Deep Research
Falcon
(SWI2-deep-research-falcon.md)
Falcon
(SWI2-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Saccharomyces cerevisiae SNF2 (SWI2/SNF2; YOR290C; UniProt P22082)
Executive summary
Saccharomyces cerevisiae Snf2 (a.k.a. Swi2; gene YOR290C; UniProt P22082) is the catalytic ATPase (“motor”) subunit of the yeast SWI/SNF chromatin-remodeling complex. Its core molecular function is ATP-dependent DNA translocation on nucleosomal substrates to reposition or destabilize nucleosomes, enabling both transcriptional activation and transcription-associated repression mechanisms (including repression by remodeling downstream of active promoters). Snf2 activity is regulated by multiple non-catalytic domains—including a conserved SnAC coupling domain and a C-terminal AT-hook module—that tune ATP hydrolysis and its coupling to nucleosome movement. (sen2011anewhighly pages 1-2, eustermann2024energydrivengenomeregulation pages 1-3, saha2023theathookis pages 1-2, morse2024swisnfchromatinremodeling pages 8-12)
1) Key concepts and definitions (current understanding)
1.1 ATP-dependent chromatin remodeling and SWI/SNF
ATP-dependent chromatin remodelers are multi-subunit machines that use ATP hydrolysis to alter nucleosome organization on DNA. In the SWI/SNF family, the principal outcomes include nucleosome sliding and histone ejection, mediated by a dedicated “motor ATPase” subunit. In yeast SWI/SNF, that motor is Snf2. (eustermann2024energydrivengenomeregulation pages 1-3)
1.2 What “SNF2” means in this report (identity verification)
This report concerns S. cerevisiae Snf2/Swi2 encoded by YOR290C (UniProt P22082), the SWI/SNF-family catalytic ATPase. The cited primary studies explicitly analyze yeast Snf2/Swi2 within SWI/SNF and experimentally manipulate domains that match the UniProt entry (HSA, QLQ, SnAC, AT-hook(s), bromodomain). (sen2011anewhighly pages 1-2, sen2011anewhighly pages 2-2, saha2023theathookis pages 1-2)
2) Molecular function: enzymatic activity, substrates, and mechanism
2.1 Enzymatic activity and substrate preference
Snf2 provides the SWI/SNF complex’s ATPase and DNA translocase activity. In mechanistic summaries and primary work, Snf2-containing SWI/SNF is described as a DNA-dependent ATPase that couples ATP hydrolysis to nucleosome mobilization/displacement; nucleosomal DNA is reported as the optimal substrate that stimulates ATP hydrolysis above naked DNA. (guoUnknownyear…thegenomewide pages 23-28)
At the SWI/SNF-family level, the activity output is nucleosome sliding and histone ejection, driven by the motor ATPase (Snf2 in yeast). (eustermann2024energydrivengenomeregulation pages 1-3)
2.2 Coupling ATP hydrolysis to nucleosome movement: the SnAC domain (primary evidence)
A highly conserved SnAC (“Snf2 ATP coupling”) domain lies between the ATPase and AT-hook region and is essential for productive remodeling. In yeast, deletion or mutation of SnAC severely impairs ATPase and nucleosome-mobilizing activities, but does not abolish complex integrity, efficient nucleosome binding, or recruitment by acidic transcription activators—supporting a specific “coupling” function between hydrolysis and mechanical work. (sen2011anewhighly pages 1-2)
Quantitatively, removal of SnAC does not substantially change apparent nucleosome binding affinity: WT nucleosome KD 2.85 ± 0.5 nM versus ΔSnAC KD 2.38 ± 0.7 nM, consistent with a catalytic/coupling defect rather than a binding defect. (sen2011anewhighly pages 4-5)
2.3 Auto-regulation by the AT-hook module: kinetic and mechanistic data (2023)
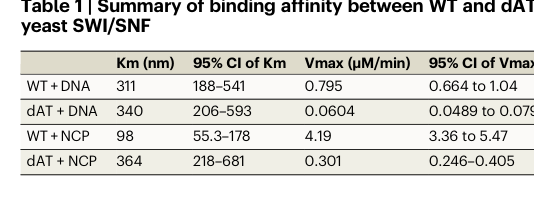
A 2023 Nature Communications study demonstrates that the C-terminal AT-hook module in yeast Snf2 is not merely a DNA tether, but an auto-regulatory element that strongly boosts ATPase catalytic velocity and remodeling rate. Deleting both AT-hooks (ΔAT) reduces ATP hydrolysis velocity ~13-fold (DNA-stimulated) and ~14-fold (nucleosome-stimulated), with Vmax decreasing from 0.795 to 0.0604 μM/min on DNA and from 4.19 to 0.301 μM/min on nucleosomes. (saha2023theathookis pages 1-2, saha2023theathookis pages 4-5)
The AT-hook deletion has modest effects on DNA KM (311 → 340 nM) but substantially worsens nucleosome KM (98 → 364 nM), indicating a key role in productive nucleosome engagement and catalysis under nucleosomal conditions. (saha2023theathookis pages 1-2, saha2023theathookis pages 4-5)
Binding effects are present but smaller than catalytic effects: free DNA KD 7.87 nM (WT) vs 12.8 nM (ΔAT), and nucleosome KD 7.67 nM (WT) vs 20.5 nM (ΔAT). (saha2023theathookis pages 1-2, saha2023theathookis pages 5-6)
The same work reports that ΔAT SWI/SNF mobilizes nucleosomes dramatically more slowly than WT (reported rates: 0.26 nM/s for WT vs 0.0019 nM/s for ΔAT), and that the remodeling reduction is tightly coupled to the ATPase reduction. (saha2023theathookis pages 2-4)
Visual evidence (kinetics and in vivo growth assays). Table 1 and Figure 4 from this study provide the kinetic parameterization and growth phenotypes supporting the above claims. (saha2023theathookis media 36b896c6, saha2023theathookis media 2a6902d6)
3) Domain architecture and experimentally supported roles
The following artifact compiles domain-level functional evidence directly supported by the retrieved primary literature and recent synthesis articles (with quantitative data where available).
