Deep Research
Falcon
(EMC2-deep-research-falcon.md)
Falcon
(EMC2-deep-research-falcon.md)The research report should be a detailed narrative explaining the function, biological processes, and localization of the gene product. Citations should be given for all claims.
You should prioritize authoritative reviews and primary scientific literature when conducting research. You can supplement
this with annotations you find in gene/protein databases, but these can be outdated or inaccurate.
We are specifically interested in the primary function of the gene - for enzymes, what reaction is catalyzed, and what is the substrate specificity? For transporters, what is the substrate? For structural proteins or adapters, what is the broader structural role? For signaling molecules, what is the role in the pathway.
We are interested in where in or outside the cell the gene product carries out its function.
We are also interested in the signaling or biochemical pathways in which the gene functions. We are less interested in broad pleiotropic effects, except where these elucidate the precise role.
Include evidence where possible. We are interested in both experimental evidence as well as inference from structure, evolution, or bioinformatic analysis. Precise studies should be prioritized over high-throughput, where available.
Research Report: Human EMC2 (UniProt Q15006; gene EMC2; aliases TTC35, KIAA0103)
Executive summary
Human EMC2 encodes a cytosolic-facing subunit of the endoplasmic reticulum (ER) membrane protein complex (EMC), a conserved machine that promotes membrane protein biogenesis. The best-supported primary function of EMC2 is architectural/assembly scaffolding of the EMC cytosolic “basket,” positioning other EMC elements (notably the EMC3/EMC6 membrane vestibule and EMC3 cytosolic loop) that directly execute insertase and topology-control activities for selected membrane proteins. Recent work (2023–2024) has refined EMC mechanisms for selectivity filtering (preventing misinsertion and enforcing topogenesis rules) and post-translational topology rectification for multipass clients; these advances update how EMC2’s scaffold role is interpreted in pathway context. (pleiner2020structuralbasisfor pages 1-3, pleiner2023aselectivityfilter pages 8-10, wu2024emcrectifiesthe pages 7-9)
1) Target identity verification (critical disambiguation)
Authoritative EMC structural and mechanistic papers explicitly identify human EMC2 as a ~35 kDa, cytosolic EMC subunit (also called TTC35) that is part of the ER membrane protein complex. This matches the provided UniProt identity (Q15006: “ER membrane protein complex subunit 2”; synonyms TTC35/KIAA0103) and places the protein in ER membrane protein biogenesis rather than in an unrelated pathway. (chitwood2019theroleof pages 2-4, pleiner2020structuralbasisfor pages 1-3)
2) Key concepts and definitions (current understanding)
2.1 The ER membrane protein complex (EMC)
The EMC is a multi-subunit ER complex implicated in insertion, folding, and assembly of membrane proteins. It has an established insertase role for certain transmembrane domains (TMDs) and additional roles in later steps of membrane protein maturation. (hegde2022thefunctionstructure pages 4-6, odonnell2020thearchitectureof pages 1-2)
2.2 “Insertase,” “topogenesis,” and “selectivity filter”
- Insertase: A factor that facilitates insertion of TMDs into the lipid bilayer in an energy-independent or low-energy manner. Purified EMC can catalyze insertion of select tail-anchored (TA) TMDs in vitro, supporting the idea that EMC itself is an insertase. (odonnell2020thearchitectureof pages 1-2, odonnell2020thearchitectureof pages 2-4)
- Topogenesis: The establishment of correct membrane protein topology (cytosolic vs luminal orientation) during biogenesis. EMC has been shown to be required for accurate membrane protein topogenesis and to prevent misinsertion/mistopology. (pleiner2023aselectivityfilter pages 1-2, wu2024emcrectifiesthe pages 7-9)
- Selectivity filter (2023): A mechanistic feature of EMC that limits inappropriate insertion events (e.g., misinsertion of mitochondrial TA proteins into the ER) and helps enforce topology rules (e.g., “positive-inside”). (pleiner2023aselectivityfilter pages 8-10, pleiner2023aselectivityfilter pages 1-2)
3) EMC2: subcellular localization and complex membership
3.1 Localization
EMC2 is cytosolic-facing and resides as part of the ER-resident EMC, rather than acting as a free cytosolic chaperone. In the human EMC cryo-EM model, EMC2 sits in the cytosolic region adjacent to other cytosolic and membrane subunits that create the substrate-entry vestibule. (pleiner2020structuralbasisfor pages 1-3, hegde2022thefunctionstructure pages 4-6)
3.2 Complex membership and paralog relationships
Human EMC contains cytosolic subunits EMC2 plus EMC8 and/or EMC9. Structural/biochemical work indicates EMC2 forms stable complexes with EMC8/9 that can be mutually exclusive in assembly contexts, consistent with functional substitution by paralogs in some settings. (pleiner2020structuralbasisfor pages 7-11, odonnell2020thearchitectureof pages 2-4)
4) Primary molecular function of EMC2 (mechanism-level annotation)
4.1 Architectural scaffold for the EMC cytosolic domain
High-resolution structural work indicates EMC2 is an architectural scaffold that organizes the cytosolic portion of EMC. In the human EMC cryo-EM structure, EMC2 “acts as an architectural scaffold for EMC8 and the cytosolic regions of EMC3, 5, and 1,” consistent with EMC2 being central for EMC integrity. (pleiner2020structuralbasisfor pages 1-3)
Mechanistically, EMC2:
* forms an α-solenoid that binds the three-helix bundle formed by the coiled-coil and C-terminus of EMC3; (pleiner2020structuralbasisfor pages 1-3)
* clamps around EMC8 via an extensive hydrophobic surface; (pleiner2020structuralbasisfor pages 1-3)
* contributes to composite interfaces that accommodate the C-terminal tail of EMC5, which traverses through the center of EMC2 to the cytosolic face. (pleiner2020structuralbasisfor pages 1-3)
Mutations at EMC2 interfaces disrupt subunit binding/assembly in vitro, supporting a non-redundant structural role. (pleiner2020structuralbasisfor pages 1-3, pleiner2020structuralbasisfor pages 7-11)
4.2 EMC2 and the substrate-entry vestibule
In the architecture model, the cytosolic vestibule that initially receives TMDs includes EMC2 (in complex with EMC8/9). EMC2 contributes conserved basic residues at the entry region (e.g., Arg26, Arg91) that may participate in substrate filtering, disfavoring passage of highly basic segments toward the intramembrane groove and thereby contributing to selectivity. (odonnell2020thearchitectureof pages 14-15)
5) Pathway context: how EMC2 fits into EMC-mediated membrane protein biogenesis
5.1 Co- and post-translational insertion and topology enforcement
The EMC is described as a co- and post-translational insertase at the ER. In the human structure, the membrane insertion pathway proceeds via an enclosed hydrophilic vestibule within the membrane formed by EMC3 and EMC6, with a methionine-rich cytosolic loop implicated in substrate capture. EMC2 scaffolding helps position these elements within a functional assembly. (pleiner2020structuralbasisfor pages 1-3)
5.2 Cotranslational engagement of multipass clients
Proteomics and ribosome profiling indicate EMC engages a range of multipass membrane proteins cotranslationally, with enrichment for transporters and other challenging substrates (e.g., TMDs with charged residues). This is a complex-level activity, but EMC2 depletion can destabilize EMC subunits, consistent with EMC2 being required for these functions by maintaining complex integrity. (shurtleff2018theermembrane pages 8-10, chitwood2019theroleof pages 2-4)
6) Recent developments (prioritizing 2023–2024)
6.1 2023: EMC selectivity filter limits misinsertion and enforces topogenesis
Pleiner et al. (Journal of Cell Biology; May 2023) report that EMC limits misinsertion at the ER via a positively charged, hydrophilic vestibule that functions as a selectivity filter. Charge repulsion disfavors translocation of positively charged segments and contributes to enforcing “positive-inside” topology rules; altering key EMC3 residues can increase misinsertion (e.g., mislocalization of RHOT1). The work used split-GFP topology reporters, glycosylation assays, and crosslinking approaches to map substrate contacts through the vestibule. (pleiner2023aselectivityfilter pages 8-10, pleiner2023aselectivityfilter pages 2-4)
