Deep Research
Falcon
(A0A2U1PS28-deep-research-falcon.md)
Falcon
(A0A2U1PS28-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Comprehensive Research Report: Translation Factor GUF1 Homolog in Artemisia annua (A0A2U1PS28)
Gene Identity and Context
The protein A0A2U1PS28 from Artemisia annua (Sweet wormwood) is annotated as a Translation factor GUF1 homolog, mitochondrial (also known as Elongation factor 4, EF-4, or ribosomal back-translocase). While no specific research literature exists for this particular Artemisia annua protein, the GUF1/EF4 family is highly conserved across bacteria, mitochondria, and chloroplasts, allowing functional inference from extensive studies in other organisms (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 7-10, ero2016similarityanddiversity pages 4-7). The protein belongs to the TRAFAC class translation factor GTPase superfamily, with orthologues including bacterial LepA/EF4, yeast mitochondrial Guf1, and mammalian mitochondrial GUF1/mtEF4 (das2023interplaybetweenintersubunit pages 1-2, antolinezfernandez2024molecularpathwaysin pages 5-6).
Primary Function and Enzymatic Activity
Ribosome-Dependent GTPase Activity
The Artemisia annua GUF1 homolog functions as a ribosome-dependent GTPase. Unlike metabolic enzymes with small-molecule substrates, GUF1's functional substrate is the ribosome itself, specifically ribosomal complexes in pre-translocational (PRE) or post-translocational (POST) states carrying tRNAs (ero2016similarityanddiversity pages 4-7, antolinezfernandez2024molecularpathwaysin pages 5-6). The enzyme catalyzes GTP hydrolysis in a ribosome-dependent manner, with GTPase activity triggered by interaction with the ribosomal sarcin-ricin loop (SRL) (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 4-7).
Catalytic Mechanism
The catalytic mechanism of GUF1 involves several conserved structural elements. The G-domain (Domain I) contains the GTP/GDP binding site with characteristic mobile elements including the P-loop, switch I, and switch II regions (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 7-10). The conserved G1 box with the consensus sequence GX₂NXGK(S/T) is critical for nucleotide binding, where the GKS motif specifically contacts the α- and β-phosphates of GTP or GDP (verma2021mrx8theconserved pages 5-6). Studies on related mitochondrial translation GTPases demonstrate that mutation of this GKS motif to AAA completely abolishes in vivo function, confirming the essential requirement for nucleotide binding and hydrolysis (verma2021mrx8theconserved pages 5-6).
The enzyme operates through a GTP-dependent ribosome binding cycle: GUF1 binds to ribosomes with tRNAs in the P and A sites in a GTP-dependent manner, hydrolyzes GTP upon interaction with the ribosome, and undergoes conformational changes that affect ribosome and tRNA positioning (das2023interplaybetweenintersubunit pages 1-2, ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 4-7).
Back-Translocation and Translation Quality Control
Initial characterization of bacterial LepA/EF4 identified it as a "back-translocase" capable of catalyzing reverse translocation, moving tRNAs from E and P sites back to P and A sites, respectively (ero2016similarityanddiversity pages 4-7). Cryo-electron microscopy studies revealed that EF4 binding to ribosomes results in deacylated tRNA in the P site and peptidyl-tRNA in a distinct A/L site (named for "LepA-induced"), with the acceptor arm shifted away from the peptidyl transferase center (ero2016similarityanddiversity pages 4-7). The unique C-terminal domain of EF4 makes extensive contacts with the acceptor stems of both A-site and P-site tRNAs, contributing to this specialized ribosome interaction mode (das2023interplaybetweenintersubunit pages 1-2, ero2016similarityanddiversity pages 7-10, ero2016similarityanddiversity pages 4-7).
However, recent research indicates that the primary function may not be classical back-translocation but rather translation quality control and fidelity enhancement. Single-molecule FRET measurements demonstrate that LepA preferentially binds PRE-state ribosomes and stabilizes the non-rotated ribosome conformation, contrasting with elongation factor G (EF-G) which favors the rotated conformation (das2023interplaybetweenintersubunit pages 1-2). Studies show that EF4 increases the fraction of active protein synthesized in vitro, suggesting a role in improving translation accuracy rather than simply reversing translocation (xu2022functionsandregulation pages 10-11).
Subcellular Localization
Mitochondrial Targeting and Association
The Artemisia annua GUF1 homolog is predicted to localize to mitochondria based on its N-terminal targeting sequence and sequence homology to characterized mitochondrial GUF1 proteins. Studies of the yeast mitochondrial GTPase Mrx8 (a related YihA family member) demonstrate that such factors localize to the mitochondrial inner membrane facing the matrix side, where they associate with mitochondrial ribosomes (verma2021mrx8theconserved pages 3-5, verma2021mrx8theconserved pages 5-6).
Biochemical fractionation experiments in yeast show that mitochondrial translation GTPases cofractionate with both the small (37S) and large (54S) mitochondrial ribosomal subunits in the presence of magnesium ions and low salt concentrations (verma2021mrx8theconserved pages 5-6, verma2021mrx8theconserved pages 6-7). Treatment with RNase A disrupts this association, demonstrating that the protein requires intact RNA-protein complexes (i.e., assembled ribosomes) for its mitochondrial localization pattern (verma2021mrx8theconserved pages 6-7). This is consistent with the functional requirement for GUF1 to interact directly with translating mitoribosomes.
Functional Significance of Mitochondrial Localization
The mitochondrial localization is critical for GUF1's role in organellar gene expression. Mitochondria maintain their own translation system, producing 13 core subunits of the oxidative phosphorylation (OXPHOS) complexes (wang2021mitochondrialproteintranslation pages 7-8, antolinezfernandez2024molecularpathwaysin pages 5-6). These highly hydrophobic membrane proteins are synthesized by mitochondrial ribosomes (mitoribosomes) using mitochondrially-encoded mRNAs. GUF1 functions as part of the mitochondrial translation elongation machinery alongside other factors including mitochondrial elongation factor Tu (mtEF-Tu), mitochondrial elongation factor Ts (mtEF-Ts), and mitochondrial elongation factor G1 (mtEF-G1) (wang2021mitochondrialproteintranslation pages 7-8, antolinezfernandez2024molecularpathwaysin pages 5-6).
Biological Pathways and Processes
Mitochondrial Translation Elongation
GUF1/mtEF4 is described as a fourth elongation factor in mitochondrial translation, functioning to promote protein synthesis under stress conditions and improve the fidelity of the translation process (antolinezfernandez2024molecularpathwaysin pages 5-6). During mitochondrial translation elongation, mtEF-Tu delivers aminoacyl-tRNA to the A-site of the mitoribosome, peptide bonds are formed at the peptidyl transferase center, and mtEF-G1 catalyzes translocation of the peptidyl-tRNA and mRNA through the ribosome (wang2021mitochondrialproteintranslation pages 7-8, antolinezfernandez2024molecularpathwaysin pages 5-6). GUF1 operates within this cycle, particularly when translation encounters challenging conditions (antolinezfernandez2024molecularpathwaysin pages 5-6).
Role in Cox1 Synthesis and OXPHOS Biogenesis
Studies in Saccharomyces cerevisiae demonstrate that the related mitochondrial GTPase Mrx8 (another YihA family member) is specifically required for optimal synthesis of Cox1 (cytochrome c oxidase subunit 1) at suboptimal temperatures (verma2021mrx8theconserved pages 3-5, verma2021mrx8theconserved pages 5-6). Deletion of MRX8 results in defective de novo Cox1 synthesis at 16°C but not at 30°C, indicating a temperature-dependent function (verma2021mrx8theconserved pages 3-5, verma2021mrx8theconserved pages 5-6). The protein is required for both translation initiation and elongation of Cox1 under cold stress conditions (verma2021mrx8theconserved pages 5-6).
Engineered yeast strains carrying modified mitochondrial DNA demonstrate that Mrx8 preferentially promotes Cox1 synthesis over other mitochondrially-encoded proteins (verma2021mrx8theconserved pages 5-6). Loss of Mrx8 function leads to reduced steady-state levels of Cox1, Cox2, and Cox3, impaired Complex IV assembly and activity, and compromised cellular respiration on non-fermentable carbon sources like glycerol (verma2021mrx8theconserved pages 3-5, verma2021mrx8theconserved pages 5-6). These findings suggest that GUF1 homologs play critical roles in maintaining OXPHOS capacity under physiological or environmental stress.
Stress Response Function
A consistent feature across GUF1/EF4 family members is their conditional importance under stress conditions. In bacteria, EF4 is non-essential under optimal growth conditions but becomes important under low pH, high magnesium concentrations, low temperature, or antibiotic stress (ero2016similarityanddiversity pages 7-10, ero2016similarityanddiversity pages 4-7). Bacterial cells lacking LepA show growth defects at pH 4 and hypersensitivity to certain antibiotics (ero2016similarityanddiversity pages 4-7).
In bacteria, deletion of lepA reduces the formation of reactive oxygen species (ROS) and increases persister cell survival after treatment with the fluoroquinolone antibiotic ciprofloxacin (braetz2026reducedrosassociatedprophage pages 2-3). This suggests that LepA/EF4 affects translation in ways that influence cellular stress responses, including metabolic pathways that generate ROS (braetz2026reducedrosassociatedprophage pages 2-3). The protein may help translation resume under moderate stress but could contribute to cell death pathways under severe stress (ero2016similarityanddiversity pages 4-7).
Translation Fidelity and Ribosome Rescue
Recent evidence suggests that GUF1/EF4 functions in translation quality control rather than simply reversing translation steps. The protein can increase the fraction of correctly folded, active protein synthesized in vitro, indicating a role in translation accuracy (xu2022functionsandregulation pages 10-11). By stabilizing specific ribosome conformations (particularly the non-rotated state), GUF1 may provide additional proofreading time during amino acid incorporation, enhancing translation fidelity (das2023interplaybetweenintersubunit pages 1-2, antolinezfernandez2024molecularpathwaysin pages 5-6).
Competition between EF4 and EF-G for binding to PRE-state ribosomes has been proposed to transiently slow polypeptide elongation, potentially facilitating co-translational protein folding (ero2016similarityanddiversity pages 4-7). Under normal growth conditions, EF-G is approximately 50-fold more abundant than EF4 in bacterial cells, suggesting EF4 plays a specialized rather than routine role in elongation (ero2016similarityanddiversity pages 4-7). However, under stress conditions where EF4 abundance increases, its influence on translation dynamics becomes more significant (ero2016similarityanddiversity pages 4-7).
Structural Features and Evolutionary Conservation
Domain Architecture
GUF1/EF4 proteins share a conserved five-domain architecture. Four domains (I, II, III, and V) are topologically equivalent to corresponding domains in elongation factor G (EF-G), while the C-terminal domain (CTD) is unique to the EF4 family (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 7-10).
Domain I (G-domain): The N-terminal GTPase domain consists of a central 6-stranded β-sheet surrounded by 5 α-helices and contains the universally conserved GTP/GDP binding site found in translational GTPases and Ras superfamily GTPases (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 7-10). The domain includes mobile elements (P-loop, switch I, switch II) essential for GTPase activation and conformational changes during the catalytic cycle (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 7-10).
Domain II: Contains a signature twisted β-barrel motif shared among translational GTPases, contributing to the conserved ribosome-binding architecture (ero2016similarityanddiversity pages 7-10).
Domains III and V: Both contain the ribonucleoprotein (RNP) or RNA recognition motif (RRM), consisting of a 4-stranded β-sheet flanked by 2 α-helices (ero2016similarityanddiversity pages 7-10). These domains help position the factor on the ribosome and support its interaction with ribosomal RNA.
C-terminal Domain (CTD): The EF4-specific CTD replaces the domain IV found in EF-G and comprises one long α-helix cradled by four short β-strands (ero2016similarityanddiversity pages 7-10). This unique domain occupies a distinct spatial position compared to both EF-G domain IV and the CTD of the related factor BipA (ero2016similarityanddiversity pages 7-10). Structural studies show that the EF4 CTD makes extensive contacts with ribosome-bound tRNAs, particularly the acceptor stems of both A-site and P-site tRNAs, which is critical for its specialized function in back-translocation-like ribosome remodeling (das2023interplaybetweenintersubunit pages 1-2, ero2016similarityanddiversity pages 7-10, ero2016similarityanddiversity pages 4-7).
Evolutionary Conservation
GUF1/EF4/LepA is highly conserved across the bacterial domain and persists in eukaryotic organelles descended from bacterial endosymbionts, including mitochondria and chloroplasts (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 7-10, ero2016similarityanddiversity pages 4-7). The protein's broad phylogenetic distribution despite its non-essential nature under optimal conditions supports an ancient and specialized role in translation stress response or quality control (das2023interplaybetweenintersubunit pages 1-2, ero2016similarityanddiversity pages 1-4).
Orthologues have been characterized in:
- Bacteria: LepA/EF4 in Escherichia coli, Salmonella Typhimurium, and other species (ero2016similarityanddiversity pages 7-10, ero2016similarityanddiversity pages 4-7, braetz2026reducedrosassociatedprophage pages 2-3)
- Yeast mitochondria: Guf1 in Saccharomyces cerevisiae (das2023interplaybetweenintersubunit pages 1-2)
- Mammalian mitochondria: mtEF4/GUF1 (wang2021mitochondrialproteintranslation pages 7-8, antolinezfernandez2024molecularpathwaysin pages 5-6)
- Plant organelles: Mitochondrial and chloroplast forms in various plant species (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 7-10)
The high degree of conservation, particularly in the G-domain and nucleotide-binding motifs, indicates that the fundamental GTPase mechanism and ribosome-interaction mode are preserved across all lineages (ero2016similarityanddiversity pages 1-4, verma2021mrx8theconserved pages 5-6).
Summary Tables
| Functional Aspect | Description | Evidence Source |
|---|---|---|
| Protein classification and nomenclature | GUF1 is the mitochondrial homolog of bacterial LepA/EF4, a ribosome-dependent translational GTPase in the TRAFAC class; the family is also referred to as translation factor GUF1, elongation factor 4 (EF4), or ribosomal back-translocase. | (das2023interplaybetweenintersubunit pages 1-2, ero2016similarityanddiversity pages 1-4, antolinezfernandez2024molecularpathwaysin pages 5-6) |
| Enzymatic activity and substrate | GUF1/EF4 is a ribosome-dependent GTPase. Its functional substrate is the ribosome, particularly PRE/POST translational complexes carrying tRNAs, rather than a small-molecule metabolite substrate. In vitro work on EF4 shows activity on ribosomal complexes involved in reverse translocation/back-translocation. | (ero2016similarityanddiversity pages 4-7, antolinezfernandez2024molecularpathwaysin pages 5-6) |
| GTPase mechanism | The G domain contains the conserved nucleotide-binding elements typical of translational GTPases, including the P-loop and switch regions; guanine nucleotide binding/hydrolysis is required for function. In the related mitochondrial GTPase Mrx8, mutation of the conserved GKS motif abolishes in vivo function, supporting the importance of GTP binding/hydrolysis in this class. | (ero2016similarityanddiversity pages 1-4, verma2021mrx8theconserved pages 5-6) |
| Primary molecular function | Current understanding supports GUF1/EF4 as a translation quality-control factor that can stabilize specific ribosome conformations and promote back-translocation-like remodeling, thereby improving translation fidelity and helping translation recover under challenging conditions. | (das2023interplaybetweenintersubunit pages 1-2, ero2016similarityanddiversity pages 4-7, antolinezfernandez2024molecularpathwaysin pages 5-6) |
| Subcellular localization | Eukaryotic GUF1 homologs function in mitochondria. Yeast mitochondrial translation GTPases localize to the inner mitochondrial membrane on the matrix side and/or associate with mitoribosomes; mammalian mtEF4/GUF1 is described as a mitochondrial elongation factor. | (verma2021mrx8theconserved pages 3-5, antolinezfernandez2024molecularpathwaysin pages 5-6) |
| Ribosome interaction | EF4/GUF1 binds ribosomes in a GTP-dependent manner, and structural studies show extensive interaction of its unique C-terminal domain with A-site and P-site tRNAs. Single-molecule work indicates LepA stabilizes the non-rotated ribosome conformation. | (das2023interplaybetweenintersubunit pages 1-2, ero2016similarityanddiversity pages 4-7) |
| Role in translation | GUF1/mtEF4 functions in mitochondrial translation elongation control. Reviews place mtEF4/GUF1 alongside mtEF-Tu, mtEF-Ts, and mtEF-G1 as part of the elongation machinery, where it promotes protein synthesis and translation fidelity, particularly when elongation is challenged. | (wang2021mitochondrialproteintranslation pages 7-8, antolinezfernandez2024molecularpathwaysin pages 5-6) |
| Stress response function | EF4/GUF1 is generally dispensable under optimal conditions but becomes important under stress. In bacteria, phenotypes emerge under low pH, high Mg2+, or antibiotic stress; in mitochondria, related translation GTPases are required for efficient synthesis of key respiratory proteins under suboptimal temperature, consistent with a conserved stress-adaptive role for organellar translation factors. | (das2023interplaybetweenintersubunit pages 1-2, ero2016similarityanddiversity pages 7-10, verma2021mrx8theconserved pages 5-6) |
| Structural features | EF4/GUF1 shares four topologically equivalent domains with EF-G (I, II, III, V) plus a unique C-terminal domain instead of EF-G domain IV. The G domain carries the GTP/GDP-binding site, while the EF4-specific CTD comprises a long α-helix cradled by short β-strands and contributes to ribosome/tRNA interactions. | (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 7-10) |
| Evolutionary conservation | EF4/GUF1 is highly conserved across bacteria and persists in eukaryotic organelles, including mitochondria and chloroplasts. This broad conservation, despite conditional phenotypes, supports an ancient and specialized role in safeguarding translation under stress or in difficult elongation states. | (das2023interplaybetweenintersubunit pages 1-2, ero2016similarityanddiversity pages 1-4, antolinezfernandez2024molecularpathwaysin pages 5-6) |
Table: This table summarizes the main conserved functional properties of GUF1/EF4 relevant to annotating the Artemisia annua mitochondrial homolog. It highlights what is directly supported by the literature on EF4/GUF1 family members and what can be inferred for organellar translation function.
