Deep Research
Falcon
(SWI3-deep-research-falcon.md)
Falcon
(SWI3-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Comprehensive Research Report: SWI3 / TYE2 (YJL176C) — SWI/SNF complex subunit Swi3 in Saccharomyces cerevisiae (UniProt P32591)
0) Target verification (critical identity check)
The requested protein is S. cerevisiae SWI3, explicitly mapped to ORF YJL176C in experimental literature, and described as a subunit of the SWI/SNF chromatin remodeling complex. In an oxygen-response localization study, SWI3 is listed in a table as “YJL176C SWI3 Subunit of the SWI/SNF chromatin remodeling complex,” aligning with the UniProt-provided organism and functional description. (dastidar2012thenuclearlocalization pages 2-4)
A key ambiguity in the literature is the presence of a Swi3 homolog/paralog Swh3, which partners with Sth1 in a distinct essential SWI/SNF-related complex (RSC-related), and does not substitute for Swi3. This distinction helps prevent confusion with non-canonical “SWI3-like” proteins. (treich1997interactionofa pages 6-7, treich1997interactionofa pages 1-2)
1) Key concepts and current understanding
1.1 SWI/SNF chromatin remodeling complexes (definition)
SWI/SNF (“Switch/Sucrose Non-Fermentable”) is an ATP-dependent chromatin remodeling complex that alters nucleosome structure to regulate DNA accessibility, thereby impacting transcription. Foundational yeast work established that the complex uses ATP to perturb nucleosome structure and assist transcriptional regulation, and that SWI3 is among the SWI/SNF gene products required for proper transcriptional control of many yeast genes. (cairns1996tfgtaf30anc1acomponent pages 1-1, treich1997interactionofa pages 1-2)
1.2 What SWI3 is (molecular function at the protein level)
SWI3 is not an enzyme; it is a core structural/regulatory subunit within SWI/SNF. Evidence supports at least three mechanistic roles:
1) Complex membership and stability/assembly: SWI3 is detected in purified SWI/SNF preparations monitored by immunoblotting, supporting that it is physically present in the complex. (cairns1996tfgtaf30anc1acomponent pages 1-1)
2) Protein–protein interaction hub: SWI3 interacts with the SWI/SNF ATPase Snf2 in two-hybrid assays, linking it to the catalytic motor subunit. (treich1997interactionofa pages 4-4, treich1997interactionofa pages 1-2)
3) DNA/nucleosome engagement via conserved domains: the SWIRM domain of Swi3 is a conserved structural module whose structure and biochemical function were experimentally characterized; it can bind DNA (and nucleosomal DNA) and contributes to SWI/SNF assembly and stability. (da2006structureandfunction pages 4-5, da2006structureandfunction pages 2-3)
2) Domain architecture and structure (evidence-based)
2.1 SWIRM domain: structure, DNA binding, and functional consequences
A high-confidence mechanistic advance for SWI3 is the direct biochemical/structural characterization of its SWIRM domain:
-
Structure: Swi3 SWIRM forms a globular antiparallel four-helix bundle (structural description from the SWIRM-domain crystal structure study). (da2006structureandfunction pages 2-3)
-
DNA binding: The Swi3 SWIRM domain binds nucleic acids with reported apparent Kd values ~72–90 nM in one set of assays, and a KD of ~105.36 ± 7.65 nM reported for binding to cruciform DNA in vitro. (da2006structureandfunction pages 4-5, da2006structureandfunction pages 2-3)
-
Mutational evidence: Structure-guided mutations in SWIRM (e.g., targeting residues implicated in binding or core stability) produce moderate to strong swi/snf phenotypes and can reduce Swi3 protein levels and defectively associate with SWI/SNF, as measured by coprecipitation with Swp73. This supports that SWIRM contributes to complex stability/assembly and likely chromatin engagement. (da2006structureandfunction pages 4-5)
2.2 SANT and coiled-coil/leucine zipper features (structure and genetics)
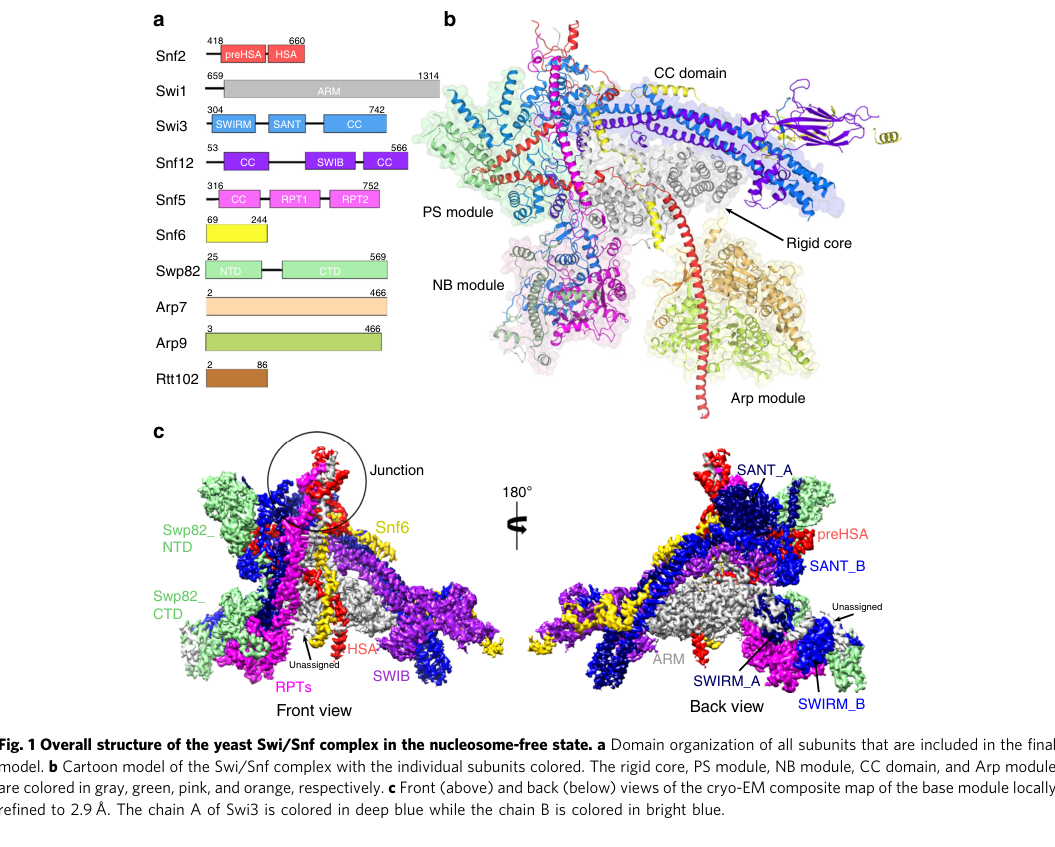
A cryo-EM structure of yeast SWI/SNF in a nucleosome-free state shows Swi3 subunits with labeled domains, including SWIRM_A/B and SANT_A/B consistent with two Swi3 copies in the base module. The figure also maps overall domain organization of subunits (including Swi3’s SWIRM and SANT regions). (wang2020structureofthe media e53f6d4b)
Earlier genetic work also emphasizes a functionally important leucine zipper motif in Swi3 (tested via fusion constructs including “most of Swi3 including the leucine zipper”) and reports the leucine zipper is required in that experimental context. (treich1997interactionofa pages 4-4)
2.3 SWI/SNF base-module architecture: two Swi3 copies as a structural backbone
Cryo-EM analysis indicates Swi3 is part of the backbone/scaffold of SWI/SNF:
- The model contains two Swi3 instances (Swi3A and Swi3B).
- Swi3 SWIRM domains contact Snf5 RPT regions and Swi3 contributes structural helices in coiled-coil organization with Snf12.