Importantly for EMC2 annotation, the study ties correct assembly/biogenesis of the insertase-competent module to cytosolic-domain interactions: deletion of EMC4’s cytosolic EMC2-binding site impaired biogenesis of a canonical EMC-dependent TA client (SQS/FDFT1), supporting the functional importance of EMC2-mediated assembly interfaces even when the “catalytic” insertion module is primarily EMC3/6. (pleiner2023aselectivityfilter pages 4-6)
6.2 2024: EMC rectifies topology of multipass membrane proteins post-translationally
Wu et al. (Nature Structural & Molecular Biology; Nov 2024) report that EMC can mediate post-translational insertion/rectification of certain TMDs near the C-terminus of multipass membrane proteins, exemplified by the final TMD insertion of SOAT1. The authors propose that some substrates are released from the ribosome–translocon in an incompletely inserted state and require EMC to rectify topology and evade quality control. The paper estimates ~250 new putative EMC substrates, indicating broader client scope than previously recognized for this topology-rectification role. (wu2024emcrectifiesthe pages 7-9)
7) Current applications and real-world implementations
7.1 Pharmacological pathway dissection: Sec61 inhibitor resistance and EMC dependence
O’Keefe et al. (Communications Biology; Jul 2021) show that type III single-pass membrane proteins (including viral HIV Vpu) can integrate into the ER via an EMC-mediated pathway that is resistant to Sec61 inhibitors such as ipomoeassin F (Ipom-F) and mycolactone. In their assays, multiple type III TMPs retained N-glycosylation in 1 µM Ipom-F, and siRNA knockdown of EMC2 (and EMC5) was used to probe EMC’s contribution and destabilized the wider EMC without broadly disrupting OST activity. This provides a practical strategy used in cell biology/pharmacology: combining Sec61 inhibitors with EMC depletion to separate Sec61- versus EMC-dependent membrane insertion routes. (o’keefe2021analternativepathway pages 1-2, o’keefe2021analternativepathway pages 2-3)
7.2 Virology (2024 synthesis): EMC as a host factor in flavivirus biogenesis
A 2024 review of ER involvement in flavivirus infection summarizes evidence that dengue virus multipass proteins NS4A/NS4B depend on EMC for biogenesis: EMC interacts with NS4B during ER translocation and supports its folding/correct topology, with context dependence on upstream NS4A. The review also notes an EMC4 role in phosphatidylserine transfer at ER–endosome contacts, impacting entry steps (fusion/RNA release). While this review discusses “EMC” rather than EMC2 specifically, EMC2 is required for complex integrity and thus is part of the host machinery underlying these phenotypes. (verhaegen2024theendoplasmicreticulum pages 3-4, chitwood2019theroleof pages 2-4)
8) Relevant statistics and data (recent and foundational)
8.1 Proteome-wide client surveys (quantitative)
- Shurtleff et al. (eLife; May 2018) used unbiased SILAC proteomics in mammalian cells with CRISPRi depletion of EMC (targeting EMC2 or EMC4). They report 37 total decreased proteins, and among 11 proteins decreased ≥2-fold in both EMC2- and EMC4-depleted cells, 10 contained at least one TMD, consistent with membrane protein-specific dependence. (shurtleff2018theermembrane pages 8-10)
- Tian et al. (Cell Reports; Sep 2019) quantified 971 UniProt-annotated transmembrane proteins and identified 36 EMC-dependent (~3.7%) and 171 EMC-independent (~17.6%) membrane proteins under their criteria. Their TMT workflow identified 5,570 proteins total, retained 4,446 for analysis, and found 81 significantly changed proteins in EMC6-KO vs WT (17 up, 64 down, p<0.01). (tian2019proteomicanalysisidentifies pages 5-6, tian2019proteomicanalysisidentifies pages 3-5)
These studies provide an empirical scope estimate: EMC dependence is substantial but not universal across the transmembrane proteome, and specific client features (e.g., polar/charged TMD residues) contribute to dependence. (tian2019proteomicanalysisidentifies pages 1-3)
8.2 Quantitative substrate-scope estimate from 2024 mechanism work
Wu et al. (2024) estimate that their sequential co-/post-translational mechanism may apply to ~250 diverse multipass proteins, including pentameric ion channel family subunits relevant for neurotransmission. (wu2024emcrectifiesthe pages 7-9)
9) Disease and phenotype links (interpretation with caution)
9.1 Association-level disease links
Open Targets lists association evidence linking EMC2 (ENSG00000104412) to traits/diseases including neurodegenerative disease, asthma, and gastroesophageal reflux disease, among others. These are association-level signals and should not be interpreted as definitive causal mechanisms without gene-level functional validation and variant-to-function mapping. (OpenTargets Search: -EMC2)
9.2 Mechanistic plausibility (expert synthesis)
Given that EMC supports the biogenesis/topology of many multipass membrane proteins—many of which are receptors, channels, and transporters—perturbations of EMC integrity (including EMC2 disruption) plausibly impact signaling and homeostasis broadly via membrane proteostasis failure; however, the most reliable gene-level statements for EMC2 remain those tied to complex assembly/stability and substrate handling demonstrated in structural and depletion studies. (chitwood2019theroleof pages 2-4, pleiner2020structuralbasisfor pages 1-3)
10) Evidence-supported functional annotation statement (recommended)
EMC2 (Q15006) is best annotated as a cytosolic scaffold subunit of the ER membrane protein complex (EMC) that is required for EMC assembly and organization of the cytosolic vestibule. Through this architectural role, EMC2 enables EMC’s insertase/topogenesis functions that promote the correct insertion, topology, and stability of subsets of TA, type III, and multipass membrane proteins, including those with challenging biophysical features (e.g., low hydrophobicity or polar/charged residues in TMDs). (pleiner2020structuralbasisfor pages 1-3, odonnell2020thearchitectureof pages 14-15, tian2019proteomicanalysisidentifies pages 1-3)
Supporting figure evidence
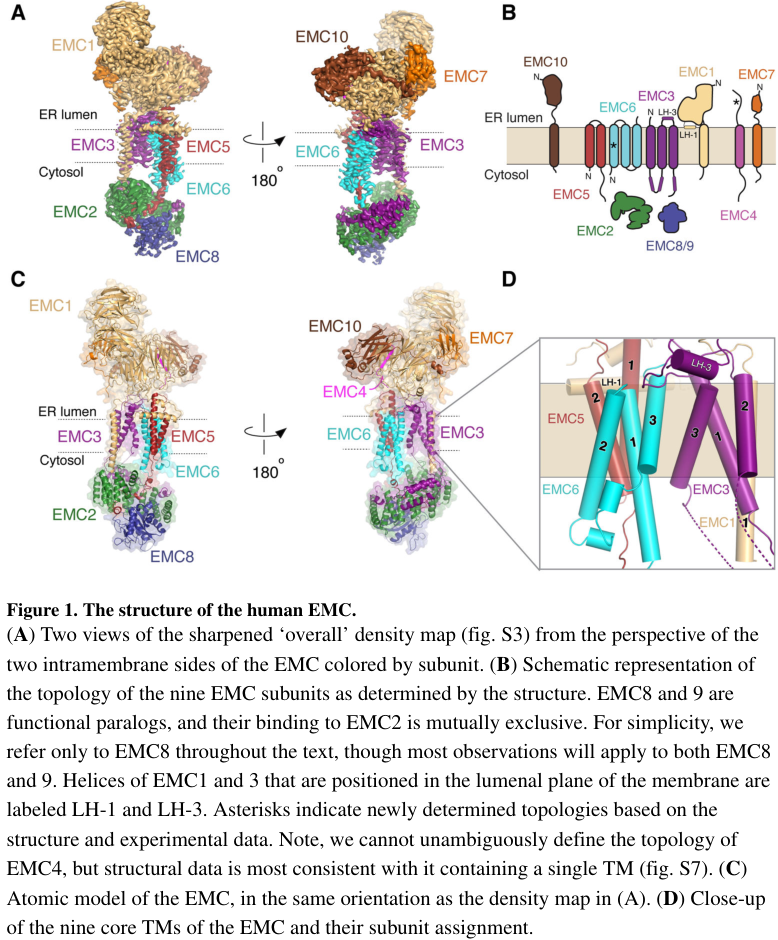
The EMC architecture and EMC2’s cytosolic placement/interfaces within the human complex are illustrated in the Pleiner et al. (Science 2020) cryo-EM figures. (pleiner2020structuralbasisfor media 452a27e0, pleiner2020structuralbasisfor media afa23ddb, pleiner2020structuralbasisfor media 9fea6c09)
Summary table (evidence map)
| Aspect | Key findings | Supporting citations |
|---|---|---|
| Identity / aliases | Human EMC2 encodes ER membrane protein complex subunit 2, also known as TTC35/KIAA0103; literature consistently identifies it as a cytosolic EMC subunit in the human ER membrane protein complex, matching UniProt Q15006. It is ~35 kDa and part of the conserved EMC core. | (chitwood2019theroleof pages 2-4, pleiner2020structuralbasisfor pages 1-3, hegde2022thefunctionstructure pages 4-6) |
| Localization | EMC2 is cytosolic-facing but tightly associated with the ER-resident EMC rather than being a free soluble factor. It sits in the cytosolic domain/vestibule of the complex adjacent to membrane subunits that form the insertase core. | (pleiner2020structuralbasisfor pages 1-3, odonnell2020thearchitectureof pages 14-15, odonnell2020thearchitectureof pages 1-2) |
| Complex membership | Human EMC is a 9-10 subunit complex depending on annotation/study context; EMC2 associates with membrane subunits plus EMC8 or EMC9 in a mutually exclusive or paralog-substitutable manner. EMC2 knockdown destabilizes other EMC components, supporting a core assembly role. | (odonnell2020thearchitectureof pages 2-4, odonnell2020thearchitectureof pages 1-2, chitwood2019theroleof pages 2-4) |
| Structural role | EMC2 forms an α-solenoid/TPR-like helical scaffold that organizes the cytosolic region. It contacts EMC3, EMC5, EMC1, and EMC8/9, and mutations at these interfaces disrupt assembly, showing EMC2 is primarily an architectural scaffold rather than the membrane-embedded catalytic insertase element. | (pleiner2020structuralbasisfor pages 1-3, pleiner2020structuralbasisfor pages 7-11) |