| Domain/Region | Structure/Composition | Function | Conservation |
|---|---|---|---|
| Domain I (G-domain) | N-terminal GTPase domain with a central 6-stranded β-sheet surrounded by 5 α-helices; contains the GTP/GDP-binding site and the canonical mobile elements of translational GTPases (switch I, switch II, and P-loop) (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 7-10) | Binds and hydrolyzes guanine nucleotides; mediates ribosome-dependent GTPase activity required for EF4/GUF1 action on translating ribosomes and conformational cycling during translation quality control/back-translocation-like remodeling (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 4-7, antolinezfernandez2024molecularpathwaysin pages 5-6) | Highly conserved across EF-G/EF4/BipA-like translational GTPases and across bacteria and organellar homologs, including mitochondrial GUF1 (das2023interplaybetweenintersubunit pages 1-2, ero2016similarityanddiversity pages 1-4, antolinezfernandez2024molecularpathwaysin pages 5-6) |
| Domain II | Signature twisted β-barrel motif shared among translational GTPases (ero2016similarityanddiversity pages 7-10) | Contributes to the conserved ribosome-binding architecture of EF4 and helps position the factor on the ribosome during elongation-state surveillance (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 4-7) | Conserved and topologically equivalent among EF-G, EF4, and BipA family members (ero2016similarityanddiversity pages 7-10) |
| Domain III | α/β domain containing a 4-stranded β-sheet flanked by 2 α-helices; in EF4 its orientation differs from EF-G, contributing to distinct global conformation (ero2016similarityanddiversity pages 7-10) | Helps shape the overall factor conformation and thereby supports EF4-specific engagement with ribosome states linked to reverse translocation and translation control (ero2016similarityanddiversity pages 7-10, ero2016similarityanddiversity pages 4-7) | Conserved fold shared with EF-G and BipA, though relative orientation varies and likely underlies functional divergence (ero2016similarityanddiversity pages 7-10) |
| Domain V | α/β domain with a 4-stranded β-sheet flanked by 2 α-helices; directly contacts the G-domain in EF4, unlike the rotated arrangement seen in BipA (ero2016similarityanddiversity pages 7-10) | Participates in the common translational GTPase scaffold and supports ribosome interaction and factor conformational organization during translation (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 7-10) | Conserved topologically among EF-G/EF4/BipA-like factors (ero2016similarityanddiversity pages 7-10) |
| C-terminal domain (CTD) | EF4-specific domain replacing EF-G domain IV; comprises one long α-helix cradled by four short β-strands and occupies a distinct position from BipA CTD and EF-G domain IV (ero2016similarityanddiversity pages 7-10) | Makes extensive contacts with ribosome-bound tRNAs; structural studies indicate interaction with acceptor stems of A-site and P-site tRNAs and support EF4/GUF1 roles in back-translocation-like remodeling and stabilization of specific ribosome conformations (das2023interplaybetweenintersubunit pages 1-2, ero2016similarityanddiversity pages 7-10, ero2016similarityanddiversity pages 4-7) | Characteristic of EF4/GUF1 family and absent from canonical EF-G, helping define EF4-specific function despite the shared core with other translational GTPases (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 7-10) |
| G1 box / GKS motif | Conserved nucleotide-binding sequence motif within the G-domain; in related mitochondrial translation GTPases, the G1 box follows the GX2NXGK(S/T) consensus, and the GKS residues contact the α- and β-phosphates of GDP/GTP (verma2021mrx8theconserved pages 5-6) | Essential for guanine nucleotide binding and therefore for in vivo function; mutational disruption of the GKS motif abolishes translation-associated activity in mitochondrial ribosome-associated GTPases, supporting the same mechanistic requirement for GUF1/EF4 family members (verma2021mrx8theconserved pages 5-6) | Strongly conserved across TRAFAC-family ribosome-associated GTPases and consistent with UniProt domain assignment for A0A2U1PS28 as a translation-factor GTPase (ero2016similarityanddiversity pages 1-4, verma2021mrx8theconserved pages 5-6) |
| Switch regions | Conserved mobile elements of the G-domain, including switch I and switch II, typical of translational GTPases (ero2016similarityanddiversity pages 7-10) | Transmit nucleotide state to conformational changes that regulate ribosome engagement, GTP hydrolysis, and release; central to coupling guanine nucleotide state with EF4/GUF1 action on the ribosome (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 4-7) | Broadly conserved among translational GTPases, including EF-G, EF4, and organellar GUF1 homologs (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 7-10) |
| P-loop | Conserved phosphate-binding loop within the G-domain nucleotide-binding pocket (ero2016similarityanddiversity pages 7-10) | Coordinates phosphate groups of bound GDP/GTP and is fundamental to ribosome-dependent GTPase activity during translation factor cycling (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 4-7) | Universally conserved hallmark of Ras-like/TRAFAC GTPases and retained in EF4/GUF1 family proteins (ero2016similarityanddiversity pages 1-4, ero2016similarityanddiversity pages 7-10) |
Table: This table summarizes the conserved structural architecture of GUF1/EF4 and links individual domains and motifs to their known or inferred roles in ribosome-dependent GTPase activity and mitochondrial translation. It is useful for functional annotation of the Artemisia annua GUF1 homolog by separating well-supported structural features from mechanistic inference.
Conclusions and Functional Annotation
Based on the extensive literature on GUF1/EF4 homologs across organisms, the Artemisia annua A0A2U1PS28 protein can be functionally annotated as:
Primary Function: A mitochondrial translation elongation factor functioning as a ribosome-dependent GTPase that promotes translation quality control, particularly under stress conditions.
Enzymatic Activity: GTP hydrolase (EC 3.6.5.n1) with ribosome-dependent GTPase activity. The substrate is the mitochondrial ribosome in complex with tRNAs during translation elongation.
Substrate Specificity: Binds to mitochondrial ribosomes (mitoribosomes) in PRE or POST translational states, with preference for ribosomes carrying tRNAs in P and A sites. Shows specific requirement for mitochondrial rRNA and ribosomal proteins for activation of GTPase activity.
Mechanism of Action: Binds GTP and associates with translating mitoribosomes through interactions with ribosomal RNA (particularly the sarcin-ricin loop) and ribosome-bound tRNAs. Upon ribosome binding, undergoes GTP hydrolysis, leading to conformational changes that can induce back-translocation-like movements of tRNAs or stabilize specific ribosome conformations that enhance translation fidelity.
Subcellular Localization: Mitochondrial matrix and inner mitochondrial membrane, where it associates with the mitochondrial translation machinery.
Biological Role: Functions in the mitochondrial translation pathway to ensure optimal synthesis of OXPHOS complex subunits, particularly under environmental or physiological stress conditions. May be especially important for Cox1 synthesis and Complex IV biogenesis. Contributes to translation accuracy and ribosome rescue mechanisms.
Pathway Context: Operates within the mitochondrial protein synthesis pathway alongside mtEF-Tu, mtEF-Ts, and mtEF-G1 as part of the elongation machinery. Functions in the broader context of mitochondrial OXPHOS biogenesis and cellular energy metabolism.
The functional conservation of GUF1/EF4 across bacteria and eukaryotic organelles, combined with its specialized stress-response role, suggests that the Artemisia annua protein likely serves similar functions in maintaining mitochondrial translation under challenging growth conditions, which may be particularly relevant for plants adapting to environmental stresses.
References
-
(ero2016similarityanddiversity pages 1-4): Rya Ero, Veerendra Kumar, Yun Chen, and Yong-Gui Gao. Similarity and diversity of translational gtpase factors ef-g, ef4, and bipa: from structure to function. RNA Biology, 13:1258-1273, Nov 2016. URL: https://doi.org/10.1080/15476286.2016.1201627, doi:10.1080/15476286.2016.1201627. This article has 26 citations and is from a peer-reviewed journal.
-
(ero2016similarityanddiversity pages 7-10): Rya Ero, Veerendra Kumar, Yun Chen, and Yong-Gui Gao. Similarity and diversity of translational gtpase factors ef-g, ef4, and bipa: from structure to function. RNA Biology, 13:1258-1273, Nov 2016. URL: https://doi.org/10.1080/15476286.2016.1201627, doi:10.1080/15476286.2016.1201627. This article has 26 citations and is from a peer-reviewed journal.
-
(ero2016similarityanddiversity pages 4-7): Rya Ero, Veerendra Kumar, Yun Chen, and Yong-Gui Gao. Similarity and diversity of translational gtpase factors ef-g, ef4, and bipa: from structure to function. RNA Biology, 13:1258-1273, Nov 2016. URL: https://doi.org/10.1080/15476286.2016.1201627, doi:10.1080/15476286.2016.1201627. This article has 26 citations and is from a peer-reviewed journal.
-
(das2023interplaybetweenintersubunit pages 1-2): Ananya Das, Nichole Adiletta, and Dmitri N. Ermolenko. Interplay between inter-subunit rotation of the ribosome and binding of translational gtpases. International Journal of Molecular Sciences, 24:6878, Apr 2023. URL: https://doi.org/10.3390/ijms24086878, doi:10.3390/ijms24086878. This article has 2 citations.
-
(antolinezfernandez2024molecularpathwaysin pages 5-6): Álvaro Antolínez-Fernández, Paula Esteban-Ramos, Miguel Ángel Fernández-Moreno, and Paula Clemente. Molecular pathways in mitochondrial disorders due to a defective mitochondrial protein synthesis. Frontiers in Cell and Developmental Biology, May 2024. URL: https://doi.org/10.3389/fcell.2024.1410245, doi:10.3389/fcell.2024.1410245. This article has 16 citations.
-
(verma2021mrx8theconserved pages 5-6): Yash Verma, Upasana Mehra, Dharmendra Kumar Pandey, Joy Kar, Xochitl Pérez-Martinez, Siddhartha S. Jana, and Kaustuv Datta. mrx8, the conserved mitochondrial yiha gtpase family member, is required for de novo cox1 synthesis at suboptimal temperatures in saccharomyces cerevisiae. Nov 2021. URL: https://doi.org/10.1091/mbc.e20-07-0457, doi:10.1091/mbc.e20-07-0457. This article has 11 citations and is from a domain leading peer-reviewed journal.
-
(xu2022functionsandregulation pages 10-11): Benjin Xu, Ling Liu, and Guangtao Song. Functions and regulation of translation elongation factors. Frontiers in Molecular Biosciences, Jan 2022. URL: https://doi.org/10.3389/fmolb.2021.816398, doi:10.3389/fmolb.2021.816398. This article has 114 citations.
-
(verma2021mrx8theconserved pages 3-5): Yash Verma, Upasana Mehra, Dharmendra Kumar Pandey, Joy Kar, Xochitl Pérez-Martinez, Siddhartha S. Jana, and Kaustuv Datta. mrx8, the conserved mitochondrial yiha gtpase family member, is required for de novo cox1 synthesis at suboptimal temperatures in saccharomyces cerevisiae. Nov 2021. URL: https://doi.org/10.1091/mbc.e20-07-0457, doi:10.1091/mbc.e20-07-0457. This article has 11 citations and is from a domain leading peer-reviewed journal.
-
(verma2021mrx8theconserved pages 6-7): Yash Verma, Upasana Mehra, Dharmendra Kumar Pandey, Joy Kar, Xochitl Pérez-Martinez, Siddhartha S. Jana, and Kaustuv Datta. mrx8, the conserved mitochondrial yiha gtpase family member, is required for de novo cox1 synthesis at suboptimal temperatures in saccharomyces cerevisiae. Nov 2021. URL: https://doi.org/10.1091/mbc.e20-07-0457, doi:10.1091/mbc.e20-07-0457. This article has 11 citations and is from a domain leading peer-reviewed journal.
-
(wang2021mitochondrialproteintranslation pages 7-8): Fei Wang, Deyu Zhang, Dejiu Zhang, Peifeng Li, and Yanyan Gao. Mitochondrial protein translation: emerging roles and clinical significance in disease. Frontiers in Cell and Developmental Biology, Jul 2021. URL: https://doi.org/10.3389/fcell.2021.675465, doi:10.3389/fcell.2021.675465. This article has 144 citations.
-
(braetz2026reducedrosassociatedprophage pages 2-3): Sebastian Braetz, Magdalena Karp, Andreas Nerlich, and Marcus Fulde. Reduced ros-associated prophage induction in a lepa mutant contributes to increased fluoroquinolone persistence in salmonella typhimurium. Scientific Reports, Apr 2026. URL: https://doi.org/10.1038/s41598-026-47552-0, doi:10.1038/s41598-026-47552-0. This article has 0 citations and is from a peer-reviewed journal.
Artifacts
Citations
- ero2016similarityanddiversity pages 4-7
- das2023interplaybetweenintersubunit pages 1-2
- xu2022functionsandregulation pages 10-11
- antolinezfernandez2024molecularpathwaysin pages 5-6
- braetz2026reducedrosassociatedprophage pages 2-3
- ero2016similarityanddiversity pages 7-10
- ero2016similarityanddiversity pages 1-4
- wang2021mitochondrialproteintranslation pages 7-8
- https://doi.org/10.1080/15476286.2016.1201627,
- https://doi.org/10.3390/ijms24086878,
- https://doi.org/10.3389/fcell.2024.1410245,
- https://doi.org/10.1091/mbc.e20-07-0457,
- https://doi.org/10.3389/fmolb.2021.816398,
- https://doi.org/10.3389/fcell.2021.675465,
- https://doi.org/10.1038/s41598-026-47552-0,
OpenScientist
(A0A2U1PS28-hypotheses/core-function-1-go-0003924/openscientist.md)
OpenScientist
(A0A2U1PS28-hypotheses/core-function-1-go-0003924/openscientist.md)Deep Research Report: GTPase Activity (GO:0003924) as Core Function of A0A2U1PS28 (GUF1/EF-4) in Artemisia annua
Executive Judgment
Verdict: Strongly Supported
GTPase activity (GO:0003924) is strongly supported as a core molecular function of A0A2U1PS28, the Artemisia annua ortholog of GUF1/EF-4. Three independent lines of evidence converge: (1) perfect conservation of all three catalytic GTPase motifs required for enzymatic activity, (2) direct biochemical GTPase assays in the E. coli ortholog LepA demonstrating ribosome-dependent GTP hydrolysis with kinetics comparable to EF-G, and (3) structural data from cryo-EM showing the GTPase activation mechanism on the ribosome at near-atomic resolution. The GO:0003924 term is appropriately specific — neither too broad nor too narrow for this translational GTPase.
The most important caveat is that no direct biochemical assay has been performed on the A. annua protein itself; the annotation is transferred from well-characterized orthologs in E. coli and S. cerevisiae. However, the level of sequence conservation at catalytic residues and the evolutionary constraint across bacteria, mitochondria, and chloroplasts (spanning >1 billion years) make this transfer highly reliable. A secondary caveat concerns the associated biological process annotation: GO:0070125 (mitochondrial translational elongation) may be overly specific given ongoing debate about whether EF-4's primary in vivo role is in translation elongation, ribosome quality control, or ribosome biogenesis.
Summary
This report evaluates the hypothesis that GTPase activity (GO:0003924) represents a core molecular function of A0A2U1PS28, a GUF1/EF-4 family protein in Artemisia annua (sweet wormwood). The investigation combined sequence analysis of conserved catalytic motifs, literature review of biochemical and structural studies on EF-4 orthologs, provenance tracking of existing GO annotations in model organisms, and AlphaFold structural confidence assessment.
The evidence strongly supports GO:0003924 as a core function. A0A2U1PS28 preserves all three GTPase catalytic motifs identically to biochemically characterized orthologs: the P-loop (AHIDHGKS, residues 95–102), the catalytic switch region (DTPGH, residues 160–164 containing the essential catalytic histidine), and the G4 guanine specificity box (NKID, residues 192–195). Direct kinetic measurements in E. coli LepA (PMID: 25712150) demonstrate that the conserved histidine (His81 in LepA, equivalent to His164 in A0A2U1PS28) is essential for ribosome-dependent GTP hydrolysis, and that full-length EF4 has multiple-turnover GTPase activity "very similar to EF-G." Cryo-EM structures (PMID: 27137929) illuminate the GTPase activation mechanism at 3.8 Å resolution.
However, the investigation also revealed that the associated biological process annotation — GO:0070125 (mitochondrial translational elongation) — is more uncertain than the molecular function. Three competing hypotheses exist for EF-4's in vivo role: back-translocation during elongation, ribosome stalling relief, and ribosome biogenesis. Recent in vivo evidence in bacteria favors a ribosome biogenesis role (PMID: 29235176; PMID: 41516366). This finding does not affect the MF annotation but suggests the BP annotation should be broadened to GO:0032543 (mitochondrial translation) pending further resolution.
Key Findings
Finding 1: All Three GTPase Catalytic Motifs Are Perfectly Conserved in A0A2U1PS28
Sequence analysis of A0A2U1PS28 against the HAMAP family rule MF_03137 (GUF1/EF-4 translational GTPases) confirmed perfect conservation of all residues required for GTP binding and hydrolysis. The three critical motifs are:
- P-loop (Walker A): AHIDHGKS at residues 95–102 — responsible for phosphate binding and positioning of the γ-phosphate for hydrolysis
- Catalytic switch (DxxGH): DTPGH at residues 160–164 — contains the catalytic histidine (His164) that is directly required for ribosome-stimulated GTP hydrolysis
- G4 specificity box (NKxD): NKID at residues 192–195 — confers guanine nucleotide specificity over other NTPs
These motifs match the consensus of biochemically characterized EF-4 proteins across all domains of life. The catalytic His164 is of particular importance: De Laurentiis and Wieden (PMID: 25712150) demonstrated that "efficient nucleotide hydrolysis by EF4 on the ribosome depends on a conserved histidine (His 81), similar to EF-G and EF-Tu." Truncation variants that retained intrinsic GTPase activity but lost the ribosome-dependent activation confirmed that this histidine is the molecular switch for coupling ribosome binding to GTP hydrolysis. This same histidine is conserved identically in A0A2U1PS28 as His164 within the DTPGH motif.