- Swi1 and Swi3 together form a major structural backbone of the complex. (wang2020structureofthe pages 1-2)
A visually accessible summary of this architecture—including the two Swi3 copies and labeled SWIRM/SANT regions—is present in the retrieved figure panel from Wang et al. 2020. (wang2020structureofthe media e53f6d4b)
3) Biological roles and pathways (process-level functional annotation)
3.1 Transcriptional regulation via chromatin remodeling
Foundational biochemical/genetic evidence supports that SWI3 functions within SWI/SNF to regulate transcription through chromatin remodeling:
-
SWI3 is one of the SWI/SNF gene products required for proper transcriptional control of many genes, and SWI/SNF uses ATP (via Snf2 ATPase) to perturb nucleosome structure and assist transcriptional activation. (cairns1996tfgtaf30anc1acomponent pages 1-1, treich1997interactionofa pages 1-2)
-
Swi3-containing complexes can activate transcription when tethered to a reporter system, and Swi3 interacts with Snf2 (two-hybrid), connecting it to the catalytic core of remodeling. (treich1997interactionofa pages 4-4, treich1997interactionofa pages 1-2)
3.2 Repression / transcriptional interference (recent 2024 mechanistic context)
Recent work highlights SWI/SNF roles beyond simple “activation,” including gene repression in specific contexts:
- A 2024 Molecular Cell study found that SWI/SNF remodeling regulates transcriptional interference and gene repression. A Swi3 truncation allele (swi3-E815X) showed relatively mild global effects among tested targets, but displayed significant upregulation of certain proximal isoforms (ODC2PROX and HNT1PROX) without significant changes in corresponding distal isoforms, consistent with SWI/SNF involvement in repression at proximal TSSs in LUTI-associated regulation. (morse2024swisnfchromatinremodeling pages 12-15)
This study provides a modern experimental anchor that SWI3 contributes to context-specific repression mechanisms, not only activation. (morse2024swisnfchromatinremodeling pages 12-15)
4) Subcellular localization and real-world cellular implementation
4.1 Nuclear localization in normoxia
SWI3 is expected to be nuclear as a chromatin remodeler subunit, and imaging experiments confirm this expectation: in air/normoxia, SWI3 (and other hypoxia-altered SWI/SNF subunits) are predominantly nuclear. (dastidar2012thenuclearlocalization pages 1-2)
4.2 Condition-dependent relocalization: hypoxia-induced cytosolic accumulation
A well-defined real-world physiological implementation of SWI3 regulation is oxygen-responsive localization:
- Under hypoxia, over 95% of Swi3 (and several other SWI/SNF components) accumulate in the cytosol. (dastidar2012thenuclearlocalization pages 1-2)
- Time-course behavior: a subset becomes predominantly cytosolic within ~6 h, and all become predominantly cytosolic after 12 h hypoxia exposure; reoxygenation can restore nuclear localization quickly (within ~1 hour for many proteins), with Swi3 shown as an example. (dastidar2012thenuclearlocalization pages 1-2)
Mechanistically, the study notes differential behavior among SWI/SNF components: Swi2/Arp7/Arp9 remain nuclear while Swi3 is affected, implying oxygen can alter nuclear SWI/SNF composition by changing which subunits are nuclear. (dastidar2012thenuclearlocalization pages 8-11)
The study also reports a quantitative gene list context: 112 oxygen-regulated target genes of Swi3 are tabulated for wild-type HAP1 cells (supporting SWI/SNF-Swi3 involvement in oxygen-regulated transcription programs). (dastidar2012thenuclearlocalization pages 6-8)
5) Interaction partners and assembly constraints
5.1 Interaction with Snf2 and functional implications
A two-hybrid screen recovered Swi3 interactions involving Snf2, supporting direct or proximal interaction between Swi3 and the ATPase motor. This reinforces a model where Swi3 helps connect regulatory/scaffold elements to the remodeling ATPase. (treich1997interactionofa pages 4-4, treich1997interactionofa pages 1-2)
5.2 SWIRM-mediated contacts in cryo-EM base module
Structural analysis indicates Swi3 SWIRM domains contact Snf5 RPT regions (electrostatic interactions) within the SWI/SNF base module, supporting a direct architectural role for SWI3 in organizing conserved core components. (wang2020structureofthe pages 1-2)
6) Quantitative/statistical highlights from recent and classic studies
Key quantitative points directly supported by the retrieved sources include:
-
SWIRM domain DNA binding: Swi3 SWIRM cruciform DNA binding KD ~105.36 ± 7.65 nM, with other apparent binding estimates in the ~72–90 nM range. (da2006structureandfunction pages 4-5, da2006structureandfunction pages 2-3)
-
Reporter assay quantitation (Swi3 homolog context): LexA-Swh3 activation is ~25-fold lower than LexA-Snf2 in a lexAop reporter assay, with β-galactosidase activities including (2 operators): LexA-Swh3 5.1 U, LexA-Snf2 120 U, LexA-Sth1 12 U; standard errors reported as <7%. Although these values are for Swh3 (a Swi3 homolog/paralog), they contextualize differential activation capacity among Swi3-like proteins and highlight the importance of not confusing Swi3 with Swh3. (treich1997interactionofa pages 4-4, treich1997interactionofa pages 1-2)
-
Hypoxia relocalization: In hypoxia, >95% of Swi3 localizes to cytosol, reversing upon reoxygenation. (dastidar2012thenuclearlocalization pages 1-2)
-
Recent mutant phenotype (2024): swi3-E815X shows significant upregulation of specific proximal isoforms (ODC2PROX, HNT1PROX) with minimal changes in corresponding distal isoforms, supporting a role in repression at proximal TSSs in transcriptional interference; the excerpt is qualitative for effect size but explicit about significance and directionality. (morse2024swisnfchromatinremodeling pages 12-15)
7) Recent developments (prioritizing 2023–2024)
7.1 2024: SWI/SNF-mediated repression via transcriptional interference
The 2024 Molecular Cell work provides direct evidence that SWI/SNF remodeling contributes to repression mechanisms in the setting of LUTI-associated regulation, with a Swi3 truncation allele showing measurable target-specific effects. (morse2024swisnfchromatinremodeling pages 12-15)
7.2 How this reframes functional annotation for SWI3
A modern view supported by the above evidence is that SWI3’s “primary function” is best annotated as:
- Structural/scaffold subunit of SWI/SNF required for proper complex assembly and targeting, enabling ATP-dependent remodeling by the Snf2 motor; and
- A contributor to both activation and repression outcomes depending on promoter architecture, transcription factor context, and chromatin state (as exemplified by transcriptional interference regulation). (cairns1996tfgtaf30anc1acomponent pages 1-1, morse2024swisnfchromatinremodeling pages 12-15, wang2020structureofthe pages 1-2)
8) Current applications and real-world implementations
Because yeast SWI3 is a core SWI/SNF subunit, its “applications” are primarily as a tool and model in chromatin biology:
-
Model for conserved SWI/SNF mechanisms: Structural biology has reached subunit/domain-level models (two Swi3 copies with labeled SWIRM/SANT regions), enabling mechanistic hypotheses about assembly and remodeling. (wang2020structureofthe pages 1-2, wang2020structureofthe media e53f6d4b)
-
Environmental response biology: The oxygen-dependent nuclear/cytosolic relocalization of Swi3 suggests a mechanism by which cells can rapidly rewire transcriptional programs by altering nuclear access of chromatin remodelers (quantified as >95% cytosolic under hypoxia). (dastidar2012thenuclearlocalization pages 1-2)
9) Expert analysis (grounded in authoritative sources)
The strongest mechanistic evidence base indicates SWI3 is a multi-domain architectural subunit that integrates three functional layers:
1) Scaffold/assembly (two copies in base module; backbone with Swi1). (wang2020structureofthe pages 1-2)
2) Chromatin engagement via SWIRM domain DNA/nucleosome binding and domain-dependent stability/assembly. (da2006structureandfunction pages 4-5)
3) Regulatory outcome shaping (activation and repression contexts), including proximal-TSS repression in transcriptional interference. (cairns1996tfgtaf30anc1acomponent pages 1-1, morse2024swisnfchromatinremodeling pages 12-15)
10) Evidence summary table
The following table consolidates the evidence themes, approaches, and key quantitative results.