| Domain/motif | Approx. location if available | Molecular role | Key experimental evidence / quantitative data | Key citations (with DOI URL) |
|---|---|---|---|---|
| HSA (helicase-SANT-associated) | N-terminal; exact residues not given in provided context | Binds actin-related proteins Arp7 and Arp9; helps build the SWI/SNF motor/ARP module | Experimental summaries report HSA is required for Arp7/Arp9 association with Snf2; ARP association contributes >90% of catalytic activity in one mechanistic summary of yeast SWI/SNF (guoUnknownyear…thegenomewide pages 23-28, sen2011anewhighly pages 2-2) | Sen et al., 2011-08, Nucleic Acids Res. DOI: https://doi.org/10.1093/nar/gkr622; mechanistic review/summary in provided context, year not stated (guoUnknownyear…thegenomewide pages 23-28) |
| QLQ | Between N-terminus and HSA | Conserved domain present in Snf2 architecture; specific function not established in the provided evidence | Domain is explicitly noted as conserved in yeast Snf2, but no direct functional assay or quantitative phenotype for QLQ is described in the retrieved context (sen2011anewhighly pages 2-2) | Sen et al., 2011-08, Nucleic Acids Res. DOI: https://doi.org/10.1093/nar/gkr622 |
| pHSA | Between HSA and ATPase region (exact residues not given) | Links ARP module to ATPase; conformational coupling on ARP binding | Mechanistic summary states pHSA connects HSA/ARP binding to the ATPase region and changes conformation upon ARP association, supporting motor regulation; no residue-level quantitative assay provided in retrieved context (guoUnknownyear…thegenomewide pages 23-28) | Mechanistic review/summary in provided context, year not stated (guoUnknownyear…thegenomewide pages 23-28) |
| ATPase / DNA translocase helicase motifs | Central motor; seven helicase motifs noted | Catalyzes ATP hydrolysis and DNA translocation on nucleosomes; powers nucleosome sliding, unwrapping, rewrapping, histone ejection/displacement | Snf2 is the catalytic subunit of SWI/SNF and a DNA-dependent ATPase whose optimal substrate is nucleosomal DNA rather than naked DNA; SWI/SNF activities summarized include nucleosome mobilization/displacement, sliding, and histone ejection. Recent review table identifies yeast Snf2 as the SWI/SNF motor ATPase with distal acidic-patch binding and bromodomain features (guoUnknownyear…thegenomewide pages 23-28, guoUnknownyear…thegenomewide pages 127-130, sen2011anewhighly pages 1-2, eustermann2024energydrivengenomeregulation pages 1-3) | Sen et al., 2011-08, Nucleic Acids Res. DOI: https://doi.org/10.1093/nar/gkr622; Eustermann et al., 2024-12, Nat. Rev. Mol. Cell Biol. DOI: https://doi.org/10.1038/s41580-023-00683-y |
| SnAC (Snf2 ATP coupling) | Between ATPase and AT-hook; deletion tested for aa1312–1444 | Couples ATP hydrolysis to remodeling; positively regulates ATPase output without major effect on ATP affinity or nucleosome binding | Deletion/mutation severely impairs ATPase and nucleosome mobilization, but not complex integrity or nucleosome binding. WT versus ΔSnAC nucleosome KD values are similar (2.85 ± 0.5 nM vs 2.38 ± 0.7 nM), supporting a catalytic-coupling rather than binding role. ΔSnAC expression profile correlates strongly with snf2Δ (r = 0.88), indicating essentiality for most Snf2-dependent gene activation (sen2011anewhighly pages 1-2, sen2011anewhighly pages 4-5, sen2011anewhighly pages 3-4) | Sen et al., 2011-08, Nucleic Acids Res. DOI: https://doi.org/10.1093/nar/gkr622 |
| AT-hook(s) | C-terminal region; ΔAT deletion spans ~aa1446–1530 in one assay set | Auto-regulatory domain that stimulates DNA- and nucleosome-stimulated ATPase activity, promotes nucleosome engagement, and supports in vivo SWI/SNF function | 2023 study shows ΔAT lowers ATPase Vmax ~13-fold on DNA (0.795 to 0.0604 μM/min) and ~14-fold on nucleosomes (4.19 to 0.301 μM/min), with modest DNA KM change (311 to 340 nM) but much worse nucleosome KM (98 to 364 nM). Binding effects are smaller but detectable: free DNA KD 7.87 vs 12.8 nM; nucleosome KD 7.67 vs 20.5 nM. Crosslinking places AT-hooks near H3 K15/K28, SnAC, and ATPase C-lobe. Growth assays indicate requirement for amino-acid biosynthesis/stress responses and ethanol metabolism, while some carbon-source switching phenotypes are less dependent on AT-hooks (saha2023theathookis pages 1-2, saha2023theathookis pages 4-5, saha2023theathookis pages 10-11, saha2023theathookis pages 2-4, saha2023theathookis pages 5-6, saha2023theathookis media 36b896c6, saha2023theathookis media 2a6902d6) | Saha et al., 2023-08, Nat. Commun. DOI: https://doi.org/10.1038/s41467-023-40386-8 |
| Bromodomain | C-terminal | Recognizes acetylated histone tails; contributes to chromatin targeting/retention and remodeler dynamics on acetylated chromatin | The bromodomain is reported to bind acetylated N-terminal histone tails. Mechanistic summary notes stimulation by tetra-acetylated H3 and acetyl-H3/H4 recognition. Recent work on stress-responsive genes showed competitive bromodomain interactions regulate Swi/Snf recruitment/release dynamics, including effects of Snf2 acetylation on binding to acetylated nucleosomes (guoUnknownyear…thegenomewide pages 23-28, sen2011anewhighly pages 2-2) | Sen et al., 2011-08, Nucleic Acids Res. DOI: https://doi.org/10.1093/nar/gkr622; Dutta et al., 2014-10, Genes Dev. DOI: https://doi.org/10.1101/gad.243584.114 |
| N-terminal activator-binding domain (ABD) | aa238–307 | Direct interaction with transcriptional activator TADs; likely contributes to recruitment but is dispensable in vivo because of redundancy with other SWI/SNF subunits | 2024 mapping study identified a ~70-aa Swi2 ABD (aa238–307) that binds Ino2 TAD1/TAD2 and also TADs from Gal4, Gcn4, Rap1, Aro80, and Swi5. In vivo, deletion variants lacking the ABD complemented swi2Δ growth phenotypes, indicating dispensability/redundancy. In reporter assays, Ino2 TAD1 activation dropped to 13.4% in swi2 null cells, whereas TAD2 retained 72.5%, supporting selective recruitment contributions (wendegatz2024transcriptionalactivationdomains pages 5-6, wendegatz2024transcriptionalactivationdomains pages 1-2, wendegatz2024transcriptionalactivationdomains pages 3-5, wendegatz2024transcriptionalactivationdomains pages 9-11) | Wendegatz et al., 2024-09, Current Genetics DOI: https://doi.org/10.1007/s00294-024-01300-x |
Table: This table summarizes the experimentally supported domain architecture of Saccharomyces cerevisiae Snf2/Swi2 (UniProt P22082; YOR290C), emphasizing what each motif contributes to SWI/SNF remodeling. It highlights where the evidence is strong and quantitative, especially for the SnAC and AT-hook regions.
Key points with highest-confidence experimental support include (i) SnAC as an ATP-to-remodeling coupling element that is dispensable for nucleosome binding but essential for remodeling output (sen2011anewhighly pages 1-2, sen2011anewhighly pages 4-5), and (ii) AT-hooks as strong catalytic accelerators (order-of-magnitude Vmax effect) and nucleosome-engagement elements (saha2023theathookis pages 1-2, saha2023theathookis pages 4-5).
4) Complex membership and interactions (SWI/SNF context)
4.1 Snf2 as the SWI/SNF motor ATPase and subunit context
A recent review table explicitly identifies yeast Snf2 as the SWI/SNF motor ATPase and associates SWI/SNF with “nucleosome sliding” and “histone ejection.” It lists canonical yeast SWI/SNF subunits that associate with the complex (including Arp7, Arp9, Snf5, Swi1, Swi3, Snf6, among others), placing Snf2 within the well-defined holo-complex. (eustermann2024energydrivengenomeregulation pages 1-3)
A primary-study context further notes SWI/SNF as a ~12-subunit complex and reports that a minimal complex (Snf2 plus Arp7 and Arp9) can exhibit in vitro activity, though with limitations relative to the full complex (e.g., reduced ability to displace H2A–H2B dimers). (sen2011anewhighly pages 1-2)
4.2 Interactions with transcriptional activators (recruitment interfaces)
A 2024 Current Genetics study provides direct biochemical evidence that transcriptional activation domains can bind to the yeast SWI/SNF ATPase subunit (Swi2/Snf2) and maps a ~70 aa N-terminal “activator-binding domain” (ABD) in Swi2 spanning aa 238–307. This ABD binds both activation domains (TAD1/TAD2) of the yeast transcription factor Ino2 and also binds TADs from several unrelated activators (Gal4, Gcn4, Rap1, Aro80, Swi5) in vitro, supporting a model in which activators can directly contact Snf2 to help recruit/engage SWI/SNF at promoters. (wendegatz2024transcriptionalactivationdomains pages 5-6, wendegatz2024transcriptionalactivationdomains pages 3-5)
Functionally, reporter assays indicate Ino2 TAD1 activation is strongly reduced in a swi2 null background (to 13.4% of WT), whereas TAD2 retains more activity (72.5%), implying differential reliance on Swi2-mediated recruitment mechanisms. Importantly, deletion variants lacking the mapped Swi2 ABD can complement growth phenotypes, supporting redundancy among SWI/SNF subunits that contain activator-binding interfaces. (wendegatz2024transcriptionalactivationdomains pages 5-6)
5) Biological processes, pathways, and cellular localization
5.1 Localization (functional inference from direct chromatin engagement)
Across primary and review sources, Snf2 is functionally characterized through nucleosome binding, ATPase assays using DNA/nucleosomes, chromatin remodeling reactions, and genome-wide chromatin occupancy measurements (ChIP-based), all of which place its function in the nucleus on chromatin. For example, Snf2 occupancy at regulatory regions changes upon promoter induction in vivo (ChIP-seq), consistent with chromatin-associated nuclear localization. (morse2024swisnfchromatinremodeling pages 12-15)
5.2 Pathways/processes: metabolic regulation and stress-responsive transcription programs (AT-hook phenotypes)
