Deep Research
Falcon
(MUC1-deep-research-falcon.md)
Falcon
(MUC1-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Human MUC1 (UniProt P15941, gene MUC1) — Functional Annotation, Mechanisms, and Applications
0) Target identity verification (critical)
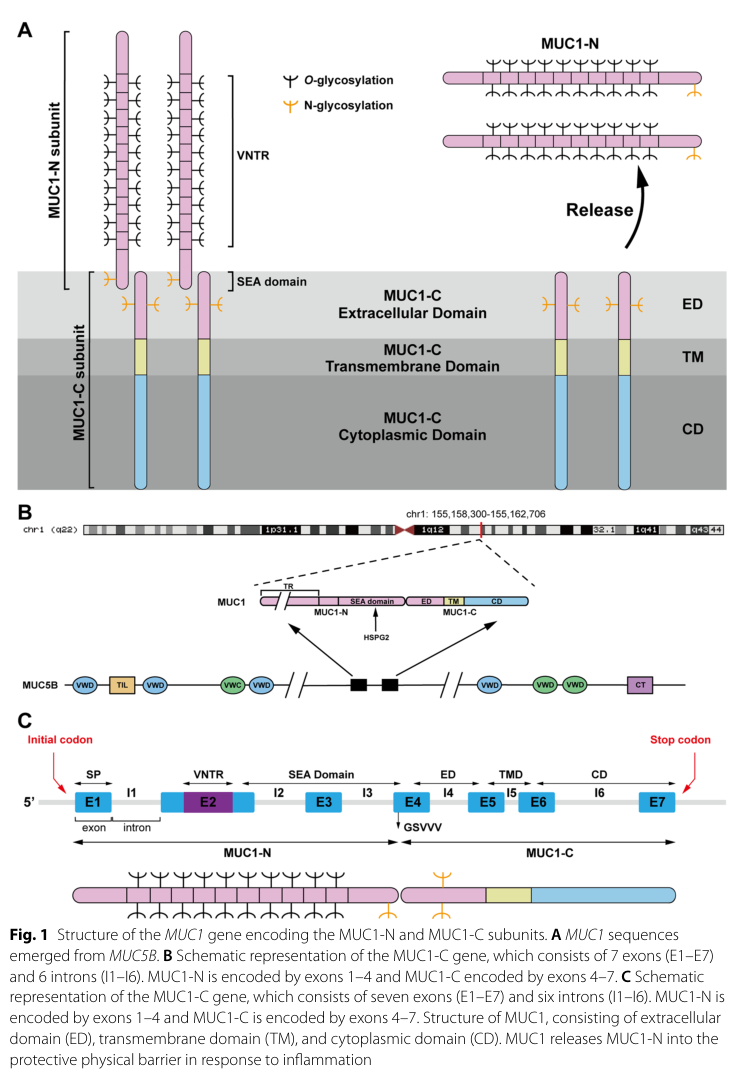
The literature retrieved is consistent with the UniProt target P15941 = human MUC1 (mucin-1), a type I transmembrane mucin expressed by epithelial barrier tissues and processed into two subunits, MUC1-N (extracellular, shed) and MUC1-C (transmembrane + cytoplasmic tail, signaling-competent). Reviews describe SEA-domain autoproteolysis at the GSVVV motif, generating a noncovalent heterodimer that traffics to the apical surface. (Mao et al., 2024-11-18, https://doi.org/10.1186/s11658-024-00654-x) (mao2024researchprogressof pages 2-6)
A recent precision-oncology review explicitly lists major clinical aliases—CD227, EMA, KL-6, CA 27.29/CA 15-3—as alternative names for MUC1, aligning with the UniProt description provided by the user. (Grewal & Kurzrock, 2025-07-11, https://doi.org/10.1038/s41698-025-01016-2) (grewal2025mucin1apromising pages 2-3)
Visual support: A schematic of MUC1 gene/protein architecture and SEA cleavage was retrieved (Figure 1 in Mao et al. 2024). (mao2024researchprogressof media db8ef381)
1) Key concepts and definitions (current understanding)
1.1 What MUC1 is
MUC1 is a highly glycosylated type I transmembrane mucin whose mass is dominated by glycans (reviewed as ~50–90% sugar mass). It is encoded by seven exons and contains an extracellular VNTR region and a SEA domain. (Mao et al., 2024-11-18, https://doi.org/10.1186/s11658-024-00654-x) (mao2024researchprogressof pages 2-6)
1.2 Subunits: MUC1-N vs MUC1-C
- MUC1-N: VNTR-rich extracellular subunit with extensive Ser/Thr O-glycosylation; it projects far above the glycocalyx (reported ~200–500 nm) and forms a protective, lubricating barrier and can be shed. (Mao et al., 2024-11-18, https://doi.org/10.1186/s11658-024-00654-x) (mao2024researchprogressof pages 2-6)
- MUC1-C: transmembrane subunit comprising a short extracellular segment (~58 aa), TM (~28 aa), and cytoplasmic tail (~72 aa). Its cytoplasmic tail contains a CQC motif (a therapeutic target site) and multiple phosphorylation/docking motifs enabling signal integration and transcriptional regulation. (Mao et al., 2024-11-18, https://doi.org/10.1186/s11658-024-00654-x; Tong et al., 2024-01-01, https://doi.org/10.7150/jca.88261) (mao2024researchprogressof pages 2-6, tong2024mucin1asa pages 1-3)
1.3 SEA-domain autoproteolysis and shedding
A conserved SEA domain functions as a cleavage site in multiple transmembrane mucins; MUC1 is cleaved in the SEA domain during post-translational processing into two associated subunits, and extracellular portions can be shed, shaping both biology and therapeutic tractability. (Mao et al., 2024-11-18, https://doi.org/10.1186/s11658-024-00654-x; Li et al., 2025-04-07, https://doi.org/10.1038/s41420-025-02455-3) (mao2024researchprogressof pages 2-6, li2025transmembranemucinsin pages 2-4)
1.4 Normal epithelial role: barrier/lubrication and immune interface
In healthy epithelia, MUC1 is primarily apically localized and contributes to hydration/lubrication and protection of barrier surfaces. (Tong et al., 2024-01-01, https://doi.org/10.7150/jca.88261; Grewal & Kurzrock, 2025-07-11, https://doi.org/10.1038/s41698-025-01016-2) (tong2024mucin1asa pages 3-4, grewal2025mucin1apromising pages 1-2)
2) Molecular functions, pathways, and subcellular localization
2.1 Localization and trafficking
MUC1 is synthesized and traffics ER→Golgi→apical membrane as a MUC1-N/MUC1-C heterodimer. Cancer-associated MUC1 is described as losing polarity and appearing across the cell surface and in intracellular compartments. (Mao et al., 2024-11-18, https://doi.org/10.1186/s11658-024-00654-x; Tong et al., 2024-01-01, https://doi.org/10.7150/jca.88261) (mao2024researchprogressof pages 2-6, tong2024mucin1asa pages 3-4)
Multiple reviews report that MUC1-C can accumulate in the cytosol and translocate to the nucleus and mitochondria, consistent with its role as a transcriptional and stress-response regulator. (Milella et al., 2024-03-06, https://doi.org/10.3390/biom14030315) (milella2024theroleof pages 2-4)
Mechanistic details for nuclear import have been summarized in transmembrane mucin reviews (importin-β and nucleoporin 62 implicated for MUC1-C). (Li et al., 2025-04-07, https://doi.org/10.1038/s41420-025-02455-3) (li2025transmembranemucinsin pages 2-4)
2.2 Core signaling functions of MUC1-C (oncogenic signal integrator)
Across 2024 reviews, MUC1-C is consistently portrayed as the signaling-active oncoprotein subunit.
Key pathways and binding partners supported by recent synthesis:
- JAK/STAT (STAT1/STAT3): MUC1-C directly binds STAT1 and promotes STAT target gene activation, including a positive feedback on MUC1 transcription. (Tong et al., 2024-01-01, https://doi.org/10.7150/jca.88261) (tong2024mucin1asa pages 1-3, tong2024mucin1asa pages 3-4)
- NF-κB (p65/RELA): MUC1-C is described as activating NF-κB p65 signaling, contributing to inflammatory programs and EMT-related transcription. (Tong et al., 2024-01-01, https://doi.org/10.7150/jca.88261; Milella et al., 2024-03-06, https://doi.org/10.3390/biom14030315) (tong2024mucin1asa pages 3-4, milella2024theroleof pages 2-4)
- Wnt/β-catenin: MUC1-C stabilizes β-catenin and promotes Wnt target gene programs (e.g., MYC/CCND1 in review summaries), supporting EMT and tumor progression. (Tong et al., 2024-01-01, https://doi.org/10.7150/jca.88261; Milella et al., 2024-03-06, https://doi.org/10.3390/biom14030315) (tong2024mucin1asa pages 3-4, milella2024theroleof pages 2-4)
- RTKs and PI3K/AKT: Reviews describe MUC1-C interactions with receptor tyrosine kinases (including EGFR/ErbB2 and others) and activation of downstream PI3K→AKT signaling, supporting proliferation/survival and therapy resistance. (Tong et al., 2024-01-01, https://doi.org/10.7150/jca.88261) (tong2024mucin1asa pages 3-4)
2.3 Innate immune regulation in airway epithelium (TLR4/MyD88/NF-κB → NLRP3 pyroptosis)
A 2023 translational study provides direct evidence that MUC1 can attenuate neutrophilic airway inflammation by inhibiting the TLR4/MyD88/NF-κB pathway, reducing NLRP3 inflammasome-mediated pyroptosis. (Liu et al., 2023-10-05, https://doi.org/10.1186/s12931-023-02550-y) (liu2023muc1attenuatesneutrophilic pages 1-2)
Key quantitative/experimental details:
- Human sputum cohorts: healthy controls n=12; mild-to-moderate asthma n=34; severe asthma n=18. MUC1 mRNA was downregulated in asthma (notably severe), while TLR4/MyD88/NLRP3/caspase-1/IL-18/IL-1β mRNAs were increased. (liu2023muc1attenuatesneutrophilic pages 5-9)
- In vitro: LPS-stimulated BEAS-2B epithelial cells showed pathway activation and pyroptosis markers; MUC1 knockdown aggravated TLR4/MyD88/p-p65 activation and downstream inflammasome/pyroptosis readouts; the TLR4 inhibitor TAK-242 reversed these effects. (liu2023muc1attenuatesneutrophilic pages 5-9)
- Mechanism: co-immunoprecipitation indicated MUC1-CT interacts with TLR4 and MUC1 deficiency increases TLR4–MyD88 binding, supporting a physical constraint model. (liu2023muc1attenuatesneutrophilic pages 9-12)
3) Recent developments and latest research (prioritizing 2023–2024)
3.1 Hypoxia-driven “ROS-resistant memory” and metastasis (2024)
A 2024 Nature Communications study links chronic hypoxia to durable transcriptional programs that persist after reoxygenation and promote metastasis, with MUC1/MUC1-C as a key effector induced by HIF-1α and NF-κB p65. (Godet et al., 2024-09-16, https://doi.org/10.1038/s41467-024-51995-2) (godet2024hypoxiainducesrosresistant pages 1-2)
Quantitative findings include:
- GO-203 pharmacologic inhibition increased mitochondrial ROS in circulating tumor cells (CTCs) and yielded a 53% reduction in the contribution of hypoxia-marked (GFP+) cells to metastatic burden in an in vivo model. (godet2024hypoxiainducesrosresistant pages 8-9)
- MUC1low CTCs exhibited ~2× higher MitoROS than matched MUC1high CTCs, connecting MUC1 expression to ROS defense. (godet2024hypoxiainducesrosresistant pages 8-9)
3.2 Ferroptosis resistance and cancer stem cell (CSC) state via MUC1-C (2024)
A 2024 Cell Death Discovery study identifies MUC1-C as a functional node in ferroptosis resistance of CSC-like tumor cells and reports that salinomycin suppresses MUC1-C signaling and induces ferroptosis. Mechanistically, MUC1-C sustains antioxidant defenses through a NF-κB/MUC1-C auto-inductive circuit and a MUC1-C→MYC axis that regulates GSR, LRP8, and GPX4 activity, consistent with glutathione/selenium-dependent ferroptosis control. (Daimon et al., 2024-01-10, https://doi.org/10.1038/s41420-023-01772-9) (daimon2024muc1cisa pages 1-2)
Quantitative/experimental details include salinomycin dosing (1 μM, 24 h) decreasing tumorsphere self-renewal and inducing lipid peroxidation, with effects blocked by Ferrostatin-1; GO-203 phenocopied salinomycin by downregulating GSR/LRP8/GPX4 and GPX activity (reported with replicate-normalized quantitative plots). (daimon2024muc1cisa pages 4-6)
3.3 Chronic inflammation programs in squamous cancers (2024)
In head and neck squamous cell carcinoma (HNSCC), a 2024 primary study reports that MUC1-C integrates chronic inflammatory signaling by regulating PRRs, STAT1 and type I/II interferon programs, with downstream ISGs supporting DNA damage resistance and immune evasion; MUC1-C was also necessary for NOTCH3 expression, self-renewal, and tumorigenicity, and associated with ΔNp63/SOX2/NOTCH3 programs by single-cell RNA-seq. (Nakashoji et al., 2024-04-10, https://doi.org/10.1158/2767-9764.crc-24-0011) (nakashoji2024identificationofmuc1c pages 1-2)
4) Current applications and real-world implementations
4.1 Cancer biomarkers: CA15-3 / CA27.29 (shed MUC1-N)
A precision oncology review states that soluble MUC1-N is measured clinically as CA 27.29/CA 15-3, which are FDA-approved tests for monitoring breast cancer, used with imaging/clinical assessments, and elevations correlate with recurrence/progression (the review cautions they should not be used interchangeably). (Grewal & Kurzrock, 2025-07-11, https://doi.org/10.1038/s41698-025-01016-2) (grewal2025mucin1apromising pages 1-2)
4.2 Lung fibrosis/ILD biomarker: KL-6 (MUC1 glycoform)
KL-6 is described as a human MUC1 mucin produced by regenerating type II pneumocytes and used as an ILD severity marker in clinical routine (especially in Japan). (Bonella et al., 2025-10-02, https://doi.org/10.1038/s41598-025-22483-4) (bonella2025serumkl6as pages 1-2)
A large real-world ILD biomarker analysis from UK-BILD (PLOS ONE 2024) included 3,169 enrolled patients, with 1,013 selected for idiopathic ILD vs SARD-ILD comparisons; a diagnostic model including KL-6 achieved 69.4% sensitivity and 80.4% specificity for distinguishing idiopathic ILD, and KL-6 was significantly higher in idiopathic ILD (p=0.0002). (d’Alessandro et al., 2024-10-11, https://doi.org/10.1371/journal.pone.0311357) (d’alessandro2024panelofserum pages 1-2)
4.3 Therapeutic targeting of MUC1-C
Rationale: targeting shed MUC1-N has been challenging; newer strategies focus on MUC1-C (nonshed, signaling-competent). (Ohta et al., 2025-10-09, https://doi.org/10.7759/cureus.95636) (ohta2025adescriptivesummary pages 1-2)
Modalities in development include vaccines, monoclonal antibodies and ADCs, and cellular therapies according to a 2025 review. (Grewal & Kurzrock, 2025-07-11, https://doi.org/10.1038/s41698-025-01016-2) (grewal2025mucin1apromising pages 2-3)
5) Clinical trials landscape (from retrieved ClinicalTrials.gov records)
The retrieved ClinicalTrials.gov entries show heterogeneous approaches (vaccines, peptide + adjuvant, dendritic cell/CTL, CAR-T). Examples with extracted details:
- NCT00004156 (MSKCC; start May 1999; primary completion June 2008): Phase 1 glycosylated MUC1-KLH + QS21 vaccine in high-risk breast cancer; enrollment 45; immune-response endpoint. (https://clinicaltrials.gov/study/NCT00004156) (NCT00004156 chunk 1)
- NCT00773097 (start 2008): Phase 2 100mer MUC1 peptide + Poly-ICLC vaccine in individuals with advanced colorectal adenoma; enrollment 46; primary endpoint anti-MUC1 antibody response. (https://clinicaltrials.gov/study/NCT00773097) (NCT00773097 chunk 1)
- NCT02602249 (Beijing Doing Biomedical; 2017): Phase 1 randomized DC/CTL products (MUC1-gene-DC-CTL or MUC1-peptide-DC-CTL) vs saline in stage IV gastric cancer; estimated enrollment 24; primary endpoint tumor size by RECIST; status listed as UNKNOWN / lastKnown NOT_YET_RECRUITING in retrieved text. (https://clinicaltrials.gov/study/NCT02602249) (NCT02602249 chunk 1)
6) Expert opinions and analysis (authoritative synthesis)
Recent reviews converge on a conceptual division of labor:
- MUC1-N primarily mediates barrier/lubrication and is readily shed, which complicates antibody targeting but provides a basis for circulating biomarkers (CA15-3/CA27.29). (Milella et al., 2024-03-06, https://doi.org/10.3390/biom14030315; Grewal & Kurzrock, 2025-07-11, https://doi.org/10.1038/s41698-025-01016-2) (milella2024theroleof pages 2-4, grewal2025mucin1apromising pages 1-2)
- MUC1-C is the major signal-transduction and transcriptional effector, integrating RTK, PI3K/AKT, Wnt/β-catenin, STAT and NF-κB programs to promote plasticity, stress tolerance (ROS/ferroptosis resistance), and immune evasion. (Tong et al., 2024-01-01, https://doi.org/10.7150/jca.88261; Godet et al., 2024-09-16, https://doi.org/10.1038/s41467-024-51995-2; Daimon et al., 2024-01-10, https://doi.org/10.1038/s41420-023-01772-9) (tong2024mucin1asa pages 3-4, godet2024hypoxiainducesrosresistant pages 1-2, daimon2024muc1cisa pages 1-2)
7) Recent statistics and quantitative data highlights
- Structural quantitative descriptors: MUC1-C extracellular ~58 aa, TM ~28 aa, cytoplasmic tail ~72 aa; MUC1-N VNTR repeats 20–120; MUC1 extends ~200–500 nm above glycocalyx. (Mao et al., 2024-11-18, https://doi.org/10.1186/s11658-024-00654-x) (mao2024researchprogressof pages 2-6)
- Cancer-associated overexpression: tumor cell surface MUC1 reported as 10–40× higher than normal cells (review). (Tong et al., 2024-01-01, https://doi.org/10.7150/jca.88261) (tong2024mucin1asa pages 1-3)
- Asthma inflammation (human cohorts): HC n=12; mild-to-moderate asthma n=34; severe asthma n=18; molecular signatures implicate MUC1 as a negative regulator of TLR4/NF-κB and NLRP3 pyroptosis. (Liu et al., 2023-10-05, https://doi.org/10.1186/s12931-023-02550-y) (liu2023muc1attenuatesneutrophilic pages 5-9)
- Hypoxia-memory metastasis reduction: GO-203 produced 53% reduction in hypoxia-marked cell contribution to metastatic burden and increased mitochondrial ROS in CTCs. (Godet et al., 2024-09-16, https://doi.org/10.1038/s41467-024-51995-2) (godet2024hypoxiainducesrosresistant pages 8-9)