| Mechanistic role in insertion | EMC as a whole is a co- and post-translational insertase for selected low/moderate-hydrophobicity TMDs; EMC2 helps form the cytosolic vestibule that initially receives substrate TMDs before transfer to the EMC3/EMC6 hydrophilic vestibule in the membrane. Conserved basic residues at/near the EMC2-containing vestibule likely contribute to substrate filtering against positively charged segments. | (pleiner2020structuralbasisfor pages 1-3, odonnell2020thearchitectureof pages 14-15, odonnell2020thearchitectureof pages 1-2) |
| Recent 2023-2024 developments | 2023: EMC was shown to contain a selectivity filter that limits ER misinsertion, using a positively charged vestibule and methionine-rich capture loops; EMC4’s EMC2-binding site was functionally important for assembly. 2024: EMC was shown to rectify topology post-translationally for some multipass proteins, with an estimated ~250 new putative substrates. | (pleiner2023aselectivityfilter pages 8-10, pleiner2023aselectivityfilter pages 4-6, wu2024emcrectifiesthe pages 7-9) |
| Known client protein classes / examples | EMC-dependent proteins are enriched for multipass transporters, ATPases, some tail-anchored proteins, and selected type III membrane proteins. Example clients/contexts include SQS/FDFT1, ATP6V0A1, FZD family proteins, CB5, SGPL1, and viral or host type III TMPs such as HIV Vpu, SMAGP, BCMA, Syt1; flaviviral NS4A/NS4B biogenesis also depends on EMC contextually. | (chitwood2019theroleof pages 2-4, tian2019proteomicanalysisidentifies pages 5-6, o’keefe2021analternativepathway pages 1-2, verhaegen2024theendoplasmicreticulum pages 3-4) |
| Quantitative stats | In unbiased mammalian proteomics, 36 of 971 transmembrane proteins were classified as EMC-dependent (~3.7%), while 171 of 971 were EMC-independent (~17.6%). Tian et al. identified 5,570 proteins total, retained 4,446 for analysis, with 81 significantly changed in EMC6-KO vs WT (17 up, 64 down). Shurtleff et al. found 37 decreased proteins, and among 11 proteins decreased ≥2-fold in both EMC2- and EMC4-depleted cells, 10 were transmembrane proteins. | (tian2019proteomicanalysisidentifies pages 5-6, tian2019proteomicanalysisidentifies pages 3-5, shurtleff2018theermembrane pages 8-10) |
| Experimental systems | EMC2/EMC function has been studied using cryo-EM, mutagenesis, co-immunoprecipitation, SEC-MALS, site-specific crosslinking, reconstituted proteoliposome insertion assays, split-GFP topology reporters, glycosylation assays, SILAC/TMT proteomics, ribosome profiling, and siRNA/CRISPRi depletion in human cell systems plus in vitro rough microsomes/reticulocyte lysates. | (pleiner2020structuralbasisfor pages 1-3, pleiner2023aselectivityfilter pages 2-4, shurtleff2018theermembrane pages 8-10, tian2019proteomicanalysisidentifies pages 3-5, o’keefe2021analternativepathway pages 1-2) |
| Disease / phenotype associations | Direct, gene-specific human disease causality for EMC2 remains limited relative to complex-level biology, but EMC perturbation affects membrane-protein homeostasis, ER stress, and client stability. Experimental evidence links EMC biology to viral infection (e.g., DENV NS4A/NS4B, HIV Vpu), proteostasis, and cancer-related phenotypes in broader EMC studies; Open Targets lists low-to-moderate evidence associations for EMC2 to traits/diseases such as neurodegenerative disease, asthma, and gastroesophageal reflux disease, which should be interpreted cautiously as association-level rather than definitive mechanism. | (chitwood2019theroleof pages 2-4, OpenTargets Search: -EMC2, verhaegen2024theendoplasmicreticulum pages 3-4) |
Table: This table summarizes the verified identity, localization, structural role, mechanism, recent advances, client scope, quantitative data, methods, and disease relevance of human EMC2 (UniProt Q15006). It is useful as a compact evidence map linking EMC2 specifically to the ER membrane protein complex and membrane-protein biogenesis.
Key sources (URLs and publication dates)
- Pleiner T et al. Structural basis for membrane insertion by the human ER membrane protein complex. Science (Published Jul 2020). https://doi.org/10.1126/science.abb5008 (pleiner2020structuralbasisfor pages 1-3)
- O’Donnell JP et al. The architecture of EMC reveals a path for membrane protein insertion. eLife (Published May 2020). https://doi.org/10.7554/eLife.57887 (odonnell2020thearchitectureof pages 2-4)
- Pleiner T et al. A selectivity filter in the ER membrane protein complex limits protein misinsertion at the ER. J Cell Biol (Published May 2023). https://doi.org/10.1083/jcb.202212007 (pleiner2023aselectivityfilter pages 8-10)
- Wu H et al. EMC rectifies the topology of multipass membrane proteins. Nat Struct Mol Biol (Issue Nov 2024). https://doi.org/10.1038/s41594-023-01120-6 (wu2024emcrectifiesthe pages 7-9)
- Tian S et al. Proteomic Analysis Identifies Membrane Proteins Dependent on the ER Membrane Protein Complex. Cell Reports (Published Sep 2019). https://doi.org/10.1016/j.celrep.2019.08.006 (tian2019proteomicanalysisidentifies pages 5-6)
- Shurtleff MJ et al. The ER membrane protein complex interacts cotranslationally to enable biogenesis of multipass membrane proteins. eLife (Published May 2018). https://doi.org/10.7554/eLife.37018 (shurtleff2018theermembrane pages 8-10)
- Hegde RS. The Function, Structure, and Origins of the ER Membrane Protein Complex. Annual Review of Biochemistry (Published Jun 2022). https://doi.org/10.1146/annurev-biochem-032620-104553 (hegde2022thefunctionstructure pages 4-6)
- Verhaegen M, Vermeire K. The endoplasmic reticulum (ER): a crucial cellular hub in flavivirus infection and potential target site for antiviral interventions. npj Viruses (Published Jun 2024). https://doi.org/10.1038/s44298-024-00031-7 (verhaegen2024theendoplasmicreticulum pages 3-4)
References
-
(pleiner2020structuralbasisfor pages 1-3): Tino Pleiner, Giovani Pinton Tomaleri, Kurt Januszyk, Alison J. Inglis, Masami Hazu, and Rebecca M. Voorhees. Structural basis for membrane insertion by the human er membrane protein complex. Jul 2020. URL: https://doi.org/10.1126/science.abb5008, doi:10.1126/science.abb5008. This article has 192 citations and is from a highest quality peer-reviewed journal.
-
(pleiner2023aselectivityfilter pages 8-10): Tino Pleiner, Masami Hazu, Giovani Pinton Tomaleri, Vy N. Nguyen, Kurt Januszyk, and Rebecca M. Voorhees. A selectivity filter in the er membrane protein complex limits protein misinsertion at the er. The Journal of Cell Biology, May 2023. URL: https://doi.org/10.1083/jcb.202212007, doi:10.1083/jcb.202212007. This article has 28 citations.
-
(wu2024emcrectifiesthe pages 7-9): Haoxi Wu, Luka Smalinskaitė, and Ramanujan S. Hegde. Emc rectifies the topology of multipass membrane proteins. Nature Structural & Molecular Biology, 31:32-41, Nov 2024. URL: https://doi.org/10.1038/s41594-023-01120-6, doi:10.1038/s41594-023-01120-6. This article has 41 citations and is from a highest quality peer-reviewed journal.
-
(chitwood2019theroleof pages 2-4): Patrick J. Chitwood and Ramanujan S. Hegde. The role of emc during membrane protein biogenesis. Trends in cell biology, 29 5:371-384, May 2019. URL: https://doi.org/10.1016/j.tcb.2019.01.007, doi:10.1016/j.tcb.2019.01.007. This article has 97 citations and is from a domain leading peer-reviewed journal.
-
(hegde2022thefunctionstructure pages 4-6): Ramanujan S. Hegde. The function, structure, and origins of the er membrane protein complex. Annual Review of Biochemistry, 91:651-678, Jun 2022. URL: https://doi.org/10.1146/annurev-biochem-032620-104553, doi:10.1146/annurev-biochem-032620-104553. This article has 65 citations and is from a domain leading peer-reviewed journal.
-
(odonnell2020thearchitectureof pages 1-2): John P O'Donnell, Ben P Phillips, Yuichi Yagita, Szymon Juszkiewicz, Armin Wagner, Duccio Malinverni, Robert J Keenan, Elizabeth A Miller, and Ramanujan S Hegde. The architecture of emc reveals a path for membrane protein insertion. May 2020. URL: https://doi.org/10.7554/elife.57887, doi:10.7554/elife.57887. This article has 121 citations and is from a domain leading peer-reviewed journal.