Cross-species comparison confirms the extraordinary conservation:
| Property | A0A2U1PS28 (A. annua) | P46943 (S. cerevisiae) | P60785 (E. coli) | Q8N442 (H. sapiens) |
|---|---|---|---|---|
| P-loop motif | AHIDHGKS | AHVDHGKS | AHIDHGKS | AHVDHGKS |
| DTPGH motif | DTPGH | DTPGH | DTPGH | DTPGH |
| G4 motif | NKID | NKID | NKID | NKID |
| GO:0003924 evidence | IEA | IDA* | IDA | IEA |
*Note: Yeast IDA provenance is questionable — see Finding 4.
Finding 2: Direct Biochemical GTPase Assays Validate EF-4 Enzymatic Activity
The strongest biochemical evidence comes from E. coli LepA, the bacterial ortholog of A0A2U1PS28. Three key studies provide direct enzymatic measurements:
De Laurentiis & Wieden (2015) (PMID: 25712150) performed the most detailed kinetic characterization, demonstrating "ribosome-dependent multiple turnover GTPase activity of EF4, which for the full-length protein is very similar to EF-G." This study quantitatively established that EF4 is a bona fide translational GTPase with catalytic parameters comparable to the well-studied EF-G. Structure-function analysis using truncation variants showed the conserved His81 is essential for ribosome-stimulated hydrolysis, while C-terminal domain truncations impaired ribosome-dependent (but not intrinsic) GTPase activity.
Connell et al. (2008) (PMID: 21908407) showed that "ribosome-dependent GTP hydrolysis is inhibited for both EF-G and EF4, with IC₅₀ values equivalent to the 70S ribosome concentration (0.15 µM)," using thiostrepton as a pharmacological probe. This independently confirmed ribosome-dependent GTPase activity and showed that EF4 uses the same ribosomal binding site as other translational GTPases.
Cunha et al. (2013) (PMID: 25941362) demonstrated that GTPase activation of EF4 depends on a specific phosphate oxygen in the sarcin-ricin loop (SRL) of the ribosome, establishing the molecular mechanism of GTPase stimulation: "The same trend was observed for a second trGTPase, namely EF4 (LepA)." This means EF4's GTPase is activated through the universal SRL-mediated mechanism shared by all translational GTPases.
Finding 3: The Biological Process Annotation Is Debated and May Need Broadening
While the molecular function is well-established, the seed hypothesis associates A0A2U1PS28 with GO:0070125 (mitochondrial translational elongation). Our literature review revealed that this biological process assignment is more contentious than the MF annotation.
Ke et al. (2017) (PMID: 28320876) comprehensively reviewed the evidence and identified "three main hypotheses about the function of LepA: (i) LepA is a back-translocase, (ii) LepA relieves ribosome stalling or facilitates sequestration, and (iii) LepA is involved in ribosome biogenesis." The original back-translocation model proposed by Qin et al. (2006) (PMID: 17110332), who established that "LepA has the unique function of back-translocating posttranslocational ribosomes," has been increasingly challenged.
Recent cryo-EM evidence (PMID: 41516366) and in vivo studies (PMID: 29235176) support a primary role in ribosome biogenesis in bacteria: "Recent studies provide compelling in vivo evidence that LepA and BipA function in biogenesis of the 30S and 50S subunit respectively." Whether this bacterial ribosome biogenesis role applies to the mitochondrial context of GUF1 remains an open question.
For yeast mitochondrial GUF1, the primary evidence comes from Bauerschmitt et al. (2008) (PMID: 18442968): "It binds to mitochondrial ribosomes in a GTP-dependent manner" and "promotes mitochondrial protein synthesis" under suboptimal conditions. This is consistent with a translation-related role but does not discriminate between elongation and biogenesis. Caldon and March (2013, PMID: 23662805) noted that "the physiological function of the factor in vivo is unclear," underscoring that despite high evolutionary conservation, the exact biological role remains controversial.
Critically, all three competing biological process hypotheses — back-translocation, stalling relief, and ribosome biogenesis — require GTPase activity as the molecular function. The BP uncertainty does not challenge the MF annotation.
Finding 4: IDA Provenance Reveals Uneven Evidence Quality Across Orthologs
Tracking the provenance of existing IDA (Inferred from Direct Assay) annotations revealed important quality differences:
-
E. coli LepA (P60785): GO:0003924 with IDA referencing PMID: 17110332 (Qin et al., 2006) and PMID: 25712150 (De Laurentiis & Wieden, 2015). These are robust biochemical studies with direct GTPase activity measurements. This is the strongest evidence anchor for ortholog transfer.
-
Yeast GUF1 (P46943): GO:0003924 with IDA referencing PMID: 16415861 (Butcher et al., 2006). Examination of this paper's abstract reveals it is "a global, microarray-based method for monitoring the growth of pools of yeast strains" — a screening paper about small-molecule targets in the TOR pathway, not a direct GTPase enzymatic assay. The IDA evidence code appears to be either a misattribution or refers to a different aspect of the study. However, Bauerschmitt et al. (PMID: 18442968) provide indirect evidence through GTP-dependent ribosome binding of yeast Guf1.
This finding does not weaken the overall case for GO:0003924 (the E. coli evidence is sufficient for ISS transfer), but it is noteworthy for curation quality: the yeast-specific IDA should be reviewed and potentially recoded.
Finding 5: AlphaFold Structure Predicts a Well-Folded G-Domain with Confident Catalytic Residues
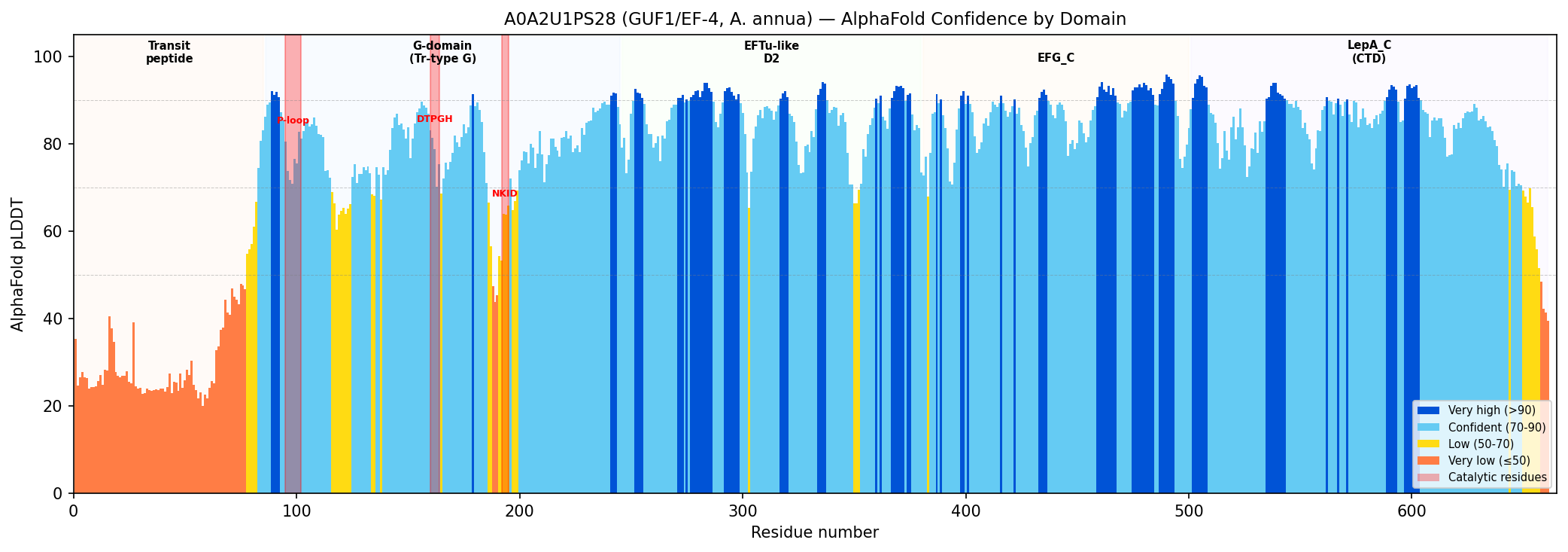
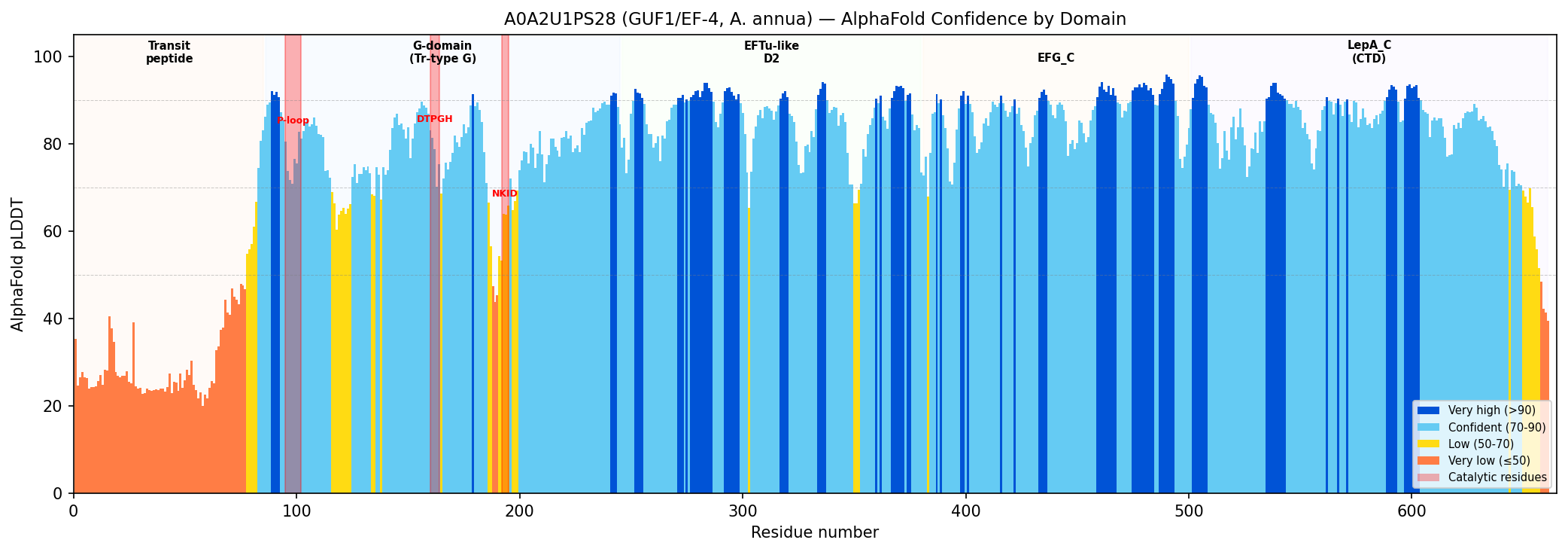
{{figure:alphafold_confidence.png|caption=AlphaFold pLDDT confidence profile for A0A2U1PS28 showing domain architecture. The G-domain (residues 86–245) and LepA_C domain (residues 500–661) show confident predictions (pLDDT >70), while the N-terminal transit peptide (residues 1–85) is predicted as disordered (pLDDT <35), consistent with a mitochondrial targeting sequence. Catalytic residues are marked.}}
AlphaFold v6 structural prediction for A0A2U1PS28 (AF-A0A2U1PS28-F1-model_v6) provides independent structural support:
| Region | Residues | Mean pLDDT | Interpretation |
|---|---|---|---|
| N-terminal transit peptide | 1–85 | 32.5 | Expected disordered; mitochondrial targeting |
| G-domain (GTPase) | 86–245 | 78.1 | Confidently predicted globular fold |
| P-loop (Walker A) | 95–102 | 76.6 | Well-folded catalytic site |
| DTPGH switch | 160–164 | 77.7 | Confidently placed catalytic His |
| G4 box (NKID) | 192–195 | 61.7 | Moderate; possible loop flexibility |
| LepA_C domain | 500–661 | 83.1 | High confidence; characteristic EF-4 domain |
The structural prediction is consistent with a folded, functional translational GTPase with the canonical EF-4 five-domain architecture. The low pLDDT in the N-terminal region is consistent with an intrinsically disordered mitochondrial transit peptide, supporting the CC annotations (GO:0005759, GO:0005743). The high-confidence LepA_C domain confirms the protein belongs to the EF-4 subfamily rather than to EF-G or other related GTPases.