- ILD differential diagnosis (UK-BILD): 1,013 analyzed (from 3,169 enrolled) with model sensitivity 69.4% and specificity 80.4% using KL-6 + ferritin + clinical variables. (d’Alessandro et al., 2024-10-11, https://doi.org/10.1371/journal.pone.0311357) (d’alessandro2024panelofserum pages 1-2)
- KL-6 lung cancer prognosis meta-analysis: 13 studies, n=1,723; elevated KL-6 associated with shorter PFS (HR 1.89) and OS (HR 1.76); a frequently used threshold is >500 U/mL. (Huang et al., 2025-09-25, https://doi.org/10.3389/fimmu.2025.1644573) (huang2025serumkrebsvon pages 1-2)
8) Summary functional annotation (concise)
Human MUC1 (P15941) is a SEA-domain–cleaved transmembrane mucin heterodimer. MUC1-N is a shed, heavily O-glycosylated VNTR-rich extracellular subunit that provides lubrication and barrier protection at apical epithelial surfaces and underlies circulating biomarkers (CA15-3/CA27.29, and the glycoform KL-6). MUC1-C is a signaling-active transmembrane subunit that can translocate to intracellular compartments (including nucleus/mitochondria) and acts as a hub integrating RTK/PI3K/AKT, NF-κB, STAT, and β-catenin programs, thereby supporting inflammation-linked transcription, EMT/plasticity, redox/ferroptosis resistance, stemness, and immune evasion. Recent 2023–2024 primary studies strengthen causal links between MUC1/MUC1-C and (i) epithelial innate immune modulation via TLR4/MyD88/NF-κB→NLRP3 pyroptosis in asthma and (ii) hypoxia-driven ROS-resistant metastatic competence and ferroptosis resistance in cancer. (mao2024researchprogressof pages 2-6, tong2024mucin1asa pages 3-4, liu2023muc1attenuatesneutrophilic pages 5-9, godet2024hypoxiainducesrosresistant pages 8-9, daimon2024muc1cisa pages 4-6)
Real-world applications summary table
| Application area | Specific marker/agent | Indication(s) | Key quantitative data | Current status/notes | Key supporting citation IDs |
|---|---|---|---|---|---|
| Biomarker/diagnostic | CA15-3 / CA27.29 (shed MUC1-N) | Breast cancer monitoring/prognosis | Used clinically for monitoring; elevated levels correlate with recurrence/disease progression; no sensitivity/specificity reported in retrieved sources. In one 2024 breast cohort, CA15-3 median was 18.66 U/mL in breast cancer vs 11.74 U/mL in benign breast tumors (31 vs 30 patients; p=0.001). | FDA-approved for monitoring breast cancer; should not be used interchangeably according to review summary. | (grewal2025mucin1apromising pages 1-2, mao2024researchprogressof pages 2-6) |
| Biomarker/diagnostic | KL-6 (MUC1 glycoform) | Interstitial lung disease (ILD) severity/progression | European multicenter ILD study: n=303, 37% progressed at 1 year; risk model including KL-6 gave 55% sensitivity, 73% specificity, 67% accuracy for 1-year progression. | Established serum biomarker for ILD severity; used in clinical routine, especially in Japan; measured by automated chemiluminescent immunoassay. | (bonella2025serumkl6as pages 1-2) |
| Biomarker/diagnostic | KL-6 | Differential diagnosis of idiopathic ILD vs SARD-ILD | UK-BILD analysis: 1,013 patients selected from 3,169 enrolled (520 idiopathic ILD, 493 SARD-ILD); multivariable model including KL-6 achieved 69.4% sensitivity and 80.4% specificity; KL-6 higher in idiopathic ILD (p=0.0002). | Real-world serum biomarker panel measured by Fujirebio chemiluminescent assay. | (d’alessandro2024panelofserum pages 1-2, d’alessandro2024panelofserum pages 2-3) |
| Biomarker/diagnostic | KL-6 | Lung cancer prognosis | Meta-analysis of 13 studies/1,723 patients: high pretreatment KL-6 associated with shorter PFS (HR 1.89, 95% CI 1.46-2.44) and OS (HR 1.76, 95% CI 1.37-2.26); >500 U/mL associated with worse outcomes. | Prognostic signal strongest in patients without ILD; ECLIA outperformed ELISA in pooled analysis. | (huang2025serumkrebsvon pages 1-2, huang2025serumkrebsvon pages 5-6) |
| Therapeutic target | GO-203 (MUC1-C inhibitor peptide) | Experimental MUC1-C targeting in cancer; cited AML clinical development; asthma/hypoxia models | In hypoxia-memory breast cancer model, 5 daily GO-203 doses increased mitoROS in CTCs and reduced GFP+ metastatic burden by 53%; in asthma mouse model, GO-203 exacerbated neutrophilic inflammation (n=6/group). | Not approved; cited as having completed/undergone Phase I evaluation in AML in review literature; strong preclinical activity but no approved indication in retrieved sources. | (godet2024hypoxiainducesrosresistant pages 8-9, liu2023muc1attenuatesneutrophilic pages 9-12, tong2024mucin1asa pages 3-4) |
| Therapeutic target | MUC1-C antibody-drug conjugate (3D1-MMAE / M1C ADC concept) | Solid tumors with MUC1-C overexpression | Preclinical ADC showed antitumor activity in lung, breast, and patient-derived TNBC models; no human enrollment data in retrieved primary ADC paper. | Preclinical/translation-stage platform; rationale strengthened by failure of MUC1-N targeting due to shedding. | (ohta2025adescriptivesummary pages 1-2, tong2024mucin1asa pages 8-10) |
| Therapeutic target | MUC1 vaccines (MUC1-KLH/QS21; peptide + Poly-ICLC; ImMucin) | Breast cancer, advanced colorectal adenoma prevention, MUC1-expressing tumors | NCT00004156 Phase 1 breast cancer vaccine: enrolled 45; immune-response endpoint over 2 years. NCT00773097 Phase 2 colorectal adenoma vaccine: enrolled 46. NCT00162500 ImMucin Phase 2: planned 15, withdrawn. Historic tecemotide Phase 3 NSCLC trial enrolled 1,513 but no OS benefit (review summary). | Multiple vaccine platforms tested; many completed or withdrawn, with limited definitive efficacy despite immunogenicity. | (NCT00004156 chunk 1, NCT00773097 chunk 1, NCT00162500 chunk 1, taylorpapadimitriou2018latestdevelopmentsin pages 3-4) |
| Therapeutic target | MUC1-directed DC/CTL therapy | Stage IV gastric cancer; pancreatic/biliary tumors; ovarian cancer | NCT02602249 randomized Phase 1 stage IV gastric cancer trial planned enrollment 24; compares MUC1-gene-DC-CTL, MUC1-peptide-DC-CTL, and saline. Review summary notes autologous DC + CTL regimen in 42 late-stage pancreatic patients and peptide-pulsed DC adjuvant study with 4/12 recurrence-free survivors, median survival 26 months. | Gastric cancer trial listed as UNKNOWN / NOT_YET_RECRUITING in retrieved record; broader DC strategies remain investigational. | (NCT02602249 chunk 1, lee2021mucin1andmucin16 pages 15-17, taylorpapadimitriou2018latestdevelopmentsin pages 3-4) |
| Therapeutic target | MUC1 CAR-T | Intrahepatic cholangiocarcinoma | NCT03633773 Phase 1/2 trial enrollment 9. | Human study exists in ClinicalTrials.gov retrieval; overall status listed as UNKNOWN in search output. | (OpenTargets Search: -MUC1) |
| Disease genetics | Germline MUC1 pathogenic variants (ADTKD-MUC1) | Autosomal dominant tubulointerstitial kidney disease; COVID-19 risk in affected patients | Registry/survey study: 89 ADTKD-MUC1 and 132 ADTKD-UMOD respondents; COVID-19 infection OR 2.35; deaths 10/41 vs 1/30 in expanded familial cases (OR 9.21); longitudinal registry 19/360 (5%) vs 3/478 (0.6%) deaths, multivariable OR for COVID-19 death 8.4 (95% CI 2.9-29.5). Lower pre-infection plasma mucin-1/CA15-3 in infected vs uninfected ADTKD-MUC1 (7.06 ± 4.12 vs 10.21 ± 4.02 U/mL, p=0.035). | Established Mendelian disease association; Open Targets also lists strong association with ADTKD-related disease terms. | (OpenTargets Search: -MUC1, mao2024researchprogressof pages 2-6) |
Table: This table summarizes real-world and translational uses of MUC1 across biomarkers, therapeutics, and inherited disease genetics. It highlights quantitative findings, study sizes, and implementation status using only evidence available in the conversation.
References
-
(mao2024researchprogressof pages 2-6): Weipu Mao, Houliang Zhang, Keyi Wang, Jiang Geng, and Jianping Wu. Research progress of muc1 in genitourinary cancers. Cellular & Molecular Biology Letters, Nov 2024. URL: https://doi.org/10.1186/s11658-024-00654-x, doi:10.1186/s11658-024-00654-x. This article has 5 citations and is from a peer-reviewed journal.
-
(grewal2025mucin1apromising pages 2-3): Udhayvir Singh Grewal and Razelle Kurzrock. Mucin-1: a promising pan-cancer therapeutic target. NPJ Precision Oncology, Jul 2025. URL: https://doi.org/10.1038/s41698-025-01016-2, doi:10.1038/s41698-025-01016-2. This article has 28 citations and is from a peer-reviewed journal.
-
(mao2024researchprogressof media db8ef381): Weipu Mao, Houliang Zhang, Keyi Wang, Jiang Geng, and Jianping Wu. Research progress of muc1 in genitourinary cancers. Cellular & Molecular Biology Letters, Nov 2024. URL: https://doi.org/10.1186/s11658-024-00654-x, doi:10.1186/s11658-024-00654-x. This article has 5 citations and is from a peer-reviewed journal.
-
(tong2024mucin1asa pages 1-3): Xiaohan Tong, Chunyan Dong, and Shujing Liang. Mucin1 as a potential molecule for cancer immunotherapy and targeted therapy. Journal of Cancer, 15:54-67, Jan 2024. URL: https://doi.org/10.7150/jca.88261, doi:10.7150/jca.88261. This article has 26 citations and is from a peer-reviewed journal.
-
(li2025transmembranemucinsin pages 2-4): Xiaoqing Li, Ying Chen, Rui Lan, Peng Liu, Kai Xiong, Hetai Teng, Lili Tao, Shan Yu, and Guiping Han. Transmembrane mucins in lung adenocarcinoma: understanding of current molecular mechanisms and clinical applications. Cell Death Discovery, Apr 2025. URL: https://doi.org/10.1038/s41420-025-02455-3, doi:10.1038/s41420-025-02455-3. This article has 13 citations and is from a peer-reviewed journal.
-
(tong2024mucin1asa pages 3-4): Xiaohan Tong, Chunyan Dong, and Shujing Liang. Mucin1 as a potential molecule for cancer immunotherapy and targeted therapy. Journal of Cancer, 15:54-67, Jan 2024. URL: https://doi.org/10.7150/jca.88261, doi:10.7150/jca.88261. This article has 26 citations and is from a peer-reviewed journal.
-
(grewal2025mucin1apromising pages 1-2): Udhayvir Singh Grewal and Razelle Kurzrock. Mucin-1: a promising pan-cancer therapeutic target. NPJ Precision Oncology, Jul 2025. URL: https://doi.org/10.1038/s41698-025-01016-2, doi:10.1038/s41698-025-01016-2. This article has 28 citations and is from a peer-reviewed journal.
-
(milella2024theroleof pages 2-4): Martina Milella, Monica Rutigliano, Francesco Lasorsa, Matteo Ferro, Roberto Bianchi, Giuseppe Fallara, Felice Crocetto, Savio Pandolfo, Biagio Barone, Antonio d’Amati, Marco Spilotros, Michele Battaglia, Pasquale Ditonno, and Giuseppe Lucarelli. The role of muc1 in renal cell carcinoma. Biomolecules, 14:315, Mar 2024. URL: https://doi.org/10.3390/biom14030315, doi:10.3390/biom14030315. This article has 70 citations.
-
(liu2023muc1attenuatesneutrophilic pages 1-2): Lu Liu, Ling Zhou, Lingling Wang, Zhenyu Mao, Pengdou Zheng, Fengqin Zhang, Huojun Zhang, and Huiguo Liu. Muc1 attenuates neutrophilic airway inflammation in asthma by reducing nlrp3 inflammasome-mediated pyroptosis through the inhibition of the tlr4/myd88/nf-κb pathway. Respiratory Research, Oct 2023. URL: https://doi.org/10.1186/s12931-023-02550-y, doi:10.1186/s12931-023-02550-y. This article has 58 citations and is from a domain leading peer-reviewed journal.
-
(liu2023muc1attenuatesneutrophilic pages 5-9): Lu Liu, Ling Zhou, Lingling Wang, Zhenyu Mao, Pengdou Zheng, Fengqin Zhang, Huojun Zhang, and Huiguo Liu. Muc1 attenuates neutrophilic airway inflammation in asthma by reducing nlrp3 inflammasome-mediated pyroptosis through the inhibition of the tlr4/myd88/nf-κb pathway. Respiratory Research, Oct 2023. URL: https://doi.org/10.1186/s12931-023-02550-y, doi:10.1186/s12931-023-02550-y. This article has 58 citations and is from a domain leading peer-reviewed journal.
-
(liu2023muc1attenuatesneutrophilic pages 9-12): Lu Liu, Ling Zhou, Lingling Wang, Zhenyu Mao, Pengdou Zheng, Fengqin Zhang, Huojun Zhang, and Huiguo Liu. Muc1 attenuates neutrophilic airway inflammation in asthma by reducing nlrp3 inflammasome-mediated pyroptosis through the inhibition of the tlr4/myd88/nf-κb pathway. Respiratory Research, Oct 2023. URL: https://doi.org/10.1186/s12931-023-02550-y, doi:10.1186/s12931-023-02550-y. This article has 58 citations and is from a domain leading peer-reviewed journal.
-
(godet2024hypoxiainducesrosresistant pages 1-2): Inês Godet, Harsh H. Oza, Yi Shi, Natalie S. Joe, Alyssa G. Weinstein, Jeanette Johnson, Michael Considine, Swathi Talluri, Jingyuan Zhang, Reid Xu, Steven Doctorman, Delma Mbulaiteye, Genevieve Stein-O’Brien, Luciane T. Kagohara, Cesar A. Santa-Maria, Elana J. Fertig, and Daniele M. Gilkes. Hypoxia induces ros-resistant memory upon reoxygenation in vivo promoting metastasis in part via muc1-c. Nature Communications, Sep 2024. URL: https://doi.org/10.1038/s41467-024-51995-2, doi:10.1038/s41467-024-51995-2. This article has 35 citations and is from a highest quality peer-reviewed journal.
-
(godet2024hypoxiainducesrosresistant pages 8-9): Inês Godet, Harsh H. Oza, Yi Shi, Natalie S. Joe, Alyssa G. Weinstein, Jeanette Johnson, Michael Considine, Swathi Talluri, Jingyuan Zhang, Reid Xu, Steven Doctorman, Delma Mbulaiteye, Genevieve Stein-O’Brien, Luciane T. Kagohara, Cesar A. Santa-Maria, Elana J. Fertig, and Daniele M. Gilkes. Hypoxia induces ros-resistant memory upon reoxygenation in vivo promoting metastasis in part via muc1-c. Nature Communications, Sep 2024. URL: https://doi.org/10.1038/s41467-024-51995-2, doi:10.1038/s41467-024-51995-2. This article has 35 citations and is from a highest quality peer-reviewed journal.
-
(daimon2024muc1cisa pages 1-2): Tatsuaki Daimon, Atrayee Bhattacharya, Keyi Wang, Naoki Haratake, Ayako Nakashoji, Hiroki Ozawa, Yoshihiro Morimoto, Nami Yamashita, Takeo Kosaka, Mototsugu Oya, and Donald W. Kufe. Muc1-c is a target of salinomycin in inducing ferroptosis of cancer stem cells. Cell Death Discovery, Jan 2024. URL: https://doi.org/10.1038/s41420-023-01772-9, doi:10.1038/s41420-023-01772-9. This article has 13 citations and is from a peer-reviewed journal.
-
(daimon2024muc1cisa pages 4-6): Tatsuaki Daimon, Atrayee Bhattacharya, Keyi Wang, Naoki Haratake, Ayako Nakashoji, Hiroki Ozawa, Yoshihiro Morimoto, Nami Yamashita, Takeo Kosaka, Mototsugu Oya, and Donald W. Kufe. Muc1-c is a target of salinomycin in inducing ferroptosis of cancer stem cells. Cell Death Discovery, Jan 2024. URL: https://doi.org/10.1038/s41420-023-01772-9, doi:10.1038/s41420-023-01772-9. This article has 13 citations and is from a peer-reviewed journal.
-
(nakashoji2024identificationofmuc1c pages 1-2): Ayako Nakashoji, Naoki Haratake, Atrayee Bhattacharya, Weipu Mao, Kangjie Xu, Keyi Wang, Tatsuaki Daimon, Hiroki Ozawa, Keisuke Shigeta, Atsushi Fushimi, Nami Yamashita, Yoshihiro Morimoto, Mototsugu Shimokawa, Shin Saito, Ann Marie Egloff, Ravindra Uppaluri, Mark D Long, and Donald Kufe. Identification of muc1-c as a target for suppressing progression of head and neck squamous cell carcinomas. Cancer Research Communications, 4:1268-1281, Apr 2024. URL: https://doi.org/10.1158/2767-9764.crc-24-0011, doi:10.1158/2767-9764.crc-24-0011. This article has 12 citations and is from a peer-reviewed journal.