-
(odonnell2020thearchitectureof pages 2-4): John P O'Donnell, Ben P Phillips, Yuichi Yagita, Szymon Juszkiewicz, Armin Wagner, Duccio Malinverni, Robert J Keenan, Elizabeth A Miller, and Ramanujan S Hegde. The architecture of emc reveals a path for membrane protein insertion. May 2020. URL: https://doi.org/10.7554/elife.57887, doi:10.7554/elife.57887. This article has 121 citations and is from a domain leading peer-reviewed journal.
-
(pleiner2023aselectivityfilter pages 1-2): Tino Pleiner, Masami Hazu, Giovani Pinton Tomaleri, Vy N. Nguyen, Kurt Januszyk, and Rebecca M. Voorhees. A selectivity filter in the er membrane protein complex limits protein misinsertion at the er. The Journal of Cell Biology, May 2023. URL: https://doi.org/10.1083/jcb.202212007, doi:10.1083/jcb.202212007. This article has 28 citations.
-
(pleiner2020structuralbasisfor pages 7-11): Tino Pleiner, Giovani Pinton Tomaleri, Kurt Januszyk, Alison J. Inglis, Masami Hazu, and Rebecca M. Voorhees. Structural basis for membrane insertion by the human er membrane protein complex. Jul 2020. URL: https://doi.org/10.1126/science.abb5008, doi:10.1126/science.abb5008. This article has 192 citations and is from a highest quality peer-reviewed journal.
-
(odonnell2020thearchitectureof pages 14-15): John P O'Donnell, Ben P Phillips, Yuichi Yagita, Szymon Juszkiewicz, Armin Wagner, Duccio Malinverni, Robert J Keenan, Elizabeth A Miller, and Ramanujan S Hegde. The architecture of emc reveals a path for membrane protein insertion. May 2020. URL: https://doi.org/10.7554/elife.57887, doi:10.7554/elife.57887. This article has 121 citations and is from a domain leading peer-reviewed journal.
-
(shurtleff2018theermembrane pages 8-10): Matthew J Shurtleff, Daniel N Itzhak, Jeffrey A Hussmann, Nicole T Schirle Oakdale, Elizabeth A Costa, Martin Jonikas, Jimena Weibezahn, Katerina D Popova, Calvin H Jan, Pavel Sinitcyn, Shruthi S Vembar, Hilda Hernandez, Jürgen Cox, Alma L Burlingame, Jeffrey L Brodsky, Adam Frost, Georg HH Borner, and Jonathan S Weissman. The er membrane protein complex interacts cotranslationally to enable biogenesis of multipass membrane proteins. eLife, May 2018. URL: https://doi.org/10.7554/elife.37018, doi:10.7554/elife.37018. This article has 257 citations and is from a domain leading peer-reviewed journal.
-
(pleiner2023aselectivityfilter pages 2-4): Tino Pleiner, Masami Hazu, Giovani Pinton Tomaleri, Vy N. Nguyen, Kurt Januszyk, and Rebecca M. Voorhees. A selectivity filter in the er membrane protein complex limits protein misinsertion at the er. The Journal of Cell Biology, May 2023. URL: https://doi.org/10.1083/jcb.202212007, doi:10.1083/jcb.202212007. This article has 28 citations.
-
(pleiner2023aselectivityfilter pages 4-6): Tino Pleiner, Masami Hazu, Giovani Pinton Tomaleri, Vy N. Nguyen, Kurt Januszyk, and Rebecca M. Voorhees. A selectivity filter in the er membrane protein complex limits protein misinsertion at the er. The Journal of Cell Biology, May 2023. URL: https://doi.org/10.1083/jcb.202212007, doi:10.1083/jcb.202212007. This article has 28 citations.
-
(o’keefe2021analternativepathway pages 1-2): Sarah O’Keefe, Guanghui Zong, Kwabena B. Duah, Lauren E. Andrews, Wei Q. Shi, and Stephen High. An alternative pathway for membrane protein biogenesis at the endoplasmic reticulum. Communications Biology, Jul 2021. URL: https://doi.org/10.1038/s42003-021-02363-z, doi:10.1038/s42003-021-02363-z. This article has 61 citations and is from a peer-reviewed journal.
-
(o’keefe2021analternativepathway pages 2-3): Sarah O’Keefe, Guanghui Zong, Kwabena B. Duah, Lauren E. Andrews, Wei Q. Shi, and Stephen High. An alternative pathway for membrane protein biogenesis at the endoplasmic reticulum. Communications Biology, Jul 2021. URL: https://doi.org/10.1038/s42003-021-02363-z, doi:10.1038/s42003-021-02363-z. This article has 61 citations and is from a peer-reviewed journal.
-
(verhaegen2024theendoplasmicreticulum pages 3-4): Marijke Verhaegen and Kurt Vermeire. The endoplasmic reticulum (er): a crucial cellular hub in flavivirus infection and potential target site for antiviral interventions. npj Viruses, Jun 2024. URL: https://doi.org/10.1038/s44298-024-00031-7, doi:10.1038/s44298-024-00031-7. This article has 37 citations.
-
(tian2019proteomicanalysisidentifies pages 5-6): Songhai Tian, Quan Wu, Bo Zhou, Mei Yuk Choi, Bo Ding, Wei Yang, and Min Dong. Proteomic analysis identifies membrane proteins dependent on the er membrane protein complex. Cell reports, 28:2517-2526.e5, Sep 2019. URL: https://doi.org/10.1016/j.celrep.2019.08.006, doi:10.1016/j.celrep.2019.08.006. This article has 79 citations and is from a highest quality peer-reviewed journal.
-
(tian2019proteomicanalysisidentifies pages 3-5): Songhai Tian, Quan Wu, Bo Zhou, Mei Yuk Choi, Bo Ding, Wei Yang, and Min Dong. Proteomic analysis identifies membrane proteins dependent on the er membrane protein complex. Cell reports, 28:2517-2526.e5, Sep 2019. URL: https://doi.org/10.1016/j.celrep.2019.08.006, doi:10.1016/j.celrep.2019.08.006. This article has 79 citations and is from a highest quality peer-reviewed journal.
-
(tian2019proteomicanalysisidentifies pages 1-3): Songhai Tian, Quan Wu, Bo Zhou, Mei Yuk Choi, Bo Ding, Wei Yang, and Min Dong. Proteomic analysis identifies membrane proteins dependent on the er membrane protein complex. Cell reports, 28:2517-2526.e5, Sep 2019. URL: https://doi.org/10.1016/j.celrep.2019.08.006, doi:10.1016/j.celrep.2019.08.006. This article has 79 citations and is from a highest quality peer-reviewed journal.
-
(OpenTargets Search: -EMC2): Open Targets Query (-EMC2, 5 results). Buniello, A. et al. (2025). Open Targets Platform: facilitating therapeutic hypotheses building in drug discovery. Nucleic Acids Research.
-
(pleiner2020structuralbasisfor media 452a27e0): Tino Pleiner, Giovani Pinton Tomaleri, Kurt Januszyk, Alison J. Inglis, Masami Hazu, and Rebecca M. Voorhees. Structural basis for membrane insertion by the human er membrane protein complex. Jul 2020. URL: https://doi.org/10.1126/science.abb5008, doi:10.1126/science.abb5008. This article has 192 citations and is from a highest quality peer-reviewed journal.
-
(pleiner2020structuralbasisfor media afa23ddb): Tino Pleiner, Giovani Pinton Tomaleri, Kurt Januszyk, Alison J. Inglis, Masami Hazu, and Rebecca M. Voorhees. Structural basis for membrane insertion by the human er membrane protein complex. Jul 2020. URL: https://doi.org/10.1126/science.abb5008, doi:10.1126/science.abb5008. This article has 192 citations and is from a highest quality peer-reviewed journal.
-
(pleiner2020structuralbasisfor media 9fea6c09): Tino Pleiner, Giovani Pinton Tomaleri, Kurt Januszyk, Alison J. Inglis, Masami Hazu, and Rebecca M. Voorhees. Structural basis for membrane insertion by the human er membrane protein complex. Jul 2020. URL: https://doi.org/10.1126/science.abb5008, doi:10.1126/science.abb5008. This article has 192 citations and is from a highest quality peer-reviewed journal.
