instead favors recruitment through protein-protein interactions with transcriptional regulators, directing chromatin remodeling activity to physiologically appropriate locations[7][16][39]. The discovery that SWI1 can undergo conformational changes to form a prion ([SWI+]) has revealed an entirely new dimension of protein regulation, demonstrating that conformational plasticity in the asparagine-rich N-terminal domain can create heritable phenotypic variants that respond to environmental changes through rapid interconversion between conformational states[1][3][4][30][43]. The connection between SWI1 function and metabolic status, achieved through both direct integration of metabolic cofactors into complex activity and through post-translational modifications that reflect metabolic state, illustrates how chromatin remodeling complexes serve as sensors of cellular metabolic condition[2][36][50]. Future research into SWI1 will likely continue to reveal additional layers of regulation and function, particularly as structural biology advances illuminate the precise molecular mechanisms by which this subunit contributes to nucleosome recognition and the coordination of chromatin remodeling activity with transcriptional regulation[12][13]. The evolutionary conservation of SWI1-like proteins from yeast to humans further underscores the fundamental importance of this protein in eukaryotic gene regulation and suggests that understanding SWI1 biology in yeast will continue to provide insights into the function of the mammalian BAF complex in development and disease[51][54].
Citations
- https://pmc.ncbi.nlm.nih.gov/articles/PMC6001882/
- https://pubmed.ncbi.nlm.nih.gov/38666605/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7346553/
- https://www.yeastgenome.org/locus/S000005937
- https://www.yeastgenome.org/complex/CPX-1150
- https://academic.oup.com/nar/article/doi/10.1093/nar/gkaf150/8074816
- https://pmc.ncbi.nlm.nih.gov/articles/PMC390273/
- https://thebiogrid.org/36161/summary/saccharomyces-cerevisiae/swi1.html
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5837817/
- https://glygen.org/protein/P09547
- http://chemgrid.org/tools/tmp_gene.php?s=4932&net=gen
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9839376/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8079448/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC317021/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7736884/
- https://academic.oup.com/nar/article/32/4/1345/1038431
- https://academic.oup.com/nar/article/35/16/5520/1280923
- https://www.uniprot.org/uniprotkb/P09547/entry
- https://pmc.ncbi.nlm.nih.gov/articles/PMC516732/
- https://www.molbiolcell.org/doi/10.1091/mbc.e12-04-0278
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1489090/
- https://www.pnas.org/doi/pdf/10.1073/pnas.91.8.2886
- https://journals.asm.org/doi/10.1128/ec.00165-06
- https://pubmed.ncbi.nlm.nih.gov/23202585/
- https://pubmed.ncbi.nlm.nih.gov/1339306/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5615180/
- https://www.science.org/doi/10.1126/science.adu5654
- https://onlinelibrary.wiley.com/doi/10.1046/j.1365-2958.1999.01390.x
- https://academic.oup.com/femsyr/article/18/6/foy061/5004851
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1459672/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC398327/
- https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2014.00190/full
- https://royalsocietypublishing.org/rsob/article/14/10/240039/91419/Opening-and-changing-mammalian-SWI-SNF-complexes
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11346436/
- https://genesdev.cshlp.org/content/13/18/2369.long
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3276066/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2715812/
- https://elifesciences.org/articles/37774
- https://academic.oup.com/nar/article/33/1/66/2401007
- https://pmc.ncbi.nlm.nih.gov/articles/PMC524348/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1271673/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2950522/
- https://journals.plos.org/plosone/article?id=10.1371%2Fjournal.pone.0056793
- https://portlandpress.com/biochemsoctrans/article/52/3/1327/234375/The-SWI-SNF-chromatin-remodeling-complex-a
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7969888/
- https://aacrjournals.org/cgd/article/13/3/95/705346/ARID-ProteinsA-Diverse-Family-of-DNA-Binding
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3003194/
- https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2021.643361/full
- https://mobidb.org/entries/P09547
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2268435/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3431495/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC85505/
- https://onlinelibrary.wiley.com/doi/10.1111/tpj.15159