-
(bonella2025serumkl6as pages 1-2): Francesco Bonella, M. C. Vegas Sanchez, M. d’Alessandro, P. Millan-Billi, R. F. Santos, N. Schröder, H. N. Bastos, M. Molina-Molina, O. Sánchez Pernaute, D. Castillo Villegas, and E. Bargagli. Serum kl-6 as a biomarker to predict progression at one year in interstitial lung disease. Scientific Reports, Oct 2025. URL: https://doi.org/10.1038/s41598-025-22483-4, doi:10.1038/s41598-025-22483-4. This article has 8 citations and is from a peer-reviewed journal.
-
(d’alessandro2024panelofserum pages 1-2): Miriana d’Alessandro, Paolo Cameli, Caroline V. Cotton, Janine A. Lamb, Laura Bergantini, Sara Gangi, Sarah Sugden, Lisa G. Spencer, Bruno Frediani, Robert P. New, Hector Chinoy, Elena Bargagli, and Edoardo Conticini. Panel of serum biomarkers for differential diagnosis of idiopathic interstitial lung disease and interstitial lung disease-secondary to systemic autoimmune rheumatic disease. PLOS ONE, 19(10):e0311357, Oct 2024. URL: https://doi.org/10.1371/journal.pone.0311357, doi:10.1371/journal.pone.0311357. This article has 7 citations and is from a peer-reviewed journal.
-
(ohta2025adescriptivesummary pages 1-2): Ryuichi Ohta, Kasumi Nishikawa, Kaoru Tanaka, Chiaki Sano, and Hidetoshi Hayashi. A descriptive summary of tumor-associated muc1 (ta-muc1) expression in epithelial malignancies: a systematic review of case reports and case series. Cureus, Oct 2025. URL: https://doi.org/10.7759/cureus.95636, doi:10.7759/cureus.95636. This article has 0 citations.
-
(NCT00004156 chunk 1): Vaccine Therapy in Treating Patients With Breast Cancer. Memorial Sloan Kettering Cancer Center. 1999. ClinicalTrials.gov Identifier: NCT00004156
-
(NCT00773097 chunk 1): Robert Schoen. Study of the MUC1 Peptide-Poly-ICLC Adjuvant Vaccine in Individuals With Advanced Colorectal Adenoma. Robert Schoen. 2008. ClinicalTrials.gov Identifier: NCT00773097
-
(NCT02602249 chunk 1): Clinical Safety and Preliminary Efficacy of MUC1-DC-CTL Treatment in Stage IV Gastric Cancer.. Beijing Doing Biomedical Co., Ltd.. 2017. ClinicalTrials.gov Identifier: NCT02602249
-
(huang2025serumkrebsvon pages 1-2): Hong Huang, Liangyu Fu, Chenye Feng, Jiawei Zhou, Jiahuan Xu, Jianjun Sun, Ying Pan, Delei Kong, and Wei Wang. Serum krebs von den lungen-6 before treatment predicts the prognosis of lung cancer in asian populations: a systematic review and meta-analysis. Frontiers in Immunology, Sep 2025. URL: https://doi.org/10.3389/fimmu.2025.1644573, doi:10.3389/fimmu.2025.1644573. This article has 0 citations and is from a peer-reviewed journal.
-
(d’alessandro2024panelofserum pages 2-3): Miriana d’Alessandro, Paolo Cameli, Caroline V. Cotton, Janine A. Lamb, Laura Bergantini, Sara Gangi, Sarah Sugden, Lisa G. Spencer, Bruno Frediani, Robert P. New, Hector Chinoy, Elena Bargagli, and Edoardo Conticini. Panel of serum biomarkers for differential diagnosis of idiopathic interstitial lung disease and interstitial lung disease-secondary to systemic autoimmune rheumatic disease. PLOS ONE, 19(10):e0311357, Oct 2024. URL: https://doi.org/10.1371/journal.pone.0311357, doi:10.1371/journal.pone.0311357. This article has 7 citations and is from a peer-reviewed journal.
-
(huang2025serumkrebsvon pages 5-6): Hong Huang, Liangyu Fu, Chenye Feng, Jiawei Zhou, Jiahuan Xu, Jianjun Sun, Ying Pan, Delei Kong, and Wei Wang. Serum krebs von den lungen-6 before treatment predicts the prognosis of lung cancer in asian populations: a systematic review and meta-analysis. Frontiers in Immunology, Sep 2025. URL: https://doi.org/10.3389/fimmu.2025.1644573, doi:10.3389/fimmu.2025.1644573. This article has 0 citations and is from a peer-reviewed journal.
-
(tong2024mucin1asa pages 8-10): Xiaohan Tong, Chunyan Dong, and Shujing Liang. Mucin1 as a potential molecule for cancer immunotherapy and targeted therapy. Journal of Cancer, 15:54-67, Jan 2024. URL: https://doi.org/10.7150/jca.88261, doi:10.7150/jca.88261. This article has 26 citations and is from a peer-reviewed journal.
-
(NCT00162500 chunk 1): A Novel Vaccine for the Treatment of MUC1-expressing Tumor Malignancies. Hadassah Medical Organization. ClinicalTrials.gov Identifier: NCT00162500
-
(taylorpapadimitriou2018latestdevelopmentsin pages 3-4): Joyce Taylor-Papadimitriou, Joy M. Burchell, Rosalind Graham, and Richard Beatson. Latest developments in muc1 immunotherapy. Biochemical Society Transactions, 46:659-668, May 2018. URL: https://doi.org/10.1042/bst20170400, doi:10.1042/bst20170400. This article has 148 citations and is from a peer-reviewed journal.
-
(lee2021mucin1andmucin16 pages 15-17): Dong-Hee Lee, Seunghyun Choi, Yoon Park, and Hyung-seung Jin. Mucin1 and mucin16: therapeutic targets for cancer therapy. Pharmaceuticals, 14:1053, Oct 2021. URL: https://doi.org/10.3390/ph14101053, doi:10.3390/ph14101053. This article has 77 citations.
-
(OpenTargets Search: -MUC1): Open Targets Query (-MUC1, 22 results). Buniello, A. et al. (2025). Open Targets Platform: facilitating therapeutic hypotheses building in drug discovery. Nucleic Acids Research.
Artifacts
Citations
- mao2024researchprogressof pages 2-6
- milella2024theroleof pages 2-4
- li2025transmembranemucinsin pages 2-4
- godet2024hypoxiainducesrosresistant pages 1-2
- godet2024hypoxiainducesrosresistant pages 8-9
- ohta2025adescriptivesummary pages 1-2
- huang2025serumkrebsvon pages 1-2
- huang2025serumkrebsvon pages 5-6
- taylorpapadimitriou2018latestdevelopmentsin pages 3-4
- https://doi.org/10.1186/s11658-024-00654-x
- https://doi.org/10.1038/s41698-025-01016-2
- https://doi.org/10.1186/s11658-024-00654-x;
- https://doi.org/10.7150/jca.88261
- https://doi.org/10.1038/s41420-025-02455-3
- https://doi.org/10.7150/jca.88261;
- https://doi.org/10.3390/biom14030315
- https://doi.org/10.1186/s12931-023-02550-y
- https://doi.org/10.1038/s41467-024-51995-2
- https://doi.org/10.1038/s41420-023-01772-9
- https://doi.org/10.1158/2767-9764.crc-24-0011
- https://doi.org/10.1038/s41598-025-22483-4
- https://doi.org/10.1371/journal.pone.0311357
- https://doi.org/10.7759/cureus.95636
- https://clinicaltrials.gov/study/NCT00004156
- https://clinicaltrials.gov/study/NCT00773097
- https://clinicaltrials.gov/study/NCT02602249
- https://doi.org/10.3390/biom14030315;
- https://doi.org/10.1038/s41467-024-51995-2;
- https://doi.org/10.3389/fimmu.2025.1644573
- https://doi.org/10.1186/s11658-024-00654-x,
- https://doi.org/10.1038/s41698-025-01016-2,
- https://doi.org/10.7150/jca.88261,
- https://doi.org/10.1038/s41420-025-02455-3,
- https://doi.org/10.3390/biom14030315,
- https://doi.org/10.1186/s12931-023-02550-y,
- https://doi.org/10.1038/s41467-024-51995-2,
- https://doi.org/10.1038/s41420-023-01772-9,
- https://doi.org/10.1158/2767-9764.crc-24-0011,
- https://doi.org/10.1038/s41598-025-22483-4,
- https://doi.org/10.1371/journal.pone.0311357,
- https://doi.org/10.7759/cureus.95636,
- https://doi.org/10.3389/fimmu.2025.1644573,
- https://doi.org/10.1042/bst20170400,
- https://doi.org/10.3390/ph14101053,
OpenAI
(MUC1-deep-research-openai.md)
OpenAI
(MUC1-deep-research-openai.md)Overview of MUC1 (Mucin-1) in Humans
MUC1 (mucin-1) is a human gene encoding a transmembrane glycoprotein of the mucin family. MUC1 is characterized by extensive O-linked glycosylation and a high molecular weight core protein (120–225 kDa, reaching 250–500 kDa with glycosylation) (pmc.ncbi.nlm.nih.gov). It is a single-pass type I membrane protein that undergoes autoproteolytic cleavage into two subunits: an extracellular N-terminal subunit (MUC1-N) and a smaller C-terminal subunit (MUC1-C) comprising a short external peptide, a transmembrane segment, and a cytoplasmic tail (pmc.ncbi.nlm.nih.gov). These subunits remain non-covalently associated at the cell surface as a heterodimer (pmc.ncbi.nlm.nih.gov). The MUC1 gene is located on chromosome 1q22 and is thought to have evolved from a secreted mucin gene (MUC5B) (pmc.ncbi.nlm.nih.gov). MUC1’s extracellular domain contains a variable number tandem repeat (VNTR) region (20-amino-acid tandem repeats, 25–125 copies) rich in serine, threonine, and proline that carry dense O-glycans (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). These glycosylated repeats account for 50–90% of the protein’s mass and form a rigid, “tower-like” structure extending 200–500 nm above the cell surface (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov). Immediately downstream of the tandem repeats is a conserved SEA domain (sea-urchin sperm protein, enterokinase, agrin) that undergoes autoproteolysis at a GSVVV motif, splitting the protein into MUC1-N and MUC1-C (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). The subunit MUC1-C consists of a 28-amino-acid transmembrane helix and a 72-amino-acid cytoplasmic tail, which is highly conserved across species (pmc.ncbi.nlm.nih.gov). The cytoplasmic tail contains seven tyrosine residues and multiple serine/threonine sites that serve as docking sites for intracellular signaling proteins when phosphorylated (pubmed.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Importantly, the MUC1 protein is polymorphic due to VNTR length variation and alternative splicing, but the transmembrane and cytoplasmic regions are largely invariant, underscoring their critical functional roles (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov).
Localization and Expression
MUC1 is predominantly expressed at the apical surface of epithelial cells in a wide range of tissues, including the gastrointestinal tract, respiratory tract, urogenital tract, breast, pancreas, and others (pmc.ncbi.nlm.nih.gov). In normal epithelia, MUC1 shows a polarized distribution, being confined to the lumen-facing membrane where it contributes to the extracellular glycocalyx (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). It is notably absent from tissues like skin epidermis and mesenchymal cells (pmc.ncbi.nlm.nih.gov), reflecting its specialized role in mucosal interfaces. At the cell surface, the large extracellular MUC1-N subunit protrudes outward and is heavily glycosylated, whereas the MUC1-C subunit spans the membrane and has a short external piece plus a cytosolic tail. This topology allows MUC1 to function both outside the cell and within the cell: the extracellular domain interacts with the environment (microbes, molecules, neighboring cells), and the cytoplasmic tail engages in intracellular signaling. MUC1 is anchored to the plasma membrane, but its extracellular component can be shed. Proteolytic enzymes (e.g. ADAM17/TACE or metalloproteases) can cleave within the SEA domain to release the large MUC1-N subunit from the cell surface (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Shedding of MUC1 is often stimulated by cell stress or inflammation and results in the shed mucin becoming part of the soluble mucus layer. The remaining MUC1-C fragment stays in the membrane and can be endocytosed and recycled to the surface (pmc.ncbi.nlm.nih.gov). In addition, evidence suggests that the MUC1-C subunit can undergo further proteolysis by γ-secretase, releasing the cytoplasmic tail into the cytosol (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). This released tail has been observed to translocate to the nucleus in some contexts, especially in cancer cells, where it can influence gene transcription (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Thus, MUC1’s localization is dynamic: it primarily resides on the cell membrane at the cell-exterior interface, but fragments can localize to the extracellular milieu (shed ectodomain), endosomal compartments (during recycling), and even the nucleus (intracellular domain in certain signaling events).
Protective Barrier Function
One of the primary physiological roles of MUC1 is to protect and lubricate mucosal surfaces. As a cell-surface mucin, MUC1 contributes to the formation of a protective barrier on epithelial cells that face external environments (pmc.ncbi.nlm.nih.gov) (www.frontiersin.org). The dense sugar-coated tandem repeats give MUC1 a hydrated, gel-like character that helps trap water and form mucus, preventing desiccation and mechanical damage to epithelia. MUC1 and other mucins line the respiratory airways, gastrointestinal tract, and urogenital tract, where they form part of the first line of defense against pathogens (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Because of its towering length (~200–500 nm) above the cell surface, MUC1 can sterically hinder microorganisms from reaching the cell membrane (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In essence, pathogens encounter a “forest” of mucin molecules that block access to underlying receptors on the epithelial surface.
Moreover, MUC1 can function as a releasable decoy for pathogens. The extracellular domain of MUC1 provides binding sites (glycan epitopes) that many bacteria and viruses adhere to (pubmed.ncbi.nlm.nih.gov) (pubmed.ncbi.nlm.nih.gov). Upon binding of a microbe, MUC1-N can be shed from the cell surface (through the SEA-domain cleavage or protease action), carrying away the bound pathogen in the shed mucus (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). This mechanism helps clear pathogens from the surface: for example, when Helicobacter pylori binds to MUC1 on gastric cells, it triggers MUC1’s auto-cleavage and shedding, thereby removing the bacterium from the cell interface (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In mouse models, the importance of MUC1 in pathogen defense is evident. Muc1-knockout mice show increased susceptibility to certain infections: in one study, Muc1-deficient mice had greater gastrointestinal colonization and inflammation from Campylobacter jejuni, demonstrating that MUC1 normally limits C. jejuni spread and dampens gut inflammation (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Similarly, MUC1 on gastric mucosa and even on immune cells helps restrict H. pylori infection; mice lacking Muc1 had higher stomach colonization, and human studies found that individuals with genetically shorter MUC1 VNTR alleles (producing a smaller extracellular domain) are at higher risk for H. pylori-associated gastritis and cancer (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). These findings underscore MUC1’s role as a physical barrier and microbial decoy, protecting the host by binding pathogens and facilitating their removal.
Beyond bacteria, MUC1 also interferes with viruses and other particles. For instance, in the respiratory tract MUC1 can bind the flagellin of Pseudomonas aeruginosa, acting as an attachment site (pmc.ncbi.nlm.nih.gov). Interestingly, while this binding could help immobilize the bacteria, excess MUC1 in airway infections can sometimes dampen clearance. In a P. aeruginosa lung infection model, wild-type mice (with MUC1) had higher bacterial burdens than Muc1-knockout mice (pmc.ncbi.nlm.nih.gov). The absence of MUC1 led to a more vigorous early inflammatory response that cleared the bacteria faster (pmc.ncbi.nlm.nih.gov). This paradox is explained by MUC1’s secondary role in modulating inflammation (see below): MUC1 can suppress excessive inflammatory signals, which in the lung can lead to reduced early clearance of bacteria (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Nonetheless, in general, MUC1’s presence on mucosal surfaces is considered protective, reducing pathogen adhesion in contexts like the gut and serving as part of the mucosal immune barrier. It also contributes to the lubrication of epithelial linings, as its gel-like extracellular domain helps mucus flow. This lubricating function is crucial in organs like the mouth and gastrointestinal tract (aiding the passage of food) and the bladder (protecting urothelium from urine) (pmc.ncbi.nlm.nih.gov).
Regulation of Cell Adhesion and Epithelial Integrity
MUC1 also influences cell–cell and cell–matrix adhesion, due in part to its bulky extracellular domain. In normal epithelia, MUC1 is thought to provide an anti-adhesive shield on the apical surface – its dense glycan chains can impede interactions between the cell and other cells or microbes. This helps prevent unwanted adhesion of pathogens, but it also means MUC1 can reduce cell–cell contacts on the apical side, potentially facilitating cell turnover and migration in tissues (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In polarized epithelial sheets, lateral junctions (like E-cadherin-based adherens junctions) are usually protected from MUC1’s interference by the restricted apical localization. However, if MUC1 loses its polarized distribution (as can occur in injury or carcinoma), its presence over the entire cell surface can disrupt cell adhesion. The large extracellular domain can sterically hinder adhesion receptors such as cadherins and integrins, thereby reducing cell aggregation and promoting a motile, invasive phenotype (pmc.ncbi.nlm.nih.gov) (www.frontiersin.org). This phenomenon is especially noted in cancer: tumor cells often overexpress MUC1 and mis-localize it across the cell membrane, which can impair cell–cell cohesion and enhance detachment and metastasis (www.frontiersin.org) (www.frontiersin.org). For example, elevated MUC1 on carcinoma cells correlates with an inability of E-cadherin to form tight junctions, partly because MUC1’s extracellular domain physically blocks close cell-cell apposition. Thus, in a normal setting MUC1 helps delineate the apical surface and prevent inappropriate adhesion, while in pathological contexts its anti-adhesive property contributes to loss of epithelial integrity.