Artifacts
Citations
- pleiner2020structuralbasisfor pages 1-3
- odonnell2020thearchitectureof pages 14-15
- pleiner2023aselectivityfilter pages 4-6
- wu2024emcrectifiesthe pages 7-9
- shurtleff2018theermembrane pages 8-10
- tian2019proteomicanalysisidentifies pages 1-3
- odonnell2020thearchitectureof pages 2-4
- pleiner2023aselectivityfilter pages 8-10
- tian2019proteomicanalysisidentifies pages 5-6
- hegde2022thefunctionstructure pages 4-6
- verhaegen2024theendoplasmicreticulum pages 3-4
- chitwood2019theroleof pages 2-4
- odonnell2020thearchitectureof pages 1-2
- pleiner2023aselectivityfilter pages 1-2
- pleiner2020structuralbasisfor pages 7-11
- pleiner2023aselectivityfilter pages 2-4
- tian2019proteomicanalysisidentifies pages 3-5
- https://doi.org/10.1126/science.abb5008
- https://doi.org/10.7554/eLife.57887
- https://doi.org/10.1083/jcb.202212007
- https://doi.org/10.1038/s41594-023-01120-6
- https://doi.org/10.1016/j.celrep.2019.08.006
- https://doi.org/10.7554/eLife.37018
- https://doi.org/10.1146/annurev-biochem-032620-104553
- https://doi.org/10.1038/s44298-024-00031-7
- https://doi.org/10.1126/science.abb5008,
- https://doi.org/10.1083/jcb.202212007,
- https://doi.org/10.1038/s41594-023-01120-6,
- https://doi.org/10.1016/j.tcb.2019.01.007,
- https://doi.org/10.1146/annurev-biochem-032620-104553,
- https://doi.org/10.7554/elife.57887,
- https://doi.org/10.7554/elife.37018,
- https://doi.org/10.1038/s42003-021-02363-z,
- https://doi.org/10.1038/s44298-024-00031-7,
- https://doi.org/10.1016/j.celrep.2019.08.006,
OpenScientist
(EMC2-deep-research-openscientist.md)
OpenScientist
(EMC2-deep-research-openscientist.md)EMC2 (ER Membrane Protein Complex Subunit 2): Comprehensive Functional Annotation
Summary
EMC2 (UniProt Q15006; synonyms TTC35, KIAA0103) is a 297-amino acid cytoplasmic scaffold protein that serves as the organizational hub of the ER membrane protein complex (EMC), an essential and evolutionarily ancient multi-subunit insertase residing at the endoplasmic reticulum (ER) membrane. EMC2 contains three tetratricopeptide repeat (TPR) motifs arranged in an α-helical solenoid fold and is classified as a peripheral membrane protein — it does not span the lipid bilayer itself but instead anchors to the cytoplasmic face of the ER through extensive interactions with the transmembrane subunits EMC3, EMC5, and the soluble partners EMC8/EMC9. Its primary molecular function is to form the cytoplasmic vestibule that captures client transmembrane domains (TMDs) from the cytosol and channels them to the membrane-embedded insertase subunit EMC3 for energy-independent membrane insertion.
The EMC inserts two major classes of substrates: (1) tail-anchored (TA) proteins with moderately hydrophobic C-terminal TMDs that cannot be engaged by the canonical GET/TRC40 pathway, and (2) the first TMD of multipass membrane proteins (including G protein-coupled receptors) in the correct N-exo/C-cyto topology, enforcing the "positive-inside rule." Through these activities, EMC2 is indirectly required for cholesterol homeostasis (via biogenesis of squalene synthase and SOAT1), GPCR signaling, rhodopsin biosynthesis and photoreceptor survival, voltage-gated ion channel assembly, and ER–mitochondria lipid transfer. EMC2 is a common essential gene in human cells, is ubiquitously expressed across more than 210 cell types, and its complex is conserved across all major eukaryotic lineages since the last eukaryotic common ancestor (LECA). Functionally, the EMC is also coupled to ER-associated degradation (ERAD) quality control and serves as a host dependency factor exploited by flaviviruses for infection.
This report synthesizes evidence from cryo-electron microscopy structural studies, site-directed mutagenesis, reconstituted biochemical assays, CRISPR genetic screens, comparative genomics, and disease genetics to provide a comprehensive functional annotation of human EMC2.
1. Gene and Protein Identity
| Property | Value |
|---|---|
| Gene | EMC2 (Ensembl: ENSG00000104412) |
| Chromosomal location | 8q23.1 (chr8:108,443,601–108,551,893, GRCh38, forward strand) |
| Synonyms | TTC35, KIAA0103 |
| UniProt | Q15006 |
| Organism | Homo sapiens |
| Protein family | EMC2 family (IPR039856) |
| Protein length | 297 amino acids |
| Key domains | Three TPR motifs (aa 87–120, 155–188, 192–225); EMC2-like domain (IPR055217); TPR-like helical domain superfamily (IPR011990) |
| Gene essentiality | Common essential gene (DepMap CRISPR screens; Chronos score ~−0.8 to −1.0) |
The gene symbol "EMC2" unambiguously refers to this ER membrane protein complex subunit in humans. The protein was originally identified as KIAA0103 in early cDNA sequencing projects and later named TTC35 based on its TPR repeats, before being renamed EMC2 following the characterization of the EMC complex.
2. The ER Membrane Protein Complex (EMC)
2.1 Discovery and Conservation
The EMC was first identified in a systematic yeast genetic screen for factors involved in ER protein folding. Wideman (2015) subsequently demonstrated through comprehensive homology searching that the EMC is "truly an ancient and conserved protein complex, present in every major eukaryotic lineage. Very few organisms have completely lost the EMC, and most, even over 2 billion years of eukaryote evolution, have retained a majority of the complex members" (PMID: 26512320). The EMC was present in the last eukaryote common ancestor (LECA), underscoring its fundamental importance to eukaryotic cell biology.
2.2 Complex Composition and Architecture
The human EMC consists of 9–10 subunits with distinct structural and functional roles:
| Subunit | Topology | Key Role |
|---|---|---|
| EMC1 | Type I TM, large lumenal β-propeller | Lumenal client folding; disease-associated |
| EMC2 | Peripheral/cytoplasmic (TPR scaffold) | Cytoplasmic substrate capture and complex organization |
| EMC3 | Multi-TM, YidC/Oxa1-like fold | Core insertase; forms lipid-exposed membrane groove |
| EMC4 | TM | Membrane groove, lipid transfer |
| EMC5/MMGT1 | TM | Hydrophilic vestibule component |
| EMC6 | TM | Gating regulation |
| EMC7 | Type I TM | Lumenal functions |
| EMC8 | Cytoplasmic | Binds EMC2, cytoplasmic subcomplex |
| EMC9 | Cytoplasmic | Binds EMC2, paralog of EMC8 |
| EMC10 | TM | Regulatory, least conserved |
Multiple cryo-EM structures of the full human EMC have been determined, including in lipid nanodiscs at 3.4 Å resolution (PDB: 6WW7; O'Donnell et al., 2020; PMID: 32459176) and in apo and VDAC-bound states (PDB: 8J0N, 8J0O; Li et al., 2024; PMID: 38517390). These structures reveal a tripartite architecture: a large lumenal domain (dominated by EMC1), a transmembrane region containing a lipid-exposed hydrophilic groove (centered on EMC3, which shares structural homology with the YidC/Oxa1 superfamily of membrane protein insertases; PMID: 35850079; PMID: 32910895), and a cytoplasmic domain forming a moderately hydrophobic vestibule for substrate capture (organized by EMC2).
3. EMC2: Structural Role and Molecular Function
3.1 Cytoplasmic Scaffold Architecture
EMC2 is entirely cytoplasmic, classified as a peripheral membrane protein associated with the ER membrane. Its three TPR repeats create a concave α-helical solenoid structure (~20 helices with two short β-strands), as revealed by the 2.2 Å crystal structure of the EMC2–EMC9 subcomplex (PDB: 6Y4L; PMID: 32459176).
The TPR motifs function as protein–protein interaction domains. In the context of the EMC, EMC2 serves as the central organizing hub for the cytoplasmic face of the complex, with experimentally validated interactions:
- EMC8 interaction: 17 experimental validations
- EMC9 interaction: 14 experimental validations (paralog of EMC8, competes for the same binding site)
- EMC3 interaction: 12 experimental validations (bridges to the membrane-embedded insertase)
- EMC5/MMGT1 interaction: 13 experimental validations
3.2 Role in Substrate TMD Capture
The cryo-EM and crosslinking studies by O'Donnell et al. (2020) revealed that "EMC's cytosolic domain contains a large, moderately hydrophobic vestibule that can bind a substrate's transmembrane domain (TMD). The cytosolic vestibule leads into a lumenally-sealed, lipid-exposed intramembrane groove large enough to accommodate a single substrate TMD" (PMID: 32459176). EMC2 forms a substantial portion of this cytoplasmic vestibule.
Pleiner et al. (2023) mapped the substrate path in detail using site-specific crosslinking, showing that client TA proteins are first captured by methionine-rich loops on the cytoplasmic face, then threaded through a hydrophilic vestibule into the membrane. "Positively charged residues at the entrance to the vestibule function as a selectivity filter that uses charge-repulsion to reject mitochondrial TA proteins. Similarly, this selectivity filter retains the positively charged soluble domains of multipass substrates in the cytosol, thereby ensuring they adopt the correct topology and enforcing the 'positive-inside' rule" (PMID: 37199759).