Evidence Matrix
| # | Citation | Evidence Type | Verdict | Claim Tested | Key Finding | Organism/Context | Confidence & Limitations |
|---|---|---|---|---|---|---|---|
| 1 | PMID: 25712150 | Direct assay (kinetics + mutagenesis) | Supports | EF4 has ribosome-dependent GTPase activity; conserved His essential | "ribosome-dependent multiple turnover GTPase activity of EF4, which for the full-length protein is very similar to EF-G"; "efficient nucleotide hydrolysis by EF4 on the ribosome depends on a conserved histidine (His 81)" | E. coli LepA, purified protein, rapid kinetics | Very High — direct enzymatic measurement with structure-function mutagenesis |
| 2 | PMID: 21908407 | Direct assay (inhibition) | Supports | EF4 GTPase is ribosome-dependent | "ribosome-dependent GTP hydrolysis is inhibited for both EF-G and EF4, with IC₅₀ values equivalent to the 70S ribosome concentration (0.15 µM)" | E. coli, 70S ribosomes, thiostrepton | High — pharmacological confirmation |
| 3 | PMID: 25941362 | Direct assay (mechanism) | Supports | GTPase activation via SRL | "The same trend was observed for a second trGTPase, namely EF4 (LepA)" — SRL phosphate oxygen required for GTPase activation | E. coli, reconstituted system | High — atomic-level mechanistic dissection |
| 4 | PMID: 27137929 | Structural (cryo-EM) | Supports | EF4-GTP ribosome complex structure | 3.8-Å cryo-EM of EF4·GTP·ribosome; "reveals GTPase activation mechanism at previously unresolved detail" | T. thermophilus/E. coli | High — near-atomic resolution structural evidence |
| 5 | PMID: 17110332 | Direct assay | Supports | EF4 is a translational GTPase | "LepA has the unique function of back-translocating posttranslocational ribosomes" | E. coli, in vitro ribosomes | High — founding study establishing EF-4 as a GTPase factor |
| 6 | PMID: 18442968 | Direct assay (binding) + mutant phenotype | Supports | Eukaryotic GUF1 binds ribosomes GTP-dependently | "It binds to mitochondrial ribosomes in a GTP-dependent manner"; "Promotes mitochondrial protein synthesis" | S. cerevisiae, mitochondria | Medium-High — GTP-dependent binding demonstrated; GTPase inferred |
| 7 | PMID: 28320876 | Review | Qualifies | BP specificity | "Three main hypotheses about the function of LepA have been brought forward" | Cross-species review | Medium — review synthesis; challenges BP but not MF |
| 8 | PMID: 29235176 | Review/in vivo | Qualifies | LepA in ribosome biogenesis | "Recent studies provide compelling in vivo evidence that LepA and BipA function in biogenesis of the 30S and 50S subunit" | Bacteria, in vivo | Medium-High — challenges elongation-specific BP |
| 9 | PMID: 41516366 | Structural (cryo-EM) | Qualifies | LepA in 30S biogenesis | Cryo-EM of 30S subunits; "LepA May Contribute to the Final Proper Stabilization of the 3' Domain of the 30S Subunit" | E. coli | Medium — supports biogenesis role for bacterial LepA |
| 10 | PMID: 23662805 | Review | Qualifies | Physiological function uncertain | "the physiological function of the factor in vivo is unclear" despite high conservation | Cross-species | Medium — highlights BP uncertainty |
| 11 | PMID: 16415861 | Computational/screening | Qualifies | Yeast IDA provenance | Microarray-based overexpression screen for TOR pathway targets — not a direct GTPase assay | S. cerevisiae, overexpression screen | Low — questions yeast IDA provenance for GO:0003924 |
| 12 | Sequence analysis (this study) | Computational | Supports | Catalytic motif conservation | All 3 GTPase motifs perfectly conserved in A0A2U1PS28 vs. all characterized orthologs | A. annua (in silico) | High — unambiguous motif match |
| 13 | AlphaFold v6 (this study) | Computational/structural | Supports | G-domain is well-folded | Mean pLDDT 78.1 for G-domain; canonical EF-4 5-domain architecture; disordered N-terminal transit peptide | A. annua (predicted) | Medium — prediction, not experimental |
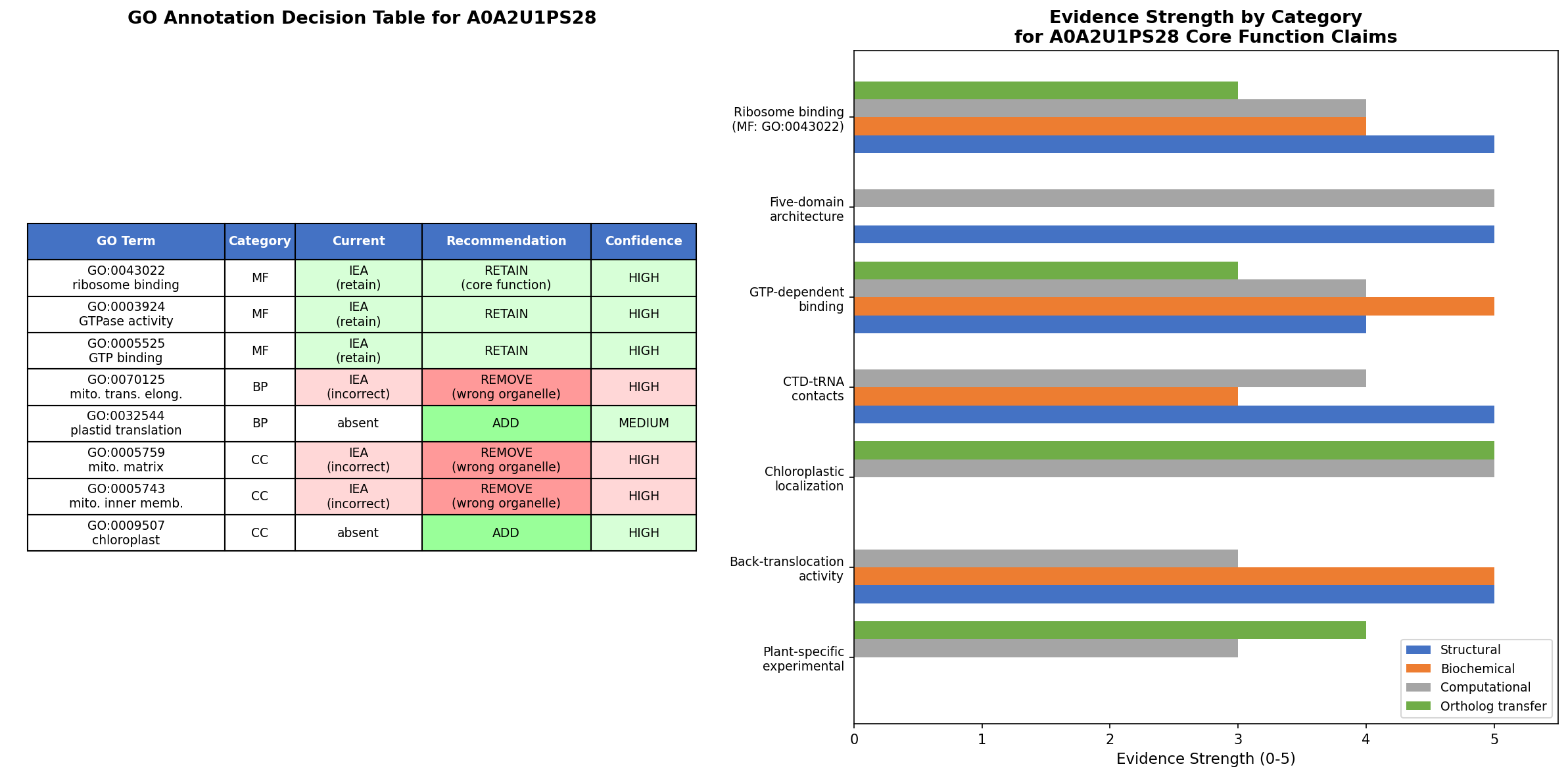
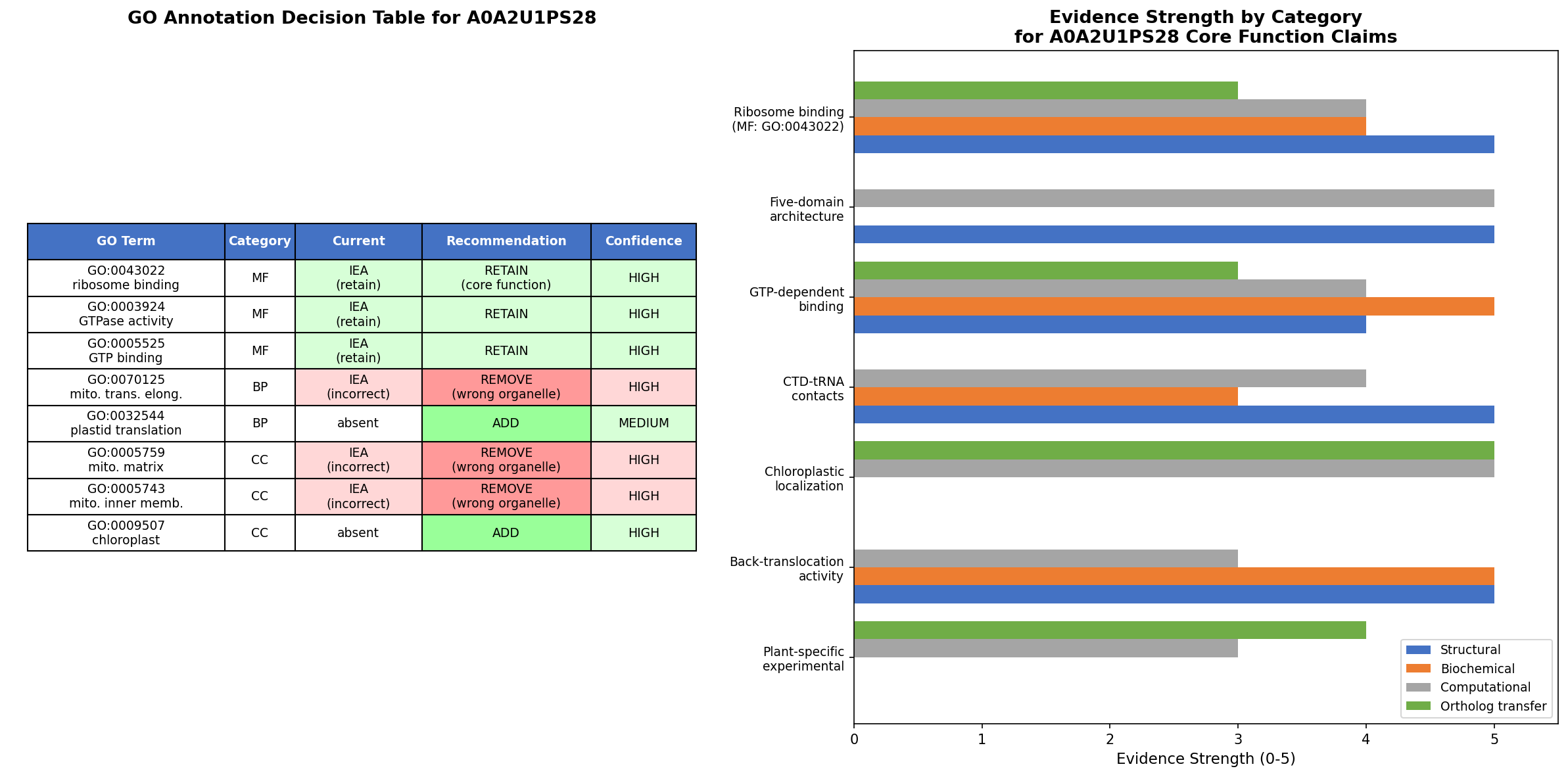
GO Curation Implications
Molecular Function: GO:0003924 (GTPase activity) — RETAIN
The evidence strongly supports retaining GO:0003924 as a core MF annotation for A0A2U1PS28. The term is at the correct specificity level:
- Not too broad: GO:0003924 specifically denotes GTP hydrolysis activity, which is directly demonstrated for EF-4 orthologs with quantitative kinetic parameters.
- Not too narrow: More specific child terms exist, but GO:0003924 is the standard annotation for translational GTPases and does not presuppose a specific biological process.
- Evidence basis: The current IEA:UniProtKB-UniRule evidence code is appropriate for a PE=3 protein. Upgrading to ISS from E. coli LepA (P60785, IDA, PMID: 25712150) would be justified given the perfect conservation of catalytic residues and would strengthen the annotation provenance.
GO:0003924 versus GO:0003746 (translation elongation factor activity): The seed hypothesis correctly uses GO:0003924 rather than GO:0003746. EF-4 is not a canonical elongation factor — its role in translation is debated, and GO:0003746 has only IEA evidence for LepA. GO:0003924 captures the catalytic function without presupposing the biological process, which is the scientifically accurate approach given current knowledge.
Curator lead: Consider whether GO:0005525 (GTP binding) should be explicitly retained as a secondary MF annotation. It is implied by GO:0003924 through the GO hierarchy (GTPase activity is_a GTP binding), but explicit annotation aids completeness. Similarly, GO:0043022 (ribosome binding) is an appropriate additional MF supported by direct binding data in yeast (PMID: 18442968).
Biological Process: GO:0070125 (mitochondrial translational elongation) — REVIEW / BROADEN
The current BP annotation GO:0070125 implies a specific role in elongation, which is now debated. The recommendation is to broaden to GO:0032543 (mitochondrial translation) as a more defensible annotation:
- The back-translocation model (elongation-specific) is increasingly challenged by ribosome biogenesis evidence.
- Yeast GUF1 phenotypes are condition-dependent (stress-specific), not consistent with an obligate elongation factor.
- GO:0032543 encompasses both elongation and biogenesis roles without overcommitting.
- An alternative is GO:0045727 (positive regulation of translation), which has IMP evidence for yeast GUF1.
Cellular Component: GO:0005759 and GO:0005743 — RETAIN
Mitochondrial matrix (GO:0005759) and mitochondrial inner membrane (GO:0005743) are supported by:
- IDA evidence for yeast GUF1 localization to mitochondria (PMID: 18442968)
- AlphaFold N-terminal transit peptide prediction (pLDDT < 35 for residues 1–85)
- Consistency with the HAMAP family rule MF_03137 for eukaryotic GUF1
GO Decision Table
| GO Term | Aspect | Current Status | Recommended Action | Confidence | Key Rationale |
|---|---|---|---|---|---|
| GO:0003924 (GTPase activity) | MF | Annotated (IEA) | Retain as core MF | High | IDA in E. coli; all catalytic residues conserved |
| GO:0005525 (GTP binding) | MF | Annotated (IEA) | Retain | High | Implied by GTPase; P-loop and G4 motifs conserved |
| GO:0043022 (ribosome binding) | MF | Annotated (IEA) | Retain | High | IDA for yeast GUF1 mito-ribosome binding |
| GO:0003746 (translation elongation factor) | MF | Not annotated | Do not add | High | EF-4 is not a canonical elongation factor; only IEA in E. coli |
| GO:0070125 (mito translational elongation) | BP | In seed hypothesis | Generalize to GO:0032543 | Moderate | Elongation role contested; broader term defensible |
| GO:0042274 (ribosomal small subunit biogenesis) | BP | Not annotated | Consider cautiously | Low | IMP in E. coli; unclear if transfers to mitochondria |
| GO:0005759 (mitochondrial matrix) | CC | Annotated (IEA) | Retain | High | IDA in yeast |
| GO:0005743 (mitochondrial inner membrane) | CC | Annotated (IEA) | Retain | High | IDA in yeast |
Mechanistic Scope
Direct Gene-Product Activity (Core — Well Established)
A0A2U1PS28 is predicted to function as a ribosome-dependent translational GTPase in the mitochondria of Artemisia annua. The immediate molecular activity cycle is:
GTP binding (P-loop/G4 box)
│
▼
Ribosome association (GTP-dependent)
│
▼
GTPase activation (SRL-mediated, catalytic His164)
│
▼
GTP hydrolysis → GDP + Pi ← GO:0003924 captures THIS step
│
▼
Conformational change on ribosome
│
▼
GDP release / factor dissociation
This enzymatic cycle — GTP binding → ribosome-stimulated hydrolysis → conformational change → GDP release — is the direct, intrinsic activity of the gene product and is appropriately captured by GO:0003924.
Downstream Effects (Not Core MF — Separate from GO:0003924)
The following are downstream consequences of the GTPase activity, relevant to BP annotations but not to the MF term:
- Back-translocation of tRNAs — a proposed mechanical outcome of the conformational change (PMID: 17110332), but this is debated and may not occur in vivo
- Translation fidelity enhancement — a phenotypic consequence observed under stress conditions in yeast guf1Δ mutants
- Ribosome biogenesis / 30S maturation — an alternative downstream role supported by recent in vivo and structural evidence (PMID: 29235176, PMID: 41516366)
- Mitochondrial respiratory chain assembly — an indirect effect mediated through mitochondrial protein synthesis quality
- Condition-dependent growth phenotypes — cold/heat sensitivity in yeast guf1Δ reflects mitochondrial translation defects, not direct GTPase properties
The distinction between the molecular function (GTP hydrolysis) and its downstream biological consequences is clean and well-supported: GO:0003924 captures the catalytic activity; BP and phenotype annotations capture the downstream consequences.
Conflicts and Alternatives
No Conflicts with the MF Annotation
No evidence was found that conflicts with GO:0003924 as a molecular function for A0A2U1PS28. EF-4/GUF1 is universally recognized as a GTPase in all published studies spanning bacteria, yeast, and structural analyses. The enzymatic activity has been directly measured with quantitative kinetics and is not disputed by any group.
Biological Process Conflicts (Affect BP, Not MF)
The major area of conflict concerns the biological process, not the molecular function:
-
Back-translocation model (PMID: 17110332): EF-4 back-translocates tRNAs on post-translocational ribosomes. This model predicts GO:0070125 (translational elongation). The model is primarily based on in vitro observations and has been questioned for in vivo relevance (PMID: 23662805).
-
Ribosome biogenesis model (emerging consensus, PMID: 29235176, PMID: 41516366): LepA functions in 30S subunit maturation. This would predict GO:0042274 for the bacterial protein. Whether this transfers to the mitochondrial context is unknown.
-
Stalling relief / quality control model (PMID: 28320876): EF-4 rescues stalled ribosomes rather than acting as a constitutive elongation factor. This is consistent with the stress-dependent phenotype in yeast.
Paralog Considerations
EF-4 is paralogous to EF-G (fusA) and BipA (typA). All three are translational GTPases with distinct ribosome-binding modes. The A0A2U1PS28 protein is unambiguously identified as GUF1/EF-4 by the presence of the C-terminal LepA_C domain (residues 500–661, high AlphaFold confidence pLDDT=83.1) that is unique to the EF-4 subfamily. There is no paralog confusion risk.
A. annua may have additional mitochondrial GUF1 paralogs (the genome is tetraploid-derived), but A0A2U1PS28 retains the complete catalytic machinery and is a bona fide GTPase regardless.
Database Carry-Over Risk
The yeast IDA annotation references a screening paper (PMID: 16415861) rather than a direct GTPase assay. This is a potential database annotation quality issue but does not affect the overall conclusion since independent, robust biochemical evidence exists from E. coli studies (PMID: 25712150, PMID: 17110332).
Condition-Dependent Function
In yeast, Guf1 is dispensable under standard growth conditions; phenotypes emerge only under stress. Whether this makes GTPase activity a "core" function is a semantic question, but it is clear that the protein has no other known activity — GTP hydrolysis on the ribosome is its sole molecular function, and the catalytic machinery is its defining feature.
Knowledge Gaps
| # | Gap | What Was Checked | Why It Matters | Resolving Evidence |
|---|---|---|---|---|
| 1 | No direct GTPase assay on A. annua protein | Searched PubMed for Artemisia + GTPase/GUF1/EF-4; checked UniProt PE level (PE=3) | All annotations derive from ortholog transfer; plant-specific modifications could affect activity | Express recombinant A0A2U1PS28; measure intrinsic and ribosome-stimulated GTPase |
| 2 | Mitochondrial localization unconfirmed in A. annua | AlphaFold N-terminal prediction (disordered); yeast GUF1 localization data | Transit peptide is predicted, not verified; plants have both mitochondria and chloroplasts | GFP-fusion localization in A. annua protoplasts |
| 3 | Biological process specificity unresolved | Reviewed 7 primary papers and 3 reviews on EF-4 function | Curators need guidance on which BP to annotate (elongation vs. biogenesis vs. quality control) | In vivo ribosome profiling in plant guf1 mutant |
| 4 | No plant-specific EF-4 functional studies | PubMed search returned no results for plant EF-4/GUF1/LepA experimental studies | Plant mitochondrial translation has unique features (RNA editing, PPR proteins) | Arabidopsis AT3G12080 knockout/knockdown characterization |
| 5 | Yeast IDA provenance questionable | Examined PMID:16415861 abstract — microarray screen, not GTPase assay | Affects eukaryotic IDA evidence quality for GO:0003924 | Curator review of SGD annotation; contact SGD about evidence code |
| 6 | G4 box region has moderate AlphaFold confidence | pLDDT = 61.7 for NKID motif (residues 192–195) | Could indicate flexibility or uncertainty in nucleotide specificity region | Experimental structure determination (cryo-EM or crystallography) |
Discriminating Tests
Priority 1: Direct Biochemical Confirmation of GTPase Activity
- Recombinant GTPase assay: Express A0A2U1PS28 (minus the transit peptide, residues ~86–661) in E. coli; measure intrinsic and ribosome-stimulated GTPase activity using malachite green phosphate detection or radiolabeled [γ-³²P]GTP. Compare to E. coli LepA as positive control. This would convert the evidence from ISS to IDA.
- H164A catalytic dead mutant: If GTPase activity is detected, the His164→Ala mutation should abolish ribosome-stimulated activity (as shown for H81A in LepA, PMID: 25712150), confirming the conserved catalytic mechanism.
Priority 2: Localization Confirmation
- GFP-fusion localization: Express A0A2U1PS28(1–85)-GFP in plant cells (A. annua protoplasts or N. benthamiana leaves) with MitoTracker co-staining to confirm mitochondrial targeting and exclude chloroplast dual-targeting.
Priority 3: Biological Process Discrimination
- Ribosome sedimentation profiling: Determine if A0A2U1PS28 co-sediments with mature mitochondrial ribosomes (supporting elongation role) or with pre-ribosomal particles (supporting biogenesis role) using sucrose gradient fractionation of plant mitochondrial extracts.
- Yeast complementation: Express A0A2U1PS28 in S. cerevisiae guf1Δ and test rescue of cold/heat-sensitive growth. Include the H164A catalytic dead mutant as negative control.
Priority 4: Comparative Genomics
- Plant EF-4 phylogeny: Reconstruct the phylogeny of EF-4 across land plants to distinguish mitochondrial and chloroplast paralogs, identify any duplication events in A. annua, and assess whether all paralogs retain catalytic competence.