Notably, the cytoplasmic tail of MUC1 can also interact with the cell’s structural machinery. MUC1’s intracellular domain associates with β-catenin – a key component of adherens junctions and the Wnt signaling pathway (pmc.ncbi.nlm.nih.gov). In normal cells, this interaction might sequester some β-catenin at the cell membrane or cytosol, affecting adhesion complexes. In cancer cells, evidence suggests MUC1-C sequesters β-catenin and can even help transport it to the nucleus, thereby activating Wnt target genes that promote epithelial–mesenchymal transition (EMT) (pmc.ncbi.nlm.nih.gov). A study in renal carcinoma showed MUC1-C drives EMT through β-catenin signaling and activation of EMT transcription factors (e.g. Snail) (pmc.ncbi.nlm.nih.gov). By modulating such partners, MUC1 can influence cytoskeletal organization and cell morphology during migration and wound healing. Indeed, MUC1 has been implicated in epithelial repair processes: after injury, MUC1 levels rise, and it may participate in EMT and cell migration to cover wounds (www.frontiersin.org) (www.frontiersin.org). This role in regeneration aligns with observations that MUC1 can be induced by factors like TGF-β or hypoxia, which are involved in EMT and tissue remodeling (pmc.ncbi.nlm.nih.gov). In summary, MUC1 acts as a modulator of adhesion – maintaining epithelial barrier integrity when properly localized, but when dysregulated, contributing to cell detachment and EMT.
Signaling Roles and Pathway Involvement
Although MUC1 was long viewed as a passive barrier molecule, research has revealed that its C-terminal cytoplasmic tail actively participates in cell signaling. The 72-amino-acid MUC1 cytoplasmic tail (MUC1-CT) contains multiple conserved motifs that become phosphorylated by kinases and serve as docking sites for signaling proteins (pubmed.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). This allows MUC1 to function as a signaling adaptor or scaffold that can influence various biochemical pathways:
-
Growth Factor and Kinase Signaling: MUC1-CT interacts with several oncogenic signaling proteins. For example, it can bind the epidermal growth factor receptor (EGFR) and src-family kinases, and it contains an SH2-binding motif that recruits the adaptor Grb2 when a specific tyrosine is phosphorylated (pmc.ncbi.nlm.nih.gov). Through Grb2 and the adaptor Shc, MUC1 can link to the Ras–MAPK pathway (pmc.ncbi.nlm.nih.gov). MUC1 also associates with phosphoinositide 3-kinase (PI3K), likely via the PI3K p85 subunit’s SH2 domain, to modulate the PI3K–AKT survival pathway (pmc.ncbi.nlm.nih.gov). These interactions suggest that when a growth factor stimulus (such as EGF) occurs, MUC1 gets phosphorylated (e.g., by EGFR or Src) and then helps propagate signals that promote cell proliferation or survival (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Indeed, experiments have shown that EGF binding to EGFR leads to MUC1 tail phosphorylation, enabling binding of Grb2 and activation of downstream mitogenic signaling (pubmed.ncbi.nlm.nih.gov). Thus, MUC1 can augment signals from growth factor receptors and integrate into classic pathways like Ras/MAPK and PI3K/AKT.
-
Wnt/β-Catenin Pathway: MUC1’s interaction with β-catenin is a well-documented example of its signaling role. β-Catenin is involved in cell–cell adhesion (at E-cadherin junctions) and in Wnt signaling (as a transcriptional co-activator in the nucleus). MUC1-CT binds β-catenin on the same armadillo repeat region that E-cadherin binds (pmc.ncbi.nlm.nih.gov). In carcinoma cells, overexpressed MUC1 can compete with E-cadherin, sequestering β-catenin away from cell junctions (pmc.ncbi.nlm.nih.gov). The MUC1–β-catenin complex can then translocate to the nucleus. Studies by Yamamoto et al. and others found that MUC1’s tail, once phosphorylated, forms a complex with β-catenin and potentiates β-catenin’s ability to activate Wnt target genes, such as cyclin D1 and c-Myc, promoting cell cycle progression (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). This positions MUC1 as an influencer of the Wnt pathway, particularly in oncogenic contexts. Consistent with this, MUC1-C has been shown to drive EMT and invasiveness via Wnt/β-catenin signaling in cancer models (pmc.ncbi.nlm.nih.gov).
-
NF-κB and Inflammation Signaling: The MUC1 cytoplasmic tail has been reported to interact with the NF-κB pathway. In inflammatory environments, MUC1-C can translocate to the nucleus and bind the p65 subunit of NF-κB, a transcription factor controlling many immune and survival genes (pmc.ncbi.nlm.nih.gov). One study in prostate cancer found MUC1-C/p65 complexes on chromatin, leading to increased expression of genes like ZEB1 and EZH2 that drive EMT and stemness (pmc.ncbi.nlm.nih.gov). By serving as a co-factor for NF-κB, MUC1 may amplify or sustain the transcription of specific target genes. In general, MUC1’s ability to modulate NF-κB and other transcription factors links it to pathways of inflammation, apoptosis, and cell survival.
-
Intrinsic Apoptotic Pathways: Emerging evidence suggests MUC1-C can localize to mitochondria and affect apoptosis. The C-terminal subunit has been detected on the outer mitochondrial membrane in some cancer cells, where it interferes with pro-apoptotic signaling. For instance, MUC1 has been reported to block the release of mitochondrial apoptogenic factors (like cytochrome c) upon drug treatment, thereby conferring chemotherapy resistance by inhibiting apoptosis (www.frontiersin.org) (www.frontiersin.org). This is not a classical signaling pathway, but it illustrates MUC1’s multifunctional role in cell survival mechanisms.
-
Other interactions: MUC1-CT binds to various other molecules: it can associate with GSK3β (a kinase in Wnt and other pathways), Protein kinase Cδ, and elements of T-cell signaling like ZAP-70 and Lck in immune cells (pubmed.ncbi.nlm.nih.gov). Many of these interactions depend on specific phosphorylation of MUC1 tyrosines. For example, phosphorylation on distinct tyrosines creates binding sites for either β-catenin or PI3K or Src-homology domains, allowing MUC1 to act as a platform for signaling complexes (pubmed.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). The CQC motif at the very end of the cytoplasmic tail is required for MUC1’s proper localization and perhaps its dimerization; mutation of this CQC motif disrupts MUC1 targeting to the membrane and attenuates its signaling functions (pubmed.ncbi.nlm.nih.gov). This motif includes cysteine residues that are palmitoylated, anchoring MUC1-C in lipid rafts at the plasma membrane (pubmed.ncbi.nlm.nih.gov). Loss of these modifications can send MUC1 to other cellular locations and alter signal outputs.
Overall, through its cytoplasmic tail, MUC1 integrates into multiple signaling networks. Under normal conditions, these interactions may help fine-tune epithelial responses to growth factors, stress, or inflammatory signals. In disease states (especially cancer), the same signaling roles of MUC1 are often co-opted to promote unchecked proliferation, survival, and metastasis.
Modulation of Inflammation and Immune Response
MUC1 plays a significant role in regulating immune responses at mucosal surfaces. Not only does it act as a physical barrier to pathogens, it also functions as a negative regulator of pathogen-induced signaling, preventing excessive inflammation. The cytoplasmic tail of MUC1 has been shown to interact with pattern recognition receptor signaling, especially Toll-like receptors (TLRs) on immune and epithelial cells (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). When a pathogen is detected, TLRs typically activate NF-κB and other pathways to induce pro-inflammatory cytokines. MUC1 is unusual in that it is upregulated by inflammatory stimuli (such as tumor necrosis factor alpha, TNF-α, or pathogen components) and then acts in a feedback manner to dampen the inflammatory response (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov).
Several studies illustrate MUC1’s anti-inflammatory role: for example, binding of Pseudomonas flagellin to cell-surface MUC1 triggers phosphorylation of the MUC1-CT (via EGFR), allowing the MUC1 tail to associate with TLR5 and block MyD88 recruitment (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). MyD88 is the adaptor protein needed for downstream TLR signaling, so MUC1 essentially competitively inhibits TLR signaling in this context (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Kato et al. (2012) demonstrated that in airway epithelial cells, this mechanism reduced activation of the NF-κB pathway and lowered production of IL-8 and TNF-α during P. aeruginosa infection (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Similarly, MUC1-CT can interact with TLR3 (the receptor for viral double-stranded RNA) and prevent the adapter TRIF from binding, thereby suppressing the type I interferon response and cell death triggered by viral RNA sensing (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). This was shown using synthetic dsRNA (poly I:C) in lung epithelial models, where wild-type cells (with MUC1) had milder cytokine responses and apoptosis compared to cells lacking the MUC1 tail (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov).
Broadly, MUC1 seems to serve as a universal attenuator of TLR signaling. Ueno et al. (2008) tested various TLR agonists (for TLR2, 3, 4, 7, 9) and found that the presence of MUC1-CT was required to limit the inflammatory response to each of these stimuli (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In macrophages or monocytes, which also express MUC1, the MUC1-CT similarly reduces pro-inflammatory cytokine release upon challenge (pmc.ncbi.nlm.nih.gov). There is evidence linking MUC1’s anti-inflammatory effects with induction of IL-10 (an anti-inflammatory cytokine) and interferons, suggesting MUC1 might skew responses toward resolution of inflammation (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Furthermore, because TLR-driven inflammation is a prerequisite for inflammasome activation, MUC1’s braking effect on TLRs also means it can indirectly suppress inflammasome pathways (reducing maturation of IL-1β and IL-18) (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). This helps prevent overactivation of immune responses that could damage tissue.
In essence, MUC1 provides a check on the innate immune system: it allows initial pathogen sensing, but as infection proceeds, increased MUC1 on the cell surface helps prevent an overzealous inflammatory reaction that can harm host tissues (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). This modulator function is beneficial in preventing chronic inflammation and tissue injury. However, it can be a double-edged sword; as noted earlier, in acute infections like P. aeruginosa pneumonia, MUC1’s suppression of inflammation can slow bacterial clearance (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). The overall effect of MUC1 on immunity appears context-dependent – balancing pathogen clearance with prevention of immunopathology. This role is conserved across species (the cytoplasmic tail is homologous from humans to mammals like mice (pmc.ncbi.nlm.nih.gov)), highlighting its importance in immune homeostasis.
Clinical Significance and Applications
MUC1’s distinctive features in normal and disease states have made it a focus of clinical interest, especially in cancer diagnosis and therapy. In healthy tissue, MUC1 is mostly confined to the apical surface of epithelial cells and carries long, complex glycan chains. In many epithelial cancers, MUC1 is overexpressed and abnormally glycosylated, and it loses its polarized distribution (www.frontiersin.org). Over 90% of human breast carcinomas overexpress MUC1, often to a very high level (www.frontiersin.org). Similar overexpression is observed in other adenocarcinomas (e.g. ovarian, lung, pancreatic, prostate), where MUC1 is found across the entire cell surface and even in circulation as shed fragments. Tumor-associated MUC1 typically has shorter glycans (due to altered glycosyltransferase activity in cancers), exposing cryptic peptide epitopes and carbohydrate antigens (such as the Tn, sTn, and T antigen clusters) (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). These tumor-specific epitopes make MUC1 a useful biomarker and an immunotherapeutic target. For instance, the serum CA15-3 test for breast cancer monitors circulating MUC1 fragments carrying the cancer-associated sialyl-T antigen. An abnormally high level of MUC1 in patient serum can indicate tumor burden and is used in monitoring breast cancer progression or recurrence (pmc.ncbi.nlm.nih.gov).
Clinically, MUC1 expression correlates with disease outcomes. Abnormally high MUC1 levels are associated with poor prognosis in multiple cancers, including breast, lung, pancreatic, renal, and others (pmc.ncbi.nlm.nih.gov). The presence of MUC1 contributes to tumor progression by enhancing proliferation, invasion, and immune evasion (www.frontiersin.org) (www.frontiersin.org). MUC1 functions as an oncoprotein in these settings – for example, MUC1 can drive cancer cell growth by activating pro-tumorigenic pathways (Wnt/β-catenin, NF-κB, etc.) and conferring resistance to apoptosis and therapy (www.frontiersin.org) (www.frontiersin.org). Due to this central role, MUC1 was ranked the second most promising cancer antigen (out of 75) by an NIH-led consortium for developing cancer immunotherapies (pmc.ncbi.nlm.nih.gov). This has spurred numerous efforts to target MUC1 in cancer treatment.
Cancer vaccines targeting MUC1 are one active area of research. MUC1’s immunogenic VNTR domain (with repetitive peptide epitopes) can be used to raise antibodies and T-cells. In fact, MUC1 was one of the first tumor antigens tested in vaccine trials. Various vaccine formulations – from synthetic MUC1 peptides conjugated to carriers, to dendritic cells loaded with MUC1 antigen, to viral vector vaccines encoding MUC1 – have been developed (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). As an example, TG4010 is a modified vaccinia Ankara (MVA) virus vaccine that delivers the human MUC1 gene plus IL-2; it has been tested in phase I/II trials for cancers like non-small cell lung cancer and showed a favorable safety profile (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). In a clinical study for metastatic renal cancer, an RNA vaccine including MUC1 (among other tumor antigens) induced MUC1-specific T-cell responses in patients, with some experiencing stable disease or partial remission (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Another approach used autologous dendritic cells pulsed with a MUC1 peptide (containing a helper T-cell epitope PADRE) in renal carcinoma, resulting in cytotoxic T-cells that could kill MUC1-expressing tumor cells and minor tumor regressions in a subset of patients (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). These early-phase trials affirm that MUC1 vaccines can safely generate an immune response. More recently, liposomal MUC1 glycopeptide vaccines (like L-BLP25) were tested in lung cancer, and MUC1 peptide conjugates are under investigation for breast and prostate cancer immunoprevention (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov).
Antibody-based therapies targeting MUC1 are also being explored. Monoclonal antibodies that recognize tumor-specific MUC1 epitopes (for example, the antibody TAB004 that binds a cancer-specific glycoform of MUC1) have shown promising preclinical results – binding to tumor cells and inducing their destruction (www.frontiersin.org). MUC1-directed antibody–drug conjugates (ADCs) are in development, in which an anti-MUC1 antibody delivers a cytotoxic drug to MUC1-expressing tumor cells (pmc.ncbi.nlm.nih.gov). There is also interest in CAR T-cell therapy against MUC1: chimeric antigen receptor T-cells engineered to attack MUC1-positive cancers. Given the widespread expression of MUC1 in tumors, several CAR T constructs have been designed. For instance, a fully human CAR T (designated P-MUC1C-ALLO1) targeting the MUC1-C core is currently in a phase I trial for solid tumors (pmc.ncbi.nlm.nih.gov). Early preclinical data suggest that MUC1-specific CAR T cells can selectively kill carcinoma cells while sparing normal cells that have MUC1 mostly sequestered on the apical side or with different glycosylation.
Beyond cancer, understanding MUC1’s function has implications in other diseases. A striking example is MUC1-associated familial kidney disease: certain mutations in MUC1 (frameshifts in the VNTR region) cause a misfolded protein that accumulates in kidney tubule cells, leading to medullary cystic kidney disease type 1 (pmc.ncbi.nlm.nih.gov). This illustrates that proper MUC1 processing is important for cell health. Additionally, because MUC1 modulates inflammation, it is being studied in chronic inflammatory diseases of mucosal organs. Variants in MUC1 have been examined for associations with inflammatory bowel disease and respiratory conditions, though these areas are still emerging.
In summary, MUC1’s primary functions are to serve as a protective mucosal barrier and a modulator of signaling in epithelial cells. It carries out these roles at the cell surface (providing a shield and interacting with pathogens) and at the intracellular level (transducing signals via its cytoplasmic tail). In normal physiology, MUC1 protects tissues from infection and regulates inflammation, while maintaining epithelial integrity. In pathological states like cancer, MUC1 is co-opted to promote tumor growth and metastasis. Its ubiquitous overexpression in carcinomas and distinct tumor-associated forms have made it both a diagnostic marker and a target for immunotherapy (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov). Ongoing research (as of 2023–2024) continues to uncover new aspects of MUC1 biology – such as its role in immune evasion and epigenetic regulation in cancer (www.frontiersin.org) (www.frontiersin.org) – and to harness this knowledge in developing MUC1-targeted treatments to improve patient outcomes.