| Evidence theme | Key findings | Key experimental approach | Representative citations with publication year + URL |
|---|---|---|---|
| Identity / complex membership | SWI3 in Saccharomyces cerevisiae corresponds to YJL176C and is a defined subunit of the Swi/Snf chromatin-remodeling complex required for transcriptional control of many yeast genes; literature also distinguishes Swi3 from the paralog/homolog Swh3 that functions with Sth1 in an essential Swi/Snf-related complex rather than canonical Swi/Snf. (treich1997interactionofa pages 1-2, dastidar2012thenuclearlocalization pages 2-4) | Biochemical purification/immunoblotting of Swi/Snf complex; genetic and two-hybrid analyses distinguishing Swi3 vs. Swh3 complexes. (cairns1996tfgtaf30anc1acomponent pages 1-1, treich1997interactionofa pages 1-2) | Cairns et al., 1996, Mol Cell Biol / https://doi.org/10.1128/mcb.16.7.3308; Treich & Carlson, 1997, Mol Cell Biol / https://doi.org/10.1128/mcb.17.4.1768 |
| Domains | Swi3 contains a SWIRM domain with direct nucleic-acid binding activity and also carries SANT and leucine-zipper/coiled-coil features noted in structural and genetic studies; SWIRM mutations impair protein stability, complex association, and growth-related Swi/Snf phenotypes. GST-Swi3 SWIRM bound cruciform DNA with reported Kd ~105.36 ± 7.65 nM; apparent nucleic-acid binding was also reported in the ~72–90 nM range for the isolated domain. (da2006structureandfunction pages 4-5, da2006structureandfunction pages 2-3) | SWIRM domain structure determination plus in vitro DNA-binding assays and structure-guided mutagenesis; mutant growth and co-precipitation assays. (da2006structureandfunction pages 4-5, da2006structureandfunction pages 2-3) | Da et al., 2006, PNAS / https://doi.org/10.1073/pnas.0510949103; Wang et al., 2020, Nat Commun / https://doi.org/10.1038/s41467-020-17229-x |
| Structure / assembly | Swi3 is a backbone/scaffold subunit of yeast Swi/Snf. Cryo-EM studies identify two Swi3 copies (Swi3A/Swi3B) in the base module; Swi3 SWIRM domains contact Snf5 RPT regions, SANT elements contribute to architecture, and Swi3 helps form the outer frame with Snf12. Earlier genetic work showed Swi3 interacts with Snf2 and that the leucine-zipper motif is functionally required. (wang2020structureofthe pages 1-2, treich1997interactionofa pages 4-4, wang2020structureofthe media e53f6d4b) | Cryo-EM structural analysis of purified Swi/Snf; yeast two-hybrid interaction mapping; functional mutational analysis of Swi3 motifs. (wang2020structureofthe pages 1-2, treich1997interactionofa pages 4-4) | Wang et al., 2020, Nat Commun / https://doi.org/10.1038/s41467-020-17229-x; Treich & Carlson, 1997, Mol Cell Biol / https://doi.org/10.1128/mcb.17.4.1768 |
| Transcriptional roles | Swi3 functions within Swi/Snf to remodel nucleosomes and support transcriptional regulation. Canonical Swi/Snf facilitates transcriptional activation, and tethering assays linked Swi3-containing complexes to activation. Recent work also supports a repressive role at selected proximal promoters: the swi3-E815X allele reduces Swi/Snf-mediated repression at some LUTI-associated TSSs, though more mildly than snf2-W935R. (treich1997interactionofa pages 1-2, treich1997interactionofa pages 4-4, morse2024swisnfchromatinremodeling pages 12-15) | Reporter/tethering assays, two-hybrid assays, and recent transcript-isoform analysis of transcriptional interference targets in swi3 mutants. (treich1997interactionofa pages 4-4, morse2024swisnfchromatinremodeling pages 12-15) | Treich & Carlson, 1997, Mol Cell Biol / https://doi.org/10.1128/mcb.17.4.1768; Morse et al., 2024, Molecular Cell / https://doi.org/10.1016/j.molcel.2024.06.029 |
| Localization | Swi3 is predominantly nuclear under normoxia but relocalizes to the cytosol under hypoxia and returns to the nucleus upon reoxygenation, indicating condition-dependent control of SWI/SNF subcellular distribution. Dastidar et al. identify Swi3 as one of six Swi/Snf proteins with this behavior. (dastidar2012thenuclearlocalization pages 11-12, dastidar2012thenuclearlocalization pages 8-11, dastidar2012thenuclearlocalization pages 1-2) | GFP/live-cell fluorescence imaging and nuclear import/localization time-course assays under hypoxia and reoxygenation. (dastidar2012thenuclearlocalization pages 11-12, dastidar2012thenuclearlocalization pages 2-4) | Dastidar et al., 2012, Cell & Bioscience / https://doi.org/10.1186/2045-3701-2-30 |
| Quantitative data | Representative quantitative findings include: isolated Swi3 SWIRM domain DNA binding Kd ~105.36 ± 7.65 nM and ~72–90 nM in related assays; LexA-Swh3 activity was ~25-fold lower than LexA-Snf2 in reporter assays, with standard errors <7% in similar measurements; under hypoxia, >95% of Swi3 and several other Swi/Snf proteins were cytosolic, versus predominantly nuclear in air; 112 oxygen-regulated target genes were listed for Swi3 in wild-type HAP1 cells; swi3-E815X caused significant upregulation of ODC2PROX and HNT1PROX with relatively mild genome-wide effects compared with snf2 mutants. (da2006structureandfunction pages 4-5, da2006structureandfunction pages 2-3, treich1997interactionofa pages 4-4, dastidar2012thenuclearlocalization pages 6-8, dastidar2012thenuclearlocalization pages 1-2, morse2024swisnfchromatinremodeling pages 12-15) | Quantitative DNA-binding assays, β-gal reporter assays, fluorescence localization scoring across counted cells, and transcript/TSS profiling in mutant strains. (da2006structureandfunction pages 4-5, treich1997interactionofa pages 4-4, dastidar2012thenuclearlocalization pages 11-12, morse2024swisnfchromatinremodeling pages 12-15) | Da et al., 2006, PNAS / https://doi.org/10.1073/pnas.0510949103; Treich & Carlson, 1997, Mol Cell Biol / https://doi.org/10.1128/mcb.17.4.1768; Dastidar et al., 2012, Cell & Bioscience / https://doi.org/10.1186/2045-3701-2-30; Morse et al., 2024, Molecular Cell / https://doi.org/10.1016/j.molcel.2024.06.029 |
Table: This table summarizes experimentally supported functional annotation evidence for yeast SWI3/YJL176C (UniProt P32591), organized by identity, domains, structure, transcriptional role, localization, and quantitative findings. It is useful as a compact evidence map tied to specific primary studies and context citations.
11) Visual evidence (SWI3 domain placement within SWI/SNF)
A key cryo-EM figure panel (Wang et al., 2020) shows the overall SWI/SNF architecture and explicitly labels SWIRM_A/B and SANT_A/B corresponding to the two Swi3 copies (Swi3A/Swi3B) within the base module. (wang2020structureofthe media e53f6d4b)
12) Limitations of this evidence set
- TYE2 synonym: Within the retrieved full-text evidence excerpts, SWI3 is mapped to YJL176C and SWI/SNF subunit roles, but the specific synonym “TYE2” did not appear in the excerpts. Therefore, TYE2 is not explicitly supported here from the gathered documents.
- Main-paper nucleosome-bound SWI/SNF cryo-EM figure: The supplementary text available for Han et al. 2020 did not contain the requested architecture figure; thus, the visual support is taken from the nucleosome-free cryo-EM structure paper (Wang et al. 2020) that explicitly provides a domain map and base-module architecture. (wang2020structureofthe media e53f6d4b)
13) Minimal functional annotation statement (recommended)
SWI3 (YJL176C; UniProt P32591) encodes a core SWI/SNF chromatin remodeling complex subunit. Swi3 contains a SWIRM domain with direct DNA/nucleosome binding activity and structural roles in complex assembly, and participates as one of (at least) two Swi3 copies in the SWI/SNF base module that forms the complex backbone with Swi1 and contacts other conserved subunits such as Snf5. Swi3 is predominantly nuclear under normoxia but undergoes regulated cytosolic relocalization under hypoxia (>95% cytosolic), consistent with dynamic control of SWI/SNF nuclear function. Functionally, Swi3 supports context-dependent transcriptional regulation, including roles in transcriptional interference-associated repression as indicated by swi3-E815X effects on proximal isoform expression. (da2006structureandfunction pages 4-5, wang2020structureofthe pages 1-2, dastidar2012thenuclearlocalization pages 1-2, morse2024swisnfchromatinremodeling pages 12-15)
References
-
(dastidar2012thenuclearlocalization pages 2-4): Ranita Ghosh Dastidar, Jagmohan Hooda, Ajit Shah, Thai M Cao, Robert Michael Henke, and Li Zhang. The nuclear localization of swi/snf proteins is subjected to oxygen regulation. Cell & Bioscience, 2:30-30, Aug 2012. URL: https://doi.org/10.1186/2045-3701-2-30, doi:10.1186/2045-3701-2-30. This article has 87 citations and is from a peer-reviewed journal.
-
(treich1997interactionofa pages 6-7): Isabelle Treich and Marian Carlson. Interaction of a swi3 homolog with sth1 provides evidence for a swi/snf-related complex with an essential function in saccharomyces cerevisiae. Molecular and Cellular Biology, 17:1768-1775, Apr 1997. URL: https://doi.org/10.1128/mcb.17.4.1768, doi:10.1128/mcb.17.4.1768. This article has 65 citations and is from a domain leading peer-reviewed journal.
-
(treich1997interactionofa pages 1-2): Isabelle Treich and Marian Carlson. Interaction of a swi3 homolog with sth1 provides evidence for a swi/snf-related complex with an essential function in saccharomyces cerevisiae. Molecular and Cellular Biology, 17:1768-1775, Apr 1997. URL: https://doi.org/10.1128/mcb.17.4.1768, doi:10.1128/mcb.17.4.1768. This article has 65 citations and is from a domain leading peer-reviewed journal.
-
(cairns1996tfgtaf30anc1acomponent pages 1-1): Bradley R. Cairns, N. Lynn Henry, and Roger D. Kornberg. Tfg/taf30/anc1, a component of the yeast swi/snf complex that is similar to the leukemogenic proteins enl and af-9. Molecular and Cellular Biology, 16:3308-3316, Jul 1996. URL: https://doi.org/10.1128/mcb.16.7.3308, doi:10.1128/mcb.16.7.3308. This article has 213 citations and is from a domain leading peer-reviewed journal.
-
(treich1997interactionofa pages 4-4): Isabelle Treich and Marian Carlson. Interaction of a swi3 homolog with sth1 provides evidence for a swi/snf-related complex with an essential function in saccharomyces cerevisiae. Molecular and Cellular Biology, 17:1768-1775, Apr 1997. URL: https://doi.org/10.1128/mcb.17.4.1768, doi:10.1128/mcb.17.4.1768. This article has 65 citations and is from a domain leading peer-reviewed journal.
-
(da2006structureandfunction pages 4-5): Guoping Da, Jeffrey Lenkart, Kehao Zhao, Ramin Shiekhattar, Bradley R. Cairns, and Ronen Marmorstein. Structure and function of the swirm domain, a conserved protein module found in chromatin regulatory complexes. Proceedings of the National Academy of Sciences of the United States of America, 103:2057-2062, Feb 2006. URL: https://doi.org/10.1073/pnas.0510949103, doi:10.1073/pnas.0510949103. This article has 139 citations and is from a highest quality peer-reviewed journal.