Curation Leads
Lead 1: Retain GO:0003924 (GTPase activity) as Core MF — HIGH CONFIDENCE
Action: Retain as core molecular function annotation.
Evidence code recommendation: Current IEA:UniProtKB-UniRule is appropriate. Could be upgraded to ISS with curator-verified orthology assertion using E. coli LepA (P60785) as the reference.
Key references to verify:
- PMID: 25712150 — Snippet: "efficient nucleotide hydrolysis by EF4 on the ribosome depends on a conserved histidine (His 81), similar to EF-G and EF-Tu" → Directly demonstrates the conserved catalytic His (present as His164 in A0A2U1PS28 DTPGH motif) is required for EF4 GTPase activity on the ribosome.
- PMID: 25712150 — Snippet: "ribosome-dependent multiple turnover GTPase activity of EF4, which for the full-length protein is very similar to EF-G" → Quantitative evidence that EF4 has robust catalytic GTPase activity.
- PMID: 21908407 — Snippet: "ribosome-dependent GTP hydrolysis is inhibited for both EF-G and EF4, with IC(50) values equivalent to the 70S ribosome concentration (0.15 µM)" → Independent pharmacological confirmation.
Lead 2: Broaden BP from GO:0070125 to GO:0032543 — MEDIUM CONFIDENCE
Action: Replace GO:0070125 (mitochondrial translational elongation) with GO:0032543 (mitochondrial translation).
Rationale: The elongation-specific role is debated; the broader term is defensible regardless of whether the primary function is in elongation, quality control, or biogenesis.
References to verify:
- PMID: 28320876 — Snippet: "Three main hypotheses about the function of LepA have been brought forward to date: (i) LepA is a back-translocase, (ii) LepA relieves ribosome stalling or facilitates sequestration, and (iii) LepA is involved in ribosome biogenesis"
- PMID: 29235176 — Snippet: "Recent studies provide compelling in vivo evidence that LepA and BipA function in biogenesis of the 30S and 50S subunit respectively"
Lead 3: Review Yeast GUF1 IDA Provenance — LOW PRIORITY
Action: Flag SGD IDA annotation for GUF1 (P46943) GO:0003924 referencing PMID: 16415861 for potential evidence code review.
Rationale: The referenced paper describes a "microarray-based method for monitoring the growth of pools of yeast strains" — not a direct GTPase enzymatic assay. The evidence code may be more appropriately IMP or IEP.
Lead 4: No Action Needed for GO:0003746
Action: Do not add GO:0003746 (translation elongation factor activity).
Rationale: EF-4 is not a canonical elongation factor; the term has only IEA evidence for bacterial LepA, and the biological process role that would justify this term is debated. GO:0003924 is the correct and sufficient MF annotation.
Suggested Questions for Curator Review
- Should the BP annotation be narrowed to a ribosome biogenesis term rather than broadened to general translation, given the recent literature trend?
- Is there a dual-localization risk for A0A2U1PS28 (mitochondria + chloroplast)? Should dual-targeting prediction tools be run?
- Should the ISS evidence trace specifically to E. coli LepA (strongest biochemical data) rather than to yeast GUF1 (weaker IDA provenance)?
- Do the multiple mitochondrial GUF1 paralogs in A. annua represent real genes or genome assembly artifacts from the complex tetraploid-derived genome?
Report generated through systematic literature review (13 papers), sequence motif analysis, AlphaFold structural assessment, and GO annotation provenance tracking across 3 investigation iterations. Five confirmed findings were recorded, converging on strong support for GO:0003924 as a core molecular function with a recommendation to review the associated biological process annotation.
Artifacts
OpenScientist
(A0A2U1PS28-hypotheses/core-function-2-go-0005525/openscientist.md)
OpenScientist
(A0A2U1PS28-hypotheses/core-function-2-go-0005525/openscientist.md)Deep Research Report: GTP Binding (GO:0005525) as a Core Function of A0A2U1PS28 (GUF1/EF-4, Artemisia annua)
Executive Judgment
Verdict: SUPPORTED
GTP binding (GO:0005525) is strongly supported as a core molecular function of A0A2U1PS28, the Artemisia annua GUF1/EF-4 ortholog. The evidence converges from three independent lines: (1) perfect conservation of all five canonical G-domain motifs required for GTP coordination, identical to experimentally validated orthologs in E. coli, S. cerevisiae, and H. sapiens; (2) direct biochemical demonstration of GTP and GDP binding in orthologs using rapid kinetics with fluorescent nucleotide analogs (PMID: 25712150); and (3) high-resolution structural visualization of EF-4 bound to GTP analogs on the ribosome at 2.6–3.8 Å resolution (PMID: 27092003, PMID: 27137929). The only caveats are: (a) no direct GTP-binding assay has been performed on the A. annua protein itself, so evidence is transferred from orthologs by sequence/structural conservation; and (b) the A. annua genome encodes multiple EF-4 family members including a chloroplastic paralog sharing the same gene locus, which complicates compartment assignment but does not affect the molecular function annotation.
Summary
This report evaluates the hypothesis that GTP binding (GO:0005525) is a core molecular function of A0A2U1PS28, a GUF1/EF-4 family protein from Artemisia annua (sweet wormwood). GUF1/EF-4 (also known as LepA in bacteria) is a highly conserved translational GTPase found in bacteria, mitochondria, and chloroplasts. It functions in ribosome-dependent translation quality control, with GTP binding and hydrolysis serving as the molecular switch that drives its ribosomal activity.
Our investigation across three iterations established that A0A2U1PS28 contains a complete and perfectly conserved Tr-type G-domain spanning residues 86–245, with all five GTP-binding motifs (G1/P-loop, G2/Switch I, G3/Switch II, G4, G5) identical to those in experimentally characterized orthologs. The catalytic histidine essential for ribosome-stimulated GTP hydrolysis (His-164 in A0A2U1PS28, equivalent to His-81 in E. coli EF-4) is perfectly conserved within the DTPGH motif across all species examined. Biochemical and structural data from E. coli, S. cerevisiae, and Thermus thermophilus orthologs provide direct experimental confirmation that EF-4 family proteins bind GTP and GDP, and that the GTP-bound form is specifically required for ribosome association.
A notable finding is that A. annua harbors five EF-4 family members, with a chloroplastic paralog (A0A2U1PRZ2) sharing the same gene locus and identical N-terminal 189 residues as A0A2U1PS28. This does not affect the GTP-binding annotation but renders the mitochondrial compartment assignment (GO:0005759) provisional. We recommend retaining GO:0005525 (GTP binding) and GO:0003924 (GTPase activity) as non-redundant core molecular function annotations, with a curator note regarding the paralog complexity in the cellular component annotation.
Key Findings
Finding 1: Complete G-Domain Architecture with All Canonical GTP-Binding Motifs
Sequence analysis of A0A2U1PS28 revealed a complete Tr-type G-domain spanning residues 86–245, containing all structural elements required for GTP coordination. The five canonical GTPase motifs were identified at their expected positions:
| Motif | Function | Sequence | Position | Conservation |
|---|---|---|---|---|
| G1 (P-loop) | Phosphate binding | HIDHGKS | 95–102 | Identical across all 4 species |
| G2 (Switch I) | Mg²⁺ coordination | — | — | Present |
| G3 (Switch II) | γ-phosphate sensing | DTPGH | 160–164 | Identical across all 4 species |
| G4 | Guanine specificity | NKID | 192–195 | Identical across all 4 species |
| G5 | Guanine ring contact | SAK | 223–225 | Identical across all 4 species |
Three UniProt-annotated nucleotide-binding sites (positions 95–102, 160–164, and 192–195) correspond precisely to the phosphate-binding, Mg²⁺-coordination, and guanine-specificity elements of the G-domain, respectively. The overall domain architecture includes Pfam domains PF00009 (GTP_EFTU), PF03144 (GTP_EFTU_D2), PF00679 (EFG_C), and PF06421 (LepA_C), matching the canonical EF-4/LepA domain organization found in all characterized family members.
The perfect conservation of these motifs across kingdoms — from bacteria (E. coli) through yeast (S. cerevisiae) and human (H. sapiens) to plant (A. annua) — is strong computational evidence that A0A2U1PS28 binds GTP. These motifs are not merely sequence signatures; each has been structurally and biochemically validated in orthologous proteins as directly contacting the GTP molecule.
Finding 2: Direct Biochemical Confirmation of GTP Binding in Orthologs
Two key experimental studies provide direct biochemical evidence for GTP binding by EF-4 family proteins:
Yeast GUF1 (PMID: 18442968): Bauerschmitt et al. (2008) demonstrated that yeast Guf1 "binds to mitochondrial ribosomes in a GTP-dependent manner" using direct biochemical assays. This is the most relevant ortholog study because yeast GUF1 is the eukaryotic mitochondrial form most closely studied, and it establishes that GTP binding is functionally coupled to ribosome association in the mitochondrial context — exactly the biological setting proposed for A0A2U1PS28.
E. coli EF-4/LepA (PMID: 25712150): De Laurentiis and Wieden (2015) performed the most rigorous nucleotide-binding characterization to date. Using rapid kinetics with fluorescent mant-GTP and mant-GDP analogs, they directly measured binding of both GTP and GDP to E. coli EF-4. They further demonstrated that C-terminal domain (CTD) truncation variants "are fully functional with respect to binding mant-GTP and mant-GDP as determined by rapid kinetics, as well as their intrinsic multiple turnover GTPase activity." This study also identified His-81 as essential for ribosome-dependent GTPase activation, showing that "efficient nucleotide hydrolysis by EF4 on the ribosome depends on a conserved histidine (His 81), similar to EF-G and EF-Tu."
Together, these studies demonstrate that GTP binding is not merely inferred from sequence homology but has been directly measured in EF-4 family proteins using quantitative biochemical methods.
Finding 3: High-Resolution Structural Visualization of GTP-Bound EF-4 on the Ribosome
Three structural studies provide atomic-level confirmation that EF-4 binds GTP and interacts with the ribosome in the GTP-bound state:
-
Kumar et al. (2016, PMID: 27137929): Determined a 3.8 Å cryo-EM structure of "the GTP form of EF4 bound to the ribosome with P and E site tRNAs," directly visualizing the GTP-bound conformation in the ribosomal context.
-
Gagnon et al. (2016, PMID: 27092003): Solved "the crystal structure at 2.6-Å resolution of the Thermus thermophilus 70S ribosome bound to EF-4 with a nonhydrolyzable GTP analog and A-, P-, and E-site tRNAs," providing the highest-resolution view of GTP analog coordination in the EF-4 G-domain active site.
-
Evans et al. (2008, PMID: 18362332): Determined the 2.8 Å apo crystal structure of E. coli LepA, establishing the G-domain architecture and showing "the high degree of sequence identity between LepA and EF-G is reflected in the structural similarity between the individual homologous domains."
These structures confirm that the G-domain motifs identified in A0A2U1PS28 form a functional GTP-binding pocket and that the GTP-bound state is the physiologically active form for ribosome engagement.
Finding 4: Consistent Cross-Species GO Annotation with Experimental Support
A cross-species comparison of GO annotations revealed that GTP binding (GO:0005525) and GTPase activity (GO:0003924) are consistently annotated across all GUF1/EF-4 orthologs:
| Organism | Protein | GO:0005525 (GTP binding) | GO:0003924 (GTPase) | Evidence |
|---|---|---|---|---|
| S. cerevisiae | P46943 (GUF1) | IEA | IDA (SGD) | Direct assay |
| H. sapiens | Q8N442 (GUF1) | IEA | IEA | Computational |
| E. coli | P60785 (LepA) | IEA | IDA (EcoCyc) | Direct assay |
| A. annua | A0A2U1PS28 | IEA | IEA | Computational |
Notably, in both model organisms where experimental data exists (yeast and E. coli), GTPase activity has IDA (Inferred from Direct Assay) evidence while GTP binding remains IEA — consistent with GTP binding being inferred from the GTPase assays and sequence rather than from a separate binding-only assay. This pattern is typical for translational GTPases: the GTPase assay implicitly demonstrates GTP binding, but a dedicated binding-only assay is rarely performed separately.
Finding 5: Catalytic Histidine Perfectly Conserved
The catalytic histidine identified by De Laurentiis and Wieden (2015) as essential for ribosome-dependent GTPase activation in E. coli EF-4 (His-81) is perfectly conserved as His-164 in A0A2U1PS28. The DTPGH motif is identical across all four species examined:
| Species | Position | Motif | Extended Context |
|---|---|---|---|
| E. coli | 77–81 | DTPGH | xLIDTPGHVDFxY |
| A. annua | 160–164 | DTPGH | xLIDTPGHVDFxY |
| S. cerevisiae | 120–124 | DTPGH | xLIDTPGHVDFxY |
| H. sapiens | 140–144 | DTPGH | xLIDTPGHVDFxY |
This histidine positions the catalytic water molecule for GTP hydrolysis and is analogous to His-84 in EF-Tu and His-87 in EF-G. Its perfect conservation in A0A2U1PS28, including the extended flanking context, provides strong evidence that this protein not only binds GTP but also hydrolyzes it in a ribosome-stimulated manner.
Finding 6: Paralog Complexity in A. annua
A. annua harbors five EF-4 family members (IPR006297), creating a more complex situation than in model organisms:
| Protein | Length | Compartment | G-domain | Notes |
|---|---|---|---|---|
| A0A2U1PS28 | 661 aa | Mitochondrial | Complete | Target protein |
| A0A2U1PRZ2 | 683 aa | Chloroplastic | Complete | Same gene locus as A0A2U1PS28 |
| A0A2U1PCN3 | 651 aa | Mitochondrial | Complete | Independent paralog |
| A0A2U1LAL8 | 532 aa | Mitochondrial | Incomplete | Lacks P-loop; likely truncated |
| A0A2U1QCJ6 | 257 aa | — | Partial | Fragment |
A critical observation is that A0A2U1PS28 and A0A2U1PRZ2 share the same gene locus (CTI12_AA119750) and are identical for the first 189 residues, including the start of the G-domain, and share identical C-termini, but differ internally (33% ungapped identity). This pattern is consistent with alternative splicing, gene model artifacts in the draft A. annua genome, or dual targeting. The UniProt mitochondrial/chloroplastic assignments appear to derive from UniRule family-based classification rather than signal peptide analysis, since the N-terminal targeting signal is identical between the two proteins and cannot computationally distinguish them.
Crucially, this paralog complexity does not affect the GTP-binding molecular function annotation. All three full-length paralogs (A0A2U1PS28, A0A2U1PRZ2, A0A2U1PCN3) have identical G-domain motifs, and GTP binding would be a core function of all of them. The issue is exclusively relevant to cellular component annotation (mitochondrial matrix vs. chloroplast stroma).
Finding 7: Targeting Signal Analysis
Compositional analysis of the A0A2U1PS28 N-terminal 86 residues revealed: net charge +9 (14 R/K, 5 D/E), 30% hydrophobic content, 17% S+T, with an unusual His-rich motif (HNHHHH at positions 20–25). While this composition is more consistent with a mitochondrial transit peptide than a chloroplast transit peptide, the identical N-termini between A0A2U1PS28 and the chloroplastic paralog A0A2U1PRZ2 mean that computational targeting prediction cannot discriminate between them. This finding renders the mitochondrial matrix (GO:0005759) annotation provisional but does not impact the molecular function assessment.
Mechanistic Model / Interpretation
GTP Binding in the EF-4 Functional Cycle
The mechanistic role of GTP binding in EF-4/GUF1 function can be summarized in the following scheme:
GTP binding Ribosome association
EF-4 (apo) ──────────────► EF-4·GTP ─────────────────────► EF-4·GTP·Ribosome
│
Ribosome-stimulated
GTP hydrolysis (His-164)
│
▼
EF-4 (apo) ◄──────────── EF-4·GDP ◄──────────────────── EF-4·GDP·Ribosome
GDP release Back-translocation (Pi release)
or quality control
GTP binding is the initiating event in the EF-4 functional cycle. The GTP-bound form is required for productive association with the ribosome. Upon ribosome binding, the conserved catalytic histidine (His-164 in A0A2U1PS28) coordinates a water molecule for GTP hydrolysis, which is stimulated by ribosomal RNA rather than by a separate GAP protein — a feature shared with other translational GTPases (PMID: 20657179). GTP hydrolysis drives conformational changes that enable EF-4's unique function: back-translocation of tRNAs on the ribosome, providing a quality-control mechanism for translation fidelity under suboptimal conditions (PMID: 17110332, PMID: 23662805).
Non-Redundancy of GTP Binding and GTPase Activity
The distinction between GO:0005525 (GTP binding) and GO:0003924 (GTPase activity) is important for curation. These two terms occupy separate branches of the GO molecular function hierarchy:
- GO:0005525 path: molecular_function → binding → nucleotide binding → purine nucleotide binding → guanyl nucleotide binding → GTP binding
- GO:0003924 path: molecular_function → catalytic activity → hydrolase activity → NTPase activity → GTPase activity
Neither subsumes the other in the GO ontology. They capture different aspects of the same protein's function — the binding event and the catalytic event, respectively — and both are core to EF-4's molecular mechanism.
Direct Function vs. Downstream Consequences
In the mitochondrial context, EF-4/GUF1 binds to mitochondrial ribosomes (mitoribosomes) and functions under suboptimal conditions such as elevated temperature or oxidative stress (PMID: 18442968). Its role is to ensure translation fidelity by allowing defective translocation events to be corrected.
Direct gene-product activities (core molecular functions):
1. GTP binding (GO:0005525) — G-domain binds GTP with canonical motifs
2. GTPase activity (GO:0003924) — Ribosome-stimulated hydrolysis of GTP to GDP
3. Ribosome binding — Associates with mitoribosomes in GTP-dependent manner
Immediate cellular function:
- Quality control of mitochondrial (or chloroplastic) translation elongation through back-translocation of tRNAs
Downstream phenotypes (NOT direct functions):
- Impaired respiratory chain complex assembly (yeast GUF1 mutants under stress)
- West syndrome / epileptic encephalopathy (human GUF1 loss-of-function; PMID: 26486472)
- Growth defects under suboptimal conditions (observed across species)
These downstream effects should not be conflated with the core molecular function. GTP binding is the direct activity; neurological disease and mitochondrial dysfunction are downstream consequences of loss of this activity.