3.3 Functional Residue Mapping by Mutagenesis
Site-directed mutagenesis studies (catalogued in UniProt Q15006, largely from O'Donnell et al. 2020) have mapped the functional surfaces of EMC2 at single-residue resolution:
| Functional Surface | Key Residues | Effect of Mutation |
|---|---|---|
| EMC5 binding | 28, 156, 160, 227 | Loss of EMC5 interaction |
| EMC8 binding | 171, 200, 227 | Decreased/abolished EMC8 interaction |
| EMC3 binding | 180, 259 | Decreased EMC3 interaction |
| Substrate TMD capture | 189, 190, 191 | Decreased TA protein TMD binding |
| No effect on TMD binding | 61, 95, 122, 193, 194 | No detectable change |
The substrate-binding residues (189–191) lie in the TPR3 region, and their proximity to the EMC3-contact residue (180) provides molecular evidence that EMC2 bridges substrate capture and membrane insertion at a defined structural junction. The EMC5 and EMC8 binding sites are distributed along the TPR solenoid, consistent with EMC2 serving as a multi-armed scaffold connecting cytoplasmic and membrane subunits.
{{figure:emc2_domain_map.png|caption=EMC2 functional domain map showing TPR repeats (blue), partner binding sites for EMC3/EMC5/EMC8, substrate TMD interaction residues (189-191), and post-translational modifications. Residue-level annotations are derived from UniProt Q15006 mutagenesis data and cryo-EM structural studies (O'Donnell et al. 2020).}}
3.4 Post-Translational Modifications
EMC2 undergoes N-terminal acetylation at Ala2 (after initiator methionine removal) and lysine acetylation at Lys255. Lys255 is located in the C-terminal helix (residues 247–274) near the EMC3-binding residue (259), suggesting potential regulation of the EMC2–EMC3 interface, though this has not been functionally tested. The best-characterized regulatory modification is ubiquitination of unassembled EMC2, which leads to proteasomal degradation and is prevented by WNK1 binding (PMID: 33964204).
3.5 EMC2 Assembly and Quality Control by WNK1
A surprising finding from Pleiner et al. (2021) revealed that the kinase WNK1 (with no lysine kinase 1) moonlights as an essential assembly factor for the EMC. WNK1 "uses a conserved amphipathic helix to stabilize the soluble subunit, EMC2, by binding to the EMC2-8 interface. Shielding this hydrophobic surface prevents promiscuous interactions of unassembled EMC2 and directly competes for binding of E3 ubiquitin ligases, permitting assembly" (PMID: 33964204). Without WNK1, free EMC2 is ubiquitinated and degraded by the proteasome. Depletion of WNK1 destabilizes both the EMC and its membrane protein clients. This quality control mechanism ensures that only properly assembled EMC complexes persist, highlighting EMC2's centrality — the entire complex depends on its successful incorporation.
4. Primary Functions of the EMC (Including EMC2)
4.1 Post-Translational Insertion of Tail-Anchored Proteins
The landmark study by Guna et al. (2018) established the EMC as a bona fide transmembrane domain insertase. They demonstrated that "known membrane insertion pathways fail to effectively engage tail-anchored membrane proteins with moderately hydrophobic transmembrane domains. These proteins are instead shielded in the cytosol by calmodulin. Dynamic release from calmodulin allowed sampling of the endoplasmic reticulum (ER), where the conserved ER membrane protein complex (EMC) was shown to be essential for efficient insertion in vitro and in cells" (PMID: 29242231). Critically, purified EMC in synthetic liposomes catalyzed the insertion of its substrates, proving direct insertase activity.
The EMC operates in parallel with the GET/TRC40 pathway but handles a distinct substrate class: TA proteins whose TMDs are moderately hydrophobic — too weak for the GET pathway but requiring assistance for membrane insertion. Jung & Zimmermann (2023) used quantitative proteomics to systematically characterize the client spectra of these pathways, confirming that each handles a distinct subset of membrane proteins (PMID: 37762469). Structural comparisons reveal that both EMC and GET insertases share a conserved hydrophilic groove mechanism, suggesting divergent evolution from a common ancestor (PMID: 35850079; PMID: 32910895).
4.2 Cotranslational Insertion of Multipass Membrane Proteins
Beyond TA proteins, the EMC plays a critical role in the cotranslational biogenesis of multipass membrane proteins. It mediates the insertion of the first TMD of multipass proteins in the correct N-exo topology (N-terminus in the ER lumen), enforcing the "positive-inside rule" that governs membrane protein topology (PMID: 37199759). Miller-Vedam et al. (2020) resolved cryo-EM structures of both yeast and human EMC that "reveal conserved intricate assemblies and human-specific features associated with pathologies. Structure-based functional studies distinguish between two separable EMC activities, as an insertase regulating tail-anchored protein levels and a broader role in polytopic membrane protein biogenesis" (PMID: 33236988).
4.3 Cooperation with the Multipass Translocon
Page et al. (2024) demonstrated that the EMC physically and genetically interacts with the back of Sec61 (BOS) complex, a component of the multipass translocon. They proposed "a unifying model for coordination between the EMC, the multipass translocon, and Sec61 for the biogenesis of diverse membrane proteins in human cells" (PMID: 38076791). This places EMC2 within a larger biogenesis network where the EMC inserts the first TMD of multipass proteins and then hands off partially inserted substrates to Sec61/BOS for completion.
4.4 Chaperone Function for Membrane Protein Complex Assembly
The most recent work by Stanton et al. (2026) demonstrated that the EMC acts as a chaperone beyond its insertase function, "facilitating the assembly of heterotrimeric voltage-gated calcium channels" at the ER membrane (PMID: 41648177). This extends EMC function beyond simple insertion to include holding and protecting unassembled membrane protein subunits, facilitating their stoichiometric assembly.
Additionally, Li et al. (2024) resolved cryo-EM structures of human EMC in apo and VDAC-bound states, revealing "a specific interaction between VDAC proteins and the EMC at mitochondria-ER contact sites, which is conserved from yeast to humans. Moreover, [they] identified a gating plug located inside the EMC hydrophilic vestibule, the substrate-binding pocket for client insertion" (PMID: 38517390). This gating plug may regulate the switch between the EMC's insertase and chaperone modes.
5. Subcellular Localization
EMC2 localizes to the cytoplasmic face of the endoplasmic reticulum membrane (GO:0072546, EMC complex; GO:0042406, extrinsic component of ER membrane). It is not itself a transmembrane protein but is tethered to the ER membrane through its extensive interactions with the transmembrane EMC subunits (EMC3, EMC5).
Within the cell, the EMC complex is found at:
- ER membrane: Primary site of function (protein insertion and chaperone activity)
- ER–mitochondria contact sites (MAMs): The EMC interacts with VDAC at these contact sites (PMID: 38517390), and with SLC25A46 to facilitate phospholipid transfer between the ER and mitochondria (PMID: 27390132)
EMC2 is ubiquitously expressed across human tissues, detected in more than 210 cell types and tissues (Bgee database, ENSG00000104412), consistent with its fundamental role in ER membrane protein biogenesis.
6. Physiological Roles and Client Specificity
6.1 Cholesterol Homeostasis
Volkmar et al. (2019) demonstrated that "insertion of the weakly hydrophobic tail-anchor (TA) of SQS into the ER membrane by the EMC ensures sufficient flux through the sterol biosynthetic pathway while biogenesis of polytopic SOAT1 promoted by the EMC provides cells with the ability to store free cholesterol as inert cholesteryl esters" (PMID: 30578317). EMC deficiency causes diminished cell viability under both limiting and excessive extracellular cholesterol, demonstrating that the EMC is a key biogenic determinant of cellular cholesterol tolerance.
6.2 Rhodopsin Biosynthesis and Photoreceptor Survival
EMC subunits are essential for rhodopsin (Rh1) stabilization. Satoh et al. (2015) showed that "dPob/EMC3, EMC1, and EMC8/9, Drosophila homologs of subunits of ER membrane protein complex (EMC), are essential for stabilization of immature Rh1 in an earlier step than that at which another Rh1-specific chaperone (NinaA) acts" (PMID: 25715730). Xiong et al. (2020) confirmed this in mammals: "Conditional knockout of the Emc3 gene in mice led to mislocalization of rhodopsin protein and death of cone and rod photoreceptor cells" (PMID: 31263175). Hiramatsu et al. (2019) further showed that the EMC specifically facilitates insertion of late-synthesized transmembrane helices of Rh1 (PMID: 31553680).
6.3 GPCR Biogenesis
GPCRs, the largest family of human membrane receptors (~800 members), are key EMC clients. The EMC inserts the first TMD of GPCRs in the correct N-exo orientation, which is essential for subsequent folding of the remaining TMDs. Page et al. (2024) showed that characteristics of a GPCR's soluble domain determine its biogenesis pathway, with the EMC, multipass translocon, and Sec61 coordinating (PMID: 38076791).
6.4 Viral Membrane Protein Biogenesis
The EMC, including EMC2/TTC35, was identified as a host dependency factor for flaviviruses. Barrows et al. (2019) showed that "TTC35 and TMEM111, which we previously demonstrated to be required for yellow fever virus (YFV) infection and others subsequently showed were also required by other flaviviruses. These proteins are components of the human endoplasmic reticulum membrane protein complex (EMC)" (PMID: 31273220). Savidis et al. (2016) independently confirmed that "both flaviviruses require the EMC for their early stages of infection" (PMID: 27342126). Bagchi et al. (2022) further showed that EMC4 specifically promotes DENV–endosomal membrane fusion by mediating ER-to-endosome transfer of phosphatidylserine (PMID: 35834589).