-
(da2006structureandfunction pages 2-3): Guoping Da, Jeffrey Lenkart, Kehao Zhao, Ramin Shiekhattar, Bradley R. Cairns, and Ronen Marmorstein. Structure and function of the swirm domain, a conserved protein module found in chromatin regulatory complexes. Proceedings of the National Academy of Sciences of the United States of America, 103:2057-2062, Feb 2006. URL: https://doi.org/10.1073/pnas.0510949103, doi:10.1073/pnas.0510949103. This article has 139 citations and is from a highest quality peer-reviewed journal.
-
(wang2020structureofthe media e53f6d4b): Chengcheng Wang, Zhouyan Guo, Xiechao Zhan, Fenghua Yang, Mingxuan Wu, and Xiaofeng Zhang. Structure of the yeast swi/snf complex in a nucleosome free state. Nature Communications, Jul 2020. URL: https://doi.org/10.1038/s41467-020-17229-x, doi:10.1038/s41467-020-17229-x. This article has 29 citations and is from a highest quality peer-reviewed journal.
-
(wang2020structureofthe pages 1-2): Chengcheng Wang, Zhouyan Guo, Xiechao Zhan, Fenghua Yang, Mingxuan Wu, and Xiaofeng Zhang. Structure of the yeast swi/snf complex in a nucleosome free state. Nature Communications, Jul 2020. URL: https://doi.org/10.1038/s41467-020-17229-x, doi:10.1038/s41467-020-17229-x. This article has 29 citations and is from a highest quality peer-reviewed journal.
-
(morse2024swisnfchromatinremodeling pages 12-15): Kaitlin Morse, Alena L. Bishop, Sarah Swerdlow, Jessica M. Leslie, and Elçin Ünal. Swi/snf chromatin remodeling regulates transcriptional interference and gene repression. Aug 2024. URL: https://doi.org/10.1016/j.molcel.2024.06.029, doi:10.1016/j.molcel.2024.06.029. This article has 21 citations and is from a highest quality peer-reviewed journal.
-
(dastidar2012thenuclearlocalization pages 1-2): Ranita Ghosh Dastidar, Jagmohan Hooda, Ajit Shah, Thai M Cao, Robert Michael Henke, and Li Zhang. The nuclear localization of swi/snf proteins is subjected to oxygen regulation. Cell & Bioscience, 2:30-30, Aug 2012. URL: https://doi.org/10.1186/2045-3701-2-30, doi:10.1186/2045-3701-2-30. This article has 87 citations and is from a peer-reviewed journal.
-
(dastidar2012thenuclearlocalization pages 8-11): Ranita Ghosh Dastidar, Jagmohan Hooda, Ajit Shah, Thai M Cao, Robert Michael Henke, and Li Zhang. The nuclear localization of swi/snf proteins is subjected to oxygen regulation. Cell & Bioscience, 2:30-30, Aug 2012. URL: https://doi.org/10.1186/2045-3701-2-30, doi:10.1186/2045-3701-2-30. This article has 87 citations and is from a peer-reviewed journal.
-
(dastidar2012thenuclearlocalization pages 6-8): Ranita Ghosh Dastidar, Jagmohan Hooda, Ajit Shah, Thai M Cao, Robert Michael Henke, and Li Zhang. The nuclear localization of swi/snf proteins is subjected to oxygen regulation. Cell & Bioscience, 2:30-30, Aug 2012. URL: https://doi.org/10.1186/2045-3701-2-30, doi:10.1186/2045-3701-2-30. This article has 87 citations and is from a peer-reviewed journal.
-
(dastidar2012thenuclearlocalization pages 11-12): Ranita Ghosh Dastidar, Jagmohan Hooda, Ajit Shah, Thai M Cao, Robert Michael Henke, and Li Zhang. The nuclear localization of swi/snf proteins is subjected to oxygen regulation. Cell & Bioscience, 2:30-30, Aug 2012. URL: https://doi.org/10.1186/2045-3701-2-30, doi:10.1186/2045-3701-2-30. This article has 87 citations and is from a peer-reviewed journal.
Artifacts
Citations
- dastidar2012thenuclearlocalization pages 2-4
- da2006structureandfunction pages 2-3
- da2006structureandfunction pages 4-5
- treich1997interactionofa pages 4-4
- wang2020structureofthe pages 1-2
- morse2024swisnfchromatinremodeling pages 12-15
- dastidar2012thenuclearlocalization pages 1-2
- dastidar2012thenuclearlocalization pages 8-11
- dastidar2012thenuclearlocalization pages 6-8
- treich1997interactionofa pages 6-7
- treich1997interactionofa pages 1-2
- dastidar2012thenuclearlocalization pages 11-12
- https://doi.org/10.1128/mcb.16.7.3308;
- https://doi.org/10.1128/mcb.17.4.1768
- https://doi.org/10.1073/pnas.0510949103;
- https://doi.org/10.1038/s41467-020-17229-x
- https://doi.org/10.1038/s41467-020-17229-x;
- https://doi.org/10.1128/mcb.17.4.1768;
- https://doi.org/10.1016/j.molcel.2024.06.029
- https://doi.org/10.1186/2045-3701-2-30
- https://doi.org/10.1186/2045-3701-2-30;
- https://doi.org/10.1186/2045-3701-2-30,
- https://doi.org/10.1128/mcb.17.4.1768,
- https://doi.org/10.1128/mcb.16.7.3308,
- https://doi.org/10.1073/pnas.0510949103,
- https://doi.org/10.1038/s41467-020-17229-x,
- https://doi.org/10.1016/j.molcel.2024.06.029,
Perplexity
(SWI3-deep-research-perplexity.md)
Perplexity
(SWI3-deep-research-perplexity.md)SWI3: A Critical Regulatory Subunit of the SWI/SNF Chromatin Remodeling Complex in Saccharomyces cerevisiae
The SWI3 gene of Saccharomyces cerevisiae, encoding a key structural and functional subunit of the SWI/SNF (SWITCH/SUCROSE NONFERMENTING) chromatin remodeling complex, represents one of the most fundamentally important regulatory proteins in eukaryotic transcriptional control and chromosome dynamics. Originally identified through genetic screens for mutants defective in mating-type switching and sucrose fermentation, SWI3 has emerged as a versatile regulatory factor whose functions extend far beyond its initial characterization, encompassing critical roles in transcriptional activation and repression, DNA replication fork stabilization, cellular stress responses, and oxygen-dependent gene regulation.[2][12][21] The SWI3 protein functions as a core structural component of the ~1.14 MDa yeast SWI/SNF complex, which contains ten to twelve polypeptides that work in concert through ATP-dependent nucleosome remodeling to control access to chromosomal DNA.[21][48] Unlike the catalytic ATPase subunits that provide the energy for chromatin remodeling, Swi3 exerts regulatory control over complex assembly, histone displacement efficiency, targeting to specific genes, and functional specialization of different SWI/SNF complexes, positioning it as a critical hub for integrating multiple regulatory signals into productive chromatin remodeling events. Recent discoveries have revealed that SWI3 also possesses unique functions independent of the canonical SWI/SNF complex, particularly in suppressing oxygen consumption and regulating the expression of genes encoding mitochondrial respiratory chain components in an oxygen-dependent manner, highlighting the multifaceted regulatory roles this protein plays in cellular physiology.
Identification and Evolutionary Context of the SWI3 Gene
The SWI3 gene was originally identified as part of the SWI gene family through classical genetic screens in budding yeast for mutations affecting mating-type switching and carbon source utilization.[2][14] The gene symbol SWI3 corresponds to the yeast chromosomal locus YJL176C, and the protein is also known by the alternative designation TYE2, reflecting its earlier identification as a transcription regulatory protein required for Ty1 transposon expression.[3][6] The discovery that TYE2 and SWI3 were the same gene represented an important convergence of findings from two independent research directions: one focused on the fundamental mechanisms of mating-type switching and transcriptional regulation, and another examining the regulation of retrotransposon activity.[3] This nomenclature reflects the historical nature of gene discovery in yeast, where researchers working on different biological phenomena often unknowingly studied the same gene product, only to have these studies merged as molecular characterization progressed.
The evolutionary conservation of SWI3 underscores its fundamental importance in eukaryotic gene regulation. SWI3 represents one of the most conserved proteins in the SWI/SNF family across all eukaryotic organisms examined to date.[2][21] In yeast, the SWI3 protein is essential for chromatin remodeling function, as demonstrated through both genetic and biochemical studies.[2][12] In plants, which have undergone early duplication events of the SWI3 gene family, Arabidopsis thaliana encodes four distinct SWI3-like proteins designated ATSWI3A, ATSWI3B, ATSWI3C, and ATSWI3D, which show remarkable functional diversification while retaining the characteristic domains present in the yeast ortholog.[2][12] In mammals, two SWI3 homologs exist, designated BAF155 and BAF170 (also known as SMARCC1 and SMARCC2), which are components of both the BAF (Brahma-associated factors) and PBAF (polybromo-associated BAF) complexes representing the mammalian equivalents of the yeast SWI/SNF complex.[34][54] This conservation across hundreds of millions of years of evolution, from unicellular fungi to complex multicellular organisms, indicates that the fundamental role of Swi3 in chromatin remodeling and transcriptional control has been preserved despite significant evolutionary divergence among these organisms.