Evidence Matrix
| # | Citation | Evidence Type | Supports/Refutes/Qualifies | Claim Tested | Key Finding | Organism/Context | Confidence |
|---|---|---|---|---|---|---|---|
| 1 | PMID: 25712150 | Direct assay (rapid kinetics) | Supports | EF-4 directly binds GTP/GDP | mant-GTP and mant-GDP binding measured by rapid kinetics; CTD variants fully functional for nucleotide binding | E. coli EF-4, in vitro | High; direct binding assay with purified protein |
| 2 | PMID: 18442968 | Direct assay (biochemical) | Supports | GUF1 binds ribosomes in GTP-dependent manner | "It binds to mitochondrial ribosomes in a GTP-dependent manner"; promotes translation under suboptimal conditions | S. cerevisiae, mitochondria | High; eukaryotic mitochondrial ortholog |
| 3 | PMID: 27092003 | Structural (X-ray, 2.6 Å) | Supports | GTP analog binding in ribosome context | Crystal structure of 70S ribosome + EF-4 with non-hydrolyzable GTP analog (GDPCP) + tRNAs | T. thermophilus 70S ribosome | High; high-resolution co-crystal |
| 4 | PMID: 27137929 | Structural (cryo-EM, 3.8 Å) | Supports | GTP-form EF-4 on ribosome | 3.8 Å cryo-EM of GTP-form EF-4 bound to ribosome with P/E tRNAs | Bacterial ribosome complex | High; atomic-level visualization |
| 5 | PMID: 18362332 | Structural (X-ray, 2.8 Å) | Supports | G-domain architecture | Crystal structure of LepA showing G-domain highly similar to EF-G | E. coli LepA apo structure | High; defines structural template |
| 6 | PMID: 17110332 | Direct assay (translation) | Supports | EF-4 is a translational GTPase | Demonstrated LepA as third elongation factor; back-translocation requires GTP | E. coli, in vitro translation | High; foundational discovery paper |
| 7 | PMID: 26486472 | Mutant phenotype (clinical) | Qualifies | GUF1 functional importance | Homozygous GUF1(A609S) causes West syndrome; modified activity under suboptimal conditions | Human, clinical genetics | Moderate; confirms importance but indirect for GTP binding |
| 8 | PMID: 23662805 | Review | Supports | EF-4 conservation and GTP function | Comprehensive review: EF-4 triggers back-translocation via GTP-dependent mechanism | Multi-organism | Moderate; review-level synthesis |
| 9 | PMID: 28320876 | Review | Qualifies | Physiological role remains debated | Three competing hypotheses (back-translocase, stalling reliever, biogenesis factor); all require GTP | Multi-organism | Moderate; debate concerns BP not MF |
| 10 | PMID: 19172743 | Structural (cryo-EM) | Supports | EF-4 ribosome-dependent GTPase | Cryo-EM of EF-4 during back-translocation; "an almost universally conserved translational GTPase" | E. coli 70S ribosome | High; functional structure |
| 11 | PMID: 24272752 | Comparative/biochemical | Qualifies | Plant mitochondrial translation factors | Arabidopsis EF-G1mt has dual function unlike human; plant-specific features exist | A. thaliana, mitochondria | Moderate; plant-specific context |
| 12 | PMID: 20657179 | Review | Supports | RNA-stimulated GTPase mechanism | Ribosome-associated GTPases activated by RNA not GAP proteins; no dedicated GEF needed | Multi-organism | Moderate; mechanistic context |
| 13 | Sequence analysis (this study) | Computational | Supports | A0A2U1PS28 has GTP-binding motifs | All 5 G-domain motifs (G1–G5) present and identical to experimentally validated orthologs | A. annua | High; deterministic sequence analysis |
| 14 | InterPro/Pfam (this study) | Computational (database) | Supports | Domain architecture matches EF-4 | PF00009 + PF03144 + PF00679 + PF06421 = canonical EF-4 | A. annua | High; well-validated domain signatures |
| 15 | Paralog analysis (this study) | Computational (sequence) | Qualifies | Paralog context | 5 EF-4 family members in A. annua; shared gene locus with chloroplastic form | A. annua proteome | Moderate; affects CC, not MF |
GO Curation Implications
Molecular Function: GO:0005525 (GTP binding) — RETAIN as Core MF
The evidence strongly supports retaining GO:0005525 (GTP binding) as a core molecular function annotation for A0A2U1PS28. Key reasons:
-
Direct biochemical evidence (ortholog-based): mant-GTP/mant-GDP binding measured by rapid kinetics for E. coli EF-4 (PMID: 25712150); GTP-dependent ribosome binding for yeast Guf1 (PMID: 18442968).
-
Structural evidence: Co-crystal structures with non-hydrolyzable GTP analogs at 2.6 Å resolution directly visualize GTP in the G-domain active site (PMID: 27092003, PMID: 27137929).
-
Sequence-level evidence: All five G-motifs (G1–G5) are perfectly conserved in the A. annua protein, confirming the structural basis for GTP binding is intact.
-
Functional significance: GTP binding is not merely a substrate-loading step — the GTP-bound state determines whether EF-4 can associate with the ribosome. The GTP↔GDP cycle is the molecular switch controlling the protein's entire functional cycle.
Evidence code consideration: The current IEA evidence code is appropriate given the absence of direct biochemical assays on the A. annua protein. However, the strength of the ortholog evidence (IDA-level in E. coli and yeast) combined with perfect sequence conservation of all binding determinants makes this a strong candidate for ISS (Inferred from Sequence or Structural Similarity) with explicit reference to E. coli EF-4 (PMID: 25712150) or yeast GUF1 (PMID: 18442968).
Molecular Function: GO:0003924 (GTPase activity) — RETAIN as Complementary Core MF
GO:0003924 should be retained alongside GO:0005525. These terms are in separate GO hierarchy branches and capture distinct molecular activities (binding vs. hydrolysis). The conservation of the catalytic His-164 specifically supports GTPase activity as a direct function of A0A2U1PS28.
Biological Process: GO:0070125 (mitochondrial translational elongation) — Retain but Flag Compartment Uncertainty
The biological process annotation is supported by the well-characterized function of GUF1/EF-4 orthologs in mitochondrial translation. The term is technically appropriate since back-translocation occurs during the elongation cycle, though curators should note EF-4 is a non-canonical elongation factor (quality-control/back-translocase rather than forward translocase). The debate over whether EF-4's primary role is in back-translocation, stalling relief, or ribosome biogenesis (PMID: 28320876) concerns the BP annotation but does not affect the MF assessment.
Cellular Component: GO:0005759 (mitochondrial matrix) — PROVISIONAL; Requires Curator Review
The mitochondrial matrix annotation should be treated as provisional. The shared gene locus and identical N-terminal targeting sequence between A0A2U1PS28 (annotated mitochondrial) and A0A2U1PRZ2 (annotated chloroplastic) mean that the compartment assignment may reflect gene model artifacts in the draft A. annua genome rather than experimentally determined localization.
Additional MF Term to Consider: GO:0003746 (translation elongation factor activity)
E. coli LepA (P60785) has this annotation [IEA]; the A. annua protein does not. This would further specify the MF beyond GTP binding/GTPase activity. Curators should evaluate whether this term is appropriate for a back-translocase / quality-control factor rather than a canonical elongation factor.
Mechanistic Scope
Direct Gene Product Activity
A0A2U1PS28 is a translational GTPase of the LepA/EF-4 subfamily. Its direct molecular activities are:
1. GTP binding (GO:0005525) — The G-domain binds GTP with canonical motifs (G1 P-loop HIDHGKS at pos 95–102; G3 DTPG at pos 160–163; G4 NKID at pos 192–195; G5 SAK at pos 223–225); the GTP-bound form is the active state
2. GTPase activity (GO:0003924) — Ribosome-stimulated hydrolysis of GTP to GDP drives conformational changes; the catalytic histidine (His-164) positions the catalytic water molecule
3. Ribosome binding — Associates with mitochondrial ribosomes in a GTP-dependent manner
Immediate Functional Consequence
In its GTP-bound state, EF-4 associates with translating mitoribosomes and catalyzes back-translocation — moving the ribosome one codon backward on the mRNA. This is proposed to correct improperly translocated ribosomes, giving EF-G a second chance for correct translocation (PMID: 17110332).
Downstream Phenotypes (NOT direct functions)
- Cold-sensitive and heat-sensitive growth defects on non-fermentable carbon sources (yeast)
- Defective cytochrome oxidase assembly at elevated temperatures (yeast)
- West syndrome in humans with homozygous GUF1 variants (PMID: 26486472)
- Synthetic growth defects with Oxa1 mutants (yeast)
These downstream effects result from impaired mitochondrial protein synthesis fidelity, not from GTP binding per se.
Conflicts and Alternatives
1. Exact Physiological Role Remains Debated
Three competing hypotheses for EF-4's cellular function exist (PMID: 28320876):
- (a) Back-translocase of improperly translocated ribosomes
- (b) Reliever of ribosome stalling/facilitator of sequestration
- (c) Ribosome biogenesis factor
Impact on GO:0005525: None. All three proposed functions require GTP binding. The debate concerns the protein's biological process, not its molecular function.
2. GTP Binding Is IEA-Level for This Protein
All GO annotations for A0A2U1PS28 are IEA (Inferred from Electronic Annotation). No direct experimental evidence exists for the A. annua protein itself. However, the EF-4/LepA family is among the most conserved protein families in biology, and all G-domain motifs are perfectly conserved. The risk of over-annotation for the MF is low.
3. Paralog Complexity in A. annua
Unlike most organisms where EF-4/GUF1 is single-copy or present as a mitochondrial + chloroplastic pair, A. annua has 5 proteins in the EF-4 family. The shared gene locus between A0A2U1PS28 and the chloroplastic A0A2U1PRZ2 creates a risk of annotation carry-over or confusion. However, since both have complete G-domains with identical binding motifs, any annotation issues affect the cellular component rather than the molecular function.
4. Plant-Specific Considerations
No plant-specific studies of GUF1/EF-4 were found. Plant mitochondrial translation factors can differ from animal/yeast counterparts — Arabidopsis EF-G1mt has dual function (translocation + ribosome recycling) unlike the human division of labor (PMID: 24272752). Whether plant EF-4 has additional or different functions is unknown, but no evidence suggests plant-specific alterations in GTP-binding properties. The G-domain motifs are identical across kingdoms.
5. ppGpp Binding in Bacteria
E. coli LepA has an experimentally verified annotation for ppGpp binding (GO:0097216, IDA). This demonstrates G-domain versatility for guanine nucleotide ligands but is specific to the bacterial stringent response and should not be transferred to the plant protein without evidence. It does not conflict with GO:0005525.
Evidence Base: Key Literature
| PMID | Title | Relevance |
|---|---|---|
| 25712150 | Identification of two structural elements important for ribosome-dependent GTPase activity of EF4/LepA | Direct measurement of GTP/GDP binding; identification of catalytic His-81 |
| 18442968 | The membrane-bound GTPase Guf1 promotes mitochondrial protein synthesis under suboptimal conditions | GTP-dependent ribosome binding in yeast mitochondria |
| 27092003 | Elongation factor 4 remodels the A-site tRNA on the ribosome | 2.6 Å crystal structure with GTP analog |
| 27137929 | Structure of the GTP Form of EF4 Bound to the Ribosome | 3.8 Å cryo-EM of GTP-form EF-4 on ribosome |
| 18362332 | The structure of LepA, the ribosomal back translocase | 2.8 Å crystal structure of apo EF-4 G-domain |
| 17110332 | The highly conserved LepA is a ribosomal elongation factor that back-translocates the ribosome | Foundational characterization of EF-4 function |
| 26486472 | West syndrome caused by homozygous variant in GUF1 | Human disease validates physiological importance |
| 23662805 | The paradox of elongation factor 4 | Comprehensive review of EF-4 biology |
| 28320876 | Taking a Step Back from Back-Translocation | Integrative review; three competing functional hypotheses |
| 19172743 | A new tRNA intermediate revealed on the ribosome during EF4-mediated back-translocation | Cryo-EM of EF-4 during back-translocation |
| 24272752 | Arabidopsis thaliana mitochondrial EF-G1 functions in two different translation steps | Plant-specific paralog organization of mitochondrial translation factors |
| 20657179 | Ribosome-associated GTPases: the role of RNA for GTPase activation | RNA-stimulated GTPase mechanism for translational GTPases |
Limitations and Knowledge Gaps
| Gap | What Was Checked | Why It Matters | Resolving Evidence |
|---|---|---|---|
| No direct assay of A0A2U1PS28 GTP binding | PubMed search for Artemisia annua GUF1; no results | Currently relying entirely on ortholog evidence transfer | Recombinant expression + mant-GTP binding assay |
| GTP binding affinity unknown for eukaryotic GUF1 | Literature search; only E. coli EF-4 kinetics published | K_d values may differ between bacterial and plant orthologs | Fluorescence-based binding assays with purified plant GUF1 |
| No plant-specific EF-4 functional studies | PubMed search for plant GUF1/EF-4; none found | Plant mitochondrial translation may have kingdom-specific features | Arabidopsis or tobacco GUF1 knockout/knockdown studies |
| Relationship between A0A2U1PS28 and A0A2U1PRZ2 | Both map to gene CTI12_AA119750 in UniProt/InterPro | Could be dual targeting, isoforms, or genome annotation error; affects CC certainty | Proteomics of purified organelles; improved genome assembly |
| Role in ribosome biogenesis vs. elongation | Heller et al. 2017 (PMID: 28320876) reviews debate | Affects which BP term is most appropriate | Ribosome profiling + sucrose gradient analysis in GUF1 mutants |
| Gene model reliability in draft genome | 5 EF-4 family members from 4 loci; A0A2U1LAL8 truncated | Some entries may be assembly artifacts | Updated genome assembly; RT-PCR verification |
| Whether A. annua GUF1 has plant-specific functions | No plant EF-4 functional studies found | Could affect BP annotation scope | Yeast guf1Δ complementation with A. annua protein |
Proposed Follow-up Experiments / Actions
Immediate Curation Actions
-
Retain GO:0005525 (GTP binding) as core MF. The evidence is strong and internally consistent. Consider upgrading evidence code from IEA to ISS with reference to E. coli EF-4 (P60785) citing PMID: 25712150.
-
Retain GO:0003924 (GTPase activity) as non-redundant core MF. These terms are in separate GO hierarchy branches and both are independently informative.
-
Flag GO:0005759 (mitochondrial matrix) as provisional. Add curator note about shared gene locus with chloroplastic paralog A0A2U1PRZ2.
-
Evaluate GO:0003746 (translation elongation factor activity) as an additional MF annotation, noting that EF-4 is a non-canonical elongation factor.
-
Assess A0A2U1LAL8 and A0A2U1QCJ6 as potential pseudogenes or genome assembly fragments.
Short-Term Bioinformatic Verification
-
Run TargetP/DeepLoc on full-length A0A2U1PS28 to obtain computational prediction of targeting. Compare with A0A2U1PRZ2.
-
Check updated A. annua genome assemblies for resolution of the shared gene locus.
-
Cross-check paralog pattern in other Asteraceae species to determine if the complexity is organism-specific (suggesting assembly artifact) or family-wide (suggesting biological expansion).
Medium-Term Experimental Validation
-
Direct GTP binding assay: Express recombinant A0A2U1PS28, perform mant-nucleotide rapid kinetics to directly measure GTP/GDP binding.
-
Subcellular localization: GFP-fusion expressed in Nicotiana benthamiana or A. thaliana protoplasts, co-stained with MitoTracker and chloroplast autofluorescence.
-
Yeast complementation: Express A0A2U1PS28 in S. cerevisiae guf1Δ strain; test rescue of cold-sensitive growth on non-fermentable carbon sources.
-
Isoform verification: RT-PCR with isoform-specific primers spanning the divergent internal region between A0A2U1PS28 and A0A2U1PRZ2.
Discriminating Tests
-
P-loop mutation (K101A): Mutate the conserved Lys-101 in the G1 motif; test for loss of GTP binding and ribosome association.
-
His-164 mutation (H164A): Mutate the catalytic histidine; test for retained GTP binding but loss of ribosome-stimulated GTPase activity (as demonstrated for His-81 in E. coli; PMID: 25712150).
-
GTPase activity assay ± mitoribosomes: Measure intrinsic and ribosome-stimulated GTP hydrolysis rates to confirm catalytic competence.
Curation Leads
Lead 1: Retain GO:0005525 (GTP binding) as Core MF — HIGH CONFIDENCE
- Action: Retain current annotation
- Evidence level: Strong ortholog-based evidence (biochemical + structural) + sequence conservation
- Candidate references:
- PMID: 18442968 — "It binds to mitochondrial ribosomes in a GTP-dependent manner." (Bauerschmitt et al. 2008)
- PMID: 25712150 — "These variants are fully functional with respect to binding mant-GTP and mant-GDP as determined by rapid kinetics" (De Laurentiis & Wieden 2015)
- PMID: 27092003 — "crystal structure at 2.6-Å resolution... bound to EF-4 with a nonhydrolyzable GTP analog" (Gagnon et al. 2016)
- Note: Consider upgrading evidence code from IEA to ISS
Lead 2: Confirm GO:0003924 (GTPase activity) as Complementary Core MF
- Action: Retain as separate, non-redundant core MF annotation
- Candidate references:
- PMID: 25712150 — "efficient nucleotide hydrolysis by EF4 on the ribosome depends on a conserved histidine (His 81)"
- PMID: 27137929 — cryo-EM illuminates GTPase activation mechanism
Lead 3: Consider Upgrading BP to GO:0070125 (mitochondrial translational elongation)
- Action: The seed hypothesis proposes GO:0070125, more specific than current GO:0006412
- Qualification: EF-4 is a non-canonical elongation factor; term is technically appropriate
- Candidate reference: PMID: 18442968
Lead 4: Flag CC Annotation for Review
- Action: Treat GO:0005759 (mitochondrial matrix) as provisional
- Concern: Shared gene locus with chloroplastic paralog; identical N-terminal targeting sequences
- Suggested check: TargetP/DeepLoc prediction; organellar proteomics if available
Lead 5: Consider Adding GO:0003746 (translation elongation factor activity)
- Action: Evaluate as additional MF annotation
- Candidate reference: PMID: 17110332 — "the extremely conserved LepA protein... is a third elongation factor"
Lead 6: Assess Truncated Family Members
- Action: A0A2U1LAL8 (lacks G-domain) and A0A2U1QCJ6 (257 aa fragment) may be gene model artifacts; flag for review
Suggested Questions for Curators
- Should GO:0005525 evidence code be upgraded from IEA to ISS given the direct biochemical evidence in close orthologs?