6.5 Functional Connection to ER-Associated Degradation (ERAD)
STRING network analysis reveals high-confidence interactions between EMC2 and ERAD components DERL2 (Derlin-2, score = 0.904) and UBAC2 (score = 0.847). A chemogenomic screen by Raj et al. (2015) found that "the set of mutants conferring sensitivity to sr7575 was strikingly narrow, affecting components of the endoplasmic reticulum-associated protein degradation (ERAD) stress response and the ER membrane protein complex (EMC). ERAD-deficient mutants were hypersensitive to sr7575 in both S. cerevisiae and A. fumigatus, indicating a conserved mechanism of growth inhibition between yeast and filamentous fungi" (PMID: 26666917). This functional coupling likely reflects the need for coordinated biogenesis (EMC) and quality control (ERAD) of membrane proteins at the ER.
7. Mechanistic Model
The following model summarizes the role of EMC2 within the EMC insertase complex:
CYTOSOL
│
┌────────────────────┼────────────────────┐
│ │ │
│ Calmodulin ──► release of TA protein │
│ │ │
│ ┌───────────▼───────────┐ │
│ │ EMC2 VESTIBULE │ │
│ │ (TPR solenoid fold) │ │
│ │ │ │
│ │ EMC8/9 ◄──► EMC2 │ │
│ │ (soluble) │ │ │
│ │ │ res │ │
│ │ WNK1 ──► │ 189- │ │
│ │ (assembly │ 191 │ │
│ │ factor) │ (TMD │ │
│ │ │ bind) │ │
│ │ ▼ │ │
│ │ res 180/259 │ │
│ │ (EMC3 contact)│ │
│ └──────────┬───────────┘ │
│ │ │
═════╪═══════════════════╪════════════════════╪═══
│ ER MEMBRANE │ │
│ ▼ │
│ ┌──────────────────┐ │
│ │ EMC3 INSERTASE │ │
│ │ (hydrophilic │ │
│ │ groove, lipid- │◄── EMC1 │
│ │ exposed) │ EMC5 │
│ │ │ EMC4 │
│ │ Selectivity │ EMC6 │
│ │ filter (+charge │ EMC7 │
│ │ repulsion) │ EMC10 │
│ └────────┬─────────┘ │
│ │ │
│ ▼ │
│ Inserted TMD in │
│ correct topology │
│ (N-exo, positive-inside) │
│ │
└────────────────────────────────────────┘
ER LUMEN
Clients: TA proteins (SQS, VAMP7, etc.)
Multipass proteins (GPCRs, rhodopsin, CaV channels)
Coupled pathways:
→ GET/TRC40 (parallel: handles strongly hydrophobic TA TMDs)
→ BOS/Sec61 (sequential: handles downstream TMDs of multipass proteins)
→ ERAD (quality control: degrades misfolded EMC clients)
Substrate selection logic: The EMC vestibule (formed largely by EMC2) captures TMDs from the cytosol. A charge-based selectivity filter at the vestibule entrance uses electrostatic repulsion to reject mitochondrial TA proteins (which have net negative flanking charges) while accepting ER-destined substrates and enforcing the positive-inside topology rule for multipass proteins. After capture, the TMD is handed off through the EMC2–EMC3 interface into a lipid-exposed, lumenally-sealed intramembrane groove in EMC3 for lateral release into the ER membrane.
8. Pathway Context
EMC2 is notably absent from canonical signaling pathway databases (no KEGG or Reactome pathway annotations). This reflects its role as core biogenesis machinery for ER membrane proteins — a "housekeeping" complex that impacts many downstream pathways indirectly:
| Pathway/Process | EMC Role | Key Clients |
|---|---|---|
| Membrane protein insertion | Primary insertase for moderate-hydrophobicity TMDs | TA proteins, first TMD of multipass proteins |
| GET/TRC40 pathway | Parallel, complementary pathway (handles strong TMDs) | TA proteins with high hydrophobicity |
| Sec61/BOS translocon | Sequential cooperator for multipass protein biogenesis | Downstream TMDs of GPCRs |
| Cholesterol biosynthesis | Biogenesis of pathway enzymes | SQS/FDFT1 (TA protein), SOAT1 (polytopic) |
| Phototransduction | Rhodopsin biogenesis | Rhodopsin/Rh1 |
| Ion channel assembly | Chaperone for complex formation | Voltage-gated Ca²⁺ channels |
| ERAD | Functional coupling; quality control of EMC clients | Misfolded membrane proteins |
| ER–mitochondria communication | Lipid transfer at contact sites | VDAC, SLC25A46 |
9. Disease Associations
9.1 EMC Subunit Mutations in Human Disease
While no Mendelian disease has been directly attributed to EMC2 mutations (likely reflecting embryonic lethality of homozygous loss, consistent with its essential gene classification), mutations in related EMC subunits cause severe disease:
- EMC1 mutations cause neurodevelopmental syndrome with global developmental delay, severe hypotonia, cerebellar atrophy, and visual impairment (PMID: 37187958; PMID: 38784058)
- EMC3 mutations have been identified as a candidate gene for inherited retinal dystrophies (PMID: 37809982)
- EMC loss causes photoreceptor degeneration in both flies and mice (PMID: 25715730; PMID: 31263175)
9.2 Cancer Prognostic Associations
EMC2 has been identified as a prognostic indicator in several cancer types:
- In acute myeloid leukemia, EMC2 was identified among significant prognostic genes, with "high transcript abundance correlating with poor outcomes" (PMID: 39311489)
- In breast cancer, EMC2 appears in ferroptosis-related prognostic gene signatures (PMID: 34956895; PMID: 34059009)
- TTC35 (EMC2) expression is significantly altered with pancreatic tumor grade (PMID: 34711879)
The mechanistic basis for these associations likely reflects EMC2's essential role in membrane protein biogenesis (including iron-handling proteins like SQS/FDFT1) rather than a direct role in ferroptosis signaling, though this remains to be directly demonstrated.
9.3 Viral Infection
As a flavivirus host dependency factor, the EMC (including EMC2) represents a potential target for antiviral intervention against dengue, Zika, and yellow fever viruses (PMID: 31273220; PMID: 27342126).
10. Key Structural Evidence
| PDB ID | Method | Resolution | Contents | Reference |
|---|---|---|---|---|
| 6Y4L | X-ray | 2.2 Å | EMC2–EMC9 subcomplex | O'Donnell et al. 2020 |
| 6WW7 | Cryo-EM | 3.4 Å | Full human EMC in nanodisc | O'Donnell et al. 2020 |
| 7ADO | Cryo-EM | 3.39 Å | Human EMC | Miller-Vedam et al. 2020 |
| 8J0N | Cryo-EM | 3.47 Å | Human EMC apo state | Li et al. 2024 |
| 8J0O | Cryo-EM | 3.32 Å | Human EMC + VDAC | Li et al. 2024 |
11. Evidence Base
Structural Studies
| Study | Key Contribution | PMID |
|---|---|---|
| O'Donnell et al. 2020 | Architecture of human EMC; cytoplasmic vestibule; EMC2–EMC9 crystal structure | 32459176 |
| Miller-Vedam et al. 2020 | Yeast and human EMC structures; dual insertase/chaperone activities | 33236988 |
| Li et al. 2024 | Human EMC apo and VDAC-bound; gating plug mechanism | 38517390 |
| McDowell et al. 2020 | GET insertase structure; structural homology with EMC | 32910895 |
Functional/Biochemical Studies
| Study | Key Finding | PMID |
|---|---|---|
| Guna et al. 2018 | EMC is a TA protein insertase for moderate-hydrophobicity TMDs | 29242231 |
| Pleiner et al. 2023 | Charge-based selectivity filter; positive-inside rule enforcement | 37199759 |
| Pleiner et al. 2021 | WNK1 as EMC2 assembly factor | 33964204 |
| Page et al. 2024 | EMC•BOS holocomplex for GPCR biogenesis | 38076791 |
| Volkmar et al. 2019 | EMC required for cholesterol homeostasis via SQS and SOAT1 | 30578317 |
| Stanton et al. 2026 | EMC chaperone activity for Ca²⁺ channel assembly | 41648177 |
| Jung & Zimmermann 2023 | Systematic characterization of EMC client spectrum | 37762469 |
Physiological/Disease Studies
| Study | Key Finding | PMID |
|---|---|---|
| Satoh et al. 2015 | EMC essential for rhodopsin biosynthesis in Drosophila | 25715730 |
| Xiong et al. 2020 | Emc3 knockout causes photoreceptor death in mice | 31263175 |
| Hiramatsu et al. 2019 | EMC facilitates late TMD insertions of Rh1 | 31553680 |
| Barrows et al. 2019 | EMC2/TTC35 is a flavivirus host dependency factor | 31273220 |
| Savidis et al. 2016 | EMC required for ZIKV/DENV infection | 27342126 |
| Wideman 2015 | EMC conserved since LECA | 26512320 |
| Wang et al. 2023 | EMC1 mutations cause neurodevelopmental disease | 37187958 |
| Raj et al. 2015 | EMC and ERAD functional coupling in chemogenomic screen | 26666917 |
| Janer et al. 2016 | SLC25A46–EMC interaction in mitochondrial lipid homeostasis | 27390132 |
12. Limitations and Knowledge Gaps
-
No direct substrate-bound structure: No structure of EMC2 bound to a client TMD has been resolved, leaving the precise substrate capture geometry partially inferred from crosslinking and mutagenesis data.