In vivo growth assays in yeast show that AT-hooks are “needed in vivo for active yeast SWI/SNF,” with phenotypes implicating SWI/SNF-dependent activation programs including amino-acid biosynthesis (e.g., rescue under amino-acid starvation conditions) and ethanol metabolism-related programs. (saha2023theathookis pages 5-6, saha2023theathookis pages 4-5, saha2023theathookis media 36b896c6)
Mechanistically, the interpretation is that some transcriptional programs require high-efficiency nucleosome mobilization (and thus strong ATPase output), whereas others tolerate attenuated remodeling. (saha2023theathookis pages 10-11)
5.3 Pathways/processes: phospholipid biosynthesis gene activation (Ino2 recruitment)
The Ino2 activator is required for expression of phospholipid biosynthetic genes in yeast, and its activation domains bind multiple SWI/SNF subunits, including Swi2/Snf2. This provides a direct path from a defined transcription factor to Snf2-containing remodeling activity at metabolic gene promoters. (wendegatz2024transcriptionalactivationdomains pages 1-2, wendegatz2024transcriptionalactivationdomains pages 5-6)
5.4 A newer functional axis (2024): transcriptional interference and gene repression
A 2024 Molecular Cell study highlights an expanded view of Snf2/SWI-SNF function in vivo: SWI/SNF not only promotes transcription initiation through promoter remodeling, but can also remodel nucleosomes downstream of active transcription start sites (TSSs) to repress proximal promoters (TSSPROX) associated with genes that also possess a distal active TSS (TSSDIST), thereby controlling transcriptional interference. (morse2024swisnfchromatinremodeling pages 66-70)
This work provides quantitative evidence of Snf2 redistribution in response to promoter induction: Snf2 occupancy at 5′ regulatory regions increased on average ~1.4-fold after distal promoter induction (paired t test p = 0.0423). (morse2024swisnfchromatinremodeling pages 12-15)
6) Recent developments and latest research (prioritizing 2023–2024)
6.1 2023: AT-hook as an evolutionarily conserved auto-regulatory domain
The 2023 AT-hook study reframes AT-hooks from passive DNA anchors to strong catalytic regulators that accelerate ATP hydrolysis and nucleosome movement (~13–14× Vmax effects) and modulate nucleosome engagement (KM shift on nucleosomes). This “attenuation” model is useful because it enables partial-function perturbations that differ from full deletion or ATPase-dead mutants, potentially revealing remodeling-efficiency thresholds for specific gene programs. (saha2023theathookis pages 1-2, saha2023theathookis pages 10-11)
6.2 2024: direct activator-binding interfaces within Swi2/Snf2
The 2024 Swi2 ABD mapping provides a contemporary biochemical framework for how intrinsically disordered transcription activation domains (TADs) can directly engage ATPase subunits of remodelers, and it experimentally supports the concept of redundancy (ABD dispensability) within SWI/SNF recruitment logic. (wendegatz2024transcriptionalactivationdomains pages 5-6, wendegatz2024transcriptionalactivationdomains pages 1-2)
6.3 2024: SWI/SNF-mediated repression via gene-body remodeling in transcriptional interference
The 2024 transcriptional interference study provides a mechanistically specific repression function for SWI/SNF in yeast, supported by genome-wide transcriptomics (including TSS mapping) and chromatin profiling (MNase-seq/ChIP-seq), expanding functional annotation beyond “coactivator” descriptions. (morse2024swisnfchromatinremodeling pages 66-70, morse2024swisnfchromatinremodeling pages 8-12)
6.4 2024: synthesis of remodeler mechanics and outputs
A 2024 Nature Reviews Molecular Cell Biology review synthesizes ATP-dependent remodeler families and explicitly associates yeast Snf2/SWI-SNF with sliding and histone ejection outputs, providing authoritative terminology and a comparative framework for mechanistic annotation. (eustermann2024energydrivengenomeregulation pages 1-3)
7) Current applications and real-world implementations
7.1 Yeast Snf2 as a model system for “tunable” chromatin remodeling perturbations
The AT-hook work demonstrates a practical experimental strategy: rather than deleting SNF2 or using ATPase-dead mutants (often lethal or broadly pleiotropic), researchers can attenuate remodeling through specific regulatory elements (AT-hooks) to dissect remodeling-efficiency requirements for distinct gene programs. This is a real-world implementation in functional genomics and mechanistic epigenetics using yeast as a tractable model. (saha2023theathookis pages 10-11, saha2023theathookis media 36b896c6)
7.2 Translational context (non-yeast): pharmacological targeting of SWI/SNF
While not yeast-specific, 2024 review literature emphasizes pharmacological targeting of SWI/SNF complexes in cancer via catalytic ATPase inhibitors, bromodomain ligands, and PROTAC-based degradation strategies. Yeast mechanistic insights (e.g., bromodomain and AT-hook regulatory logic in a homologous ATPase family) inform hypotheses about conserved regulatory surfaces, even though subunit composition differs between yeast and mammals. (eustermann2024energydrivengenomeregulation pages 1-3)
8) Expert opinions and analysis (authoritative interpretations)
-
The 2024 Nature Reviews Molecular Cell Biology synthesis frames chromatin remodelers as “energy-driven genome regulation” machines and explicitly categorizes SWI/SNF output modes (sliding and ejection) with a motor ATPase at the center (Snf2 in yeast), reflecting the field’s current consensus model. (eustermann2024energydrivengenomeregulation pages 1-3)
-
The 2023 AT-hook study’s analysis argues that understanding partial attenuation (rather than complete inactivation) is critical for separating remodeling “efficiency” requirements across biological programs, an approach aligned with modern quantitative mechanistic biology. (saha2023theathookis pages 10-11)
9) Key statistics and quantitative data (recent studies)
-
ATPase kinetics (2023): Deleting Snf2 AT-hooks reduces Vmax ~13× on DNA and ~14× on nucleosomes (0.795→0.0604 μM/min on DNA; 4.19→0.301 μM/min on nucleosomes). Nucleosome KM worsens ~3.7× (98→364 nM), while DNA KM changes minimally (311→340 nM). (saha2023theathookis pages 4-5, saha2023theathookis media 36b896c6)
-
Binding affinities (2023): ΔAT weakens nucleosome binding more than DNA binding (KD nucleosome 7.67→20.5 nM; KD DNA 7.87→12.8 nM). (saha2023theathookis pages 5-6)
-
Remodeling rate (2023): Reported nucleosome mobilization rates differ dramatically between WT and ΔAT (0.26 nM/s vs 0.0019 nM/s). (saha2023theathookis pages 2-4)
-
Gene regulation (2011): SnAC deletion yields gene expression changes strongly correlated with snf2Δ (correlation r = 0.88), consistent with SnAC being required for most Snf2-dependent gene activation. (sen2011anewhighly pages 3-4)
-
Transcriptional interference / repression (2024): Snf2 occupancy at 5′ regulatory regions increases ~1.4-fold upon distal promoter induction (p = 0.0423). SER3 mRNA is upregulated ~40-fold in the snf2-Q928K allele in the context of interference regulation at the SER3 locus. (morse2024swisnfchromatinremodeling pages 12-15, morse2024swisnfchromatinremodeling pages 8-12)
10) Practical functional annotation summary (actionable)
Primary molecular function: ATP-dependent chromatin remodeling as the SWI/SNF motor ATPase, translocating on nucleosomal DNA to drive nucleosome sliding and histone ejection/destabilization. (eustermann2024energydrivengenomeregulation pages 1-3, guoUnknownyear…thegenomewide pages 23-28)
Key regulatory/coupling elements:
* SnAC: essential ATP-to-remodeling coupling; large functional impact without major nucleosome binding change. (sen2011anewhighly pages 1-2, sen2011anewhighly pages 4-5)
* AT-hooks: strong catalytic accelerators and nucleosome engagement elements (order-of-magnitude Vmax effects). (saha2023theathookis pages 1-2, saha2023theathookis pages 4-5)
Localization: Nuclear and chromatin-associated, supported by nucleosome engagement assays and in vivo chromatin occupancy profiling. (morse2024swisnfchromatinremodeling pages 12-15)
Pathways/processes:
* Activator-driven metabolic transcription programs (e.g., amino-acid biosynthesis/stress; phospholipid biosynthesis via Ino2). (saha2023theathookis pages 4-5, wendegatz2024transcriptionalactivationdomains pages 1-2)
* Chromatin-mediated repression via transcriptional interference (gene-body remodeling). (morse2024swisnfchromatinremodeling pages 66-70, morse2024swisnfchromatinremodeling pages 8-12)
11) Source list (URLs and publication dates)
- Saha D. et al. (2023-08). “The AT-hook is an evolutionarily conserved auto-regulatory domain of SWI/SNF required for cell lineage priming.” Nature Communications. https://doi.org/10.1038/s41467-023-40386-8 (saha2023theathookis pages 1-2)
- Morse K. et al. (2024-08). “Swi/Snf chromatin remodeling regulates transcriptional interference and gene repression.” Molecular Cell. https://doi.org/10.1016/j.molcel.2024.06.029 (morse2024swisnfchromatinremodeling pages 12-15)
- Wendegatz E.-C. et al. (2024-09). “Transcriptional activation domains interact with ATPase subunits of yeast chromatin remodelling complexes SWI/SNF, RSC and INO80.” Current Genetics. https://doi.org/10.1007/s00294-024-01300-x (wendegatz2024transcriptionalactivationdomains pages 5-6)
- Eustermann S. et al. (2024; published in Nat Rev Mol Cell Biol with 2024-12 metadata in retrieved record). “Energy-driven genome regulation by ATP-dependent chromatin remodellers.” https://doi.org/10.1038/s41580-023-00683-y (eustermann2024energydrivengenomeregulation pages 1-3)
- Sen P. et al. (2011-08). “A new, highly conserved domain in Swi2/Snf2 is required for SWI/SNF remodeling.” Nucleic Acids Research. https://doi.org/10.1093/nar/gkr622 (sen2011anewhighly pages 1-2)
References
-
(sen2011anewhighly pages 1-2): Payel Sen, Sujana Ghosh, B. Franklin Pugh, and Blaine Bartholomew. A new, highly conserved domain in swi2/snf2 is required for swi/snf remodeling. Nucleic Acids Research, 39:9155-9166, Aug 2011. URL: https://doi.org/10.1093/nar/gkr622, doi:10.1093/nar/gkr622. This article has 70 citations and is from a highest quality peer-reviewed journal.