Structural Organization and Protein Domain Architecture
The SWI3 protein is composed of several evolutionarily conserved domains that mediate its multiple functions within the SWI/SNF complex and in chromatin regulation more broadly. The defining structural features of Swi3 include the SWIRM domain (Swi3/Rsc8/Moira), a SANT domain (Swi3/Ada2/N-CoR/TFIIIB), a leucine zipper region, and various interconnecting sequences whose precise functions remain to be fully elucidated.[2][11][12] The SWIRM domain, encompassing approximately 85 amino acids and localized to residues 311-395 in the yeast Swi3 protein, forms a four-helix bundle containing a pseudo two-fold axis and a helix-turn-helix motif commonly found in DNA-binding proteins.[11][45] This domain was initially thought to directly contact DNA, but structural and biochemical evidence now indicates that the SWIRM domain primarily mediates protein-protein interactions essential for the assembly and functional integrity of chromatin-remodeling complexes, rather than serving a direct DNA-binding function.[45]
The SANT domain, located at residues 527-576 in yeast Swi3, represents one of the most commonly encountered protein-interaction domains in chromatin regulatory proteins and functions in facilitating the binding and modification of histone tails.[2][11][43] The SANT domain is found in numerous chromatin-associated proteins including the Swi3 subunit itself, the Ada2 component of the SAGA histone acetyltransferase complex, the N-CoR corepressor complex, and TFIIIB, indicating its widespread involvement in chromatin regulation.[43][44] The SANT domain possesses predicted DNA-binding activity based on sequence analysis, with regions showing homology to the DNA-binding domain of the myb proto-oncogene, although biochemical evidence for direct DNA binding by the SANT domain remains limited in most contexts.[44][50] The leucine zipper region at the C-terminus of Swi3 functions in mediating dimerization and higher-order protein-protein interactions, allowing Swi3 to form homodimers with itself and heterodimers with other Swi3-related proteins, which is particularly important in organisms with multiple SWI3 gene copies such as plants.[2][12][50]
The spatial arrangement and functional integration of these domains allow Swi3 to serve as a molecular organizer of the SWI/SNF complex. The relatively large size and multi-domain architecture of Swi3 (approximately 56 kD in yeast) suggests that it functions not merely as a passive structural component but as an active regulatory hub integrating signals from multiple sources and coordinating the assembly and function of the complex.[48] In this regard, Swi3 resembles other protein scaffolds and adaptors that organize multi-subunit enzyme complexes or signaling machineries through multiple simultaneous protein-protein interaction interfaces. The modular nature of Swi3, with distinct domains dedicated to specific functions, has allowed for the evolution of specialized SWI/SNF complexes with distinct regulatory properties through the selective combination of different Swi3 variants with different ATPase subunits or regulatory factors in organisms where gene duplication has produced multiple Swi3 isoforms.
Composition and Architecture of the SWI/SNF Chromatin Remodeling Complex
The SWI/SNF complex represents one of the most extensively characterized ATP-dependent chromatin remodeling machines, and understanding Swi3's role requires appreciation of the complex's overall composition and molecular architecture. The minimal functional core of the conserved SWI/SNF complex consists of four essential components: a SWI2/SNF2 ATPase providing the ATP hydrolysis energy, SNF5, SWP73, and a pair of SWI3 subunits.[2][12][21] The presence of a pair of SWI3 subunits in the minimal core complex reflects the importance of Swi3 dimerization for complex stability and function, as demonstrated through biochemical and structural studies.[2][12] The complete yeast SWI/SNF complex, as characterized through biochemical purification and mass spectrometry, contains eleven subunits: Snf2 (the ATPase motor), Swi3, Swi1, Snf5, Swp82, Snf12, Arp7, Arp9, Snf6, Snf11, and Taf14, with the Snf2 subunit serving as the catalytic center that uses ATP hydrolysis energy to drive nucleosome remodeling.[7][20] Individual subunits contribute distinct molecular functions: the Snf2 ATPase domain contains two RecA-like lobes that wrap around DNA, the polybasic SnAc domain couples ATP hydrolysis to nucleosome sliding, and the bromodomain recognizes acetylated histone tails to target the complex to active chromatin.[22]
The structural organization of the SWI/SNF complex has been illuminated through recent cryo-electron microscopy and crosslinking mass spectrometry studies that have revealed the complex possesses a rigid tripartite core containing the motor module with the ATPase, an actin-related protein (ARP) module featuring Arp7 and Arp9, and a recruitment module comprising structural subunits including Swi3.[22][32] Swi3 appears to be embedded within this rigid core, physically interacting with multiple other subunits to form the structural backbone of the complex, while flexible lobes extending from the core provide additional regulatory functions and substrate recognition surfaces.[32] This architecture explains how the complex can maintain structural integrity while simultaneously accommodating conformational changes necessary for nucleosome substrate engagement and remodeling.[22][32] The presence of Swi3 homodimers in the minimal core complex suggests that the two Swi3 molecules may form a structural scaffold within this core, potentially helping to organize the positioning of the catalytic Snf2 ATPase and facilitating proper orientation of the complex with respect to nucleosome substrates.
Molecular Mechanism of SWI/SNF-Dependent Chromatin Remodeling
The fundamental mechanism by which the SWI/SNF complex, operating with Swi3 as a critical regulatory component, remodels chromatin involves using the energy of ATP hydrolysis to alter DNA-histone contacts and reposition nucleosomes along the DNA backbone. The process begins with the binding of the remodeling complex to a nucleosome substrate with nanomolar affinity, an event that involves multiple subunits recognizing different features of the nucleosome structure.[19][22] The motor domain of the Snf2 ATPase binds nucleosomal DNA at approximately two turns from the nucleosome dyad (a location designated SHL2 for Superhelix Location 2), while the polybasic SnAc domain contacts the acidic patch of histone H2A-H2B, and additional subunits including Swi3 may interact with histone H4 tails and other nucleosomal features.[19][22][26]
Upon ATP binding and hydrolysis by the Snf2 ATPase, the complex undergoes conformational changes that translocate DNA around the histone octamer through a mechanism involving the creation of transient DNA loops that propagate around the nucleosome.[19][22] The tracking domain of the ATPase ensures directional movement of these loops in only one direction, preventing backward movement that would be energetically wasteful, while the torsion subdomain executes the actual directional DNA translocation.[19][22] As the DNA loop reaches the exit site on the opposite side of the nucleosome, it resolves, resulting in net nucleosome repositioning along the DNA sequence without complete dissociation of the histone octamer from the DNA.[19][22] This sliding mechanism allows the complex to expose or occlude regulatory DNA sequences by moving nucleosomes relative to fixed genomic features such as transcription factor binding sites.
Critically, Swi3 has been identified as playing a key regulatory role in this remodeling mechanism beyond simple structural scaffolding. Studies examining mutations in the Swi3 C-terminus and specific motifs within Swi3 have revealed that Swi3 controls not only the assembly of the complex but also the efficiency of ATP-dependent histone H2A-H2B dimer displacement, a process related to but distinct from simple nucleosome sliding.[37][48] The mechanism by which Swi3 exerts this regulatory effect likely involves its control over the conformational dynamics of the complex and its ability to sense nucleosome substrate properties and transduce this information to modulate ATPase activity and remodeling outcomes. Furthermore, Swi3 plays a direct role in recruiting the SWI/SNF complex to target genes and chromatin regions through its interactions with transcription factors and histone-modifying enzymes, suggesting that Swi3 functions as a key hub integrating targeting signals with catalytic remodeling activity.