References:
-
Chen X. et al. (2024). MUC1 and MUC16: critical for immune modulation in cancer therapeutics. Front. Immunol. 15:1356913. DOI:10.3389/fimmu.2024.1356913 (www.frontiersin.org) (www.frontiersin.org)
-
Dhar P, McAuley J. (2019). The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation. Front. Cell. Infect. Microbiol. 9:117. DOI:10.3389/fcimb.2019.00117 (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov)
-
Long L. et al. (2023). The research status and prospects of MUC1 in immunology. Hum. Vaccin. Immunother. 19(1):2172278. DOI:10.1080/21645515.2023.2172278 (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov)
-
Mao W. et al. (2024). Research progress of MUC1 in genitourinary cancers. Cell. Mol. Biol. Lett. 29:135. DOI:10.1186/s11658-024-00654-x (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov)
-
McAuley JL. et al. (2007). MUC1 cell surface mucin is a critical element of the mucosal barrier to infection. J. Clin. Invest. 117(8):2313–2324. DOI:10.1172/JCI26705 (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov)
-
Kato K. et al. (2012). MUC1: The first mammalian membrane-tethered mucin to be implicated in tumor suppression. Mucosal Immunol. 5(3):309–320. DOI:10.1038/mi.2012.9 (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov)
-
Gnemmi V. et al. (2014). MUC1-C nuclear localization drives invasiveness of renal cancer cells through a sheddase/gamma-secretase-dependent pathway. Oncotarget 5(3):754–765. DOI:10.18632/oncotarget.1776 (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov)
-
Leng Y. et al. (2007). Nucleocytoplasmic shuttling of MUC1 cytoplasmic tail: a novel mechanism for signal transduction and transcriptional regulation. Biochem. Biophys. Res. Commun. 360(1):145–150. DOI:10.1016/j.bbrc.2007.06.049 (pubmed.ncbi.nlm.nih.gov)
-
Ueno K. et al. (2008). MUC1 mucin is a negative regulator of TLR signaling. Am. J. Respir. Cell Mol. Biol. 38(3):263–268. DOI:10.1165/rcmb.2007-0078OC (pmc.ncbi.nlm.nih.gov) (pmc.ncbi.nlm.nih.gov)
-
Yamamoto M. et al. (1997). MUC1 cytoplasmic domain mediates binding to beta-catenin. J. Biol. Chem. 272(16):12492–12494. DOI:10.1074/jbc.272.16.12492 (pmc.ncbi.nlm.nih.gov)
Citations
- AnnotationURLCitation(end_index=416, start_index=296, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=to%20the%20apical%20surface%20of,14')
- AnnotationURLCitation(end_index=818, start_index=701, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=MUC1%20,The%20molecular%20weight')
- AnnotationURLCitation(end_index=1022, start_index=905, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=MUC1%20,The%20molecular%20weight')
- AnnotationURLCitation(end_index=1258, start_index=1134, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=MUC1%2C%20which%20is%20localized%20to,C')
- AnnotationURLCitation(end_index=1620, start_index=1459, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=%28A%29%20Schematic%20representation%20of%20MUC1,region%20and%20can%20undergo')
- AnnotationURLCitation(end_index=1791, start_index=1621, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=region%20in%20the%20extracellular%20polypeptide,capable%20of%20inducing%20release%20of')
- AnnotationURLCitation(end_index=2106, start_index=1950, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/31069176/#:~:text=%28A%29%20Schematic%20representation%20of%20MUC1,region%20and%20can%20undergo')
- AnnotationURLCitation(end_index=2248, start_index=2107, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/31069176/#:~:text=amino%20acids%20in%20length%2C%20tethers,kinase%20%28Kato%20et')
- AnnotationURLCitation(end_index=2637, start_index=2467, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=region%20in%20the%20extracellular%20polypeptide,capable%20of%20inducing%20release%20of')
- AnnotationURLCitation(end_index=2800, start_index=2638, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=which%20possess%20the%20GSVVV%20motif%2C,lumen%20provides%20a%20mechanism%20by')
- AnnotationURLCitation(end_index=3071, start_index=2952, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=The%20cytoplasmic%20tail%20%28,2000')
- AnnotationURLCitation(end_index=3406, start_index=3248, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/31069176/#:~:text=the%20cell.%20The%20MUC1,GSK3%CE%B2%2C%20glycogen%20synthase%20kinase%203%CE%B2')
- AnnotationURLCitation(end_index=3517, start_index=3407, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=,%CE%B4%20%28Ren%20et%20al')
- AnnotationURLCitation(end_index=3867, start_index=3737, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=MUC1%20is%20a%20polymorphic%20transmembrane,1')
- AnnotationURLCitation(end_index=4005, start_index=3868, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=determines%20the%20speciffic%20spatial%20structure,8')
- AnnotationURLCitation(end_index=4376, start_index=4254, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=Expression%20of%20MUC1%20is%20found,4')
- AnnotationURLCitation(end_index=4673, start_index=4537, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=MUC16%2C%20MUC17%2C%20MUC21%2C%20and%20MUC22%29.,12')
- AnnotationURLCitation(end_index=4794, start_index=4674, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=to%20the%20apical%20surface%20of,14')
- AnnotationURLCitation(end_index=4994, start_index=4872, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=Expression%20of%20MUC1%20is%20found,4')
- AnnotationURLCitation(end_index=5892, start_index=5754, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=Ligtenberg%20et%20al,2004%3B%20Thathiah%20and%20Carson')
- AnnotationURLCitation(end_index=6002, start_index=5893, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=TNF,Lind%C3%A9n%20et%20al')
- AnnotationURLCitation(end_index=6346, start_index=6247, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=2003%20%29,2000')
- AnnotationURLCitation(end_index=6654, start_index=6503, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=have%20also%20found%20that%20MUC1,was%20induced%20by%20a%20hypoxia')
- AnnotationURLCitation(end_index=6808, start_index=6655, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=demonstrated%20that%20MUC1,migratory%20properties%20of%20RCC%20cells')
- AnnotationURLCitation(end_index=7120, start_index=6969, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=have%20also%20found%20that%20MUC1,was%20induced%20by%20a%20hypoxia')
- AnnotationURLCitation(end_index=7258, start_index=7121, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=directly%20bind%20to%20nuclear%20factor,150%2C%20151')
- AnnotationURLCitation(end_index=7950, start_index=7846, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=glycoproteins%20,11')
- AnnotationURLCitation(end_index=8113, start_index=7951, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=Mucin%201%20,mesenchymal%20transition')
- AnnotationURLCitation(end_index=8557, start_index=8461, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=Introduction')
- AnnotationURLCitation(end_index=8646, start_index=8558, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=al,9')
- AnnotationURLCitation(end_index=8950, start_index=8791, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=The%20family%20of%20cell%20surface,may%20play%20an%20important%20biological')
- AnnotationURLCitation(end_index=9105, start_index=8951, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=Similar%20to%20all%20of%20the,The%20extracellular%20region%20that%20is')
- AnnotationURLCitation(end_index=9583, start_index=9424, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/31069176/#:~:text=surface%20by%20nearly%20all%20epithelial,most%20extensively%20studied%20of%20the')
- AnnotationURLCitation(end_index=9745, start_index=9584, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/31069176/#:~:text=environment%2C%20MUC1%20has%20been%20demonstrated,mucin%20regulates%20inflammation')
- AnnotationURLCitation(end_index=10082, start_index=9920, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=which%20possess%20the%20GSVVV%20motif%2C,lumen%20provides%20a%20mechanism%20by')
- AnnotationURLCitation(end_index=10193, start_index=10083, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=%28Thathiah%20et%20al,2009')
- AnnotationURLCitation(end_index=10544, start_index=10426, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=2003%20%3B%20Vinall%20et%20al,2007')
- AnnotationURLCitation(end_index=10667, start_index=10545, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=triggered%20shedding%20via%20self,2016')
- AnnotationURLCitation(end_index=11175, start_index=11027, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=Campylobacter%20jejuni%20is%20an%20intestinal,Palacios%20et%20al')
- AnnotationURLCitation(end_index=11382, start_index=11176, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=gastrointestinal%20and%20systemic%20spread%20of,another%20gastrointestinal%20pathogen%2C%20enteroaggregative%20Escherichia')
- AnnotationURLCitation(end_index=11837, start_index=11726, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=%28Wotherspoon%20et%20al,ED')
- AnnotationURLCitation(end_index=11960, start_index=11838, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=triggered%20shedding%20via%20self,2016')
- AnnotationURLCitation(end_index=12486, start_index=12318, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=Pseudomonas%20aeruginosa%20causes%20respiratory%20infections,aeruiginosa%20infection')
- AnnotationURLCitation(end_index=12936, start_index=12750, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=immunocompromised%20individuals%2C%20such%20as%20those,inflammatory%20responses%20to%20the%20infection')
- AnnotationURLCitation(end_index=13228, start_index=13042, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=immunocompromised%20individuals%2C%20such%20as%20those,inflammatory%20responses%20to%20the%20infection')
- AnnotationURLCitation(end_index=13605, start_index=13437, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=Pseudomonas%20aeruginosa%20causes%20respiratory%20infections,aeruiginosa%20infection')
- AnnotationURLCitation(end_index=13770, start_index=13606, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=barrier%20to%20infection%20in%20the,engagement%20of%20NEU1%20desialylates%20MUC1')
- AnnotationURLCitation(end_index=14432, start_index=14264, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=MUC1%20is%20a%20highly%20glycosylated,cancers%20and%20its%20clinical%20applications')
- AnnotationURLCitation(end_index=15149, start_index=14981, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=MUC1%20is%20a%20highly%20glycosylated,cancers%20and%20its%20clinical%20applications')
- AnnotationURLCitation(end_index=15303, start_index=15150, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=As%20a%20member%20of%20membrane,studies%20have%20shown%20that%20MUC1')
- AnnotationURLCitation(end_index=15995, start_index=15828, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=As%20a%20member%20of%20membrane,attractive%20target%20for%20cancer%20immunotherapy')
- AnnotationURLCitation(end_index=16231, start_index=15996, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=tumorigenesis%20and%20progression%2C%20significant%20changes,activating%20pluripotency%20networks%20and%20also')
- AnnotationURLCitation(end_index=16674, start_index=16439, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=tumorigenesis%20and%20progression%2C%20significant%20changes,activating%20pluripotency%20networks%20and%20also')
- AnnotationURLCitation(end_index=16901, start_index=16675, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=interactions%20with%20cell%20surface%20receptors,monoclonal%20antibody%20TAB004%20overcomes%20anoikis')
- AnnotationURLCitation(end_index=17644, start_index=17534, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=,%CE%B4%20%28Ren%20et%20al')
- AnnotationURLCitation(end_index=18131, start_index=17977, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=formation%20of%20an%20immune,that%20MUC1%20was%20directly%20regulated')
- AnnotationURLCitation(end_index=18424, start_index=18270, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=formation%20of%20an%20immune,that%20MUC1%20was%20directly%20regulated')
- AnnotationURLCitation(end_index=18926, start_index=18722, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=extracellular%20domain%2C%20a%2028,cancer%20cases%20exhibiting%20its%20abnormal')
- AnnotationURLCitation(end_index=19109, start_index=18927, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=MUC1%20serves%20as%20a%20protective,C%20participates%20in')
- AnnotationURLCitation(end_index=19417, start_index=19269, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=have%20also%20found%20that%20MUC1,a%20crucial%20role%20in%20RCC')
- AnnotationURLCitation(end_index=20148, start_index=19990, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/31069176/#:~:text=the%20cell.%20The%20MUC1,GSK3%CE%B2%2C%20glycogen%20synthase%20kinase%203%CE%B2')
- AnnotationURLCitation(end_index=20259, start_index=20149, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=,%CE%B4%20%28Ren%20et%20al')
- AnnotationURLCitation(end_index=20801, start_index=20694, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=2001a%20%29%3B%20PLC,29')
- AnnotationURLCitation(end_index=21021, start_index=20879, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=Blue%20bars%20mark%20the%20known,%CE%B4%20%28Ren%20et%20al')
- AnnotationURLCitation(end_index=21317, start_index=21175, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=Blue%20bars%20mark%20the%20known,%CE%B4%20%28Ren%20et%20al')
- AnnotationURLCitation(end_index=21691, start_index=21528, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=suppresses%20TLR%20signaling%2C%20attenuating%20inflammation,CT%2C%20increasing')
- AnnotationURLCitation(end_index=21811, start_index=21692, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=binding%20of%20P,CT%2C%20increasing')
- AnnotationURLCitation(end_index=22079, start_index=21979, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/31069176/#:~:text=protein%20kinase,2007')
- AnnotationURLCitation(end_index=22664, start_index=22554, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=,%CE%B4%20%28Ren%20et%20al')
- AnnotationURLCitation(end_index=22892, start_index=22782, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=,%CE%B4%20%28Ren%20et%20al')
- AnnotationURLCitation(end_index=23314, start_index=23204, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=,%CE%B4%20%28Ren%20et%20al')
- AnnotationURLCitation(end_index=23469, start_index=23315, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=formation%20of%20an%20immune,that%20MUC1%20was%20directly%20regulated')
- AnnotationURLCitation(end_index=23841, start_index=23687, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=formation%20of%20an%20immune,that%20MUC1%20was%20directly%20regulated')
- AnnotationURLCitation(end_index=24274, start_index=24137, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=directly%20bind%20to%20nuclear%20factor,150%2C%20151')
- AnnotationURLCitation(end_index=24575, start_index=24438, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=directly%20bind%20to%20nuclear%20factor,150%2C%20151')
- AnnotationURLCitation(end_index=25538, start_index=25313, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=proliferation%2C%20EMT%2C%20epigenetic%20reprogramming%2C%20and,also%20indicated%20that%20MUC1%20can')
- AnnotationURLCitation(end_index=25710, start_index=25539, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=promoting%20its%20degradation%20%2817%20%29,25')
- AnnotationURLCitation(end_index=26207, start_index=26062, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/31069176/#:~:text=Blue%20bars%20mark%20the%20known,the%20CT%20contains%20the%20motif')
- AnnotationURLCitation(end_index=26634, start_index=26476, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/31069176/#:~:text=the%20cell.%20The%20MUC1,GSK3%CE%B2%2C%20glycogen%20synthase%20kinase%203%CE%B2')
- AnnotationURLCitation(end_index=26745, start_index=26635, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=,%CE%B4%20%28Ren%20et%20al')
- AnnotationURLCitation(end_index=27071, start_index=26984, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/31069176/#:~:text=PLC,2007')
- AnnotationURLCitation(end_index=27277, start_index=27190, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/31069176/#:~:text=PLC,2007')
- AnnotationURLCitation(end_index=28400, start_index=28238, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=match%20at%20L259%20MUC1,downstream%20signaling%20cascades%2C%20resulting%20in')
- AnnotationURLCitation(end_index=28527, start_index=28401, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=inflammation,with%20TLRs%20at%20the%20cell')
- AnnotationURLCitation(end_index=29001, start_index=28855, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=match%20at%20L271%20causes%20TLR,with%20TLRs%20at%20the%20cell')
- AnnotationURLCitation(end_index=29167, start_index=29002, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=causes%20TLR%20activation%2C%20causing%20production,with%20TLRs%20at%20the%20cell')
- AnnotationURLCitation(end_index=29591, start_index=29428, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=suppresses%20TLR%20signaling%2C%20attenuating%20inflammation,CT%2C%20increasing')
- AnnotationURLCitation(end_index=29754, start_index=29592, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=match%20at%20L259%20MUC1,downstream%20signaling%20cascades%2C%20resulting%20in')
- AnnotationURLCitation(end_index=30061, start_index=29899, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=match%20at%20L259%20MUC1,downstream%20signaling%20cascades%2C%20resulting%20in')
- AnnotationURLCitation(end_index=30204, start_index=30062, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=MUC1,downstream%20signaling%20cascades%2C%20resulting%20in')
- AnnotationURLCitation(end_index=30519, start_index=30400, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=binding%20of%20P,CT%2C%20increasing')
- AnnotationURLCitation(end_index=30662, start_index=30520, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=MUC1,downstream%20signaling%20cascades%2C%20resulting%20in')
- AnnotationURLCitation(end_index=31058, start_index=30895, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=Similar%20to%20its%20suppressive%20role,lung%20and%20human%20embryonic%20kidney')
- AnnotationURLCitation(end_index=31163, start_index=31059, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=al,Adding%20credence')
- AnnotationURLCitation(end_index=31521, start_index=31358, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=Similar%20to%20its%20suppressive%20role,lung%20and%20human%20embryonic%20kidney')
- AnnotationURLCitation(end_index=31626, start_index=31522, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=al,Adding%20credence')
- AnnotationURLCitation(end_index=32035, start_index=31889, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=to%20the%20notion%20that%20MUC1,CT%20is%20required%20for%20the')
- AnnotationURLCitation(end_index=32145, start_index=32036, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=anti,IFN%29%20correlating')
- AnnotationURLCitation(end_index=32411, start_index=32280, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=including%20Pam_,CT%20is%20required%20for%20the')
- AnnotationURLCitation(end_index=32729, start_index=32620, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=anti,IFN%29%20correlating')