-
(eustermann2024energydrivengenomeregulation pages 1-3): Sebastian Eustermann, Avinash B. Patel, Karl-Peter Hopfner, Yuan He, and Philipp Korber. Energy-driven genome regulation by atp-dependent chromatin remodellers. Nature reviews. Molecular cell biology, 25:309-332, Dec 2024. URL: https://doi.org/10.1038/s41580-023-00683-y, doi:10.1038/s41580-023-00683-y. This article has 113 citations.
-
(saha2023theathookis pages 1-2): Dhurjhoti Saha, Solomon Hailu, Arjan Hada, Junwoo Lee, Jie Luo, Jeff A. Ranish, Yuan-chi Lin, Kyle Feola, Jim Persinger, Abhinav Jain, Bin Liu, Yue Lu, Payel Sen, and Blaine Bartholomew. The at-hook is an evolutionarily conserved auto-regulatory domain of swi/snf required for cell lineage priming. Nature Communications, Aug 2023. URL: https://doi.org/10.1038/s41467-023-40386-8, doi:10.1038/s41467-023-40386-8. This article has 9 citations and is from a highest quality peer-reviewed journal.
-
(morse2024swisnfchromatinremodeling pages 8-12): Kaitlin Morse, Alena L. Bishop, Sarah Swerdlow, Jessica M. Leslie, and Elçin Ünal. Swi/snf chromatin remodeling regulates transcriptional interference and gene repression. Aug 2024. URL: https://doi.org/10.1016/j.molcel.2024.06.029, doi:10.1016/j.molcel.2024.06.029. This article has 21 citations and is from a highest quality peer-reviewed journal.
-
(sen2011anewhighly pages 2-2): Payel Sen, Sujana Ghosh, B. Franklin Pugh, and Blaine Bartholomew. A new, highly conserved domain in swi2/snf2 is required for swi/snf remodeling. Nucleic Acids Research, 39:9155-9166, Aug 2011. URL: https://doi.org/10.1093/nar/gkr622, doi:10.1093/nar/gkr622. This article has 70 citations and is from a highest quality peer-reviewed journal.
-
(guoUnknownyear…thegenomewide pages 23-28): X Guo. … the genome-wide interplay between swi/snf and tup1-cyc8 complexes in transcriptional regulation and chromatin remodeling in saccharomyces cerevisiae. Unknown journal, Unknown year.
-
(sen2011anewhighly pages 4-5): Payel Sen, Sujana Ghosh, B. Franklin Pugh, and Blaine Bartholomew. A new, highly conserved domain in swi2/snf2 is required for swi/snf remodeling. Nucleic Acids Research, 39:9155-9166, Aug 2011. URL: https://doi.org/10.1093/nar/gkr622, doi:10.1093/nar/gkr622. This article has 70 citations and is from a highest quality peer-reviewed journal.
-
(saha2023theathookis pages 4-5): Dhurjhoti Saha, Solomon Hailu, Arjan Hada, Junwoo Lee, Jie Luo, Jeff A. Ranish, Yuan-chi Lin, Kyle Feola, Jim Persinger, Abhinav Jain, Bin Liu, Yue Lu, Payel Sen, and Blaine Bartholomew. The at-hook is an evolutionarily conserved auto-regulatory domain of swi/snf required for cell lineage priming. Nature Communications, Aug 2023. URL: https://doi.org/10.1038/s41467-023-40386-8, doi:10.1038/s41467-023-40386-8. This article has 9 citations and is from a highest quality peer-reviewed journal.
-
(saha2023theathookis pages 5-6): Dhurjhoti Saha, Solomon Hailu, Arjan Hada, Junwoo Lee, Jie Luo, Jeff A. Ranish, Yuan-chi Lin, Kyle Feola, Jim Persinger, Abhinav Jain, Bin Liu, Yue Lu, Payel Sen, and Blaine Bartholomew. The at-hook is an evolutionarily conserved auto-regulatory domain of swi/snf required for cell lineage priming. Nature Communications, Aug 2023. URL: https://doi.org/10.1038/s41467-023-40386-8, doi:10.1038/s41467-023-40386-8. This article has 9 citations and is from a highest quality peer-reviewed journal.
-
(saha2023theathookis pages 2-4): Dhurjhoti Saha, Solomon Hailu, Arjan Hada, Junwoo Lee, Jie Luo, Jeff A. Ranish, Yuan-chi Lin, Kyle Feola, Jim Persinger, Abhinav Jain, Bin Liu, Yue Lu, Payel Sen, and Blaine Bartholomew. The at-hook is an evolutionarily conserved auto-regulatory domain of swi/snf required for cell lineage priming. Nature Communications, Aug 2023. URL: https://doi.org/10.1038/s41467-023-40386-8, doi:10.1038/s41467-023-40386-8. This article has 9 citations and is from a highest quality peer-reviewed journal.
-
(saha2023theathookis media 36b896c6): Dhurjhoti Saha, Solomon Hailu, Arjan Hada, Junwoo Lee, Jie Luo, Jeff A. Ranish, Yuan-chi Lin, Kyle Feola, Jim Persinger, Abhinav Jain, Bin Liu, Yue Lu, Payel Sen, and Blaine Bartholomew. The at-hook is an evolutionarily conserved auto-regulatory domain of swi/snf required for cell lineage priming. Nature Communications, Aug 2023. URL: https://doi.org/10.1038/s41467-023-40386-8, doi:10.1038/s41467-023-40386-8. This article has 9 citations and is from a highest quality peer-reviewed journal.
-
(saha2023theathookis media 2a6902d6): Dhurjhoti Saha, Solomon Hailu, Arjan Hada, Junwoo Lee, Jie Luo, Jeff A. Ranish, Yuan-chi Lin, Kyle Feola, Jim Persinger, Abhinav Jain, Bin Liu, Yue Lu, Payel Sen, and Blaine Bartholomew. The at-hook is an evolutionarily conserved auto-regulatory domain of swi/snf required for cell lineage priming. Nature Communications, Aug 2023. URL: https://doi.org/10.1038/s41467-023-40386-8, doi:10.1038/s41467-023-40386-8. This article has 9 citations and is from a highest quality peer-reviewed journal.
-
(guoUnknownyear…thegenomewide pages 127-130): X Guo. … the genome-wide interplay between swi/snf and tup1-cyc8 complexes in transcriptional regulation and chromatin remodeling in saccharomyces cerevisiae. Unknown journal, Unknown year.
-
(sen2011anewhighly pages 3-4): Payel Sen, Sujana Ghosh, B. Franklin Pugh, and Blaine Bartholomew. A new, highly conserved domain in swi2/snf2 is required for swi/snf remodeling. Nucleic Acids Research, 39:9155-9166, Aug 2011. URL: https://doi.org/10.1093/nar/gkr622, doi:10.1093/nar/gkr622. This article has 70 citations and is from a highest quality peer-reviewed journal.