Function in Transcriptional Regulation and Gene Activation
The primary and best-characterized function of Swi3, in the context of the SWI/SNF complex, is the regulation of transcription through modulation of chromatin structure at gene promoters and regulatory regions. The initial characterization of SWI3 mutants identified defects in the transcription of a diverse set of genes, with mutations in SWI/SNF subunit genes causing defects affecting the transcription of approximately five percent of yeast genes, indicating that SWI/SNF functions as a global but selective regulator of transcription.[2][14] The SWI/SNF complex mediates transcriptional activation of target genes through a multi-step process: transcription factor binding to promoter sequences recruits the SWI/SNF complex through direct protein-protein interactions with specific transcription factors; upon recruitment, SWI/SNF remodels nucleosomes at the promoter to expose transcription factor binding sites and core promoter elements; this chromatin remodeling facilitates the binding of additional transcription factors and the assembly of the transcription pre-initiation complex containing RNA polymerase II and general transcription factors such as TFIID, TFIIB, and TFIIE; nucleosome repositioning then permits productive initiation of transcription.[38][41]
The recruitment of SWI/SNF to specific promoters occurs through direct interaction between Swi3 and/or other SWI/SNF subunits with transcription factors acting as activation domains. Studies employing photo-cross-linking label transfer methods have demonstrated that the Snf5, Swi1, and Snf2/Snf2 subunits can directly contact acidic activation domains of transcription factors such as Gcn4 and Hap4, while Swi3 itself was not significantly biotin labeled in parallel experiments, suggesting that Swi3 may not directly contact the activation domains of acidic activators but instead plays a regulatory role in complex assembly and function.[33] However, Swi3 does interact with transcription factors in other contexts, particularly with factors like Rlm1, which is responsible for cell wall stress-responsive gene activation, and this interaction may be mediated through different molecular interfaces than those recognized by acidic activation domains.[41] The recruitment of SWI/SNF to promoters requires an activation domain but does not require promoter sequences, TBP, or RNA polymerase II holoenzyme, indicating that SWI/SNF can be targeted independently of the general transcription machinery and that targeting occurs through direct recognition of DNA-bound transcription factors rather than through promoter sequence recognition.[38]
A particularly well-characterized example of Swi3's function in transcriptional activation comes from studies of the HO gene, which encodes the endonuclease responsible for catalyzing mating-type switching in homothallic yeast strains.[29] Activation of HO requires the coordinated action of Swi5 and Swi4/Swi6 transcription factors, and this transcriptional activation process requires Swi3 as a component of the SWI/SNF complex.[29] The HO promoter is encompassed by positioned nucleosomes when the gene is repressed, including a nucleosome that covers one of the two Swi5 binding sites and the TATA box region, thus occluding access of transcription factors and the general transcription machinery to their binding sites.[29] Cell-cycle regulated acetylation of histone H3 and H4 at the HO promoter occurs in a cell-cycle dependent manner, requiring Swi5 and the SWI/SNF complex, suggesting a sequential model in which histone acetylation facilitates SWI/SNF recruitment and subsequent nucleosome remodeling.[29] The evidence indicates that Swi5-dependent recruitment of the Gcn5 histone acetyltransferase requires chromatin remodeling events by the SWI/SNF complex, with Swi3 playing a key role in coupling these remodeling events to productive gene activation.
Another exemplary function of Swi3-containing SWI/SNF complexes occurs at the PHO5 gene, encoding acid phosphatase, whose expression is induced under conditions of phosphate starvation through the combined action of the Pho2 and Pho4 transcription factors.[59] The PHO5 promoter is repressed in high-phosphate conditions through the presence of positioned nucleosomes covering critical regulatory regions, and upon phosphate starvation, these nucleosomes are disrupted in a Swi3-dependent manner to permit gene activation.[59] Pioneering studies on PHO5 established the principle that nucleosomes physically block access of transcription factors to their binding sites and that nucleosome disruption is required for transcriptional induction, providing fundamental insights into chromatin-mediated gene regulation that have proven applicable across many genes and organisms.[59] The requirement for Swi3 at PHO5 demonstrates that Swi3 function is not limited to a subset of particularly complex gene promoters but is broadly required for transcriptional activation of genes whose promoters are repressed by nucleosomes.
Role in Transcriptional Repression and Negative Regulation
While the predominant characterization of SWI/SNF function has focused on transcriptional activation, recent evidence has revealed that Swi3 and other SWI/SNF subunits also play important roles in transcriptional repression and negative regulation of gene expression. This counterintuitive function, wherein a complex best known for chromatin opening also represses transcription, was illuminated through studies of genes containing tandem or upstream intergenic noncoding RNA-containing (LUTI) promoters, where chromatin remodeling by SWI/SNF at a distal promoter can interfere with transcription from a downstream proximal promoter through a mechanism of transcriptional interference.[9][24] The SWI/SNF complex, when recruited to distal promoters, performs nucleosome remodeling downstream of the active transcription start site for target loci, and this downstream remodeling interferes with coding-sequence-proximal promoters, leading to gene repression for select LUTI-regulated genes.[9][24] This discovery adds a new layer of complexity to understanding Swi3 function, indicating that the same chromatin remodeling activity that facilitates transcription initiation at one promoter can, when occurring at a different genomic location or under different cellular contexts, lead to the repression of nearby genes through transcriptional interference mechanisms.
A classic example of Swi3/SWI/SNF-mediated transcriptional repression occurs at the SER3 gene, which encodes one of the enzymes responsible for serine biosynthesis.[9][24] In serine-rich conditions, the SWI/SNF complex is activated at the upstream SRG1 locus to drive transcription of an upstream intergenic noncoding RNA; this SRG1 transcript then reads through the SER3 promoter in an orientation that occludes the binding of activators required for SER3 expression, thereby repressing SER3 transcription through a transcriptional interference mechanism that is dependent on SWI/SNF function.[9][24] Notably, mutations in Snf2 and Swi3 that reduce but do not eliminate canonical transcription initiation activity by the SWI/SNF complex can still support transcriptional repression at SER3 and LUTI-regulated genes, indicating that transcriptional repression may require nucleosome remodeling through a mechanism distinct from transcriptional activation, possibly involving reduced efficiency of nucleosome displacement or nucleosome repositioning rather than complete nucleosome ejection.
Role in Cellular Responses to Cell Wall Stress
A particularly well-characterized non-transcription function of Swi3 involves its critical role in mediating cellular transcriptional responses to cell wall stress, an area of investigation that has revealed how Swi3 and the SWI/SNF complex integrate environmental stress signals into appropriate chromatin remodeling activities. The yeast cell wall integrity (CWI) pathway includes the mitogen-activated protein kinase (MAPK) Slt2/Mpk1, which upon sensing cell wall perturbations activates the transcription factor Rlm1 through phosphorylation, leading to the induction of genes whose protein products are required for cell wall repair and maintenance.[7][39][41] The SWI/SNF complex is a key element of this CWI MAPK pathway, and Swi3 plays an essential role in this process, as swi3Δ mutants show drastically reduced expression of cell wall stress-responsive genes and hypersensitivity to cell wall-interfering compounds including calcofluor white and zymolyase.[7][39][41]
The mechanism by which Swi3 and the SWI/SNF complex function in cell wall stress responses involves the direct recruitment of the complex to promoters of stress-responsive genes through interaction with the activated Rlm1 transcription factor.[7][39][41] Upon cell wall stress, Rlm1 is phosphorylated and activated by the Slt2 MAPK, and this activated Rlm1 directly interacts with SWI/SNF subunits, including Snf2 and Snf6, recruiting the complex to the promoters of target genes such as MLP1, SRL3, and YLR194C.[7][39][41] The recruitment of SWI/SNF is followed by nucleosome displacement at the promoter regions, which occurs in a manner dependent on Swi3, facilitating the binding of additional transcription factors and RNA polymerase II to these promoters.[7][39][41] Importantly, the kinetics of nucleosome remodeling at stress-responsive genes occur remarkably quickly, with nucleosome displacement at the HNT1 locus occurring within five minutes of cell wall stress induction, suggesting that Swi3 and other SWI/SNF components are rapidly mobilized and targeted to stress-responsive genes in response to environmental perturbations.[24] The requirement for Swi3 is particularly stringent at these loci, as the global expression profile in response to cell wall stress is severely altered in swi3Δ mutants, with approximately 76% of cell wall stress-responsive genes showing Swi3-dependent induction, 19% showing Swi3-dependent basal expression, and only 5% being induced by cell wall stress independent of Swi3.[39][41]
This cell wall stress function extends to multiple targets of the CWI pathway, and the SWI/SNF complex has been identified as essential for generating appropriate transcriptional responses upon a variety of cell wall perturbations, including treatment with calcofluor white (which interferes with chitin biosynthesis) and zymolyase (which digests the cell wall).[7][39][41] The discovery of SWI/SNF as a key element of the CWI MAPK pathway provides a mechanistic link between signal transduction and chromatin remodeling, demonstrating how Swi3 and other SWI/SNF subunits function at the nexus of environmental sensing and gene expression control. This role has implications for understanding how cells integrate multiple types of environmental information and coordinate appropriate adaptive responses.