-
Regulation of EMC2 function is poorly understood: The acetylation at Lys255 near the EMC3-binding interface hints at post-translational regulation, but the responsible enzyme(s) and functional consequences have not been characterized.
-
Client specificity determinants incompletely defined: While it is known that EMC handles moderate-hydrophobicity TMDs and the GET pathway handles strongly hydrophobic ones, the precise biophysical thresholds and how EMC2's vestibule discriminates substrates remain incompletely defined.
-
Disease associations are largely correlative: EMC2's appearances in ferroptosis/cancer prognostic gene signatures likely reflect its essential role in membrane protein biogenesis rather than a direct role in ferroptosis. Mechanistic studies are needed.
-
Gating plug dynamics unresolved: The gating plug inside the EMC hydrophilic vestibule may regulate switching between insertase and chaperone modes, but its regulation and dynamics during substrate engagement have not been captured.
-
No EMC2-specific Mendelian disease: This likely reflects embryonic lethality of homozygous loss, but hypomorphic alleles or mosaic states have not been systematically searched for.
13. Proposed Follow-up Experiments
-
Characterize Lys255 acetylation: Use acetylation-mimicking (K255Q) and acetylation-dead (K255R) mutants to test whether this modification regulates EMC2–EMC3 binding affinity and insertase activity in reconstituted assays.
-
Resolve substrate-bound EMC structure: Use cryo-EM with stalled substrates (e.g., dominant-negative TA proteins) to capture the EMC with a client TMD in the vestibule.
-
Define EMC2-specific client spectrum: Perform TMT-based quantitative proteomics comparing EMC2-depleted versus EMC3-depleted cells to determine whether EMC2 has functions independent of the insertase.
-
Screen for EMC2 disease variants: Mine ClinVar and gnomAD for rare EMC2 missense variants at functionally critical residues (189–191, 180, 259) and test their effects on EMC assembly and client protein levels.
-
Test EMC2 in ferroptosis directly: Determine whether EMC2 knockdown sensitizes cells to ferroptosis inducers and whether this is mediated through loss of specific client biogenesis.
-
Explore antiviral potential: Determine whether partial EMC inhibition can suppress flavivirus replication without lethal cytotoxicity, leveraging residual insertion capacity from the parallel GET pathway.
References
- Guna A, Volkmar N, Christianson JC, Hegde RS. The ER membrane protein complex is a transmembrane domain insertase. Science. 2018;359(6374):470–473. PMID: 29242231.
- O'Donnell JP, Phillips BP, Yagita Y, et al. The architecture of EMC reveals a path for membrane protein insertion. eLife. 2020;9:e57887. PMID: 32459176.
- Miller-Vedam LE, Brauning B, Popova KD, et al. Structural and mechanistic basis of the EMC-dependent biogenesis of distinct transmembrane clients. eLife. 2020;9:e62611. PMID: 33236988.
- Pleiner T, Hazu M, Tomaleri GP, et al. WNK1 is an assembly factor for the human ER membrane protein complex. Mol Cell. 2021;81(13):2730–2742. PMID: 33964204.
- Pleiner T, Hazu M, Pinton Tomaleri G, et al. A selectivity filter in the ER membrane protein complex limits protein misinsertion at the ER. J Cell Biol. 2023;222(6):e202212007. PMID: 37199759.
- Volkmar N, Thezenas ML, Louie SM, et al. The ER membrane protein complex promotes biogenesis of sterol-related enzymes maintaining cholesterol homeostasis. J Cell Sci. 2019;132(2):jcs223453. PMID: 30578317.
- Li Y, Zhang Y, Xu L, et al. Structural insights into human EMC and its interaction with VDAC. EMBO J. 2024;43:1377–1398. PMID: 38517390.
- Wideman JG. The ubiquitous and ancient ER membrane protein complex (EMC): tether or not? F1000Res. 2015;4:624. PMID: 26512320.
- Barrows NJ, Anglero-Rodriguez Y, Kim B, et al. Dual roles for the ER membrane protein complex in flavivirus infection. J Virol. 2019;93(22):e01010-19. PMID: 31273220.
- Savidis G, McDougall WM, Meraner P, et al. Identification of Zika virus and dengue virus dependency factors using functional genomics. Cell Rep. 2016;16(1):232–246. PMID: 27342126.
- Satoh T, Ohba A, Liu Z, Inagaki T, Bhatt VS. dPob/EMC is essential for biosynthesis of rhodopsin and other multi-pass membrane proteins in Drosophila photoreceptors. eLife. 2015;4:e06306. PMID: 25715730.
- Xiong L, Zhang L, Yang Y, et al. ER complex proteins are required for rhodopsin biosynthesis and photoreceptor survival in Drosophila and mice. J Cell Biol. 2020;219(3):e201909094. PMID: 31263175.
- Sinning I, McDowell MA. Cryo-EM insights into tail-anchored membrane protein biogenesis in eukaryotes. Curr Opin Struct Biol. 2022;75:102428. PMID: 35850079.
- Page BM, Nguyen HT, Pleiner T, et al. Role of a holo-insertase complex in the biogenesis of biophysically diverse ER membrane proteins. Science. 2024;383(6684):eadj7880. PMID: 38076791.
- Stanton AC, Singal N, Biswal M, et al. The ER membrane protein complex acts as a chaperone to promote the biogenesis of multi-bundle membrane proteins. 2026. PMID: 41648177.
- Janer A, Prudent J, Paupe V, et al. SLC25A46 is required for mitochondrial lipid homeostasis and cristae maintenance. EMBO Mol Med. 2016;8(9):1019–1038. PMID: 27390132.
- Jung S, Bhatt P, Zimmermann R. Quantitative mass spectrometry characterizes client spectra of components for targeting and insertion into the ER membrane. Int J Mol Sci. 2023;24(19):14869. PMID: 37762469.
- Wang L, Wang Y, Gao S, Xie F. Novel compound heterozygous variants in EMC1. Mol Genet Genomic Med. 2023;11(7):e2177. PMID: 37187958.
- Alzayed S, Alzuabi M, Alqusaimi N, et al. Tribal founder EMC1 variant. Clin Genet. 2024;106(2):194–199. PMID: 38784058.
- Liu X, Yang J, Han R, Zhou H, Qu S, Shi Y. ncRNA-mediated overexpression of ferroptosis-related gene EMC2. Immun Inflamm Dis. 2021;9(4):1510–1527. PMID: 34956895.
- Shangguan Y, Huang L, Chen Y, et al. Prognostic assessment value of immune escape-related genes in AML. Sci Rep. 2025;15(1):33094. PMID: 39311489.
- Bagchi P, Speckhart K, Kennedy A, Tai AW, Bhatt VS. A specific EMC subunit supports dengue virus infection. PLoS Pathog. 2022;18(7):e1010717. PMID: 35834589.
- Raj S, Krishnan K, Askew DS, et al. The toxicity of a novel antifungal compound is modulated by ERAD components. mBio. 2015;6(6):e01861-15. PMID: 26666917.
- Hiramatsu N, Tago T, Satoh T, Satoh AK. ER membrane protein complex is required for the insertions of late-synthesized transmembrane helices of Rh1 in Drosophila photoreceptors. Mol Biol Cell. 2019;30(20):2577–2589. PMID: 31553680.
- McDowell MA, Heimes M, Fiorentino F, et al. Structural basis of tail-anchored membrane protein biogenesis by the GET insertase complex. Mol Cell. 2020;80(1):72–86. PMID: 32910895.
- Park JM, Mau CZ, Chen YC, et al. A case-control study in Taiwanese cohort and meta-analysis of serum ferritin in pancreatic cancer. Sci Rep. 2021;11(1):21242. PMID: 34711879.
- Rapaport D, Herrmann JM. Chasing the right tail: How the ER membrane complex recognizes its substrates. J Cell Biol. 2023;222(7):e202306065. PMID: 37436711.
Citations
- PMID:26512320
- PMID:32459176
- PMID:38517390
- PMID:35850079
- PMID:32910895
- PMID:37199759
- PMID:33964204
- PMID:29242231
- PMID:37762469
- PMID:33236988
- PMID:38076791
- PMID:41648177
- PMID:27390132
- PMID:30578317
- PMID:25715730
- PMID:31263175
- PMID:31553680
- PMID:31273220
- PMID:27342126
- PMID:35834589
- PMID:26666917
- PMID:37187958
- PMID:38784058
- PMID:37809982
- PMID:39311489
- PMID:34956895
- PMID:34059009
- PMID:34711879
- PMID:37436711