-
(saha2023theathookis pages 10-11): Dhurjhoti Saha, Solomon Hailu, Arjan Hada, Junwoo Lee, Jie Luo, Jeff A. Ranish, Yuan-chi Lin, Kyle Feola, Jim Persinger, Abhinav Jain, Bin Liu, Yue Lu, Payel Sen, and Blaine Bartholomew. The at-hook is an evolutionarily conserved auto-regulatory domain of swi/snf required for cell lineage priming. Nature Communications, Aug 2023. URL: https://doi.org/10.1038/s41467-023-40386-8, doi:10.1038/s41467-023-40386-8. This article has 9 citations and is from a highest quality peer-reviewed journal.
-
(wendegatz2024transcriptionalactivationdomains pages 5-6): Eva-Carina Wendegatz, Maike Engelhardt, and Hans-Joachim Schüller. Transcriptional activation domains interact with atpase subunits of yeast chromatin remodelling complexes swi/snf, rsc and ino80. Current Genetics, Sep 2024. URL: https://doi.org/10.1007/s00294-024-01300-x, doi:10.1007/s00294-024-01300-x. This article has 3 citations and is from a peer-reviewed journal.
-
(wendegatz2024transcriptionalactivationdomains pages 1-2): Eva-Carina Wendegatz, Maike Engelhardt, and Hans-Joachim Schüller. Transcriptional activation domains interact with atpase subunits of yeast chromatin remodelling complexes swi/snf, rsc and ino80. Current Genetics, Sep 2024. URL: https://doi.org/10.1007/s00294-024-01300-x, doi:10.1007/s00294-024-01300-x. This article has 3 citations and is from a peer-reviewed journal.
-
(wendegatz2024transcriptionalactivationdomains pages 3-5): Eva-Carina Wendegatz, Maike Engelhardt, and Hans-Joachim Schüller. Transcriptional activation domains interact with atpase subunits of yeast chromatin remodelling complexes swi/snf, rsc and ino80. Current Genetics, Sep 2024. URL: https://doi.org/10.1007/s00294-024-01300-x, doi:10.1007/s00294-024-01300-x. This article has 3 citations and is from a peer-reviewed journal.
-
(wendegatz2024transcriptionalactivationdomains pages 9-11): Eva-Carina Wendegatz, Maike Engelhardt, and Hans-Joachim Schüller. Transcriptional activation domains interact with atpase subunits of yeast chromatin remodelling complexes swi/snf, rsc and ino80. Current Genetics, Sep 2024. URL: https://doi.org/10.1007/s00294-024-01300-x, doi:10.1007/s00294-024-01300-x. This article has 3 citations and is from a peer-reviewed journal.
-
(morse2024swisnfchromatinremodeling pages 12-15): Kaitlin Morse, Alena L. Bishop, Sarah Swerdlow, Jessica M. Leslie, and Elçin Ünal. Swi/snf chromatin remodeling regulates transcriptional interference and gene repression. Aug 2024. URL: https://doi.org/10.1016/j.molcel.2024.06.029, doi:10.1016/j.molcel.2024.06.029. This article has 21 citations and is from a highest quality peer-reviewed journal.
-
(morse2024swisnfchromatinremodeling pages 66-70): Kaitlin Morse, Alena L. Bishop, Sarah Swerdlow, Jessica M. Leslie, and Elçin Ünal. Swi/snf chromatin remodeling regulates transcriptional interference and gene repression. Aug 2024. URL: https://doi.org/10.1016/j.molcel.2024.06.029, doi:10.1016/j.molcel.2024.06.029. This article has 21 citations and is from a highest quality peer-reviewed journal.
Artifacts
Citations
- eustermann2024energydrivengenomeregulation pages 1-3
- sen2011anewhighly pages 1-2
- sen2011anewhighly pages 4-5
- saha2023theathookis pages 2-4
- sen2011anewhighly pages 2-2
- wendegatz2024transcriptionalactivationdomains pages 5-6
- morse2024swisnfchromatinremodeling pages 12-15
- saha2023theathookis pages 10-11
- morse2024swisnfchromatinremodeling pages 66-70
- saha2023theathookis pages 5-6
- sen2011anewhighly pages 3-4
- saha2023theathookis pages 1-2
- morse2024swisnfchromatinremodeling pages 8-12
- saha2023theathookis pages 4-5
- wendegatz2024transcriptionalactivationdomains pages 1-2
- wendegatz2024transcriptionalactivationdomains pages 3-5
- wendegatz2024transcriptionalactivationdomains pages 9-11
- https://doi.org/10.1093/nar/gkr622;
- https://doi.org/10.1093/nar/gkr622
- https://doi.org/10.1038/s41580-023-00683-y
- https://doi.org/10.1038/s41467-023-40386-8
- https://doi.org/10.1101/gad.243584.114
- https://doi.org/10.1007/s00294-024-01300-x
- https://doi.org/10.1016/j.molcel.2024.06.029
- https://doi.org/10.1093/nar/gkr622,
- https://doi.org/10.1038/s41580-023-00683-y,
- https://doi.org/10.1038/s41467-023-40386-8,
- https://doi.org/10.1016/j.molcel.2024.06.029,
- https://doi.org/10.1007/s00294-024-01300-x,
Perplexity
(SWI2-deep-research-perplexity.md)
Perplexity
(SWI2-deep-research-perplexity.md)Comprehensive Functional Annotation of SNF2/SWI2: An ATP-Dependent Chromatin Remodeling Enzyme in Saccharomyces cerevisiae
The SNF2 gene, also known as SWI2, encodes a transcription regulatory protein that serves as the catalytic subunit of the SWI/SNF chromatin remodeling complex in the baker's yeast Saccharomyces cerevisiae[1][2]. This approximately 170 kDa protein functions as an ATP-dependent helicase-like enzyme that couples ATP hydrolysis to mechanical remodeling of chromatin structure, thereby regulating access to genomic DNA for transcription, replication, and repair processes[2][3]. The SNF2/SWI2 gene was originally identified through genetic screens examining mutations that affected sucrose non-fermentation (SNF) and mating type switching (SWI), making it one of the founding members of the ATP-dependent chromatin remodeling enzyme family[1][5]. Research over the past three decades has established that SNF2 and its mammalian homologs are central regulators of gene expression and genome stability, with mutations in human SNF2 family members implicated in cancer and other genetic disorders[2][5]. The present report provides a comprehensive examination of the molecular mechanisms, cellular functions, and biological roles of this critical chromatin remodeling enzyme.
Structural Organization and Domain Architecture of the SNF2 Protein
The SNF2 protein is organized into several distinct functional domains that collectively enable its diverse chromatin remodeling activities. At the core of SNF2 lies the Snf2 helicase-like ATPase domain, which consists of two tandem RecA-like folds representing conserved structural elements found across diverse ATP-dependent enzymes[3][25]. This domain contains seven conserved helicase-related sequence motifs (motifs I through VI) that classify SNF2 as a member of the Superfamily 2 (SF2) of helicase-like proteins[3][25][30]. Although SNF2 family proteins contain these characteristic helicase motifs, they do not exhibit the classical DNA unwinding activity that defines true helicases; instead, they utilize ATP energy to translocate along duplex DNA without requiring strand separation[2][6][30]. The structural organization of the Snf2 domain includes an N-terminal subdomain I (comprising motifs I, Ia, II, and III) that is primarily responsible for ATP binding and hydrolysis, and a C-terminal subdomain II (comprising motifs IV to VI) that plays a critical role in energy transduction and converting the chemical energy of ATP hydrolysis into mechanical work[2].
A particularly significant structural element of SNF2 is the bromodomain, which functions as a reader of histone acetylation marks[8][11]. The bromodomain of SNF2 specifically recognizes acetylated lysine residues on histone H3 and H4 tails, with the protein showing relatively high binding affinity for various acetylated histone peptides compared to other bromodomain-containing proteins[8]. This domain is critical for recruiting the SWI/SNF complex to acetylated chromatin regions, as deletion of the Snf2 bromodomain severely impairs binding to hyperacetylated nucleosomes and reduces the remodeling activity of the complex on acetylated substrates[8][11]. The bromodomain-histone acetylation interaction represents an important regulatory mechanism by which the SWI/SNF complex is preferentially targeted to active or poised chromatin regions. Beyond these two major domains, SNF2 also contains an AT-hook DNA-binding motif, which facilitates DNA recognition and binding, enhancing the protein's interaction with nucleosomal and non-nucleosomal DNA substrates[2].