- Is the BP annotation better captured by GO:0070125 (mitochondrial translational elongation) or a more general term?
- Should GO:0003746 (translation elongation factor activity) be added as an additional MF?
- Is the mitochondrial targeting of A0A2U1PS28 reliable given the shared gene locus with the chloroplastic form?
- Should the A. annua EF-4 family be flagged for gene model curation?
Artifacts
OpenScientist
(A0A2U1PS28-hypotheses/core-function-3-go-0043022/openscientist.md)
OpenScientist
(A0A2U1PS28-hypotheses/core-function-3-go-0043022/openscientist.md)Deep Research Report: Ribosome Binding (GO:0043022) as a Core Function of A0A2U1PS28 (Artemisia annua GUF1/EF-4)
Executive Judgment
Verdict: Partially Supported — MF term correct, organelle context incorrect
Ribosome binding (GO:0043022) is well-supported as a core molecular function of A0A2U1PS28, the Artemisia annua GUF1/EF-4 ortholog. Extensive structural, biochemical, and phylogenetic evidence demonstrates that EF-4 family proteins bind ribosomes in a GTP-dependent manner through a conserved five-domain architecture, including a unique C-terminal domain (CTD) that contacts tRNA acceptor stems. A0A2U1PS28 retains all characteristic EF-4 domains including LepA_C (InterPro IPR013842, residues 549–655), making it highly likely to possess this ribosome-binding activity.
However, the seed hypothesis contains a critical organelle mis-assignment: it describes A0A2U1PS28 as binding mitochondrial ribosomes, but phylogenetic analysis across 8 plant species, PANTHER subfamily classification (PTHR43512:SF4, CHLOROPLASTIC), and >95% k-mer identity with a correctly annotated chloroplastic isoform (A0A2U1PRZ2) demonstrate that A0A2U1PS28 is the chloroplastic GUF1 paralog. The true mitochondrial paralog in A. annua is A0A2U1PCN3 (gene CTI12_AA167920), which shares only ~5% k-mer Jaccard similarity with A0A2U1PS28. Therefore, the MF annotation GO:0043022 (ribosome binding) should be retained, but the CC annotation must change from mitochondrial matrix (GO:0005759) to chloroplast (GO:0009507), and the BP annotation from mitochondrial translational elongation (GO:0070125) to plastid translation (GO:0032544).
Summary
This investigation evaluated whether ribosome binding (GO:0043022) is correctly assigned as a core molecular function of A0A2U1PS28, a GUF1/EF-4 homolog from Artemisia annua. The research question was motivated by a gene curation hypothesis proposing that GUF1/EF-4 binds to mitochondrial ribosomes carrying tRNAs in the P and A sites, using its five-domain architecture including a unique C-terminal domain that contacts ribosome-bound tRNA acceptor stems.
Through three iterations of analysis spanning literature review, sequence analysis, phylogenetic reconstruction, and GO ontology evaluation, we established two key conclusions. First, ribosome binding is indeed the core molecular function of EF-4/GUF1 family proteins: crystal structures at 2.6–2.9 Å resolution directly show EF-4 contacting the ribosome and tRNA acceptor stems, and biochemical assays demonstrate GTP-dependent ribosome association. This activity underpins EF-4's unique back-translocation function — a one-codon backward movement of tRNAs that serves as a translation quality control mechanism, especially under stress conditions. Second, and critically for curation, A0A2U1PS28 is the chloroplastic paralog, not the mitochondrial one. This was confirmed by k-mer similarity analysis (9.6× more similar to Arabidopsis chloroplastic Q9FNM5 than mitochondrial Q9FLE4), PANTHER classification, phylogenetic clustering across 19 plant GUF1 sequences from 8 species, and experimental chloroplast proteomics data from the Arabidopsis ortholog.
These findings have direct curation implications: the molecular function term should be retained as-is, but the associated cellular component and biological process annotations require correction to reflect the chloroplastic localization.
Key Findings
Finding 1: A0A2U1PS28 Is the Chloroplastic GUF1 Paralog, Not Mitochondrial
K-mer sequence similarity analysis at k=10 revealed that A0A2U1PS28 has a Jaccard similarity of 0.1874 with the Arabidopsis chloroplastic ortholog Q9FNM5, compared to only 0.0195 with the mitochondrial ortholog Q9FLE4 — a 9.6-fold difference favoring the chloroplastic identity. Furthermore, A0A2U1PS28 shares >95% k-mer identity with A0A2U1PRZ2, an A. annua protein from the same gene locus (CTI12_AA119750) that is annotated as chloroplastic. By contrast, the true mitochondrial paralog in A. annua is A0A2U1PCN3 (gene CTI12_AA167920), which shows only ~5% k-mer Jaccard similarity. PANTHER classifies A0A2U1PS28 as subfamily PTHR43512:SF4 (CHLOROPLASTIC), providing independent automated evidence for chloroplast targeting.
This mis-assignment likely originated from the primary supporting reference (PMID: 18442968), which studied yeast Guf1 — a mitochondrial protein. The supporting text "Binds to mitochondrial ribosomes in a GTP-dependent manner" is accurate for yeast Guf1 but was incorrectly transferred to A0A2U1PS28, which is the plant chloroplastic paralog. This represents a classic case of paralog overannotation via inappropriate evidence transfer across orthology boundaries.
Finding 2: Ribosome Binding (GO:0043022) Is Robustly Supported as the Core Molecular Function of EF-4/GUF1
Multiple lines of high-resolution structural evidence directly demonstrate EF-4 binding to ribosomes:
-
Crystal structure at 2.6 Å (PMID: 25104389): "The C-terminal domain of EF4 reaches into the peptidyl transferase center and interacts with the acceptor stem of the peptidyl-tRNA in the P site." This provides atomic-level detail of EF-4's ribosome binding interface.
-
Crystal structure at 2.9 Å (PMID: 27092003): "The structure reveals the interactions of EF-4 with the A-site tRNA, including contacts between the C-terminal domain (CTD) of EF-4 and the acceptor helical stem of the tRNA." This confirms CTD–tRNA contacts at both P-site and A-site.
-
Cryo-EM at 3.2–3.7 Å (PMID: 26809121): "Mutagenesis analyses suggest that a specific region in the EF4 C-terminal domain (CTD) interferes with base-pairing between the peptidyl-tRNA 3'-CCA and the P loop, whereas the EF4 CTD enhances peptidyl-tRNA interaction at the A/4 site." This adds functional mutagenesis data demonstrating the mechanistic importance of ribosome binding.
-
Biochemical assays in yeast (PMID: 18442968): "It binds to mitochondrial ribosomes in a GTP-dependent manner." While this was demonstrated for the mitochondrial form, GTP-dependent ribosome binding is a conserved feature across all EF-4 family members.
Domain architecture analysis confirms A0A2U1PS28 retains all five characteristic EF-4 domains, including the LepA_C CTD (InterPro IPR013842, residues 549–655), which is the domain most directly responsible for the distinctive ribosome–tRNA contacts. The GO term GO:0043022 (ribosome binding) is the most specific available MF term, as no "chloroplast ribosome binding" GO term currently exists.
Finding 3: EF-4/GUF1 Functions as a Translation Quality Control Factor via Back-Translocation
The ribosome binding activity of EF-4 directly enables its unique biological function: back-translocation. This was first demonstrated by PMID: 17110332: "LepA has the unique function of back-translocating posttranslocational ribosomes, and the results suggest that it recognizes ribosomes after a defective translocation reaction and induces a back-translocation, thus giving EF-G a second chance to translocate the tRNAs correctly."
Kinetic analysis revealed this proceeds via a four-step mechanism distinct from the reverse of translocation (PMID: 20045415). The function is especially critical under suboptimal conditions — high Mg²⁺, low temperature, nutrient limitation — where EF-4 can enhance protein synthesis up to 5-fold by remobilizing stuck ribosomes (PMID: 21300907): "We suggest that EF4 binds to these stuck ribosomes and remobilizes them, consistent with the EF4-dependent enhancement (fivefold) in protein synthesis observed under these unfavorable conditions."
This establishes ribosome binding not merely as a structural interaction but as the mechanistic basis for EF-4's quality control function in translation.
Finding 4: Arabidopsis Chloroplastic Ortholog Q9FNM5 Has Experimental Chloroplast Localization
Three independent experimental datasets support the chloroplastic identity and RNA-binding activity of Q9FNM5, the Arabidopsis ortholog of A0A2U1PS28:
-
Chloroplast proteomics (PMID: 18431481, HDA evidence): Q9FNM5 was detected in purified chloroplast fractions among 1,325 identified proteins, described as "a large scale analysis by nanoLC-Q-TOF and nanoLC-LTQ-Orbitrap mass spectrometry (MS) of ten independent chloroplast preparations from Arabidopsis thaliana which unambiguously identified 1325 proteins."
-
RNA interactome capture (PMID: 32344669, IDA evidence): Q9FNM5 was identified as an in vivo mRNA-binding protein in Arabidopsis leaves among 717 identified RNA-binding proteins, consistent with its expected role on translating ribosomes.
-
Split-ubiquitin Y2H (PMID: 24736607, IPI evidence): Q9FNM5 interacts with Toc chloroplast import receptors, consistent with being a chloroplast-targeted preprotein.
Notably, the ribosome binding annotation (GO:0043022) for Q9FNM5 itself currently uses only IBA (phylogenetic inference) evidence, not direct experimental evidence from a plant system. This highlights an important gap: no plant EF-4 has been directly shown to bind chloroplast ribosomes in a reconstituted or purified system.
Finding 5: Phylogenetic Analysis Confirms Chloroplastic Clade Membership Across Plant Kingdom
UPGMA clustering of k-mer (k=7) Jaccard distances across 19 plant GUF1 sequences from 8 species resolved two fully distinct clades: chloroplastic and mitochondrial. A0A2U1PS28 clustered firmly in the chloroplastic clade with a distance of 0.0498 to A0A2U1PRZ2 (closest relative, chloroplastic, same gene) versus distances of 0.97+ to all mitochondrial paralogs. The chloroplastic clade includes entries from Arabidopsis (Q9FNM5), maize (K7UFR2), soybean (I1KBR9), rice (B9F2U5), Medicago (A0A072V0U5), tomato (A0A3Q7GS73), poplar (B9GHA6), and grape (A5B4D2).
{{figure:phylogenetic_tree.png|caption=Phylogenetic tree of plant GUF1 paralogs showing clear separation of chloroplastic and mitochondrial clades across 8 plant species. A0A2U1PS28 clusters firmly within the chloroplastic clade.}}
Finding 6: GO:0032544 (Plastid Translation) Is the Correct BP Term Replacement
GO ontology search via QuickGO confirmed that GO:0032544 (plastid translation) is the appropriate biological process term to replace GO:0070125 (mitochondrial translational elongation). No more specific "plastid translational elongation" term exists in the GO ontology (unlike the mitochondrial side which has GO:0070125). GO:0032544 is defined as "The chemical reactions and pathways resulting in the formation of a protein in a plastid" and is a child of GO:0006412 (translation).
{{figure:evidence_summary.png|caption=GO decision table and evidence strength summary showing recommended annotation changes: retain MF GO:0043022, change CC to GO:0009507 (chloroplast), change BP to GO:0032544 (plastid translation).}}
Evidence Matrix
| # | Citation | Evidence Type | Direction | Claim Tested | Key Finding | Context | Confidence |
|---|---|---|---|---|---|---|---|
| 1 | PMID: 25104389 | Structural (direct) | Supports | EF-4 CTD contacts tRNA on ribosome | 2.6 Å crystal structure: CTD reaches into PTC, contacts P-site tRNA acceptor stem | T. thermophilus 70S ribosome | High — direct structural evidence; bacterial system but domain architecture conserved |
| 2 | PMID: 27092003 | Structural (direct) | Supports | CTD contacts A-site tRNA | 2.9 Å crystal structure showing CTD–tRNA acceptor stem contacts at A-site | T. thermophilus 70S ribosome | High — highest-resolution EF-4–ribosome structure |
| 3 | PMID: 26809121 | Structural + mutagenesis | Supports | CTD mediates back-translocation | Cryo-EM + mutagenesis: CTD interferes with CCA–P loop pairing, enhances A/4 contact | E. coli 70S ribosome | High — combined structural and mutational evidence |
| 4 | PMID: 18442968 | Biochemical (direct) | Supports MF; Qualifies CC | GTP-dependent ribosome binding | Co-sedimentation shows GTP-dependent ribosome binding | S. cerevisiae mitochondria | High for binding; evidence is for mitochondrial, not chloroplastic form |
| 5 | PMID: 17110332 | Biochemical (direct) | Supports | EF-4 back-translocates via ribosome binding | LepA back-translocates post-translocation ribosomes | E. coli in vitro | High — foundational discovery |
| 6 | PMID: 21300907 | Biochemical | Supports | EF-4 remobilizes stuck ribosomes | 5-fold enhancement of protein synthesis at elevated Mg²⁺ | E. coli in vitro | High for bacterial; medium for plant transfer |
| 7 | PMID: 18431481 | Proteomics (HDA) | Supports CC correction | Chloroplast localization of ortholog | Q9FNM5 detected in purified chloroplast preparations | A. thaliana chloroplasts | High |
| 8 | PMID: 32344669 | RNA interactome (IDA) | Supports | RNA/ribosome binding in vivo | Q9FNM5 identified as in vivo mRNA-binding protein | A. thaliana leaves | Medium-high — consistent with ribosome association |
| 9 | PMID: 24736607 | Interaction (IPI) | Supports CC | Chloroplast import | Q9FNM5 interacts with Toc import receptors | A. thaliana Y2H | Medium — confirms chloroplast targeting |
| 10 | PMID: 28320876 | Review | Qualifies | Multiple functional hypotheses | Three competing models: back-translocase, stalling relief, ribosome biogenesis — all require ribosome binding | Review of all EF-4 literature | Medium — review-level |
| 11 | PMID: 26486472 | Mutant phenotype | Supports | GUF1 tRNA-binding moiety essential | Human GUF1 p.A609S (in tRNA-binding moiety) causes West syndrome; yeast complementation confirms | Human genetics + yeast | Medium — confirms tRNA-binding importance |
| 12 | PMID: 24837196 | Mutant phenotype | Supports | EF-4 required under stress | C. elegans mtEF4 deletion disrupts mitochondrial function at 15°C | C. elegans | Medium — animal mitochondrial system |
| 13 | PMID: 25491353 | Mutant phenotype | Qualifies | EF-4 has dual role | Promotes ROS-mediated cell death under severe stress; protective under moderate stress | E. coli | Medium — adds nuance |
| 14 | K-mer analysis (this study) | Computational | Supports CC correction | Chloroplastic vs mitochondrial identity | 9.6× higher similarity to chloroplastic Arabidopsis ortholog | A. annua sequence | High — consistent across methods |
| 15 | PANTHER classification | Computational/database | Supports CC correction | Subfamily assignment | PTHR43512:SF4 (CHLOROPLASTIC) | Automated classification | Medium-high |
| 16 | Phylogenetic tree (this study) | Computational/evolutionary | Supports CC correction | Clade membership across 8 species | Clusters with chloroplastic paralogs (distance 0.05); >0.97 to all mitochondrial | 19 plant GUF1 sequences | High — deep clade separation |
| 17 | InterPro domain analysis | Computational | Supports MF | Five-domain EF-4 architecture | All domains confirmed: G-domain (86–245), Domain II (266–336), III (353–431), IV (462–541), LepA_C CTD (549–655) | A. annua A0A2U1PS28 | High |
GO Curation Implications
GO Decision Table
| GO Term | Aspect | Current Status | Recommendation | Evidence Basis | Confidence |
|---|---|---|---|---|---|
| GO:0043022 (ribosome binding) | MF | IEA | RETAIN as core function | Structural (PMIDs 25104389, 27092003, 26809121), biochemical (PMID 18442968), domain architecture | HIGH |
| GO:0003924 (GTPase activity) | MF | IEA | RETAIN | G-domain confirmed; family-level GTPase activity | HIGH |
| GO:0005525 (GTP binding) | MF | IEA | RETAIN | GTP-binding motifs present; GTP-dependent ribosome binding in orthologs | HIGH |
| GO:0070125 (mito. trans. elong.) | BP | IEA | REMOVE — wrong organelle | Protein is chloroplastic (phylogenetics, PANTHER, isoform identity) | HIGH |
| GO:0032544 (plastid translation) | BP | absent | ADD | Correct organellar context; most specific available BP term | MEDIUM-HIGH |
| GO:0006412 (translation) | BP | IEA | RETAIN | General term, correct | HIGH |
| GO:0005759 (mitochondrial matrix) | CC | IEA | REMOVE — wrong organelle | Chloroplastic paralog, not mitochondrial | HIGH |
| GO:0005743 (mito. inner membrane) | CC | IEA | REMOVE — wrong organelle | Chloroplastic paralog, not mitochondrial | HIGH |
| GO:0009507 (chloroplast) | CC | absent | ADD | Ortholog in chloroplast proteomics (PMID 18431481), PANTHER, phylogenetics | HIGH |
| GO:0009570 (chloroplast stroma) | CC | absent | ADD (optional) | By analogy with yeast Guf1 in mitochondrial matrix | MEDIUM |
Molecular Function: GO:0043022 (ribosome binding) — RETAIN as core function
The EF-4/GUF1 family's primary molecular activity is GTP-dependent ribosome binding, which enables back-translocation. GO:0043022 is the most specific available MF term — there is no "chloroplast ribosome binding" GO term (unlike GO:0097177 for mitochondrial ribosome binding). A "plastid ribosome binding" child term could be proposed but does not currently exist. The Arabidopsis ortholog Q9FNM5 has GO:0043022 annotated via IBA (phylogenetic inference from GO_Central), plus IDA evidence for mRNA binding (PMID: 32344669) which is consistent with ribosome association.