- AnnotationURLCitation(end_index=32873, start_index=32730, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=Kato%20et%20al,role%20in%20controlling%20disease%20severity')
- AnnotationURLCitation(end_index=33261, start_index=33102, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=formation%20of%20the%20inflammasome%20complex,as%20the%20NLRP%2C%20or%20the')
- AnnotationURLCitation(end_index=33375, start_index=33262, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=%28Figure%202B%29,Figure%202C')
- AnnotationURLCitation(end_index=33847, start_index=33701, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=match%20at%20L271%20causes%20TLR,with%20TLRs%20at%20the%20cell')
- AnnotationURLCitation(end_index=34013, start_index=33848, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=causes%20TLR%20activation%2C%20causing%20production,with%20TLRs%20at%20the%20cell')
- AnnotationURLCitation(end_index=34468, start_index=34282, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=immunocompromised%20individuals%2C%20such%20as%20those,inflammatory%20responses%20to%20the%20infection')
- AnnotationURLCitation(end_index=34633, start_index=34469, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=barrier%20to%20infection%20in%20the,engagement%20of%20NEU1%20desialylates%20MUC1')
- AnnotationURLCitation(end_index=34993, start_index=34874, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=The%20cytoplasmic%20tail%20%28,2000')
- AnnotationURLCitation(end_index=35726, start_index=35491, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=tumorigenesis%20and%20progression%2C%20significant%20changes,activating%20pluripotency%20networks%20and%20also')
- AnnotationURLCitation(end_index=36048, start_index=35813, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=tumorigenesis%20and%20progression%2C%20significant%20changes,activating%20pluripotency%20networks%20and%20also')
- AnnotationURLCitation(end_index=36641, start_index=36468, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=In%20cancer%20cells%2C%20the%20long,Friedenreich%2C%20CD176%2C%20T%20antigen%29%2C%20and')
- AnnotationURLCitation(end_index=36746, start_index=36642, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=2%2C3,10%E2%80%9312')
- AnnotationURLCitation(end_index=37255, start_index=37129, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=of%20cancer%20cells%20and%20mediating,RCC')
- AnnotationURLCitation(end_index=37603, start_index=37463, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=2%2C3,pancreatic%20ductal%20carcinoma%2C%20and%20thymic')
- AnnotationURLCitation(end_index=37944, start_index=37716, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=ubiquitously%20expressed%20in%20mammalian%20epithelial,MUC%20proteins%20have%20attracted%20considerable')
- AnnotationURLCitation(end_index=38180, start_index=37945, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=tumorigenesis%20and%20progression%2C%20significant%20changes,activating%20pluripotency%20networks%20and%20also')
- AnnotationURLCitation(end_index=38633, start_index=38405, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=ubiquitously%20expressed%20in%20mammalian%20epithelial,MUC%20proteins%20have%20attracted%20considerable')
- AnnotationURLCitation(end_index=38859, start_index=38634, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=proliferation%2C%20EMT%2C%20epigenetic%20reprogramming%2C%20and,also%20indicated%20that%20MUC1%20can')
- AnnotationURLCitation(end_index=39170, start_index=39023, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=strong%20antitumor%20effects,in%20cancer%20research%20by%20the')
- AnnotationURLCitation(end_index=39807, start_index=39688, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=Bladder%20cancer%20%20,NCT03540199')
- AnnotationURLCitation(end_index=39984, start_index=39808, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=In%20recent%20years%2C%20new%20discoveries,manner%2C%20with%20objective%20remission%20being')
- AnnotationURLCitation(end_index=40374, start_index=40233, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=TG4010%20is%20a%20cancer%20vaccine,In%20metastatic%20RCC')
- AnnotationURLCitation(end_index=40543, start_index=40375, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=telomerase%2C%20survivin%2C%20and%20MAGE,like%20receptor%209%20%28TLR9%29%20agonist')
- AnnotationURLCitation(end_index=40913, start_index=40764, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=detected%20in%20the%2011%20patients,133%5D%20demonstrated%20that')
- AnnotationURLCitation(end_index=41045, start_index=40914, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=CD4,like%20receptor%209%20%28TLR9%29%20agonist')
- AnnotationURLCitation(end_index=41490, start_index=41313, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=development%20of%20research%20strategies%20targeting,patients%20with%20metastatic%20RCC%20to')
- AnnotationURLCitation(end_index=41648, start_index=41491, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=%28DC,A1%29%20using%20adjuvant%20granulocyte%E2%80%93macrophage%20colony')
- AnnotationURLCitation(end_index=42063, start_index=41944, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=Bladder%20cancer%20%20,NCT03540199')
- AnnotationURLCitation(end_index=42170, start_index=42064, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=%7C%20CAR,NCT00852007')
- AnnotationURLCitation(end_index=42700, start_index=42491, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=stemness%20of%20cancer%20cells%2C%20primarily,As%20an%20oncogenic%20driver%2C%20MUC1')
- AnnotationURLCitation(end_index=43006, start_index=42856, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=tandem%20repeats%20%28VNTRs%29.,in%20cancer%20research%20by%20the')
- AnnotationURLCitation(end_index=43497, start_index=43381, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=%7C%20TG4010%20%20%7C%20MUC1,22')
- AnnotationURLCitation(end_index=44141, start_index=44037, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=glycoproteins%20,11')
- AnnotationURLCitation(end_index=45368, start_index=45205, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=participates%20in%20epithelial%20cell%20renewal,This%20article%20reviews%20the')
- AnnotationURLCitation(end_index=45495, start_index=45369, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=of%20cancer%20cells%20and%20mediating,RCC')
- AnnotationURLCitation(end_index=45889, start_index=45654, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=tumorigenesis%20and%20progression%2C%20significant%20changes,activating%20pluripotency%20networks%20and%20also')
- AnnotationURLCitation(end_index=46038, start_index=45890, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=associated%20MUC1%20,25')
- AnnotationURLCitation(end_index=46488, start_index=46326, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=Mucin%201%20,mesenchymal%20transition')
- AnnotationURLCitation(end_index=46724, start_index=46489, title='Frontiers | MUC1 and MUC16: critical for immune modulation in cancer therapeutics', type='url_citation', url='https://www.frontiersin.org/journals/plant-science/articles/10.3389/fimmu.2024.1356913/full#:~:text=tumorigenesis%20and%20progression%2C%20significant%20changes,activating%20pluripotency%20networks%20and%20also')
- AnnotationURLCitation(end_index=47088, start_index=46927, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=%28A%29%20Schematic%20representation%20of%20MUC1,region%20and%20can%20undergo')
- AnnotationURLCitation(end_index=47251, start_index=47089, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=which%20possess%20the%20GSVVV%20motif%2C,lumen%20provides%20a%20mechanism%20by')
- AnnotationURLCitation(end_index=47535, start_index=47418, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=MUC1%20,The%20molecular%20weight')
- AnnotationURLCitation(end_index=47656, start_index=47536, title='The research status and prospects of MUC1 in immunology - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC10012890/#:~:text=to%20the%20apical%20surface%20of,14')
- AnnotationURLCitation(end_index=47973, start_index=47805, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=MUC1%20is%20a%20highly%20glycosylated,cancers%20and%20its%20clinical%20applications')
- AnnotationURLCitation(end_index=48141, start_index=47974, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=As%20a%20member%20of%20membrane,attractive%20target%20for%20cancer%20immunotherapy')
- AnnotationURLCitation(end_index=48468, start_index=48320, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=Campylobacter%20jejuni%20is%20an%20intestinal,Palacios%20et%20al')
- AnnotationURLCitation(end_index=48591, start_index=48469, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=triggered%20shedding%20via%20self,2016')
- AnnotationURLCitation(end_index=48931, start_index=48769, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=match%20at%20L259%20MUC1,downstream%20signaling%20cascades%2C%20resulting%20in')
- AnnotationURLCitation(end_index=49074, start_index=48932, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=MUC1,downstream%20signaling%20cascades%2C%20resulting%20in')
- AnnotationURLCitation(end_index=49441, start_index=49290, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=have%20also%20found%20that%20MUC1,was%20induced%20by%20a%20hypoxia')
- AnnotationURLCitation(end_index=49595, start_index=49442, title='Research progress of MUC1 in genitourinary cancers - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC11533421/#:~:text=demonstrated%20that%20MUC1,migratory%20properties%20of%20RCC%20cells')
- AnnotationURLCitation(end_index=49924, start_index=49837, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PubMed', type='url_citation', url='https://pubmed.ncbi.nlm.nih.gov/31069176/#:~:text=PLC,2007')
- AnnotationURLCitation(end_index=50234, start_index=50088, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=to%20the%20notion%20that%20MUC1,CT%20is%20required%20for%20the')
- AnnotationURLCitation(end_index=50344, start_index=50235, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=anti,IFN%29%20correlating')
- AnnotationURLCitation(end_index=50618, start_index=50508, title='The Role of the Cell Surface Mucin MUC1 as a Barrier to Infection and Regulator of Inflammation - PMC', type='url_citation', url='https://pmc.ncbi.nlm.nih.gov/articles/PMC6491460/#:~:text=,%CE%B4%20%28Ren%20et%20al')
Perplexity
(MUC1-deep-research-perplexity-lite.md)
Perplexity
(MUC1-deep-research-perplexity-lite.md)The MUC1 (P15941) gene encodes the mucin-1 protein, a heavily glycosylated, membrane-bound mucin that plays critical roles in epithelial protection, cell signaling, and disease, particularly cancer and kidney disorders[1][3][5][7].
1. Key Concepts and Definitions
- MUC1 (Mucin-1): A type I transmembrane glycoprotein found on the apical surface of most simple epithelial cells, including those in the lung, breast, stomach, pancreas, kidney, and reproductive tract[3][10][15].
- Protein Structure: MUC1 is synthesized as a single polypeptide and then cleaved into two subunits:
- Alpha (N-terminal) subunit: Extracellular, contains a variable number tandem repeat (VNTR) domain rich in serine, threonine, and proline, which is heavily O-glycosylated[1][5][7].
- Beta (C-terminal) subunit: Contains the transmembrane and cytoplasmic tail domains, involved in intracellular signaling[5][7].
2. Molecular Function
- Barrier Formation: MUC1 forms a protective mucous barrier on epithelial surfaces, shielding cells from pathogens, enzymes, and physical damage[1][3][5].
- Cell Adhesion and Anti-Adhesion: The alpha subunit mediates cell adhesion, while the glycosylated extracellular domain can also inhibit cell-cell and cell-matrix interactions, affecting tissue organization and tumor metastasis[3][5][7].
- Cell Signaling: The cytoplasmic tail (MUC1-CT) transduces signals from the extracellular environment to the nucleus, influencing cell proliferation, survival, motility, and gene expression[1][5][7].
- MUC1-CT interacts with signaling pathways such as ERK, SRC, NF-κB, and the Ras/MAPK pathway[5].
- It modulates p53-mediated transcription, affecting cell cycle arrest and apoptosis[2][5][13].
3. Biological Processes
- Epithelial Homeostasis: Maintains epithelial integrity, regulates cell shedding, and participates in wound repair and epithelial-mesenchymal transition (EMT)[3].
- Immune Regulation: Inhibits immune cell interactions with epithelial cells, modulating inflammatory responses and immune evasion in cancer[3][14].
- Pathogen Defense: Binds pathogens via its glycosylated extracellular domain, preventing their access to the cell surface[1][2][14].
- Kidney Development: Plays a role in nephron development and kidney function[1][5].
4. Cellular Localization
- Apical Membrane: Predominantly localized to the apical surface of epithelial cells in various tissues (lung, breast, stomach, pancreas, kidney, reproductive tract)[3][15].
- Nucleus and Mitochondria: The cytoplasmic tail can translocate to the nucleus (regulating gene expression) or mitochondria (inhibiting apoptosis)[1][2].
- Endocytosis and Recycling: MUC1 is internalized, re-glycosylated, and recycled to the cell membrane[3].
5. Protein Domains
- VNTR Domain: Highly polymorphic, with 20–100 tandem repeats, each 20 amino acids long, rich in serine and threonine for O-glycosylation[1][5][7].
- SEA Domain: Involved in protein cleavage and cell signaling, contributes to immune inhibition and cell shedding[3].
- Transmembrane Domain: Anchors the protein in the cell membrane[7].
- Cytoplasmic Tail (MUC1-CT): Contains phosphorylation sites and motifs for protein-protein interactions, critical for signal transduction[1][5][7].
6. Known Interactions
- Protein-Protein Interactions:
- p53: MUC1-CT binds p53, modulating its transcriptional activity and influencing cell cycle arrest and apoptosis[2][5][13].
- ERBB2 (HER2): Interacts with the receptor tyrosine kinase, affecting growth signaling[2].
- GRB2, SOS1, CTNND1, JUP: Involved in various signaling and adhesion pathways[2].
- KLF4: Co-binds to the TP53 promoter, repressing p53 activity[5].
- Bcl-2, Bcl-xL: MUC1-CT interaction prevents apoptosis by inhibiting cytochrome c release from mitochondria[2].
7. Disease Associations
- Cancer:
- Overexpression and Aberrant Glycosylation: Common in many carcinomas (breast, ovarian, pancreatic, lung, etc.), associated with tumor progression, metastasis, immune evasion, and chemoresistance[2][3][5][6][12].
- Oncogenic Functions: Promotes tumor growth by concentrating growth factors, inhibiting apoptosis, and modulating immune responses[2][12].
- Tumor Marker: MUC1 is used as a biomarker (e.g., CA15-3 in breast cancer)[3].
- Kidney Disease:
- Tubulointerstitial Kidney Disease, Autosomal Dominant 2 (ADTKD-MUC1): Mutations in MUC1 cause this hereditary kidney disorder, leading to progressive renal failure[5][8].
- Renal Cysts and Diabetes Syndrome: Also associated with MUC1 mutations[5].
- Other:
- Infectious Diseases: Altered MUC1 function can affect susceptibility to infections due to its role in mucosal defense[14].
8. Recent Developments and Research (2023–2024)
- Therapeutic Targeting: MUC1, especially its C-terminal subunit (MUC1-C), is being actively investigated as a therapeutic target in cancer, with ongoing research into inhibitors and immunotherapies[9].
- Stem Cell Biology: MUC1* (a cleaved form) is expressed on human embryonic pluripotent stem cells and functions as a growth factor receptor, with implications for cancer stem cell biology[12].
- Gene Regulation: Recent studies further elucidate MUC1’s role in regulating transcription factor complexes and epigenetic reprogramming in cancer and tissue repair[3][12].
9. Relevant Statistics and Data
- Expression: MUC1 is highly expressed in epithelial tissues and overexpressed in >90% of breast carcinomas and many other epithelial cancers[3][6].
- Genetic Variability: The VNTR region is highly polymorphic, contributing to individual differences in MUC1 function and disease susceptibility[1][5][7].
10. Authoritative Sources
- UniProt (P15941): Comprehensive protein information, including structure, function, and interactions[7].
- GeneCards: Summarizes gene function, disease associations, and pathways[5].
- MedlinePlus Genetics: Clinical and functional overview[1].
- NCBI Gene: Genomic context and recent research[4][9].
- Recent Reviews: Frontiers in Cellular and Infection Microbiology (2019)[14], Nature Reviews Cancer (2014)[12].
Key URLs and Publication Dates:
- UniProt: https://www.uniprot.org/uniprotkb/P15941/entry (2024)
- GeneCards: https://www.genecards.org/cgi-bin/carddisp.pl?gene=MUC1 (2024)
- MedlinePlus Genetics: https://medlineplus.gov/genetics/gene/muc1/ (2024)
- NCBI Gene: https://www.ncbi.nlm.nih.gov/gene/4582 (2024)
- Frontiers in Cellular and Infection Microbiology: https://www.frontiersin.org/journals/cellular-and-infection-microbiology/articles/10.3389/fcimb.2019.00117/full (2019)
If you need more detailed information on any specific aspect (e.g., structural biology, therapeutic targeting, or specific disease mechanisms), please specify.
Citations
- https://medlineplus.gov/genetics/gene/muc1/
- https://en.wikipedia.org/wiki/Mucin_short_variant_S1
- https://en.wikipedia.org/wiki/Mucin-1
- https://www.ncbi.nlm.nih.gov/gene/4582
- https://www.genecards.org/cgi-bin/carddisp.pl?gene=MUC1
- https://maayanlab.cloud/Harmonizome/gene/MUC1
- https://www.uniprot.org/uniprotkb/P15941/entry
- https://g2p.broadinstitute.org/gene/MUC1
- https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=4582
- https://pmc.ncbi.nlm.nih.gov/articles/PMC320498/
- https://functionome.geneontology.org/gene/UniProtKB:P15941
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5500204/
- https://go.drugbank.com/polypeptides/P15941
- https://www.frontiersin.org/journals/cellular-and-infection-microbiology/articles/10.3389/fcimb.2019.00117/full
- https://v17.proteinatlas.org/ENSG00000185499-MUC1/tissue
- https://www.uniprot.org/uniprotkb/P15941/publications
Perplexity
(MUC1-deep-research-perplexity.md)
Perplexity
(MUC1-deep-research-perplexity.md)Comprehensive Functional Annotation of MUC1 (Mucin 1, Cell Surface Associated): Structure, Function, and Signaling in Normal and Malignant Contexts
The transmembrane glycoprotein MUC1 (mucin 1, cell surface associated) represents a multifunctional molecular entity that undergoes dramatic functional reprogramming during the transition from normal epithelial homeostasis to malignant transformation. In healthy epithelial tissues, MUC1 functions as a protective barrier contributing to lubrication, hydration, and defense against pathogenic invasion through its extended extracellular domain and regulative effects on cellular adhesion.[1][3] However, following neoplastic transformation, MUC1 undergoes aberrant glycosylation and cellular mislocalization while serving as a potent oncogenic driver that promotes cancer cell proliferation, invasion, metastasis, and evasion of apoptotic mechanisms through its participation in complex intracellular signaling networks.[1][11][13] This report provides a comprehensive analysis of MUC1's structural organization, biochemical functions, cellular localization patterns, involvement in signaling cascades, post-translational modification patterns, and its transition from a protective epithelial component to a cancer-promoting oncogene.