The DNA-stimulated ATPase activity of SNF2 demonstrates approximately 30-fold stimulation in the presence of double-stranded DNA, indicating that the protein has evolved to sense and respond to DNA substrates[27]. When bound to nucleosomal DNA, SNF2 shows even greater catalytic efficiency, suggesting that the protein has specific recognition elements for nucleosomal substrates. Critically, mutations within the Walker A box (motif I) of SNF2 completely eliminate ATP hydrolysis and render the protein catalytically inactive, demonstrating the absolute requirement for ATP binding and hydrolysis for all known functions of SNF2[2]. The conserved motif V appears particularly important for coupling ATP hydrolysis to actual chromatin remodeling activity; amino acid substitutions within this motif reduce nucleosome mobilization and restrict enzyme accessibility while maintaining ATP hydrolysis capacity, indicating that motif V specifically transduces chemical energy into mechanical remodeling work[2].
The SWI/SNF Complex: Architecture, Composition, and Modularity
The SNF2 protein does not function in isolation but rather serves as the catalytic engine of the approximately 1 megadalton (MDa) SWI/SNF chromatin remodeling complex, one of the first ATP-dependent chromatin remodeling enzymes to be identified and characterized[2][4]. The complete yeast SWI/SNF complex comprises at least twelve subunits that work together to facilitate diverse chromatin remodeling activities[13]. Recent proteomic and biochemical studies have revealed that the SWI/SNF complex exhibits internal modularity, with subunits organizing into at least four functionally and structurally distinct modules: an Arp module (containing Arp7, Arp9, and Rtt102), a catalytic module (containing Snf2 and Snf11), a regulatory module (subdivided into a Snf6-Snf12-Swi3 sub-module and a Snf5-Swp82-Taf14 sub-module), and a Swi1 module[13]. This internal architecture reveals that while some subunits are crucial for maintaining complex integrity, others function as peripheral components that can be dispensed with under certain circumstances.
The modularity of the SWI/SNF complex suggests functional specialization within the larger assembly, where different combinations of subunits may be utilized for distinct chromatin remodeling tasks[13]. Importantly, Snf2 occupancy and recruitment to promoters can occur independently of complete complex integrity and even in the absence of high levels of histone acetylation for certain gene targets, indicating that multiple regulatory pathways control SNF2 function[13]. The Snf5 subunit, which is part of the regulatory module, plays a crucial role in stabilizing the complex and regulating Snf2 occupancy at specific genomic loci, particularly at transposable element loci where high histone acetylation is present[13]. In contrast, at protein-coding genes with lower histone acetylation levels, Snf5 and other regulatory module subunits show greater importance for Snf2 recruitment, suggesting that the composition of the SWI/SNF complex or the specific auxiliary subunits present can influence which genes are targeted for remodeling[13].
Biochemical Mechanism of ATP-Dependent Chromatin Remodeling
The fundamental biochemical role of SNF2 is to utilize the energy stored in adenosine triphosphate (ATP) hydrolysis and convert it into mechanical work that alters nucleosome structure and positioning[3][25]. The mechanism begins when SNF2 binds to nucleosomal DNA, typically at superhelical location (SHL) +2 or adjacent positions, and subsequently hydrolyzes ATP in a DNA-stimulated manner[56]. Recent cryo-electron microscopy (cryo-EM) structures of SNF2 homologs in complex with nucleosomes have provided unprecedented structural insights into this mechanism, revealing that SNF2-mediated chromatin remodeling involves asymmetric deformation of the histone octamer core[56]. Specifically, the structures captured show increased disorder in histone residues near the nucleosome dyad axis and within the H2A-H2B acidic patch when SNF2 is bound in the presence of ADP-BeF₃ (a non-hydrolyzable ATP analog)[56].
The ATP-dependent translocation mechanism involves ATP-driven conformational changes between the two RecA-like domains of the Snf2 helicase-like region, which result in closure of the interdomain cleft and advancement of DNA by one base pair at a time[30]. During this translocation, the conserved arginine residue within motif VI maintains contact with the γ-phosphate of ATP, stabilizing the closed interdomain configuration[30]. The translocation of DNA along the octamer surface, combined with the conformational flexibility induced in the histone core, eventually results in propagation of DNA around the nucleosome, causing the nucleosome to slide along the DNA in a directional manner[56]. SNF2 appears to amplify intrinsic nucleosome dynamics during the sliding reaction, using the histone octamer as a deformable medium for allosteric control rather than treating it simply as a rigid barrier to be overcome[56].
A critical distinction between SNF2-type remodelers and other remodeler subfamilies like ISWI and Mi-2 is that SNF2 shows equal stimulation of ATPase activity by both free and nucleosomal DNA[27]. This contrasts with ISWI and Mi-2 subfamilies, which show strong preference for nucleosomal DNA substrates[27]. The ability of SNF2 to respond equivalently to both DNA types, combined with evidence that SNF2 preferentially interacts with nucleosomal DNA and generates superhelical torsion on nucleosomal templates more efficiently than on naked DNA, suggests that SNF2 may have evolved specialized recognition mechanisms for nucleosomal substrates involving both the histone core and the DNA component[27].
Chromatin Remodeling Activities: Nucleosome Sliding, Eviction, and Histone Exchange
The SWI/SNF complex catalyzes multiple distinct chromatin remodeling reactions, each with different biological consequences[3][7][25]. The most well-characterized activity is nucleosome sliding, in which SNF2 ATP hydrolysis drives the translocation of nucleosomes along DNA, moving positioned nucleosomes toward DNA ends or neighboring nucleosomes in a directional manner[2][3][7]. This sliding activity is quantifiable through restriction enzyme accessibility assays, in which ATP-dependent movement of nucleosomes exposes previously occluded restriction sites, and the accessibility increases proportionally with the kinetics of nucleosome mobilization[2]. A single deletion within SNF2 motif V (ΔSTRAGGLG) dramatically impairs nucleosome mobilization while maintaining ATP hydrolysis capacity, demonstrating that the energy transduction function of motif V is specifically required for this mechanical remodeling activity[2].
Nucleosome eviction or disassembly represents another critical SWI/SNF function in which the ATP-dependent remodeling leads to complete removal of nucleosomes from DNA[7][22]. This eviction activity is particularly important during gene regulation, as it ensures that promoter and enhancer DNA sequences become accessible to transcription factors, histone-modifying enzymes, and RNA polymerase II[7][22]. The conditions under which SNF2 catalyzes sliding versus eviction remain incompletely understood, but likely depend on complex composition, the presence of histone chaperones, and the specific nature of the nucleosomal substrate.
In addition to sliding and eviction, SNF2 participates in histone variant incorporation and exchange, though this function may be particularly prominent for specific SNF2 family members in the SWR1-like subfamily rather than SNF2 itself[3]. However, SNF2 does play an important regulatory role in managing histone variant distribution across the genome. The INO80 Snf2 family member can replace nucleosomal H2A.Z-H2B dimers with canonical H2A-H2B dimers, a specificity opposite to that of the SWR1 complex, and this bidirectional regulation appears essential for maintaining proper genomic distribution of H2A.Z and genome stability[3]. SWI/SNF has also been shown to restrict the distribution of Cse4-containing (centromeric histone variant) nucleosomes to centromeric regions, preventing ectopic spreading that would compromise chromosome segregation[25].
Gene Regulation and Transcriptional Activation
The primary biological role of SNF2 in vivo is to facilitate transcriptional activation of genes by promoting accessibility of DNA to transcription regulatory proteins[1][5]. The SWI/SNF complex is recruited to promoters of specific target genes through direct physical interactions between SNF2 and other SWI/SNF subunits (particularly Snf5, Swi1, and Snf2 itself) with acidic activation domains of transcription factors[58]. This direct targeting mechanism requires that the transcription factor bind to DNA adjacent to the nucleosome or promoter region, as recruitment of SWI/SNF requires more than 53 base pairs of DNA adjacent to the transcription factor binding site[55]. The recruitment process occurs independently of core promoter recognition machinery, as SWI/SNF can be recruited to promoters in the absence of TFIID and other preinitiation complex components[55].
Early genetic studies identified SNF2 as required for expression of the SUC2 gene, which encodes invertase involved in sucrose fermentation and represents a paradigmatic example of glucose-repressed gene regulation[1][5][43]. In wild-type yeast, the SUC2 gene is strongly repressed in the presence of glucose but rapidly derepressed when glucose is depleted, and this glucose-responsive regulation absolutely requires functional SNF2[43]. The mechanism involves an acidic transcription factor that binds to the SUC2 promoter and recruits SWI/SNF through direct protein-protein interactions, which then remodels chromatin at the promoter to allow transcription factor and RNA polymerase II binding[5][8].