Oxygen-Dependent Localization and Regulation of Respiration Gene Expression
A more recently discovered and highly specialized function of Swi3 involves its unique role in regulating oxygen consumption and the expression of genes encoding mitochondrial respiratory chain components in an oxygen-dependent manner. This function appears to be somewhat independent of the canonical SWI/SNF complex function, as certain properties distinguish Swi3's role from that of other SWI/SNF subunits.[1][16][37][40][48] Under conditions of hypoxia, Swi3 along with several other SWI/SNF components including Snf5, Snf6, Snf11, Snf12, and Swp82 undergo rapid and dramatic changes in subcellular localization, accumulating in the cytosol rather than maintaining their typical nuclear localization.[1][16][37][40] Upon reoxygenation, these proteins rapidly relocalize back to the nucleus with kinetics that are faster than the transcriptional responses to hypoxia and reoxygenation, suggesting that changes in SWI/SNF protein localization precede and likely contribute to transcriptome responses to oxygen availability.[37]
The mechanism underlying this oxygen-dependent localization appears to involve direct sensing of oxygen availability, possibly through heme-binding proteins or other oxygen-sensing mechanisms that are coupled to the subcellular localization machinery controlling SWI/SNF proteins.[37][40] Notably, the changes in Swi3 localization in response to hypoxia and reoxygenation occur more rapidly than the bulk transcriptome response, with localization changes occurring between 6-12 hours or 1-2 generations, while transcriptome changes are completed after 5-6 generations, indicating that alterations in Swi3 localization represent an early step in the hypoxia response pathway.[37][40] The relocation of Swi3 to the cytosol during hypoxia may functionally alter the composition and properties of the SWI/SNF complex present in the nucleus, as the availability of nuclear Swi3 is reduced and the complex may associate with alternative subunits or exist in alternative conformational states.[37]
In terms of direct effects on respiration, computational analysis of SWI/SNF target genes reveals that Swi3 targets, but not Swi2 targets, are significantly enriched in genes encoding metabolic enzymes involved in oxidation and reduction reactions, while Swi2 targets are preferentially enriched in genes involved in ribosome biogenesis and translation, indicating functional specialization between different SWI/SNF subunits.[48] When the SWI3 gene is deleted, the rate of oxygen consumption is significantly elevated compared to wild-type cells, and the levels of mitochondrial respiratory chain complexes are substantially increased.[1][48] Conversely, Swi3 suppresses oxygen consumption, suggesting that under normal aerobic conditions, Swi3 functions to moderate respiration and mitochondrial activity, potentially allowing cells to adapt their respiratory metabolism to changing oxygen availability and metabolic demands.[1][48] Swi3 strongly affects heme and oxygen-dependent activation of respiration gene promoters, whereas Swi2 affects only basal, heme-independent activities of these promoters, indicating that Swi3 has a unique role in oxygen-dependent transcriptional regulation that is distinct from the broader function of Swi2 and other SWI/SNF components in transcriptional activation.[1][48] In fact, Swi3 has been identified as a dominant regulator in the control of respiratory gene expression, with the effect of swi3 deletion being stronger than that of known respiratory regulators including Hap2/3/4/5, Mot3, and Rox1, classic oxygen-responsive transcription factors.[1][48]
Interactions with Histone Modifications and Recruitment Signals
The recruitment and functional activity of the SWI/SNF complex at specific promoters and regulatory regions depends critically on the recognition of histone modifications and other chromatin-associated signals, processes in which Swi3 plays important regulatory roles. Acetylation of the histone H3 N-terminal tail dramatically facilitates the recruitment and nucleosome mobilization by the SWI/SNF and RSC (Remodel the Structure of Chromatin) complexes, two closely related members of the SWI/SNF family of chromatin remodelers.[26] The mechanism by which H3 acetylation enhances SWI/SNF recruitment involves the bromodomain of the Snf2 ATPase subunit, which recognizes acetylated lysine residues on the H3 tail and brings the complex into closer proximity with nucleosome substrates.[26] Tetra-acetylated H3 tails increase the affinity of SWI/SNF for nucleosomes approximately 7-9 fold while also changing the subunits of SWI/SNF that interact with the H3 tail, such that H3 tail acetylation enhances crosslinking of the Snf2/Snf2 subunit and alters crosslinking patterns of other core subunits including Swi3.[26]
Biochemical analysis of SWI/SNF subunit interactions with acetylated H3 tails has revealed that acetylation of H3 by histone acetyltransferases such as Gcn5 changes the pattern and efficiency of crosslinking between SWI/SNF subunits and the histone H3 tail.[26] Specifically, H3 tail acetylation enhances crosslinking of the Snf2/Snf2 subunit at H3 residues 7 and 22, while the lower amount of Swi3 crosslinking observed at positions 3 and 7 becomes dependent on acetylation, with Snf5 and Snf6 showing reduced crosslinking to the H3 tail at residue 3 upon acetylation.[26] These results collectively demonstrate that acetylation of H3 tails alters the interactions of SWI/SNF with nucleosomes in a way specific to the H3 tail and could be the basis for the increased affinity of SWI/SNF for H3-acetylated nucleosomes observed in biochemical assays.[26] The combined effect of H3 acetylation and transcription activator recruitment is greater than either separately, suggesting that these two modes of SWI/SNF recruitment act in parallel pathways that together have an overall combined effect, indicating that Swi3 and other SWI/SNF components integrate multiple independent targeting signals to achieve specificity and efficiency in nucleosome remodeling at target genes.[26]
Protein-Protein Interactions and Complex Assembly
Swi3 plays a central role in organizing the SWI/SNF complex through its extensive network of protein-protein interactions with other subunits and with non-complex proteins that regulate or recruit the complex. Within the SWI/SNF complex, Swi3 interacts with multiple core subunits including Snf2, Snf5, Snf6, Swp73, and Arp7/Arp9, forming the structural backbone of the complex.[2][12] Swi3 exists as a homodimer in the yeast SWI/SNF complex, and this dimerization is mediated in part through the C-terminal leucine zipper domain, suggesting that the formation of Swi3 homodimers is essential for proper complex assembly and function.[2][12] Outside the complex, Swi3 interacts with various transcription factors and regulatory proteins that recruit the complex to specific genes, including direct interactions with Rlm1 in the context of cell wall stress responses.[41]
The regulatory significance of Swi3-mediated interactions extends to organisms with multiple SWI3 gene copies, where different Swi3 variants can form homodimers or heterodimers with each other, allowing for the assembly of functionally distinct SWI/SNF complexes with different regulatory properties and target gene specificities. In Arabidopsis thaliana, which encodes four SWI3-like proteins, ATSWI3A and ATSWI3B show similar protein interaction properties, both capable of homodimerization and formation of heterodimers with each other, ATSWI3C, and the core SWI/SNF component BSH/SNF5.[2][12][21] However, ATSWI3D exhibits more restricted interaction properties, recognizing only ATSWI3B as a binding partner and failing to interact with BSH, suggesting that ATSWI3D may recruit BSH only in a heterodimeric complex with ATSWI3B.[2][12][21] These differential protein interaction properties among the four Arabidopsis Swi3 homologs suggest that the plant SWI/SNF complexes have undergone considerable functional diversification from the single yeast complex, with different Swi3 variants potentially recognizing distinct transcriptional activators, histone modifications, or other recruiting signals.
Interestingly, ATSWI3B interacts not only with core SWI/SNF components but also with FCA, a putative RNA-binding protein that acts as a positive regulator of flower transition, suggesting that plant SWI/SNF complexes may be specialized for distinct regulatory functions including those related to developmental transitions and the control of flowering time.[2][12][21] This interaction points to a role for SWI/SNF in coordinating chromatin remodeling with RNA-mediated signaling pathways in plants, a concept that may have broader implications for understanding SWI/SNF function in other contexts. The multiplicity of SWI3 gene copies and the differential protein interaction properties of different Swi3 variants provide a molecular basis for the assembly of multiple SWI/SNF complexes with distinct subunit compositions, target gene specificities, and regulatory properties, allowing for a single molecular machine to be adapted for diverse regulatory purposes.
Mating-Type Switching and SWI3 Function
The original identification of SWI3 came from genetic studies of mating-type switching in homothallic strains of S. cerevisiae, where swi3 mutations were shown to cause defects in the process of mating-type switching, a fundamental cellular process in which cells switch between the two alternative mating types, MATa and MATα.[2][13][14][18] Mating-type switching occurs through a site-specific homologous recombination process known as gene conversion, in which an HO endonuclease creates a double-strand break at the MAT locus, and the resulting DNA lesion is repaired through replacement of the MAT allele sequences with a copy of the opposite mating type information harbored at one of two heterochromatic donor loci, HMLα or HMRa.[15] The SWI/SNF complex is required for efficient expression of the HO endonuclease gene, whose transcription is subject to strict cell-cycle regulation such that HO is expressed only during G1 phase of the cell cycle, ensuring that mating-type switching occurs at the appropriate time and that daughter cells in a lineage are equally likely to switch or maintain their original mating type, promoting conjugation between cells of opposite mating types and diploid formation.[15][29][50]
The involvement of Swi3 in HO gene regulation was discussed earlier in the context of Swi5 and Swi4/Swi6-mediated transcriptional regulation, but the broader principle illustrated by this example—that Swi3 and the SWI/SNF complex are required for proper cell-cycle regulation of gene expression—extends to numerous other genes subject to cell-cycle dependent regulation.[29] The cell-cycle dependent pattern of histone acetylation at the HO promoter requires Swi5 and an intact SWI/SNF complex, demonstrating that Swi3 is involved in coupling cell-cycle regulatory signals, transmitted through cell-cycle dependent transcription factors such as Swi5, to chromatin remodeling activities that permit transcription of HO at the appropriate phase of the cell cycle.[29] This role in cell-cycle dependent transcriptional regulation may extend beyond mating-type switching to other cell-cycle regulated genes, suggesting that Swi3 functions broadly in coordinating chromatin remodeling with cell-cycle progression.