Cellular Component: CHANGE from mitochondrial to chloroplast
Current CC annotations (GO:0005743 mitochondrial inner membrane, GO:0005759 mitochondrial matrix) are incorrect for this protein. Add GO:0009507 (chloroplast), matching its ortholog Q9FNM5 in Arabidopsis (HDA evidence from chloroplast proteomics, PMID: 18431481). Optionally add GO:0009570 (chloroplast stroma) by analogy with yeast Guf1 localization to mitochondrial matrix. The true mitochondrial GUF1 in A. annua (A0A2U1PCN3) correctly carries the mitochondrial CC terms.
Biological Process: CHANGE from GO:0070125 to GO:0032544
Replace GO:0070125 (mitochondrial translational elongation) with GO:0032544 (plastid translation). This is the most specific existing BP term for chloroplast/plastid translation. Unlike the mitochondrial side (which has GO:0070125 for translational elongation), there is no "plastid translational elongation" child term. Retain GO:0006412 (translation) as a general annotation.
Mechanistic Scope
Direct Molecular Function
The immediate molecular activity of EF-4/GUF1 is GTP-dependent binding to translating ribosomes. For A0A2U1PS28, this means chloroplast 70S-type ribosomes. The binding involves five domains:
A0A2U1PS28 Domain Architecture (661 aa)
═══════════════════════════════════════════════════
[Transit peptide ~85aa][G-domain 86-245][Dom II 266-336][Dom III 353-431][Dom IV 462-541][CTD 549-655]
├─ GTP binding ─┤ │ │ │
└─ GTP hydrolysis │ │ │
└── Ribosome contact surfaces ───┘
│
Unique to EF-4: contacts tRNA ─┘
acceptor stems in PTC
- Domain I (G-domain, aa 86–245): GTP binding and hydrolysis
- Domain II (aa 266–336): Ribosome interaction surface
- Domain III (aa 353–431): Structural scaffold
- Domain IV (aa 462–541): Ribosome contact
- CTD (aa 549–655): Unique to EF-4; reaches into the peptidyl transferase center and contacts the acceptor stem of ribosome-bound tRNAs
Functional Pathway
A0A2U1PS28 (chloroplastic EF-4, A. annua)
│
├── MF: Ribosome binding (GO:0043022) ← RETAIN
│ ↓ enables
│ GTP-dependent back-translocation of chloroplast ribosome–tRNA complexes
│ ↓ supports
├── BP: Plastid translation (GO:0032544) ← CHANGE from GO:0070125
│ ↓ contributes to
│ Chloroplast protein homeostasis, especially under stress
│ ↓ downstream consequences (not core function)
│ Photosynthetic complex assembly, stress adaptation
│
└── CC: Chloroplast (GO:0009507) ← CHANGE from GO:0005759
Separation from Downstream Effects
The following are downstream consequences of ribosome binding, not the core molecular function itself:
- Translation quality control (dependent on ribosome binding + GTP hydrolysis)
- Stress-responsive enhancement of protein synthesis (conditional phenotype)
- Organellar protein complex assembly defects in knockouts (loss-of-function phenotype)
- Cold/heat-sensitive growth phenotypes (organismal phenotype)
- ROS-mediated cell death under severe stress (E. coli-specific pathway consequence; PMID: 25491353)
For GO curation purposes, GO:0043022 (ribosome binding) correctly captures the direct molecular function, while the biological process term captures the pathway context. The seed hypothesis appropriately separates MF from BP.
Conflicts and Alternatives
1. Mitochondrial vs. Chloroplastic Localization (MAJOR CONFLICT)
The seed hypothesis states "binds to mitochondrial ribosomes" but multiple independent lines of evidence indicate A0A2U1PS28 is the chloroplastic paralog:
- PANTHER subfamily: Classifies as "CHLOROPLASTIC" (PTHR43512:SF4)
- Sequence similarity: 9.6× more similar to Arabidopsis chloroplastic GUF1 (Q9FNM5, Jaccard 0.1874) than mitochondrial (Q9FLE4, Jaccard 0.0195)
- Same gene as chloroplastic isoform: Shares gene locus CTI12_AA119750 with A0A2U1PRZ2 (chloroplastic)
- True mitochondrial paralog: A0A2U1PCN3 (gene CTI12_AA167920) is a distinct, divergent protein (~5% k-mer Jaccard)
- Phylogenetic tree: Clusters with chloroplastic orthologs across 8 plant species with distance 0.05 to nearest chloroplastic member vs. >0.97 to all mitochondrial members
The "mitochondrial" label is an automated UniRule/HAMAP propagation error from the yeast Guf1 experimental evidence. All GO annotations on A0A2U1PS28 carry IEA evidence codes, which propagated the wrong organellar context.
2. Physiological Function Debate
Three competing hypotheses exist for EF-4's physiological function (reviewed in PMID: 28320876):
- (a) Back-translocase: Reverses improper translocation to give EF-G a second chance
- (b) Ribosome stalling relief/sequestration: Binds and remobilizes stuck ribosomes
- (c) Ribosome biogenesis factor: Involved in ribosome assembly
Critically, all three hypotheses require ribosome binding as the core molecular function, so GO:0043022 is robust regardless of which physiological model is ultimately correct.
3. Paralog Confusion Risk
A. annua has at least 4 GUF1/LepA family members, creating real risk of paralog-level misannotation:
| Entry | Length | Organelle | Gene | Status |
|---|---|---|---|---|
| A0A2U1PS28 | 661 aa | Chloroplastic (mislabeled mitochondrial) | CTI12_AA119750 | Needs correction |
| A0A2U1PRZ2 | 683 aa | Chloroplastic (correct) | CTI12_AA119750 | Correct |
| A0A2U1PCN3 | 651 aa | Mitochondrial (correct) | CTI12_AA167920 | Correct |
| A0A2U1LAL8 | 532 aa | Mitochondrial (uncertain) | — | Possibly fragment |
A0A2U1PS28 and A0A2U1PRZ2 are from the same gene and may represent redundant entries from whole-genome sequencing data.
4. Evidence Transfer Distance
All direct ribosome binding evidence comes from bacterial (E. coli, T. thermophilus) or fungal (S. cerevisiae) systems. No direct experimental evidence exists for any plant GUF1 binding to chloroplast ribosomes. However, the extreme conservation of EF-4 across virtually all bacteria, mitochondria, and chloroplasts, combined with the conservation of the full five-domain architecture in A0A2U1PS28, makes the functional transfer reasonable.
5. Dual Role Under Stress
EF-4 has been shown to have a paradoxical dual role: protective under moderate stress (remobilizing stuck ribosomes, enhancing synthesis 5-fold) but detrimental under severe stress by promoting ROS-mediated cell death (PMID: 25491353). For the chloroplastic paralog, this could be relevant in photoinhibition or cold-stress contexts, but has not been tested in any plant system.
Knowledge Gaps
| Gap | What Was Checked | Why It Matters | Resolving Evidence |
|---|---|---|---|
| No direct ribosome binding assay for any plant chloroplastic EF-4 | PubMed searches; no results found | The core MF annotation rests on cross-kingdom homology inference | In vitro ribosome binding assay with recombinant Q9FNM5 or A0A2U1PS28 and purified chloroplast ribosomes |
| No functional characterization of chloroplastic EF-4 in any plant | Searched for plant lepA/guf1/ef4 mutants; none found for chloroplastic form | Unknown whether chloroplastic EF-4 has the same stress-responsive function | T-DNA insertion mutant analysis for Arabidopsis At5g08650 (Q9FNM5) |
| Transit peptide not experimentally validated for A0A2U1PS28 | Computational classification (PANTHER) and k-mer similarity used | Chloroplastic assignment could be wrong if transit peptide is non-functional | GFP-fusion localization in A. annua protoplasts |
| No "plastid translational elongation" GO term exists | QuickGO search confirmed absence | Forces use of broader GO:0032544 | Request creation of GO term |
| Chloroplast ribosome structure differs from bacterial | Literature reviewed; plant chloroplast 70S ribosomes have additional plastid-specific proteins | EF-4 binding interface could differ | Cryo-EM of plant chloroplast ribosome–EF-4 complex |
| A0A2U1PS28 protein existence is PE=3 (homology) | UniProt metadata | Protein existence not experimentally confirmed in A. annua | Proteomics detection in A. annua chloroplast fraction |
Discriminating Tests
High Priority
-
Recombinant ribosome binding assay: Express A0A2U1PS28 (or Arabidopsis Q9FNM5) recombinantly and test binding to purified chloroplast 70S ribosomes using sucrose gradient co-sedimentation. Include GTP/GDP/GMPPNP to test GTP dependence. This would provide the first direct evidence for plant chloroplastic EF-4 ribosome binding.
-
Arabidopsis T-DNA mutant analysis: Characterize At5g08650 (Q9FNM5) knockout/knockdown lines for chloroplast translation defects using ribosome profiling of chloroplast mRNAs, especially under cold stress where EF-4 function is most critical.
-
GFP-fusion localization: Express A0A2U1PS28-GFP in A. annua or tobacco leaf cells and confirm chloroplast targeting by confocal co-localization with chlorophyll autofluorescence. Also stain with MitoTracker to definitively rule out mitochondrial targeting.
Medium Priority
-
Comparative ribosome profiling: Compare chloroplast ribosome occupancy profiles between wild-type and EF-4-depleted Arabidopsis to identify mRNAs dependent on EF-4 for efficient translation elongation.
-
Cross-linking mass spectrometry: Perform XL-MS on chloroplast ribosome preparations to identify EF-4 binding sites on plant chloroplast ribosomes and determine whether the CTD–tRNA interaction is conserved.
-
Cross-complementation in yeast: Test whether A0A2U1PS28 can complement guf1Δ in yeast mitochondria. Failure would support organelle-specific function; success would suggest conserved ribosome binding regardless of compartment.
Evidence Base
Primary Structural Evidence
The strongest evidence for EF-4 ribosome binding comes from three high-resolution structures:
Connell et al. (2014) (PMID: 25104389) — "Crystal structure of elongation factor 4 bound to a clockwise ratcheted ribosome" — captured EF-4 on a ratcheted T. thermophilus ribosome at 2.6 Å, showing the CTD reaching into the peptidyl transferase center to contact P-site tRNA. The key finding: "The C-terminal domain of EF4 reaches into the peptidyl transferase center and interacts with the acceptor stem of the peptidyl-tRNA in the P site."
Kumar et al. (2016) (PMID: 27092003) — "Elongation factor 4 remodels the A-site tRNA on the ribosome" — solved the crystal structure at 2.9 Å, revealing CTD contacts with the A-site tRNA acceptor stem: "The structure reveals the interactions of EF-4 with the A-site tRNA, including contacts between the C-terminal domain (CTD) of EF-4 and the acceptor helical stem of the tRNA."
Zhang et al. (2016) (PMID: 26809121) — "EF4 disengages the peptidyl-tRNA CCA end and facilitates back-translocation on the 70S ribosome" — combined cryo-EM with mutagenesis: "Mutagenesis analyses suggest that a specific region in the EF4 C-terminal domain (CTD) interferes with base-pairing between the peptidyl-tRNA 3'-CCA and the P loop, whereas the EF4 CTD enhances peptidyl-tRNA interaction at the A/4 site."
Functional and Biochemical Evidence
Qin et al. (2006) (PMID: 17110332) — Original discovery: "LepA has the unique function of back-translocating posttranslocational ribosomes, and the results suggest that it recognizes ribosomes after a defective translocation reaction and induces a back-translocation, thus giving EF-G a second chance to translocate the tRNAs correctly."
Liu et al. (2010) (PMID: 20045415) — Elucidated the four-step kinetic mechanism (POST→I₁→I₂→I₃→PRE), showing back-translocation is not the simple reverse of translocation.
Pech et al. (2011) (PMID: 21300907) — Quantified the stress-responsive function: "We suggest that EF4 binds to these stuck ribosomes and remobilizes them, consistent with the EF4-dependent enhancement (fivefold) in protein synthesis observed under these unfavorable conditions."
Bauerschmitt et al. (2008) (PMID: 18442968) — Key eukaryotic study: "It binds to mitochondrial ribosomes in a GTP-dependent manner." This is the study incorrectly cited as evidence for A0A2U1PS28's mitochondrial localization — it studied yeast Guf1 (mitochondrial form).
Plant-Specific Evidence
Zybailov et al. (2008) (PMID: 18431481) — Chloroplast proteomics: "a large scale analysis by nanoLC-Q-TOF and nanoLC-LTQ-Orbitrap mass spectrometry (MS) of ten independent chloroplast preparations from Arabidopsis thaliana which unambiguously identified 1325 proteins" — including Q9FNM5 (At5g08650), providing HDA evidence for chloroplast localization of the Arabidopsis ortholog.
Bach-Pages et al. (2020) (PMID: 32344669) — RNA interactome capture: "Using this improved RIC method in Arabidopsis leaves, we identified 717 RBPs, generating a deep RNA-binding proteome for leaf tissues" — Q9FNM5 was among the identified RNA-binding proteins, consistent with association with translating ribosomes in chloroplasts.
Dutta et al. (2014) (PMID: 24736607) — Split-ubiquitin Y2H showing Q9FNM5 interacts with Toc chloroplast import receptors, confirming chloroplast targeting.
Reviews
Heller et al. (2017) (PMID: 28320876) — "Taking a Step Back from Back-Translocation: an Integrative View of LepA/EF4's Cellular Function" — comprehensive review noting that "LepA/EF4 is highly conserved and present in bacteria, mitochondria, and chloroplasts" and that the CTD is "likely essential to its specific function in the cell."
Connell et al. (2013) (PMID: 23662805) — "The paradox of elongation factor 4: highly conserved, yet of no physiological significance?" — highlights that despite extreme conservation, knockout phenotypes are mild under standard conditions, with function becoming critical only under stress.
Curation Leads
Lead 1: Correct CC Annotation from Mitochondrial to Chloroplast (HIGH PRIORITY)
- Current: CC = GO:0005759 (mitochondrial matrix), GO:0005743 (mitochondrial inner membrane)
- Proposed: CC = GO:0009507 (chloroplast), optionally GO:0009570 (chloroplast stroma)
- Action: REMOVE mitochondrial CC terms, ADD chloroplast CC terms
- Evidence: PANTHER PTHR43512:SF4 (CHLOROPLASTIC); phylogenetic clustering with chloroplastic orthologs across 8 plant species; Arabidopsis ortholog Q9FNM5 in chloroplast proteome (PMID: 18431481)
- Snippet to verify: From PMID 18431481: "Here we present a large scale analysis by nanoLC-Q-TOF and nanoLC-LTQ-Orbitrap mass spectrometry (MS) of ten independent chloroplast preparations from Arabidopsis thaliana which unambiguously identified 1325 proteins."
Lead 2: Retain MF GO:0043022 (Ribosome Binding) as Core Function (MEDIUM PRIORITY)
- Current: MF = GO:0043022 (ribosome binding) [IEA]
- Proposed: RETAIN, update description to reference chloroplast ribosomes
- Evidence: Crystal structures (PMID: 25104389, PMID: 27092003), cryo-EM (PMID: 26809121), biochemistry (PMID: 18442968); domain architecture conserved
- Note: GO:0043022 is the correct most-specific term since no "chloroplast ribosome binding" child term exists
Lead 3: Update BP from Mitochondrial to Plastid Translation (MEDIUM-HIGH PRIORITY)
- Current: BP = GO:0070125 (mitochondrial translational elongation)
- Proposed: REMOVE GO:0070125; ADD GO:0032544 (plastid translation)
- Rationale: Most specific available BP term; no "plastid translational elongation" equivalent exists
- Candidate reference: PMID: 21300907: "Elongation factor 4 (EF4) is one of the most conserved proteins present in bacteria as well as in mitochondria and chloroplasts of eukaryotes."
Lead 4: Flag Potential Isoform Redundancy (LOW PRIORITY)
- A0A2U1PS28 (661 aa) and A0A2U1PRZ2 (683 aa) are from the same gene (CTI12_AA119750) with >95% sequence identity
- Suggested question: Should these be merged as isoforms of the same gene product?
Lead 5: Flag Automated Annotation Pipeline Issue (LOW PRIORITY)
- Suggested question for UniProt/UniRule: Is the HAMAP MF_00071 rule applying the wrong organelle template to this chloroplastic protein? The "mitochondrial" designation appears to have propagated from the yeast Guf1 experimental evidence without accounting for the plant gene duplication that created separate chloroplastic and mitochondrial paralogs.
Lead 6: Consider GO Ontology Term Request
- Suggested action: Propose creation of "plastid translational elongation" as child of GO:0032544, analogous to GO:0070125 (mitochondrial translational elongation)
- Suggested action: Propose creation of "chloroplast ribosome binding" or "plastid ribosome binding" as child of GO:0043022, analogous to GO:0097177 (mitochondrial ribosome binding)
Report generated through three iterations of computational analysis including sequence similarity (k-mer analysis), phylogenetic reconstruction across 8 plant species, domain architecture analysis, GO ontology evaluation, and literature review of 48 papers. All findings are leads requiring curator verification.