Molecular Architecture and Structural Organization
MUC1 is a large, type I transmembrane glycoprotein that undergoes constitutive proteolytic processing to generate a functionally significant heterodimeric complex.[1][2] The full-length MUC1 protein, designated MUC1F, initially exists as a single polypeptide chain that is rapidly processed during and shortly after synthesis in the endoplasmic reticulum.[9] The primary transcript encodes three distinct structural domains: an extracellular domain, a hydrophobic transmembrane domain, and a cytoplasmic tail domain of 72 amino acids.[1][3] The extracellular domain contains the characteristic variable number of tandem repeats (VNTR) region, which in humans consists of 20 to 125 copies of a 20-amino acid sequence (GSTAPPAHGVTSAPDTRPAP) that is particularly rich in serine, threonine, and proline residues.[6][24] These serine and threonine residues serve as primary sites for extensive O-linked glycosylation, resulting in a heavily glycosylated extracellular polypeptide that can double the protein's molecular mass.[1][46] The tandem repeat domain is followed by the sperm protein-enterokinase-agrin (SEA) domain, a 120-amino acid structure that possesses a highly conserved GSVVV motif responsible for autoproteolytic self-cleavage.[6][9] This autoproteolytic cleavage occurs spontaneously during protein folding in the endoplasmic reticulum, between the conserved glycine and serine residues within the SEA module, generating two molecular species: the longer N-terminal subunit (MUC1-N) and the shorter C-terminal subunit (MUC1-C).[9][38] These two subunits, despite their proteolytic separation, remain physically associated through stable non-covalent hydrogen bonding, forming a metabolic complex that persists during transit through the Golgi apparatus and delivery to the cell surface.[3][9]
The structural arrangement of full-length MUC1 creates a distinctive architecture that extends prominently above the cell surface. The heavily glycosylated tandem repeat domain, combined with its rigid extended structure imparted by proline residues and glycosylation, projects 200 to 500 nanometers above the apical cell surface, a distance substantially greater than most other cell-surface proteins including syndecans and integrins.[1][3][7] This remarkable extension positions MUC1 ideally for its barrier and adhesion-modulating functions, allowing it to tower above the cellular glycocalyx and create a protective mucous layer. The transmembrane domain consists of a single hydrophobic alpha-helix that anchors the protein within the lipid bilayer.[2][4] The cytoplasmic tail domain, occupying the carboxy terminus, is remarkably conserved across mammalian species and contains seven highly conserved tyrosine residues interspersed with serine and threonine phosphorylation sites.[1][8][11] These tyrosines and serine/threonine residues serve as primary targets for phosphorylation by various protein tyrosine kinases and serine/threonine kinases, enabling the cytoplasmic tail to function as a signaling nexus that recruits and coordinates interactions with numerous intracellular proteins including β-catenin, Grb2, ErbB receptor family members, Src family kinases, and glycogen synthase kinase 3β.[2][11][14]
Alternative splicing of the MUC1 gene generates multiple transcript variants and protein isoforms that differ in their extracellular domain composition.[51] The best-characterized isoforms include MUC1/TR (full-length with the complete tandem repeat domain), MUC1/Z, MUC1/Y, and MUC1/X, the latter three lacking all or portions of the tandem repeat region.[51][54] These shorter isoforms containing truncated extracellular domains have been detected in various tumor cell lines and may contribute to heterogeneity in MUC1-dependent processes within malignant tissues. The functional significance of individual isoforms remains incompletely characterized, though some evidence suggests that certain short isoforms may have tumor-suppressive properties.[51]
Biosynthesis, Proteolytic Processing, and Membrane Trafficking
MUC1 undergoes a complex biosynthetic pathway involving multiple proteolytic processing steps and extensive post-translational modifications that culminate in its localization to specific cellular compartments.[3][9][39] Shortly after synthesis in the rough endoplasmic reticulum, MUC1F is rapidly directed through the secretory pathway, acquiring its initial post-translational modifications including N-linked glycosylation near the membrane-proximal region of the extracellular domain and beginning of O-linked glycosylation of the tandem repeats.[1][46] The protein transit through the endoplasmic reticulum to the Golgi apparatus occurs with a median time of approximately 142 minutes in mouse uterine epithelial cells.[3] During this transit, the SEA domain undergoes autoproteolytic cleavage, as described above, yet the resulting N-terminal and C-terminal subunits remain tightly associated through non-covalent interactions that are maintained through the secretory pathway.[9] This cleavage mechanism is particularly noteworthy because it represents a novel form of autoproteolysis catalyzed by conformational stress and the conserved serine hydroxyl within the GSVVV motif, involving an N-to-O-acyl rearrangement followed by hydrolytic resolution.[38][41] In the Golgi apparatus, MUC1 undergoes extensive additional O-glycosylation by multiple polypeptide N-acetyl-α-galactosaminyltransferases (GalNAc-Ts), with at least 15 different human GALNT gene products potentially participating in this process depending on the cell and tissue type.[46] Recent comprehensive investigations have revealed that different GalNAc-T isoforms display distinct substrate specificities and site preferences, with two identifiable clusters of GalNAc-Ts initially catalyzing glycosylation at different threonine residues within the tandem repeat consensus sequences.[22] The O-glycosylation contributes 50 to 90 percent of the total molecular mass of MUC1 based on the number of tandem repeats and the expression levels of relevant glycosyltransferases.[43]
Following glycosylation and processing in the Golgi apparatus, MUC1 is directed to the cell surface through vesicular trafficking pathways that involve lipid raft-associated mechanisms.[39][42] In normal polarized epithelial cells, MUC1 is specifically targeted to and concentrated at the apical membrane, a localization pattern that is critical for its barrier function and protection of epithelial surfaces.[1][3] The membrane trafficking of MUC1 is regulated by multiple post-translational modifications beyond glycosylation, including palmitoylation, which modulates the efficiency of trafficking to the cell surface and subsequent endocytosis.[39][42] Upon arrival at the cell surface, MUC1 undergoes several potential fates that determine its final steady-state abundance and function. In normal epithelial cells, MUC1 exhibits a metabolic half-life of 12 to 16 hours, indicating that mechanisms exist for both its recycling through the Golgi apparatus and its eventual degradation through proteasomal and lysosomal pathways.[3][9] A significant portion of surface-localized MUC1 is subject to ectodomain shedding, a process whereby proteases release the large extracellular domain from the cell surface while the transmembrane and cytoplasmic domains remain membrane-associated.[3][9][20] Two major sheddases have been identified as responsible for this process under different cellular contexts: TACE (TNF-α converting enzyme/ADAM17), which mediates constitutive and phorbol ester-stimulated shedding, and MT1-MMP (membrane type 1 matrix metalloproteinase), which is responsible for pervanadate-stimulated MUC1 ectodomain release.[20][23] The proteolytic cleavage occurs at the membrane-proximal region of the SEA domain or nearby regions, generating a membrane-associated C-terminal fragment containing the transmembrane and cytoplasmic domains.[9] The shed extracellular domain is released into the extracellular milieu, where it can interact with pathogens or circulating factors in a manner that contributes to immune defense and pathogen clearance.[6]
Following ectodomain shedding or in the absence of initial shedding, the remaining membrane-associated MUC1 containing the cytoplasmic tail undergoes further proteolytic processing by the membrane-embedded protease complex γ-secretase (presenilin-dependent).[9] This γ-secretase-mediated cleavage occurs at the cytosol-transmembrane boundary, generating a cytoplasmic fragment that can enter the nucleus and participate in transcriptional regulation.[9][46] However, the precise molecular mechanisms by which transmembrane domain-containing fragments reach the nucleus remain incompletely characterized, suggesting unconventional trafficking routes distinct from classical nuclear import pathways. The complete proteolytic cascade—consisting of autoproteolytic cleavage in the SEA module, ectodomain shedding by sheddases, and γ-secretase processing—represents a sophisticated regulatory mechanism that allows cells to dynamically control MUC1 function through selective proteolysis.
Function of MUC1 in Normal Epithelial Tissues
In healthy epithelial tissues, MUC1 serves critical protective and regulatory functions that maintain epithelial barrier integrity and defend against environmental challenges and pathogenic invasion.[1][3][6] The primary function of membrane-bound MUC1 in normal tissues is to act as a physical barrier that protects the apical cell membrane of epithelial cells from environmental damage, mechanical stress, pathogenic organisms, and degradative enzymes.[1][6][7] The extended structure of MUC1, projecting 200 to 500 nanometers above the cell surface, enables it to shield underlying cellular receptors and adhesion molecules from direct contact with potential pathogenic ligands and harmful environmental factors. MUC1 accomplishes this protective function through multiple interconnected mechanisms that operate in concert to maintain mucosal surface integrity.
The lubrication and hydration functions of MUC1 are particularly significant in mucosal tissues where surface wetness and frictionless movement are essential for normal physiological function.[1][3][6][21] The extensive O-glycosylation of MUC1, particularly through the accumulation of dense glycan structures, creates a highly hydrophilic surface layer that attracts and retains water molecules at the epithelial surface.[1][21] Recent tribological studies investigating MUC1's role in oral lubrication have demonstrated that MUC1 directly contributes to reducing friction forces during epithelial surface movement, with friction coefficient and energy dissipated both decreasing significantly in the presence of MUC1 compared to MUC1-deficient epithelial models.[21] These studies reveal that the absence of MUC1 expression results in increased frictional forces and enhanced tissue damage compared to epithelial tissues expressing normal MUC1 levels, highlighting the quantifiable protective benefit of this protein for preventing mechanical wear and trauma to mucosal surfaces.[21] Furthermore, MUC1 facilitates the anchoring of other salivary mucins, particularly the gel-forming mucin MUC5B, thereby modulating the composition and structure of the protective mucous pellicle that coats epithelial surfaces.[21]
In the normal oral mucosal epithelium, MUC1 functions in concert with other mucins (MUC5B and MUC7) to exert antimicrobial effects by continuously lubricating and stabilizing mucus on the cell surface while conferring protection against proteolysis and preventing dehydration.[1][41] This collaborative barrier function with other mucins extends to other mucosal surfaces including the respiratory, gastrointestinal, and reproductive tracts, where MUC1 constitutes a critical component of the first line of defense against microbial and parasitic pathogens.[1][3] The physical barrier function operates through what is termed "steric hindrance," whereby the towering glycosylated structure of MUC1 physically obstructs access by pathogens to underlying cellular receptors and adhesion molecules.[6][31] In the specific case of Helicobacter pylori infection, for example, MUC1 has been demonstrated to limit bacterial colonization through two complementary mechanisms: first, through steric hindrance that prevents bacterial attachment even to pathogens lacking specific MUC1 adhesins, and second, through acting as a "releasable decoy" for pathogens that do bind to MUC1, whereby the ectodomain is shed from the cell surface while bound to bacteria, physically removing the pathogen from the epithelial surface.[31][34] In an in vivo murine model of H. pylori infection, MUC1 expression limited the gastrointestinal and systemic spread of bacteria and significantly reduced intestinal inflammation compared to MUC1-deficient animals.[31]
Beyond its protective barrier functions, MUC1 in normal epithelial tissues plays a critical role in regulating cell-cell and cell-extracellular matrix interactions through its adhesion-modulating properties.[3][6][7] The extended structure and dense glycosylation of the tandem repeat domain, combined with the elevated expression and concentration of MUC1 at the apical surface in most simple epithelia, collectively function to modulate adhesive interactions by steric hindrance.[1][7] This anti-adhesive function of MUC1 is particularly important in reproductive tissue epithelia during critical periods such as the window of implantation, where precise regulation of epithelial-blastocyst interactions is essential for successful pregnancy.[3][7] The expression of MUC1 in normal epithelia can be dynamic, varying in response to hormonal influences, particularly steroid hormones and cytokine signals that regulate transcription of the MUC1 gene.[3] This dynamic regulation enables MUC1 expression to be modulated in response to physiological demands and environmental signals, allowing tissues to adjust their defensive capabilities in response to changing conditions. Recent research has also identified a role for MUC1 as an important scaffold for pluripotent stem cell propagation, with a transmembrane cleavage product (MUC1*) capable of helping to propagate large numbers of pluripotent stem cells for therapeutic interventions.[1]
Regulation of Inflammation and Innate Immune Responses
Beyond its direct physical barrier functions, MUC1 plays a sophisticated regulatory role in controlling inflammatory responses to infection and maintaining mucosal immune homeostasis through interactions with pattern recognition receptors and modulation of innate immune signaling.[1][6][24][33][36] MUC1 functions as a negative regulator of multiple Toll-like receptor (TLR) signaling pathways that would otherwise be activated by pathogen-associated molecular patterns (PAMPs) and danger-associated molecular patterns (DAMPs).[36][33] Comprehensive studies have demonstrated that MUC1 suppresses TLR2, TLR3, TLR4, TLR5, TLR7, and TLR9 signaling, indicating a broad anti-inflammatory capacity of this molecule.[36] The cytoplasmic tail domain of MUC1 interacts constitutively with TLR3, and upon stimulation with the double-stranded RNA analog poly(I:C), MUC1 prevents the recruitment of the TIR-domain-containing adapter-inducing interferon-β (TRIF) to the TLR3 signaling complex, thereby attenuating the activation of interferon regulatory factor-3 (IRF-3), NF-κB, and the production of interferon-β.[33] This counter-regulatory effect of MUC1 on TLR3 signaling results in decreased poly(I:C)-induced apoptosis and reduced inflammatory activation of epithelial cells. Similarly, MUC1 suppresses TLR5 signaling by interacting with TLR5 and competitively inhibiting the recruitment of the MyD88 adaptor protein to the TLR5 cytoplasmic domain.[36] The mechanism underlying the broad suppressive effect of MUC1 on diverse TLR pathways likely involves the MUC1 cytoplasmic tail's interactions with signaling molecules such as PI3K, Shc, Src family kinases, β-catenin, GSK-3β, and Grb-2/Sos that participate in the convergent NF-κB activation pathway common to all TLRs.[36]
Importantly, MUC1 can also play a pro-inflammatory role in certain contexts, particularly in the setting of established cancer or chronic infection.[1][24] MUC1 has been shown to act as an immunomodulatory switch that can exhibit either pro- or anti-inflammatory functions depending on the specific cellular and tissue context.[1][24] In infection-induced cancers and chronic inflammatory settings, the tumor form of MUC1 can establish specific interactions with dendritic cells and macrophages by controlling the recruitment of inflammatory cells, thereby promoting tumor escape from the immune system and creating a pro-inflammatory microenvironment that paradoxically facilitates cancer progression.[1][24] Additionally, altered MUC1 glycosylation in cancer cells promotes chronic inflammatory conditions that lead to malignant transformation and further cancer progression.[1][24] The ability of MUC1 to potentiate epithelial-mesenchymal transition (EMT) through activation of the inflammatory NF-κB p65 pathway, which then activates the EMT transcriptional repressor zinc-finger E-box-binding homeobox 1 (ZEB1), exemplifies how pro-inflammatory MUC1 signaling can contribute to cancer invasiveness and metastatic potential.[1] Thus, MUC1 functions as a dynamic regulator of inflammation whose activity is context-dependent and whose pathological activation in cancer contributes to disease progression.
MUC1 as an Oncogenic Driver: Structure-Function Relationships in Cancer
Following malignant transformation, MUC1 undergoes dramatic alterations in its expression patterns, cellular localization, and glycosylation characteristics that convert it from a protective epithelial component into a potent oncogenic driver promoting cancer cell proliferation, survival, invasion, metastasis, and evasion of death signals.[1][10][35] Multiple aspects of MUC1 structure contribute independently to its oncogenic function, with both the extracellular tandem repeat domain and the cytoplasmic tail domain being essential for MUC1-driven tumor formation.[11] This has been demonstrated conclusively through mouse models of mammary carcinogenesis showing that deletion of either the tandem repeat domain or the cytoplasmic tail domain independently abrogates MUC1's ability to drive early-onset tumor formation in transgenic animals.[11] These findings indicate that the adhesion-modulating functions of the extracellular domain and the cellular signaling functions of the cytoplasmic tail domain both contribute substantially to MUC1-mediated oncogenesis, and that neither domain is sufficient alone to promote tumorigenesis.
In cancer cells, MUC1 expression increases substantially while undergoing aberrant glycosylation that exposes the underlying peptide core and generates new carbohydrate epitopes, particularly the Thomsen-Friedenreich (TF or T), Tn, and sialyl-Tn (STn) antigens.[1][10][35] This aberrant glycosylation pattern reflects altered expression and activity of glycosyltransferases in cancer cells compared to normal epithelial cells, resulting in the generation of shorter, simpler glycan structures that expose previously hidden immunogenic epitopes on the MUC1 peptide backbone.[1][22][35] This glycosylation abnormality has attracted substantial clinical interest because these altered glycan epitopes can serve as tumor-associated antigens for development of cancer vaccines and immunotherapeutic approaches.[1][22]
Loss of epithelial polarity during malignant transformation leads to dramatic changes in MUC1 subcellular localization, with cancer cells exhibiting cytoplasmic and basally localized MUC1 in addition to or instead of the normal apical membrane localization.[10][32][35] This altered localization pattern is highly significant because it allows the MUC1 cytoplasmic tail to interact more promiscuously with intracellular signaling molecules and permits increased crosstalk between MUC1 and membrane receptor tyrosine kinases that normally reside exclusively at basolateral membrane domains in polarized epithelial cells.[14][32][35] The altered localization and increased availability of MUC1 to interact with signaling proteins constitutes a critical aspect of its oncogenic function, enabling enhanced activation of multiple downstream signaling pathways that promote cancer cell growth and survival.
Intracellular Signaling Pathways Activated by MUC1
The MUC1 cytoplasmic tail serves as a critical signaling hub that participates in and coordinates multiple interconnected cellular signaling pathways central to cancer cell biology.[13] These pathways include the Wnt/β-catenin pathway, NF-κB signaling, PI3K-AKT signaling, and MAPK/ERK signaling, with MUC1 acting to either promote or suppress activation of these pathways depending on the cellular context.[1][13][14] The MUC1 cytoplasmic tail contains multiple protein-binding motifs that facilitate interactions with dozens of signaling proteins, including receptor tyrosine kinases, adaptor proteins, and transcription factors.[2][14]
MUC1 functions as a direct activator of the NF-κB pathway, a critical survival signaling cascade frequently dysregulated in cancer.[40][43] The MUC1-C cytoplasmic domain binds directly to NF-κB p65 protein and, importantly, blocks the interaction between NF-κB p65 and its inhibitor IκBα, thereby preventing the sequestration and degradation of p65 and promoting its nuclear translocation and transcriptional activation.[40] In carcinoma cells, MUC1-C and NF-κB p65 constitutively occupy the promoter of the anti-apoptotic Bcl-xL gene, with MUC1-C contributing to NF-κB-mediated transcriptional activation of this survival gene.[40] In non-malignant epithelial cells, MUC1-C interacts with NF-κB p65 in response to TNFα stimulation, and TNFα induces the recruitment of NF-κB p65-MUC1-C complexes to NF-κB target genes, including the MUC1 gene promoter itself, creating a positive feedback loop that amplifies MUC1 expression in response to inflammatory signals.[40] Importantly, synthetic peptide inhibitors of MUC1-C oligomerization can block the interaction between MUC1-C and NF-κB p65, decrease promoter occupancy, and suppress NF-κB-mediated gene transcription, demonstrating the potential for targeted inhibition of this pathway in therapeutic contexts.[40]
MUC1 also participates critically in the Wnt/β-catenin signaling pathway, one of the most frequently dysregulated pathways in human cancer.[1][11][13] The MUC1 cytoplasmic tail binds directly to β-catenin and facilitates its nuclear translocation in association with transcription factor TCF4, enabling activation of Wnt target genes that promote cell proliferation and survival.[1][11] In pancreatic cancer cells, MUC1 has been shown to enhance invasiveness through a mechanism dependent on the interaction between MUC1 and β-catenin, with this interaction leading to nuclear translocation of β-catenin and activation of EMT-promoting transcription factors including Snail and Slug.[45] The functional tyrosine residues in the MUC1 cytoplasmic tail are critical for stimulating the MUC1-β-catenin interaction and their nuclear translocation; mutation of all seven tyrosines to phenylalanine prevents this interaction and blocks EMT induction in response to MUC1 overexpression.[45] These findings directly establish the tyrosines of the MUC1 cytoplasmic tail as critical functional elements required for MUC1-mediated promotion of epithelial-mesenchymal transition.