Role in DNA Replication and Genome Stability
In addition to its well-characterized roles in transcriptional regulation, Swi3 has been identified as playing important functions in DNA replication and the maintenance of genome stability. In the fission yeast Schizosaccharomyces pombe, a close relative of the budding yeast S. cerevisiae that shares many fundamental cellular processes, Swi1 and Swi3 define a fork protection complex that coordinates leading- and lagging-strand synthesis and stabilizes stalled replication forks.[25] The Swi1-Swi3 complex associates with chromatin specifically during S phase and is required for proficient DNA replication in the absence of agents that cause genotoxic stress, as swi1Δ mutants accumulate foci containing the Rad22 DNA repair protein during S phase, indicating replication abnormalities.[25] Furthermore, Swi3 is required for survival of fork arrest and for the replication checkpoint, as swi3Δ cells display strong sensitivity to hydroxyurea (HU), a drug that blocks DNA synthesis and stalls replication forks, and this HU sensitivity is additive with mutations in checkpoint kinases like Chk1, indicating that Swi3 has a role in tolerating HU-induced replication arrest that is partially independent of checkpoint signaling.[25]
The specific requirement for Swi3 in replication fork stabilization and survival of genotoxic stress appears to involve functions distinct from those of Swi1, as swi1Δ swi3Δ double mutants are more sensitive to HU than swi1Δ single mutants, indicating that Swi3 has additional functions beyond those it shares with Swi1.[25] In addition, the large reduction of Swi3 abundance in swi1Δ cells despite normal Swi1 expression in swi3Δ cells suggests that Swi1 and Swi3 are interdependent for stability, possibly through the formation of a stable protein complex.[25] The extension of SWI/SNF function to DNA replication adds a new dimension to understanding Swi3's biological roles, indicating that the complex is involved in coordinating nucleosome remodeling with multiple DNA-dependent processes beyond transcription, potentially helping to maintain an appropriate balance between chromatin compaction that is necessary for genome protection and chromatin accessibility necessary for DNA replication machinery function.
Evolutionary Expansion and Specialization of SWI3 Genes in Plants and Animals
The characterization of SWI3 homologs in different eukaryotic organisms has revealed a fascinating pattern of evolutionary expansion and functional specialization that illuminates how a single ancestral SWI/SNF subunit has been adapted for diverse regulatory purposes in different cellular and developmental contexts. In mammals, two SWI3 homologs, BAF155 and BAF170, copurify with distinct SWI/SNF complexes, indicating that they are components of the same mammalian SWI/SNF complexes, while two distinct SWI/SNF-like complexes in mammals—BAF (Brahma-associated factors) and PBAF (polybromo-associated BAF)—contain different catalytic ATPase subunits (Brahma or BRG1) combined with the same core structural subunits including BAF155 and BAF170.[34][54] This organization suggests that mammalian cells have evolved distinct SWI/SNF complexes with different ATPase motors but shared structural components, allowing for specialization through the selection of different catalytic subunits rather than exclusively through the selection of different structural subunits.
In plants, particularly in Arabidopsis thaliana, the evolutionary trajectory has been quite different, with the early duplication of the SWI3 gene family resulting in four distinct SWI3-like proteins (ATSWI3A, ATSWI3B, ATSWI3C, and ATSWI3D) that show remarkable functional diversification compared with yeast and animal orthologs.[2][12][14][21][50] The mutations in ATSWI3A and ATSWI3B cause similar blocks of embryo development at the early globular stage, indicating that these two genes have partially redundant essential functions in early plant development.[2][12][14][21][50] In contrast, mutations in ATSWI3C cause more subtle defects including semidwarf stature, inhibition of root elongation, leaf curling, aberrant stamen development, and reduced fertility, suggesting that ATSWI3C has specialized functions in post-embryonic development and reproductive processes.[2][12][14][21][50] Mutations in ATSWI3D result in severe dwarfism, alterations in the number and development of flower organs, and complete male and female sterility, indicating that ATSWI3D is specialized for developmental processes controlling plant size, flower development, and reproductive competence.[2][12][21][50] The root system of homozygous atswi3c seedlings is greatly reduced, because the elongation of primary roots is inhibited and the seedlings develop several side roots and secondary root branches, revealing a specific role for ATSWI3C in regulating root development and branching patterns.[2][12]
These functional specializations among the four Arabidopsis SWI3 homologs far exceed the functional specialization observed in mammals, where the two SWI3 homologs have been maintained as largely interchangeable components of distinct complexes distinguished primarily by their associated ATPase motors. This difference in evolutionary strategies reflects the different developmental complexities of animals versus plants; animals have built specialized developmental processes into the regulatory properties of different genes and their transcriptional control mechanisms, while plants have evolved specialized SWI/SNF complexes containing different SWI3 variants to perform specialized developmental functions. The multiplicity of SWI3 variants in plants, combined with the finding that ATSWI3B interacts with FCA and other regulatory factors, suggests that plants may utilize specialized SWI/SNF complexes for specific developmental programs, such as the control of flowering time through the interaction of ATSWI3B with FCA, an RNA-binding protein involved in the flowering time regulatory network.
Functional and Clinical Implications
Understanding the function of Swi3 and the SWI/SNF complex has profound implications for understanding eukaryotic gene regulation and, by extension, for understanding the basis of diseases in which SWI/SNF function is disrupted. In mammals, mutations affecting core components of SWI/SNF complexes, including the two BAF155 and BAF170 SWI3 homologs, lead to tumorigenesis in somatic tissues of mice and humans, indicating their roles in tumor suppression.[2][12][14][21] This finding has spurred extensive investigation into the mechanisms by which disrupted SWI/SNF function leads to cancer, with evidence suggesting that loss of SWI/SNF function can alter the chromatin landscape in ways that permit the expression of oncogenes or the silencing of tumor suppressor genes, contributing to malignant transformation. The discovery of inactivating mutations in BAF subunits, including the SWI3 homologs, in a substantial fraction of human cancers has made SWI/SNF a major focus of cancer genomics research, with many studies aimed at understanding how specific mutations in individual subunits contribute to cancer development and progression.
Beyond cancer, dysfunction of SWI/SNF complexes has been implicated in neurological and developmental disorders, reflecting the broad importance of SWI/SNF function in controlling gene expression during development and in mature cells. In particular, mutations in BAF subunits have been linked to neurodevelopmental disorders including autism spectrum disorder, intellectual disability, and schizophrenia, suggesting that proper SWI/SNF function is essential for normal brain development and neural gene expression patterns. The characterization of SWI3 function in yeast and plants has provided crucial insights into the molecular mechanisms by which SWI/SNF dysfunction might lead to these diverse pathological conditions, enabling the development of mechanistic models that help explain how specific mutations in SWI3 or other SWI/SNF subunits could disrupt critical gene expression programs.
Conclusion
The SWI3 protein represents a fascinating example of a versatile regulatory factor whose functions span multiple fundamental cellular processes, from transcriptional control to DNA replication and oxygen-dependent gene regulation. As a core structural component of the SWI/SNF chromatin remodeling complex, Swi3 exerts its effects through its capacity to organize and regulate the assembly of this multi-subunit complex, mediate its recruitment to target genes through interactions with transcription factors, and modulate the efficiency and outcomes of ATP-dependent nucleosome remodeling. The discovery of additional specialized functions of Swi3 in oxygen-dependent regulation and its integration into multiple cellular signaling pathways reveal a protein of remarkable functional depth and regulatory sophistication. The evolutionary conservation of Swi3 across all eukaryotes, combined with the functional diversification observed in organisms with multiple SWI3 gene copies, demonstrates that this protein embodies fundamental principles of eukaryotic gene regulation while simultaneously being capable of specialization for particular developmental and environmental contexts. Future research into the mechanisms by which Swi3 integrates multiple regulatory signals, and into the consequences of Swi3 dysfunction in disease states, promises to yield important insights into eukaryotic gene regulation and the basis of human disease.
Citations
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5293592/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1197427/
- https://pubmed.ncbi.nlm.nih.gov/8221926/
- https://www.uniprot.org/uniprotkb/P32591/entry
- https://www.yeastgenome.org/locus/S000003712
- https://thebiogrid.org/33585/summary/saccharomyces-cerevisiae/swi3.html
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3395667/
- https://grassius.org/family/Maize/SWI/SNF-SWI3
- https://pmc.ncbi.nlm.nih.gov/articles/PMC11419397/
- https://www.yeastgenome.org/complex/CPX-1150
- https://pubmed.ncbi.nlm.nih.gov/26104712/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2924208/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1168809/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9839376/
- https://www.yeastgenome.org/locus/S000005816
- https://pmc.ncbi.nlm.nih.gov/articles/PMC516732/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3201869/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2839515/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC316758/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5312515/
- https://elifesciences.org/articles/54449
- https://pmc.ncbi.nlm.nih.gov/articles/PMC135607/
- https://genesdev.cshlp.org/content/10/17/2117
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1456245/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3489556/
- https://genesdev.cshlp.org/content/13/18/2369.long
- https://www.molbiolcell.org/doi/10.1091/mbc.e12-04-0278
- https://www.ncbi.nlm.nih.gov/research/pubtator3/publication/22621902?text=22621902
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1257399/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3645734/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1413740/
- https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2022.1027071/full
- https://www.yeastgenome.org/blog/yeast-researchers-turn-up-the-heat-on-essential-genes
- https://onlinelibrary.wiley.com/doi/abs/10.1002/(SICI)1097-0061(199708)13:10%3C917::AID-YEA148%3E3.0.CO;2-G
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4130347/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC85505/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4399242/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1360160/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5378177/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC232324/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3276623/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4860946/