SNF2 is similarly required for transcription of many other glucose-repressed genes including members of the GAL regulon and additional carbohydrate utilization genes[5]. Beyond glucose metabolism, SNF2 has been implicated in regulating genes involved in amino acid biosynthesis through cooperation with other chromatin remodeling complexes[5]. A particularly novel recent discovery reveals that SNF2 plays an unexpected role in transcriptional interference, a regulatory mechanism in which transcription from one promoter can interfere with transcription from a downstream promoter on the same DNA molecule[19]. Specifically, SWI/SNF can repress select promoters that are subject to transcriptional readthrough from an upstream promoter by performing nucleosome remodeling downstream of the active transcription start site, thereby interfering with downstream promoter accessibility[19].
Role in Metabolic Gene Regulation and Nutrient Sensing
Beyond its general role in transcriptional activation, SNF2 has emerged as a critical metabolic regulator that couples gene expression to nutrient availability and cellular metabolic status[5]. The SWI/SNF complex senses and responds to changes in carbon source availability by dynamically regulating the expression of genes involved in carbohydrate metabolism. Studies in both yeast and mammalian cells have demonstrated that disruption of specific SNF2 family members or SWI/SNF subunits leads to dysregulation of glucose metabolism genes[5]. In patient-derived bladder cancer tumors, reduced expression of SMARCA2 (the mammalian BRM homolog of yeast SNF2) correlated with increased expression of the glycolytic enzyme pyruvate kinase M2 and with cancer metastasis, suggesting that SNF2-mediated metabolic regulation has direct implications for tumor biology[5].
Recent evidence indicates that SNF2 function is dynamically regulated by acetylation of the Snf2 protein itself, and this post-translational modification plays a critical role in controlling both recruitment and release of SWI/SNF from stress-responsive metabolic genes[38]. Upon nutrient deprivation-induced stress, Snf2 becomes acetylated by histone acetyltransferases, and this acetylation regulates both the occupancy and residency time of SWI/SNF at promoters of stress-responsive genes[38]. The bromodomain of Snf2, in addition to recognizing histone acetylation marks, can engage in intramolecular interactions with acetylated lysine residues on Snf2 itself, and this self-interaction negatively regulates binding and remodeling of acetylated nucleosomes, thereby fine-tuning SWI/SNF activity[38]. The increased acetylation of Snf2 during stress specifically facilitates release of the complex from genes that are repressed by stress while simultaneously increasing occupancy at genes that are upregulated, suggesting an elegant regulatory mechanism by which post-translational modification of the catalytic subunit itself serves as a switch for dynamic gene regulation[38].
Beyond glucose and carbon metabolism, emerging work has identified roles for SNF2 in lipid metabolism and amino acid metabolism regulation[5]. The SNF2 complex also plays a role in sensing sulfur metabolic status, with the methyltransferase Hmt1 targeting SNF2 for arginine methylation, and loss of Hmt1 catalytic activity reducing Snf2-mediated nucleosome remodeling at the promoters of stress response genes[5]. These findings collectively demonstrate that SNF2 functions not merely as a general transcriptional activator but as a sophisticated metabolic sensor that couples gene expression to the nutritional and biochemical status of the cell.
Roles in DNA Repair and Genome Stability
Beyond transcriptional regulation, SNF2 and related Snf2 family members play critical roles in maintaining genomic integrity and responding to DNA damage[7][49][50][52]. The Rad54 protein, which belongs to the Snf2/Swi2 family of SF2 helicase-like ATPases, functions as a potent and processive motor protein essential for homologous recombination-based DNA repair[52]. Rad54 acts in concert with Rad51 recombinase, the central protein of homologous recombination, facilitating the key reactions of homology search and DNA strand invasion[17][52]. Unlike SNF2, which is primarily involved in chromatin remodeling for transcriptional regulation, Rad54 executes distinct functions in recombinational DNA repair, including displacement of nucleosomes at sites of double-strand breaks and facilitation of Rad51 filament formation[17][52].
The ATP hydrolysis activity of Rad54 is absolutely essential for its function in DNA repair, as mutations in the Walker A box that abolish ATP hydrolysis display phenotypes identical to null mutations, indicating that mechanical translocation on DNA is central to the repair function[17][52]. Rad54 translocates on double-stranded DNA as a potent processive motor, and this translocation activity appears critical for its multiple roles in homologous recombination, including chromatin remodeling at recombination sites and stabilization of Rad51-ssDNA filaments[17][52].
Several other Snf2 family members have been specifically implicated in the DNA damage response. The INO80 Snf2 complex is involved in repair of DNA lesions, with deletion of INO80 leading to aberrant spreading of H2A.Z histone variants and resulting in defects in DNA repair and replication fork stability[3]. The mammalian SMARCAL1 Snf2 family member is recruited to sites of DNA damage through recruitment by replication protein A (RPA) and plays a role in fork reversal, a critical response to replication stress in which stalled replication forks are remodeled into four-way DNA junctions[50][53]. SMARCAL1 shows specific protective functions for telomeres during replication stress that are not shared by other fork remodelers ZRANB3 or HLTF[53].
Regulation by Post-Translational Modifications
SNF2 function is subject to extensive regulation through post-translational modifications that modulate its activity, cellular localization, and protein-protein interactions. Acetylation of Snf2 itself represents a particularly important regulatory mechanism, with the protein being acetylated by the Gcn5 histone acetyltransferase[38]. As noted above, this acetylation regulates both recruitment and release of the SWI/SNF complex from target genes, serving as a molecular signal of metabolic status and stress conditions. The acetylation of Snf2 also affects the binding capacity of the Snf2 bromodomain, as the intramolecular interaction between the bromodomain and acetylated Snf2 lysine residues can competitively inhibit binding to acetylated histones[38].
Arginine methylation of Snf2 has also been documented, with the methyltransferase Hmt1 targeting Snf2 in yeast[5]. Interestingly, loss of Hmt1 catalytic activity specifically reduces Snf2-mediated nucleosome remodeling at the promoters of stress response genes, suggesting that methylation of Snf2 facilitates its chromatin remodeling activity under stress conditions[5]. The expression of Hmt1 is itself reduced under stress conditions, potentially allowing for increased transcriptional stochasticity and phenotypic heterogeneity in cell populations facing nutrient limitation, which could facilitate survival of subpopulations through altered metabolic gene expression patterns[5].
Other post-translational modifications affecting Snf2 and SWI/SNF function include phosphorylation by checkpoint kinases in response to DNA damage, which modulates recruitment of the complex to DNA damage sites[49]. The dynamic regulation of SNF2 through multiple post-translational modification pathways demonstrates that the protein does not function as a static catalytic enzyme but rather as an information-processing node that integrates multiple cellular signals.
Cellular Localization and Subcellular Distribution
SNF2 functions exclusively within the nucleus, where it has access to chromatin substrates[3][4][7]. The protein localizes to both euchromatic regions, where it facilitates transcription of actively expressed genes, and to heterochromatic regions, where it may play roles in maintaining proper chromatin architecture. Recent studies have revealed that SNF2 occupancy at promoters is highly dynamic, with the protein showing rapid recruitment to promoters upon activation of target genes and relatively fast release when genes are repressed[4][38]. This dynamic behavior is particularly evident during stress responses, where Snf2 recruitment to stress-responsive genes increases rapidly upon induction and then decreases as cells adapt to stress conditions[38].
The bromodomain of Snf2 plays a critical role in directing SNF2 localization to specific chromatin regions, as this domain binds to acetylated histone marks that are enriched at active gene promoters and at histone variant-containing nucleosomes[8]. Loss of the Snf2 bromodomain reduces occupancy of the SWI/SNF complex at genes with high histone acetylation levels, particularly during stress responses[38]. In addition to active gene promoters, SNF2 is recruited to regulatory regions including enhancers and silencers, where it facilitates binding of transcription factors and modulation of chromatin accessibility[7].
Evolutionary Conservation and SNF2 Family Structure
SNF2 is highly conserved among eukaryotes, with homologous proteins identified in organisms ranging from unicellular fungi to plants, invertebrates, and mammals[1][3][6][7][25]. This remarkable evolutionary conservation reflects the fundamental importance of ATP-dependent chromatin remodeling for eukaryotic gene regulation and genome stability. In mammals, at least two major SNF2 family members directly homologous to yeast SNF2 have been identified: SMARCA4 (also called BRG1, the human SNF2 ortholog) and SMARCA2 (also called BRM)[7][22]. Both of these mammalian SNF2 proteins function as catalytic subunits of BAF (Brg1-associated factor) complexes that are functionally analogous to yeast SWI/SNF[7][22].