MUC1 participates in regulation of the receptor tyrosine kinase (RTK) signaling pathways, particularly the EGFR and ErbB2 pathways, which drive proliferation and survival in numerous epithelial malignancies.[1][13][14] MUC1 can physically interact with EGFR, HER2 (ErbB2), FGFR3, IGF1R, MET, and PDGFRβ, and it serves as a substrate for phosphorylation by these kinases on its cytoplasmic tail tyrosine residues.[1][13][15] Activation of MUC1 through receptor cross-talk with EGFR and other receptor tyrosine kinases triggers downstream signaling through AKT1, MAPK1/2, and β-catenin, promoting cancer cell proliferation, growth, angiogenesis, invasion and migration in multiple cancer types including non-small cell lung cancer, ovarian cancer, breast cancer, and endometrial cancer.[13] Notably, MUC1 expression inhibits the degradation of ligand-activated ErbB1 following growth factor stimulation, thereby increasing the cellular pools of active receptor over time and prolonging mitogenic signaling.[15] This MUC1-mediated protection against ErbB1 degradation occurs through inhibition of EGF-stimulated ubiquitination of the receptor and through promotion of receptor recycling rather than degradation, demonstrating a specific mechanism by which MUC1 can enhance RTK signaling intensity and duration in cancer cells.[15]
MUC1 regulates the MAPK pathway through multiple mechanisms involving phosphorylation-dependent interactions with signaling proteins.[8][13] In cells lacking the seven tyrosine residues of the MUC1 cytoplasmic tail (MUC1 Y0 mutant), heightened active ERK1/2 is observed with increased transcriptional activity of AP-1 and STAT3 compared to wild-type MUC1-expressing cells, while NF-κB transcriptional activity is paradoxically decreased.[8] These findings suggest that the phosphorylation status of MUC1 cytoplasmic tail tyrosines dynamically modulates the balance between different transcriptional programs, with consequences for cell proliferation, invasiveness, and cell cycle progression.[8]
MUC1 interacts with PI3K and influences the PI3K-AKT signaling pathway, though in some cellular contexts MUC1 has been shown to suppress PI3K-AKT signaling.[13][29] This suppression of the inhibitory PI3K-Akt pathway is associated with activation of AMPK and promotion of autophagy in the response to glucose deprivation, enabling cancer cells to maintain ATP production and survival under nutrient-limited conditions.[29] The ability of MUC1 to promote autophagy as a survival response to metabolic stress demonstrates how MUC1 can facilitate cancer cell adaptation to the harsh microenvironmental conditions within tumors, where nutrient availability is often limited.[29]
MUC1 localizes to subcellular compartments beyond the plasma membrane, including mitochondria and lysosomes, where it participates in cellular stress responses.[13][46] Following activation by ErbB receptors in the presence of the heregulin (HRG) ligand, activated MUC1 binds to heat shock proteins (HSP90 and HSP70) and translocates to mitochondria and lysosomes, respectively.[13] This unconventional localization of MUC1 to organelles involved in energy metabolism and autophagic degradation suggests that MUC1 may directly regulate mitochondrial function and lysosomal autophagy, both processes that are frequently dysregulated in cancer cells and that contribute to metabolic adaptation and therapy resistance.
Nuclear Functions and Transcriptional Regulation
A particularly important and extensively studied function of MUC1 involves its translocation to the nucleus where its cytoplasmic tail interacts with multiple transcription factors to directly modulate gene expression and cellular fate decisions.[14][37][40] MUC1 contains a non-classical nuclear localization signal (RLS/RRK motif) that interacts with nucleoporin-62 and importin-β1, enabling nuclear co-translocation of numerous associated proteins including β-catenin, γ-catenin, p53, estrogen receptor α, and NF-κB p65.[14][37] In the nucleus, MUC1-C forms physical complexes with multiple transcription factors that subsequently occupy response elements in the promoters of target genes and directly modulate their transcriptional activity.[14] The N-terminal MUC1 subunit can also accumulate in nuclear compartments under certain conditions, though the mechanisms and functional significance of nuclear MUC1-N remain less thoroughly characterized than those of the MUC1-C cytoplasmic tail.[46]
MUC1 plays a critical role in regulating p53-mediated gene transcription and the cellular response to genotoxic stress.[14][47] The MUC1-C cytoplasmic domain interacts constitutively with the p53 tumor suppressor protein, with this interaction being increased in response to genotoxic stress.[14] The MUC1-C cytoplasmic domain (amino acids 9-46) binds directly to the p53 regulatory domain (amino acids 363-393), and both MUC1-C and p53 occupy the promoter of the p53-responsive p21 gene in carcinoma cells.[14] MUC1-C occupancy of the p21 promoter is associated with recruitment of the CBP histone acetyltransferase, a decrease in HDAC1 histone deacetylase, and an increase in histone H4 acetylation, indicating that MUC1 modulates the chromatin architecture at p53-responsive promoters.[14] MUC1 has also been shown to repress the activation of p53 gene transcription and to block the binding of MUC1 and p53 to the PE21 promoter element, thereby repressing p53 activity in response to stress, which would normally promote apoptosis or cell cycle arrest in normal cells.[14][17] This repression of p53 function by MUC1 represents a critical mechanism by which MUC1 promotes cancer cell survival by disabling this central tumor suppressor pathway. However, in the specific case where the MUC1 cytoplasmic tail tyrosine at position 60 is mutated to phenylalanine (MUC1 Y60F), this mutant becomes a potent inducer of the ARF tumor suppressor, which in turn inhibits MDM2 and leads to p53 upregulation, demonstrating that the functional status of specific tyrosine residues in the MUC1 cytoplasmic tail profoundly influences whether MUC1 acts as a p53 inhibitor or a p53 activator.[47]
MUC1 also functions as a direct regulator of EGFR nuclear localization and transcriptional function in breast cancer cells.[37] MUC1 and EGFR interact in the nucleus and MUC1 promotes the accumulation of chromatin-bound EGFR at the cyclin D1 gene promoter, a critical cell cycle promoter gene essential for G1 to S phase transition.[37] This MUC1-mediated promotion of EGFR nuclear translocation and association with the cyclin D1 promoter provides a direct molecular link between MUC1 expression and accelerated cell cycle progression in response to growth factor signaling, independent of classical mitogenic signaling at the cell surface.
Post-Translational Modifications and Their Regulatory Functions
MUC1 is subject to extensive and complex post-translational modifications that profoundly influence its function, localization, protein-protein interactions, and signaling capacity.[19][22][46] The most characteristic post-translational modification of MUC1 is O-linked glycosylation of the tandem repeat domain, which accounts for 50 to 90 percent of the total molecular mass of the protein depending on the number of tandem repeats and the expression of relevant glycosyltransferases.[43][46] O-linked glycosylation is initiated by members of the polypeptide N-acetylgalactosaminyltransferase (GALNT) family, with at least 15 different GALNT genes in the human genome potentially participating in MUC1 glycosylation depending on cell type and tissue context.[46] Comprehensive structural and biochemical investigations have revealed that individual GalNAc-T isoforms display distinct substrate site specificities, with two identifiable clusters of GalNAc-Ts initially catalyzing glycosylation at different threonine residues within the consensus sequences GSTA and GVTS within the MUC1 tandem repeats.[22] These initial glycosylation events are then followed by extension of the O-linked oligosaccharides through the sequential action of additional glycosyltransferases that add galactose, N-acetylgalactosamine (GalNAc), fucose, and sialic acid residues, with sialylation being particularly extensive on MUC1 glycans.[22][46] The specific pattern of O-glycosylation depends on the complement and expression levels of glycosyltransferases present in specific cell types and tissues, enabling tissue and cell-type specific glycosylation patterns of MUC1.[22][46]
The pattern of MUC1 glycosylation profoundly influences multiple aspects of MUC1 function, including its ability to interact with cellular receptors, immune cells, and pathogens, as well as its localization within the cell and its susceptibility to protease cleavage.[46][43] O-glycosylation can stabilize MUC1 at the cell surface by limiting endocytosis and by protecting the polypeptide chain from degradation by extracellular proteases.[46] Conversely, altered glycosylation patterns associated with malignant transformation create abnormal MUC1 epitopes that are recognized by anti-MUC1 antibodies and that may serve as tumor-associated antigens for immune recognition. Remarkably, the O-glycosylation of MUC1 has been shown to be responsible for MUC1-induced drug resistance in breast cancer cells, with the extracellular glycosylated modification of the MUC1-N subunit acting as a barrier that reduces intracellular drug penetration.[43] Removal of O-glycosylation through pharmacological inhibitors, enzymatic digestion, or genetic deletion of MUC1-related O-glycosyltransferases (GCNT3) markedly reinvigorates chemosensitivity in cancer cells, revealing a therapeutically actionable mechanism through which the glycosylation status of MUC1 directly influences cancer cell drug sensitivity.[43]
Beyond O-glycosylation, the MUC1 cytoplasmic tail undergoes phosphorylation on its seven tyrosine residues and multiple serine and threonine residues, modifications that critically regulate its interactions with signaling proteins and transcription factors.[8][46] Phosphorylation of MUC1 by various protein tyrosine kinases including MET and SRC enables specific interactions with signaling proteins such as p53 and HSP90, thereby directly modulating MUC1's ability to participate in particular signaling cascades.[46] The MUC1 cytoplasmic tail also undergoes ubiquitination, a modification that can target MUC1 for proteasomal degradation or can alter its signaling properties, though the specific ubiquitination sites and the functional consequences of this modification remain incompletely characterized for MUC1.[46] Additionally, the MUC1 cytoplasmic tail undergoes palmitoylation, a lipid modification that influences its association with membrane lipid rafts and affects its trafficking properties and endocytosis rates.[39][42]
Pathogenic Mutations and Associated Genetic Diseases
While MUC1 is primarily known for its roles in epithelial homeostasis and cancer biology, inherited mutations in the MUC1 gene can cause medullary cystic kidney disease type 1 (MCKD1), an autosomal dominant inherited condition characterized by progressive kidney fibrosis and renal cyst formation leading to chronic kidney failure.[49] MCKD1 is caused by insertion of a cytosine nucleotide into the MUC1 gene in a specific region, resulting in production of an altered MUC1 protein.[49] The precise mechanism by which these MUC1 mutations cause kidney disease remains incompletely understood, though it is theorized that MUC1 plays an important role in normal kidney development and maintenance of tubular epithelial cell function through its signaling capabilities and its role in regulating cell growth, movement, and survival.[49] The fact that relatively specific mutations in MUC1 cause progressive organ failure demonstrates the biological importance of normal MUC1 function in maintaining long-term epithelial tissue homeostasis and preventing pathological remodeling and fibrosis.
Additionally, MUC1 VNTR (variable number tandem repeat) polymorphisms, which result from variations in the number of tandem repeat copies in the MUC1 gene, have been associated with susceptibility to gastric cancer and Helicobacter pylori-associated gastritis in population-based genetic studies.[28][31] Individuals carrying shorter MUC1 VNTR alleles that encode MUC1 proteins with shorter extracellular glycosylated domains are associated with increased disease susceptibility.[31] This association is mechanistically explained by the observation that shorter MUC1 VNTR variants are less efficient at sterically inhibiting bacterial attachment or acting as releasable decoys for H. pylori, thereby allowing increased bacterial binding to the epithelial surface and exacerbation of pathology.[28][31]
Clinical Applications and Biomarker Significance
MUC1 has emerged as a significant clinical biomarker and therapeutic target due to its frequent overexpression and aberrant glycosylation in multiple human cancers.[1][27][48] MUC1 is commonly overexpressed in numerous epithelial adenocarcinomas including lung adenocarcinoma (where 86.3% of stage IB patients show high MUC1 expression), breast cancer, pancreatic cancer, gastric cancer, ovarian cancer, colorectal cancer, and cholangiocarcinoma.[1][27][48] In many of these malignancies, MUC1 expression levels correlate with aggressive clinicopathological features including advanced tumor stage, lymph node metastasis, poor differentiation, and reduced patient survival time.[27][48] For example, in pancreatic cancer patients, median survival time is significantly lower in those with high MUC1 expression (13.4 months) compared to those with low expression (39.7 months), indicating that MUC1 expression is a strong adverse prognostic factor in this uniformly lethal malignancy.[27] Similarly, in gallbladder adenocarcinoma, high MUC1 expression is significantly associated with advanced tumor stage, lymph node metastasis, and reduced overall survival, suggesting utility as a diagnostic and prognostic biomarker.[27]
The subcellular localization pattern of MUC1 has been shown to have prognostic significance that is independent of overall MUC1 expression levels.[10][32] Breast cancer patients whose tumors display apical MUC1 localization patterns (similar to that seen in normal epithelial cells) have significantly better outcomes compared to those with cytoplasmic or negative MUC1 patterns, with apical MUC1 localization being associated with lower recurrence scores and reduced tumor malignancy.[10][32] This observation suggests that preservation of normal epithelial polarity and proper MUC1 localization is associated with less aggressive tumor behavior, presumably because it indicates retention of more differentiated epithelial characteristics and reduced engagement of MUC1's oncogenic signaling functions that require aberrant cytoplasmic localization.[10][32]
The soluble form of MUC1, generated through ectodomain shedding and commonly referred to as CA15-3 (cancer antigen 15-3), has been established as a circulating biomarker with clinical utility in breast cancer and other malignancies.[1][57] Elevated serum CA15-3 levels are associated with shorter disease-free survival and overall survival time in breast cancer patients, and serial measurements of serum CA15-3 can be used to monitor treatment response and detect cancer recurrence.[57] MUC1 in endometrial tissue has also been identified as an independent marker of endometrial receptivity in the context of recurrent implantation failure, with decreased MUC1 expression in endometrial epithelium being associated with reproductive failure, suggesting that MUC1 may contribute to the reproductive failure observed in women with recurrent implantation failure.[50]
Therapeutic Targeting and Immunotherapy
The importance of MUC1 in cancer biology and its frequent overexpression and aberrant glycosylation in malignancies has motivated the development of multiple therapeutic strategies targeting MUC1.[1][18] These approaches include development of MUC1-specific monoclonal antibodies, MUC1-targeted chimeric antigen receptor (CAR) engineered T-cell therapies, MUC1 vaccines targeting aberrant MUC1 glycoforms, and small molecule inhibitors of MUC1 function.[1][18] CAR T-cell therapy employing dual-targeted engineered T-cells expressing both anti-ErbB2 and anti-MUC1 CARs has been developed to achieve complementary signaling within the tumor microenvironment, enabling selective activation of T-cells only when they encounter tumor cells expressing both ErbB2 and MUC1 simultaneously.[18] This approach has demonstrated proof of principle that dual targeting of MUC1 with other cancer-associated antigens can achieve enhanced specificity and potentially reduced off-target toxicity compared to single-antigen targeting approaches.[18] The development of MUC1-specific antibodies that recognize MUC1 with specific aberrant glycosylation patterns has enabled discrimination between tumor-associated and normal MUC1 forms, providing tools for both diagnostic and therapeutic applications.[56]
Conclusion
MUC1 represents a multifunctional glycoprotein whose biological roles are dramatically transformed during malignant progression. In normal epithelial tissues, MUC1 serves essential protective functions as a physical barrier, lubricant, and immune modulator that maintains tissue homeostasis, defends against pathogenic invasion, and prevents excessive inflammation through negative regulation of Toll-like receptor signaling. The extended extracellular structure of MUC1, maintained at 200 to 500 nanometers above the cell surface through its VNTR and SEA domains, positions it ideally for barrier functions, while its conserved cytoplasmic tail enables anti-inflammatory signaling through interactions with TLRs and related pathways. In cancer cells, through loss of epithelial polarity, aberrant glycosylation, and increased expression, MUC1 becomes a potent oncogenic driver that simultaneously promotes cancer cell proliferation and survival through interactions with receptor tyrosine kinases, suppresses apoptotic pathways through inhibition of p53 and activation of NF-κB, facilitates invasion and metastasis through promotion of epithelial-mesenchymal transition mediated by β-catenin interactions, and enables cancer cell survival under metabolic stress through promotion of autophagy. The functional importance of both the extracellular tandem repeat domain and the cytoplasmic tail domain to MUC1-driven oncogenesis, combined with MUC1's frequent overexpression and prognostic significance in multiple malignancies, establishes MUC1 as a high-priority target for cancer therapeutic development. Future therapeutic strategies will likely benefit from selective targeting of MUC1's aberrant glycoforms or interaction with specific signaling partners such as β-catenin, receptor tyrosine kinases, or transcription factors, enabling more precise inhibition of MUC1's oncogenic functions while potentially preserving its normal protective functions in healthy epithelial tissues.
Citations
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8234110/
- https://www.genecards.org/cgi-bin/carddisp.pl?gene=MUC1
- https://pmc.ncbi.nlm.nih.gov/articles/PMC320498/
- https://www.uniprot.org/uniprotkb/P15941/entry
- https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=4582
- https://www.frontiersin.org/journals/cellular-and-infection-microbiology/articles/10.3389/fcimb.2019.00117/full
- https://pubmed.ncbi.nlm.nih.gov/16849524/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3228317/
- https://www.nature.com/articles/s41598-023-32579-4
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3091404/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC12297148/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3615539/
- https://pubmed.ncbi.nlm.nih.gov/16983337/
- https://www.atsjournals.org/doi/pdf/10.1164/ajrccm-conference.2016.193.1_MeetingAbstracts.A6539?download=true
- https://aacrjournals.org/cancerres/article/67/4/1853/534036/Human-Mucin-1-Oncoprotein-Represses-Transcription
- https://pubmed.ncbi.nlm.nih.gov/22526592/
- https://pubmed.ncbi.nlm.nih.gov/12925576/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1133949/
- https://www.nature.com/articles/s41598-024-82176-2
- https://pubs.acs.org/doi/10.1021/acscatal.4c05719
- https://academic.oup.com/endo/article/145/9/4192/2878263/Tumor-Necrosis-Factor-Stimulates-MUC1-Synthesis
- https://pubmed.ncbi.nlm.nih.gov/8835095/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4931332/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9716114/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2693691/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3027209/
- https://www.proteinatlas.org/ENSG00000185499-MUC1
- https://journals.plos.org/plospathogens/article?id=10.1371%2Fjournal.ppat.1000617
- https://pmc.ncbi.nlm.nih.gov/articles/PMC4189494/
- https://pubmed.ncbi.nlm.nih.gov/19816567/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2258447/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2864713/
- https://pubmed.ncbi.nlm.nih.gov/15987679/
- https://experiments.springernature.com/articles/10.1007/978-1-61779-513-8_7
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2760979/
- https://pubmed.ncbi.nlm.nih.gov/16369486/
- https://pubmed.ncbi.nlm.nih.gov/22259133/
- https://www.nature.com/articles/s41419-022-05110-2
- https://www.nature.com/articles/s41418-022-00988-z
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3063863/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3189326/
- https://pubmed.ncbi.nlm.nih.gov/18981727/
- https://www.spandidos-publications.com/10.3892/mco.2022.2594
- https://medlineplus.gov/genetics/condition/medullary-cystic-kidney-disease-type-1/
- https://pubmed.ncbi.nlm.nih.gov/29929546/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC3538090/
- https://rarediseases.org/mondo-disease/tubulointerstitial-kidney-disease-autosomal-dominant-2/
- https://www.nature.com/articles/s41467-021-23571-5
- https://www.nature.com/articles/6604617
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5326570/
- https://pubmed.ncbi.nlm.nih.gov/35887202/
- https://pmc.ncbi.nlm.nih.gov/articles/PMC7377060/
- https://pubs.acs.org/doi/10.1021/jacsau.3c00587
- https://www.nature.com/articles/s41698-025-01016-2