The broader SNF2 family has been classified into multiple subfamilies based on structural features and functional characteristics[25][51]. These subfamilies include the Swi2/Snf2-like proteins (including the yeast SNF2 and mammalian SMARCA4/SMARCA2), the Swr1-like proteins (involved in histone variant deposition), the SS01653-like proteins, the Rad54-like proteins (involved in DNA recombination), the Rad5/6-like proteins, and the distant SMARCAL1-like proteins[59]. Each subfamily appears to have distinct properties of the ATPase motor that are tuned to specific functional roles, as evidenced by the observation that helicase-like regions can be functionally swapped within but not between subfamilies[51].
The existence of multiple SNF2 family members in higher eukaryotes appears to reflect evolutionary specialization and expansion of the chromatin remodeling machinery. In mammals, the duplication of SNF2 into SMARCA4 and SMARCA2 has allowed for specialization of function, with both proteins capable of forming multiple distinct complexes containing different combinations of auxiliary subunits, thereby creating a more complex and nuanced chromatin remodeling machinery than exists in yeast[7][39].
Disease Relevance and Cancer Mutations
The medical significance of SNF2 and its mammalian homologs has become increasingly apparent with the discovery that mutations and dysregulation of SNF2 family proteins are associated with human diseases, particularly cancers[2][5][7]. The motif V region of human BRG1 (the SMARCA4 ortholog of yeast SNF2) has been identified as a potential mutation hotspot for amino acid substitutions associated with various cancers[2]. More broadly, components of the SWI/SNF chromatin remodeling complex are mutated in greater than 20% of all human cancers, making SWI/SNF one of the most frequently altered complexes in human malignancies[5].
Several specific disease associations have been documented. In T-cell differentiation, mutations in SMARCA4 block normal T-cell development, resulting in predominantly double-negative (CD8− and CD4−) T-cell populations and T-cell lymphomas[7][39]. In murine models, conditional mutations in Brg1 (SMARCA4) block T-cell differentiation, and complete ablation results in predominantly double-negative T-cell populations[7][31][39]. Mutations in SMARCB1, another integral BAF complex component, result in bone marrow failure and T-cell lymphoma in humans[39]. Loss of the auxiliary subunit SMARCC1 impairs hematopoietic stem cell maintenance and leads to hematopoietic dysfunction[39].
The finding that SWI/SNF dysfunction often leads to cancer rather than loss of cellular viability suggests that dysregulation of metabolic gene expression and chromatin accessibility at oncogenic pathways may contribute to tumorigenesis[5]. This insight has opened therapeutic opportunities, as the metabolic vulnerabilities created by SWI/SNF dysregulation in cancer cells may be exploitable for targeted therapy[5]. Understanding the precise mechanisms by which SWI/SNF mutations contribute to cancer phenotypes remains an active area of investigation, but dysregulation of metabolic gene expression, altered epigenetic landscapes, and aberrant chromatin accessibility appear to be central features[5].
Specialized Functions in Distinct Biological Processes
Beyond the central roles in transcriptional regulation and chromatin remodeling, SNF2 and related Snf2 family members have been implicated in specialized biological processes. In Tetrahymena thermophila, a ciliate organism with nuclear dualism, specific Snf2 family proteins including IswiG_Tt and Rad5_Tt are required for selection of the developing micronucleus (hMIC) and subsequent generation of gametic pronuclei[35]. These Snf2 proteins promote euchromatin formation at selected micronuclei through remodeling of DNA lesions and chromatin structure changes, and their disruption leads to persistent DNA damage markers and degradation of all potential gametic nuclei[35].
In Drosophila embryos, the Lodestar (Lds) Snf2 family protein, the Drosophila ortholog of human TTF2, exhibits a dual function during mitosis in both driving removal of nascent transcripts from mitotic chromatin and facilitating sister chromatid resolution[57]. The ATPase activity of Lds is essential for prompt eviction of engaged transcripts as cells enter mitosis, and Lds also cooperates with Topoisomerase 2 to ensure efficient DNA decatenation and mitotic fidelity[57]. This dual function uncovered in Drosophila suggests that SNF2 family proteins may have unexpected roles in coordinating multiple cellular processes simultaneously.
Conclusion and Significance
The SNF2 protein represents a paradigmatic example of how cells have evolved sophisticated machinery to solve the fundamental problem of chromatin-templated gene regulation. Operating as the catalytic engine of the SWI/SNF chromatin remodeling complex, SNF2 couples the hydrolysis of ATP to mechanical remodeling of nucleosome structure and positioning, thereby making DNA sequences accessible to transcription factors and regulatory enzymes[1][3][7]. The protein's structural organization, with its helicase-like ATPase domain, bromodomain for recognizing histone modifications, and other regulatory domains, reflects millions of years of evolutionary refinement for coordinating access to genomic information with cellular metabolic and developmental signals[3][25][27][30].
The mechanisms by which SNF2 executes chromatin remodeling have been illuminated through structural, biochemical, and genetic studies, revealing an intricate process involving ATP-driven conformational changes between RecA-like domains, translocation along DNA, and orchestrated deformation of the histone octamer[2][27][30][56]. The regulation of SNF2 through post-translational modifications, transcription factor-mediated recruitment, and complex composition provides multiple layers of control ensuring that chromatin remodeling activities occur at appropriate genomic locations and in response to cellular signals[13][38][58].
Beyond its canonical role in transcriptional activation, SNF2 plays increasingly appreciated roles in metabolic gene regulation, coupling gene expression to nutrient availability and cellular energy status[5]. The discovery that SNF2 regulates metabolic pathways through dynamic control of gene expression suggests that understanding SNF2 function provides insights into fundamental aspects of cellular metabolism and adaptation to environmental changes. The conservation of SNF2 function from yeast to mammals, combined with the medical significance of SNF2 mutations in human cancer and genetic diseases, underscores the biological importance of this enzyme and the widespread consequences when chromatin remodeling goes awry[5][7].
The remaining outstanding questions about SNF2 function include the precise mechanisms determining whether SNF2 catalyzes nucleosome sliding versus eviction on specific substrates, the full extent of SNF2 regulatory roles in processes beyond transcription, and how multiple post-translational modifications integrate to control SNF2 activity in response to diverse cellular signals. Continued investigation of SNF2 structure, mechanism, and biology promises to yield further insights into the regulation of eukaryotic gene expression and the pathogenic consequences of its dysregulation in human disease. The functional annotation of SNF2 thus remains an active area of research with direct implications for understanding both normal cellular physiology and the molecular basis of cancer and other genetic diseases.
Citations
- https://pubmed.ncbi.nlm.nih.gov/1516829/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1168809/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4162295/
- https://www.yeastgenome.org/locus/S000005816
- https://portlandpress.com/biochemsoctrans/article/52/3/1327/234375/The-SWI-SNF-chromatin-remodeling-complex-a
- https://genesdev.cshlp.org/content/7/4/583.short
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4348595/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1783998/
- https://portlandpress.com/essaysbiochem/article/63/1/45/173/Nucleosome-remodelling-structural-insights-into
- https://pubmed.ncbi.nlm.nih.gov/18837912/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5837817/
- https://portlandpress.com/biochemsoctrans/article/41/6/1640/68103/A-snapshot-of-Snf2-enzymes-in-fission-yeast
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3105400/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3171615/
- https://genesdev.cshlp.org/content/6/12a/2288.full.pdf
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11419397/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4004355/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC369633/
- https://academic.oup.com/nar/article/51/16/8496/7231339
- https://www.epigenhub.com/p/5421/swi2snf2-polyclonal-antibody/
- https://pubmed.ncbi.nlm.nih.gov/7608126/
- https://academic.oup.com/nar/article/34/15/4160/3111884
- https://www.uniprot.org/uniref/UniRef50_P22082
- https://www.uniprot.org/uniprotkb/P22082/entry
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9786623/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC232117/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1461034/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4201291/
- https://www.pnas.org/doi/10.1073/pnas.92.21.9722
- https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2022.910663/full
- https://pubmed.ncbi.nlm.nih.gov/14690608/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC367125/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3338269/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1204454/
- https://journals.plos.org/plosone/article?id=10.1371%2Fjournal.pone.0008111
- https://pubmed.ncbi.nlm.nih.gov/1648006/
- https://pubmed.ncbi.nlm.nih.gov/32971328/
- https://academic.oup.com/nar/article/34/10/2887/2543556
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8092973/
- https://pubmed.ncbi.nlm.nih.gov/27399259/
- https://genesdev.cshlp.org/content/13/18/2369.long
- https://elifesciences.org/articles/46057
- https://pmc.ncbi.nlm.nih.gov/articles/PMC10481674/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC135607/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7074175/